Karya Ilmiah
PENGGUNAAN MANNANOLIGOSAKARIDA DARI
BUNGKIL INTI SAWIT SEBAGAI PENGENDALI
Salmonella sp
PADA TERNAK UNGGAS
Di susun oleh
Dr. Nevy Diana Hanafi, SPt, MSi.
Dr. Ir. Ma’ruf Tafsin, MSi
DEPARTEMEN PETERNAKAN
FAKULTAS PERTANIAN
UNIVERSITAS SUMATERA UTARA
MEDAN
PENDAHULUAN
Mikroba seperti virus atau bakteri berpotensi membahayakan ternak yang dapat ditemukan di udara, makanan atau air. Bakteri yang sering mengkontaminasi ternak ayam, baik pada saat penetasan, pembesaran dan pascapanen diantaranya kelompok Salmonella sp. Bakteri tersebut selain akan berpengaruh terhadap kesehatan ternak juga akan berpengaruh terhadap aspek keamanan pangan ditinjau dari segi mikrobiologis. Potensi yang ditimbulkannnya yaitu dapat mengkontaminasi produk daging atau telur yang akan dikonsumsi manusia.
Beberapa upaya telah ditempuh untuk mengatasi hal tersebut seperti melakukan vaksinasi, sanitasi ataupun penggunaan antibiotik. Upaya tersebut disamping mempunyai banyak manfaat juga mempunyai keterbatasan, sebagai contoh untuk antibiotik sekarang ini ditemukan beberapa strain bakteri yang resisten terhadap antibiotik. Selain itu penggunaannya terutama pada negara maju pengaturannya sangat ketat karena akan berpengaruh pada aspek keamanan pangan untuk manusia. Upaya alternatif dicoba untuk mengatasi keterbatasan tersebut, diantaranya dengan menggunakan karbohidrat. Devegowda et al. (1997) melaporkan bahwa tiga oligosakarida utama yang dapat memperbaiki produksi ternak, yaitu mannanoligosakarida (MOS), fruktooligosakarida, dan galaktooligosakarida, dan MOS dilaporkan memberikan hasil yang paling baik.
Pendekatan baru untuk mencegah infeksi mikroba ditemukan dengan diketahuinya pentingnya proses penempelan pada saluran pencernaan. Diketahui bahwa fimbriae tipe 1 yang sensitif terhadap mannosa berperan dalam menempelnya mikroba patogen. Bakteri seperti Salmonella, E. coli, dan
Vibrio cholera mempunyai lektin pada permukaan selnya yang penempelannya
spesifik terhadap mannosa, dengan demikian mannosa dapat menghambat proses penempelan mikroba merugikan pada saluran pencernaan (Center for Food and Nutrition Policy (CFNP) Technical Advisory Panel (TAP) Review 2002).
Bungkil inti sawit (BIS) adalah hasil ikutan dari industri pengolahan kelapa sawit dan di Indonesia ketersediaannya sangat tinggi. Luas lahan kelapa sawit pada tahun 2004 di proyeksikan sekitar 4.4 juta ha (Jakarta Future Exchange 2001) dan pada tahun 2006 mencapai luas 5.2 juta ha
(Kompas 2006). Produksi tandan buah segar kelapa sawit sekitar 12.5 – 27.5 ton/ha, dan sekitar 2 persennya berupa bungkil inti sawit (Sinurat 2001). Penggunaan BIS sebagai salah satu pakan potensial telah banyak dilaporkan baik pada ternak ruminansia ( Elisabeth dan Ginting 2003; Mathius et al. 2003), ayam (Sundu dan Dingle 2005), bahkan ikan (Keong dan Chong 2005).
Makalah ini mencoba menjelaskan mengenai potensi bungkil inti sawit serta bahan sejenis yang dapat menghasilkan mannanoligosakarida yang dapat dimanfaatkan sebagai pengendali bakteri Salmonella sp.
MIKROFLORA PADA SALURAN PENCERNAAN UNGGAS
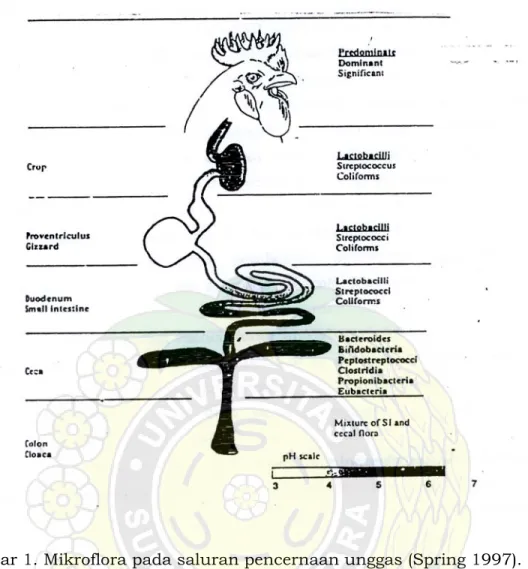
Saluran pencernaan pada unggas yang baru ditetaskan umumnya steril. Sesaat setelah menetas unggas yang masih muda secara alami mikroflora saluran pencernaannya berkembang melalui kontaminasi dari material feses yang berasal dari ayam dewasa. Faktor lain yang berpengaruh yaitu transfer mikroba dari induk pada anak, dan kontak dengan bakteri dari lingkungan. Saluran pencernaan unggas apabila dilihat dari aspek mikrobiologis dapat dikelompokkan menjadi lima bagian yaitu : tembolok (crop); rempela; usus halus; sekum; kolon dan kloaka (Gambar 1).
