STRUKTUR GENETIK POPULASI MANGGIS (Garcinia
mangostana L.) INDONESIA
Abstrak
Keragaman dan struktur genetik populasi manggis (Garcinia mangostana
L.) Indonesia dipelajari menggunakan 13 primer yang terdiri dari 8 primer RAPD (Random Amplified Polymorphysm DNA) dan 5 primer ISSR (Inter-Simple Sequence Repeat). Tujuan penelitian ini adalah untuk mengungkap struktur genetik empat populasi manggis Indonesia berbasis marka molekuler. Bahan tanaman yang digunakan berasal dari 4 populasi manggis Indonesia yaitu Purwakarta (Jawa Barat), Kerinci (Jambi), Tembilahan (Riau) dan Bulukumba (Sulawesi Selatan). Jumlah sampel per populasi adalah sebanyak 21-33 tanaman dengan jumlah total 106 individu. Diversitas genetik populasi diobservasi melalui jumlah alel (Na), jumlah alel efektif, (Ne), Shannon's information indeks (I), persentase loki polimorfik (PPL) menggunakan program GenAlex 6.2. Perbedaan antar populasi dan dalam populasi dihitung menggunakan AMOVA pada program yang sama. Hasil penelitian menunjukkan bahwa populasi Purwakarta memiliki perbedaan gemetik tertinggi dan konsisten untuk semua parameter genetik yang diamati (Na=1.44, Ne=1.32, dan I=0.29), dan terendah pada populasi Kerinci (Na=1.00, Ne=1.17 dan I=0.15). AMOVA menunjukkan bahwa perbedaan genetik antar populasi sama dengan perbedaan genetik dalam populasi yaitu masing-masing sebesar 50%. Polymorfisme DNA yang dijumpai pada sejumlah aksesi dalam populasi mampu memberikan perbedaan genetik. Berdasarkan nilai PhiPT ke empat populasi manggis menunjukkan perbedaan genetik yang nyata satu sama lain pada taraf 1%. Pasangan populasi Bulukumba dan Kerinci memiliki jarak genetik terdekat (D=0.08) dan identitas genetik tertinggi (Nei I=0.92). Sebaliknya pasangan populasi Bulukumba dan Tembilahan menunjukkan jarak genetik yang terjauh (D=0.16) dan identitas genetik terendah (Nei I=0.85). Populasi Purwakarta dan Tembilahan masing-masing terbagi menjadi dua kelompok genetik berbeda.
POPULATION GENETIC STRUCTURE OF APOMICTIC
MANGOSTEEN (Garcinia mangostana L.) IN INDONESIA
Abstract
Population genetic structure in mangosteen (Garcinia mangostana L.) was studied using 13 primers consists of 8 Random Amplified Polymorphysm DNA (RAPD) and 5 Inter-Simple Sequence Repeat (ISSR) primers. The objective of this study was to study genetic structure of mangosteen populations as apomictic plant. The total of 106 samples was collected from seven mangosteen populations of four locations in Indonesia: Purwakarta (West Java), Kerinci (Jambi), Tembilahan (Riau) and Bulukumba (South Sulawesi). Molecular data were analyzed by GenAlex 6.2 program. The primers generated 132 bands of which 95 (72.2%) were polymorphic and 37 (28.8%) monomorphic. Cluster analysis
showed that the samples grouping based on their geographical origin. Genetic diversity observed at population levels found the number of locally common alleles (Na) ranging from 1.0 to 1.44, number of effective alleles (Ne) from 1.171 to 1.315, percentage of polymorphic loci from 30% to 62%, and Shannon’s information index (I) from 0.154 to 0.293. The highest level of genetic diversity within populations found in Purwakarta population (Na=1.32 ,Ne=1.32, and
I=0.28) and the lowest in Kerinci ((Na=1.00, Ne=1.17, dan I=0.15). Pairwise populations of Kerinci and Bulukumba showed the closest genetic distance (D= 0.08) with the highest uniformity (Nei I = 0.92). In contrast, the couple of Bulukumba and Tembilahan populations shows the furthest genetic distance (D= 0.164) with the lowest genetic uniformity (Nei I = 0.85). AMOVA showed that genetic diversity within population equal to among populations, i.e., 50%. Each of Purwakarta and Tembilahan populations divided into two distinct genetic groups. Keywords : apomictic, mangosteen, population genetic structure
Pendahuluan
Keragaman genetik manggis telah cukup banyak dipelajari dan sebagian besar diantaranya masih berbasis individu tanaman (Mansyah et al. 2003b; Sinaga et al. 2007a; Ramage et al. 2004). Hasil penelitian tersebut menunjukkan bahwa secara individual tanaman manggis menunjukkan variasi secara genetik. Penelitian berbasis individu dirasa belum cukup karena belum dketahui kapasitasnya dalam memberikan gambaran tentang keragaman genetik pada tingkat populasi. Untuk mengoptimalkan penggalian informasi tentang potensi keragaman sumberdaya genetik manggis, perlu dilakukan penelitian yang lebih intensif dengan menggunakan materi tanaman berbasis populasi pada daerah sentra produksi lainnya.
Banyak informasi menyebutkan bahwa daerah Kerinci (Propinsi Jambi) diduga merupakan daerah asal tanaman manggis karena pada daerah ini banyak ditemukan kerabat liarnya. Tembilahan diketahui memiliki populasi manggis dengan karakter morfologi yang khas (Mansyah et al. 2005), dan Purwakarta merupakan sentra produksi manggis yang sangat potensial di Jawa Barat. Sulawesi Selatan merupakan daerah yang melintasi garis Wallace dan memiliki luas panen manggis terbesar di pulau Sulawesi (BPS 2009) serta merupakan daerah asal dari G. celebica yang diduga sebagai progenitor manggis (Sulassih 2010).
Ahli genetika populasi telah lama mengenal bahwa keragaman genetik yang ada pada spesies merupakan struktur bertingkat (hierarchically structured). Perbedaan genetik mungkin terdapat antar individu didalam satu populasi, antar populasi didalam daerah geografi yang sama, antar populasi dari daerah geografi berbeda, dan antar seluruh daerah geografi (Holsinger & Mason-Gamer 1996). Penelitian berbasis populasi mempunyai beberapa keuntungan diantaranya dapat mendeteksi terjadinya percampuran genetik, menduga asal usul populasi dan cukup akurat untuk estimasi frekuensi alel didalam tiap populasi dan antar populasi (Palush et al. 2003). Pendekatan ini memegang peranan penting dalam mengurangi erosi genetik dan memungkinkan untuk identifikasi varietas lokal yang dijumpai pada daerah tertentu (Van der Hulst 2000).
Spesies tanaman menunjukkan variasi morfologi yang luas dan sebagian besar diantaranya bersifat adaptif (Wright & Brandon 2005). Setiap spesies dibatasi oleh jarak geografi dan kombinasi faktor biotik dan abiotik yang bekerjasama mempengaruhi dinamika populasi. Studi struktur genetik populasi bertujuan untuk mendefinisikan populasi dari suatu spesies, kuantifikasi besarnya penyebaran antar populasi, dan memahami hubungan kekerabatannya. Perbedaan populasi secara genetik biasanya didefinisikan berdasarkan signifikansi heterogenitas secara statistik dalam frekuensi alel antar sampel (Tessier & Bernatches.1999, dalam Wright & Brandon 2005).
Secara teori struktur genetik populasi kebanyakan tersedia untuk tanaman seksual. Pengetahuan tentang struktur genetik pada populasi tanaman apomiksis masih sangat terbatas. Ford & Richards (1985) dan van Oostrum et al (1985) telah meneliti tentang struktur genetik populasi agamosperm obligat pada Taraxacum dan memberikan gambaran bahwa diversitas genetiknya sangat rendah dengan heterozigositas (H) yang tinggi dibandingkan tipe seksual. Studi genetika populasi pada tanaman apomiksis dapat digunakan untuk menjelaskan diversitas klonal, lokasi geografi yang menunjukkan perbedaan genetik, kuantifikasi variasi dan hubungan antar populasi (Ellstrand & Roose 1987). Informasi ini penting dalam strategi pengembangan untuk koleksi, dan konservasi material tanaman sebagai sumberdaya genetik dan peningkatan pemanfaatannya.
Pemahaman struktur populasi pada beberapa daerah geografi hampir tidak memungkinkan untuk dilakukan tanpa bantuan pendekatan genetika molekular. Beberapa alasan diantaranya adalah bahwa data DNA polimorfisme jumlahnya sangat banyak, mewakili informasi genotipik yang lengkap, dapat mengintergrasikan informasi dalam periode waktu yang panjang dan dapat memberikan gambaran secara histori (Kreitman 1983). Berbagai teknik molekuler seperti RAPD (Random Amplified Polymorphysm DNA), AFLP (Amplified Fragment Length Polymorphysm), RFLP (Restriction Fragment Length Polymorphysm), SSR (Simple Sequence Repeat) dan ISSR (Inter Simple Sequence Repeat). Analisis RAPD telah digunakan pada studi apomiktik Hypericum perforatum dan memberikan identifikasi perbedaan individu. Marka
ISSR sangat baik untuk individu berkerabat dekat dan dapat diaplikasikan pada studi variasi di dalam populasi (Gonzales et al. 2005).
