INDUKSI MUTASI TANAMAN GANDUM (
Triticum aestivum
L.)
MELALUI IRADIASI SINAR GAMMA SECARA
IN VITRO
UNTUK TOLERANSI TERHADAP SUHU TINGGI
RYAN BUDI SETIAWAN
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA*
Dengan ini saya menyatakan bahwa tesis berjudul Induksi Mutasi Tanaman Gandum (Triticum aestivum L.) Melalui Iradiasi Sinar Gamma Secara In vitro Untuk Toleransi Terhadap Suhu Tinggi adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
RINGKASAN
RYAN BUDI SETIAWAN. Induksi Mutasi Tanaman Gandum (Triticum aestivum L.) Melalui Iradiasi Sinar Gamma Secara In vitro Untuk Toleransi Terhadap Suhu Tinggi. Dibimbing oleh NURUL KHUMAIDA dan DINY DINARTI.
Dalam rangka mengurangi tingginya nilai impor gandum perlu dilakukan upaya penanaman dengan tetap memperhatikan faktor kesesuaian lingkungan. Program pemuliaan tanaman penting dilakukan untuk perbaikan genetik dan karakter sehingga berpengaruh positif terhadap peningkatan kualitas, kuantitas dan pengembangan gandum di Indonesia. Embriogenesis somatik memainkan peranan yang sangat besar dalam perbanyakan tanaman. Integrasi dengan program pemuliaan konvensional maupun teknik molekular/bioteknologi memungkinkan embriogenesis somatik menjadi suatu alat untuk meningkatan laju perbaikan genetik tanaman. Syarat utama dalam melakukan rekayasa genetik tanaman secara in vitro adalah kemampuan untuk menumbuhkan sel somatik dalam kondisi steril pada media pertumbuhan dan meregenerasikannya menjadi tanaman utuh.
Salah satu kendala pengembangan tanaman gandum di Indonesia adalah terbatasnya varietas yang beradaptasi terhadap lingkungan tropis. Terjadinya perubahan iklim akibat pemanasan global yang menyebabkan suhu permukaan bumi diperkirakan akan meningkat sekitar 0.3 oC per dekade. Persaingan lahan di dataran tinggi dengan tanaman hortikultura juga menghambat pengembangan gandum di Indonesia. Oleh karena itu perlu dilakukan upaya perakitan varietas gandum tropika yang mampu beradaptasi di dataran menengah (400-700 mdpl) dan toleran terhadap suhu tinggi. Pemuliaan tanaman menggunakan teknik mutasi melalui iradiasi sinar gamma dapat digunakan untuk peningkatan dan perbaikan genetik tanaman.
Tujuan umum dari penelitian ini adalah untuk memperoleh galur mutan gandum yang memeliki toleransi terhadap suhu tinggi. tujuan khusus penelitian ini adalah: mendapatkan metode induksi kalus embriogenik, embrio somatik dan proliferasi dari tiga varietas gandum (Dewata, Selayar dan Nias), menentukan nilai radiosensitivitas yang akan digunakan sebagai dasar induksi mutasi melalui iradiasi sinar gamma pada kalus embriogenik, serta melakukan uji cepat ketahanan suhu tinggi fase kecambah pada tiga varietas gandum. Embrio muda dan tua digunakan sebagai eksplan. Media dasar MS dengan penambahan kombinasi zat pengatur tumbuh (ZPT) 2.4D, picloram dan NAA digunakan dalam studi embriogenesis somatik. Hasil penelitian menunjukkan bahwa media induksi kalus embriogenik terbaik adalah media dasar MS dengan penambahan kombinasi ZPT 2.0 mg L-1 2.4D + 1.0 mg L-1 picloram dengan persentase pembentukan kalus embriogenik sebesar 60% pada varietas Dewata menggunakan eksplan immature embrio (embrio muda).
radiosensitivitas menggunakan lima dosis iradiasi sinar gamma (10, 20, 30, 40 dan 50 Gy). Seleksi ketahanan terhadap suhu tinggi dilakukan menggunakan inkubator pada suhu 25, 30 dan 35 oC. Nilai radiosensitivitas yang diperoleh bervariasi untuk setiap varietas yang berkisar (LD20: 7.79-18.96 Gy, LD50: 24.29-33.63 Gy).
Selayar merupakan varietas yang memiliki nilai sensitivitas lebih tinggi dibandingkan Dewata dan Nias. Meningkatnya dosis iradiasi dan suhu seleksi menurunkan bobot, diameter dan daya hidup kalus embriogenik, jumlah embrio, serta persentase planlet yang berkecambah. Berdasarkan seleksi in vitro menggunakan suhu tinggi (25, 30 dan 35 oC) diperoleh 19 planlet mutan putatif yang berasal dari embrio somatik yang muncul pada permukaan kalus embriogenik yang bertahan hidup pada varietas Dewata. Uji cepat ketahanan suhu tinggi pada fase kecambah dari tiga varietas gandum menunjukkan bahwa meningkatnya suhu hingga 35 oC mempengaruhi semua peubah yang diamati meliputi persentase daya berkecambah, panjang akar, jumlah akar, jumlah daun, panjang tunas, bobot basah tunas dan akar. Bedasarkan pengujian cepat tahap kecambah menunjukkan varietas selayar memiliki tingkat toleransi lebih baik terhadap suhu tinggi dibandingkan dengan varietas lainnya.
SUMMARY
RYAN BUDI SETIAWAN. Mutation Induction of Wheat (Triticum aestivum L.) through Gamma Ray Irradiation by In vitro culture for High Temperature Tolerance. Supervised by NURUL KHUMAIDA and DINY DINARTI
In order to reduce the high import value of wheat, it is necessary to planting with regard to the environmental suitability factors. Ultilization of plant breeding programs for genetic improvement give positive effect to improving the quality and quantity of wheat development in Indonesia. Somatic embryogenesis plays a large role in the plant propagation. Integration with conventional breeding and molecular engineering/biotechnology, allows somatic embryogenesis become a tool to increase the rate of plant genetic improvement. The main requirement in genetic modified plants as in vitro is the ability to regenerate somatic cells under sterile conditions in growth media and regenerate into whole plants.
One of the problem to developing wheat in Indonesia is limited varieties adapted to tropical environments. Climate change by global warming causes the earth's surface temperature is expected to rise about 0.3 oC per decade. Competition in the highland area with horticultura crops also inhibits the wheat development in Indonesia. Therefore, necessary to assembly of tropical wheat varieties which are able to adapt in the middle area (400-700 meter above sea level) and tolerant for high temperatures. The technique mutation through gamma ray irradiation may be used for plant genetic improvement.
The general objective of this research was to obtain wheat promising lines tolerant to high temperature. Specifically, this study has objectives that include the following: to obtain the methods for embryogenic callus, somatic embryo induction, proliferation and regeneration of three wheat variety (Dewata, Selayar and Nias), to obtain the radiosensitivity value that will be use as basic for mutation induction through gamma ray irradiation on embryogenic callus, to obtain the information about effect of high temperature for wheat sprout. Mature and immature embryos used as explants. MS with the addition plant growth regulator 2.4D, picloram and NAA used in the medium.
The best embryogenic callus induction medium on Dewata obtained by responsive varietiy to produce somatic embryos with normal planlet germination of 79.46%. Radiosensitivity test used five irradiation doses (10, 20, 30, 40 and 50 gray). Level gamma irradiation radiosensitivity from embryogenic callus are different among three wheat varieties (LD20: 7.79-18.96 gray, LD50: 24.29-33.63
gray).
mutants were derived from embryos in embryogenic callus survival after irradiation and high temperature selection. Testing the high temperature in phase of the three varieties of wheat sprouts showed that increasing the temperature to 35 °C greatly affects all observed variables include the percentage of germination, root length, number of roots, number of leaves, length of the shoots, fresh weight of the shoots and roots. Selayar is varietie which indicates the level of tolerance to high temperatures compared with other varieties.
© Hak Cipta Milik IPB, Tahun 2015
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains
pada Program Studi Pemuliaan dan Bioteknologi Tanaman
INDUKSI MUTASI TANAMAN GANDUM (
Triticum aestivum
L.)
MELALUI IRADIASI SINAR GAMMA SECARA
IN VITRO
UNTUK TOLERANSI TERHADAP SUHU TINGGI
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
BOGOR 2015
Judul Tesis : Induksi Mutasi Tanaman Gandum (Triticum aestivum L.) Melalui Iradiasi Sinar Gamma Secara In vitro Untuk Toleransi Terhadap Suhu Tinggi
Nama : Ryan Budi Setiawan NIM : A253120061
Disetujui oleh Komisi Pembimbing
Dr Ir Nurul Khumaida, MSi Ketua
Dr Ir Diny Dinarti, MSi Anggota
Diketahui oleh
Ketua Program Studi
Pemuliaan dan Bioteknologi Tanaman
Dr Ir Yudiwanti Wahyu EK, MS
Dekan Sekolah Pascasarjana
Dr Ir Dahrul Syah, MScAgr
Tanggal Ujian : 19 November 2014
PRAKATA
Puji dan syukur penulis ucapkan kepada Allah subhanahu wa ta’ala atas segala karunia dan nikmat-Nya sehingga tesis ini berhasil diselesaikan. Shalawat dan salam tercurah untuk Rasulullah Muhammad SAW atas semua perjuangan dan dakwah beliau. Tema yang dipilih dalam penelitian yang dilaksanakan sejak bulan Maret 2013 sampai Agustus 2014 ini adalah Induksi Mutasi Tanaman Gandum (Triticum aestivum L.) Melalui Iradiasi Sinar Gamma Secara In vitro Untuk Toleransi Terhadap Suhu Tinggi. Dengan terselesaikannya penulisan tesis ini, penulis ucapkan terima kasih yang sebesar-besarnya kepada:
1. Dr Ir Nurul Khumaida MSi dan Ibu Dr Ir Diny Dinarti MSi selaku komisi pembimbing yang telah banyak memberikan arahan, bimbingan, motivasi dan nasehat demi terselesaikannya tesis ini.