Gambar 1 menunjukkan bahwa faktor utama yang menentukan populasi mikroba adalah pH. Escherichia coli dan Enterococci merupakan organisme yang dominan yang ditemukan pada unggas yang baru menetas. Pada bagian tembolok, Lactobacillus menjadi dominan pada lima hari pertama, sedangkan pada usus halus memerlukan waktu dua minggu. Kolonisasi bakteri pada usus halus lebih lambat dibandingkan pada bagian lain dari saluran pencernaan dan pada hari pertama konsentrasinya dibawah 105
CFU/g (Coloni Forming Unit). Pada bagian sekum, pada umur unggas sekitar dua sampai empat minggu bakteri obligat aerob meningkat. Pada saat ini bakteri Bifidobacteria, Bacteroides, Eubacteria, Peptostreptococci, dan Clostridia menjadi predominan. Selain itu pada sekum ditemukan juga kelompok bakteri selulolitik pada tingkat diatas 103 CFU/g (Spring 1997).
Gambar 1. Mikroflora pada saluran pencernaan unggas (Spring 1997). Sekarang ini telah diketahui bahwa mikroflora yang secara alami sudah ada dalam saluran pencernaan (indegenous) pada hewan dan manusia dapat memberikan perlindungan terhadap infeksi mikroorganisme yang bersifat patogen. Istilah yang menjelaskan perlindungan tersebut dikenal dengan nama ‘colonization resistance’. Penelitian yang menunjukkan hal tersebut diantaranya dilakukan pada mencit dan diamati pada tiga fase yaitu sebelum, selama, dan sesudah pemberian antibiotik (streptomycin dan neomycin). Hasil penelitian menunjukkan bahwa sebelum pemberian antibiotik ‘colonization resistance’ tinggi terhadap tiga mikroba (E coli; Klebsiela pneumoniae;
Pseudomonas aeroginosa). Selama pemberian antibiotik akan menurunkan
resistensi dan mencit lebih mudah terinfeksi tiga mikroba patogen tersebut karena hilangnya flora pada usus. Selanjutnya pada fase setelah pemberian
antibiotik resistensi ini kembali menuju normal karena terjadinya repopulasi flora saluran pencernaan yang tahan terhadap antibiotik (Hentges 1992).
Hentges (1992) menjelaskan beberapa hipotesis muncul untuk menjelaskan mekanisme yang dapat menekan bakteri patogen. Beberapa faktor tersebut diantaranya muncul teori kompetisi terhadap nutrien; merubah kondisi lingkungan yang tidak ideal bagi patogen seperti dihasilkannya asam lemak terbang oleh flora usus ; dan kompetisi untuk menempati ruang yang ada pada saluran pencernaan. Selanjutnya Mulder et al. (1997) menjelaskan teori “competitive exclusion (CE)” yaitu perlakuan terhadap anak ayam (DOC) yang diberi mikroflora yang menghasilkan resistensi terhadap mikroorganisme yang berpotensi patogen. Beberapa percobaan telah dilakukan menggunakan kultur mikroba murni maupun kultur campuran (undefined microflora). Hasil penelitian menunjukkan bahwa dibandingkan dengan kultur murni ternyata pemberian dengan “undefined microflora” yang berasal dari sekum ayam memberikan hasil yang lebih baik. Kultur tersebut mengandung sejumlah besar mikroba aerobik yang telah diketahui dan banyak bakteri anaerobik yang belum diketahui.
Teori “competitive exclusion (CE)” pertama kali dikemukakan oleh Rantala dan Nurmi (1973) dan banyak mengilhami peneliti selanjutnya untuk mengamati pencegahan bakteri merugikan seperti Salmonella pada ternak unggas. Beberapa hasil positif ditemukan yaitu dengan menurunnya kolonisasi bakteri Salmonella pada ayam broiler dengan digunakannya kultur yang mengandung 29 strain bakteri dari sekum (Corrier et al. 1995). Selanjutnya Ziprin dan Deloach (1993) meneliti pada ayam broiler dan petelur dengan menggunakan bakteri normal dari sekum. Hasil penelitian menunjukkan bahwa bakteri Salmonella menurun meskipun kultur mikroba dari sekum tersebut diberikan tiga hari setelah dilakukan uji tantang terhadap
Salmonella typhimurium.
Selanjutnya Spring (1997) merangkum beberapa mekanisme pengaturan bakteri yang mempengaruhi mikroflora pada saluran pencernaan. Tabel 1 menjelaskan bahwa mekanisme yang tercakup dalam CE sangat kompleks dan dapat dilihat bahwa populasi bakteri mempunyai pendekatan berbeda dalam melakukan kompetisi terhadap bakteri pendatang. Secara garis besar
mekanisme yang terjadi dapat dibedakan secara tidak langsung dan secara langsung. Secara tidak langsung merupakan akibat dari mikroflora normal meningkatkan respon fisiologis inang dan akan mempengaruhi interaksi antara inang dengan mikroba. Mekanisme secara langsung adalah terjadinya saling penekanan antara suatu populasi bakteri terhadap populasi bakteri lainnya. Tabel 1. Mekanisme pengaturan bakteri terhadap mikroflora saluran
pencernaan pada unggas
Mekanisme Pengaturan Faktor Pengontrol
Perangsangan proses kekebalan Ig pada usus halus
Modifikasi garam empedu Asam empedu tak berkonjugasi Stimulasi peristalsis Laju lintas
Penggunaan nutrient Kompetisi nutrien atau faktor pertumbuhan
Pemanfaatan nutrient sinergis Penempelan Kompetisi tempat reseptor
Stimulasi pergantian epitel sel Pembentukan lingkungan terbatas pH
Asam laktat
VFA
Hidrogen sulfida
Modifikasi garam empedu
Perangsangan proses kekebalan Produksi substansi antimikroba Ammonia
Hidrogen peroksida hemolisin Enzim bakteri Bakteriofage Bakteriosin Antibiotik Sumber : Spring (1997).