Tujuan penelitian ini adalah: (1) mengetahui variasi genetik pada empat populasi manggis di Indonesia, (2) mengungkap struktur genetik populasi manggis sebagai tanaman yang diperbanyak secara vegetatif melalui biji apomiktik.
Bahan dan Metode
Tempat dan Waktu
Penelitian dilaksanakan di Laboratorium Pusat Kajian Buah Tropika (PKBT-IPB), Laboratorium Balai Penelitian Tanaman Buah Tropika, dan Laboratorium Biologi Molekuler Balai Besar Penelitian dan Pengembangan Bioteknologi dan Sumberdaya Genetik Pertanian, mulai Januari 2010 sampai Maret 2011 .
Bahan dan Alat
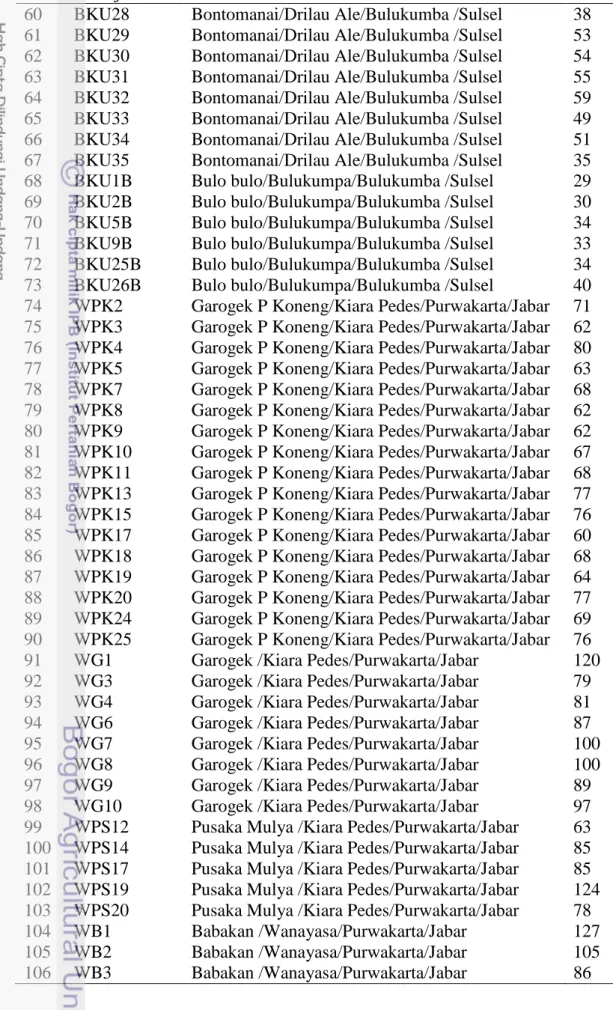
Materi tanaman yang digunakan adalah 106 sampel daun tanaman manggis (Tabel 8) yang berasal dari empat populasi manggis Indonesia yaitu Kabupaten Purwakarta (Jawa Barat), Kabupaten Kerinci (Jambi), Kabupaten Tembilahan (Riau) dan serta Kabupaten Bulukumba (Sulawesi Selatan) (Gambar 24). Jumlah tanaman sampel pada tiap populasi adalah sebanyak 21-33 individu. Karakteristik umum dari lokasi pengambilan sampel disajikan Pada Tabel 9.
Tabel 8 Aksesi manggis yang digunakan untuk analisis struktur genetik populasi No Kode aksesi Daerah asal (Desa/kecamatan
kabupaten/propinsi)
Lingkar batang (cm)
1 TMB1 P. Palas/Tembilahan Hulu/Tembilahan /Riau 108
2 TMB2 P. Palas/Tembilahan Hulu/Tembilahan /Riau 104
3 TMB3 P. Palas/Tembilahan Hulu/Tembilahan /Riau 85
4 TMB5 P. Palas/Tembilahan Hulu/Tembilahan /Riau 88
5 TMB7 P. Palas/Tembilahan Hulu/Tembilahan /Riau 103
6 TMB8 P. Palas/Tembilahan Hulu/Tembilahan /Riau 124
7 TMB9 P. Palas/Tembilahan Hulu/Tembilahan /Riau 104
8 TMB10 P. Palas/Tembilahan Hulu/Tembilahan /Riau 72
9 TMB13 P. Palas/Tembilahan Hulu/Tembilahan /Riau 117
10 TMB14 P. Palas/Tembilahan Hulu/Tembilahan /Riau 85
11 TMB19 P. Palas/Tembilahan Hulu/Tembilahan /Riau 92
Tabel 8 Lanjutan
13 TMB22 P. Palas/Tembilahan Hulu/Tembilahan /Riau 100
14 TMB24 P. Palas/Tembilahan Hulu/Tembilahan /Riau 95
15 TMB25 P. Palas/Tembilahan Hulu/Tembilahan /Riau 96
16 TMB26 P. Palas/Tembilahan Hulu/Tembilahan /Riau 97
17 TMB28 P. Palas/Tembilahan Hulu/Tembilahan /Riau 113
18 TMB29 P. Palas/Tembilahan Hulu/Tembilahan /Riau 60
19 TMB34 P. Palas/Tembilahan Hulu/Tembilahan /Riau 93
20 TMB35 P. Palas/Tembilahan Hulu/Tembilahan /Riau 97
21 TMB36 P. Palas/Tembilahan Hulu/Tembilahan /Riau 30
22 KRM Semerap/Keliling danau/Kerinci /Jambi 152
23 KRI1 Semerap/Keliling danau/Kerinci /Jambi 47
24 KRI2 Semerap/Keliling danau/Kerinci /Jambi 37
25 KRI3 Semerap/Keliling danau/Kerinci /Jambi 84
26 KRI4 Semerap/Keliling danau/Kerinci /Jambi 36
27 KRI5 Semerap/Keliling danau/Kerinci /Jambi 76
28 KRI7 Semerap/Keliling danau/Kerinci /Jambi 68
29 KR10 Semerap/Keliling danau/Kerinci /Jambi 72
30 KR11 Semerap/Keliling danau/Kerinci /Jambi 53
31 KR12 Semerap/Keliling danau/Kerinci /Jambi 66
32 KR13 Semerap/Keliling danau/Kerinci /Jambi 60
33 KR14 Semerap/Keliling danau/Kerinci /Jambi 57
34 KR15 Semerap/Keliling danau/Kerinci /Jambi 67
35 KR17 Semerap/Keliling danau/Kerinci /Jambi 78
36 KR18 Semerap/Keliling danau/Kerinci /Jambi 56
37 KR19 Semerap/Keliling danau/Kerinci /Jambi 66
38 KR21 Semerap/Keliling danau/Kerinci /Jambi 61
39 KR22 Semerap/Keliling danau/Kerinci /Jambi 61
40 KR23 Semerap/Keliling danau/Kerinci /Jambi 62
41 KR24 Semerap/Keliling danau/Kerinci /Jambi 33
42 KR25 Semerap/Keliling danau/Kerinci /Jambi 29
43 KR26 Semerap/Keliling danau/Kerinci /Jambi 38
44 KR28 Semerap/Keliling danau/Kerinci /Jambi 35
45 KR29 Semerap/Keliling danau/Kerinci /Jambi 40
46 KR30 Semerap/Keliling danau/Kerinci /Jambi 37
47 BKU5 Sawere/Gangking/Bulukumba /Sulsel 114
48 BKU7 Sawere/Gangking/Bulukumba /Sulsel 104
49 BKU8 Bontomanai/Drilau Ale/Bulukumba /Sulsel 37
50 BKU13 Bontomanai/Drilau Ale/Bulukumba /Sulsel 75
51 BKU14 Bontomanai/Drilau Ale/Bulukumba /Sulsel 64
52 BKU15 Bontomanai/Drilau Ale/Bulukumba /Sulsel 76
53 BKU16 Bontomanai/Drilau Ale/Bulukumba /Sulsel 68
54 BKU17 Bontomanai/Drilau Ale/Bulukumba /Sulsel 49
55 BKU19 Bontomanai/Drilau Ale/Bulukumba /Sulsel 64
56 BKU21 Bontomanai/Drilau Ale/Bulukumba /Sulsel 74
57 BKU23 Bontomanai/Drilau Ale/Bulukumba /Sulsel 54
58 BKU26 Bontomanai/Drilau Ale/Bulukumba /Sulsel 36
Tabel 8 Lanjutan
60 BKU28 Bontomanai/Drilau Ale/Bulukumba /Sulsel 38
61 BKU29 Bontomanai/Drilau Ale/Bulukumba /Sulsel 53
62 BKU30 Bontomanai/Drilau Ale/Bulukumba /Sulsel 54
63 BKU31 Bontomanai/Drilau Ale/Bulukumba /Sulsel 55
64 BKU32 Bontomanai/Drilau Ale/Bulukumba /Sulsel 59
65 BKU33 Bontomanai/Drilau Ale/Bulukumba /Sulsel 49
66 BKU34 Bontomanai/Drilau Ale/Bulukumba /Sulsel 51
67 BKU35 Bontomanai/Drilau Ale/Bulukumba /Sulsel 35
68 BKU1B Bulo bulo/Bulukumpa/Bulukumba /Sulsel 29
69 BKU2B Bulo bulo/Bulukumpa/Bulukumba /Sulsel 30
70 BKU5B Bulo bulo/Bulukumpa/Bulukumba /Sulsel 34
71 BKU9B Bulo bulo/Bulukumpa/Bulukumba /Sulsel 33
72 BKU25B Bulo bulo/Bulukumpa/Bulukumba /Sulsel 34
73 BKU26B Bulo bulo/Bulukumpa/Bulukumba /Sulsel 40
74 WPK2 Garogek P Koneng/Kiara Pedes/Purwakarta/Jabar 71
75 WPK3 Garogek P Koneng/Kiara Pedes/Purwakarta/Jabar 62
76 WPK4 Garogek P Koneng/Kiara Pedes/Purwakarta/Jabar 80
77 WPK5 Garogek P Koneng/Kiara Pedes/Purwakarta/Jabar 63
78 WPK7 Garogek P Koneng/Kiara Pedes/Purwakarta/Jabar 68
79 WPK8 Garogek P Koneng/Kiara Pedes/Purwakarta/Jabar 62
80 WPK9 Garogek P Koneng/Kiara Pedes/Purwakarta/Jabar 62
81 WPK10 Garogek P Koneng/Kiara Pedes/Purwakarta/Jabar 67
82 WPK11 Garogek P Koneng/Kiara Pedes/Purwakarta/Jabar 68
