www.elsevier.com / locate / livprodsci
Effects of an additional iron injection on growth and humoral
immunity of weanling pigs
a ,
*
a b bE.M.A.M. Bruininx
, J.W.G.M. Swinkels , H.K. Parmentier , C.W.J. Jetten ,
c b
J.L. Gentry , J.W. Schrama
a
Research Institute for Pig Husbandry, P.O. Box 83, 5240 AB Rosmalen, The Netherlands
b
Department of Animal Husbandry, Wageningen Institute of Animal Science, Wageningen Agricultural University, 6700 AH Wageningen,
The Netherlands
c
Department of Animal Sciences, University of Kentucky, Lexington, KY 40546-0215, USA Received 21 July 1999; received in revised form 1 February 2000; accepted 14 March 2000
Abstract
In a 5-week study, 120 crossbred weaning pigs (age 4 weeks) were used to examine the effects of an additional Fe-injection and immunization moment on the growth and humoral immune response. Pigs were allotted to one of eight treatments based on body weight and litter origin. Factors included: (1) single intramuscular injection (day 3 after birth) vs. double intramuscular injection (days 3 and 21 after birth) of 200 mg Fe; (2) antigen challenge (Keyhole limpet hemocyanin (KLH) and ovalbumin (OA) vs. placebo); and (3) immunization moment (day 21 vs. 1 after weaning). All pigs had free
21
access to prestarter diets supplemented with 80 mg kg Fe as FeSO4?7H O. Pigs were housed in a room consisting of 122
pens (ten pigs per pen). During the experiment, blood hemoglobin (Hb) levels and body weight were determined weekly and total Ig, IgG and IgM titers to KLH and OA twice a week. At day 6 after weaning, the blood Hb levels of double injected pigs were higher (P,0.01) than the blood Hb-levels of the pigs that had received a single Fe-injection. Prior to day 6 and thereafter, blood Hb levels were similar both for single and double injected groups. Throughout the experiment, the additional Fe-injection, and immunization moment had no effect (P.0.1) on growth and total Ig response to KLH and OA. But the genotype of the pigs affected (P,0.05) the kinetics of the total Ig response to KLH and OA. In conclusion, an additional injection of Fe one week prior to weaning does not affect body growth, and does not enhance or support the humoral immunity to a T-cell dependent antigen. Furthermore, the suppressing effect of weaning on humoral immunity was not affected by additional Fe. 2000 Elsevier Science B.V. All rights reserved.
Keywords: Weaning; Iron; Growth; Immune response; Pigs
1. Introduction
*Corresponding author. Tel.:131-73-528-6555; fax:
131-73-The first 2 weeks after weaning are a critical
521-8214.
period in the nursery phase of pigs. This period is
E-mail address: [email protected] (E.M.A.M.
Bruininx). often associated with depressed feed intake and
growth as well as health problems. Social, dietary minal boars (GY-s). Based on this rotational breed-and immunological stressors resulting from the pro- ing system, pigs were divided into three groups that cess of weaning together with the occurrence of differed in major genotype: DL, FL and GY-z (Table pathogens (e.g. specific Escherichia coli strains) are 1). The sow line that contributed 28.1% to the considered as main risk factors (Bourne, 1986; genetic composition of the pig was considered the Nabuurs, 1991; Van Beers-Schreurs et al., 1992; major genotype. All pigs received an intramuscular
Makkink, 1993). Fe injection (200 mg Fe, Fe dextran; AUV Cuijk) on
Iron (Fe) is a nutrient related to health and day 3 of age. Pigs had no access to supplemental Fe immunity (Brock, 1994). Administration of iron (200 (i.e. creep feeding, soil) during the suckling period. mg Fe as iron dextran per piglet) on day 2 or 3 after
birth to prevent anemia is a common practice in 2.2. Experimental design modern pig husbandry. Kamphues et al. (1992)
showed that an additional Fe-injection on day 21 The experiment was conducted following a 23
after birth resulted in a higher growth post weaning. 232 factorial arrangement of treatments. The first Furthermore, they suggested that the improved factor was an additional intramuscular injection with
growth was associated with a lower incidence of 200 mg Fe (FE; 2 ml Fe-dextran; AUV Cuijk) on
diarrhoea. Recently, it was shown that a minor day 21 of age or 2 ml of a placebo (SALINE; PBS difference in Fe status has an effect on the metabo- solution). The second factor was an intramuscular lism (Gentry et al., 1997) and immunity (Schrama et injection with 2 ml of an antigen cocktail (IMMU; 1
al., 1997) of weaning pigs. The role of iron in mg Keyhole limpet hemocyanin (KLH)), 4 mg
maintaining health is, apart from anemia prevention, ovalbumin (OA), and 15 mg Lf tetanus toxoid (TT) still not completely clear. Hershko (1993) showed an in a 1:1 v / v Specoll:PBS solution (ID-Lelystad, optimal immune responsiveness at physiological Lelystad, The Netherlands) or 2 ml of a placebo normal levels of iron in human blood. An increase as (PLAC; 1:1 v / v Specoll:PBS solution). The third well as a decrease in blood iron levels had negative factor was immunization 1 day before weaning (D2
effects on immune responsiveness (De Sousa et al., 1) or 1 day after weaning (D1). The antigen cocktail 1991; Brock, 1994). Additionally, it was shown was used to elicit a T-cell-dependent humoral
im-(Gwazdauskas et al., 1978; Haye and Kornegay, mune challenge.