PENGGUNAAN ANTIBIOTIK PADA TERNAK
Kemajuan peternakan ayam broiler sekarang ini menuntut optimalisasi baik dari segi pertumbuhan, perbaikan konversi ransum, dan kepadatan ternak per satuan luas. Meningkatnya kepadatan akan membawa akibat semakin mudahnya ayam akan terkena serangan penyakit. Upaya pencegahan dan pengobatan yang dilakukan sekarang ini masih bergantung pada penggunaan antimikroba, bahkan dapat dikatakan secara ekonomis tidak mungkin mengembangkan ternak ayam broiler komersial tanpa antimikroba. Pada negara-negara maju seperti Masyarakat Uni Eropa penggunaan antibiotik sebagai imbuhan pakan pengaturannya sangat ketat, dan sampai sekarang penggunaan beberapa antibiotik seperti virginiamycin, tylosin, spiramycin, dan zinc bacitracyn telah dilarang. Pelarangan penggunaan antibiotik yang bersifat pencegahan ini akan membawa akibat meningkatnya penggunaan antibiotik yang bersifat terapetik (menggunakan dosis tinggi), lebih banyak ayam yang akan terkena bakteri patogen dan pada akhirnya kerugian ekonomis akan lebih besar (Bouliane 2003).
Penggunaan antibiotik secara tidak terkontrol akan membawa dampak negatif diantaranya terjadinya resistensi dan ternak yang mengkonsumsi pakan yang mengandung antibiotik, juga akan mengekskresikannya. Levy (2000a). mengemukakan bahwa pada beberapa kasus ditemukan bahwa 80 persen antibiotik yang diberikan secara oral akan lewat dan tidak mengalami perubahan oleh hewan dan masuk ke kolam limbah yang kaya akan bakteri. Selanjutnya akan menyebar ke lahan pertanian karena digunakan sebagai pupuk, dan mengakibatkan pencemaran air permukaan dengan membawa baik obat tersebut maupun bakteri yang resisten ke dalam tanah dan air. Todar (2000) menjelaskan bahwa resistensi mikroba dapat diakibatkan beberapa hal. Pertama, resistensi alamiah, sebagai contoh streptomycete mempunyai gen yang bertanggung jawab untuk resistensi terhadap antibiotiknya sendiri; atau bakteri gram negatif mempunyai membran luar yang menghambat permeabilitas terhadap antibiotik; atau organisme tersebut mempunyai keterbatasan dalam sistem transport terhadap antibiotik; atau terbatasnya target atau reaksi yang akan dicapai oleh antibiotik. Kedua, resistensi buatan, bakteri akan mengembangkan resistensi terhadap antibiotik, yaitu bakteri yang
dahulunya sensitif menjadi resisten. Resistensi seperti ini dihasilkan dari perubahan gen dan dicapai dengan dua cara yaitu ; (1)mutasi dan seleksi; dan (2) pertukaran gen antara strain dan spesies.
Selanjutnya Levy (2000b) menjelaskan bahwa di Amerika Serikat (AS) lebih dari 40 persen antibiotik yang diproduksi diberikan pada hewan baik untuk pencegahan dan pengobatan infeksi, dan pemacu pertumbuhan. Penggunaan antibiotik sebagai pemacu pertumbuhan kadarnya sangat kecil untuk melawan infeksi dan diberikan dalam jangka waktu yang lama (beberapa minggu sampai bulan). Pemberian dalam jangka waktu yang lama dan dosis rendah ini menjadikan bakteri terseleksi dan menjadi resisten. Environmental Media services (EMS) (2000) menjelaskan bahwa bakteri Salmonella umum ditemukan pada produk ternak (daging dan telur) dan di AS dilaporkan infeksi
Salmonella pada manusia lebih dari 40.000 kasus setiap tahun. Selain itu
ditemukan pula strain Salmonella DT-104 yang resisten terhadap lima antimikroba: ampicillin, chloramphenic, streptomycin, sulfonamide, dan tetrasiklin.
MANNANOLIGOSAKARIDA (MOS)
Sumber Mannanoligosakarida (MOS)
MOS dapat diperoleh dari beberapa sumber yaitu dari fungi (dinding sel fungi) dan dari sumber lain seperti dinding sel tanaman atau berupa limbah pertanian. Uraian berikut ini menjelaskan mengenai beberapa sumber yang dapat digunakan untuk memproduksi MOS. Faktor yang perlu diperhatikan dalam memproduksi MOS yaitu kandungan komponen gula mannosa yang dikandung sumber bahan yang akan diekstraksi.
Hasil penelitian Tafsin (2000) menunjukkan bahwa Dinding sel fungi
Penicillium sp didominasi oleh mannosa. Urutan selengkapnya komponen gula
dari dinding sel Penicillium sp adalah tersusun atas glukosa; mannosa; galaktosa; asam glukoronat; arabinosa : dan glukosamin dengan perbandingan konsentrasi berturut-turut 119 ; 169; 11; 15; 1; 1 . Penelitian lanjutan mengenai derajat antigenisitas dengan mengukur produksi antibodi poliklonal dengan menggunakan metode ELISA (Enzymes Link Immunosorbant Assay) menunjukkan bahwa baik glikoprotein maupun polisakarida yang diekstraksi
dari miselium fungi tersebut bersifat imunogenik. Hal tersebut dapat dilihat dari nilai absorbansi yang lebih tinggi (300-400 persen) dibandingkan dengan hewan kontrol. Percobaan tersebut menggunakan hewan kelinci sebagai model percobaannya.
Bungkil inti sawit tinggi akan serat kasar yakni berkisar antara 13.0– 15.7% dan ADF (Acid Detergent Fiber) 31.7% (Daud et al. 1993). Total dinding sel terbanyak adalah mannosa sebesar 56.4%. Formasi linier mannan berbentuk kristal yang cukup tinggi dan ikatan β-(1-4) sulit untuk dipecah. Adapun secara lengkap komponen dinding sel dari bungkil inti sawit tertera pada Tabel 2.
