EFEK SUPLEMENTASI ASAM LEMAK
VOLATIL
BERCABANG
DAN
KAPSUL LISIN
SERTA TREONIN
TERHADAP NUTRISI PROTEIN SAPI HOLSTEIN
Oleh
:
WARDHANA SURYAPRATAMA
PROGRAM PASCASARJANA
INSTII'CIT I'EKTANIAN
BOGOH
THE EFFECTS OF BRANCHED CHAIN VOLATILE FATTY ACIDS AND
ENCAPSULATED LYSINE AND THREONINE SUPPLEMENTS ON
PROTEIN NUTRITION OF GROWING HOLSTEIN
Wardhana Suryapratama
Under Supervision of Toha Sutardi as Chairman of Dissertation Committee, Djokowoerjo Sastradipradja, H. Suryahadi, Kartiarso, and H. Mohammad Winugroho as Member of the
Committee.
ABSTRACT
Protein nutrition in ruminants is frequently limited by inadequate bacterial growth in the rumen and the lack of amino acids supplies for postruminal absorption. This study tried to stimulate bacterial growth through supplementation of isoacids and enrichment of postruminal supply of lysine, threonine, and essential fatty acids.
Trial 1 was a 5x5 Latin Square experiment in young Holstein bulls, 348
*
29 kg, where treatments were A: basal diet, B: A+
139 mg urealkg w 7 5 , C: B+
28 mg CaS0,Ikgw7',
D: C+
0.05 mmol isobutyrate+
0.05 mmol P-methylbutyrate, and E: D+
0.05 mmol a-methylbutyrate. The basal diet composed of 55% forage and 45% concentrate containing 10.5 MJ MEIkg and 15% crude protein (CP). Dry matter intake was not affected by the treatments but significant changes were noted in the rumen fermentation parameters. Urea addition (treatment B) increased cultivable rumen bacteria from 6.44 x 10" to 8.50 x 10" colonieslml (p<0.05). Addition of CaSO, (treatment C) reduced rurnen bacteria to the similar level as that of treatment A (p<0.05). The reduction might be due to an excess of Ca that resulted in mineral imbalance. Addition of isobutyrate and b-methylbutyrate (treatment D)increased rumen bacteria from 5.93 x 10" to 7.54 x 10" colonies/ml, suggesting that the isoacids stimulated bacterial growth. The failure of a-methylbutyrate to stimulate further bacterial growth indicated that the best proportion of the isoacids was not equimolar. The number of bacterial colonies was in a good agreement with the quantity of allantoin excreted in urine. Regression of urinary allantoin excretion (A, mglday) on the square root of bacterial colonies (B) was A = (2.6.B 0.445)/(2.62
+
B 0.445) with an R2 = 0.551 and Sb = 0.0817.SUMMARY
Wardhana Suryapratama. The Effects of Branched Chain Volatile Fatty Acids and Encapsulated Lysine and Threonine Supplements on Protein Nutrition o f Growing Holstein (Under Supervision o f Toha Sutardi a s C h a i r m a n of D i s s e r t a t i o n Committee, Djokowoerjo Sastradipradja, H.
Suryahadi, Kartiarso, and H. Mohammad Winugroho as Member of the Committee).
!
Protein nutrition i n ruminants i s frequently limited by inadequate bacterial growth i n the rumen and t h e lack of amino acids supplies for postruminal absorption. I n fact, t y p e of feeding on ruminants often ignores serving sufficient nutrient t o biomass synthesis of rumen microbes. This phenomenon mainly will influence feed digestion in rumen and in common will influence supply o f protein nutrient to host.
Supply of amino acids into postruminal intestine of ruminants mainly come from microbial protein that is synthesized in rumen, also feeds' protein that is undigested in rumen. This figures explained that ruminants' feed should not only support the growth of rumen microbial but also capable introduce amino acids needed by ruminants and the feed should be easy t o be absorbed in small intestine. Base o n the figure above, a sequence experiment has been done in order t o unify nutrient supplementation technology t o optimize bioprocess in rumen and essential amino acids introducing technology into post-rumen through rumen degradation resistant capsulation.
I
containing 10.5 MJ MEIkg and 15% crude protein (CP). Treatment was urea, calcium sulfate, isobutyrate, P-methylbutyrate, and a-methyl- butyrate. So, there were five dietary treatments where A: basal diet, B: A
+
139 mg urealkg w ~ . ~ ~ , C: B+
28 mg CaS041kgw ~ . ~ ~ ,
D: C+
0.05 mmol isobutyrate+
0.05 mmol P-methylbutyrate, and E: D+
0.05 mmol a-methylbutyrate. Rearing period was 30 days, consists of feed adaptation period 20 days, then growth observation was done within the last 10 days. Total collection data was done within the last 5 days of observation period.
The result of the trial 1 showed that nutrient consumption did not show the significant respond t o each treatment ration. This result figured that no nutrient supplementation given met the minimal limit that disturbs the appetite of animals. In addition, nutrient digestibility has not been affected by the treatment. The rations were capable providing rumen ammonia in an ideal concentration t o support the growth of rumen microbial. I t was about 8 mM. Total volatile fatty acid (VFA) concentration was in an ideal concentration t o rumen microbial growth. I t was about 110 mM.
There was an indication that the supplementation has been effected fermenta-tion in rumen mainly VFA profile both total and individual. The total VFA was A: 103, B: 102, C: 98, D: 99.3, and E: 95.6 mM. However, the individual VFA profiles represented by acetic, propionic and butyric acids. The average of acetic acid concentration was A: 65.2, B: 61.4, C: 60.2, D: 60.1, and E: 60.5 mM. The average of propionic acid was A: 27.3, B: 27.3, C: 24.6, D: 27.1, and E: 25.8 mM; and the average of
w
butyric acid was A: 4.86, B: 4.66, C: 4.75, D: 4.75, and E: 4.02 mM. Urea addition (treatment B) increased cultivable rumen bacteridfrom 6.44
by decreasing total VFA concentration. Addition of isobutyrate and
P-
methylbutyrate (treatment D) increased rumen bacteria from 5.93 x 10" t o7.54 x 10" colonies/ml, suggesting that the isoacids stimulated bacterial growth. The failure of a-methyl-butyrate t o stimulate further bacterial growth indicated that the best proportion of the isoacids was not equimolar. The number of bacterial colonies was in a good agreement with the quantity of allantoin excreted in urine. Regression of urinary allantoin excretion (A, mglday) on t h e square root of bacterial colonies (B) was A = (2.6.B 0.445)/(2.62
+
B 0.445) with anR~
= 0.55 1 and Sb = 0.0817. In D and E treatments, the body weights gain significantly decreased ( p < 0 . 0 5 ) .There was two possibilities, the amount of isoacid added (0.05 mmol) maybe did not enough t o fulfill the rumen microbe requirement, also maybe it is caused by unqualified basal ration figured out by the high concentration of urine nitrogen and decreasing nitrogen retention. This fact shows that the released dietary protein was very low, so the ruminant was deficient t o protein t o be stored by the body.
The trial 2, it aimed t o manipulate post-rumen absorption system using 5 Holstein heifers with initial weight of 160
*
43 kg in Latin Square 5 x 5 . The basal diet composed of 45% Napier grass (Pennisetum purpureum var. Africa) and 55% concentrate (2 part's coconut meal and 1part pollard) containing 10.5 MJ ME/kg and 15% crude protein (CP). Five treatments were A: treatment E a s in Trial 1
+
1.5% corn oil, B: A+
1.5 ml corn oil enclosed in formaldehyde treated gelatin capsule, C: B
+
encapsulated lysine, 28 mg/kgw'.~',
D: C+
100 mg encapsulated threonine, and E: D+
1.5% cod liver oil. Rearing period was 30 days, consists of feed adaptation period 20 days, then growth observation was done within the last 10 days. Total collection data was donflwithin the last 5 days of observation period.appetite. Dry matter and protein digestibility were significantly influenced by the treatments. Generally, the ration used in this experiment was capable t o serve rumen ammonia in an ideal concentration t o support the growth of rumen microbial. I t was about 8 mM. Also, total VFA concentration was in an ideal range t o support the growth of rumen microbial, it was 110 mM. Addition of encapsulated corn oil (treatment B) increased dry matter digestibility from 73.9 t o 77.5% (pC0.05) and crude protein digestibility from 87.8 t o 92.0% (p<0.05). Presumable, corn oil addition (treatment A) will induce saponification reaction of calcium so the negative effect of exceed calcium can be reduce. Addition of
I
encapsulated lysine (treatment C) and threonine (treatment D) increased the corresponding amino acid concentration in the blood plasma, indicating an increase in the absorption. However, comparisons of plasma amino acid ratio prior t o and 3-4 hr after feeding showed that lysine was not the first limiting amino acids. Threonine appeared as the first limiting amino acid only in the E treatment. Addition of encapsulated lysine and threonine then did not change nitrogen retention as'well as liveweight gain. Good agreement between bacterial colonies and urinary allantoin excretion was
also apparent in Trial 2. Regression of A on B was A = ( 2 . 9 . ~ ~ . ~ ~ ~ ) / ( 1 . 7 7 + ~ 0 . 5 5 0
) with an
R~
= 0.905 and Sb = 0.0663.Base on the trial 1 can be concluded that fermentation process in rumen can be manipulated by nutrient addition for microbial growth that is urea and isoacid. Calcium sulfate supplementation decreased the number of bacteria colonies by 15.12%. Isoacid supplementation was capable increasing the number of bacteria colonies by 11.1 I%, but it was less than
I
the number before calcium sulfate addition. The given ration should be able support the rumen microbial growth a s well as serve dietary protein passed from degradation that easily absorbed in post-rumen intestine. Thus, if the ration used in the trial 1 is aimed t o support the animal growth, the best treatment urea supplementation (ration B). Isoacid supplementation in fact halts the trial animal growth due t o poor growth of
rumen bacteria caused by poor ration quality. Poor quality of ration is represented by high content of urinary nitrogen and low number of N
retention.