1979; Blecha and Kelley, 1981) that weaning re- The experimental design resulted in eight treat-sulted in reduced immunoglobulin production, proba- ment groups. The four control groups that received a bly due to the stress associated with weaning. placebo injection (PLAC) consisted each of ten pigs, In this study, the effects of an additional iron whereas, the four experimental treatments consisted injection were examined on growth and humoral each of 20 pigs. Pigs were assigned according to immune response. A secondary aim was to examine weight, genotype, and blood Hb status to one of the whether the humoral immune responsiveness was eight treatment groups at the end of the third week in
affected by weaning. the suckling period.
Table 1
For this experiment, 120 weanling barrows and
Finnish Landrace 14.1 7.8 28.1 50
gilts of approximately 4 weeks of age with a mean Dutch Landrace 7.8 28.1 14.1 50
BW of 7.6 kg (S.E.M.50.11) were used. Pigs Great Yorkshire sow-line 28.1 14.1 7.8 50
originated from four Dutch Landrace (DL), five a
GY-z, percentage of Great Yorkshire sow-line; DL, Dutch
Finnish Landrace (FL), and six Great Yorkshire Landrace; FL, Finnish Landrace; GY-s, Great Yorkshire
2.3. Housing and feeding dilutions of serum were applied to antigen-coated wells (1mg KLH, 1mg OA, or 0.2mg TT per ml) of
After weaning pigs were housed in 12 pens a microtiter plate and incubated for 1 h at room
(2.6531.25 m; ten pigs per pen). All pens accom- temperature. Subsequently 1:8000 diluted horseradis-modated pigs of all treatment groups. The pens had h-peroxidase (PO) conjugated rabbit anti-swine H1
partially solid, electrically heated, floors and tri-bar L (R?Sw-IgG (H1L) / PO; Cappel, Gaithersburg, metal slats. The room was equipped with a com- MD) for total immunoglobulin (Ig), 1:8000 diluted puter-controlled heating and mechanical ventilation rabbit anti-IgG Fc (R?Sw-IgG (Fc) / PO; Nordic, system. The first 2 weeks after weaning a room Tilburg, The Netherlands) for IgG, or 1:8000 diluted temperature of 278C was maintained. Afterwards, goat anti-swine IgM Fc (GaSW-IgM(Fc) / PO; Nor-room temperature was gradually decreased to 208C dic) for IgM was added during 1 h at room tempera-(1.58C per week). The first two weeks of the ture. After washing, tetramethylbenzidine (TMB; experimental period floor temperature was set at Sigma, St. Louis, MO) was added as a substrate. 328C. Thereafter, floor heating was turned off. Color formation was stopped with 2.5 N sulfuric acid Pigs had free access to water and pelleted com- after 10 min. All absorbances were expressed rela-mercial diets using nipple drinkers and dry feeders, tive to the absorbance of a standard positive control respectively. The first two weeks after weaning pigs serum. The titer of sera against KLH, TT, or OA in a
21
were fed a prestarter diet (14.3 MJ ME kg and sample was that dilution that showed an extinction 1.03% apparent ileal digestible lysine). Thereafter, closest to 50% of EMAX, where EMAX represents the
21
all pigs were fed a starter diet (13.6 MJ ME kg highest mean extinction of a standard positive and 0.93% apparent ileal digestible lysine). Both (pooled) serum (Frankena, 1987).
diets were supplemented with 80 mg Fe as FeSO4?