Tabel 2 Persentase komponen gula netral pada bungkil inti sawit (BIS) Komponen gula netral Persentase dari dinding sel (%)
Mannosa 56.4 ± 7.0 Selulosa 11.6 ± 0.7
Xylosa 3.7 ± 0.1
Galaktosa 1.4 ± 0.2
Total 73.1 ± 7.2
Sumber : Daud et al. (1993).
Turner et al. (2000) menyebutkan bahwa sumber yang paling umum
yang dapat digunakan untuk menghasilkan MOS adalah dari Saccharomyces
cerevisae. Hal tersebut dipakai karena kandungan gula mannosanya yang
tinggi yang mencapai 45% dari keseluruhan dinding selnya. Sumber lain yaitu CFNP TAP Review (2002) menyebutkan kandungannnya dapat mencapai 50 persen.
Ishihara et al. (2000) menjelaskan sumber MOS dari tumbuhan yaitu
dari guar gum (Cyamoposis tetragonolobus). Guar gum diperoleh dari biji guar yang selanjutnya diproses dengan menggunakan enzim β-D-mannanase untuk memecah ikatan tulang punggung (backbone) , dan mengandung galaktomannan dengan bobot molekul 20.000 Da.
Peranan MOS sebagai Pengendali Salmonella.
Polisakarida dari nilai nutrisinya secara umum dikenal sebagai penyumbang sumber energi untuk ternak disamping sebagai bagian integral struktur seperti asam nukleat, glikolipid dan glikoprotein. Devegowda et al. (1997) melaporkan bahwa ada tiga oligosakarida utama yang dapat memperbaiki produksi ternak, yaitu Mannanoligosakarida, fruktooligosakarida, dan galaktooligosakarida. Mannanoligosakarida (MOS) dilaporkan memberikan hasil yang paling baik. Selanjutnya pada ayam broiler yang dilakukan uji tantang menggunakan strain liar Salmonella menunjukkan hasil yang lebih baik pada ayam yang diberi MOS. Selain itu MOS juga mempunyai fungsi untuk mengikat mikotoksin seperti zearalenone dan aflatoksin (Lyons 1997; Power 1997).
Pendekatan baru untuk mencegah infeksi mikroba ditemukan dengan diketahuinya pentingnya proses penempelan pada saluran pencernaan. Diketahui bahwa fimbriae tipe 1 yang sensitif terhadap mannosa berperan dalam menempelnya patogen, dengan demikian mannosa dapat menghambat proses penempelan mikroba merugikan pada saluran pencernaan. Kemampuan lain dari MOS adalah dapat merangsang sistem kekebalan dan efek ini juga berperan dalam melawan bakteri Salmonella (Spring 1997).
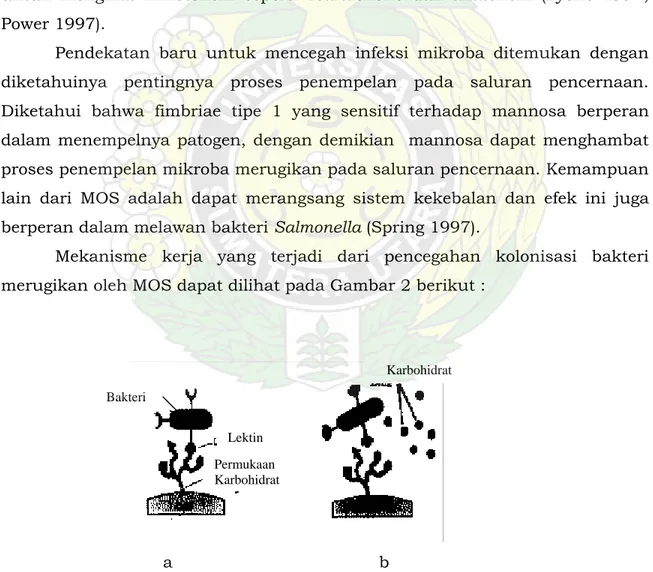
Mekanisme kerja yang terjadi dari pencegahan kolonisasi bakteri merugikan oleh MOS dapat dilihat pada Gambar 2 berikut :
Karbohidrat Lektin Permukaan Karbohidrat Bakteri a b
Gambar 2. Mekanisme kerja MOS mencegah kolonisasi bakteri merugikan (CFNP Technical Advisory Panel (TAP) Review 2002).
Gambar 2a menjelaskan mekanisme terjadinya kolonisasi bakteri pada saluran pencernaan, sedangkan Gambar 2b menunjukkan efek penggunaan karbohidrat seperti MOS dalam mencegah kolonisasi bakteri yang merugikan. Karbohidrat pada permukaan sel merupakan faktor utama yang bertanggung jawab dalam pengenalan oleh sel. Bakteri mempunyai lektin pada permukaan selnya yang dapat mengenal gula spesifik dan membiarkan sel untuk menempel pada gula tersebut. Gula tersebut dapat ditemukan pada permukaaan sel epitel. Pengikatan Salmonella, E. coli, dan Vibrio cholera dimediasi oleh substansi seperti lektin yang spesifik terhadap mannosa dari permukaan sel bakteri. MOS akhirnya dapat mencegah penempelan bakteri patogen pada usus halus sehingga tidak terjadi kolonisasi yang dapat menimbulkan penyakit, dan dapat menjadi sumber makanan terhadap bakteri lain yang menguntungkan (CFNP TAP review 2002).
Turner et al. (2000) menunjukkan adanya efek yang menguntungkan
dari MOS terhadap kesehatan pada saluran pencernaan dan sistem kekebalan. Sebagai contoh terhadap Salmonella thypimurium invitro akan dihambat dengan adanya mannosa, dan selanjutnya setelah dilakukan pemberian melalui air minum pada ayam ternyata menurunkan kolonisasi S. thypimurium pada sekumnya. Selanjutnya pada ternak kalkun, ternyata penggunaan MOS akan meningkatkan level plasma IgG dan konsentrasi IgA pada cairan empedu.
