BAB II. TINJAUAN PUSTAKA
2.1 Biologi ulat sutera Bombyx mori L. (Lepidoptera: Bombycidae)
Ulat sutera termasuk kelompok serangga holometabola, yaitu hewan yang
mengalami metamorfosis sempurna. Hal ini berarti bahwa setiap generasi
me-lewati 4 stadia yaitu telur, larva, pupa dan imago. Selama metamorfosis, stadia
ulat adalah masa ulat makan. Masa ini merupakan masa yang sangat penting untuk
sintesis protein sutera dan pembentukan telur (Atmosoedarjo et al. 2000). Ulat
sutera mempunyai lebih dari 4000 strain. Ulat sutera B. mori berasal dari tetua B.
mandarina yang ditemukan di pohon murbei Cina, Jepang dan negara lain di Asia
Timur (Jingade et al. 2011).
Ulat sutera didomestikasi sudah cukup lama, menyebabkan kehilangan
ke-mampuan untuk hidup mandiri di alam bebas. Indra penciuman sudah sangat
ti-dak berfungsi, titi-dak mengenal lagi tanaman murbei dalam jarak beberapa
senti-meter, dan tidak dapat bergerak dari batang ke batang lain untuk mendapatkan
daun, karena kemampuan mobilitasnya sudah lemah. Daya pegangnya juga sangat
lemah, sehingga tidak mampu mempertahankan diri dari goncangan batang oleh
angin, atau oleh sebab-sebab lain. Ulat sutera sudah tidak dapat lagi melindungi
diri dari musuh dan tidak bisa bergerak cepat. Ngengatnya tidak bisa terbang
untuk berkopulasi dan sulit bertelur di daun murbei (Atmosoedarjo et al. 2000).
Setelah dipelihara selama beberapa ribu tahun di berbagai negara, maka ulat
sutera mempunyai banyak varietas yang diklasifikasikan berdasarkan sifat, yang
berhubungan erat dengan ekonomi dan teknik pemeliharaan sebagai berikut (1)
klasifikasi berdasarkan negara asal yaitu ras Cina, Jepang, Eropa, dan Tropis, (2)
polivoltin, dan (3) klasifikasi berdasarkan frekuensi ganti kutikula, yaitu three
moulter, four moulter, dan five moulter (Guntoro 1994).
2.1.1 Morfologi ulat sutera
Bentuk tubuh ulat terbagi menjadi 3 bagian utama, yaitu kepala, toraks, dan
abdomen (Gambar 2.1). Ulat sutera yang biasa dipelihara mempunyai bintik hitam
kecoklatan yang disebut bintik mata.
Gambar 2.1 Ulat sutera (Bombyx mori) instar V; A. thorax, B. segmen abdomen C. Crescent, D. Eye spots (mata), E. head, F. caudal, G. thorax legs, H. spiracles, I. stars spots, J. abdominal legs dan K. caudal legs
(Komoeniczak 2010)
a. Kepala
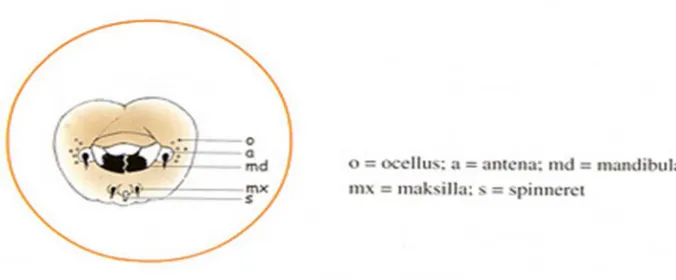
Kepala ulat sutera meskipun kecil memiliki struktur yang kompleks. Di
bagian kepala terdapat sepasang antena berfungsi sebagai alat indera. Antena
terdiri dari 3 ruas pendek. Di pangkal antena terdapat sepasang mandibula
(ra-hang) bersebelahan yang bergerak ke sisi untuk mengigit dan mengunyah daun
murbei. Daun yang digigit masuk melalui bagian mulut. Satu lingkaran kecil di
tengah di bawah mulut terdapat spinneret (Gambar 2.2). Spineret adalah saluran
tempat keluarnya filamen sutera. Filamen sutera keluar dari lubang yang ada di
tampak seperti antena kecil disebut maksila yaitu indera perasa yang berfungsi
untuk mengidentifikasi pakan. Disisi yang lain di dasar antena, terdapat 6 pasang
lingkaran kecil berbentuk setengah bulatan, yaitu oseli atau mata tunggal. Oseli
atau mata tunggal, tidak mengenal bentuk dari objek yang nampak, tetapi hanya
bisa melihat antara terang dan gelap. Untuk lebih jelasnya dapat dilihat pada
Gambar 2.2 di bawah ini:
Gambar 2.2 Kepala ulat sutera dilihat dari depan (Omura 1980)
Ulat makan terus menerus apabila terdapat komponen gula (glukosida) pada
daun murbei (Nayar dan Fraenkel 1962). Respon makan dipengaruhi oleh jumlah
atau rasio dari stimulan makan. Interaksi sinergis maupun antagonis dari
campur-an rcampur-angscampur-angcampur-an kimia merupakcampur-an penentu ycampur-ang scampur-angat mempengaruhi palatabilitas
atau respon palatum terhadap pakan (Ishikawa et al. 1969).
