PROFIL GEN CYT B SEBAGAI ACUAN KONSERVASI GENETIK TARSIUS SULAWESI (Tarsius tarsier kompleks)
Decky D. W. Kamagi
Jurusan Pendidikan Biologi FMIPA Universitas Negeri Manado deckykamagi@yahoo.com
ABSTRAK
Status konservasi beberapa spesies Tarsius Sulawesi (Tarsius tarsier kompleks) sekarang ini ada yang rentan punah bahkan ada yang hampir punah. Oleh karena itu usaha konservasi terutama konservasi genetik perlu dilakukan agar keberadaan spesies Tarsius Sulawesi tetap lestari. mtDNA telah banyak digunakan untuk mempelajari keragaman genetik dan kekerabatan beberapa spesies. Kajian ini menganalisis karakter gen cyt b pada beberapa spesies Tarsius Sulawesi dan menyidik keragaman genetik dan hubungan kekerabatan di antara beberapa spesies berbasis gen cyt b parsial. Gen cyt b parsial beberapa spesies Tarsius tipe lokalitas Sulawesi Utara, diamplifikasi dan disekuensing, dan selajutnya sekuen parsial gen cyt b yang diperoleh disejajarkan dengan gen cyt b yang selaras dari beberapa taksa berkerabat famili
Tarsiidae yang diambil dari GenBank. Ekstraksi DNA total menggunakan innuPREP DNA Micro kit, dan amplifikasi gen cyt b menggunakan primer umum L 14841 dan H15149. Rekonstruksi
pohon filogenetik dilakukan melalui metode Maximum Likelihood (ML). Hasil yang diperoleh menunjukkan bahwa rata-rata frekuensi basa pada sekuen gen cyt b parsial adalah T = 32.90%, A = 28.73%, C =22.42 dan G = 15.95%. Rekonstruksi pohon filogenetik menunjukkan bahwa Tarsius tipe lokalitas Sulawesi Utara bersekutu dengan kelompok Tarsius tarsier kompleks dan terpisah dari kelompok Cephalopahcus bancanus dan Carlito syrichta. Dengan demikian gen cyt b telah mengalami peristiwa substitusi basa dan dapat dipergunakan untuk membedakan antar spesies Tarsius dalam usaha konservasi genetik.
Kata Kunci : konservasi, Tarsius, Sulawesi, punah, DNA,gen cyt b
PENDAHULUAN
Sekuen DNA mitokondria DNA (mtDNA), seperti gen sitokrom b (cyt b) dan beberapa gen lainnya secara luas telah digunakan untuk kajian populasi genetik dan filogenetik (Avise et al., 1979, Brown et al., 1982). Sekuen tersebut telah terbukti berguna untuk memperkirakan waktu divergensi spesies dan populasi, perbandingan laju relatif evolusi, dan inferensia filogenetik dalam dan antar spesies vertebrata (Nei 1987; Avise 1994). Gen cyt b mitokondria diketahui sebagai gen yang lebih cepat berkembang dan dapat lebih bervariasi baik di dalam maupun antar spesies serta dapat digunakan untuk mengkaji filogeni dan biogeografi (Roos et al., 2008; Lim, et al., 2010; Randi, 1996; Karanth et al., 2008). Gen cyt b telah secara luas digunakan untuk kajian molekuler pada vertebtara karena ketersediaan primer-primer kekal yang bekerja dengan baik untuk amplifikasi pada berbagai spesies (Kocher et al., 1989). Gen cyt b mengandung kelas karakter diskrit yaitu, ketiga posisi basa pada kodon, yang memaparkan laju mutasi dimulai dari yang konservatif sampai dengan yang cepat (Irwin et al., 1991), sehingga sering digunakan untuk mengkaji evolusi molekuler. Informasi tentang variasi dari empat nukleotida yang disebabkan oleh pola substitusi dan variabilitas tingkat substitusi diantara tapak, sangat diperlukan dalam kajian evolusi molekuler. Karena gen-gen mitokondria memiliki tingkat substitusi yang lebih besar daripada yang terjadi pada gen-gen inti dan pada mtDNA primata kejadian transisi lebih sering daripada transversi (Brown, 1983), maka estimasi pola dan laju substitusi nukleotida diantara tapak dapat ditentukan. Kondo et al., (1993), dalam kajian mereka pada enam spesies primata menemukan bahwa tingkat transisi
lebih cepat 17 kali daripada tingkat transversi. Proses substitusi nukleotida umumnya dianggap sebagai transisi dan transversi. Transisi adalah substitusi diantara sesama basa purin (A↔G) atau diantara sesama pirimidin (C↔T), sedangkan transversi adalah substitusi basa purin oleh pirimidin atau sebaliknya (A↔C, A↔T, G↔C dan G↔T).