7H O per kg feed. All nutrients met or exceeded2 2.5. Statistical analyses
NRC (1998) recommended levels.
All data were analyzed with a pig as the ex-2.4. Measurements
perimental unit. The effect of the additional Fe injection, immunization procedure and immunization Pigs were weighed 1 week before weaning (d
moment on growth, blood Hb levels and antibody
28) and on the day of weaning (d 0). After
production were tested with the GLM procedures of weaning, pigs were weighed on days 6, 13, 20, 27
SAS (1989) by means of F-tests using a split-plot and 34. On the days of weighing, blood samples
model, with values in time of individual animals were taken using an ear prick and capillary tube (5
taken as repeated measurements:
ml) for analysis of blood hemoglobin (Hb)
con-centration. Blood Hb was determined using a mini- Y 5m 1Fe 1I 1T 1G1interaction
ijklmn i j k l photometer (AMESE Minilab; Miles Inc., Elkhart,
1e 1D 1interaction1e (1)
IN) and a reagent kit (AMESE Mini-pak hemoglo- 1,ijklm n 2,ijklmn
bin test pack; Miles, Inc.).
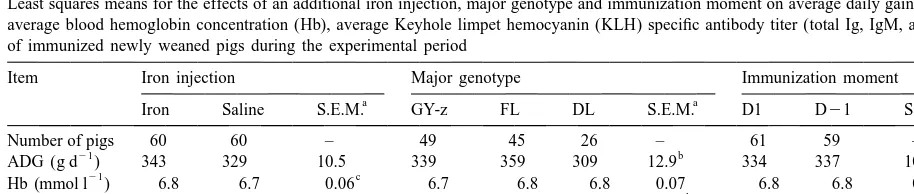
effect within animals between sampling days. The humoral immune response. Averaged over the total experimental period Hb concentration of FE pigs was effects of the additional iron injection, immunization
higher (P,0.05, Table 2) than that of SALINE pigs procedure, immunization moment, genotype and
21
(6.8 vs. 6.7 mmol l ). Averaged over the
ex-their interactions were tested against error term 1.
perimental period, Hb concentration was not affected The effect of sampling day and all interaction effects
by immunization procedure, immunization moment with sampling day were tested against error term 2.
and genotype (Table 2). Although blood Hb level of FE pigs was higher (P,0.01; Fig. 1) at day 6 after weaning, there was no interaction (P.0.1) between
3. Results
an additional iron injection and time (sampling moment). Both immunization procedure and im-Preliminary analysis of the various antibody titres
munization moment did also not interact with time with model (1) showed the presence of several three
(P.0.1; Fig. 1). The pattern (kinetics) of the blood way interaction effects between immunization
pro-Hb concentration was different among the three cedure, sampling day and the other effects.
Further-major genotypes as indicated by the interaction effect more, preliminary analysis within the PLAC
treat-between genotype and time (P,0.05; Fig. 1) con-ment showed that there were nearly no effects of an
centration. additional iron injection, immunization moment and
genotype. Therefore, the effects of an additional iron
3.2. Growth injection, immunization moment and genotype were
tested only in immunized animals using a similar
Averaged over the total experiment, ADG was not model as (1) omitting the effect of immunization.
affected (P.0.1) by an additional iron injection, In general, immunized pigs mounted similar
sys-immunization procedure and sys-immunization moment temic antibody responses directed to KLH and OA.
(Table 2). With regard to genotype, the highest ADG Therefore only the results of the KLH challenge will
was found in the predominantly Finnish Landrace be presented. The antibody response to TT will not
21
pigs (359 g day ; Table 2). Averaged over all be shown because it was masked by TT resembling
treatments, ADG increased over time (P,0.001) antigens at the time of analysis (cross-reactivity).
21 21
from 131 g day in week 1 to 471 g day in week
3.1. Blood hemoglobin concentration 5. This increase in ADG with time was not affected
(P.0.1) by an additional iron injection, immuniza-The primary aim of this study was to assess the tion procedure or immunization moment. Growth effect of an additional iron injection for weanling pattern was affected (P,0.001) by genotype. During pigs on day 21 on their Hb status, growth and the fifth week after weaning, pigs that had
pre-Table 2
Least squares means for the effects of an additional iron injection, major genotype and immunization moment on average daily gain (ADG), average blood hemoglobin concentration (Hb), average Keyhole limpet hemocyanin (KLH) specific antibody titer (total Ig, IgM, and IgG) of immunized newly weaned pigs during the experimental period
Item Iron injection Major genotype Immunization moment
a a a
Iron Saline S.E.M. GY-z FL DL S.E.M. D1 D21 S.E.M.