Daun murbei hibrida untuk pakan tidak berpengaruh secara nyata terhadap
persentase kematian larva instar I-V, berat kokon, berat kulit kokon, persentase
kulit kokon, panjang filamen, berat filamen, persentase filament dan daya gulung
tetapi nyata berpengaruh terhadap rendemen pemeliharaan. Secara umum semua
jenis murbei dapat digunakan untuk pakan ulat sutera (Pudjiono dan Na’iem
2007). Pada saluran pencernaan B. mori terdapat senyawa bersifat alkalin dan
Jumlah bakteri selulolitik dalam saluran pencernaan meningkat pada setiap instar
(Anand et al. 2009).
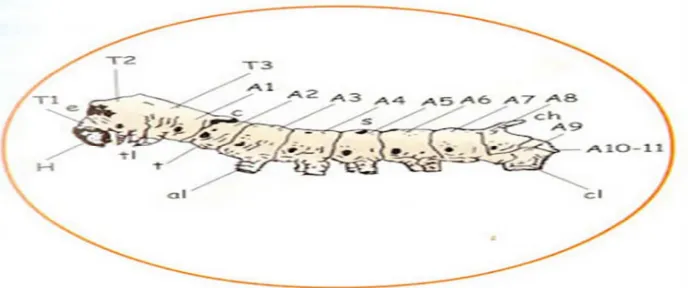
b. Toraks dan abdomen
Tubuh ulat sutera terdiri dari 13 segmen yang terdiri dari satu segmen pada
kepala, tiga segmen toraks dan sembilan segmen abdomen (Gambar 2.3). Pada
bagian paling depan, kepala dibungkus kulit keras yang berwarna hitam
kecok-latan. Selanjutnya ada 3 segmen tersusun berurutan yaitu protoraks, mesotoraks,
dan metatoraks. Sembilan segmen terakhir disebut abdomen. Segmen ke 3,4,5,6
dan segmen terakhir masing-masing mempunyai sepasang kaki, yang disebut kaki
abdomen.
Gambar 2.3 Toraks dan abdomen ulat sutera (Omura 1980)
Kaki abdomen terakhir yang paling besar disebut kaki kaudal. Kaki
abdo-men tidak mempunyai segabdo-men dan disebut proleg, sedangkan tanduk yang ada
pa-da segmen ke-8 disebut tanduk kaupa-dal.
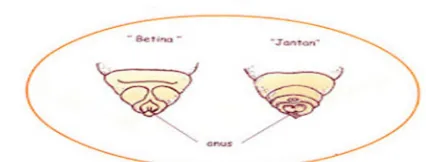
2.1.2 Perbedaan antara ulat jantan dan betina
Pada Gambar 2.4 terlihat perbedaan ulat jantan dan betina. Pada ulat jantan
terdapat satu titik dari kelenjar Herold pada abdomen berbatasan antara segmen
abdomen ke-8 dan 9, sedangkan ulat betina sepasang bintik kecil pada bagian
dan belakang. Waktu yang paling tepat untuk membedakan kedua jenis kelamin
adalah pada awal instar V, tepat setelah molting yang ke-4. Karakteristik ini sulit
dilihat dengan mata biasa, tetapi dapat dibantu dengan menggunakan kaca
pembe-sar atau hanya dapat dilakukan oleh para ahli.
Gambar 2.4 Perbedaan ulat jantan dan betina bagian perut segmen belakang (Omura 1980)
Siklus hidup ulat sutera antara 50 hingga 60 hari termasuk periode
inku-basi, tahapan ulat, pupa, dan imago serta akan berakhir saat setelah imago kawin.
Telur ulat sutera berbentuk kecil, rata, dan elips, dilapisi dengan lapisan keras
(kulit telur). Bentuk dan ukurannya sangat kecil. Ukuran telur berdiameter 1 mm
(Lamarque dan Gros 1945). Pada ujung telur terdapat micropyle yaitu tempat
sperma memasuki sel telur. Telur yang baru menetas berwarna putih susu atau
ku-ning keruh yang terdiri dari warna chorion (kulit telur), serosa dan kuning telur
(komponen dalam isi telur). Setelah hari ke-2 atau ke-3, warna telur mulai
beru-bah, hari ke-6 dan ke-7, warna telur berubah menjadi abu-abu sampai ungu gelap
(Sinchaisri 1993).
Ulat yang baru menetas mempunyai panjang 3 mm, diselimuti oleh
rambut-rambut tipis dan berwarna hitam. Selama stadia ulat mengalami pergantian
kuti-kula sebanyak empat kali. Selama masa pergantian kutikuti-kula, ulat mengalami masa
(moulting). Selama pergantian kutikulapertama, ulat sutera memproduksi kutikula
baru untuk menggantikan kutikula lamanya. Setelah itu ulat kembali makan,
tum-buh dan memasuki instar selanjutnya. Instar I sampai instar III disebut ulat kecil,
sedangkan instar IV dan V disebut ulat besar. Total periode ulat dari penetasan
hingga membentuk kokon yaitu 25 hingga 30 hari (Sinchaisri 1993).