Result of the trial 2 can be concluded that corn oil supple-mentation is useful t o reduce negative effect caused by calcium sulfate. The highest growth of the animal experiment was revealed by ration B (encapsulated corn oil supplementation). Lysine and threonine were not the first and second limiting amino acids. Threonine appeared a s the first limiting amino acid only in the E treatment. Addition of encapsulated Iysine and threonine then did not change nitropen retention as'well as liveweight gain.
...
RINGKASAN
Wardhana Suryapratama. Efek suplementasi asam lemak volatil bercabang dan kapsul lisin serta treonin terhadap nutrisi protein sapi Holstein (dibawah bimbingan Toha Sutardi selaku ketua, Djokowoerjo Sastradipradja, H. Suryahadi, Kartiarso, dan H. Mohammad Winugroho, masing-masing sebagai anggota).
Nutrisi protein pada hewan ruminansia sering dibatasi oleh adanya perkembangan mikroba rumen yang terganggu dan pasokan asam amino pada pasca rumen yang kurang mencukdpi. Dalam praktek pola pemberian pakan pada hewan ruminansia sering mengabaikan penyediaan nutrien yang c u k u p untuk sintesis biomassa mikroba rumen. Hal ini terutama akan mempengaruhi pencernaan pakan di dalam rumen dan secara umum akan mempengaruhi pasokan nutrien protein bagi induk semang.
Pasokan asam amino ke dalam usus pasca rumen hewan ruminansia terutama berasal dari protein mikroba yang disintesis di rumen, dan juga dari protein pakan yang tidak didegradasi di rumen. Hal ini memberikan pengertian bahwa pakan ruminansia selain harus mampu mendukung pertumbuhan mikroba rumen, juga harus mampu memasok kebutuhan asam amino bagi hewan ruminansia dan pakan tersebut mudah diserap diusus halus. Untuk i t u dilakukan suatu rangkaian percobaan yang ditujukan untuk memadukan teknologi suplementasi nutrien untuk optimasi bioproses d i dalam rumen dan teknologi pasokan asam amino esensial ke pasca rumen melalui kapsulasi yang tahan degradasi di rumen.
Percobaan pertama bertujuan unt'uk manipulasi bioproses di dalam rumen, menggunakan 5 ekor sapi jantan Holstein dengan bobot a y a l 348 h
bagian dedak gandum), dengan kandungan energi 10.5 MJ ME dan 15% protein kasar. Sebagai perlakuan adalah suplementasi urea, kalsium sulfat, isobutirat, P-metilbutirat, dan a-metilbutirat, sehingga terdapat 5 jenis pakan perlakuan, yaitu A = ransum kontrol, B = A+urea (139 mg/kg
w'.'~),
C
= B+kalsium sulfat (28 mglkgw ' . ~ ~ ) ,
D = C+isobutirat danP-
metilbutirat (masing-masing 0.05 mmol), dan E = D+a-metilbutirat (0.05 mmol). Periode pemeliharaan selama 30 hari, terdiri dari periode adaptasi ransum selama 2 0 hari, kemudian dilakukan pengamatan pertumbuhan selama 1 0 hari. Koleksi t o t a l dilakukan selama 5 hari terakhir dari periodepengamatan. i
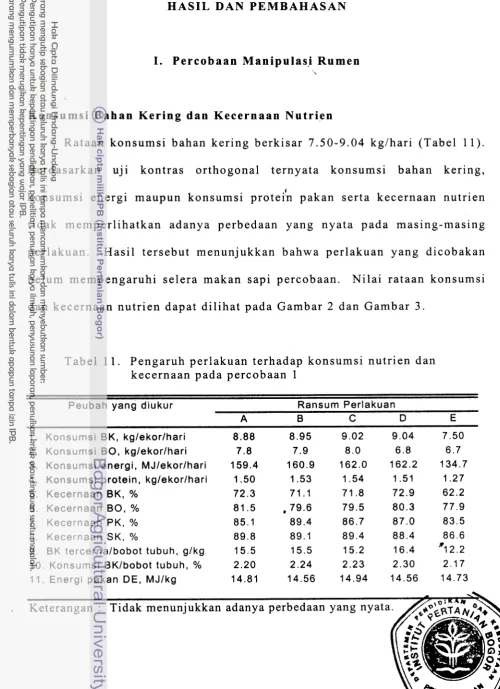
Hasil percobaan 1 menunjukkan bahwa konsumsi nutrien tidak memperlihatkan respon yang nyata pada masing-masing ransum perlakuan. Hasil tersebut mencerminkan bahwa seluruh suplementasi nutrien yang dilakukan belum sampai pada taraf yang mengganggu selera makan hewan percobaan. Demikian pula terhadap kecernaan nutrien secara keseluruhan belum terpengaruh oleh perlakuan yang diberikan. Secara umum ransum yang digunakan dapat dikatakan mampu menyediakan amonia cairan rumen dalam kadar yang cukup ideal untuk mendukung pertumbuhan mikroba rumen, yaitu sekitar 8 mM. Konsentrasi VFA total juga terdapat dalam ki- q saran yang ideal untuk pertumbuhan mikroba rumen, yaitu sekitar 110 mM.
Mulai perlakuan C (suplementasi kalsium sulfat) terdapat indikasi bahwa alantoin urin, populasi bakteri maupun pertumbuhan sapi percobaan memberikan respon yang menurun, ha1 ini diduga karena penambahan kalsium sulfat mengakibatkan kelebihan jumlah kalsium yang sebenarnya pada ransum dasar sudah melebihi kebutuhan. Akibat dari kelebihan kalsium dapat mempengaruhi ketidakseimbangan (imbalans) mineral dalam ransum. Ketidakseimbangan mineral tersebut dapat mempengaruhi aktifitas enzim-enzim mikroorganisme rumen, antara lain fosfohidrolase, fosfotransferase dan mempengaruhi reaksi-reaksi ATP, sehingga pertumbuhan bakteri terhambat, selanjutnya mempengaruhi kecernaan pakan yang dicerminkan dengan menurunnya konsentrasi VFA total. Suplementasi isoacid (perlakuan D dan E) nyata meningkatkan kembali populasi bakteri dan alantoin urin. Hal ini diduga karena asam lemak volatil bercabang merupakan sumber kerangka karbon untuk pembentukan asam amino bercabang bagi bakteri selulolitik. Pada perlakuan D dan E (suplementasi isoacid) pertambahan bobot tubuh nyata menurun (P<0.05). Terdapat d u a kemungkinan yang diduga menyebabkan penurunan tersebut, yaitu jumlah isoacid yang ditambahkan 0.05 mmol diduga kurang mencukupi kebutuhan mikroba rumen, selain itu diduga pula karena kualitas ransum dasar kurang baik, yang tercemin dari tingginya nitrogen urin dan menurunnya retensi N. Kenyataan tersebut menunjukkan bahwa protein ransum yang 1010s degradasi terlalu rendah, sehingga ruminansia kekurangan protein untuk di deposisi oleh tubuh.