Ishihara et al. (2000) melakukan penelitian MOS yang diperoleh dari
Guar gum dan mengamati efeknya terhadap Salmonella enteridis (SE) pada ayam broiler dan ayam petelur. Hasil penelitian menunjukkan penambahan MOS secara oral menurunkan adanya SE pada organ, Peningkatan ekskresi SE pada feses, menurunkan titer antibodi terhadap SE pada serum. Efek lain yang ditimbulkan yaitu meningkatkan jumlah bakteri Bifidobacterium spp dan
Lactobacillus spp. Keadaan yang sama ditemui pada ayam petelur dengan
menurunnya SE baik pada permukaan kerabang, putih dan kuning telur. Kadar optimum MOS pada penelitian ini yaitu 0.025% dari ransum.
Spring et al. (2000) meneliti efek MOS pada ayam broiler menemukan
bahwa MOS dapat mengaglutinasikan lima dari tujuh strain E.coli dan 7 dari 10 strain Salmonella thypimurium dan Salmonella enteridis. Sedangkan terhadap strain Salmonella pullorum, Salmonella choleraesuis, dan
Campylobacter tidak terjadi agglutinasi. Selanjutnya dilakukan uji tantang
terhadap bakteri Salmonella thypimurium 29E sebanyak 104 cfu pada umur
anak ayam tiga hari. Kadar MOS yang diberikan sebanyak 4000 ppm, dan hasil penelitian menunjukkan terjadinya penurunan konsentrasi Salmonella
thypimurium dari 5.40 menjadi 4.01 log cfu pada hari ke sepuluh. Hasil
penelitian lainnya yang diperoleh menunjukkan bahwa MOS tidak menurunkan konsentrasi coliform pada sekum, dan tidak mempunyai efek terhadap konsentrasi laktobacillus, enterococcus, bakteri anaerob, laktat, VFA, dan pH sekum.
Secara umum, Ferket et al. (2002) membandingkan antara penggunaan antibiotik dengan MOS dan terlihat pada Tabel 3. Tabel 3 menjelaskan bahwa penggunaan antibiotik jelas akan memperbaiki efisiensi pertumbuhan dan kesehatan ternak, tetapi potensi bahaya yang ditimbulkan akibat penggunaan yang tidak tepat sangat besar, diantaranya semakin meningkatnya ancaman dari bakteri patogen yang resisten terhadap antibiotik. Alasan tersebut mendorong industri peternakan untuk menggunakan bahan alternatif yang lebih aman. MOS dapat dikatakan menjadi alternatif terbaik terhadap antibiotik sebagai pemacu pertumbuhan, dan MOS dapat lebih menguntungkan daripada antibiotik jika digunakan secara strategis bersama-sama dengan bahan non obat-obatan seperti probiotik, fruktoosoligosakarida, bio-aktif peptide, dan daun-daunan (‘herb’).
Tabel 3 Perbandingan penggunaan antibiotik dengan MOS
Antibiotik Mannanoligosakarida (MOS) • Menghambat viabilitas dan
proliferasi beberapa mikroflora patogen dan mikroba pencernaan yang menguntungkan
• Mencegah penempelan dan kolonisasi beberapa bakteri pada saluran pencernaan, tapi tidak membunuhnya
• Mempunyai aktivitas spektrum luas terhadap bakteri gram positif
• Mempunyai aktivitas spesifik terhadap bakteri gram negatif yang mempunyai Fimbriae tipe I yang spesifik terhadap mannose
• Menurunkan efek merugikan dari metabolit mikroba dengan menekan mikrofloranya
• Menurunkan efek merugikan dari metabolit mikroba dengan meningkatkan profil mikroflora
• Menurunkan stress imunologis dengan cara menurunkan masuknya mikroba pada saluran pencernaan
• Merangsang sistem kekebalan dengan jalan berlaku seperti antigen mikroba yang bersifat non patogen • Penggunaan secara jangka panjang
dan tidak tepat dapat menghasilkan patogen yang resisten
• Tidak menghasilkan bakteri yang resiten baik terhadap antibiotik atau MOS
• Memberikan keuntungan pada inang untuk menyerap zat makanan penting dengan jalan menekan kompetisi dari mikroba saluran cerna.
• Memberikan keuntungan pada inang untuk menyerap zat makanan penting dengan jalan memperbaiki kesehatan ‘brush borders’.
• Memperbaiki ketersedian Energi Netto (EN) untuk produksi dengan jalan memperbaiki Energi Metabolis (EM) pakan dan menurunkan kebutuhan energi tubuh untuk hidup pokok.
• Memperbaiki ketersedian Energi Netto (EN) untuk produksi dengan jalan memperbaiki Energi Metabolis (EM) pakan.
• Secara konsisten memperbaiki penampilan pertumbuhan pada kondisi lingkungan yang berbeda.
• Memperbaiki penampilan pertumbuhan terutama ketika
dilakukan uji tantang dengan patogen dari saluran pencernaan.
• Menurunkan perlindungan mukosa yang non spesifik dengan jalan menurunkan kolonisasi bakteri yang menguntungkan (sebagai contoh ; laktobasilus)
• Meningkatkan perlindungan mukosa yang non spesifik dengan jalan peningkatan relatif jumlah sel goblet dan sekresi mucus dan meningkatnya koloni bakteri yang menguntungkan.