Tipe yang paling banyak dikaji dari semua jenis tingkat substitusi di antara empat nukleotida tersebut adalah bias transisi. Sejauh mana bias transisi itu biasanya diestimasi dengan menghitung jumlah perbedaan transisi dan transversi dari sekuen yang menyimpang dan mengambil rasio mereka. Bias transisi ini disebabkan oleh kurangnya mekanisme perbaikan mtDNA, atau karena suatu tautomerik pasangan basa yang dengan ikatan G-T dan A-C yang menyimpang dari model Watson-Crick (Wakeley, 1994). Masalah yang dihadapi pada hasil pengamatan sekuen nukleotida dimana pada tapak yang sama terjadi subsitusi ganda, yang akan mengaburkan bias transisi/transversi (Kimura, 1980; Wakeley, 1994; Yang dan Yoder, 1998).
Beberapa metode dengan pendekatan berbeda telah dikembangkan untuk mengestimasi bias transisi untuk keperluan analisis evolusioner molekuler. Tingkat substitusi diantara empat nukleotida dapat diestimasi melalui metode berdasarkan jarak, metode likehood dan parsymony (Kimura, 1980; Kumar, 1996; Yang dan Yoder, 1998). Kajian genom mitokondria utuh telah dilakukan pada C. syrichta (Matsui et al., 2009), C. bancanus (Andrews, et al. 1998; Schmitz et al. 2002), dan genom cyt b utuh juga telah dikaji pada, T. wallacei (Merker et al. 2010), T. lariang (Merker et al. 2009), T. dentatus (Merker et al., 2009), dan T. dentatus x T. lariang (Mereker et al., 2009) sehingga sebagian gen cyt b homolog mereka dapat disejajarkan dengan sekuen gen cyt b parsial dari Tarsius Sulawesi Utara: T. sangirensis, T. tarsier dan T. tumpara untuk mengungkap keragaman dan hubungan kekerabatan diantara mereka untuk kepentingan konservasi genetik.
MATERI DAN METODE
Pengumpulan dan Perlakuan Sampel
Spesimen sampel Tarsius Sulawesi Utara yang dikumpulkan terdiri dari: T. sangirensis, T. tarsier dan T. tumpara berasal dari 3 lokasi berbeda. Sekuen gen cyt b parsial beberapa spesies Tarsius dan aksesinya diambil dari GenBank.
Ekstraksi DNA dan Amplifikasi dan Sekuensing Amplikon Gen cyt b
DNA total diekstraksi menggunakan InnuPREP DNA micro Kit. Komponen dan kondisi PCR, kami optimasi sehingga gen cyt b dapat teramplifikasi. Primer yang digunakan adalah primer universal: L14841 dan H15149 (Kocher et al. 1989). Amplifikasi gen cyt b menggunakan mesin Boimetra Tpersonal. Produk amplifikasi dikirim ke perusahaan (First BASE, Laboratories Sdn. Bhd. Selangor, Malaysia) untuk disekuensing. Alat yang digunakan ABI PRISM 3730xl Genetic Analyzer. Biosystem USA.