Percobaan k e dua, bertujuan untuk manipulasi sistem absorpsi pasca rumen, menggunakan 5 ekor sapi betina Holstein dengan bobot awal 160
*
43 kg, dan rancangan Bujur Sangkar Latin 5x5. Ransum dasar yang digunakan terdiri dari rumput gajah yang tumbuh liar 45% (Pennzsetum
sehingga terdapat 5 jenis pakan perlakuan, yaitu A=ransum E percobaan 1
+
minyak jagung (1.5% BK), B=A+kapsul minyak jagung (1.5 mi), C=B+ kapsul lisin ( 2 8 mglkgwO."),
D=C+kapsul treonin (0.10 g), dan E=D+ minyak ikan (1.5% BK). Periode pemeliharaan selama 3 0 hari, terdiri dari periode adaptasi ransum selama 2 0 hari, kemudian dilakukan pengamatan pertumbuhan selama 1 0 hari. Koleksi t o t a l dilakukan selama 5 hari terakhir dari periode pengamatan. Kapsul yang digunakan adalah kapsul yang diperoleh dipasaran secara mudah kemudian kapsul diselaputi gelatin dan dicelupkan kedalam formaldehid 3 6 % kemudian dikeringkan dan baru diberikan pada hewan percobaan.I
Hasil percobaan 2 menunjukkan bahwa konsumsi nutrien tidak memperlihatkan respon yang nyata pada masing-masing ransum perlakuan. Hasil tersebut mencerminkan bahwa seluruh suplementasi nutrien yang dilakukan belum sampai pada taraf yang mengganggu selera makan hewan percobaan. Kecernaan bahan kering maupun protein nyata dipengaruhi oleh perlakuan yang diuji. Secara umum ransum yang digunakan dapat dikatakan mampu menyediakan amonia cairan rumen dalam kadar yang cukup ideal untuk mendukung pertumbuhan mikroba rumen, yaitu sekitar 8 mM. Konsentrasi VFA t o t a l juga terdapat dalam kisaran yang ideal untuk pertumbuhan mikroba rumen, yaitu sekitar 110 mM.
Perlakuan ransum B (suplementasi kapsul minyak jagung) cenderung meningkatkan produksi alantoin urin dimana A=1.42, B=1.47, C=1.45, D=1.40, dan E = 1 . 1 8 glhari, dan cenderung meningkatkan pertambahan bobot tubuh harian, yaitu A=0.820, B=0.860, C=0.725, D=0.760, dan E=0.640 kglhari. Hal tersebut karena dalam ransum B terdapat
I
suplementasi minyak jagung. Diduga dengan adanya penambahan minyak jagung akan terjadi reaksi penyabunan kalsium, sehingga pengafuh negatif kalsium dapat dikurangi, selanjutnya bakteri rumen dapat berkembang dan pertumbuhan meningkat.
Terdapat hubungan yang erat antara alantoin urin dengan jumlah koloni bakteri rumen seperti halnya pada percobaan 1, dengan formula A =
(2.9 ~ ~ . ~ ~ ~ ) / ( 1 . 7 7 + ~ ~ . ~ ~ ~ ) , R~ = 0.905 dan S b = 0.0663, dimana A adalah alantoin urin yang diproduksi dan B adalah jumlah populasi bakteri rumen.
Suplementasi kapsul lisin dan treonin belum memberikan respon yang nyata pada retensi N dan pertumbuhan. Hal ini menegaskan bahwa suplementasi kedua asam amino tersebut harus ada jaminan kecukupan metionin.
Berdasarkan percobaan 1 dapat disimpulkan bahwa proses fermentasi dalam rumen dapat dimanipulasi dengan menambah nutrien bagi pertumbuhan mikroba rumen, yaitu suplementasi urea dan isoacid. Suplementasi kalsium sulfat menurunkan jumlah koloni bakteri sebesar
i
15.12%. Suplementasi isoacid mampu meningkatkan jumlah koloni bakteri (1 1 . 1 1 % ) namun belum mampu menyamai jumlah koloni bakteri sebelum diberi kalsium sulfat. Ransum yang diberikan selain harus mampu mendukung perkembangan mikroorganisme rumen, juga mampu menyedia- kan protein ransum 1010s degradasi yang mudah diserap dalam saluran pencernaan pasca rumen. Oleh karena itu jika penggunaan ransum pada percobaan 1 ditujukan untuk pertumbuhan maka suplementasi yang terbaik adalah penambahan urea (ransum B). Penambahan isoacid justru menurunkan pertumbuhan sapi karena perkembangan bakteri rumen tidak didukung oleh pakan yang berkualitas baik. Rendahnya kualitas ransum yang diuji tercermin dari tingginya N urin dan rendahnya retensi N.
Hasil percobaan 2 dapat disimpulkan bahwa suplementasi minyak jagung dapat bermanfaat untuk mengurangi pengaruh negatif yang ditimbulkan oleh kalsium sulfat. Pertumbuhan sapi percobaan tertinggi dicapai oleh penggunaan ransum B (suplementasi kapsul minyak jagung),<
*
Rasio konsentrasi asam amino plasma darah sebelum dan sesudah makan menunjukkan bahwa lisin tidak merupakan asam amino pembatas yang utama. Adapun treonin diketahui merupakan asam amino pembatas yang utama pada ransum E. Suplementasi lisin dan treonin cenderung memberikan respon pada peningkatan retensi N dan penurunan jumlah N urin namun secara statistik belum memberikan respon yang nyata.
RIWAYAT HIDUP
Penulis dilahirkan di kota nan sejuk Salatiga Jawa Tengah pada tanggal 21 Juni 1954. Merupakan anak ke tiga dari empat bersaudara.
~ y a h bernama dr. Soegeng Wirjodipoero dan ibu bernama Sri Ernawati Suparini.
Lulus Sekolah Dasar Sokanegara I di Purwokerto pada tahun 1966, lulus SMP Negeri I11 di Purwokerto tahun 1969, dan lulus SMA Negeri I
Purwokerto tahun 1972. Tahun 1973 melanjutkan pendidikan ke Fakultas Peternakan Gadjah Mada Yogyakarta dan mendapat gelar Sarjana Peternakan tahun 198 1. Tahun 1983 penulis mulai mengabdikan diri
sebagai staf pengajar pada Fakultas Peternakan Universitas Jenderal
Soedirman Purwokerto, dan tahun 1985 mendapat kesempatan mengikuti pendidikan S2 di Fakultas Pascasarjana Universitas Gadjah Mada
Yogyakarta, lulus Magister Sains tahun 1989. Tahun 1993 diterima sebagai mahasiswa program S3 di Program Pascasarjana Institut Pertanian
Bogor pada program studi Ilmu Ternak dengan beasiswa dari Tim Manajemen Program Doktor.
Penulis dikaruniai dua orang anak bernama Latief Wikantadi dan Edi
I
Wibowo dari seorang istri bernama FM Suhartati.
I
UCAPAN TERIMA KASIH
Puja dan puji syukur penulis panjatkan kehadirat Allah s. w.t., karena berkat limpahan rakhmat dan kasih sayangNya penulis dapat menyelesaikan
laporan penelitian dalam bentuk disertasi yang berjudul "Efek Suplementasi Asam Lemak Volatil Bercabang dan Kapsul Lisin Serta
Treonin Terhadap Nutrisi Protein Sapi Holsteinn.
i
Penelitian beserta penulisannya dapat diselesaikan bukan karena
usaha penulis semata, namun karena adanya interaksi dan bantuan dari berbagai pihak. Oleh karena itu pada kesempatan ini penulis menghaturkan
terima kasih dan penghargaan yang setinggi-tingginya kepada yang terhormat Bapak Prof. Dr. Toha Sutardi, M.Sc. selaku ketua komisi
pembimbing, Bapak Prof Dr. Djokowoerjo Sastradipradja, Bapak Dr.Ir. H.
Suryahadi, DEA, Bapak Dr.Ir. Kartiarso, M.Sc. dan Bapak Dr. Ir. H.
Mohammad Winugroho, selaku anggota komisi pembimbing, yang dengan tulus ikhlas telah membimbing dan memberi petunjuk kepada penulis.
Terima kasih dan penghargaan yang sama penulis haturkan kepada yang terhormat Bapak Rektor IPB, Pimpinan Program Pascasarjana IPB dan Bapak Ketua Program Ilmu Terlfak yang telah berkenan menerima
Terima kasih dan penghargaan yang sama, penulis haturkan kepada
para dosen yang telah menularkan ilmunya kepada penulis, juga kepada para karyawan Program Pascasarjana IPB, yang telah banyak membantu
sehingga proses belajar mengajar dapat berjalan lancar.
Terima kasih dan penghargaan yang tinggi juga penulis haturkan
kepada yang terhormat Bapak Rektor UNSOED dan Bapak Dekan Fakultas Peternakan UNSOED yang telah berkenan memberi ijin kepada penulis
untuk mengikuti program S3 di IPB, demikian pula kepada istri dan anak- anak tercinta yang telah banyak berkorban dan memberikan dorongan
sehingga penulis dapat menyelesaikan pendidikan S3.
Hanya Allahlah yang memiliki kesempurnaan dan penulis menyadari
sepenuhnya bahwa tulisan ini masih jauh dari sempurna. Meskipun demikian penulis tetap berharap semoga disertasi ini dapat bermanfaat bagi perkembangan Ilmu Peternakan pada umumnya.
Akhir kata penulis berharap, semoga amal dan budi baik dari semua pihak yang telah diberikan kepada penulis mendapat balasan dan pahala yang berlimpah dari Allah s.w. t ., Amin.