2. Dr Ir Yudiwanti Wahyu EK MS selaku ketua mayor Pemuliaan dan Bioteknologi Tanaman dan Seluruh Dosen, karyawan serta teknisi atas semua Ilmu dan bantuannya.
3. Dr Sintho Wahyuning Ardie SP MSi selaku penguji luar komisi atas semua kritik dan sarannya.
4. Prof Dr Ir Irfan Suliansyah MS, Prof Dr Ir Ardi MSc, dan Prof Dr Ir Warnita MS atas surat rekomendasi untuk melanjutkan studi pascasarjana di IPB. 5. DIKTI atas Beasiswa Unggulan tahun 2012-2014.
6. Bapak Prayitno dari Badan Tenaga Nuklir Nasional (BATAN) yang telah membantu dalam iradiasi sinar gamma.
7. Bapak, Mamak, dan adik-adik serta seluruh keluarga atas segala doa dan kasih sayangnya
8. Rekan-rekan pascasarjana PBT angkatan 2012 untuk semua kebersamaan dan perjuangannya.
9. Rekan-rekan Laboratorium Kultur Jaringan 3 untuk semua bantuannya 10. Azfani Nelza buat semua dukungan dan semangatnya.
11. Keluarga besar BDP angkatan 2007 (last generation) Universitas Andalas atas semua kebersamaanya.
Penulis menyadari bahwa tulisan ini masih jauh dari sempurna, namun penulis berharap semoga tesis ini bermanfaat bagi perkembangan ilmu pengetahuan, khususnya pengembangan gandum tropika di Indonesia.
DAFTAR ISI
DAFTAR ISI v
DAFTAR TABEL vi
DAFTAR GAMBAR vii
DAFTAR LAMPIRAN viii
1 PENDAHULUAN 1
Latar Belakang 1
Perumusan Masalah 2
Tujuan Penelitian 3
Manfaat Penelitian 3
Hipotesis Penelitian 3
Ruang Lingkup Penelitian 5
2 TINJAUAN PUSTAKA 7
3 EMBRIOGENESIS SOMATIK TIGA VARIETAS GANDUM 17
Pendahuluan 17
Bahan dan Metoda 18
Prosedur Analisis Data 20
Hasil dan Pembahasan 21
Simpulan 39
4 RADIOSENSITIVITAS DAN INDUKSI MUTASI SINAR GAMMA
UNTUK KETAHANAN TERHADAP SUHU TINGGI 40
Pendahuluan 40
Bahan dan Metoda 41
Prosedur Analisis Data 43
Hasil dan Pembahasan 43
Simpulan 56
5 UJI CEPAT KETAHANAN TERHADAP SUHU TINGGI PADA
FASE KECAMBAH 58
Pendahuluan 58
Bahan dan Metode 59
Prosedur Analisis Data 59
Hasil dan Pembahasan 59
Simpulan 64
6 PEMBAHASAN UMUM 66
7 SIMPULAN DAN SARAN 73
DAFTAR PUSTAKA 74
DAFTAR TABEL
1 Tipe, sumber dan deskripsi berbagai macam iradiasi pengion 12 2 Pengaruh media dan varietas terhadap persentase induksi kalus dari
eksplan immature embrio gandum 21
3 Pengaruh media dan varietas terhadap waktu muncul kalus gandum dari
eksplan immature embrio (hari) 22
4 Pengaruh media dan varietas terhadap persentase kalus embriogenik
dari eksplan immature embrio gandum 23
5 Pengaruh media dan varietas terhadap diameter kalus dari eksplan immature embrio gandum (mm) 25 6 Pengaruh media dan varietas terhadap persentase induksi kalus dari
eksplan mature embrio gandum 26
7 Pengaruh media dan varietas terhadap waktu muncul kalus dari eksplan mature embrio gandum (hari) 27 8 Pengaruh media dan varietas terhadap persentase kalus embriogenik
dari eksplan mature embrio gandum 27
9 Pengaruh media dan varietas terhadap diameter kalus dari eksplan mature embrio gandum (mm) 29 10 Skoring uji kruskal walis terhadap penutupan eksplan oleh kalus
gandum 32
11 Karakteristik embrio somatik gandum varietas Dewata dan Nias yang
berasal dari eksplan immature embrio 33
12 Pengaruh zat pengatur tumbuh terhadap pertambahan diameter kalus
embriogenik gandum (mm) 35
13 Pengaruh zat pengatur tumbuh terhadap pertambahan bobot kalus
embriogenik gandum (mg) 35
14 Bobot kalus embriogenik (KE) dan jumlah embrio somatik varietas Dewata hasil proliferasi pada perbedaan konsentrasi ZPT dan jenis
media 36
15 Bobot kalus embriogenik (KE) dan jumlah embrio somatik varietas Nias hasil proliferasi pada perbedaan konsentrasi ZPT dan jenis media 37 16 Total pertambahan diameter kalus gandum selama 6 minggu setelah
iradiasi sinar gamma (mm) 43
17 Pertambahan bobot kalus gandum (mg) pada 6 MSK setelah iradiasi
sinar gamma 45
18 Skoring perubahan warna kalus gandum hasil iradiasi sinar gamma
umur 6 MSK 46
19 Nilai LD20 dan LD50 kalus embriogenik tiga varietas gandum dari
eksplan immature embrio gandum hasil iradiasi sinar gamma 46 20 Pertambahan diameter kalus gandum selama 4 minggu setelah iradiasi
sinar gamma pada beberapa agen seleksi in vitro cekaman suhu tinggi
(mm) 49
21 Pertambahan bobot kalus selama 4 minggu setelah iradiasi sinar gamma pada beberapa agen seleksi in vitro cekaman suhu tinggi (mg) 50 22 Persentase dan jumlah total kalus gandum yang bertahan hidup selama
23 Karakteristik mutan putatif gandum toleran suhu tinggi yang dihasilkan
pada varietas Dewata 56
24 Pengaruh suhu dan varietas terhadap persentase daya berkecambah, panjang akar dan jumlah akar tiga varietas gandum 60 25 Pengaruh suhu dan varietas terhadap jumlah daun dan panjang tunas
tiga varietas gandum 62
26 Pengaruh suhu dan varietas terhadap bobot basah, bobot kering tunas
dan akar tiga varietas gandum 63
27 Batas suhu tinggi beberapa tanaman 71
DAFTAR GAMBAR
1 Kerangka berfikir penelitian studi embriogenesis dan seleksi in vitro
gandum toleran terhadap cekaman suhu tinggi 4
2 Diagram alir tahap penelitian studi embriogenesis dan seleksi in vitro
gandum toleran terhadap cekaman suhu tinggi 6
3 Perbandingan perkembangan embriogenesis zigotik dengan embriogenesis somatik pada tanaman dikotil dan monokotil 9 4 Model konsep mekanisme toleransi suhu tinggi yang dipengaruhi oleh
tiga faktor 16
5 Eksplan yang digunakan dalam induksi kalus gandum 19 6 Tipe kalus gandum yang terbentuk pada media induksi kalus 19
7 Skoring penutupan eksplan oleh kalus gandum 19
8 Perkembangan kalus gandum selama 8 MSK 22
9 Proporsi tipe kalus gandum yang berasal dari eksplan immature embrio 24 10 Embrio somatik varietas Dewata yang berasal dari beberapa media pada
umur 8 MSK 24
11 Proporsi tipe kalus gandum yang berasal dari eksplan mature embrio 28 12 Pengaruh jenis ekplan terhadap embriogenesis somatik gandum varietas
Dewata dan Selayar 30
13 Perkecambahan embrio somatik gandum 34
14 Proliferasi embrio somatik gandum 36
15 Jumlah embrio somatik gandum per fase histodiferensiasi 38 16 Tahapan perkembangan embrio somatik gandum dan histodiferensiasi
embrio di bawah mikroskop 39
17 Pertambahan diameter kalus pada uji radiosensitivitas tiga varietas
gandum 44
18 Skoring perubahan warna kalus gandum akibat iradiasi pada uji
radiosensitivitas 45
19 Grafik radiosensitivitas sinar gamma pada tiga varietas gandum 47 20 Perubahan warna kalus gandum akibat iradiasi dan cekaman suhu tinggi 51 21 Jumlah dan persentase perkecambahan embrio somatik setelah iradiasi
dan pengujian suhu tinggi varietas Dewata 54
22 Perkecambahan mutan putatif gandum pada varietas Dewata 55 23 Planlet mutan putatif toleran suhu tinggi hasil perkecambahan embrio
somatik. 56
DAFTAR LAMPIRAN
1 Komposisi media Murashige dan Skoog. 83
2 Penelitian embriogenesis somatik beberapa tanaman sereal. 84 3 Penelitian induksi mutasi beberapa tanaman sereal 87
1
PENDAHULUAN
Latar Belakang
Gandum (Triticum aestivum L.) merupakan salah satu tanaman pangan utama yang ditanam hampir di seluruh dunia. Luas penanaman gandum mencakup sekitar 30% areal penanaman sereal di dunia, dengan lebih dari 220 juta ha areal budidaya (Cossani dan Reynolds 2012). Kebutuhan dan konsumsi gandum masyarakat Indonesia sangat tinggi, hal ini ditunjukkan oleh impor pada tahun 2012 yang mencapai 6 300 000 ton (BPS 2013) setara dengan US$ 298 516 200 (Kemenperin 2013). Meningkatnya nilai impor mengindikasikan bahwa permintaan pasar terhadap gandum semakin tinggi, yang berarti peluang dan potensi pengembangannya di Indonesia sangatlah besar. Ketergantungan pangan dari luar negeri dikhawatirkan mengancam ketahanan pangan khususnya berkaitan dengan tanaman gandum.