BUNGKIL INTI SAWIT SEBAGAI SUMBER MOS
Bungkil inti sawit dapat digunakan sebagai salah satu bahan untuk menghasilkan mannan. Penelitian Tafsin (2007) yaitu mencoba kombinasi perlakuan fisik dan kimia dicoba untuk mendapatkan komponen tersebut. Kandungan total gula terekstrak yang dihasilkan mencapai sekitar 2-3 persen dari total BIS (as fed), sedangkan jumlah mannosa terekstrak dari total mannan BIS pada perlakuan kombinasi ekstraksi menggunakan kaca mencapai 5.49% dan 7.58%, berturut-turut pada pelarut menggunakan air dan NaOH 0.05 N. Hasil tersebut masih lebih rendah dibandingkan yang dilaporkan oleh Yokomizo (2005) serta Morikoshi dan Yokomizo (2006) yang menggunakan enzim mannanase terhadap BIS yang mencapai 10-20 % dari total mannan yang ada pada BIS yang diperoleh dengan waktu reaksi selama 24-72 jam dan pada temperatur 60 oC.
Analisis dengan menggunakan kromatografi filtrasi gel (Sephadex G-50) menunjukkan komponen terekstrak berupa polisakarida. Selanjutnya, analisis komponen monosakarida dengan HPLC (Carbohydrate column) menunjukkan bahwa komponen polisakarida tersusun atas glukosa, galaktosa, dan mannosa, dengan komponen dominan berupa galaktomannan. Penggunaan pelarut yang berbeda yaitu air dan NaOH (0.05 dan 0.1N) menghasilkan rasio komponen gula yang berbeda. Penggunaan NaOH ternyata meningkatkan komponen galaktosa, sehingga rasio komponen gula antara mannosa:galaktosa menjadi lebih kecil dibandingkan menggunakan pelarut air. Hal tersebut menunjukkan komponen galaktosa lebih mudah terekstrak karena dalam struktur polisakarida BIS, komponen galaktosa ada pada rantai sisi (side chain) berupa -D-galactopyranosil dengan ikatan (1→6) (Kennedy dan White 1988b). Penggunaan NaOH 0.1 N selain menghasilkan kandungan total gula terekstrak paling rendah, juga menunjukkan kandungan mannosa yang terkecil dibandingkan perlakuan lainnya. Komponen utama dari dinding sel bungkil inti sawit (BIS) adalah linear (1-4)- β –D-Manp (Carre 2002), dan menunjukkan kesamaan dengan guar gum, tetapi dengan rasio galaktomannan yang berbeda. Rasio galaktomannan pada guaran adalah 1:2 (Kennedy dan White 1988).
Respons penggunaan bahan sejenis Mannan terhadap pertumbuhan ayam menunjukkan hasil yang beragam. Laporan Menocal et al. (2005) yang
menggunakan S cerevisiae menunjukkan efek positif dan menyamai penggunaan antibiotik (avilamisin) sebagai pemacu pertumbuhan pada ayam broiler. Efek positif penggunaan MOS terhadap pertumbuhan pada ternak babi dilaporkan oleh Davis et al. (2002), selanjutnya pada kalkun (Zdunczyk et al. 2005) dan pada ayam broiler (Waldroup et al. 2003). Dosis MOS yang digunakan pada penelitian tersebut berkisar 0.05-0.4% dari ransum. Selanjutnya beberapa laporan menyebutkan tidak adanya pengaruh penggunaan MOS terhadap pertumbuhan ayam broiler (Ma et al. 2006; Flemming et al. 2004; dan Shafey et al. 2001) dengan dosis penggunaan berkisar 0.05-0.3%. Hooge (2004) mengumpulkan informasi beberapa penelitian dari tahun 1993-2003 tentang penggunaan MOS menunjukkan bahwa terjadi perbaikan relatif terhadap pertumbuhan sebesar 1.61% dibandingkan pakan kontrol.
Penggunaan mannan dari BIS dari penelitian Tafsin (2007) juga menunjukkan hasil yang beragam terhadap pertumbuhan ayam. Pada keadaan tanpa infeksi, penggunaan mannan dari BIS tidak memberikan pengaruh terhadap PBB. Hasil sebaliknya ditunjukkan pada perlakuan infeksi
S thyphimurium yaitu ayam yang diberi mannan dari BIS menunjukkan
pertumbuhan yang lebih baik dibandingkan perlakukan kontrol infeksi. Secara konsisten terlihat bahwa pada penggunaan sebanyak 2 000-3 000 ppm menunjukkan PBB sekitar 10% dan 20% lebih baik dibandingkan perlakuan kontrol infeksi, berturut-turut pada tingkat infeksi 104 dan 107 CFU S
thyphimurium.
Hasil tersebut menunjukkan bahwa mannan memberikan efek positif pada kondisi lingkungan yang buruk (ancaman mikroba pathogen tinggi) dan mungkin dapat menjelaskan mengapa penggunaan bahan sejenis seperti MOS memberikan hasil yang beragam terhadap pertumbuhan ayam. Efek tersebut diakibatkan penekanan terhadap pertumbuhan akibat kehadiran bakteri yang merugikan seperti S thyphimurium dapat dihindari, dan hal tersebut terlihat pada hasil penelitian yang menunjukkan terjadinya penurunan jumlah koloni dan insiden Salmonella pada penelitian ini.
Penggunaan mannan dari BIS dalam ransum tidak mempengaruhi tingkat konsumsi ransum ayam. Selanjutnya pengamatan terhadap konversi
ransum ayam menunjukkan hasil yang beragam. Nilai konversi ransum pada percobaan 1 dipengaruhi oleh penggunaan mannan, dan penggunaan pada tingkat 4 000 ppm menunjukkan hasil lebih buruk dibandingkan perlakuan lainnya. Hasil sebaliknya ditunjukkan pada percobaan 3 yang menunjukkan tidak adanya pengaruh terhadap konversi ransum ayam. Perbedaan perlakuan yang diberikan yaitu pada percobaan ke 3 dilakukan periode penyesuaian selama 5 hari dengan menggunakan ransum kontrol, sedangkan pada percobaan 1 ransum perlakuan sudah diberikan pada hari pertama. Dampak buruk penggunaan mannan dalam dosis yang tinggi (4 000 ppm) terlihat lebih berat ketika digunakan pada umur ayam satu hari(DOC).