Pergantian kutikula adalah suatu proses yang kompleks dan dikendalikan
oleh hormon-hormon tertentu dalam tubuh. Molting meliputi lapisan kutikula
din-ding tubuh, lapisan kutikula trakea, saluran pencernaan depan dan belakang, serta
struktur endoskeleton. Proses pergantian kutikula serangga dipengaruhi oleh
hor-mon. Hormon merupakan sinyal kimia (chemical signals) atau pembawa pesan
ki-mia (chemical messenger) yang dikirim dari sel dalam bagian tubuh tertentu ke
sel-sel dalam bagian tubuh lainnya pada individu organisme yang sama. Moulting
pada serangga diatur oleh hormon 20-hydroxyecdysone steroids (ecdysterone atau
ecdysteroids, 20HE), JH-sesqui terpenoid, hormon eclosion, dan hormon bursicon
(Klowden 2007).
Proses moulting terdiri dari tiga tahap, yaitu apolysis, ecdysis, dan
skleroti-nisasi. Ketiga tahap tersebut dapat dijelaskan sebagai berikut:
a. Apolysis yaitu pelepasan kutikula lama. Pada tahap ini, hormon moulting
dile-paskan ke dalam hemolimf dan kutikula lama terpisah dari sel epidermis yang
berada di bawahnya. Ukuran epidermis akan meningkat karena mitosis dan
kemudian kutikula baru dihasilkan. Enzim yang disekresikan oleh sel
epider-mis mencerna endokutikula lama.
b. Ecdysis yaitu pembentukan kutikula baru. Tahap ini dimulai dengan pemisahan
kutikula lama, biasanya dimulai pada garis tengah sisi dorsal toraks. Pemisahan
oleh kontraksi otot abdomen yang disebabkan karena serangga menerima udara
atau air. Setelah ini, serangga akan keluar dari kutikula lama.
c. Sklerotinisasi yaitu pengerasan kutikula baru. Kutikula baru yang baru
terben-tuk sangat lembut dan pucat sehingga ini merupakan saat yang sangat rentan
bagi serangga. Dengan demikian, serangga harus melakukan pengerasan (
har-dening) terhadap kutikula baru tersebut. Sklerotinisasi terjadi setelah satu atau
dua jam, dimana eksokutikula akan mengeras dan menjadi gelap. Pada
serang-ga dewasa, sayap akan berkembang karena kekuatan hemolimf yang masuk
melalui vena sayap (Klowden 2007).
Pertumbuhan dan perkembangan serangga diselingi oleh periode moulting
yang diatur oleh steroid 20-hydroxyecdysone (20-HE, hormon moulting,
ecdyste-rone) dan JH sesquiterpenoid (Klowden 2007). Pada tahap dewasa, kedua
hor-mon ini juga terlibat dalam pengaturan pematangan reproduksi (Dhadialla et al.
1997).
Proses pembentukan eksoskeleton yang baru, diawali dengan adanya input
sensorik dari tubuh serangga mengaktifkan sel-sel saraf (neurosecretory cells)
tertentu dalam otak. Sel saraf ini menanggapinya dengan mengeluarkan hormon
otak yang memicu corpora cardiaca untuk melepaskan prothoracicotropic
hor-mone (PTTH) ke dalam sistem peredaran darah. PTTH selanjutnya merangsang
kelenjar prothoracic (prothoracic glands) untuk mengeluarkan hormon moulting,
yaitu ecdysteroids atau 20-hydroxyecdysone steroids (Meyer 2005). Dari sinilah
proses moulting mulai berlangsung, diawali dengan peningkatan titer 20HE dan
diakhiri dengan penurunan titer 20HE dan pelepasan hormon eclosion (Klowden
2007). Peningkatan titer 20HE mengakibatkan epidermis terpisah dari kutikula
eksu-vial), selanjutnya ruang eksuvial diisi oleh cairan molting yang mengandung
en-zim inaktif, chitinolytic (chitinase dan protease) yang mampu mencerna kutikula
lama begitu teraktivasi (Klowden 2007). Sementara itu, sel-sel epidermis
teror-ganisir kembali untuk sintesis sejumlah besar protein serta sekresi epikutikula dan
kutikula baru. Setelah titer 20HE mulai menurun, enzim dalam cairan moulting
di-aktifkan untuk memulai proses pencernaan prokutikula (endokutikula yang tidak
tersklerotisasi). Setelah proses ini selesai, cairan moulting diserap kembali dan
terjadi pengerasan preecdysial kutikula baru (Reynolds 1987).
Apabila titer 20HE menurun ke tingkat basal, kutikula lama terlepas (
ec-dysis) dengan diawali pelepasan crustaceancardioaktive peptide (CCAP), hormon
eclosion, dan ecdysis-triggering hormone, yang bersama-sama bekerja pada
se-jumlah target yang memastikan selesainya proses moulting. Hormon Eclosion
(EH) memulai pelepasan CCAP dari sel-sel ventral ganglion yang menonaktifkan
perilaku preecdysis dan bersama-sama dengan EH mengaktifkan perilaku
ecdy-sis. CCAP bertanggung jawab sebagai motor pemicu dalam menyelesaikan
ecdy-sis. EH juga terlibat bersama hormone bursicon untuk pengerasan kutikula.