Number of pigs 60 60 – 49 45 26 – 61 59 –
S.E.M., pooled standard error of the mean.
b
Major genotype main effect, P,0.05.
c
Fig. 1. Least-square means of blood hemoglobin (Hb) concentration in newly weaned pigs during the experimental period. Panel A: the main effect of iron treatment (pigs injected with additional iron [FE] vs. placebo pigs [SALINE]); panel B: the main effect of immunization procedure (immunized pigs [IMMU] vs. placebo pigs [PLAC]); panel C: main effect of immunization moment (pigs immunized 1 day before [D21] or 1 day after [D1] weaning); and panel D: the main effect of major genotype (predominantly Finnish Landrace pigs [FL] vs. predominantly Dutch Landrace pigs [DL] vs. predominantly Great Yorkshire pigs [GY-z]).
dominantly the FL genotype achieved the highest moment (Table 2). Immunized pigs that had
pre-21
ADG (550 g day ), whereas, pigs that had pre- dominantly the GY-z genotype had a higher total Ig
dominantly the DL genotype achieved the lowest and IgG response to KLH than pigs that had
21 21
ADG (378 g day ). The ADG (485 g day ) of predominantly the FL or DL genotype (P,0.05;
pigs that had predominantly the GY-z genotype was Table 2). Genotype had no effect on the IgM
intermediate (Fig. 2). response to KLH.
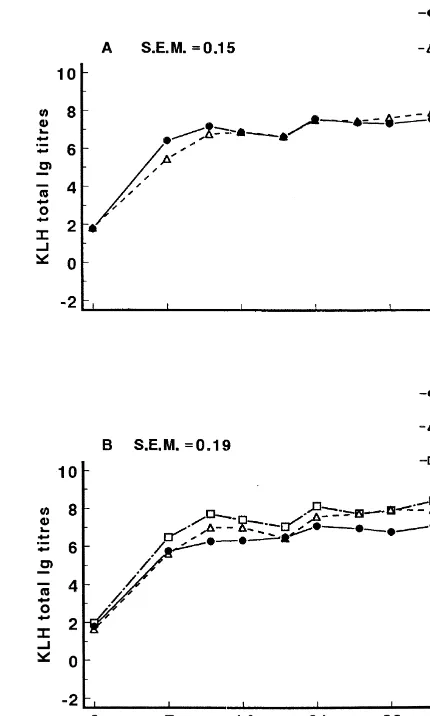
Total antibody titers and titers of the IgM and IgG
3.3. Keyhole limpet hemocyanin isotypes were affected by time (P,0.001). The
Fig. 2. Least-square means of ADG of newly weaned pigs during the experimental period. The main effect of major genotype (predominantly Finnish Landrace pigs [FL] vs. predominantly Dutch Landrace pigs [DL] vs. predominantly Great Yorkshire pigs [GY-z]).
responses (Figs. 3A, 4A and 5A). At day 7 the total Ig titer to KLH of pigs immunized 1 day before weaning (D21) was higher than of pigs immunized 1 day after weaning (D1). The pattern of total Ig response to KLH was also affected (P,0.005) by genotype (Fig. 3B). At day 11, the total Ig titer for
2
GY-z pigs was approximately 1.4 units ( log-scale) higher than that of FL pigs. This difference was reduced to approximately 1.1 units from day 14
Fig. 3. Least-square means of Keyhole limpet hemocyanin
onwards. The development of the total Ig titer (KLH)-specific total antibody titer of immunized newly weaned against KLH of DL pigs was intermediate to GY-z pigs during the experimental period. Panel A: the main effect of and FL pigs. In contrast, the kinetics of IgM and IgG immunization moment (pigs immunized 1 day before [D21] or 1 day after [D1] weaning); and panel B: the main effect of major
responses to KLH were not affected by genotype
genotype (predominantly Finnish Landrace pigs [FL] vs.
pre-(P.0.1; Figs. 4B and 5B).
dominantly Dutch Landrace pigs [DL] vs. predominantly Great Yorkshire pigs [GY-z]).