Hal tersebut diakibatkan mannan terekstrak dari BIS ada dalam bentuk komponen yang terlarut dan tergolong pada polisakarida bukan pati (NSP; Non
Starch Polysacharides) dan bersifat antinutritif yang mengakibatkan gangguan
pencernaan dan absorpsi nutrien dengan jalan meningkatnya viskositas dari digesta. Efek buruk tersebut berkurang pada periode penyesuaian pemberian selama 5 hari, dan hal tersebut diduga terkait dengan telah berkembangnya mikroba yang ada dalam saluran pencernaan ayam yang dapat mengeluarkan enzim yang dapat memotong rantai polisakarida menjadi lebih pendek sehingga viskositasnya menurun dan mengakibatkan dampak buruk efek anti nutrisinya menjadi berkurang.
Efek antinutritif NSP sangat tergantung pada ukuran berat molekulnya. Percobaan Choct (2002) menunjukkan hal tersebut yang menggunakan NSP berupa arabinoxylan dan diuji dalam bentuk utuh (BM 758 000 Da) dibandingkan dengan yang sudah didegradasi oleh xylanase sehingga viskositasnya menurun empat kali lipat dan ternyata ketika diuji pada level yang sama (3%), bentuk yang terdegradasi tidak menunjukkan efek antinutrisi pada ayam broiler. Lin dan Hsu (2006) yang meneliti pada angsa menyebutkan bahwa pakan yang mengandung 9% NSP nyata menurunkan penampilan ternak.
Pengujian mannan (in vitro) terhadap Salmonella menunjukkan bahwa mannan dari BIS tidak mempunyai aktivitas yang bersifat membunuh bakteri (bakterisidal) maupun menghambat pertumbuhan bakteri (bakteriostatik), tetapi aktivitas yang ditimbulkannya berupa penempelan antara komponen
mannosa dari mannan dengan reseptor bakteri (fimbriae tipe I) yang tampak secara mikroskopik. Tidak terjadinya aktivitas yang bersifat bakterisidal maupun bakteriostatik tersebut ditinjau dari perspektif lingkungan dan penggunaannya dimasa akan datang sangat menguntungkan. Hal tersebut diakibatkan tidak terjadinya proses adaptasi oleh mikroba yang memungkinkan terjadinya mutasi. Efek terjadinya mutasi bakteri yang membahayakan pada manusia terlihat pada penggunaan antibiotik seperti dilaporkan Environmental Media services (EMS) (2000) yang menjelaskan ditemukannya strain Salmonella DT-104 yang resisten terhadap beberapa antimikroba seperti ampisilin, kloramfenikol, streptomisin, dan tetrasiklin.
PENUTUP
Bungkil Inti Sawit (BIS) sebagai produk samping dari industri pengolahan kelapa sawit mempunyai potensi ketersediaan dan kontinuitas yang tinggi di Indonesia. Nilai manfaat BIS sampai sekarang ini masih lebih banyak ditujukan sebagai salah satu komponen dari pakan baik untuk ternak ruminansia maupun monogastrik. Struktur polisakarida yang spesifik dari BIS didominasi komponen gula mannosa, dan hal ini dapat menjadi nilai tambah untuk mengendalikan bakteri merugikan seperti dari kelompok Salmonella
sp.dan dapat menjadi prebiotik. Hasil tersebut menjadikan potensi BIS tidak
hanya sebagai bahan pakan tetapi dapat ditingkatkan nilai manfaatnya sebagai pengendali bakteri yang merugikan pada ternak.
DAFTAR PUSTAKA
Bouliane M. 2003. Can We Farm Poultry Without Antimicrobials?. Quebec; Fac. of Veterinary Medicine, Univ. of Montreal.
[CFNP TAP] Center for Food and Nutrition Policy Technical Advisory Panel Review. 2002. Cell Wall Carbohydrates : Livestock. Virginia; CFNP. Choct M. 2002. Non-starch Polysaccharides: effect on nutritive value. Di
dalam: McNab JM and Boorman KN, editor. Poultry Feedstuff: supply,
composition, and nutritive value. Nottingham; CAB international.
Corrier DE et al. 1995. Treatment of commercial broiler chickens with a characterized culture of caecal bacteria to reduce salmonella colonization. Poult Sci 74: 1093-1101.
Daud MJ, Jarvis MC, Rasidah A. 1993. Fibre of PKC and its potential as poultry feed. Proceeding. 16th MSAP Annual Conference, Kuala Lumpur,
Davis ME et al. 2002. Effect of mannan oligosaccharides and (or) pharmacological additions of copper sulphate on growth performance and immunocompetence of weanling and growing/finishing pig. J Anim
Sci 80:2887-2894.
Devegowda G et al. 1994. Biotechnology in the feed industry. Proc. Alltech’s
10th Annual Symposium. Nottingham University Press. 235-245.
Elisabeth W, Ginting SP. 2003. Pemanfaatan hasil samping industri kelapa sawit sebagai bahan pakan ternak sapi potong. Pros. Lokakarya
Nasional Sistem Integrasi Kelapa-Sawit Sapi (9-10 September 2003),
Bengkulu.
[EMS] Environmental Media services. 2000. Public Health Concerns. http:www.ems.org/antibiotics/antibiotics_public.html. [31 Januari 2000] Ferket PR, Park CW, Grimes JL. 2002. Benefits of dietary antibiotics and
mannanoligosaccharide supplementation for poultry. Multi-State Poultry
meeting. Raleigh; North Carolina State Univ.
Flemming JS, Freitas JRS, Fontoura P, Monthaninhi NR, Arruda JS. 2004. Use of Mannanoligosaccharides in Broiler Feeding. Braz J Poult Sci. 6(2):159-161.