Penjajaran Sekuen dan Analisis Data
Sekuens gen cyt b dijajarkan dengan penjajar otomatis ClustalW melalui aplikasi piranti lunak MEGA5 versi 5.2.2. Estimasi laju dan pola substitusi menggunakan dua pendekatan berbeda yaitu metode berdasarkan likehood dan parsimony. Data sekuens gen cyt b parsial dari C. bancanus, C. syrichta, T. wallacei, T. lariang, T. dentatus dan T. dentatus x T. lariang diperoleh dari GenBank.
HASIL PENELITIAN Amplifikasi sekuen
Primer L 14841 dan H 15149 mampu mengamplifikasi sekuen gen cyt b sampai dengan panjang sampai 306 bp sekuen gen cyt b. Panjang sekuen gen cyt b parsial yang teramplifikasi terletak mulai pada nukleotida ke-96 sampai ke-402 pada sekuen gen cyt b utuh atau terletak pada nukleotida ke-14265 sampai 14570 genom mtDNA utuh Tarsius.
Penjajaran sekuen
Sekuen gen cyt b parsial dengan panjang 306 pb dari T. sangirensis, T. tarsier dan T. tumpara disejajarkan dengan sekuen gen cyt b homolog dari beberapa spesies Tarsius yang diambil dari GenBank. Hasil penjajaran sekuen menunjukkan situs tak-beragam = 224 (73%), situs tak-beragam = 83 (27%), parsimoni informatif (Pi) = 83 (27%). Komposisi basa
Rata-rata frekuensi basa pada sekuens gen cyt parsial Tarsius, adalah T = 32.90%, A = 28.73%,C =22.42 dan G = 15.95%. Frekuensi basa pada masing-masing spesies yang tertinggi adalah basa T, sedangkan frekuensi yang paling rendah adalah basa G. Frekuensi basa T pada T. lariang dan T. dentatus x lariang adalah yang paling tinggi diantara diantara spesies lainnya, sedangkan frekuensi basa G pada T. dentatus adalah paling rendah diantara spesies lainnya. Frekuensi basa di setiap posisi pada kodon dengan kerangka pembacaan dimulai dari basa kedua pada sekuen - disesuaikan dengan kerangka bacaan pada proses tranlasi gen cyt b - menunjukkan variasi, dimana frekuensi basa T di posisi pertama dan kedua pada kodon adalah yang tertinggi, sedangkan frekuensi basa C di posisi pertama dan basa A pada posisi kedua adalah yang paling rendah. Frekuensi basa G di posisi ketiga pada kodon adalah yang paling rendah.
Hasil analisis sekuen gen cyt b parsial secara lengkap ditampilkan pada Tabel 1. Diversitas nukleotida (Pi) = 0.069833, dengan total jumlah mutasi = 97 dan tapak polimorpik = 83. Rasio ts/tv= (R) = 4.978 dan rasio ts/tv = (k) antara basa purin = 5.253 dan antara basa pirimidin = 13.418.
Tabel 1. Profil Sekuen Gen Cyt b Parsial
Parameter Posisi pada Kodon
Pertama Kedua Ketiga Total
Frekuensi tirosin 29.9 42.2 26.7 306
Frekuensi sitosin 17.6 21.6 28.1 306
Frekuensi adenin 25.8 17.6 42.7 306
Frekuensi guanin 26.7 18.6 2.5 306
Frekuensi tapak invariabel 73%
Frekuensi tapak parsimoni informatif 27%
Diversitas nukleotida (Pi) 0.069833
Jumlah haplotipe 9
Total jumlah mutasi 97
Situs polimorpik 83
Rasio ts/tv (k) Purin = 5.253, Pirimidin = 13.418
Rasio ts/tv (R) 4.978
Distribusi Gamma diskrit
(Model Tamura-Nei) 0.1882,
Rata-rata laju evolusioner 0.00, 0.01, 0.10, 0.63, dan 4.26 substitusi per situs
Catatan: Analisis pada posisi basa pada kodon hanya menggunakan panjang sekuen 306 nt, sedangkan parameter lainnya tetap menggunakan panjang sekuen 307 nt.