Bogor, Januari 1999
KATA PENGANTAR
Salah satu peran bakteri rumen adalah sebagai pemasok asam amino
bagi hewan inang, yaitu ternak sapi, kerbau, kambing dan domba, akan tetapi peran tersebut sering diabaikan oleh peternak sehingga pola pembe-
rian pakan pada hewan ruminansia sering tidak mendukung pertumbuhan bakteri rumen. Bahkan a d a anggapan bahwa tanpa diberi pakan yang ber- kualitas baik hewan ruminansia masih dapat memberikan kontribusi yang
besar bagi kehidupan manusia. Namun bagi hewan ruminansia yang mempunyai potensi genetik yang tinggi memerlukan pakan dengan kualitas
yang unggul. Untuk itu penulis mencoba melakukan suatu rangkaian penelitian tentang suplementasi nutrien untuk mendukung pertumbuhan
bakteri rumen, maupun suplementasi nutrien yang dipasok langsung k e dalam saluran pencernaan pasca rumen.
Hasil yang menggelitik dari percobaan ini yaitu penggunaan asam
lemak volatil bercabang sebagai nutrien pendukung pertumbuhan bakteri rumen mempunyai potensi yang positif namun harus diimbangi dengan
ransum yang berkualitas tinggi, sehingga perlu perhatian yang saksama dalam penggunaannya.
e
Hasil penelitian ini diharapkan dapat memberikan informasi dan
I
suatu teknik memanipulasi pakan pada hewan ruminansia.
EFEK SUPLEMENTASI ASAM LEMAK VOLATkL
BERCABANG DAN KAPSUL LISIN SERTA TREONIN
TERHADAP NUTRISI PROTEIN SAP1 HOLSTEIN
WARDEANA SU~YAPRATAMA NRP. 93514
U a t v k memenuhi persyaratad memperoleh gelar Doktor dalam bidang firnu Ternak
PROGRAM
PA~CASARJANAIW$TITUT
PERTANIAN
BOGORJudul Penelitian : EFEK SUPLEMENTASI ASAM LEMAK VOLATIL BERCABANG DAN KAPSUL LISIN SERTA TREONIN TERHADAP NUTRISI PROTEIN SAP1 HOLSTEIN
Nama Mahasiswa : Wardhana Suryapratama
Nomor Pokok : PTK93514
Menyetujui
1. Komisi Pembimbing
,-
Prof-Dr. Toha Sutardi, M.Sc. i
Ketua
'-.
Prof.Dr. D. Sastradipradja Dr.Ir. H. Suryahadi, DEA
Anggota .
Dr.Ir. Kartiarso, M. Sc. Anggota
2. Ketua Program Studi Ilmu
Ternak
.Prof.Dr. Adi Sudono, M.Sc.
Dr.1r.H. M. Winugroho
Anggota -
rogram Pascasarjana
DAFTAR IS1
KATA PENGANTAR
DAFTAR TABEL
DAFTAR GAMBAR
PENDAHULUAN Latar Belakang Tujuan Penelitian Hipotesis Kegunaan Penelitian TINJAUAN PUSTAKA
Mikroorganisme Rumen Hewan Ruminansia Suplementasi Asam Amino
Suplementasi Asam Lemak Volatil Bercabang Sulfur Bagi Hewan Ruminansia
MATERI DAN METODE PENELITIAN
I. Percobaan Manipulasi Rumen (Sapi Jantan)
11. Percobaan Manipulasi Pasca Rumen (Sapi Betina)
Halaman
xvii
xxi
xxii
HASIL DAN PEMBAHASAN 40
I. Percobaan Manipulasi Rumen 40
Konsumsi Bahan Kering dan Kecernaan Nutrien 40
Parameter Metabolisme Rumen 4 3
Bakteri Rumen, Alantoin Urin, Retensi N, dan Pertumbuhan 50
XI. Percobaan Manipulasi Pasca Rumen 59
Konsumsi Bahan Kering dan Kecernaan Nutrien 59
Parameter Met abolisme Rumen 61
Bakteri Rumen, Alantoin Urin, Retensi N, dan Pertumbuhan 65
Diskusi Umum
KESIMPULAN
*
75DAFTAR TABEL
Nomor Judul Tabel Halaman
1. Komposisi asam amino bakteri rumen maupun isi rumen 12 2. Pengaruh suplementasi asam lemak volatil bercabang terha-
dap kecernaan dinding sel dan amonia (24 jam inkubasi) 14 Pengaruh suplementasi asam lemak volatil bercabang terha-
dap kecernaan dinding sel pada beberapa substrat tanaman
(24 jam inkubasi) 15
Pengaruh suplementasi asam lemak volatil bercabang terha- dap kecernaan bahan kering dan retensi N pada sapi laktasi
i
16 Ketersediaan relatif berbagai sumber sulfur diukur berdasar-
kan sintesis protein mikroba (in vztro)
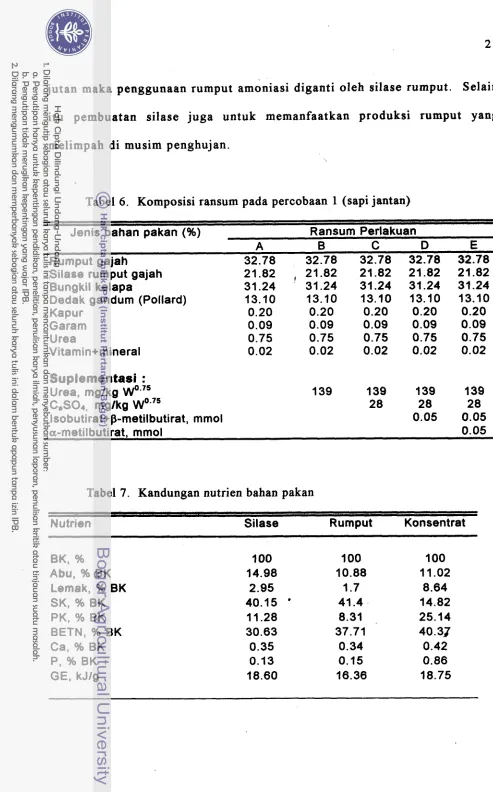
Komposisi ransum pada percobaan 1 (sapi jantan)
Kandungan nutrien bahan pakan
Denah hasil pengacakan pada percobaan 1
Komposisi ransum pada percobaan 2 (sapi betina) Denah hasil pengacakan pada percobaan 2
Pengaruh perlakuan terhadap konsumsi nutrien dan kecernaan
pada percobaan 1 40
Pengaruh perlakuan terhadap parameter metabolisme rumen
pada percobaan 1 44
Pengaruh perlakuan terhadap bakteri rumen, protozoa, retensi
N, dan pertumbuhan sapi pada percobaan 1 50 a
Pengaruh perlakuan terhadap konsumsi nutrien dan kecernaan
nutrien pada percobaan 2 59
I Pengaruh perlakuan terhadap parameter metabolisme rumen
pada percobaan 2 62
Pengaruh perlakuan terhadap bakteri rumen, protozoa, retensi
N, dan pertumbuhan sapi pada percobaan 2 66
DAFTAR GAMBAR
Nomor Judul Gambar Halaman
1. Suplai asam amino pada usus halus hewan ruminansia
(Egan, 1986) 11
2. Rataan konsumsi nutrien percobaan 1 41
3. Rataan kecernaan nutrien pada percobaan 1 42
4. Hubungan antara konsumsi BK dengan kecernaan pada
percobaan 1 42
5. Rataan VFA total pada percobaan 1 45
I
6. Hubungan antara bakteri rumen dengan VFA total pada
percobaan 1 46
7. Rataan konsentrasi amonia pada percobaan 1 47
8. Hubungan antara CS+isoacids dengan VFA, NH,, dan pH
pada percobaan 1 48
9. Hubungan antara bakteri dengan CS+isoacid 53
10. Respons bakteri dan alantoin urin pada percobaan 1 55
1 1. Hubungan antara bakteri dengan alantoin urin pada per-
cobaan 1 55
12. Respons retensi N dan pertumbuhan sapi pada percobaan 1 5 8
13. Hubungan antara retensi N dan pertumbuhan sapi pada per-
cobaan 1 58
14. Rataan konsumsi nutrien pada percobaan 2 60
15. Rataan kecernaan nutrien pada perdobaan 2 60
16. Rataan konsentrasi VFA total pada percobaan 2
*
6317. Hubungan antara bakteri dengan VFA total pada percobaan 2 64
Nomor Judul Gambar Halaman
19. Hubungan antara bakteri dengan alantoin pada percobaan 2 67
20. Respons koloni bakteri, ekskresi alantoin urin dan pertum-
PENDAHULUAN
Latar Belakang
Selain berperan pada proses pencernaan pakan secara fermentatif di rumen, bakteri rumen juga berperan dalam pasokan protein pada hewan inang. Besarnya pasokan protein biomassa mikroba rumen k e dalam usus
domba yang mendapat pakan dedak Lucerne dapat mencapai 10.3 g Nlhari dari masukan protein pakan 20.0 g Nlrhari, sedangkan pasokan protein
pakan yang 1010s degradasi di rumen dan masuk k e abomasum sebanyak 6.5 g Nlhari (Wallace dan Cotta, 1988). Begitu pula Demeyer (1991) melaporkan bahwa 50-80% nitrogen yang sampai k e usus halus dapat
berasal dari biomassa mikroba rumen. Hal ini mengakibatkan adanya
anggapan bahwa kualitas protein bagi hewan ruminansia tidaklah penting. Namun bagi ruminansia yang baru melahirkan, sapi perah masa awal lak- tasi maupun masa pertumbuhan yang pesat pada hewan muda, lebih banyak
membutuhkan nutrien dan hewan-hewan tersebut akan mempunyai kinerja yang lebih baik jika mendapat pasokan protein yang berkualitas tinggi dan
tahan degradasi di rumen. Hal ini menunjukkan bahwa komposisi asam amino dari protein pakan menjadi penting bagi hewan ruminansia seperti
1
juga pada hewan non-ruminansia. Asam amino metionin, lisin dan treonin
I
merupakan asam amino pembatas bagi ruminansia (Felix e t a l . , 1980). Selain ketiga asam amino tersebut, masih a d a asam amino lain yang sangat
aktivitas bakteri selulolitik (Czerkawski e t a l . , 1984; Russell dan Sniffen,
1984), kerangka karbon asam amino tersebut dapat berasal dari asam
lemak volatil bercabang (isoacids), yaitu asam isobutirat, P-metilbutirat,
dan a-metilbutirat. Hasil penelitian E r w a n t o (1995) menunjukkan bahwa
suplementasi asam amino bercabang bermanfaat untuk memacu perkem-
bangan mikroba rumen.