Pengembangan tanaman gandum di Indonesia terkendala oleh berbagai hal yaitu terbatasnya varietas yang beradaptasi terhadap lingkungan tropis. Di daerah asalnya, gandum dibudidayakan pada suhu 8-20 oC dan menghendaki suhu 10-21
o
C sebagai suhu optimal untuk pertumbuhannya (Fischer dan Maurer 1978), sehingga pengembangan dan budidaya gandum di Indonesia lebih sesuai dilakukan di dataran tinggi (>900 m dpl) dengan suhu sekitar 22 °C. Pengembangan tanaman gandum juga terhambat akibat terbatasnya lahan penanaman di dataran tinggi, karena adanya persaingan dengan komoditas hortikultura yang lebih memiliki nilai komersial. Selain itu terjadinya perubahan iklim akibat pemanasan global juga menjadi kendala pengembangan gandum di Indonesia. Peningkatkan suhu sekitar 1.8 sampai 5.8 oC diprediksi akan terjadi diakhir abad ini (IPCC 2007), atau naik sekitar 1-3 oC dari keadaan suhu sekarang berturut-turut pada tahun 2025 dan 2100 (Wahid et al. 2007). Strategi adaptasi yang paling mendesak dilakukan untuk menanggulangi perubahan iklim dalam dunia pertanian menurut World Development Report (2008) antara lain: menanam varietas yang memiliki daya adaptasi tinggi dengan masa tanam yang lebih singkat, mengubah masa tanam, selain itu diperlukan penelitian yang intensif untuk perakitan varietas yang memiliki daya tahan tinggi terhadap kekeringan, banjir dan peningkatan temperatur. Upaya yang dapat dilakukan untuk pengembangan gandum di Indonesia adalah dengan perakitan varietas gandum tropika yang dapat beradaptasi dengan baik di dataran menengah dan toleran terhadap suhu yang lebih tinggi.
Teknik mutasi dapat digunakan untuk peningkatkan keragaman genetik sehingga memungkinkan pemulia untuk melakukan seleksi pada genotipe sesuai dengan tujuan yang dikehendaki. Mutagen yang biasanya digunakan dapat digolongkan menjadi dua kelompok yaitu mutagen fisik dan mutagen kimia. Mutagen fisik dapat berupa mutagen pengion yaitu sinar gamma, sinar x, sinar beta, sinar alpha dan sinar ultraviolet, sedangkan mutagen kimia yang paling umum digunakan adalah EMS (Ethyl methane sulfonate), DES (diethyl sulfonate), EI (ethyleneimine), ENV (ethyl nitroso urethan), ENH (Ethyl nitroso urea) dan MNH (methyl nitroso urea) (Jamsari 2008).
Iradiasi sinar gamma merupakan radiasi pengion yang mempunyai kekuatan daya tembus tinggi yang dipancarkan oleh sumber dari 60Co, 137Cs. Kemampuan penetrasi yang cukup kuat ke dalam jaringan tanaman ini dapat menyebabkan perubahan baik pada tingkat gen maupun kromosom. Perubahan yang terjadi dapat berupa delesi, duplikasi, insersi, dan translokasi. Terjadinya perubahan pada gen maupun kromosom akan mengakibatkan perubahan pada karakter tanaman (Shu et al. 2012).
Penggunaan sinar gamma sebagai mutagen untuk menginduksi mutasi dan menghasilkan karakter-karakter tertentu yang diinginkan telah banyak dipejari. Yusnita (2005) menginduksi variasi somaklonal dan melakukan seleksi in vitro untuk resistensi penyakit busuk batang sclerotium pada tanaman kacang tanah (Arachis hypogea L). El sayeed et al. (2007) mengunakan iradiasi sinar gamma untuk meningkatkan toleransi cekaman salinitas pada tanaman gandum (Triticum aestivum L.) secara in vitro. Ajijah (2009) mengaplikasikan induksi mutasi menggunakan sinar gamma pada tanaman purwoceng (Pimpinella pruatjan Molk.) untuk mendapatkan tanaman tahan terhadap suhu tinggi. Nur (2013) melakukan iradiasi sinar gamma pada benih gandum (Triticum aestivum L.) untuk peningkatan keragaman genetik dan toleransi terhadap suhu tinggi.
Perumusan Masalah
Ketahanan pangan nasional masih merupakan isu yang strategis bagi Indonesia mengingat kecukupan produksi, distribusi dan konsumsi pangan memiliki keterkaitan yang erat dengan masalah sosial, ekonomi dan politik. Oleh karena itu, ketahanan pangan merupakan program utama dalam pembangunan pertanian saat ini dan masa mendatang. Ketahanan pangan merupakan suatu sistem yang terintegrasi yang terdiri atas berbagai subsistem, subsistem utamanya adalah ketersediaan pangan, distribusi dan konsumsi pangan. Terwujudnya ketahanan pangan merupakan sinergi dari interaksi ketiga subsistem tersebut.
Permasalahan pangan di Indonesia sangat komplek. Penurunan pembangunan sektor pangan dan pertanian pasca reformasi menyebabkan Indonesia mengalami kesulitan untuk memenuhi dan menjaga ketahanan pangan nasional yang terlihat dari meningkatnya impor pada berbagai produk pertanian. Gandum merupakan tanaman pangan yang kebutuhannya secara keseluruhan di impor gandum dari luar negeri. Nilai impor yang tinggi dan memiliki kecenderungan meningkat setiap tahunnya dikhawatirkan mengganggu stabilitas dan ketahanan pangan nasional jika seandainya pasokan gandum impor berkurang.
beradaptasi pada lingkungan tropis dan memiliki toleransi terhadap suhu tinggi. Program pemuliaan tanaman penting dilakukan untuk perbaikan genetik dan karakter sehingga berpengaruh positif terhadap peningkatan kualitas, kuantitas dan pengembangan gandum di Indonesia. Embriogenesis somatik memainkan peranan yang sangat besar dalam perbanyakan tanaman. Integrasi dengan program pemuliaan konvensional maupun teknik molekular/ bioteknologi memungkinkan embriogenesis somatik menjadi suatu alat dalam peningkatan laju perbaikan genetik tanaman. Kerangka berfikir penelitian disajikan pada Gambar 1.
Tujuan Penelitian
Secara umum penelitian ini bertujuan untuk mendapatkan varian gandum yang dapat dibudidayakan di dataran menengah (400-700 m dpl) dan toleran terhadap suhu tinggi. Tujuan khusus penelitian ini adalah sebagai berikut:
1. Mempelajari pengaruh berbagai jenis dan konsentrasi zat pengatur tumbuh (ZPT) terhadap induksi kalus embriogenik dan embrio somatik pada beberapa varietas gandum menggunakan eksplan immature dan mature embrio.
2. Mendapatkan media proliferasi kalus embriogenik dan embrio somatik yang optimum pada beberapa varietas gandum.
3. Mendapatkan nilai LD20 dan LD50 pada kalus embriogenik dari setiap
varietas gandum.
4. Mempelajari karakter morfologi kecambah gandum pada cekaman suhu tinggi.
Manfaat Penelitian
1. Penelitian ini diharapkan dapat memperoleh protokol embriogenesis somatik dari setiap varietas gandum meliputi metode induksi serta proliferasi kalus embriogenik dan embrio somatik.
2. Penelitian ini diharapkan dapat memperoleh nilai radiosensitivitas kalus embriogenik dari setiap varietas gandum yang akan digunakan sebagai dasar untuk melakukan induksi mutasi dan seleksi in vitro.
3. Penelitian ini diharapkan dapat memperoleh mutan putatif yang memiliki ketahanan terhadap suhu tinggi.
4. Penelitian ini diharapkan dapat memperoleh informasi ketahanan suhu tinggi lebih awal menggunakan pengujian cepat fase kecambah benih dari setiap varietas gandum.
Hipotesis Penelitian
1. Terdapat media induksi kalus embriogenik dan embrio somatik serta media proliferasi embrio somatik yang optimal pada setiap varietas gandum.
2. Terdapat nilai LD20 dan LD50 kalus embriogenik dari setiap varietas
3. Terdapat mutan putatif gandum yang toleran cekaman suhu tinggi.
4. Terdapat perbedaan karakter morfologi gandum fase kecambah pada cekaman suhu tinggi.
Gambar 1 Kerangka berfikir penelitian studi embriogenesis dan seleksi in vitro gandum toleran terhadap cekaman suhu tinggi
Kandidat galur harapan toleran suhu tinggi
Konvensional
Introduksi
Hibridisasi Konsumsi
gandum tinggi Nilai impor
meningkat
Pertambahan jumlah penduduk Pemborosan devisa negara
Program kemandirian dan kedaulatan pangan
Perluasan areal penanaman 1 Perbaikan teknik budidaya
2 Pengembangan varietas unggul gandum tropika
Perbaikan varietas
Bioteknologi
Variasi somaklonal
Induksi mutasi
Kultur in vitro
Rekayasa genetika Pemanfaatan lahan di
dataran menengah (400-700 m dpl)
Seleksi in vitro dengan inkubator suhu tinggi Persaingan lahan
dataran tinggi
Cekaman suhu tinggi
Ruang Lingkup Penelitian
Penelitian diawali dengan sterilisasi benih muda dan tua untuk menghasilkan kultur asenik. Immature dan mature embrio diisolasi dari benih dan dijadikan sebagai eksplan yang dikulturkan pada sepuluh jenis media induksi kalus embriogenik dan embrio somatik. Proliferasi kalus embriogenik dilakukan menggunakan tiga jenis media media. Tahap selanjutnya, embrio somatik yang telah dihasilkan diproliferasikan pada dua jenis media, yaitu media padat dan cair dengan komposisi ZPT yang berbeda. Proses perkecambahan embrio somatik dari setiap media induksi kalus menjadi planlet menggunakan media MS tanpa ZPT (MS0).