Substitusi Nukleotida
Tapak-tapak beragam yang ditemukan di sepanjang sekuen 307 nukleotida berjumlah 83 tapak. Tapak-tapak beragam tersebut disebabkab oleh kejadian substitusi basa disepanjang sekuen. Macam substitusi di 83 tapak beragam tersebut terdiri dari substitusi transisi dan tranversi dan sementara itu pada tapak yang sama juga terjadi substitusi-tunggal ganda (multi single-substitution). Kejadian substitusi transisi jauh lebih besar dari pada kejadian tranversi. Kejadian substitusi basa di posisi ketiga pada kodon ditemukan jauh lebih besar daripada substitusi basa di posisi pertama dan kedua, sedangkan substitusi basa di posisi pertama lebih besar daripada substitusi basa di posisi kedua. Kejadian substitusi transisi pada basa di posisi ketiga lebih besar daripada substitusi yang sama pada basa di posisi pertama dan kedua, begitu pula kejadian substitusi transversi basa pada posisi ketiga lebih besar daripada substitusi yang sama pada basa di posisi pertama dan kedua. Kejadian subsitusi transisi dan transversi pada basa di posisi kedua, jauh lebih rendah daripada substitusi yang sama pada basa pertama dan ketiga. Penghitungan substitusi tersebut diperoleh dengan mengabaikan substitusi-tunggal ganda pada tapak yang sama. Pada kejadian substitusi transisi, T↔C jauh lebih besar dibanding dengan A↔G dan sedangkan pada kejadian substitusi tranversi, C↔A lebih besar daripada kejadian substitusi yang sama pada pasangan basa lainnya.
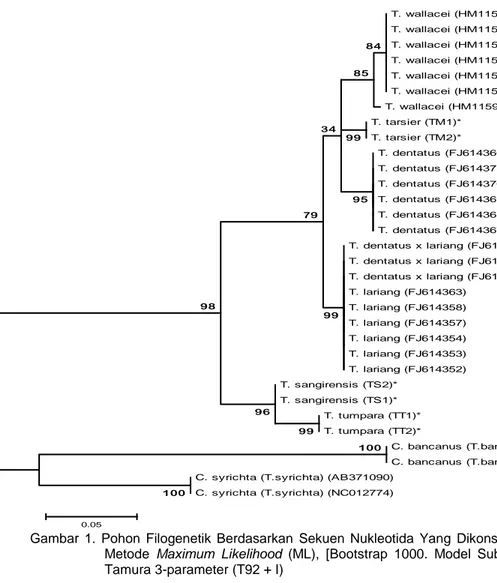
Gambar 1. Pohon Filogenetik Berdasarkan Sekuen Nukleotida Yang Dikonstruksi Dengan Metode Maximum Likelihood (ML), [Bootstrap 1000. Model Substitusi: Model Tamura 3-parameter (T92 + I) T. wallacei (HM115983) T. wallacei (HM115980) T. wallacei (HM115977) T. wallacei (HM115978) T. wallacei (HM115979) T. wallacei (HM115984) T. wallacei (HM115982) T. tarsier (TM1)* T. tarsier (TM2)* T. dentatus (FJ614366) T. dentatus (FJ614371) T. dentatus (FJ614370) T. dentatus (FJ614369) T. dentatus (FJ614368) T. dentatus (FJ614367) T. dentatus x lariang (FJ614361) T. dentatus x lariang (FJ614359) T. dentatus x lariang (FJ614356) T. lariang (FJ614363) T. lariang (FJ614358) T. lariang (FJ614357) T. lariang (FJ614354) T. lariang (FJ614353) T. lariang (FJ614352) T. sangirensis (TS2)* T. sangirensis (TS1)* T. tumpara (TT1)* T. tumpara (TT2)*
C. bancanus (T.bancanus) (AF348159) C. bancanus (T.bancanus) (AB011077) C. syrichta (T.syrichta) (AB371090)
C. syrichta (T.syrichta) (NC012774) 99 100 99 96 100 98 99 79 84 85 34 95 0.05
Rekonstruksi pohon filogenetik berbasis gen cyt b parsial melalui metode Maximum Likelihood (Gambar 1), menunjukkan bahwa bahwa Tarsius Sulawesi Utara berada pada klade yang berbeda dengan kelompok Tarsius Sulawesi lainnya dan kelompok C. bancanus dan C. syrichta.