Dalam praktek, pola pemberian pakan pada sapi sering mengabaikan penyediaan semua nutrien untuk sintesis 'biomassa mikroba rumen. Keter-
batasan pasokan salah satu nutrien dapat menurunkan laju pertumbuhan mikroba. Laju sintesis protein mikroba rumen selain ditentukan oleh
ketersediaan energi (dalam bentuk ATP), juga ditentukan oleh ketersediaan amonia. Amonia merupakan produk utama katabolisme protein pakan dan
juga merupakan substrat utama untuk sintesis protein mikroba. Bryant dan Robinson (1962) melaporkan bahwa 8 0 % mikroba yang diisolasi dari rumen
dapat tumbuh baik dengan amonia sebagai sumber nitrogen. Hasil
I S
percobaan i n vitro dengan teknik perunutan i s o t o p N menunjukkan bahwa
8 0 % nitrogen pada sel mikroba dapat berasal dari substrat 1 5 ~ ~ 4 ' (Al-
Rabbat e t a l . , 1971). Hal ini menegaskan kembali bahwa amonia merupakan sumber utama nitrogen yang digunakan untuk pertumbuhan
*
biomassa mikroba rumen. Selain berasal dari katabolisme protein, amonia R
Pasokan asam amino k e dalam usus pasca rumen hewan ruminansia
selain berasal dari protein mikroba yang disintesis di rumen, juga dari protein pakan yang tidak didegradasi di rumen dan dari protein endogen (Stern et a l . , 1994). Hal ini memberikan pengertian bahwa pakan rumi-
nansia selain harus mampu memacu pertumbuhan mikroba juga harus
mampu memasok kebutuhan asam amino bagi hewan ruminansia dan pakan tersebut mudah diserap di usus halus. Untuk mencapai tujuan tersebut perlu adanya teknologi manipulasi pakran untuk optimasi bioproses di
rumen dan pasca rumen yang dilakukan secara t e p a t . Manipulasi pakan untuk optimasi bioproses di rumen dapat dilakukan dengan suplementasi nutrien dan kerangka karbon yang c o c o k dan tepat, sedangkan manipulasi
pasokan asam amino pasca rumen dapat menggunakan beberapa cara diantaranya perlakuan panas, pencampuran dengan tanin, reaksi dengan
asetaldehid, glutaraldehid, maupun formaldehid, esterifikasi g u g u s kar- boksil dari asam amino, pelapisan asam amino dengan lemak atau bahan
polimer yang resistan dari degradasi di dalam rumen (Arora, 1983; Leng, 1 9 9 1) s e r t a kapsulasi asam amino dengan gelatin yang diformaldehid (Sutardi, 1996: konsultasi pribadi). Teknologi kapsulasi menggunakan
gelatin yang mendapat perlakuan formaldehid bertujuan agar asam amino tidak dirombak di dalam rumen tetapi kapsul gelatin baru akan didegradasi
Tujuan Penelitian
Berdasarkan pemikiran di atas, penelitian yang dilakukan ditujukan
untuk memadukan teknologi suplementasi nutrien untuk optimasi bioproses di rumen dan pasokan asam amino esenssial k e pasca rumen, dengan
harapan sapi percobaan mempunyai pertumbuhan yang optimal.
Hipotesis
Suplementasi urea, kalsium sulfat dan asam lemak volatil bercabang mampu meningkatkan bioproses (cacahan bakteri rumen, kecernaan nutri-
en, dan produk fermentasi rumen) di dalam rumen dan pertumbuhan sapi. Suplementasi minyak jagung, kapsul minyak jagung, kapsul lisin dan
treonin, dan minyak ikan mampu meningkatkan cacahan bakteri rumen, produk fermentasi rumen, kecernaan nutrien, retensi nitrogen, dan pertumbuhan sapi.
Kegunaan Penelitian
Hasil penelitian diharapkan sebagai bahan informasi mengenai kebutuhan protein hewan ruminansia, perpaduan teknologi defaunasi untuk
peningkatan bioproses di dalam rumen dengan pasokan asam amino esensial
*
di u s u s pasca rumen. Selain itu berguna pula bagi kalangan industriawan f l
TINJAUAN PUSTAKA
Mikroorganisme Rumen Hewan Ruminansia
Rumen merupakan saluran pencernaan utama pada sapi, dan
mempertahankan suatu medium yang mendukung kepadatan dan variasi populasi mikroorganisme. Mikroorganisme melakukan proses fermentasi material pakan untuk memproduksi asam organik rantai pendek (short
c h a i n o r g a n i c a c i d s ) atau asam l e m a i volatil (Volatile F a t t y Acids = VFA), g a s metan (CHd), karbon dioksida (COz) dan proses tersebut
menyediakan substrat dan ATP untuk pertumbuhan mikroorganisme. Agen utama yang memecah serat, gula, pati dan protein di dalam rumen adalah bakteri, protozoa dan fungi (Leng, 199 1).
Bakteri rumen termasuk kelompok mikroba yang mempunyai
beberapa fungsi vital yang menguntungkan bagi hewan inang, yaitu : (1) Pakan serat dan material polimerik tanaman lainnya tidak dapat didegra- dasi oleh enzim hewan inang, akan tetapi difermentasi menjadi asam lemak
volatil (VFA), karbon dioksida dan g a s metan. VFA merupakan sumber energi utama bagi ternak ruminansia yaitu menyediakan 70-80% kebutuhan energi ternak (Robert, 1981; Maurice, 1987; France dan S i d d ~ n , 1993).
(2) Fermentasi berkaitan erat dengan pertumbuhan mikroba, dan prc)tein sel
mikroba yang terbentuk merupakan sumber protein yang utama bagi hewan inang. (3) Mikroba rumen juga membentuk beberapa vitamin tertentu yang
dapat dimanfaatkan oleh hewan inang. (4) Beberapa bakteri rumen dapat
yang terbaik adalah degradasi asam amino toksik seperti mimosin dan
derivatifnya yang berasal dari komponen tanaman legum L e u c a e n a (Stewart, 1991). Bakteri selulolitik termasuk bakteri anaerob, kebanyakan
membutuhkan NH3 sebagai sumber N, laju pertumbuhannya dipengaruhi pula oleh adanya asam lemak volatil bercabang ( b r a n c h e d c h a i n f a t t y
acids/isoacids) yaitu isobutirat P-metilbutirat dan a-metilbutirat ( a r s k o v ,
1982).
P r o t o z o a yang hidup di rumen, terutama dari jenis siliata dengan dua
familia, yaitu Isotrichidae (holotrik) dan Ophryoscolecidae (oligo-trik). P r o t o z o a rumen adalah anaerob yang memperoleh energi untuk tumbuhnya
melalui fermentasi karbohidrat. Bakteri rumen menyediakan sumber nitrogen, dan lemak untuk p r o t o z o a rumen.
Secara kuantitatif keberadaan p r o t o z o a siliata di dalam rumen penting untuk mencerna komponen karbohidrat, protein dan lipid pakan
yang di makan oleh ruminasia (Bird, 1991). Walaupun demikian, p r o t o z o a rumen nampaknya tidak sepenting bakteri rumen, ha1 tersebut karena
hewan yang bersangkutan dapat hidup dengan baik dan proses fermentasi t e t a p d a p a t berjalan dengan baik tanpa keberadaan p r o t o z o a . Meskipun populasi p r o t o z o a selalu lebih rendah* dari bakteri, tetapi protozoa
mempunyai ukuran yang lebih besar dari bakteri. Jika p r o t o z o a djkeluar- kan ataupun dihambat pertumbuhannya di dalam rumen, maka populasi
menelan dan mencerna bakteri rumen. Dengan demikian dapat dijelaskan
bahwa produksi protein mikroba akan lebih rendah pada hewan-hewan faunasi daripada hewan-hewan yang mengalami defaunasi ( a r s k o v , 1982).