Percobaan induksi mutasi diawali dengan melakukan penentuan Lethal Doses 20 (LD20) dan LD50 dengan perlakuan iradiasi sinar gamma pada lima
dosis iradiasi yaitu: 10, 20, 30, 40, 50 gray. Bahan yang digunakan adalah kalus embriogenik varietas Dewata, Nias dan Selayar. Nilai radiosensitivitas (LD20 dan
LD50) yang diperoleh digunakan sebagai acuan dalam mutasi in vitro pada kalus
Gambar 2 Diagram alir tahap penelitian studi embriogenesis dan seleksi in vitro gandum toleran terhadap cekaman suhu tinggi
Uji cepatcekaman suhu tinggi fase kecambah (25, 30 dan 35 oC) Sterilisasi immature dan mature seed
Induksi kalus embriogenik dan embrio somatik pada 10 jenis media
Proliferasi embrio somatik pada 2 jenis media (padat dan cair)
Penentuan radiosensitivitas, LD20 dan LD50 kalus
embriogenik varietas Dewata, Nias dan Selayar (iradiasi sinargamma dengan dosis 10, 20, 30, 40, dan
50 gray)
Protokol embriogenesis somatik Penyiapan Immature dan mature seed tiga varietas gandum
(Dewata, Nias dan Selayar)
Perkecambahan embrio somatik Mutasi induksi sinar gamma dan seleksi in vitro
ketahanan terhadap suhu tinggi (menggunakan agen seleksi cekaman suhu (25, 30, 35 oC)
Perkecambahan embrio somatik hasil seleksi in vitro menjadi planlet utuh (MS tanpa ZPT)
Karakterisasi mutan putatif planlet
Mutan putatif gandum toleran terhadap suhu tinggi
Benih tiga varietas gandum (Dewata, Selayar dan Nias)
Informasi dan uji cepat skrining gandum terhadap cekaman suhu tinggi Proliferasi kalus embriogenik pada 3
2
TINJAUAN PUSTAKA
Botani Tanaman Gandum
Budidaya tanaman gandum telah berkembang sejak 7500- 6500 sebelum masehi (SM) di daerah Timur Tengah, 5000 SM di sekitar sungai Nil dan 3000 SM di Cina. Negara produsen utama gandum adalah Rusia, USA, Cina, India, Perancis, dan Kanada. Tahun 1529 Spanyol memperkenalkan gandum ke Amerika yang merupakan benua baru (Briggle 1980). Masyarakat prasejarah sudah mengenal gandum dan berbagai biji-bijian lainnya sebagai sumber makanan. Berdasarkan penggalian arkeolog diperkirakan gandum berasal dari daerah sekitar laut merah dan laut mediterania, yaitu daerah sekitar Turki, Syam, Irak, dan Iran (Sleper dan Poehlman 2006).
Gandum termasuk divisi Spermatophyta, kelas Angiospermae, subkelas Monocotylodenae, ordo Graminae, famili Poaceae, dan genus Triticum. Spesies-spesies yang termasuk di dalam genus Triticum dikelompokkan ke dalam tiga kelas ploidi yaitu diploid (2n=2x=14), tetraploid (2n=4x= 28) dan heksaploid (2n=6x=42). Saat ini terdapat 11 spesies diploid, 12 spesies tetraploid, dan 6 spesies heksaploid yang sudah diidentifikasi dan dideskripsikan (Sleper dan Poehlman 2006). Namun hanya dua spesies dari genus Triticum yang memiliki nilai ekonomis penting yaitu
Triticum aestivum L. dan Triticum turgidum L.. Triticum aestivum L. merupakan
gandum yang umum dikenal yang dimanfaatkan untuk bahan baku roti. Triticum
turgidum L. yang dikenal dengan gandum durum digunakan untuk membuat pasta.
Sleper dan Poehlman (2006) mengklasifikasikan gandum berdasarkan kegunaannya yang meliputi gandum keras (hard wheat) yang memiliki kandungan gluten dan protein tinggi serta cocok untuk pembuatan roti, gandum lunak (soft wheat) yang memiliki kandungan gluten dan protein yang lebih rendah, cocok untuk pembuatan kue-kue kering, biskuit, dan crackers, dan gandum durum yang memiliki kandungan gluten dan protein sangat rendah, cocok untuk pembuatan macaroni dan spaghetti.
Gandum termasuk tanaman semusim yang menyerbuk sendiri ( self-polinated), penyerbukan silang hanya 1-4% yang biasanya disebabkan oleh angin atau serangga. Bunga-bunganya bermekaran pada pertengahan pagi menjelang siang. Kemampuan reseptif stigma berkisar 4-13 hari sedangkan viabilitas serbuk sari hanya sekitar 30 menit. Kondisi masak fisiologis dicapai apabila kandungan kelembaban dari keseluruhan bulir yang terbentuk telah menurun antara 25-35% (Ginkel dan Villareal 1996).
Umumnya, kernel berbentuk oval dengan panjang 6-8 mm dan diameter 2-3 mm. Seperti jenis serealia lainnya, gandum memiliki tekstur yang keras. Biji gandum terdiri dari tiga bagian yaitu bagian kulit (bran), bagian endosperma, dan bagian lembaga (germ). Bagian kulit dari biji gandum sebenarnya tidak mudah dipisahkan karena merupakan satu kesatuan dari biji gandum tetapi bagian kulit ini biasanya dapat dipisahkan melalui proses penggilingan (Ginkel dan Villareal 1996).
Bran merupakan kulit luar gandum dan terdapat sebanyak 14.5% dari total keseluruhan gandum. Bran terdiri dari 5 lapisan yaitu epidermis (3.9%), epikarp (0.9%), endokarp (0.9%), testa (0.6%), dan aleuron (9%). Bran memiliki kandungan protein dan kadar serat tinggi sehingga baik dikonsumsi ternak besar. Epidermis merupakan bagian terluar bijigandum, mengandung banyak debu yang apabila terkena air akan menjadi liat dan tidak mudah pecah. Fenomena inilah yang dimanfaatkan pada penggilingan gandum menjadi tepung terigu agar lapisan epidermis yang terdapat pada biji gandum tidak hancur dan mengotori tepung terigu yang dihasilkan. Endosperma merupakan bagian terbesar dari biji gandum (80-83%) yang banyak mengandung protein dan pati. Pada proses penggilingan, bagian inilah yang akan diambil sebanyak banyaknya untuk diubah menjadi tepung terigu dengan tingkat kehalusan tertentu (Ginkel dan Villareal 1996).
Kultur Jaringan
Kultur jaringan adalah suatu metode untuk mengisolasi bagian dari tanaman seperti protoplasma, sel, jaringan dan organ yang kemudian menumbuhkannya dalam lingkungan yang aseptik, sehingga bagian-bagian tanaman tersebut dapat memperbanyak diri dan beregenerasi menjadi tanaman utuh kembali. Salah satu penerapan kultur jaringan adalah perbanyakan mikro. Perbanyakan mikro secara umum dapat diartikan sebagai usaha menumbuhkan bagian tanaman dalam media aseptik dan memperbanyaknya hingga menghasilkan tanaman sempurna. Tujuan utama penerapan perbanyakan mikro adalah produksi tanaman dalam jumlah besar dalam waktu yang singkat terutama untuk varietas-varietas unggul serta memperoleh tanaman yang terbebas dari serangan patogen. Eksplan yang ditanam pada media tumbuh yang tepat dapat beregenerasi melalui proses yang disebut organogenesis atau embriogenesis (Neumann et al. 2009).
Embriogenesis Somatik
Embriogenesis somatik merupakan metode perbanyakan masal yang banyak diterapkan pada berbagai macam tanaman. Menurut Corredoira et al. (2006), pemahaman tentang embriogenesis tanaman penting diketahui yaitu melalui pengamatan proses seluler yang mendasari diferensiasi. Hal tersebut akan bermanfaat untuk memperoleh metode yang tepat pada aplikasi perbanyakan bibit secara masal, kryopreservasi, induksi mutasi dan transformasi genetik.
Kultur jaringan memungkinkan regenerasi sel somatik melalui dua alternatif jalur yaitu organogenesis dan embriogenesis somatik. Organogenesis adalah pembentukan tunas adventif secara langsung dari eksplan yang menghasilkan struktur unipolar, sedangkan embriogenesis somatik adalah proses regenerasi sel somatik yang membelah dan berkembang menjadi struktur bipolar atau embrio. Struktur unipolar terdiri atas tunas atau akar yang terpisah jaringan pembuluhnya, sedangkan struktur bipolar terdiri atas meristem tunas dan akar yang padu (Gunawan 1992).
Menurut Davies (2004) embrio somatik dapat terbentuk melalui 2 cara yaitu embriogenesis secara langsung dan tidak langsung. Embriogenesis langsung terjadi pada saat embrio terbentuk langsung dari jaringan eksplan atau Pre Embryonically Determined Cells (PEDC). Embriogenesis tidak langsung terjadi melalui fase kalus atau kultur suspensi yang biasa disebut Induced Embryonically Determined Cells (IEDC). Menurut Rose et al. (2010) tahap perkembangan embrio somatik dan embrio zigotik memliki pola yang sama yaitu dimulai dari fase globular, hati, torpedo dan planlet untuk tanaman dikotil dan globular, hati, scutelar dan coleoptilar pada monokotil (Gambar 3).
Gambar 3 Perbandingan perkembangan embriogenesis zigotik dengan embriogenesis somatik pada tanaman dikotil dan monokotil (Greer 2008)
somatik menjadi sebuah hal yang sangat penting dalam mempelajari dan menganalisis berbagai peristwa biokimia dan molekuler, sehingga memiliki nilai potensial yang besar bagi ilmu biologi dan pertanian. Gray (2005) menyatakan bahwa sel/jaringan somatik pada kondisi normal akan berkembang menjadi jaringan parenkim, tetapi dapat beralih menjadi embriogenik pada kondisi khusus. Embrio mampu diinduksi dari sel somatik diduga karena dikendalikan oleh gen dan adanya kemampuan totipotensi sel untuk tumbuh menjadi tanaman lengkap. Wattimena (1992) menyebutkan bahwa kemampuan jaringan eksplan membentuk kalus dan laju pertumbuhan kalus dapat berbeda-beda pada setiap tanaman. Kemampuan jaringan membentuk kalus dan laju pertumbuhan kalus tergantung pada media, ZPT yang digunakan dan faktor lingkungan lainnya.