PEMBAHASAN
Gen individual maupun genom keseluruhan secara signifikan beragam dalam komposisi nukleotida sudah diketahui. Keragaman nukleotida secara luas ditemukan pada DNA inti maupun DNA ekstra-kromosomal seperti mtDNA. Gen pengkode protein pada DNA mitokondria menunjukkan heterogenitas yang cukup besar pada perubahan sekuen (Bielawski dan Gold, 1996). Keragaman nukleotida suatu gen atau genom muncul karena adanya perubahan dari basa-basa nitrogen di situs tertentu pada sekuen nukleotida tersebut.
Komposisi basa dimana basa G pada level terendah merupakan ciri khas gen cyt b sebagai gen penyandi protein pada mamalia (Avise, 1994). Temuan Irwin et al., (1991) pada beberapa mamalia ternyata frekuensi basa G di semua posisi kodon adalah rendah. Keragaman komposisi basa pada nukleotida biasanya paling menonjol terdapat pada posisi basa pada kodon sinonim dari gen, karena redundansi dalam kode genetik, dan variasi dalam kandungan DNA yang mungkin memiliki sedikit efek pada kandungan asam amino yang menyandikan protein (Singer and Hickey, 2000). Keragaman komposisi basa pada nukleotida antara lain disebabkan oleh kejadian substitusi basa di beberapa tapak. Kejadian substitusi di sepanjang sekuen 307 bp sampel ditemukan di 83 situs berbeda. Adanya kejadian substitusi yang ditemukan di beberapa situs sekuen menyebabkan frekuensi basa di sepanjang sekuen Tarsius sp. menjadi beragam. Substitusi pada gen-gen mitokondria adalah suatu proses yang kompleks dan dapat bervariasi pada kelompok-kelompok taksonomik (Bielawski dan Gold, 1996). Kejadian substitusi dapat berupa substitusi transisi yaitu substitusi antara dua basa purin (adenin ↔ guanin) atau dua basa pirimidin (timin ↔ sitosin) dan substitusi transversi yaitu substitusi basa antara purin dan pirimidin (A↔T, A↔C, T↔G, dan C↔G). Kejadian substitusi di sejumlah tapak pada sekuen gen cyt parsial Tarsius adalah berupa substitusi transisi dan transversi, dengan rasio bias transisi ts/tv = 4. 978, menunjukkan bahwa perbandingan kejadian transisi lebih besar daripada transversi. Tingginya kejadian substitusi transisi dibandingkan transversi adalah biasa terjadi pada sekuen mtDNA eutharian (Brown, et al, 1984, Avise, 1994). Kocher et. al., (1989) mengemukakan bahwa biasanya pada mtDNA substitusi transisi lebih dominan daripada substitusi transversi, dan saturasi T ↔ C akan jauh lebih besar dibanding A ↔ G.
Secara luas telah diakui bahwa tingkat mutasi pada mtDNA adalah sangat tinggi. Pada banyak kasus ditemukan bahwa tingkat substitusi lebih tinggi terjadi pada posisi basa di kodon sinonim daripada posisi basa di kodon non-sinonim. Tingkat substitusi pada kodon non-sinonim bervariasi diantara gen-gen, dan pada beberapa kasus, kejadian itu lebih rendah pada mitokondria daripada gen inti (Saccone et al. 1991). Kejadian substitusi transisi lebih dominan daripada substitusi transversi adalah biasa terjadi pada mtDNA (Kocher et. al., (1997). Perolehan substitusi transisi lebih dominan daripada substitusi transversi, menyebabkan saturasi T↔C menjadi jauh lebih besar dibanding A↔G. Perbandingan kejadian substitusi transisi yang jauh lebih besar
daripada substitusi transversi sejalan dengan yang dikatakan Brown et. al., (1982) bahwa pada gen cyt b kejadian substitusi transisi lebih dominan daripada substitusi transversi.