Pertumbuhan dan perkembangan serangga diatur oleh 20HE, JH, EH, dan
neuro-hormon lainnya (Klowden 2007).
Perubahan morfologi dan ultra-struktural yang terjadi pada epidermis
se-lama pertumbuhan dan perkembangan serangga tergantung pada pengaturan
eks-presi gen dengan titer yang berbeda dari 20HE dengan ada atau tanpa JH
(Rid-diford 1996). Setiap gangguan dalam homeostasis dari satu atau lebih hormon ini
dengan sumber eksogen dari hormon atau dengan analog sintetis (agonis atau
antagonis) akan mengakibatkan gangguan atau pertumbuhan dan perkembangan
hor-mon-hormon yang terlibat dalam sintesis dan reabsorbsi kutikula akan merugikan
kelangsungan hidup pada tahap perkembangan (Dhadialla et al. 1997).
Saluran pencernaan tengah atau mesenteron adalah bagian utama untuk
pen-cernaan makanan, penyerapan nutrisi dan penyedia substansi barrier. Pada ulat
instar V sangat penting untuk pertumbuhan, perkembangan dan produksi sutera.
Pada mesenteron ditemukan 96 jenis protein dan memproduksi 10 kDa protein
dan prekusor hormon diapause (Zhang et al. 2011).
2.1.3 Tahapan pupa
Sekitar lima atau enam hari ulat mulai membentuk kokon. Ulat sutera
ber-ubah bentuk di dalam kokon menjadi pupa (Gambar 2.5).
Gambar 2.5 Perkembangan proses pembentukan pupa ulat sutera (Atmosoedarjo
et al. 2000)
Pupa berwarna kuning keputihan dan lembek namun secara bertahap akan
mengeras. Periode pupa selama 11 hingga 12 hari (Sinchaisri 1993). Struktur
da-lam dari pupa berbeda dengan ulat atau imago, akan tetapi lebih mendekati imago.
Tubuh pupa histolisis dari organ ulat dan histogenesis dari organ imaginal. Organ
ulat yang mengalami histolisis adalah kelenjar sutera, kaki abdomen, oseli,
ke-lenjar ganti kulit dan tanduk kaudal. Bagian mulut, kaki toraks, organ
pen-cernaan, tubula malpigi, otot dan badan lemak mengalami perubahan besar
di stadia pupa, dan pada stadia ini dapat dibedakan antara jantan dan betina
(Gambar 2.6).
Gambar 2.6 Organ seksual dari pupa (Shimizu dan Tajima 1972)
2.1.4 Tahapan imago
Waktu keluarnya ngengat biasanya terjadi di pagi hari. Ngengat membasahi
kulit kokon dengan sekresi alkalin dan merusak kokon, mendorong kokon hingga
dapat keluar. Ngengat kemudian melebarkan sayap dan mengeringkannya.
Nge-ngat betina kemudian akan membiarkan kelenjar seksual mengembung untuk
me-mikat ngengat jantan (Sinchaisri 1993).
2.2 Ras ulat sutera
Ras ulat sutera unggul memiliki produksi kokon yang tinggi dan dapat
menghasilkan benang sutera dengan kualitas yang baik. Ras ulat sutera yang
se-ring dilakukan budidaya adalah ras Cina, Jepang, Eropa dan Tropika.
Masing-masing ras memiliki keunggulan dan kelemahan yang berbeda. Di Indonesia yang
telah banyak dikembangkan adalah ulat sutera ras Cina, Jepang, dan hasil
persi-langan dari ras Jepang dengan Cina. Ras Cina dan Jepang ini disamping memiliki
keunggulan juga memiliki beberapa kelemahan, seperti kokon yang tipis, tidak
rentan terhadap penyakit dan umur produksi yang panjang. Tetapi dengan
menyi-langkan kedua ras tersebut kelemahan-kelemahannya dapat dikurangi dan sifat
ras dapat dilihat pada Tabel 2.1. Kombinasi kemampuan ras ulat sutera B. morí
seperti ras KA, CC1, CA2, NB4D2 dan NB18 telah disilangkan dan dianalisis.
Analisis karakter kualitas benang masing-masing ras menunjukkan hasil yang
berbeda (Bhargava et al. 1995).
Tabel 2.1 Ciri-ciri dan karakteristik ras ulat sutera
No. Jenis Ras Ciri-ciri dan Karakteristik
1 Ras
Jepang
Imago bertelur banyak, siklus hidup panjang, bentuk ulat kecil, kokon berwarna kuning atau hijau berlekuk di tengahnya, tebal seperti kacang tanah sehingga produksi kokon lebih tinggi, umur produksinya relatif lebih panjang atau lama, varietas univoltin dan bivoltin. Ras ini memiliki kecepatan tumbuh yang medium.