83 WPK13 Garogek P Koneng/Kiara Pedes/Purwakarta/Jabar 77
84 WPK15 Garogek P Koneng/Kiara Pedes/Purwakarta/Jabar 76
85 WPK17 Garogek P Koneng/Kiara Pedes/Purwakarta/Jabar 60
86 WPK18 Garogek P Koneng/Kiara Pedes/Purwakarta/Jabar 68
87 WPK19 Garogek P Koneng/Kiara Pedes/Purwakarta/Jabar 64
88 WPK20 Garogek P Koneng/Kiara Pedes/Purwakarta/Jabar 77
89 WPK24 Garogek P Koneng/Kiara Pedes/Purwakarta/Jabar 69
90 WPK25 Garogek P Koneng/Kiara Pedes/Purwakarta/Jabar 76
91 WG1 Garogek /Kiara Pedes/Purwakarta/Jabar 120
92 WG3 Garogek /Kiara Pedes/Purwakarta/Jabar 79
93 WG4 Garogek /Kiara Pedes/Purwakarta/Jabar 81
94 WG6 Garogek /Kiara Pedes/Purwakarta/Jabar 87
95 WG7 Garogek /Kiara Pedes/Purwakarta/Jabar 100
96 WG8 Garogek /Kiara Pedes/Purwakarta/Jabar 100
97 WG9 Garogek /Kiara Pedes/Purwakarta/Jabar 89
98 WG10 Garogek /Kiara Pedes/Purwakarta/Jabar 97
99 WPS12 Pusaka Mulya /Kiara Pedes/Purwakarta/Jabar 63
100 WPS14 Pusaka Mulya /Kiara Pedes/Purwakarta/Jabar 85
101 WPS17 Pusaka Mulya /Kiara Pedes/Purwakarta/Jabar 85
102 WPS19 Pusaka Mulya /Kiara Pedes/Purwakarta/Jabar 124
103 WPS20 Pusaka Mulya /Kiara Pedes/Purwakarta/Jabar 78
104 WB1 Babakan /Wanayasa/Purwakarta/Jabar 127
105 WB2 Babakan /Wanayasa/Purwakarta/Jabar 105
3 3 1 2 Desa Semerap Danau Kerinci Tembilahan Kerinci Purwakarta Bulukumba
Tabel 9 Karakteristik umum lokasi populasi manggis. (Desa/kecamatan
kabupaten/propinsi)
Habitat/ketinggian Jumlah
sampel Semerap, Keliling Danau,
Kerinci, Jambi
Lahan kering, berbukit / ± 875 m dpl 25
Pulau Palas, Tembilahan, Riau Rawa pasang surut, datar/ ± 16 m dpl 21 Wanayasa, Kiara Pedes,
Purwakarta, Jawa Barat
Lahan kering, datar / ± 783 m dpl 33
Bontomanai dan Bulo-bulo, Bulukumba, Sulawesi Selatan
Lahan kering, datar / ± 30 m dpl 27
m dpl = meter diatas permukaan laut
Ekstraksi, Purifikasi, Penentuan Kuantitas DNA dan Amplifikasi
Prosedur isolasi DNA, purifikasi, penentuan kuantitas DNA, amplifikasi sama dengan kegiatan 2. Primer yang digunakan terdiri dari 8 primer RAPD dan 5 primer ISSR seperti tercantum pada Tabel 6.
Analisis Data
Data hasil amplifikasi diskor sebagai data biner yaitu pita-pita yang dimiliki bersama diberi skor 1 (ada) dan jika tidak diberi skor 0. Keragaman genetik dianalisis dengan program NTSYS pc 2.1 (Rohlf 2000). Struktur genetik populasi dianalisa menggunakan program GenAlex 6.2 (Peakall & Smouse 2006). Pada program ini data biner marka dominan RAPD dan ISSR dapat digunakan untuk menghitung frekuensi alel melalui asumsi seperti marka kodominan sebagai berikut:
(1) atau ada pita mewakili genotipe AA / Aa, (0) atau tidak ada pita mewakili genotipe aa . Frekuensi alel A=. p = 1 – q , frekuensi alel a = q = 1 − p
Frekuensi genotip aa = q2
= frekuensi pita tidak ada = 1− frekuensi pita ada q = √(frekuensi pita tidak ada )
Diversitas genetik populasi ditentukan berdasarkan parameter berikut : 1. Jumlah alel berbeda (Na), nilainya >1.
Dihitung langsung berdasarkan proporsi alel berbeda. Mewakili semua lokus dengan jumlah pita dengan frekuensi .>5%.
2. Jumlah alel efektif (Ne), Nilainya berkisar antara 1 sampai < n alel.
Mengukur perbandingan diversitas alel antar lokus dengan distribusi frekuensi alel yang luas. Menyediakan pendugaan jumlah frekuensi alel yang sama pada populasi ideal dengan homozigositas setara dengan populasi aktual. Dihitung dengan rumus: Ne= 1 / (p2 + q2)
3. Persentase lokus polimorfik (PPL): Dihitung sebagai persentase lokus polimorfik dari seluruh lokus.
4. Shannon’s information index (I): Nilainya > 0, dihitung berdasarkan frekuensi
pita ada (p) dan tidak ada (q) dengan rumus: I = -1* (p*ln (p)+q*ln(q)).
5. Variasi Antar populasi (AP) dihitung menggunakan AMOVA pada program GenAlex 6.2
6. Variasi dalam populasi (WP) = dihitung menggunakan AMOVA (molecular analysis of variance)
7. PhiPT (perbedaan genetik antara pasangan populasi). Nilainya berkisar antara -1.0 sampai 1.0. Mewakili korelasi antara individu dalam populasi, terhadap total (analog dengan Fst ). Dihitung sebagai proporsi varians antar populasi (VAP), terhadap total varians (VWP) pada AMOVA:
8. Jarak genetik (GD) , nilainya > 0, dihitung dengan rumus
2nxy = jumlah karakter sama n = jumlah total karakter biner.
9. Identitas genetik (Nei’s Genetic Identity/Nei I). Nilainya 0.0 – 1.0, dihitung untuk menentukan keseragaman genetik dengan rumus:
pix dan piy = frekuensi alel ke i dalam populasi x dan y. Untuk multipel lokus , Jxy, Jx dan Jy dihitung dengan menjumlahkan semua loki dan alel dan dibagi dengan jumlah lokus.
Untuk menguji apakah perbedaan genetik antara pasangan populasi (PhiPT) berkorelasi nyata dengan jarak genetik antar populasi geografi digunakan uji F statistik (1%). Selanjutnya kladogram dikonstruksi menggunakan metode UPGMA dengan koeffisien Jaccard pada program Free Tree dan TreeV32.
Hasil dan Pembahasan
Polimorfisme Marka RAPD dan ISSR pada Empat Populasi Manggis
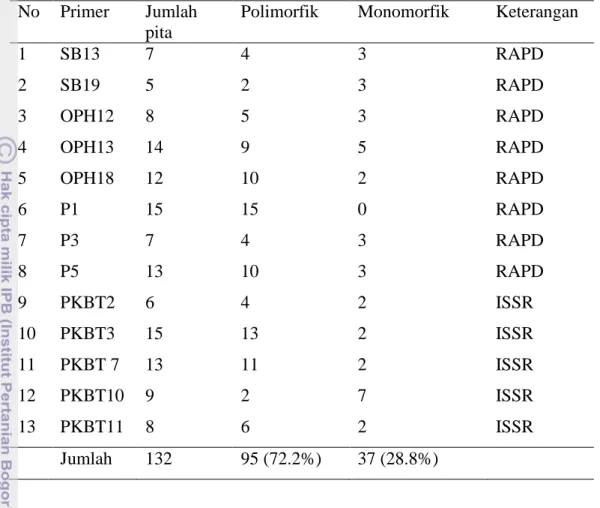
Hasil analisis PCR dari semua primer yang digunakan berupa 132 pita DNA yang terdiri dari 95 (72.2%) pita polimorfik dan 37 (28.8%) pita monomorfik. Jumlah pita per primer antara 5–15 pita yang terdiri dari beberapa pita yang umum dan pita spesifik dengan ukuran antara 250 sampai 2400 bp. Produk amplifikasi dari 8 primer RAPD dan 5 primer ISSR disajikan pada Tabel 10 dan bagian dari analisis PCR pada Lampiran 5, 6, 7, dan 8.