Protozoa mempunyai pengaruh negatif di dalam rumen, terutama sekali bila ruminansia diberi pakan hijauan yang rendah kandungan proteinnya (Bird et a l . , 1990). Protozoa menelan dan mencerna bakteri
serta mengurangi biomassa bakteri di dalam rumen, sebagai akibatnya mengurangi suplai protein bagi hewan i n k g (Leng, 199 1).
Sintesis protein mikrobial meningkat dengan tanpa adanya protozoa.
Sintesis N mikrobial cairan rumen yang diukur secara in vitro (cairan rumen diambil dari kambing yang faunasi dan defaunasi) 15% lebih tinggi
pada inkubasi yang bebas protozoa dan sintesis protein mikrobial meningkat 33% dalam inkubasi yang bebas protozoa (cairan rumen diambil
dari domba yang faunasi dan defaunasi). Studi perbandingan antara domba yang faunasi dan defaunasi menunjukkan bahwa suplai protein mikrobial
post rumen 20% lebih tinggi pada hewan yang di defaunasi (Bird, 1991). Hal tersebut berarti, apabila di dalam rumen tidak terdapat protozoa, maka
lebih banyak protein mikrobial yang tersedia untuk hewan inang, dan diharapkan mampu meningkatkan produktivitas ruminansia.
Hasil penelitian in vitro (Satter dan Slyter, 1974) menggunakan cairan rumen sapi, pada fermentasi kultur kontinyu menunjukkan bahwa produksi protein mikroba rumen mencapai laju yang maksimum pada
konsentrasi amonia di a t a s nilai tersebut tidak mempunyai pengaruh yang berarti terhadap protein mikroba rumen.
Jika hewan ruminansia mendapat pakan yang mengandung serat
relatif tinggi seperti bahan pakan hijauan pada umumnya, maka teknologi defaunasi untuk mengendalikan populasi p r o t o z o a perlu dilakukan. Hal ini
telah terbukti dari beberapa penelitian yang menunjukkan bahwa perlakuan defaunasi dapat meningkatkan pertumbuhan bakteri selulolitik (Jalaludin,
I
1994; Erwanto, 1995). Minyak jagung dapat digunakan sebagai agensia defaunasi. Perlakuan defaunasi akan lebih efektif jika diikuti dengan pasokan nutrien untuk mendukung perkembangan mikroorganisme rumen
Suplementasi Asam Amino
Laju pertumbuhan dan produksi mikroorganisme rumen sangat menentukan penampilan hewan ruminansia. Mikroorganisme rumen meru-
pakan sumber protein yang penting untuk hewan inang dan juga mempe- ngaruhi laju fermentasi di dalam rumen. Laju pertumbuhan mikroorga-
nisme rumen yang rendah, dapat menghambat laju fermentasi pakan di dalam rumen dan selanjutnya dapat mengurangi konsumsi ransum bagi
hewan inang yang akhirnya masukan energi juga akan rendah. Pengaruh rendahnya laju pertumbuhan mikroorganisme rumen terhadap k e t e ~ s e d i a a n asam amino bagi hewan inang dan terhadap kosumsi ransum sangat nyata
itu laju pertumbuhan mikroorganisme rumen perlu dimaksimumkan, diantaranya dengan cara suplementasi asam amino. Wilkerson et a l . (1993) melaporkan bahwa metionin adalah asam amino pembatas utama. Dengan
demikian kebutuhan asam amino tersebut bagi pertumbuhan mikroorga- nisme rumen harus tercukupi.
P a d a umumnya pakan hijauan kekurangan asam amino metionin, lisin, valin, isoleusin dan leusin. Kekurangan metionin dapat dipasok melalui
t
analognya ataupun melalui amonium sulfat (Erwanto, 1995), sedangkan untuk memenuhi kebutuhan asam amino lisin perlu dicari bentuk pasokan
yang t e p a t . Hal tersebut karena tubuh hewan maupun mikroorganisme rumen tidak mempunyai sistem transaminasi bagi lisin. Lisin dapat dipasok
dalam bentuk kapsul lisin, kapsul terbuat dari bahan gelatin yang tahan degradasi di dalam rumen, yaitu kapsul dilapisi gelatin yang diformaldehid dahulu sebelum diberikan pada hewan, dengan harapan suplementasi lisin
dapat lebih dimanfaatkan oleh hewan inang melalui proses pencernaan pasca rumen.
Kekurangan asam amino rantai cabang (valin, leusin dan isoleusin)
dapat dipasok melalui kerangka karbonnya. Kerangka karbon yang sesuai
adalah kerangka karbon dari asam lemak volatil bercabang yaitu isobutirat,
P-metilbutirat dan a-metilbutirat. Jika pasokan asam lemak volatil I
bercabang tersebut masuk k e dalam rumen maka diharapkan dapat dimanfaatkan untuk pertumbuhan bakteri. Selanjutnya sapi percobaan
Pasokan terhadap asam amino treonin pada hewan ruminansia juga
perlu diperhatikan karena asam amino tersebut selain tidak mempunyai sistem transaminasi di dalam sel hewan, juga di dalam rumen akan menga-
lami reduksi. Pada dasarnya rumen merupakan organ pereduksi karena mempunyai sistem redoks potensial yang negatif sehingga cenderung mere-
duksi senyawa-senyawa, akibatnya treonin akan kehilangan gugus O H dan pasokan treonin ke dalam rumen akan hilang dan tidak dapat dimanfaatkan
i
oleh hewan inang (Sutardi, 1980). Untuk itu perlu diatasi dengan teknologi kapsulasi menggunakan gelatin yang diformaldehid agar tahan
degradasi di dalam rumen.
Suplai asam amino pada usus hewan ruminansia dapat berasal dari
tiga sumber yaitu pertama berasal dari protein pakan yang 1010s dari
degradasi di dalam rumen, kedua berasal dari sel-sel mikroorganisme yang keluar dari retikulo-rumen (sering disebut mikroba rumen) dan ketiga berasal dari protein endogenus (Richardson dan Hatfield, 1978; Merchen et
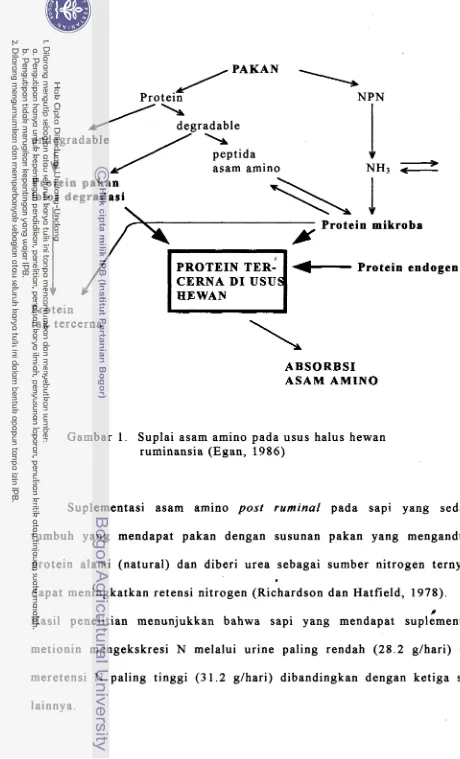
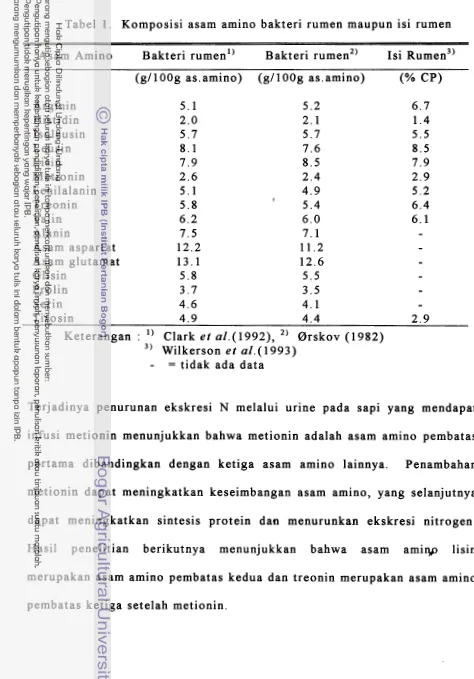
al., 1986; Egan, 1986; Wilkerson et al., 1993) (Gambar 1). Komposisi asam amino mikroorganisme rumen dapat dilihat pada Tabel 1.