2 Ras Cina Imago peka terhadap kelembaban yang tinggi, lebih lemah sehingga lebih rentan terhadap serangan penyakit, kokonya berbentuk jorong berwarna putih, kuning emas, kehijauan dan merah jambu. Bentuk kokon bulat, lapisan kokon tipis sehingga produksi serat suteranya lebih rendah dibandingkan ulat sutera ras Jepang, serat suteranya halus dan mudah di pintal, umur produksinya lebih pendek atau cepat, univoltin dan bivoltin serta memiliki kecepatan tumbuh yang cepat.
3 Ras
Eropa
Telur dan ulatnya berukuran besar dengan siklus hidup yang panjang, kokonnya berukuran besar dengan sedikit lekuk di tengahnya, warna kokon putih atau kemerahan, serat suteranya panjang, imago tidak tahan hidup di daerah panas, jenis univoltin, tumbuh lambat dan tidak kuat, sehingga hanya dapat dipelihara di musim semi yang hangat di daerah subtropis saja.
4 Ras
Tropis
Ngengat hidup di daerah tropis, tahan terhadap suhu panas, dan kokon berukuran kecil, polivoltin, mempunyai telur kecil dan ringan, ulat kecil tetapi kuat dan tumbuh sangat cepat, bentuk kokon seperti kumparan, mempunyai banyak serabut (floss) dan kulit kokon tipis, sehingga produksinya rendah.
2.3 Genetika dan pemuliaan ulat sutera
Ulat sutera merupakan hewan penelitian yang ideal sehingga mendapat
per-hatian besar dari para ahli genetika di seluruh dunia. Hal ini dapat dilihat dari
ban-yaknya sifat yang diturunkan, baik pada telur, larva, pupa, maupun imago. Ulat
gene-rasi pertahun. Ulat sutera mempunyai sifat kualitatif dan kuantitatif, dengan
jum-lah kromosom 56 buah yang terdiri dari 27 pasang kromosom tubuh dan 1 pasang
kromosom seks. Kromosom seks betina adalah heterogamet dengan formula
kro-mosom ZW, sedangkan yang jantan homogamet dengan formula ZZ. Krokro-mosom
yang menentukan jenis kelamin betina adalah kromosom W (Atmosoedarjo et al.
2000).
Menurut Brasla dan Matei (1997) tujuan dan sasaran pemuliaan ulat sutera
adalah untuk meningkatkan hasil kokon dan benang sutera serta untuk
menda-patkan jenis ulat sutera yang sesuai dengan masing-masing kondisi lingkungan.
Peningkatan kualitas bibit ulat sutera perlu dilaksanakan di Indonesia, karena bibit
yang digunakan berasal dari daerah sub-tropis, yang biasa dipelihara pada kondisi
optimum. Untuk kondisi tropis seperti Indonesia yang agroklimatnya berfluktuasi,
kualitas pakan rendah dan kemampuan para pemelihara ulat terbatas, diperlukan
jenis ulat yang lebih kuat. Untuk meningkatkan kualitas bibit ulat sutera ada
bebe-rapa cara yang sudah dikenal yaitu seleksi, persilangan, gabungan antara
persi-langan dan seleksi, serta rekayasa genetika (Atmosoedarjo et al. 2000).
2.3.1 Seleksi
Dipandang dari segi genetik, seleksi diartikan sebagai suatu tindakan untuk
membiarkan ternak tertentu bereproduksi, sedangkan ternak lainnya tidak diberi
kesempatan bereproduksi. Seleksi dapat dibagi menjadi dua, yaitu seleksi alam
dan seleksi buatan. Seleksi alam meliputi kekuatan-kekuatan alam yang
menentu-kan ternak-ternak amenentu-kan bereproduksi dan menghasilmenentu-kan keturunan untuk
melanjut-kan proses reproduksi. Ternak yang dapat beradaptasi dengan lingkungannya dan
bereproduksi. Kemampuan ternak untuk bertahan hidup dipengaruhi oleh faktor
genetik.
Pada ulat sutera, seleksi dilakukan bertahap pada galur induk, dimulai dari
telur, ulat, kokon, pupa, dan imago, sehingga hanya individu yang baik saja yang
terpilih untuk bibit. Tujuan seleksi pada setiap stadia berlainan. Pada stadia telur,
bertujuan untuk mendapatkan jumlah telur per induk yang tinggi, penetasan yang
seragam dan persentase penetasan yang baik. Seleksi ulat bertujuan untuk
men-dapatkan keseragaman pertumbuhan, umur ulat yang pendek, dan rendemen
pe-meliharaan yang tinggi. Dalam seleksi harus diperhatikan, bahwa sifat yang
pen-ting secara ekonomi dipengaruhi oleh faktor genetik dan lingkungan. Oleh karena
itu, dalam pemeliharaan untuk galur terseleksi, efek lingkungan harus diusahakan
sekecil mungkin, sedang variasi genetik harus dianalisis dan dievaluasi dalam
memilih galur yang spesifik secara efisien, ketika didapatkan telur dari induk
unggul, kokon galur induk berbentuk kacang dan oval disilangkan untuk menguji
sifat dari hibrid, karena meskipun galur murninya unggul tidak selalu
menghasil-kan hibrid yang berkualitas baik (Atmosoedarjo et al. 2000).