Menurut Purnamaningsih (2002), faktor yang mempengaruhi pembentukan embrio somatik adalah: 1) Jenis eksplan, penggunaan eksplan yang bersifat meristematik umumnya memberikan keberhasilan pembentukan embrio somatik yang lebih tinggi. Eksplan yang digunakan dapat berupa aksis embrio zigotik muda dan dewasa, kotiledon, mata tunas, epikotil maupun hipokotil. Eksplan yang digunakan dapat berbeda tergantung jenis tanaman dan tahap perkembangan dari eksplan. 2) Sumber nitrogen, embriogenesis somatik mengalami proses perkembangan morfologi seperti yang terjadi pada embrio zigotik. Faktor yang penting dalam induksi dan perkembangan embriogenesis somatik adalah komposisi nutrisi pada media kultur. Nitrogen merupakan faktor utama dalam memacu morfogenesis secara in vitro. 3) Gula, gula merupakan salah satu komponen organik yang harus diberikan ke dalam media tumbuh. Gula berfungsi disamping sebagai sumber karbon, juga berguna untuk mempertahankan tekanan osmotik media. 4) ZPT.
Pemuliaan Mutasi
Upaya perbaikan kuantitas dan kualitas suatu varietas tanaman dapat ditempuh melalui program pemuliaan tanaman. Pemuliaan tanaman bertujuan untuk memperbaiki karakter tanaman sehingga menjadi lebih unggul dalam beberapa sifat dibandingkan dengan tanaman asalnya, misalnya tanaman menjadi lebih tahan terhadap serangan hama dan penyakit, tahan terhadap cekaman lingkungan, berproduksi lebih tinggi dan memiliki kandungan gizi yang lebih baik (Shu et al. 2012).
Pemuliaan tanaman merupakan pekerjaan yang berkesinambungan mulai dari proses peningkatan keragaman genetik tanaman, dilanjutkan dengan seleksi, pemurnian benih, pengujian hingga pelepasan varietas unggul tanaman. Adanya keragaman genetik yang tinggi sangat diharapkan dalam program pemuliaan tanaman sehingga peluang keberhasilan dalam proses seleksi tanaman unggul menjadi lebih besar. Metoda yang umum digunakan dalam upaya peningkatan keragaman genetik tanaman melalui program pemuliaan adalah melalui introduksi, hibridisasi, bioteknologi, induksi mutasi dan rekayasa genetika (Shu et al. 2012).
molekular. Secara umum penampilan mutan dapat dilihat dengan tiga cara: perbedaan morfologi, perbedaan tingkat biokimia dan molekular, dan perbedaan tingkat adaptasi terhadap lingkungan tumbuh. Hasil mutasi yang paling mudah dilihat bila terjadi perubahan morfologi seperti bentuk, ukuran dan warna (Jusuf 2001).
Menurut Shu et al. (2012) mutagen yang sering digunakan terbagi menjadi tiga yaitu: 1) radiasi pengionan, 2) radiasi non pengionan, 3) radiasi menggunakan bahan kimia. Mutagen kimia yang banyak digunakan oleh pemulia adalah yang berasal dari kelompok alkylating agents. Senyawa ini mengandung satu atau lebih kelompok alkil reaktif yang dapat ditransfer ke molekul lain pada posisi dimana kerapatan elektronnya tinggi. Alkylating agents ini akan bereaksi dengan DNA dengan cara mengalkilasi kelompok fosfat, termasuk basa-basa purin dan pirimidin. Mutagen kimia yang biasa digunakan diantaranya adalah EMS (ethyl methane sulphonate), DES (diethylsulfate), El (ethylenimine), NMUT (nitroso methyl urethane), NMU (nitro methyl urea), NEUT (nitrose ethyl urethane) dan NEU (nitrose ethyl urea).
Mutasi berdasarkan tingkatan terjadinya dapat digolongkan menjadi mutasi genom, mutasi kromosom, mutasi gen dan mutasi diluar inti sel. Mutasi genom disebabkan karena terjadinya perubahan jumlah dari genom tanaman, misalnya akibat euploid maupun aneuploid. Mutasi kromosom diakibatkan karena pecahnya benang kromosom (chromosome breakage). Perubahan yang terjadi pada struktur kromosom tanaman digolongkan dalam empat macam proses yaitu: 1) Duplikasi yaitu terjadinya penambahan segmen kromosom, 2) Delesi yaitu terjadinya pengurangan segmen kromosom, 3) Inversi yaitu pemutaran segmen kromosom, 4) Translokasi yaitu pindah silang antar kromosom nonhomolog (Aisyah 2006).
Iradiasi Sinar Gamma
Teknologi nuklir berperan dalam pemuliaan tanaman terutama terkait dengan kemampuannya untuk menimbulkan mutasi. Kemampuan tersebut dikarenakan tenaga nuklir memiliki energi yang cukup tinggi sehingga jika sinar tersebut memapar pada materi genetik akan menimbulkan perubahan struktur pada komposisi pada materi genetik. Mutasi adalah perubahan yang terjadi pada materi genetik sehingga menyebabkan perubahan ekspresi. Perubahan dapat terjadi pada tingkat pasangan basa, tingkat satu ruas DNA, bahkan pada tingkat kromosom (Jusuf 2001).
paling efektif dalam menghasilkan varietas mutan unggul berbagai jenis tanaman. Berbagai macam tipe radiasi pengion dapat dilihat pada tabel 1.
Tabel 1 Tipe, sumber dan deskripsi berbagai macam iradiasi pengion
Tipe radiasi Sumber Deskripsi Energi Resiko Penetrasi
Sinar X Mesin X-ray Radiasi
Sinar Gamma Radioisotop Radiasi
elektromagnetik
Beberapa MeV
Berbahaya Seluruh
bagian
Neutron Reaksi nuklir Hanya dapat dilihat
jika berintekasi
Partikel alpha Radioisotop Inti helium, ion berat 2-9 MeV Sangat
berbahaya
Beberapa mm
Proton/ deuteron
Reaksi nuklir Inti hidrogen Beberapa
GeV
Iradiasi sinar gamma merupakan teknologi radiasi bagian dari teknologi nuklir yang menggunakan radioisotop. Dibandingkan mutagen kimia, radioisotop memiliki kelebihan sifat fisik yaitu memancarkan sinar radioaktif. Keberhasilan perlakuan iradiasi sangat ditentukan oleh sensitivitas genotipe yang diiradiasi terhadap dosis radiasi yang diberikan. Tingkat sensitivitas tanaman dipengaruhi oleh jenis tanaman, fase tumbuh, ukuran, dan bahan yang akan dimutasi, serta sangat bervariasi antar jenis tanaman dan antar genotype. Shu et al. (2012) menyatakan bahwa semakin banyak kadar oksigen dan molekul air (H2O) berada dalam materi yang diradiasi, maka semakin banyak pula radikal bebas yang terbentuk sehingga materi menjadi semakin sensitif.
Sensitivitas terhadap radiasi dapat diukur berdasarkan nilai LD50 (lethal
dose 50) yaitu dosis yang menyebabkan kematian 50% dari populasi tanaman yang diradiasi. Dalam induksi mutasi, beberapa studi menunjukkan bahwa dosis optimum yang dapat menghasilkan mutan dengan tingkat keragaman tertinggi umumnya diperoleh disekitar LD50 (Datta 2001). Untuk mendapatkan nilai LD50
menyebabkan perubahan abnormal pada fenotipe tanaman. Pengaruh dosis radiasi terhadap persen kematian, pertumbuhan, dan fertilitas telah banyak dilaporkan. Penelitian yang dilakukan oleh Puchooa (2005) mengenai iradiasi sinar gamma pada tanaman Anthurium menyatakan dosis iradiasi sekitar 15 Gy menyebabkan kematian 50% pada kalus. Mahadevamma et al. (2012) menunjukkan dosis iradiasi sebesar 30 Gy menyebabkan sekitar 50% kalus pepaya mengalami kematian yang ditunjukkan oleh perubahan warna dan berhentinya pertumuhan kalus. Sejalan dengan itu penelitian Ling et al. (2008) menyatakan LD50 kalus
jeruk sekitar 27 Gy. Efek iradiasi sinar gamma sangat dipengaruhi oleh jenis tanaman, variteas serta kandungan air dan oksigen pada sel kalus.
Terdapat beberapa teknik yang biasa digunakan dalam pengaplikasian induksi mutasi dengan mutagen fisik (radiasi), yaitu (a) iradiasi tunggal (acute irradiation), (b) chronic irradiation, (c) Iradiasi terbagi (fractionated irradiation), dan (d) Iradiasi berulang. Teknik iradiasi tunggal (acute irradiation) dilakukan dengan cara pemberian dosis secara sekaligus dalam satu kali penembakan iradiasi. Chronic irradiation adalah teknik iradiasi dengan cara memberikan dosis iradiasi yang sangat rendah secara terus menerus dalam waktu beberapa bulan. Teknik iradiasi terbagi (fractionated irradiation) dapat dilakukan dengan pemberian dosis iradiasi sebanyak dua kali, dimana tiap perlakuan diberikan setengah dosis. Sedangkan teknik iradiasi berulang adalah teknik iradiasi yang dilakukan melalui pemberian dosis secara berulang dengan jarak waktu yang tidak terlalu lama (intermittent irradiation) atau dapat pula diberikan dengan jarak waktu yang cukup lama (recurrent irradiation) (Aisyah 2006).
Pengaruh Cekaman Suhu Tinggi dan Mekanisme Ketahanannya
Peningkatan suhu sangat berpengaruh pada fase pertumbuhan vegetatif dan reproduktif tanaman gandum. Phasad et al. (2008) mengemukakan bahwa cekaman suhu tinggi diatas 22 oC secara signifikan menurunkan hasil panen gandum, suhu tinggi selama masa antesis dapat menyebabkan aborsi dari floret, meningkatkan sterilitas polen, dehidrasi jaringan, menurunkan proses asimilasi karbon dan meningkatkan fotorespirasi. Meningkatnya suhu malam diatas 20 oC juga menurunkan fertilitas polen sehingga mengurangi jumlah dan ukuran biji. Meningkatnya suhu sangat mempengaruhi proses fotosintesis, reduksi fotosintesis berpengaruh pada fase vegetatif maupun generatif pada tanaman gandum. rendahnya fotosintesis akibat cekaman suhu tinggi disebabkan rusaknya struktur dan fungsi dari kloroplas sehingga mengurangi jumlah klorofil, selain itu meningkatnya suhu menyebabkan inaktifasi enzim yang berperan pada proses metabolisme seperti mengganggu fungsi enzim rubisco yang berperan dalam mengatur karboksilasi selama proses fotosintesis.
berperan dalam proses biosintesis pati. Sucrose sintase dan SSS sangat penting dalam menentukan laju biosintesis pati sedangkan granule bound starch sintase berperan mengontrol biosintesis amilosa. Suhu tinggi dapat menurunkan aktifitas enzim pada tanaman gandum sehingga menghambat perkembangan biji dan akumulasi pati (Prakash et al. 2004). Suhu tinggi sangat berkaitan dengan laju pengisian biji dimana waktu pengisian biji akan berkurang.