Pengamatan bias transisi di tapak sinonim pada banyak kelompok taksa berkerabat dekat, menunjukkan bahwa substitusi transisional berkembang lebih cepat daripada substitusi transversional (Bielawski dan Gold, 1996). Lebih lanjut Kocher et al., (1989), mengemukakan bahwa substitusi nukleotida pada tingkat spesies sebagian besar adalah transisi sedangkan pada tingkat genus adalah transversi. Brown et. al., (1982) mengemukakan bahwa timbulnya kejadian transisi tinggi pada mtDNA adalah sebagai manisfestasi dari meningkatnya tekanan mutasi, namun demikian laju substitusi nukleotida tidak berkorelasi dengan laju perubahan struktural genom dari organela. Pada mamalia, mtDNA memang pesat dalam substitusi nukleotida, tetapi susunan dan ukuran gen dari genom tetap untuk setiap spesies (Castro, et al., 1998). Brown et. al., (1982) mengatakan bahwa faktor yang bertanggung jawab sehingga tingginya tingkat mutasi mtDNA melampaui DNA inti antara lain: a) kecenderungan rusaknya sistem replikasi, b) kurang efisiensinya fungsi pengeditan dan c) tingginya tingkat pertukaran (turnover). Sementara itu (Reyes et al., 1998) mengatakan bahwa tingginya tingkat mutasi yang diamati dalam mitokondria dapat dikaitkan dengan kerusakan oksidatif oleh radikal bebas yang dihasilkan selama pengangkutan elektron untuk oksigen yang terjadi pada kompleks rantai respirasi dalam membran mitokondria bagian dalam. Penyebab lain karena rendahnya aktivitas proof-reading DNA polymerase selama replikasi dan kurangnya sistem perbaikan DNA.
DAFTAR RUJUKAN
Andrews, T. D., L. S. Jermiin and S. Easteal. 1998. Accelerated evolution of cytochrome b in simian primates: adaptive evolution in concert with other mitochondrial proteins. J. Mol. Evol. 47 (3), 249-257.
Avise, J. C., R. A., Lansman, and R. O., Shade, 1979. Use endonuclease to Measure Mitochondrial DNA Sequence Relatedness in Natural Population. I. Population Structure and Evolution in Genus Peromyscus. Genetics 92:279-295.
Bielawski J. P. and J. R. Gold. 1996. Unequal Synonymous Substitution Rates Within and Between Two Protein-Coding Mitochondrial Genes. Mol. Biol. Evol. 13(6):889-892.
Brown, W. M., E.M. Prager, A.Wang, and A.C.Wilson, 1982. Mitochondrial DNA sequences of Primates: Tempo and Mode of Evolution. J. Mol.Evol. 18:225-239. Castro, J. A., A. Picornell, M. Ramon. 1998. Mitochondrial DNA: a tool for populational
genetics studies. Int. Microbiol. 1:327–332.
Clayton. D. A. 2000. Transcription and replication of mitochondrial DNA. Human Reproduction, Vol. 15. (Suppl. 2), pp. 11-17
Farias I. P., G. Ortı, I. Sampaio, H. Schneider, A. Meyer. 2001. The Cytochrome b Gene as a Phylogenetic Marker: The Limits of Resolution for Analyzing Relationships Among Cichlid Fishes. J. Mol Evol 53:89–103
Foster P. G., L. S. Jermiin, and D. A. Hickey. 1997. Nucleotide composition bias affects amino acid content in proteins coded by animal mitochondria. J. Mol. Evol. 44:282–288.
Groves C. and M. Shekelle, 2010. Genera and Spesies of Tarsiidae. Int. J. Primatol. 31:1071-1082.
Irwin D.M., T.D. Kocher and A.C. Wilson. 1991. Evolution of Cytochrome b Gene Mammals. J. Mol.Evol 32:128-144.
Karanth, K.P., S. Lalji, R. V. Collura and S.C. Beth. 2008. Molecular phylogeny and biogeography of langurs and leaf monkey of South Asia., Mol. Phylogenet., Evol., 46:683-694.