2.3.2 Persilangan
Pendekatan yang digunakan untuk memperbaiki kualitas genetik hewan atau
ternak, yaitu dengan sistem persilangan. Persilangan adalah perkawinan antar
in-dividu, yang tidak memiliki hubungan kekerabatan dalam populasi. Persilangan
biasanya berdampak pada peningkatan daya hidup. Persilangan memiliki tingkat
kesuburan, daya tumbuh dan daya tahan yang lebih tinggi. Gejala ini disebut
Pada ulat sutera, persilangan dilakukan antar galur yang berasal dari daerah
yang berbeda agar sifat-sifat unggul atau karakteristik yang dimiliki
masing-masing galur dapat bergabung pada hibridnya. Persilangan digunakan secara luas
dalam rangka memperbaiki kualitas jenis ulat dengan mengeksploitasi gen-gen
unggul (Atmosoedarjo et al. 2000).
Menurut Razdan et al. (1994) yang paling penting diperhatikan dalam
per-silangan untuk membentuk galur baru maupun hibrid baru adalah karakter spesifik
dari masing-masing ras. Hal-hal lain yang perlu diperhatikan dalam persilangan
adalah sifat yang dipilih, sistem persilangan, induk yang digunakan, lingkungan
dan seleksi. Untuk mendapatkan galur unggul terdapat masalah dalam pemilihan
induk untuk persilangan, mengingat daya gabung tergantung dari interaksi yang
kompleksitas dari gen-gen yang tidak dapat ditentukan hanya dari penampilan
induk-induknya. Oleh karena itu perlu dicoba sebanyak mungkin persilangan,
sebelum hasil terbaik dapat ditentukan.
2.3.3 Efek heterosis dan maternal
Silang luar berpengaruh dalam meningkatkan proporsi gen-gen yang
hetero-zigot dan menurunkan proporsi gen yang homohetero-zigot namun tidak mempengaruhi
frekuensi genotip. Perubahan derajat heterozigositas tergantung dari hubungan
kerabatan ternak yang disilangkan. Jika ternak yang tidak memiliki hubungan
ke-luarga disilangkan, maka keturunannya cenderung menampilkan performa yang
lebih baik dari rataan performa tetuanya untuk sifat-sifat tertentu. Fenomena ini
disebut hibrid vigor yang nilainya dapat diukur. Pengukuran kuantitatif hibrid
vigor disebut heterosis. Heterosis didefinisikan sebagai persentase peningkatan
dikatakan ada jika rataan performa ternak hasil persilangan melebihi rataan tetua.
Laju peningkatan heterozigositas akibat silang luar tergantung pada perbedaan
genetik dari tetuanya. Makin jauh hubungan kekerabatannya antara kedua ternak
tersebut maka makin sedikit kesamaan gen-gennya dan makin besar pula tingkat
heterozigositasnya (Noor 2005). Nilai heterosis untuk setiap sifat berbeda dan
tingkat heterosis bagi masing-masing sifat pun ternyata tidak konsisten atau
bervariasi, karena susunan genetik dari induk yang terlibat dalam persilangan
berlainan (Atmosoedarjo et al. 2000). Pewarisan maternal terdapat apabila faktor
yang menentukan sifat keturunan terdapat di luar inti nukleus dan pemindahan
faktor itu hanya berlangsung melalui sitoplasma. Pengaruh maternal ada apabila
genotip diwariskan dari induk betina menentukan fenotip dari keturunan.
Faktor-faktor keturunan berupa gen-gen yang berasal dari inti dipindahkan oleh kedua
jenis kelamin, dan dalam persilangan-persilangan tertentu sifat-sifat keturunan itu
mengalami segregasi mengikuti pola Mendel. Pengaruh maternal berasal dari
sito-plasma sel telur yang telah dimodifikasi oleh gen-gen yang dipindahkan secara
kromosomal (Suryo 1995).
Cara untuk mengetahui adanya pengaruh maternal, biasanya para pemulia
ulat sutera melakukan perkawinan secara resiprokal untuk menghasilkan
persi-langan yang paling baik. Menurut Welsh (1991) persipersi-langan resiprokal adalah
per-silangan antara dua induk, dimana kedua induk berperan sebagai pejantan dalam
suatu persilangan, dan sebagai betina dalam persilangan yang lain. Seleksi
ber-ulang resiprokal memperbaiki kemampuan berkombinasi spesifik maupun umum.
Cara yang ditempuh adalah dengan melakukan seleksi terhadap dua populasi
da-lam waktu yang bersamaan. Bukti-bukti adanya fenomena pewarisan terpaut
Tazima (1964) terhadap galur yang menghasilkan bobot kulit kokon. Persilangan
resiprokal menghasilkan bobot kulit kokon berbeda pada keturunan pertama dan
kedua pada masing-masing kelamin. Pengaruh gen terpaut kelamin diindikasikan
bervariasi tinggi terhadap bobot kulit kokon pada kedua jenis kelamin
keturu-nannya. Penyebabnya adalah adanya gen utama yang mengontrol sifat-sifat
dewasa terpaut pada kromosom Z yang mempengaruhi ekspresi karakter
kuan-titatif dan aksinya dimodifikasi oleh gen autosomal (Tazima 1964).