Meningkatnya suhu 5 oC diatas 20 oC akan meningkatkan laju pengisian biji tetapi mengurangi durasi pengisian biji sekitar 12 hari pada tanaman gandum (Yin et al. 2009). Saat suhu tinggi suplai fotosintat ke dalam biji mungkin juga terhambat. Hasil penelitian Streck (2005) menunjukkan naiknya suhu 1 oC diatas suhu optimal (15-20 oC) menyebabkan rata-rata waktu pengisian biji menurun sekitar 2.8 hari. Suhu tinggi selama masa pengisian biji juga berpengaruh terhadap kualitas biji, melalui berkurangnya jumlah protein dan terjadinya reduksi pati (Gooding et al. 2003).
Farooq et al. (2011) menyatakan bahwa mekanisme ketahanan tanaman gandum terhadap suhu tinggi dipengaruhi oleh tiga faktor yaitu: 1) sistem pertahanan antioksidan, 2) ekspresi gen HSPs, dan 3) stay green. Cekaman suhu tinggi akan menyebabkan terbentuknya radikal bebas yang lebih dikenal dengan reaktif oxygen species (ROS) misalnya superoksida, hidroksil dan hidrogen perosikda. Genotipe yang toleran akan mampu menangani dan mendetoksifikasi radikal bebas yang terbentuk sehingga tidak berpengaruh buruk terhadap proses metabolisme. Berbagai macam enzim antioksidan yang berperan dalam mekanisme ketahanan terhadap suhu tinggi diantaranya superoksida dismutase yang mampu merubah O2- pada hidrogen peroksida, catalase yang mampu
mengeliminasi hidrogen peroksida melalui dekomposisi H2O dan O2, serta
askorbat peroksidase yang mampu mengurangi kandungan hidrogen peroksida di dalam sel (Goyal dan Asthir 2010). Balla et al. (2009) menunjukkan bahwa saat tercekam suhu tinggi selama fase reproduktif aktivitas enzim antioksidan seperti catalase dan superoksida dismutase akan mengalami peningkatan pada genotipe gandum yang toleran.
Ekspresi heat shock protein (HSPs) telah banyak dipelajari sebagai respon awal saat tanaman tercekam suhu tinggi. HSPs berperan untuk melindungi protein dari kerusakan akibat peningkatan suhu. Meningkatnya suhu akan menyebabkan sintesis protein akan berkurang tetapi HSPs akan diproduksi di dalam sel (Rampino et al. 2009). Nguyen et al. (1994) menyatakan bahwa mRNA yang mengkode gen HSPs kelas utama dengan bobot molekul rendah yaitu HSP16 akan disintesis pada tanaman gandum yang ditanam pada suhu 32-35 oC. Penelitian lain juga menyatakan bahwa terjadi ekspresi HSP18 saat terjadi peningkatan suhu 3.2-3.6 oC diatas suhu normal pada genotipe gandum yang toleran terhadap suhu tinggi (Harma-Natu et al. 2010), dan penelitian yang dilakukan oleh Sumesh et al. (2008) juga menyatakan bahwa terjadi ekspresi HSP100 saat peningkatan suhu pada genotipe gandum yang toleran. Prorein dehidrin yang termasuk dalam kelompok 2 late embryogenesis abundant protein (LEA) diketahui juga mampu menstabilkan makro molekul dan melindungi enzim-enzim yang berperan pada awal metabolisme saat meningkatnya suhu (Brini et al. 2010).
lebih dari 20 gen HSF telah diidentifikasi pada tingkat sequen DNA, dimana 20 gen ini memiliki struktur dan fungsi yang berbeda (Nover et al. 2001). Fungsi HSF adalah memediasi ekspresi dan melindungi fungsi dari gen target seperti gen yang mengkode HSP. Meskipun HSP penting dalam toleransi terhadap cekaman tapi gen tersebut tidak bisa melalukan proteksi komponen sel secara terus menerus tanpa bantuan gen HSF. Gen HSF tidak hanya terlibat dalam regulasi HSP tapi juga dibutuhkan saat kondisi tidak tercekam (Panchuk et al. 2002). Menurut Xu et al. (2012) Gen HSF yang berikatan dengan heat shock element (HSE) mampu mengaktifkan fungsi dari gen yang mengkode HSP. Dalam kondisi normal, monomer HSF akan berkombinasi dengan HSP90, tetapi dibawah kondisi tercekam monomer HSF akan berubah menjadi trimer dan secara tepat akan berikatan dengan HSE untuk menginisiasi proses transkripsi dari gen HSP. Gen HSP tidak hanya diinduksi oleh cekaman suhu tinggi tetapi juga oleh banyak faktor cekaman lainnya.
Penuaan daun (leaf senescens) merupakan indikator kerusakan kloroplas saat terjadinya peningkatan suhu terutama jika peningkatan suhu terjadi pada saat pembungaan dan pengisian biji. Oleh karena itu, Peningkatan dan penjagaan klorofil serta efisiensi kapasitas fotosintesis atau yang dikenal dengan stay green dapat menjadi penanda toleransi terhadap suhu tinggi. Kehilangan klorofil akan berpengaruh terhadap proses fotosintesis dan pengisian biji. Genotipe yang tetap hijau (stay green) harusnya lebih bisa menjaga pengisian biji saat terjadi peningkatan suhu (Farooq et al. 2011)
Menurut Cossani dan Reynolds (2012) kualitas dan kuantitas hasil gandum saat terkena cekaman suhu tinggi dan mekanisme adaptasinya dipengaruhi oleh beberapa faktor yaitu: 1) Perubahan karakter yang berkaitan dengan intersepsi cahaya matahari misalnya, terjadinya penggulungan daun secara cepat dan kemampuan tanaman untuk menjaga kloroplas dari kerusakan sehingga tanaman tetap hijau (stay green). 2) Perubahan karakter yang berkaitan dengan efisiensi penggunaan radiasi matahari, meliputi perubahan morfologi daun, munculnya lapisan lilin, peningkatan produksi klorofil a/b, caratenoid dan antioksidan, efisiensi penggunaan air melalui pengaturan transpirasi, peningkatan fiksasi CO2,
3
EMBRIOGENESIS SOMATIK TIGA VARIETAS GANDUM
Pendahuluan
Program pemuliaan tanaman perlu dilakukan dalam rangka perbaikan genetik dan karakter unggul sehingga berpengaruh positif terhadap peningkatan kualitas, kuantitas dan pengembangan gandum di Indonesia. Berbagai macam cara dapat dilakukan dalam rangka perbaikan genetik seperti hibridisasi, kultur in vitro, mutasi dan tranformasi. Embriogenesis somatik memainkan peranan yang sangat penting dalam perbanyakan tanaman. Integrasi dengan program pemuliaan konvensional maupun teknik molekular dan bioteknologi memungkinkan embriogenesis somatik menjadi suatu alat dalam peningkatan laju perbaikan genetik tanaman. Embriogenesis somatik merupakan proses dimana sel somatik dalam kondisi induktif akan mampu menghasilkan sel embriogenik yang menjalani serangkaian perubahan morfologi dan biokimia dalam pembentukan embrio somatik (Komamine et al. 2005).
Perbaikan genetik secara in vitro dari berbagai tanaman sereal termasuk gandum dapat mengatasi berbagai cekaman biotik dan abiotik yang menyebabkan penurunan hasil (Jones et al. 2005). Namun, hal ini sangat tergantung pada metode keberhasilan regenerasi eksplan melalui teknik kultur in vitro. Oleh karena itu penetapan protokol yang tepat seperti induksi kalus, proliferasi, perkecambahan dan aklimatisasi sangat diperlukan (Noor et al. 2009; Yu et al. 2008). Syarat utama dalam merekayasa genetik tanaman secara in vitro adalah kemampuan untuk menumbuhkan sel somatik dalam kondisi steril pada media pertumbuhan dan meregenerasikannya menjadi tanaman utuh.
Pembentukan kalus embrigenik pada tanaman umumnya sangat tergantung pada genotipe, sumber eksplan, kondisi fisiologis tanaman donor, tipe jaringan, media, jenis dan konsentrasi ZPT, serta interaksi faktor-faktor tersebut (Gray 2005). Berbagai macam sumber eksplan telah banyak dipelajari dalam rangka studi embriogenesis somatik tanaman gandum seperti immature infloresence (Barro et al. 1999; He dan Lazzeri 2001), leaf basal (Chugh dan Khurana 2003; Mahalakshmi et al. 2003), mikrospora (Zheng 2003), mature embrio (Malik et al. 2004; Rahman et al. 2008; Munazir et al. 2010; Nasab et al. 2012), dan immature embrio (Jones et al. 2005; Fahmi et al. 2006; Yasmin et al. 2009). Sarker dan Biswas (2002) melaporkan bahwa eksplan immature embrio merupakan eksplan yang paling responsif untuk menginduksi terbentuknya kalus embriogenik dan regenerasinya menjadi planlet dibandingkan dengan sumber eksplan lain.
dilaporkan seperti aplikasi tranformasi genetik (Jones et al. 2005), peningkatan toleransi terhadap cekaman kekeringan (Mahmood et al. 2012), salinitas (Zair et al. 2003; El Sayeed et al. 2007), suhu tinggi (Benderradji et al. 2012) dan ketahanan terhadap penyakit (Svabova dan Lebeda 2005). Tujuan dari percobaan ini adalah mempelajari dan mendapatkan media serta eksplan yang terbaik untuk menginduksi kalus embriogenik, proliferasi kalus embriogenik, proliferasi embrio somatik dan perkecambahannya pada setiap varietas gandum.