4. Discussion
21 21
(1.5 g dl 50.9 mmol l ) suggests that the mag-The additional iron injection on day 21 after birth nitude of the increase in blood Hb was higher in a did increase the blood Hb concentrations during the study of Kamphues than in our study.
nursery period (Table 2). The highest difference (0.4
21
mmol l ) was found at day 6 after weaning. In a 4.1. Effect of iron status on growth and immunity study of Kamphues et al. (1992), pigs were also
given an additional injection with iron at day 21 after Although the iron status was improved in our
21
Fig. 5. Least-square means of Keyhole limpet hemocyanin Fig. 4. Least-square means of Keyhole limpet hemocyanin (KLH)-specific antibody titer (IgG) of immunized newly weaned (KLH)-specific antibody titer (IgM) of immunized newly weaned pigs during the experimental period. Panel A: the main effect of pigs during the experimental period. Panel A: the main effect of immunization moment (pigs immunized 1 day before [D21] or 1 immunization moment (pigs immunized 1 day before [D21] or 1 day after [D1] weaning); and panel B: the main effect of major day after [D1] weaning); and panel B: the main effect of major genotype (predominantly Finnish Landrace pigs [FL] vs. pre-genotype (predominantly Finnish Landrace pigs [FL] vs. pre- dominantly Dutch Landrace pigs [DL] vs. predominantly Great dominantly Dutch Landrace pigs [DL] vs. predominantly Great Yorkshire pigs [GY-z]).
Yorkshire pigs [GY-z]).
21
7.6 mmol l , respectively, during a 6-week ex-Kamphues et al. (1992) and Schrama et al. (1997) perimental period. The lack of growth response to reported an improvement in growth resulting from an additional iron in this study may be due to either the increase in iron status. Like in our study, Kamphues small difference in iron status or the relatively low et al. (1992) injected pigs with additional iron at day growth response of the pigs (Table 2). The latter is 21 after birth. The difference in iron status in the supported by Kamphues et al. (1992) who found that study of Schrama et al. (1997) was created by the growth response to an additional iron injection injection of 100 or 200 mg of Fe (Fe-dextran) at day was most profound in the heaviest pigs.
the results of Schrama et al. (1997) and Kleinbeck However, they found that Dutch Landrace pigs and McGlone (1999). They also found no effects of showed the highest antibody response to KLH. The
iron status on, respectively the humoral immune Finnish Landrace and Norwegian Landrace pigs
response to KLH and plasma IgG concentrations showed intermediate antibody responses, whereas
using non-anemic pigs. However, reduced serum IgG Yorkshire pigs showed the lowest antibody response levels in iron deficient children were reported by to KLH.
´
Galan et al. (1988). This might suggest that iron In addition to antibody response to KLH, growth status only affects humoral immunity of pigs which was also affected by major genotype (Table 2). The
are anemic. genotype with the lowest antibody response had the
In contrast to the humoral immune response, highest growth (Finnish Landrace), whereas the
cellular immune response is known to be affected by genotypes with the highest antibody responses to the iron status of animals (De Sousa et al., 1988, KLH had the lowest growth (Great Yorkshire and 1991; Brock, 1994). They demonstrated decreased Dutch Landrace).
T-cell-dependent immune responses during iron de-ficiency. It remains questionable, however, that a
decrease in cellular immune response would have References
been demonstrated with the difference in iron status
achieved in this study using pigs without iron Blecha, F., Kelley, K.W., 1981. Effects of cold and weaning stressors on the antibody mediated immune response of pigs. J.
deficiency.
Anim. Sci. 53, 439–447.
Blecha, F., Charley, B., 1990. Rationale for using
immuno-4.2. Effect of immunization moment potentiators in domestic food animals. In: Blecha, F., Charley,
B. (Eds.), Immunomodulation in Domestic Food Animals.
Weaning can be considered a stressful event for Academic Press, San Diego, CA, pp. 3–19.
Blecha, F., Pollman, D.S., Nichols, D.A., 1983. Weaning pigs at an
pigs that is accompanied with reduced performance
early age decreases cellular immunity. J. Anim. Sci. 56, 396–
(Le Dividich and Herpin, 1994) and decreased
400.
immune responsiveness (Blecha et al., 1983; Blecha Bourne, F.J., 1986. The nutritional challenge of weaning. Nutr. and Charley, 1990). In the present study, all pigs Factor 2, 4–10.
experienced the stressful weaning process. Although Brock, J.H., 1994. Iron in infection, immunity, inflammation and neoplasia. In: Brock, J.H., Halliday, J.W., Pippard, M.J.,
both immunization moments were near this stressful
Powell, L.W. (Eds.), Iron Metabolism in Health and Disease.
experience, the results of the present study suggest
W.B. Saunders, London, pp. 354–389.
that immunization of pigs following a stressor can De Sousa, M., Breedvelt, F., Dynesius-Trentham, R.D., Trentham, affect the development of a humoral immune re- D., Lum, J., 1988. Iron, iron-binding proteins and immune
sponse (Figs. 3A, 4A and 5A). However, it should be system cells. Ann. NY Acad. Sci. 526, 310–322.