Salah satu faktor yang mengakibatkan pewarisan maternal adalah
keberada-an mitokondria pada sel telur. Mitokondria memiliki perkeberada-angkat genetik sendiri
yaitu DNA mitokondria atau sering disingkat mtDNA. mtDNA ini mempunyai
karakteristik yang khas dan diwariskan secara maternal atau pola pewarisannya
hanya melalui garis ibu. Hal ini disebabkan karena sel telur memiliki jumlah
mito-kondria yang lebih banyak dibandingkan sel sperma. Mitomito-kondria dalam sel
sper-ma banyak terkandung di bagian ekor karena bagian ini sangat aktif bergerak
se-hingga membutuhkan banyak ATP. Pada saat terjadi pembuahan sel telur, bagian
ekor sperma dilepaskan sehingga hanya sedikit atau hampir tidak ada mtDNA
yang masuk ke dalam sel telur. Selain itu, dalam proses pertumbuhan sel jumlah
mtDNA secara paternal semakin berkurang. Oleh karena itu dapat dianggap tidak
terjadi rekombinasi sehingga dapat dikatakan bahwa mtDNA bersifat haploid, dan
diturunkan dari ibu ke seluruh keturunannya (Wallace et al. 1996).
2.4 Fenomena heat shock
Heat shock adalah proses cedera termal yang disebabkan oleh peningkatan
suhu secara mendadak dalam molekul biologis seperti DNA, RNA, lipid, dan
ke-lainan pada tingkat sel. Pola yang normal menghentikan sintesis protein. RNA
transfer dan RNA ribosomal memiliki konformasi yang longgar sehingga
menye-babkan degradasi. DNA mengalami kehilangan kemampuan untuk berfungsi
dengan baik. Ada agregasi protein berfilamen pada inti membentuk sitoskeleton
dan pada saat yang sama pH cairan tubuh turun. Kenaikan suhu menyebabkan
pe-ningkatan energi kinetik dari makromolekul, penurunan ikatan ion, ikatan
hidro-gen, Van-der-Wals dan meningkatkan interaksi hidrofobik. Protein mengalami
denaturasi dan membatasi akses enzimatik DNA menyebabkan skala besar
keru-sakan DNA. Stres panas akhirnya menyebabkan kematian sel. Efek lain yang
pa-ling penting dari pengaruh suhu atau stres dalam bentuk apapun adalah pada
protein seluler. Protein seluler biasanya berfungsi dalam sel pada 1) sintesis de
novo dari polipeptida dan perakitan protein, 2) transportasi intra seluler dan
organel, 3) selama atau setelah terpapar stres protein mengalami denaturasi. Pada
perakitan protein dapat mengalami interaksi yang tidak pantas satu sama lain atau
dengan komponen seluler lainnya. Interaksi tersebut dapat menghasilkan agregat
protein dan mengurangi protein fungsional dan akibat terburuk adalah sitotoksik
(Feder 1996).
Paparan singkat pada sel dengan suhu tinggi diharapkan akan memberikan
perlindungan kepada organisme dari paparan suhu berikutnya yang lebih parah.
Kejut panas yang diberikan pada Drosophila subobscura yang bertujuan untuk
mengadaptasikan perubahan suhu pemeliharaan. Selanjutnya dinyatakan bahwa
pemberian kejut panas secara bertahap menunjukkan kelangsungan hidup dan
aklimatisasi serangga pada suhu yang lebih tinggi (Dingley dan Smith 1968).
Ritossa (1962) melaporkan bahwa panas dapat meningkatkan metabolisme
Hal ini diamati ketika larva Drosophila pada suhu 27oC-37oC muncul kromosom
politen. Penemuan ini mengawali diidentifikasinya heat-shock protein (Hsp). Pada
pertengahan tahun 1980-an, peneliti mengakui bahwa Hsp berfungsi sebagai
mo-lekul chaperone. Molekul chaperone adalah jenis protein yang memungkinkan sel
untuk mengatasi masalah protein setelah stres. Protein heat shock melakukan
peran ini dengan membentuk polipeptida baru atau protein yang terungkap selama
proses seluler normal, sedangkan yang diinduksi Hsp berfungsi dalam
menang-gapi denaturasi protein akibat stres. Menurut Kregel (2002), faktor kejut panas
(Hsfs) terdapat dalam sitosol, terikat oleh protein heat shock (Hsp) dalam keadaan
tidak aktif. Sebuah rangsangan fisiologis (stres) diperkirakan dapat mengaktifkan
Hsfs, untuk memisahkan diri dari Hsp. Hsfs terfosforilasi oleh protein kinase dan
membentuk trimer di sitosol. Hsf trimer kompleks memasuki inti dan
memanas-kan elemen kejut panas (Hse) di wilayah promotor dari gen Hsp, mRNA Hsp
di-transkripsi dan meninggalkan inti menuju sitosol, kemudian Hsp baru disintesis.
Induksi gen Hsp dapat dilakukan dengan memberikan rangsang berupa etanol,
logam berat, hipoksia, hiperoksia, perubahan pH, radikal bebas, berbagai racun
dan toksin, iskemia, kejutan osmotik, radiasi pengion dan faktor lingkungan
lainnya (Feder 1996).