Untuk mendapatkan penampilan hewan yang lebih baik, maka jumlah
asam amino yang sampai ke usus halus eharus lebih banyak. Penggunaan
protein yang tahan terhadap degradasi ruminal merupakan strategi untuk meningkatkan suplai protein post ruminal bagi hewan inang (Merchen et
/
pAKANProtein
1
NPN/
h e g r a d a b l e undegradableJ
peptidaasam amino
/I
I
-
Protein pakan
1010s degradasi
\
i"'-
Y
Protein mikrobaProtein endogen
Protein t a k tercerna
CERNA DI USU HEWAN
ABSORBS1 ASAM AMINO
Gambar 1. Suplai asam amino pada usus halus hewan ruminansia (Egan, 1986)
Suplementasi asam amino post ruminal pada sapi yang sedang
tumbuh yang mendapat pakan dengan susunan pakan yang mengandung protein alami (natural) dan diberi urea sebagai sumber nitrogen ternyata
*
dapat meningkatkan retensi nitrogen (Richardson dan Hatfield, 1978). f l
Hasil penelitian menunjukkan bahwa sapi yang mendapat suplementasi
[image:139.566.43.503.22.781.2]Tabel 1. Komposisi asam amino bakteri rumen maupun isi rumen
--
Asam Amino Bakteri rumen1' Bakteri rumen2) Isi ~ ~ m e n ~ '
(g1100g as.amino) (gI100g as.amino) (% CP)
Arginin 5 . 1 5 . 2 6 . 7
Histidin 2 . 0 2 . 1 1.4
Isoleusin 5 . 7 5.7 5.5
Leusin 8 . 1 7 . 6 8.5
Lisin 7 . 9 8.5 7 . 9
Metionin 2 . 6 2 . 4 2 . 9
Fenilalanin 5 . 1 4 . 9 5 . 2
I
Treonin 5.8 5.4 6.4
Valin 6 . 2 6 . 0 6 . 1
Alanin 7.5 7 . 1
-
Asam aspartat 12.2 11.2
-
Asam glutamat 13.1 12.6
-
Glisin 5 . 8 5.5
-
Prolin 3 . 7 3.5
-
Serin 4 . 6 4 . 1
-
Tirosin 4 . 9 4 . 4 2 . 9
Keterangan : " Clark et a1.(1992), 2' Orskov (1982) 3' Wilkerson e t aZ.(1993)
-
= tidak a d a d a t aTerjadinya penurunan ekskresi N melalui urine pada sapi yang mendapat infusi metionin menunjukkan bahwa metionin adalah asam amino pembatas
pertama dibandingkan dengan ketiga asam amino lainnya. Penambahan metionin dapat meningkatkan keseimbangan asam amino, yang selanjutnya
dapat meningkatkan sintesis protein dan menurunkan ekskresi nitrogen. Hasil penelitian berikutnya menunjukkan bahwa asam amiqp lisin
[image:140.561.41.515.81.760.2]Suplementasi Asam Lemak Volatil Bercabang
Keberadaan mikroorganisme di dalam rumen selain berperan dalam proses pencernaan pakan secara fermentatif juga berperan sebagai pemasok
sumber protein bagi hewan inang. Untuk dapat berkembang dan melakukan
pencernaan fermentatif dengan baik, mikroorganisme rumen membutuhkan pasokan nutrien yang cukup dan tidak berlebihan. Satu diantara nutrien yang diperlukan untuk perkembangan mikroorganisme rumen adalah asam
I
lemak volatil bercabang (isoacids) (Felix e t a l . , 1980; Gorosito e t a l . ,
1985; Johnson e t a l . , 1994). Adanya pasokan nutrien tersebut secara kontinyu dan pembuangan produk fermentasi yang lancar maka diperoleh
tingkat konsentrasi amonia (NH3) dan volatile fatty a c i d s (VFA) yang tidak berlebihan, sehingga dapat diharapkan perkembangan mikroorganisme
rumen mencapai maksimal, dan tentunya produktivitas hewan inang akan maksimal pula.
Asam lemak volatil bercabang (branched-chain fatty acids = BCFA)
yang terdiri dari isobutirat (i-C4), P-metilbutirat (i-Cs) dan a-metilbutirat
(2Me-C4) merupakan prekursor untuk sintesis protein mikroorganisme rumen (Russell dan Sniffen, 1984). Rantai karbon dari asam lemak volatil
bercabang tersebut digunakan oleh baktkri selulolitik untuk sintesis asam
amino esensial valin, leusin dan isoleusin. Selain untuk sintesis asam amino tersebut, isoacid1BCFA (dan asam lemak n-valerat) juga digunakan oleh bakteri selulolitik untuk sintesis asam lemak rantai panjang dan
sebagai prekursor dalam sintesis asam amino prolin (Amos e t a l . , 1971).
Asam lemak volatil berantai cabang tersebut disintesis dari protein dan sumber karbon lain selama proses fermentasi di dalam rumen (Czerkawski
e t a l . , 1984). Adanya daur ulang protein bakteri di dalam rumen juga meningkatkan asam lemak volatil bercabang (Klusmeyer e t a l . , 1987).
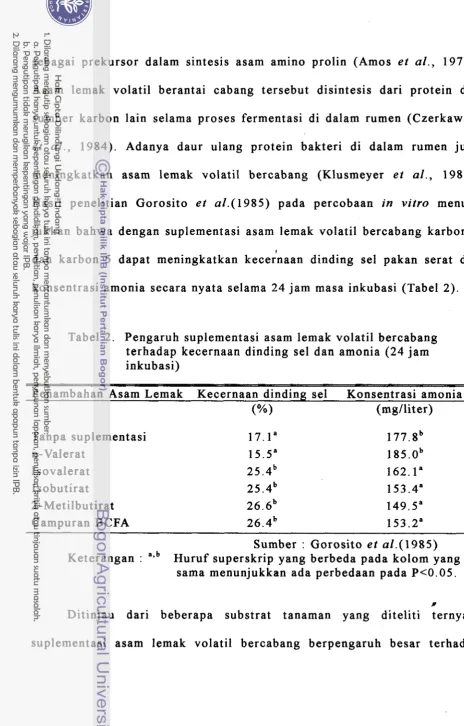
Hasil penelitian Gorosito e t a1.(1985) pada percobaan in vitro menun- jukkan bahwa dengan suplementasi asam lemak volatil bercabang karbon-4
i
dan karbon-5 dapat meningkatkan kecernaan dinding sel pakan serat dan
konsentrasi amonia secara nyata selama 24 jam masa inkubasi (Tabel 2 ) .
Tabel 2. Pengaruh suplementasi asam lemak volatil bercabang terhadap kecernaan dinding sel dan amonia (24 jam inkubasi)
Penambahan Asam Lemak Kecernaan dinding sel Konsentrasi a m o i a (mg/liter) Tanpa suplementasi
n-Valerat Isovalerat Isobutirat 2-Metilbutirat Campuran BCFA
Sumber : Gorosito e t a1.(1985) Keterangan : ~ u r u f superskrip yang berbeda pada kolom yang
sama menunjukkan ada perbedaan pada P c 0 . 0 5 .
I
Ditinjau dari beberapa substrat tanaman yang diteliti ternyata
[image:142.566.42.507.29.756.2]k e c e r n a a n d i n d i n g s e l s u b s t r a t y a n g t i d a k m e n g a n d u n g a s a m a m i n o , s e p e r t i
p a d a k e r t a s s a r i n g m a u p u n j e r a m i g a n d u m ( T a b e l 3 ) .
T a b e l 3 . P e n g a r u h s u p l e m e n t a s i a s a m l e m a k v o l a t i l b e r c a b a n g t e r h a d a p k e c e r n a a n d i n d i n g s e l p a d a b e b e r a p a s u b s t r a t t a n a m a n ( 2 4 j a m i n k u b a s i )
S u b s t r a t t a n a m a n N o n - s u p l e m e n t a s i C - 4 d a n C - 5 a s a m S i g n i f i k a n s i - l e m a k b e r c a b a n g '
(%I
(%)I
A l f a l f a h a y 3 4 . 1 3 8 . 7
*
T i m o t h y h a y 4 8 . 5 5 2 . 6 N S R u m p u t R e e d C a n a r y 3 2 . 1 3 7 . 8 N S R u m p u t B e r m u d a 1 8 . 2 2 1 . 8 NS R u m p u t O r c h a r d 3 0 . 2 3 8 . 0*
S i l a s e J a g u n g 3 4 . 1 3 6 . 7*
J e r a m i G a n d u m 1 0 . 4 1 3 . 0
* *
K e r t a s s a r i n g 1 . 3 1 1 . 9* *
S u m b e r : G o r o s i t o et n1.(1985) K e t e r a n g a n : ' P e n a m b a h a n a s a m l e m a k 1 . 7 6 m M
*
i n e n u n j u k k a n p e r b e d a a n p a d a P c 0 . 0 5* *
m e n u n j u k k a n p e r b e d a a n p a d a P < 0 . 0 1 N S m e n u n j u k k a n t i d a k a d a b e d a .B e r d a s a r k a n d a t a p a d a T a b e l 3 m e m b e r i k a n p e n g e r t i a n b a h w a
k e b e r h a s i l a n m e m a c u p e n c e r n a a n f r a k s i s e r a t p a k a n m e l a l u i s u p l e m e n t a s i
a s a m l e m a k t e r s e b u t s a n g a t p e n t i n g , k a r e n a k e l o m p o k b a k t e r i s e l u l o l i t i k
s a n g a t d i u n t u n g k a n o l e h s u p l e m e n t a s i t e r s e b u t
P a d a k e d u a t a b e l t a d i j u g a n a m p a k b a h w a s u p l e m e n t a s i a s a m lemak
I
v o l a t i l r a n t a i k a r b o n l u r u s ( n - v a l e r a t ) t i d a k m e m b e r i k a n p e n g a r u h y a n g
[image:143.561.39.518.7.775.2]p e n g e r t i a n b a h w a n-valerat tidak menjadi faktor p e m b a t a s pertumbuhan dan p e r k e m b a n g a n m i k r o o r g a n i s m e rumen.