Tingkat polimorfisme gabungan marka RAPD dan ISSR pada penelitian ini (72.2%), lebih tinggi daripada analisis ISSR pada kegiatan 1 yaitu sebesar 58%. Hasil penelitian sebelumnya menggunakan 5 primer RAPD pada 23 sampel manggis diperoleh 42% pita polimorfis (Mansyah et al. 2003b). Hasil ini hampir sama dengan studi menggunakan marka ISSR pada tanaman apomiksis Psammochloa villosa (Poaceae) dengan 70.52% pita polimorfik (Li & Ge 2001) dan marka AFLP pada Rubus alceifolius dengan 78% pita polimorfik (Amsellem et al. 2000). Ramage et al. (2004) melaporkan 42.45% pita polimorfis pada 37 aksesi manggis dengan menggunakan teknik Randomly Amplified DNA Fingerprinting (RAF).
Tabel 10 Produk amplifikasi dari 8 primer RAPD dan 5 primer ISSR pada 106 aksesi manggis
No Primer Jumlah
pita
Polimorfik Monomorfik Keterangan
1 SB13 7 4 3 RAPD 2 SB19 5 2 3 RAPD 3 OPH12 8 5 3 RAPD 4 OPH13 14 9 5 RAPD 5 OPH18 12 10 2 RAPD 6 P1 15 15 0 RAPD 7 P3 7 4 3 RAPD 8 P5 13 10 3 RAPD 9 PKBT2 6 4 2 ISSR 10 PKBT3 15 13 2 ISSR 11 PKBT 7 13 11 2 ISSR 12 PKBT10 9 2 7 ISSR 13 PKBT11 8 6 2 ISSR Jumlah 132 95 (72.2%) 37 (28.8%)
Perbedaan polimorfisme ini dapat disebabkan oleh perbedaan primer dan materi tanaman yang digunakan. Pada penelitian ini digunakan kombinasi antara RAPD dan ISSR dengan materi tanaman berbeda dan jumlah sampel yang lebih banyak dari penelitian sebelumnya. Fomeck 2005 (dalam Wegscheider 2009) telah melakukan penelitian variasi klonal didalam kultivar anggur dan mendapatkan bahwa tingkat keluasan genetik tergantung pada marka yang diaplikasikan dan tipe sampel tanaman. Retrotransposon-based marker systems, MSAP (methyl-sensitive amplified length polymorphism) atau ISTR ( Inter-Retrotransposon Amplified Polymorphism) menunjukkan level polimorfisme yang tinggi daripada AFLP. Variasi klonal yang dipelajari pada klon Pinot sering tidak terlihat tetapi berhasil pada klon yang lain. Klon Traminer menunjukkan perbedaan variabilitas klonal dengan kultivar yang lain (Imazio et al. 2002).
Polimorfisme dan diversitas genetik pada tanaman apomiksis merupakan hasil dari mutasi poin meliputi insersi, delesi, inversi, non disjunction,
rekombinasi somatik oleh aktivitas transposon, dan mutasi atau perubahan kromosom pada genom maternal (Richards 1997). Transposable element diketahui sebagai gen loncat (jumping gene) yang merupakan sekuen DNA bersifat mobil yang dijumpai pada banyak organisme hidup, baik prokaryotik maupun eukaryotik. Gen ini mempunyai kemampuan untuk meloncat dan merubah lokasinya di dalam genom, serta mengamplifikasi jumlah kopinya. Keberadaan dan aktifitasnya dapat menyebabkan mutasi dan perubahan morfologi (Capy et al. 1998 dalam Wegscheider 2009).
Alasan lain yang tidak dapat diabaikan adalah bahwa penggunaan mikrosatelit sering menghasilkan variasi yang lebih tinggi dari marka lainnya (Paun & Horandl 2006). Inter-simple sequence repeat (ISSR) merupakan semiarbitrary marker yang komplemen dengan mikrosatelit dengan tingkat polimorfisme tinggi, dan menghasilkan marka dominan (Zietkiewicz et al. 1994; Mishra et al. 2003). SSR merupakan bagian non fungsional pada kromosom dengan kecepatan mutasi yang lebih tinggi daripada fragmen lainnya yaitu sekitar 2.5x10-5 sampai 1x10-2 mutasi per lokus per gamet per generasi, dengan variasi tergantung pada lokus dan organisme (Hong et al. 2006).
Pada anggur klon Pinot Noir retrotransposon terdapat dimana-mana pada genom tanaman dan menyebabkan mutasi (Wegscheider 2009). Tingginya tingkat polimorfisme menunjukkan perbedaan sekuen retrotransposon. Retrotransposon ini menginduksi mutasi dan mungkin bertanggung jawab terhadap variasi klonal antar Pinot noir, yang menyebabkan perbedaan genetik dan bahkan fenotipik. Dalam genom tanaman retrotransposon terdapat dalam jumlah kopi yang besar dan bervariasi. Genom kecil seperti Arabidopsis thaliana (125 Mb), mengandung sekitar 4-8% (Kapitonov & Jurka 1999), Oryza sativa sekitar 10% (Mao et al. 2000), Zea mays kira-kira 50% (Bennetzen 2000), dan genom besar seperti Hordeum vulgäre (5000 Mb), mengandung 70% (Vicient et al. 1999, dalam Bennetzen 2000). Bennetzen & Kellogg (1997) mengatakan bahwa ukuran genom berkorelasi positif dengan jumlah kopi transposon dan retrotransposon memegang peranan penting dalam sejarah evolusi tanaman. Diperkirakan manggis juga memiliki kopi transposon yang tinggi karena memiliki level poliploidi yang tinggi.
Adanya pita-pita spesifik seperti OPH-12 1400 bp, OPH-13/2400 bp, 950 bp dan OPH-18/1800 bp, 950 bp dan 850 bp, PKBT-3/1000 bp, PKBT-7/775 bp, dan PKBT-2 1900 bp yang berasosiasi dengan beberapa individu populasi Tembilahan (Lampiran 6) merupakan hal yang menarik. Tiga aksesi dengan pola pita DNA yang unik (WPK2, WPK8, dan WPK 11) juga dijumpai dalam populasi Purwakarta (Lampiran 7).
Analisis Kluster
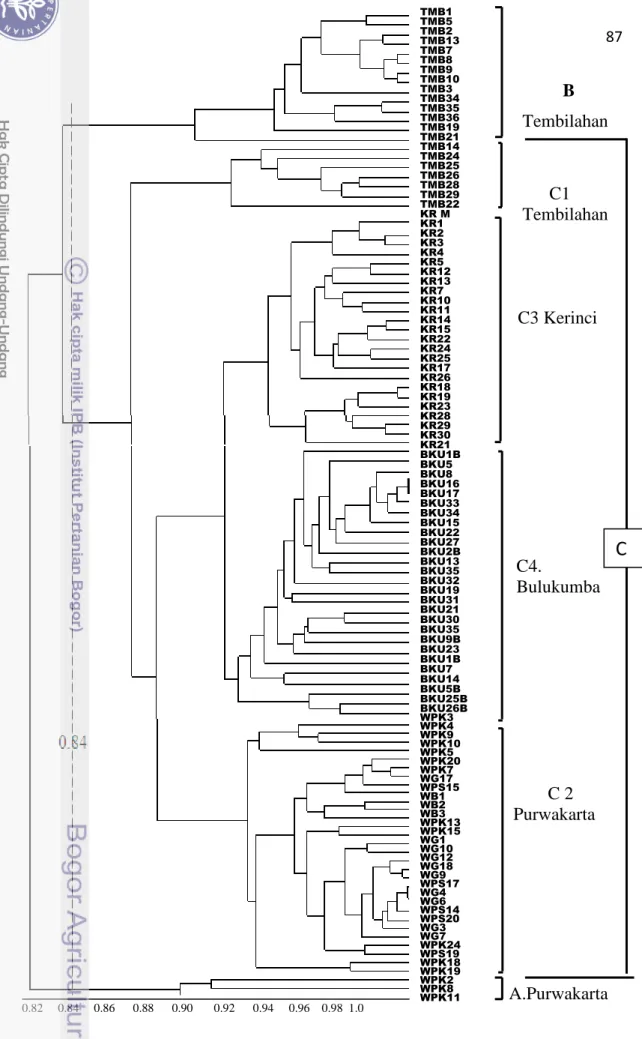
Koefisien kemiripan genetik dari 106 aksesi manggis berdasarkan kombinasi analisis RAPD dan ISSR bervariasi antara 0.82-0.99 (Gambar 25).
Tingkat kemiripan genetik keempat populasi manggis ini lebih tinggi dibandingkan dengan penelitian menggunakan analsis RAPD (Mansyah et al
2003b) yaitu sebesar 0.75-1.0, analysis RAPD (Sinaga et al. 2008) sebesar 0.71 – 1.00), dan AFLP (Sobir et al 2009) sebesar 46-77%. Hasil ini hampir sama dengan Ramage et al. (2004) yang memperoleh koefisien kemiripan genetik sebesar 0.86-0.98 pada 37 aksesi manggis. Perbedaan koefisien kemiripan ini dapat diebabkan oleh perbedaan materi tanaman yang digunakan, jumlah sampel, jumlah primer dan tingkat polimorfisme primer yang digunakan. Nilai korelasi antar pita adalah sebesar r=0.91 (Lampiran 9), yang menunjukkan bahwa pengelompokan berdasarkan pita DNA sangat baik).