De Sousa, M., Reimao, R., Porto, G., Grady, R.W., Hilgartner,
realized that in this experiment there was a 2-day
M.W., Giardina, P., 1991. Iron and lymphocytes: reciprocal
difference in the development of the immune
re-regulatory interactions. In: Albertini, A., Lenfant, C.L.,
Man-sponses that could be responsible for the observed nucci, P.M., Sixma, J.J. (Eds.), Biotechnology of Plasma differences between both immunization moments. Proteins. Current Studies in Hematology and Blood
Transfu-The absence of the three-way interaction between sion. Karger, Basel, pp. 171–177, No. 58.
Frankena, K., 1987. The interaction between Cooperia spp. and
immunization moment, day and iron status suggests
Ostertagia spp. (Nematoda, Trichostrongylidae) in cattle. PhD
that additional iron did not affect the immune
thesis, Agricultural University Wageningen, The Netherlands.
responsiveness even under stressful conditions. Galan, P., Mekki, M., Hereberg, N., Hereberg, S., 1988. Iron´
deficiency, inflammatory processes and humoral immunity in
4.3. Effect of genotype children. Int. J. Vitam. Nutr. Res. 58, 225–230.
Gentry, J.L., Swinkels, J.W.G.M., Lindemann, M.D., Schrama, J.W., 1997. Effect of hemoglobin and immunization status on
In this study, antibody responses to KLH were
energy metabolism of weanling pigs. J. Anim. Sci. 75, 1032.
affected by major genotype of the pig (Table 2). Gwazdauskas, F.C., Gross, W.B., Bibb, T.L., McGillard, M.C., Genotype effects on immune responses of purebred 1978. Antibody titers and corticoids near weaning in steer and
Haye, S.N., Kornegay, E.T., 1979. Immunoglobin G, A and M and Makkink, C.A., 1993. Of pigs, dietary proteins, and pancreatic antibody response in sow reared and artificially-reared pigs. J. proteases. PhD dissertation, Department of Animal Nutrition, Anim. Sci. 48, 1116–1122. Wageningen Agricultural University, The Netherlands. Hershko, C., 1993. Iron, infection and immune function. Proc. Nabuurs, M., 1991. Etiologic and pathogenic studies on post
Nutr. Soc. 52, 165–174. weaning diarrhoea. PhD dissertation, State University, Utrecht, Joling, P., Mok, K.S., De Vries Reilingh, G., Wever, P.J.M., The Netherlands.
Cornelis, R.S., Oskam, J.P.H., Henken, H.M., 1993. An evalua- National Research Council (NRC), 1998. Nutrient requirements of tion of immune competence in different swine breeds. Vet. Q. swine, 10th Revised Edition. Subcommittee on Swine Nutri-15, 9–15. tion, Committee on Animal Nutrition, Board on Agriculture.
¨
Kamphues, J., Manner, K., Netzer, C., 1992. Effects of a 2nd iron SAS, 1989. SAS / STAT User’s Guide, Version 6, 4th Edition. injection in suckling pigs on iron retention and performance SAS Institute Inc, Cary, NC.
before and after weaning. In: Proc. 12th IPVS Cong., August Schrama, J.W., Schouten, J.M., Swinkels, J.W.G.M., Gentry, J.L., 1992, The Hague, The Netherlands, p. 601. De Vries Reilingh, G., Parmentier, H.K., 1997. Effect of Kleinbeck, S.N., McGlone, J.J., 1999. Intensive indoor versus hemoglobin status on humoral immune response of weanling outdoor swine production systems: genotype and supplemental pigs differing in coping styles. J. Anim. Sci. 75, 2588–2596. iron effects on blood hemoglobin and selected immune mea- Van Beers-Schreurs, L., Vellenga, T., Wensing, B., Breukink, H.J., sures in young pigs. J. Anim. Sci. 77, 2384–2390. 1992. The pathogenesis of the post-weaning syndrome in Le Dividich, J., Herpin, P., 1994. Effects of climatic conditions on weaned pigs: a review. Vet. Q. 14, 29–34.