Lim, L. S., K. C. Ang, M.C. Mahani, A.W. Shahrom anf B.M. Md-Zain, 2010. Mitochondrial DNA polymorphism and phylogenetic relationships of proto malays in peninsular Malaysia. J. Biol.Sci.,10:71-83.
Kimura M. 1980. A simple method for estimating evolutionary rates of bases substitution trough comparative studies of nucleotide sequences, J. Mol. Evol. 16: 111 – 120.
Kocher, T. D., W.K. Thomas, A. Meyer, S.V. Edwards, S. Paabo, FX. Villablanca, A.C. Wilson, 1989, Dynamics of mitokondrial DNA evolution in animals: Amplification and sequencing with conserved primers. J. Proc. Natl. Acad. Sci. USA. 86:6196-6200.
Kocher R.D and K.L. Carleton. 1997. Base substitution in fish mitochondrial DNA: patters and rates. In Molecular systematics of Fish. Kocher, TD and Ca Stepien (Eds.). Academic Press: San Diego.
Kondo, R., S. Horay, Y. Satta, and N. Takahata. 1993. Evolution of hominid mitochondrial DNA with special reference to the silent substitution rate over the genome. J. Mol. Evol. 36:517-531.
Matsui, A., F. Rakotondraparany, I. Munechika, M. Hasegawa and S. Horai. 2009. Molecular phylogeny and evolution of prosimians based on complete sequences of mitocondrial DNAs. J. Gene 441 (1-2), 53-66..
Merker, S., Driller, C., Perwitasari-Farajallah, D., Pamungkas, J. and Zischler, H. 2009. Elucidating geological and biological processes underlying the diversification of Sulawesi tarsiers. J. Proc. Natl. Acad. Sci. U.S.A. 106 (21), 8459-8464.
Merker S., C. Driller, H. Dahruddin, Wirdateti, W. Sinaga, D. P. Farajallah and M. Shekelle. 2010. Tarsius wallacei: A New Tarsier Species from Central Sulawesi Occupies a Discontinuous Range. Int. journal Primate, Vol. 31 (6):1107-1122. Randi, E. 1996, A mitochondrial cytochrome phylogeny of the Alectoris partridges. J.
Mol. Phyl. Evol. 6:34-41.
Reyes, A., C. Gissi, G. Pesole, and C. Saccone, 1998. Asymmetrical Directional Mutation Pressure in the Mitochondrial Genome of Mammals. Mol. Biol. Evol. 15(8):957–966.
Roos, C., T. Nadler and L. Walter, 2008. Mitochondrial phylogeny, taxonomy and biogeography of silver langur spesies group (Trachypithecus cristatus). Mol. Phylogenet. Evol.,47:629-636.
Saccone, C., G. Pesole and B. Kadenbach, 1991. Evolutionary analysis of the nucleus-encoded subunits of mammalian cytochrome c oxidase. Eur. J. Biochem. 195:151– 156.
Schmitz, J. M., Ohme, H. Zischler, 2002. The complete mitochondrial sequence of Tarsius bancanus: Evidence for an extensive nucleotide compositional plasticity of primate mitochondrial DNA. Mol. Biol. Evol.,19:544-553.
Shekelle M. 2008. Distribution of Trasiers Acoustic Forms, North and Central Sulawesi: with Notes on The Primary Taxonomy of Sulawesi’s Tarsiers. Primates of The Oriental Night p.:35 – 50.
Singer G. A. C. and D. A. Hickey. 2000. Nucleotide Bias Causes a Genomewide Bias in the Amino Acid Composition of Proteins. Mol. Biol. Evol. 17(11):1581–1588. Wakeley, J. 1994. Substitution-Rate Variation among Sites and the Estimation of
Transition Bias. J. Mol. Biol. Evol, 11(3):436-441.
Yang Z. and A. Yoder, 1999. Estimation of the Transition/Transversion Rate Bias and Species Sampling. J. Mol Evol (1999) 48:274–283.