Pada koefisien kemiripan 0.84 analisis kluster membagi aksesi kedalam tiga kelompok yaitu: 1) Tiga aksesi unik dari Purwakarta (kluster A); 2). Empat belas aksesi dari Tembilahan (cluster B); 3). dan 90 aksesi lainnya (kluster C). Di dalam kluster C, terdapat empat subkluster yang terbentuk berdasarkan lokasi geografi yaitu Tembilahan (subkluster C1), Purwakarta (subkluster C2), Kerinci (subkluster C3), dan Bulukumba (subkluster C4). Beberapa variasi juga terlihat didalam tiap subkluster. Koefisien kemiripan terendah atau variasi genetik tertinggi terlihat pada populasi Purwakarta (0.82-0.99) dan Tembilahan (0.84-0.99). Hal ini dapat disebabkan oleh adanya genotipe dengan pola pita DNA yang unik dibandingkan dengan individu lainnya (Lampiran 5 dan 6). Populasi Purwakarta dan Tembilahan masing-masing terbagi menjadi dua subkluster yaitu A dan C2 serta B dan C1.
B Tembilahan C1 Tembilahan C3 Kerinci C4. Bulukumba C 2 Purwakarta A.Purwakarta
Gambar 25 Dendrogram 106 aksesi manggis berdasarkan 5 primer RAPD and 11 primer ISSR.
C
0.82 0.84 0.86 0.88 0.90 0.92 0.94 0.96 0.98 1.0 TMB7 TMB8 TMB9 TMB10 TMB3 TMB34 TMB35 TMB36 TMB19 TMB21 TMB14 TMB24 TMB25 TMB26 TMB28 TMB29 TMB22 KR M KR1 KR2 KR3 KR4 KR5 KR12 KR13 KR7 KR10 KR11 KR14 KR15 KR22 KR24 KR25 KR17 KR26 KR18 KR19 KR23 KR28 KR29 KR30 KR21 BKU1B BKU5 BKU8 BKU16 BKU17 BKU33 BKU34 BKU15 BKU22 BKU27 BKU2B BKU13 BKU35 BKU32 BKU19 BKU31 BKU21 BKU30 BKU35 BKU9B BKU23 BKU1B BKU7 BKU14 BKU5B BKU25B BKU26B WPK3 WPK4 WPK9 WPK10 WPK5 WPK20 WPK7 WG17 WPS15 WB1 WB2 WB3 WPK13 WPK15 WG1 WG10 WG12 WG18 WG9 WPS17 WG4 WG6 WPS14 WPS20 WG3 WG7 WPK24 WPS19 WPK18 WPK19 WPK2 WPK8 WPK11Secara keseluruhan, ke empat populasi manggis tersebut menunjukkan jumlah klonal yang sangat tinggi. Hampir semua tanaman sampel (98.2%) merupakan individu klonal. Hubungan antar populasi menunjukkan individu klonal tersebut mengelompok berdasarkan daerah geografinya. Urutan koefisien kemiripan populasi adalah Kerinci > Bulukumba > Purwakarta > Tembilahan (Ganbar 25). Tingginya kemiripan genetik (0.83-1.00) serta pengelompokan berdasarkan geografi menunjukkan bahwa tanaman manggis dibudidayakan secara tradisional melalui perbanyakan klonal dengan menggunakan tanaman induk dari daerah setempat.
Diversitas klonal dapat terjadi karena mutasi menghasilkan klon satelit yang mungkin terdiri dari mutasi normal (substitusi, penyusunan kembali dan autosegregasi) (Richards 2003). Diversitas klonal yang tinggi juga dijumpai pada Psammochloa villosa, tanaman klonal yang diperbanyak secara cepat melalui rizoma. Pada tanaman ini dideteksi adanya 98 klon diantara 157 individu menggunakan 12 primer ISSR (Li & Ge 2001).
Variasi genetik pada tanaman apomiksis berasosiasi dengan tingkat ploidi (Bhat et al. 2005). Otto (2007) menyatakan bahwa poliploidisasi adalah penambahan set kromosom lengkap pada genom dan merupakan sumber mutasi yang paling besar. Poliploid mengalami lebih banyak deleterious mutasi daripada diploid. Adams & Jonathan (2005) menyatakan bahwa poliploidi merupakan proses yang lama dan berulang, menyebabkan kehilangan gen berbeda, perubahan genome yang cepat, dan pengaruh ekstensif terhadap ekspresi gen. Variasi genetik pada apomiktik poliploid obligat telah dilaporkan pada Taraxacum menggunakan isozim esterase pada rata-rata 22% progeninya. Informasi ini menggambarkan bahwa variasi yang muncul melalui apomiksis terjadi dengan kecepatan yang lebih besar dari mutasi (Ford & Richards 1985; Hughes & Richards 1985).
Wendel (2000) menjelaskan bahwa pada beberapa poliploid yang baru terbentuk genomnya tidak stabil dan mengalami penyusunan kembali secara cepat (rapid reparterning). Mansyah et al. (2008) menyatakan adanya variasi genetik yang cukup besar antara antara pohon induk manggis dan keturunannya. Berdasarkan hasil Analisis RAPD pada satu tanaman induk manggis dengan 18 individu keturunannya hanya 4 individu (22%) yang identik dengan induknya.
Empat belas tanaman lainnya (78%) berbeda secara genetik. Paun & Hörandl (2006) melaporkan variasi alel yang ekstensif melalui mutasi di dalam keturunan aseksual hexaploid R. carpaticola, yang merupakan sumber penting variasi klonal.
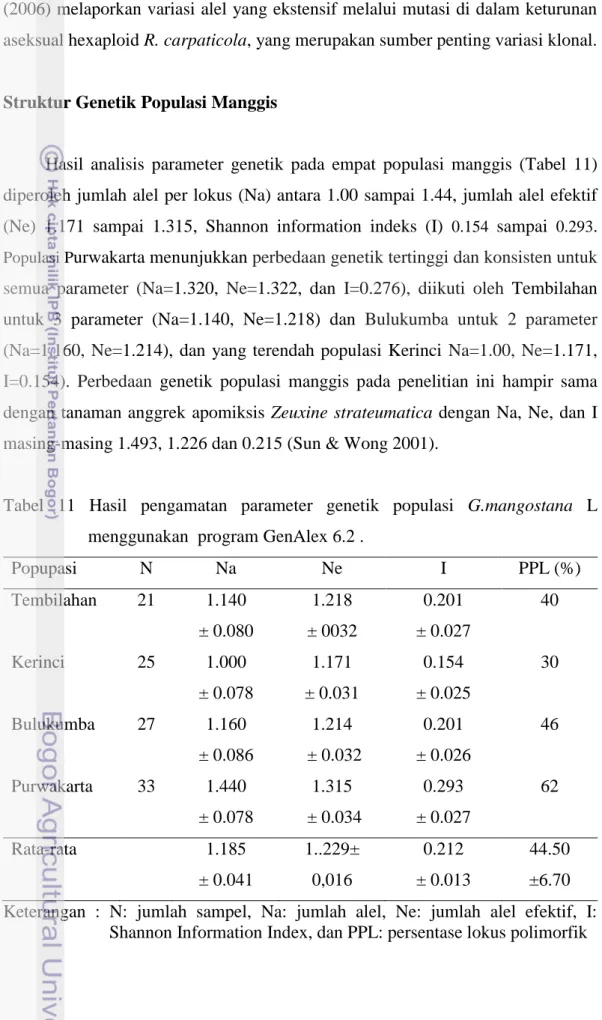
Struktur Genetik Populasi Manggis
Hasil analisis parameter genetik pada empat populasi manggis (Tabel 11) diperoleh jumlah alel per lokus (Na) antara 1.00 sampai 1.44, jumlah alel efektif (Ne) 1.171 sampai 1.315, Shannon information indeks (I) 0.154 sampai 0.293. Populasi Purwakarta menunjukkan perbedaan genetik tertinggi dan konsisten untuk semua parameter (Na=1.320, Ne=1.322, dan I=0.276), diikuti oleh Tembilahan untuk 3 parameter (Na=1.140, Ne=1.218) dan Bulukumba untuk 2 parameter (Na=1.160, Ne=1.214), dan yang terendah populasi Kerinci Na=1.00, Ne=1.171,
I=0.154). Perbedaan genetik populasi manggis pada penelitian ini hampir sama dengan tanaman anggrek apomiksis Zeuxine strateumatica dengan Na, Ne, dan I masing-masing 1.493, 1.226 dan 0.215 (Sun & Wong 2001).
Tabel 11 Hasil pengamatan parameter genetik populasi G.mangostana L menggunakan program GenAlex 6.2 .