2.4.1 Jenis dan fungsi heat shock proteins (Hsp)
Heat-shock proteins diklasifikasikan berdasarkan urutan homologi dan berat
molekul terdiri atas : Hsp110, Hsp100, Hsp90, Hsp70, Hsp40, Hsp10 dan
kelom-pok protein heat-shock kecil. Suhu ambang batas untuk induksi Hsp berkorelasi
dengan suhu yang khas di mana spesies hidup. Spesies termofilik memiliki batas
pola karakteristik dan ekspresi Hsp khas (atau non ekspresi) selama berbagai
ta-hap perkembangan, termasuk gametogenesis, embriogenesis dan metamorfosis.
Studi ekstensif telah dilakukan pada respon heat shock oleh banyak peneliti pada
berbagai spesies serangga seperti Drosophila sp. (Tissiers et al. 1974; Lindquist
1980; Gilchrist dan Huey 1999; Karunanidhi et al. 1999), Locusta migratoria
(Whyard et al. 1986), Anopheles stephensi (Nath dan Lakhotia 1989), Manduca
hornworm-Sexta (Fittinghoff dan Riddiford, 1990), Sarcophaga crassipalpis
(Joplin dan Denlinger 1990) dan Lymantria dispar (Denlinger et al. 1992).
Studi ekspresi Hsp di alam dan respons terhadap rangsangan laboratorium
masih sedikit. Hal ini menunjukkan bahwa pola ekspresi gen Hsp berkorelasikan
dengan lingkungan alam spesies tersebut (Lindquist 1980; Huey dan Bennet 1990;
Sarge et al. 1995; Somero 1995). Sejumlah peneliti menyatakan bahwa Hsp
bertanggungjawab pada proses thermotolerance (Morimoto et al. 1994).
Per-cobaan yang dilakukan dengan cara menghilangkan gen Hsp atau menghambat
ekspresi menunjukkan penurunan thermotolerance (Sanchez dan Lindquist 1990;
Craig dan Jacobsen 1984; Johnston dan Kucey, 1988). Demikian juga penyisipan
gen Hsp-70 meningkatkan thermotolerance pada sel (Li et al. 1991; Solomon et
al. 1991; Li dan Duncan 1995) dan organisme (Welte et al. 1993; Feder et al.
1996).
2.4.2 Bibit larva toleran terhadap panas.
Banyak faktor dikaitkan dengan kinerja yang buruk dari jenis ulat bivoltin
pada kondisi tropis. Aspek utama adalah banyak karakter kuantitatif menurun
ta-jam ketika suhu lebih tinggi dari 28oC. Risiko hibridisasi polivoltin ke bivoltin
mendapatkan hibrida yang produktif sejauh ini belum ditemukan hasil yang
me-muaskan. Pemeliharaan ulat pada suhu tinggi menurunkan kualitas kepompong
(Tazima dan Ohuma 1995). Satu-satunya cara adalah mengembangkan keturunan
bivoltin dengan teknik gen ketahanan serta mengubah masa pemeliharaan pada
kondisi suhu tinggi.
Evegnev et al. (1987) mempelajari respon panas dalam sel B. mori dan
me-nemukan bahwa peningkatan suhu dapat mengaktifkan transkripsi mRNA heat
shock dalam sel yang terpapar, tetapi pada tingkat translasi heat shock gagal
menginduksi sintesis Hsp dan tidak mampu menghambat produksi polyhedrin
da-lam sel tersebut. Joy dan Gopinathan pada tahun 1995 melaporkan munculnya pita
protein pada 93, 70, 46 dan 28 kDa untuk paparan suhu tinggi pada B. mori strain
bivoltin dan multivoltin, tetapi dengan kinetika yang bervariasi. Pada sel hemosit
dari ras multivoltin terdapat induksi protein 70 kDa. Lee et al. (2003) menyatakan
bahwa kloning fragmen DNA mengandung daerah promotor untuk gen
pengko-dean homolog Hsc70-4 dari urutan cDNA parsial yang terdaftar di basis data EST
Bombyx mori. Urutan asam amino 649 pb adalah 89% sampai 96% identik dengan
yang berasal dari Drosphilla melanogaster Hsc-4 dan Meduca sexta Hsc70-4.
Analisis ekspresi menunjukkan bahwa transkripsi mRNA terjadi di semua
jaring-an yjaring-ang diperiksa djaring-an tidak dirjaring-angsjaring-ang oleh heat shock. Jadi Hsc70-4 adalah
mo-lekul chaperon yang terdapat di setiap jaringan B. mori. Dalam penelitian terbaru
Vasudha et al. (2006) menyatakan ekspresi diferensial dari Hsps pada larva
dengan ukuran 90 kDa pada instar pertama, kedua dan ke-tiga, 84 kDa pada instar
keempat dan 84, 62, 60, 47 kDa dan 33 kDa pada instar V. Untuk mencapai
keberhasilan dibutuhkan (1) pemahaman mekanisme toleransi suhu pada ulat, (2)
menginduksi ekspresi, (3) memahami pola diferensial ekspresi Hsp berbagai
bivoltin dan ras polivoltin dan (4) menemukan gen Hsp yang bertanggung jawab