Bahan dan Metoda
Penelitian ini dilaksanakan di Laboratorium Kultur Jaringan 3 Departemen Agronomi dan Hortikultura Fakultas Pertanian, Institut Pertanian Bogor dan Laboratorium Biologi Sel Universitas Gajah Mada, mulai bulan Maret 2013 sampai Februari 2014. Eksplan yang digunakan adalah immature dan mature embrio dari tiga varietas gandum yaitu Dewata, Selayar dan Nias (Gambar 4).
Induksi kalus embriogenik tiga varietas gandum
Eksplan yang digunakan adalah immature dan mature embrio yang diisolasi dari benih muda dan tua (Gambar 5). Benih muda yang berumur ± 21 hari setelah antesis (HSA) disterilkan dengan merendamnya dalam larutan fungisida (kandungan bahan aktif markozeb) dan bakterisida (kandungan bahan aktif achimidol) dengan konsentrasi 2 g L-1 selama 30 menit, kemudian dibilas dengan aquadest steril dan direndam dengan 20% larutan chlorox (kandungan bahan aktif natrium hipoklorit 5.25%) selama 30 menit dan dibilas lagi, selanjutnya dicelupkan dalam alkohol 70% selama tiga menit. benih tua (masak fisiologis) disterilkan dengan cara dicuci menggunakan detergen kemudian dibilas dengan air mengalir selama 60 menit. Benih kemudian direndam dalam larutan fungisida dan bakterisida dengan konsentrasi 2 g L-1 selama 60 menit, kemudian direndam dengan aquadest steril selama 1 malam. Selanjutnya benih direndam dengan 100% larutan chlorox selama 45 menit dan dibilas lagi, benih direndam dalam alkohol 70% selama tiga menit dan dibilas tiga kali dengan aquadest steril. Untuk memisahkan embrio dari benih dilakukan dengan cara menjepit benih menggunakan pinset hingga embrio keluar.
gelap selama 4 minggu, kemudian dipindahkan ke ruangan dengan penyinaran 16 jam/hari selama 4 minggu dengan tingkat penyinaran 1500 lux.
Peubah yang diamati meliputi: persentase eksplan membentuk kalus, waktu induksi kalus, tipe kalus, tipe 1: kalus remah transparan dan berair, tipe 2: kalus kompak transparan dan berair, tipe 3: kalus kompak kuning kecoklatan, tipe 4: kalus remah kuning kehijauan (Gambar 6), pertumbuhan kalus yang diukur dengan skoring, skor 1: eksplan tidak membentuk kalus, skor 2: ≤ 25% kalus menutupi eksplan, skor 3: 25-50% kalus menutupi eksplan, skor 4: 50-75% kalus menutupi eksplan, skor 5: 75-100% kalus menutupi eksplan (Gambar 7), diameter clump kalus, persentase kalus embriogenik dan analisis histodiferensiasi.
Gambar 5 Eksplan yang digunakan dalam induksi kalus gandum.a) biji muda, b) biji tua (masak fisiologis), c) Immature embrio. Bar berlaku untuk semua gambar.
Gambar 6 Tipe kalus gandum yang terbentuk pada media induksi kalus. a) tipe 1: kalus remah, transparan dan berair, b) tipe 2: kalus kompak, transparan dan berair, c) tipe 3: kalus kompak kuning kecoklatan, d) tipe 4: kalus remah kuning kehijauan.Bar berlaku untuk semua gambar
Gambar 7 Skoring penutupan eksplan oleh kalus gandum. a) embrio, b) skor 1: eksplan tidak membentuk kalus, c) skor 2: < 25% kalus menutupi eksplan, d) skor 3: 25-50% kalus menutupi eksplan), e) skor 4: 50-75% kalus menutupi eksplan, f) skor 5: 75-100% kalus menutupi eksplan. Bar berlaku untuk gambar pada baris yang sama
d c
b a
b c
1 cm
0.5 cm
a b c
f e
d
c b
a 1 mm
Proliferasi kalus embriogenik tiga verietas gandum
Media dasar yang digunakan yaitu media MS dengan penambahan ZPT 2.4D dan picloram, sukrosa 30 g L-1 serta gelrite 2 g L-1. Rancangan percobaan disusun berdasarkan rancangan acak lengkap. Perlakuan yang digunakan adalah komposisi media proliferasi kalus embriogenik yaitu media MS dengan penambahan: 1.0 mg L-1 2.4D, 1.0 mg L-1 2.4D + 1.0 mg L-1 picloram dan 2.0 mg L-1 2.4D + 1.0 mg L-1 picloram. Setiap perlakuan diulang sebanyak 5 kali sehingga terdapat 15 satuan percobaan dan setiap satuan percobaan dikulturkan 5 clump kalus embriogenik yang berukuran ±0.5 cm x 0.5 cm. Kultur diinkubasi pada suhu 20±4 ºC dengan penyinaran selama 16 jam/ hari selama 4 minggu. Peubah yang diamati adalah diameter dan bobot kalus embriogenik.
Proliferasi embrio somatik
Media dasar yang digunakan yaitu media MS dengan penambahan ZPT 2.4D, picloram, sukrosa 30 g L-1 dan gelrite 2 g L-1. Rancangan percobaan disusun berdasarkan rancangan acak lengkap faktorial. Faktor pertama adalah jenis media dengan dua taraf (padat dan cair), sedangkan faktor kedua adalah komposisi media proliferasi embrio somatik yaitu media MS dengan penambahan:1.0 mg L-1 2.4D + 0.5 mg L-1 picloram dan 0.5 mg L-1 2.4D + 0.25 mg L-1 picloram. Terdapat 4 kombinasi perlakuan dengan 5 kali ulangan sehingga diperoleh 20 satuan percobaan dan setiap satuan percobaan dikulturkan 1 clump kalus embriogenik yang mengandung embrio somatik dengan ukuran ±0.5 cm x 0.5 cm. Kultur diinkubasi pada suhu 20±4 ºC dengan penyinaran selama 16 jam/hari selama 8 minggu. Peubah yang diamati adalah bobot kalus embriogenik dan jumlah embrio somatik yang terbentuk per fase histodiferensiasi.
Perkecambahan embrio somatik
Embrio somatik yang telah diperoleh kemudian dipindahkan ke media perkecambahan yaitu MS0, 30 g L-1 sukrosa, 2 g L-1 gelrite. Kultur diinkubasikan
dengan penyinaran 1 000 lux selama 16 jam sehari pada suhu 20±4 ºC. Peubah yang diamati meliputi: persentase perkecambahan embrio somatik, persentase planlet normal dan abnormal.
Prosedur Analisis Data
Hasil dan Pembahasan
Induksi kalus embriogenik dan embrio somatik pada beberapa varietas gandum tropika menggunakan eksplan immature embrio
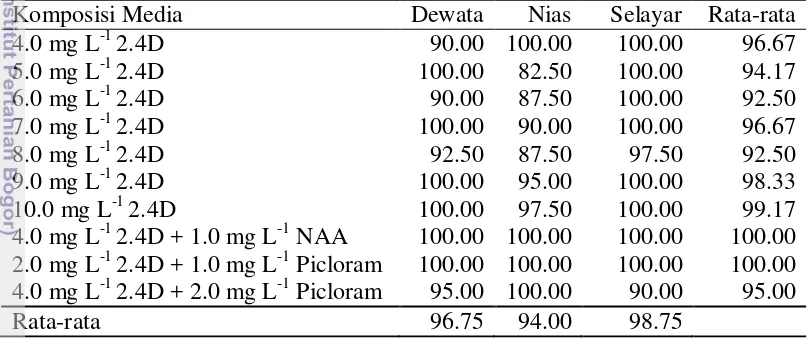
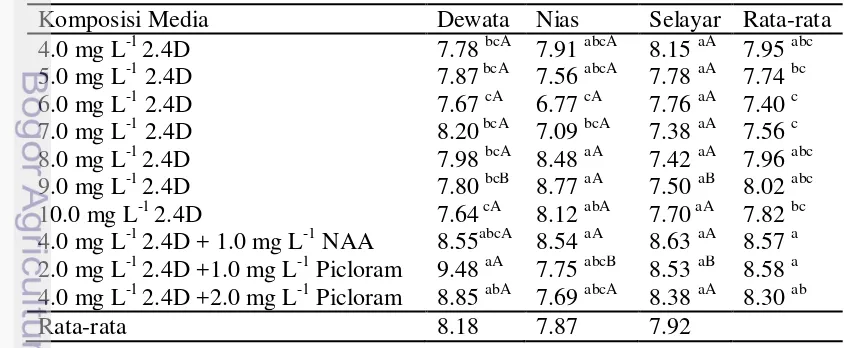
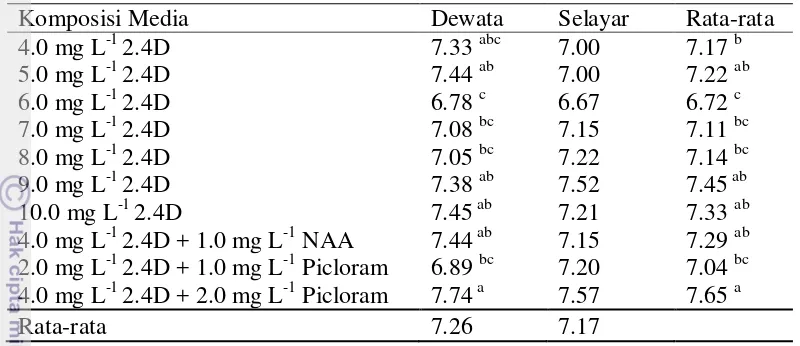
Sepuluh macam media yang digunakan berhasil untuk menginduksi pembentukan kalus tanaman gandum dari tiga varietas yaitu Dewata, Nias dan Selayar. Hasil analisis ragam menunjukkan bahwa perlakuan jenis varietas dan media yang digunakan secara tunggal maupun interaksinya tidak memberikan pengaruh terhadap persentase induksi kalus. Rata-rata persentase eksplan membentuk kalus pada ketiga varietas sangat tinggi yaitu 96.75% pada Dewata, 94.00% pada Nias dan 98.75% pada Selayar. Rata-rata persentase eksplan membentuk kalus pada kesepuluh media yang digunakan berkisar 92.50-100.00% (Tabel 2).