Hentges JD. 1992. Gut Microflora and Disease Resistance. Di dalam: Fuller R., editor. Probiotics : The Scientific Basis. Chapman and Hall, London. Hooge DM. 2004. Meta-analysis of broiler chicken pen trials evaluating dietary
mannan oligosaccharides, 1993-2003. Int J Poult Sci 3(3): 163-174.
Ishihara N, Shu DC, Akachi S, Juneja LR. 2000. Preventive effect of partially hydrolyzed guar gum on infection of Salmonella enteridis in young and laying hen. Poult Sci 79:689-697.
Jakarta Future Exchange. 2001. Perkembangan produk minyak goreng sawit
di Indonesia. http/www.bbj.jfx.com.
Keong NW, Chong KK. 2002. The nutritive value of palm kernel meal and the effect of enzyme supplementation in practical diets for red hybrid tilapia (Oreochromis sp.). Asian Fish Sci 15:167-176.
Kennedy JF, White CA. 1988. The Plant, Algae, and Microbial Polysaccharides. Di dalam: Kennedy JF, editor. Carbohydrate Chemistry. Oxford; Clarendon Press.
[KOMPAS]. 2006. Malaysia kuasai 330.000 hektar lahan sawit, 25 Agustus 2006.
Levy BS. 2000a. Environmental Concerns:The antibiotics paradox; How miracle
drugs are destroying the miracle. http:www.ems.org/antibiotics/
antibiotics-concerns.html. [30 oktober 2000].
Levy BS. 2000b. The Challenge of Antibiotics Resistance. http/www.sciam.com/1998/0398issue/0398levy.html. [31 Oktober 2000].
Lin PH, Hsu JC. 2006. Effects of different sources of dietary non-starch polysaccharides on the growth performance and carbohydrase development in intestine of goslings. XIIth AAAP Animal science congress
2006 Busan, Korea.
Ma D, Shan A, Li Q, Du J. 2006. Influence of mannan oligosaccharide,
Ligustrum lucidum and Schsandra chinesis on antioxidant and immunity
of chicken. XIIth AAAP Animal Science Congress 2006, Busan.Korea. Mathius IW, Sitompul D, Manurung BP, Azmi. 2003. Produk samping
komplit untuk sapi: suatu tinjauan. Pros. Lokakarya Nasional Sistem
Integrasi Kelapa-Sawit Sapi (9-10 September 2003), Bengkulu.
Menocal JA et al. 2005. Effect of Saccharomyces cerevisiae cell walls on productive parameters in broiler chicks. Tẻc Pecu Mẻx 43(2):155-162. Morikoshi T, Yokomizo F, penemu; 6 April 2006. β-1,4 mannobiose- containing
composition. US patent 2006 0073191.
Mulder RW, Havenaar R, Huis in’t Veld JHJ. 1997. Interventions Strategies: The Use of Probiotics and Competitive Exclusion Microfloras Againts Contamination with Pathogens in Pig and Poultry. Di dalam: Fuller, R, editor. Probiotics 2; Application and Practical Aspect. London; Chapman and Hall.
Rantala M, Nurmi E. 1973. Prevention of the growth Salmonella infantis in chick by the flora of the allimentary tract of chicken. Br Poult Sci 14:627-630.
Shafey TM, Al-mufarez M, Shalaby I, Jarlenabi, AJ. 2001. The effect of feeding mannanoligosaccharides (BioMos) on the performance of meat chicken under two different vaccination programs. Asian-Aust J Anim Sci 14: 559-563.
Sinurat AP. 2001. Pemanfaatan bahan pakan lokal. Makalah pada dies
natalis HIMASITER III, Fapet IPB Bogor.
Spring P. 1997. Understanding the development of the avian gastrointestinal microflora : an essential key for developing competitive exclusion products. Proc. Alltech 11th Annual Asia Pacific Lecture Tour. 149-160. Spring P, Wenk C, Dawson KA, Newman KE. 2000. The effects of dietary
mannanoligosaccharides on cecal parameters and the concentration of enteric bacteria in the ceca of salmonella-challenge broiler chicks. Poult
Sci 79: 205-211.
Sundu B, Dingle J. 2005. Use of enzyme to improve the nutritional value of palm kernel meal and copra meal. Proc. Quensland Poult Sci Symp, Australia 11:1-15.
Tafsin M. 2000. Studi Kimia Polisakarida Dinding Sel Cunninghamella spp dan Penicillium spp yang Diisolasi dari Pakan serta Efek Toksiknya pada Mencit [tesis]. Bogor; Program Pascasarjana, Institut Pertanian Bogor. Todar K. 2000. Bacterial resistance to antibiotics. Dept. of bacteriology, Univ.of
Wisconsin. http/www.bact.wise.edu/Bact330/lecturebactres. [31 Oktober 2000].
Turner JL, Dritz PAS, Minton JE. 2000. Alternatives to conventional microbials in swine diets. Prof Anim Sci 17:217-226.
Waldroup PW, Edgar O, Oviedo-Rondon, Fritts CA. 2003. Comparison of Bio-MOS and antibiotic feeding programs in broiler diet containing cooper sulfate. Int J Poult Sci 2(1):28-31.
Yokomizo F, penemu; Fuji Oil Co. LTD; 24 Mei 2005. Mannose-containing palm kernel meal. US patent 6 896 918.
Zdunczyk Z, Juskiewicz J, Jankowski J, Biedrzycka E, Koncicky A. 2005. Metabolic response of the Gastrointestinal tract of turkeys to diet with different level of mannan-oligosaccharides. Poult Sci 84:903-909
Ziprin RL, Deloach JR. 1993. Comparison of proboitic maintained by in vivo passage through laying hens and broiler. Poult Sci 72: 628-635.