Popupasi N Na Ne I PPL (%) Tembilahan 21 1.140 ± 0.080 1.218 ± 0032 0.201 ± 0.027 40 Kerinci 25 1.000 ± 0.078 1.171 ± 0.031 0.154 ± 0.025 30 Bulukumba 27 1.160 ± 0.086 1.214 ± 0.032 0.201 ± 0.026 46 Purwakarta 33 1.440 ± 0.078 1.315 ± 0.034 0.293 ± 0.027 62 Rata-rata 1.185 ± 0.041 1..229± 0,016 0.212 ± 0.013 44.50 ±6.70 Keterangan : N: jumlah sampel, Na: jumlah alel, Ne: jumlah alel efektif, I:
Persentase lokus polimorfik (PPL) tertinggi juga ditunjukkan oleh populasi Purwakarta (62%.) diikuti oleh Bulukumba (46%), Tembilahan (40%) dan Kerinci (30%) (Tabel 11). Variasi PPL di dalam dan antar populasi dengan marka RAPD pada tanaman apomiktik juga dijumpai pada tujuh populasi Hypericum perforatum L. Tiga populasi mempunyai tingkat keragaman yang rendah (PPL 47.6 - 61%) dan empat populasi lainnya lebih tinggi dengan PPL 69.5 – 85.4% (Pilepic et al. 2008). Studi pada tiga spesies anggrek dengan sistem reproduksi berbeda menunjukkan PPL yang berbeda. Spesies Zeuxine Strateumatica dengan reproduksi apomiksis memiliki PPL terendah (49.25%), Z. gracilis (self compatible) 53.25% dan Eulophia sinensis (menyerbuk silang)menunjukkan PPL tertinggi yaitu 79.38% (Sun & Wong 2001).
AMOVA pada Tabel 12 menunjukkan bahwa manggis mempunyai struktur populasi dengan perbedaan genetik di dalam populasi yang sama besarnya dengan perbedaan genetik antar populasi yaitu sebesar 50%, yang berarti bahwa laju perubahan genetik didalam populasi sama dengan antar populasi AMOVA pada Psammochloa villosa, tanaman klonal yang diperbanyak secara cepat melalui produksi rhizoma menunjukkan variasi genetik yang besar (87.46%) antar populasi dan hanya 12.54% antar individu dalam populasi. Faktor utama yang bertanggung jawab terhadap tingginya perbedaan antar populasi dan rendahnya perbedaan dalam populasi karena tanaman ini mempunyai kapasitas untuk reproduksi seksual melalui penyerbukan sendiri (Li & Ge 2001).
Tabel 12 Hasil analisis varians molekuler (AMOVA) populasi manggis.
Sumber keragaman db JK JKT Varians Persentase
Antar Populasi 3 441.481 147.160 5.400 50%
Dalam Populasi 102 544.425 5.337 5.337 50%
Total 105 985.906 10.737 100%
db = derajat bebas, JK= Jumlah Kuadrat, JKT= Jumlah Kuadrat Tengah
Struktur genetik yang membentuk populasi dipengaruhi terutama oleh sistem reproduksinya. Richards (1997) menjelaskan bahwa tiga model dasar
reproduksi yaitu outbreeding, inbreeding dan apomixis mempunyai perbedaan konsekuensi pada genetika populasi. Sebagai contoh penelitian pada tiga spesies anggrek menunjukkan bahwa kolonisasi pada Z. strateumatica (apomiktik) menghasilkan perbedaan antar populasinya yang sangat tinggi (0.924) dibandingkan dengan Z. gracilis (self compatible) yaitu 0.539 dan E. Sinensis (menyerbuk silang) sebesar 0.653. Sangat tingginya perbedaan genetik atau kurangnya gene flow antar populasi Z. strateumatica konsisten dengan sifat apomiktik dan kolonisasi. Apomixis mencegah rekombinasi seksual dalam populasi dan gene flow melalui pollen antar populasi sehingga menghasilkan variasi genetik yang rendah dalam populasi dan variasi genetik yang tinggi antar populasi. Pola struktur populasi pada Z. strateumatica sesuai dengan teori sistem reproduksi dan kolonisasi(Sun & Wong 2001).
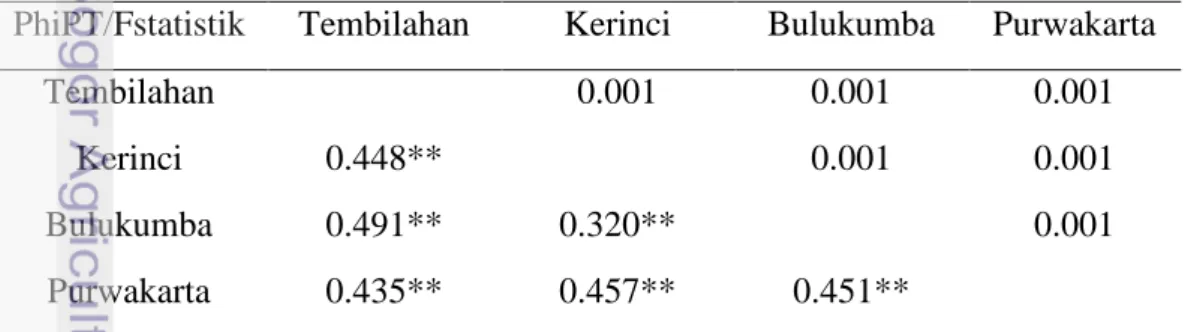
Data pada Tabel 13 memberikan informasi bahwa walaupun variasi didalam dan antar populasi sama besarnya, tiap pasangan populasi menunjukkan perbedaan yang nyata. Pasangan nilai diversitas genetik antar populasi (PhiPT) dan statistik ujinya (P_value) ditampilkan pada Tabel 13. Ke enam pasangan populasi menunjukkan perbedaan yang signifikan (P<0.001), yang berarti bahwa struktur genetik tiap populasi berbeda nyata. Pasangan populasi Kerinci dan Bulukumba menunjukkan nilai perbedaan antar populasi terendah (PhiPT=0.320) dan pasangan populasi Bulukumba dan Tembilahan mempunyai perbedaan genetik tertinggi tertinggi (PhiPT= 0.491).
Tabel 13 Pasangan nilai PhiPT dan uji statistik antar populasi G. mangostana . PhiPT/Fstatistik Tembilahan Kerinci Bulukumba Purwakarta
Tembilahan 0.001 0.001 0.001
Kerinci 0.448** 0.001 0.001
Bulukumba 0.491** 0.320** 0.001
Purwakarta 0.435** 0.457** 0.451**
Keterangan: Angka di bawah diagonal adalah nilai PhiPT dan di atas diagonal adalah nilai statistik uji berdasarkan 999 permutasi. * P < 0.05 ** P< 0.01
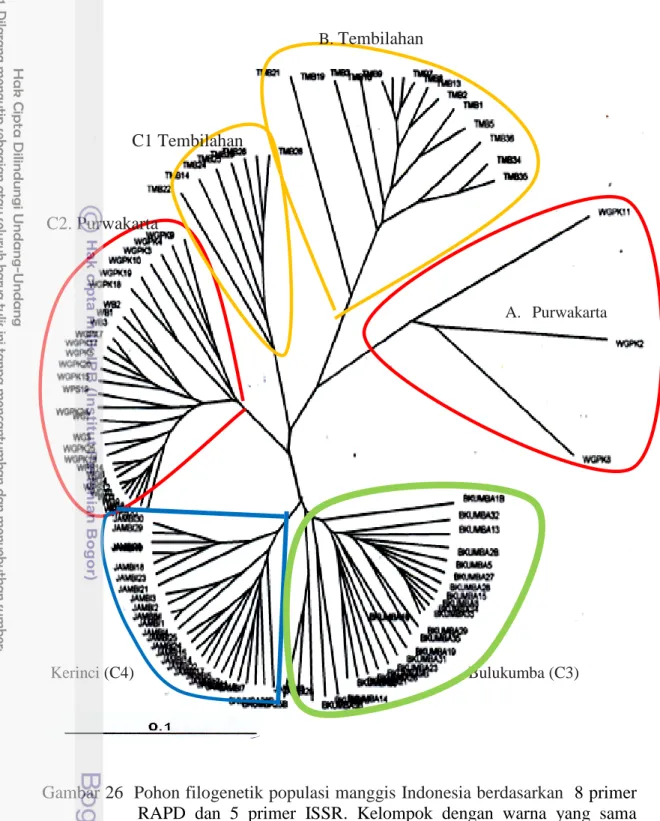
Analisis Filogenetik
Pohon filogenetik pada Gambar 26 dapat dijelaskan melalui pasangan jarak genetik dan identitas genetiknya (Tabel 14). Jarak genetik adalah ukuran yang menyatakan kedekatan hubungan genetik antara dua populasi, sedangkan identitas genetik merupakan nilai yang menyatakan tingkat kemiripan genetik antar dua populasi. Jarak genetik (D) antar populasi manggis pada penelitian ini berkisar antara 0.079 – 0.164 dengan identitas genetik yang tinggi (Nei I=0.849-0.924). Jarak genetik antara populasi manggis Bulukumba dan Kerinci (D=0.079) merupakan yang terdekat dengan identitas genetik tertinggi (Nei I=0.924) Sebaliknya jarak genetik antara populasi Bulukumba dan Tembilahan merupakan yang terjauh (D=0.164) dengan identitas genetik terendah (Nei I=0.849).
Populasi Purwakarta konsisten menunjukkan tingkat perbedaan genetik yang terbesar untuk semua parameter genetik yang diamati dan terbagi menjadi dua kelompok genetik berbeda yaitu kelompok A dan C2. Pemisahan kedua kelompok ini berdasarkan perbedaan pola pita DNAnya (Lampiran 7). Sumber keragaman genetik populasi Purwakarta dapat disebabkan oleh mutasi dan adanya percampuran genetik oleh migrasi tanaman dari daerah lain. Tiga aksesi pada kelompok A diduga berasal dari daerah lain yang berkontribusi dalam meningkatkan variasi genetik dalam populasi. Kedua kelompok genetik tersebut sulit dibedakan secara morfologi.