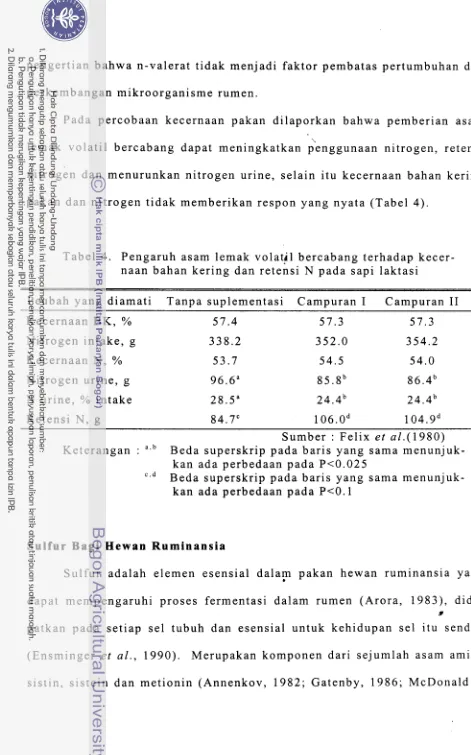
P a d a p e r c o b a a n kecernaan pakan dilaporkan b a h w a pemberian asam lemak v o l a t i l b e r c a b a n g d a p a t meningkatkan pknggunaan nitrogen, retensi nitrogen dan m e n u r u n k a n nitrogen urine, selain itu kecernaan bahan kering pakan dan n i t r o g e n t i d a k memberikan respon yang nyata ( T a b e l 4 ) .
T a b e l 4 . P e n g a r u h asam lemak volatil bercabang t e r h a d a p kecer- naan b a h a n kering dan retensi N pada s a p i laktasi
P e u b a h y a n g d i a m a t i T a n p a suplementasi Campuran I Campuran I1 Kecernaan B K , % 5 7 . 4 5 7 . 3 5 7 . 3 Nitrogen i n t a k e , g 3 3 8 . 2 3 5 2 . 0 3 5 4 . 2
Kecernaan N, % 5 3 . 7 5 4 . 5 5 4.0
Nitrogen u r i n e , g 96.6" 8 5 . 8 b 86.4b N urine, % i n t a k e 28.5" 2 4 . 4 b 2 4 . 4 b R e t e n s i N , g 84.7" 1 0 6 . 0 d 1 0 4 . 9 ~
S u m b e r : Felix et a1.(1980) K e t e r a n g a n : "." B e d a superskrip pada b a r i s yang s a m a menunjuk-
kan ada perbedaan pada P < 0 . 0 2 5
c * d B e d a s u p e r s k r i p p a d a b a r i s yang s a m a menunjuk-
kan ada perbedaan pada P c 0 . 1
S u l f u r B a g i H e w a n R u m i n a n s i a
S u l f u r a d a l a h e l e m e n esensial dalam pakan hewan ruminansia yang dapat m e m p e n g a r u h i p r o s e s fermentasi dalam rumen ( A r o r a , 1983), dida-
I
[image:144.568.47.518.14.769.2]a l . , 1 9 8 8 ; E n s m i n g e r et a l . , 1 9 9 0 ) , dan merupakan b a g i a n y a n g penting dari bakteri r u m e n ( H u n g a t e , 1966). J u g a sebagai k o m p o n e n d u a vitamin yaitu t i a m i n d a n b i o t i n d a n s e b a g a i k o m p o n e n rambut, wol dan bulu, terdapat d a l a m s a l i v a , e m p e d u , hormon i n s u l i n ( ~ n s m i n ' g e r e t a l . , 1 9 9 0 ) dan dalam koenzirn A ( R i i s , 1 9 8 3 ) .
H a n y a s e d i k i t s u l f u r dalam tubuh berada dalam b e n t u k anorganik m e s k i p u n d i k e t a h u i ada sedikit sulfat dalam d a r a h (McDonald et a l . , 1988). K a d a r s u l f u r d a l a m biomassa m i k r o b a rumen d a p a t mencapai 8glkg bahan k e r i n g dan s e b a g i a n besar terdapat dalam protein ( B i r d , 1 9 7 2 ) . Sekitar 0.15 p e r s e n d a r i bobot badan d a n 1 0 persen dari k a n d u n g a n mineral tubuh adalah s u l f u r ( E n s m i n g e r et a l . , 1990).
D e n g a n a d a n y a m e t a b o l i s m e s u l f u r oleh mikroba dalam rumen maka r u m i n a n s i a d a p a t m e n g g u n a k a n s u l f u r dalam bentuk organik dan anorganik. S u l f u r d i b u t u h k a n untuk m e n s i n t e s i s asam amino yang m e n g a n d u n g sulfur dari m i k r o b a r u m e n (Goodrich dan Garrett, 1 9 8 6 ) . J u m l a h s u l f u r yang d i b u t u h k a n o l e h m i k r o o r g a n i s m e rumen tergantung k e p a d a laju metabo- l i s m e p r o t e i n d a n b e r b a n d i n g l u r u s d e n g a n k e b u t u h a n nitrogennya. A g r i c u l t u r e R e s e a r c h C o u n c i l ( 1 9 8 0 ) menyarankan b a h w a untuk setiap gram k e b u t u h a n n i t r o g e n d i b u t u h k a n 0 . 0 7 gram s u l f u r yang dapat d i h i - d r o l i s a d a l a m r u m e n . Secara t r a d i s i o n a l e k e c u k u p a n s u l f u r dapat disuplai dalam b e n t u k p r o t e i n tetapi dengan m e n i n g k a t n y a penggunaan senyawa N
Bahan pakan yang kaya akan sulfur antara lain alfalfa, tepung darah,
tepung biji kapas, tepung bulu, tepung ikan dan limbah hasil laut, tepung
biji rami, tepung daging, hasil sampingan ternak unggas, dan tepung
kedelai (Ensminger e t a l . , 1990). Molases juga mengandung cukup banyak
sulfur (0.3 0 persen). Sebagai sumber suplemen biasanya digunakan elemen
sulfur, ragi, bermacam-macam garam sulfat (Preston dan Leng, 1987),
antara lain amonium sulfat, natrium sulfat dan kalsium sulfat. Metionin
1
dan hidroksi analognya merupakan sumber sulfur yang baik, sedangkan
sulfur elemental memiliki efisiensi pemanfaatan yang rendah (Kahlon et a l . ,
1975). Nilai ketersediaan relatif berbagai senyawa sumber sulfur
dibandingkan dengan asam amino L-metionin dapat dilihat pada Tabel 5.
Tabel 5 . Ketersediaan relatif berbagai sumber sulfur diukur berdasarkan sintesis protein mikroba
(in
vitro)Sumber Sulfur Ketersediaan (%)
L-metionin Kalsium sulfat Amonium sulfat DL-metionin
Natrium
sulfatNatrium
sulfida
Sulfur elemental
Analog hidroksi metionin (AHM)
Sumber : Kahlon e t a l . ( l P 7 5 )
Tabel 5 tersebut menunjukkan bahwa sulfur anorganik dalam bentuk
[image:146.576.47.534.17.789.2] [image:146.576.40.535.38.773.2]ketersediaan relatif tinggi, hampir mendekati L-metionin dan bahkan
melebihi DL-metionin. Sulfur dalam bentuk elemen mempunyai nilai
ketersediaan relatif rendah, ha1 tersebut mungkin disebabkan oleh
kelarutannya yang rendah dalam cairan rumen (Hungate, 1966).
Oleh karena sulfur berfungsi dalam sintesis asam amino yang
mengandung sulfur dan beberapa vitamin B selama pencernaan di dalam
rumen, maka mikroorganisme rumen yang kekurangan sulfur tidak dapat
1
berfungsi secara normal. Penambahan sulfur dalam kondisi demikian dapat
meningkatka