Tabel 2 Pengaruh media dan varietas terhadap persentase induksi kalus dari eksplan immature embrio gandum
Komposisi Media Dewata Nias Selayar Rata-rata
4.0 mg L-1 2.4D 90.00 100.00 100.00 96.67
5.0 mg L-1 2.4D 100.00 82.50 100.00 94.17
6.0 mg L-1 2.4D 90.00 87.50 100.00 92.50
7.0 mg L-1 2.4D 100.00 90.00 100.00 96.67
8.0 mg L-1 2.4D 92.50 87.50 97.50 92.50
9.0 mg L-1 2.4D 100.00 95.00 100.00 98.33
10.0 mg L-1 2.4D 100.00 97.50 100.00 99.17
4.0 mg L-1 2.4D + 1.0 mg L-1 NAA 100.00 100.00 100.00 100.00 2.0 mg L-1 2.4D + 1.0 mg L-1 Picloram 100.00 100.00 100.00 100.00 4.0 mg L-1 2.4D + 2.0 mg L-1 Picloram 95.00 100.00 90.00 95.00
Rata-rata 96.75 94.00 98.75
Gambar 8 Perkembangan kalus gandum selama 8 MSK. a-h pertumbuhan kalus berturut-turut pada umur 1-8 MSK. Bar berlaku untuk gambar pada baris yang sama
Hasil analisis ragam menunjukkan bahwa perlakuan komposisi media dan jenis varietas secara tunggal tidak memberikan pengaruh terhadap waktu munculnya kalus, sedangkan interaksi kedua faktor menunjukkan pengaruh yang nyata. Rata-rata waktu muinculnya kalus pada varietas Dewata berkisar 4.40 hari dengan menggunakan media 4.0 mg L-1 2.4D + 1.0 mg L-1 NAA sampai 4.86 hari pada media 7.0 mg L-1 2.4D, pada varietas Selayar kalus muncul setelah 4.26 dengan menggunakan media 8.0 mg L-1 2.4D sampai 4.74 hari menggunakan media 4.0 mg L-1 2.4D + 2 mg L-1 picloram, sedangkan pada pada varietas Selayar berkisar dari 4.23 hari menggunakan media 2.0 mg L-1 2.4D + 1.0 mg L-1 picloram sampai 4.94 pada media 9.0 mg L-1 2.4D (Tabel 3). ZPT dari golongan auksin seperti 2.4D, NAA dan picloram sangat berperan terhadap insiasi kalus. Munculnya kalus ditandai diawali dengan pembengkakan eksplan, kemudian terbentuk kalus yang berwarna putih bening disekitar eksplan. Perkembangan kalus diakibatkan adanya pembelahan pada sel-sel kalus.
Tabel 3 Pengaruh media dan varietas terhadap waktu muncul kalus gandum dari eksplan immature embrio (hari)
Komposisi Media Dewata Nias Selayar Rata-rata
4.0 mg L-1 2.4D 4.48 aA 4.30 aA 4.68 aA 4.49 5.0 mg L-1 2.4D 4.78 aA 4.39 aA 4.63 aA 4.60 6.0 mg L-1 2.4D 4.56 aA 4.59 aA 4.90 aA 4.68 7.0 mg L-1 2.4D 4.86 aA 4.50 aB 4.56 aB 4.64 8.0 mg L-1 2.4D 4.55 aAB 4.26 aB 4.75 aA 4.52 9.0 mg L-1 2.4D 4.67 aA 4.56 aA 4.93 aA 4.72 10.0 mg L-1 2.4D 4.79 aA 4.54 aA 4.59 aA 4.64 4.0 mg L-1 2.4D + 1.0 mg L-1 NAA 4.40 aA 4.72 aA 4.65 aA 4.59 2.0 mg L-1 2.4D + 1.0 mg L-1 Picloram 4.53 aA 4.68 aA 4.23 aA 4.48 4.0 mg L-1 2.4D + 2.0 mg L-1 Picloram 4.60 aA 4.74 aA 4.68 aA 4.67
Rata-rata 4.62 4.53 4.66
Keterangan: Angka-angka yang diikuti oleh huruf kecil yang sama pada kolom yang sama dan huruf besar yang sama pada baris yang sama menunjukkan tidak berbeda nyata
berdasarkan uji DMRT pada taraf α = 5%.
a b c d
e f g h 0.5 cm
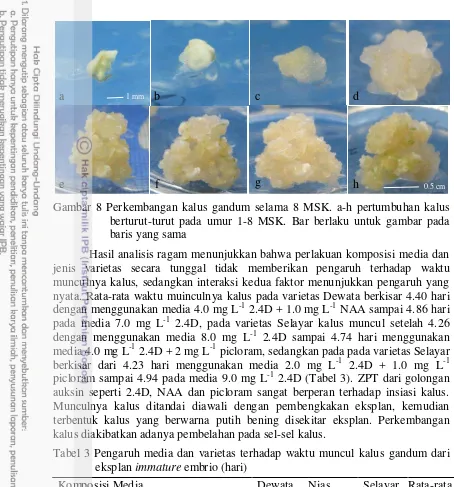
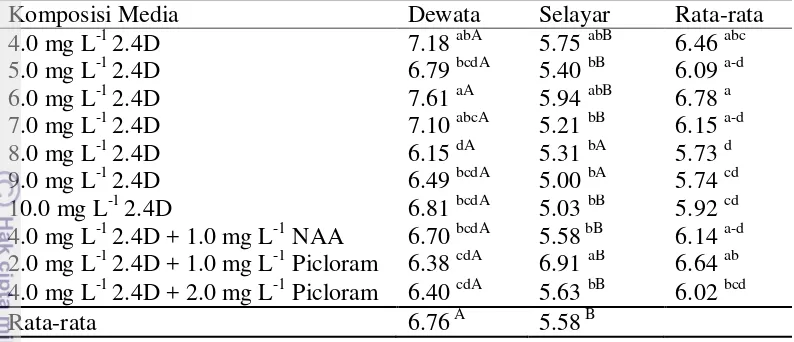
Hasil analisis ragam menunjukkan bahwa perlakuan komposisi media dan jenis varietas secara tunggal maupun interaksinya berpengaruh nyata terhadap persentase kalus embriogenik. Persentase kalus embriogenik pada varietas Dewata berkisar 60.00% menggunakan media 2.0 mg L-1 2.4D + 1.0 mg L-1 picloram hingga 3.13% menggunakan media 10.0 mg L-1 2.4D, pada varietas Nias sekitar 18.75% menggunakan media 4.0 mg L-1 2.4D + 2 mg L-1 picloram, dan pada varietas Selayar sekitar 13.89% menggunakan media 8.0 mg L-1 2.4D. Rata-rata persentase kalus embriogenik tertinggi dari semua media sebesar 22.85% pada varietas Dewata, 7.26% pada varietas Selayar dan 6.24% pada varietas Nias Varietas Dewata menunjukkan skoring kalus tipe 2 dengan proporsi rata-rata pada semua media sekitar 65.72%, varietas Selayar menunjukkan kalus skoring tipe 2 dengan sebesar 77.9%, sedangkan varietas Nias dengan proporsi 76.28% (Gambar 9).
Tabel 4 Pengaruh media dan varietas terhadap persentase kalus embriogenik dari eksplan immature embrio gandum
Komposisi Media Dewata Nias Selayar Rata-rata
Pembentukan kalus yang regenerable pada tanaman umumnya sangat tergantung pada genotipe, sumber eksplan, kondisi fisiologis tanaman donor, tipe jaringan, media, jenis dan konsentrasi ZPT, serta interaksi faktor-faktor tersebut (Gray 2005). Kalus tipe 4 yang terbentuk saat induksi diduga berpotensi menjadi kalus embriogenik karena memiliki struktur remah berwarna kuning kehijauan. Warna kalus yang kuning kehijauan dengan struktur yang remah memperlihatkan adanya kapasitas embriogenik dan potensinya membentuk embrio somatik (Gambar 10).
2-3 MSK. Kalus embriogenik dapat dilihat dari perubahan morfologi kalus dan perubahan inti sel yang terlihat padat dan jelas pada pengujian histologi.
Gambar 9 Proporsi tipe kalus gandum yang berasal dari eksplan immature embrio Pembentukan kalus embriogenik dan embrio somatik pada media yang ditambah picloram dan NAA menunjukkan bahwa penambahan kedua jenis auksin ini mampu menginduksi sel-sel yang mempunyai kemampuan membentuk embrio somatik dibandingkan dengan hanya menggunakan 2.4D tunggal. Kemampuan sel-sel membentuk embrio somatik berhubungan erat dengan aktivitas gen yang spesifik yang mempengaruhi embriogenesis somatik (Jones et al. 2005). Faktor lain yang mempengaruhi embriogenesis adalah faktor endogen. Umehara et al. (2007) menyatakan bahwa embriogenesis tanaman dipengaruhi oleh gen transkripsi seperti LEC1, LEC2, FUS3, ABI3/VP1. Faktor endogen lain yang juga mempunyai peran dalam embriogenesis adalah kandungan molekul yang rendah seperti auksin, giberellin (GA), asam absisik (ABA) yang dapat mendorong embriogenesis somatik dan senyawa fenolik yang dapat menghambat embriogenesis somatik.
Gambar 10 Embrio somatik varietas Dewata yang berasal dari beberapa media pada umur 8 MSK. a) 4.0 mg L-1 2.4D+1.0 mg L-1 NAA, b) 4.0 mg L-1 2.4D+2.0 mg L-1 picloram, c) 2.0 mg L-1 2.4D+1.0 mg L-1 picloram
Induksi kalus embriogenik dan embrio somatik dipengaruhi oleh varietas, jenis dan konsentrasi auksin yang digunakan. Dalam hal ini terdapat hubungan sensitivitas sel atau jaringan terhadap auksin (picloram atau 2.4D) yang