Populasi Tembilahan terpisah kedalam 2 kluster (B dan C1) berdasarkan pita pita spesifik (Lampiran 6) dan karakter morfologinya (Gambar 7c, 9, 13c, 16b, 18c, 20c). Populasi Tembilahan pada kluster B lebih dekat jarak genetiknya dengan tiga genotipe unik Purwakarta (Kluster A), sedangkan populasi Tembilahan pada kluster C1 lebih dekat dengan populasi Purwakarta pada kluster C2. Populasi Tembilahan memiliki genotipe yang unik tetapi perbedaan genetiknya lebih rendah dari Purwakarta dan Bulukumba yang dapat dilihat dari parameter genetiknya. Populasi Tembilahan memiliki jumlah alel efektif dan PPL yang lebih rendah daripada Purwakarta (Tabel 11). Kondisi ini berkemungkinan disebabkan karena genotipe unik tersebut merupakan hasil introduksi dari daerah lain ke Tembilahan. Amsellem et al. (2000) menyatakan bahwa secara teoritis adanya kolonisasi tanaman pada daerah baru akan mengurangi diversitas genetik populasi awal dan perbedaan genetik antar populasi.
B. Tembilahan C1 Tembilahan C2. Purwakarta A. Purwakarta Kerinci (C4) Bulukumba (C3)
Gambar 26 Pohon filogenetik populasi manggis Indonesia berdasarkan 8 primer RAPD dan 5 primer ISSR. Kelompok dengan warna yang sama berasal dari lokasi yang sama.
Perbedaan geografi kelihatannya tidak mencerminkan jarak genetik antar populasi. Sebagai contoh populasi Kerinci dan Bulukumba yang dipisahkan oleh pulau dan lautan mempunyai jarak genetik yang dekat (0.079), sedangkan populasi Kerinci dan Tembilahan dengan jarak geografi yang dekat mempunyai jarak genetik yang relatif lebih jauh (0.151). Tidak adanya hubungan antara jarak
geografi dengan jarak genetik ini juga terjadi pada Gentianella germanica, species tanaman berumur pendek dan selfcompatible dari famili Gentianaceae (Fischer & Matthies 1998).
Tabel 14. Pasangan jarak genetik Nei (D) dan identitas genetik (Nei I) pada 4 populasi manggis
Tembilahan Kerinci Bulukumba Purwakarta
Tembilahan 0.151 0.164 0.148
Kerinci 0.860 0.079 0.137
Bulukumba 0.849 0.924 0.132
Purwakarta 0.862 0.872 0.876
D (diatas diagonal) dan Nei I (dibawah diagonal)
Kladogram pada Gambar 27 menjelaskan hubungan antara populasi berdasarkan sejarah evolusinya. Kladogram berakar pada tiga genotipe unik dari Purwakarta (Kelompok A), diikuti oleh kelompok Tembilahan (grup B), Tembilahan (grup C1), Purwakarta (C2), Kerinci (C3) dan Bulukumba (C4). Konsisten dengan analisis kluster (Gambar 28) dan filogenetik (Gambar 29) kladogram juga menghasilkan tiga kluster utama. Dua kluster utama (A dan B) mewakili daerah dengan individu yang sangat berbeda secara genetik dengan daerah lainnya. Kluster ke tiga (grup C) pada kladogram terdiri dari individu tiap populasi yang mengelompok bersama sama berdasarkan daerah geografinya.
Kladogram juga memisahkan populasi Purwakarta dan Tembilahan menjadi dua kelompok genetik berbeda. Ke dua kelompok dalam lokasi Purwakarta dan Tembilahan tersebut tidak dipisahkan oleh habitat yang nyata dengan jarak sampel 5 m sampai 1 km, tetapi dipisahkan oleh pemilik yang berbeda. Keragaman genetik serupa juga dijumpai pada klon ubi kayu seperti yang dilaporkan oleh Elias et al. 2000). Petani menemukan varietas baru melalui pertukaran material tanaman di dalam komunitas atau dengan desa lainnya. Penemuan ini membuktikan bahwa variabilitas genetik di dalam dan antar populasi terjadi melalui introduksi, yang ditunjukkan oleh diversitas morfologi dan molekuler.
C4 BULUKUMBA C3 KERINCI C2. PURWAKARTA C1 TEMBILAHAN B.TEMBILAHAN A.PURWAKARTA Gambar 27 Kladogram populasi manggis yang dikonstruksi dari 8 primer RAPD
dan 5 primer ISSR.
BKU1B BKU32 BKU13 BKU2B BKU5 BKU27 BKU34 BKU26 BKU15 BKU9 BKU34 BKU33 BKU16 BKU17 BKU28 BKU35 BKU19 BKU31 BKU23 BKU9B BKU26 BKU21 BKU30 BKU14 BKU7 BKU5B BKU25B BKU26B KR 26 KR13 KR7 KR10 KR11 KR12 KR5 KR17 KR22 KR14 KR15 KR24 KR25 KR4 KR1 KRM KR2 KR3 KR21 KR23 KR18 KR19 KR28 KR29 KR30 WPS20 WPS17 WG8 WG9 WG1 WG10 WPS12 WG4 WG5 WPS14 WPK13 WPK25 WG3 WPG7 WPK24 WPS19 WPK15 WPK20 WPK5 WPK17 WPK7 WB3 WB1 WB2 WP18 WPK19 WPK10 WPK3 WPK4 WPK9 TMB22 TMB14 TMB24 TMB25 TMB29 TMB28 TMB26 TMB21 TMB19 TMB3 TMN10 TMB9 TMB7 TMB8 TMB13 TMB2 TMB1 TMB5 TMB36 TMB34 TMB35 WPK11 WPK2 WPK8 C
Kasus ini juga terjadi pada spesies tanaman berkayu terutama yang menyerbuk sendiri atau apomiksis yang diintroduksi oleh manusia ke daerah baru (Husband & Barrett 1991). Diversitas genetik melalui seed dispersal telah dilaporkan pada apomiktik Cratageous douglasii sehingga terjadi percampuran gene pool antara populasi dan meningkatkan diversitas lokal (Van Dijk 2003). Hasil penelitian ini menunjukkan bahwa pola struktur genetik populasi berhubungan dengan sejarah gene flow oleh penyebaran materi perbanyakan tanaman karena aktifitas manusia (Elias et al. 2000).
Berdasarkan pohon filogenetik dan kladogram dapat disimpulkan bahwa pembentukan populasi manggis diawali dari Tembilahan, kemudian menyebar ke Purwakarta, Kerinci dan Bulukumba. Secara geografi populasi tersebut dipisahkan oleh pulau dan lautan melintasi garis Wallaceae, namun tetap memungkinkan terjadinya pertukaran material tanaman dengan perantaraan manusia. Studi filogenetik menggunakan ITS (internal transcribed spacer region) menunjukkan bahwa kebanyakan spesies Garcinia dari Timur garis Wallace mengelompok di dalam spesies dari Barat. Hal ini mencerminkan penyebaran spesies melintasi daerah biogeografi dari barat ke Timur ketika dataran dataran Sahul dan Sunda bersatu, kira-kira 20 juta tahun yang lalu. Hipotesis ini dikuatkan oleh perkiraan penyebaran aksesi G. rigida di Timur garis Wallace' dari nenek moyangnya di sebelah Barat garis Wallaceae kira- kira 2l.58±2.90 juta tahun lalu (Nazre 2006)
Hasil penelitian ini memberikan gambaran bahwa spesies Garcinia mangostana yang relatif sukar dibedakan tetap mempunyai struktur genetik tersendiri. Hasil yang sama diketahui pada spesies tanaman endemik Astragalus bibullatus (Fabaceae) di Tennessee Georgia. Tanaman ini mirip secara genetik yang ditunjukkan oleh identitas genetik yang tinggi (0.976 – 0.991), variasi antar populasi 10%, dan 28% marka polimorfik yang terbagi menjadi 2 atau 3 kelompok genetik (Baskauf & Burke 2009).
Selain mengungkapkan stuktur genetik tanaman manggis, penelitian ini juga memberikan informasi adanya beberapa klon lokal yang merupakan sumber keragaman genetik tanaman manggis. Keberadaan klon-klon ini merupakan interaksi antar manusia, faktor ekologi, dan faktor budidaya, seperti yang terjadi pada cassava (Elias et al. 2000). Purwakarta dan Tembilahan merupakan pusat
keragaman sumberdaya genetik manggis yang penting di Indonesia. Informasi ini dapat digunakan untuk menyusun strategi penanganan plasma nutfah manggis dan konservasinya.
Kesimpulan
1 Populasi manggis Indonesia mempunyai koefisien kemiripan genetik sebesar 0.82-1.00 dan terdiri dari tiga kelompok genetik yang terpisah berdasarkan lokasi geografi.
2 Struktur genetik populasi manggis menunjukkan bahwa perbedaan genetik antar populasi sama dengan perbedaan genetik dalam populasi yaitu sebesar sebesar 50% yang merupakan ciri tanaman klonal dan apomiktik.
3 Semua parameter genetik populasi (jumlah alel, Shannon Information index, dan jumlah lokus polimorfik) menunjukkan bahwa populasi Purwakarta mempunyai perbedaan genetik tertinggi dan yang terendah pada populasi Kerinci.
4 Hubungan antar populasi menunjukkan bahwa pasangan populasi Tembilahan dan Bulukumba mempunyai perbedaan genetik terbesar dan identitas genetik terendah. Sebaliknya pasangan populasi Kerinci dan Bulukumba menunjukkan perbedaan genetik terkecil dan identitas genetik tertinggi.
5 Populasi Purwakarta dan Tembilahan masing-masing terbagi menjadi dua kelompok genetik berbeda yang menunjukkan kedua daerah tersebut memiliki klon lokal yang berpotensi untuk dikembangkan lebih lanjut.
6 Pembentukan populasi manggis diawali dari wilayah Tembilahan, kemudian menyebar ke Purwakarta, Kerinci dan Bulukumba.
Daftar Pustaka
Adams K, Jonathan FW. 2005. Polyploidy and genome evolution in plants. Curr. Opinion Plant Biol. 8: 135–141.
Amsellem L, Noyer JL, Bourgeois TL Hossaert-Mickey M. 2003. Comparison of genetic diversity of the invasive weed Rubus alceifolius Poir. (Rosaceae) in its native range and in areas of introduction, using amplified fragment length polymorphism (AFLP) markers. Molec. Ecol. 9: 443–455
Baskauf CJ, Burke JM. 2009. Population Genetics of Astragalus bibullatus (Fabaceae) Using AFLPs. J. Heredity 100(4):424–431
Bennetzen JL. 2000. Transposable element contributions to plant gene and genome evolution. Plant Mol. Biol. 42: 251-269.
Bennetzen JL. Kellogg EA. 1997. Do plants have a one-way ticket to genomic obesity? Plant Cell. 9: 1509-1514.
Bhat V, Dwivedi KK, Khurana JP, Sopory SK. 2005. Apomixis: An enigma with potential applications. Curr. Sci. 89(11): 1879-1893.
[BPS] Badan Pusat Statistik. 2009. Statistik Pertanian Indonesia 2009. Jakarta. Ellstrand NC, Roose ML 1987. Patterns of genotypic diversity in clonal plant
species. A. J. Bot. 74: 121–131.
Ellias M, Panaud O, Robert T. 2000. Assessment of genetic variability in a traditional cassava (Manihot esculenta Crantz) farming system using AFLP markers. Heredity 85: 219-230.
Fischer M, Matthies D. 1998. RAPD variation in relation to population size and plant fitness in the rare Gentianella Germanica (Gentianaceae) Amer. J. Bot. 85(6): 811–819.
Ford H, Richards AJ. 1985. Isozyme variation within and between Taraxacum agamospecies in a single locality. Heredity 55:289–291.
Gonzalez A, Wong A, Delgado-Salinas A, Papa R, Gepts P. 2005. Assessment of Inter simple sequence repeat markers to differentiate sympatric wild and domesticated Populations of Common Bean. Crop Sci. 45: 606–615.
Holsinger KE, Mason-Gamer RJ. 1996. Hierarchical analysis of nucleotide diversity in geographically structured populations. Genetics 142 629-639
Hong X, Scofield DG, Lynch M. 2006. Intron size, abundance, and distribution within untranslated regions of genes. Mol. Biol. Evol. 23:2392-404.
Hughes J, Richards AJ. 1985. Isozyme inheritance in diploid Taraxacum Hybrids. Heredity 54:245–249.
Husband BC, Barrett SCH. 1991. Colonisation history and population genetic structure of Eichornia paniculata in Jamaica. Heredity. 66: 287–296.
Kapitonov VV, Jurka J. 1999. Molecular paleontology of transposable elements from Arabidopsis thaliana. Genetica. 107: 27-37.
Li A, Ge S. 2001. Genetic variation and clonal diversity of Psammochloa villosa (Poaceae) detected by ISSR Markers. Ann. Bot. 87: 585-590.
Lim AL. 1984. The embryology of Garcinia mangostana L. (Clusicaeae). Garden Bulletin Singapore. 37: 93–103.
Liu B, Wendel JF. 2003. Epigenetic phenomena and the evolution of plant allopolyploids. Mol. Phylogenet. Evol. 29:365-379.
Mackanzie S. 2005. The Mitochondrial Genome of Higher Plants: a target for natural adaptation. p: 78-88. In: Robert J. Henry (Ed): Plant Diversity and Evolution: Genotypic and Phenotypic Variation in Higher Plants. CABI Publishing, Cambridge, USA.
Mansyah E, Baihaki A, Setiamihardja R, Darsa JS, Sobir, Poerwanto R (2003b). Analisis variabilitas genetik manggis (Garcinia mangostana L.) di Jawa dan Sumatera Barat menggunakan teknik RAPD. Zuriat 4(1): 35-44.
Mansyah E, Prasetyo BW, Jawal MAS, Rusdianto U, Muas I. 2005. Manggis unik dari Tembilahan. Warta Penelitian dan Pengembangan Pertanian. 27(2): -8.
Mansyah E, Santoso PJ, Muas I, Sobir. 2008. Evaluation of genetic diversity between and within mangosteen (Garcinia mangostana) trees. 4th International Symposium on Tropical and Subtropical Fruits. November 3– 7. Bogor, West Java. Indonesia (In press)
Mao L, Wood TC, Yu Y, Budiman MA, Tomkins J., Woo S, Sasinowski M., Presting G, Frisch D, Goff S, Dean RA, Wing RA. 2000. Rice transposable elements: a survey of 73,000 sequence-tagged-connectors. Genome Res. 10: 982-990.
Mishra PK, Fox RTV, Culham A. 2003. Inter-simple sequence repeat and aggressiveness analyses revealed high genetic diversity, recombination and long-range dispersal in Fusarium culmorum. School of Plant Sciences, The University of Reading, Whiteknights, Reading RG6 6AS, UK..
Oostrum HV,. Sterk AA, Wijsman HJW. 1985. Genetic variation in agamospermous microspecies of Taraxacum sect. Erythrosperma and sect obliqua. Heredity 55:223–228.
Otto SP. 2007. The evolutionary Consequences of Polyploidy. Cell. 131: 452-462
Paun O. Ho¨randl E. 2006. Evolution of Hypervariable Microsatellites in Apomictic Polyploid Lineages of Ranunculus carpaticola: Directional Bias at Dinucleotide Loci. Genetics 174: 387–398.
Pilepic KH, Males Z, Plazibat M. 2008. Population structure in Hypericum perforatum. L., a hybrid apomictic plant species of medicinal importance,
was studied using RAPD markers. Periodicum Biologorum 110 (4): 367– 371.
Ramage CM, Sando L, Peace CP, Caroll BJ, Drew RA. 2004. Genetic diversity revealed in the apomictic fruit species Garcinia mangostana L. (mangosteen). Euphytica. 136(1):1-10.
Richards AJ. 1997. Plant Breeding Systems. Second Edition. Departemen of Agricultural and Environtmental Science University of Newcastle Upon Tyne. Chapman and Hall. London. 529 pp.
Richards AJ. 2003. Apomixis in flowering plants: an overview. Phil. Trans. R. Soc. Lond. B (2003) 358: 1085–1093.
Sabar. 2005. Kebijakan Departemen Perdagangan di bidang ekspor buah-buahan. Temu Pelaku Agribisnis Mangga dan Manggis. Bandung, 29-30 November 2005. 9 hal.
Sinaga S, Sobir, Poerwanto, Aswidinnoor H, Duryadi D. 2007a. Genetic variability analysis on apomictic mangosteen (Garcinia mangostana) in Indonesian and its close related species by using RAPD markers. Floribunda 3(4): 77 – 83.
Sinaga S. 2008. Analisis Keanekaragaman genetik dan fenotip manggis (Garcinia mangostana L.) dan kerabat dekatnya. Disertasi. Sekolah Pasca sarjana.Institut Pertanian Bogor.
Sulassih. 2011. Analisis hubungan kekerabatan manggis (Garcinia mangostana l. ) menggunakan penanda morfologi, dan molekuler (ISSR) terhadap kerabat liarnya. Thesis. Sekolah Pasca Sarjana. Institut Pertanian Bogor. 75 hal. Sun M, Wong KC. 2001. Genetic structure of three orchid species with
constrating breeding system using RAPD and allozyme markers. Amer. J. Bot. 88(12): 2180–2188.
Van Der Hulst RGM, Mes THM, Falque M, Stam P, Den Nijs JCM, Bachmann K 2003. Genetic structure of a population sample of apomictic dandelions. Heredity 90: 326–335
van Dijk PJ. 2003. Ecological and evolutionary opportunities of apomixis: insights from Taraxacum and Chondrilla Phil. Trans. R. Soc. Lond. B 358, 1113–1121.
Wegscheider E, Benjak A, Forneck A. 2009. Clonal variation in Pinot noir revealed by S-SAP involving universal retrotransposon-based sequences Am. J. Enol. Vitic. 60:1:104-109.
Wright SI, Brandon SG. 2005. Molecular Population Genetics and the Search for Adaptive Evolution in Plants. Mol. Biol. Evol. 22(3):506–519.
Zietkiewicz E, Rafalski A, Labuda D. 1994. Genome finger printing by Simple Sequence Repeats (SSR)-anchored polymerase chain reaction amplification. Genomics 20: 176-183.