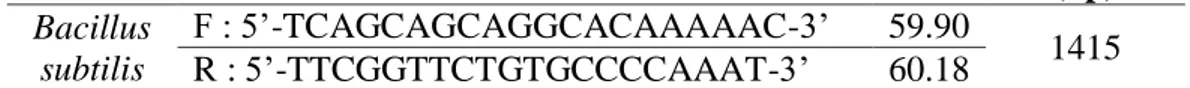41
Peremajaan biakan dan kultur padat pada bakteri B.subtilis dan B.thuringiensis dilakukan dengan memindahkan atau memperbarui biakan bakteri dari biakan lama ke medium tumbuh yang baru secara berkala. Machmud (2001) menyatakan bahwa teknik peremajaan biakan merupakan cara paling tradisional yang digunakan peneliti untuk memelihara koleksi isolat mikroba di laboratorium. Peremajaan biakan ini juga bertujuan untuik menyelamatkan isolat bakteri B.subtilis dan B.thuringiensis dari kontaminasi oleh bakteri lain dan memberikan penyegaran pada nutrien yang dibutuhkan untuk pertumbuhan bakteri.
Peremajaan kultur bakteri dengan menggunakan medium segar dengan jenis yang sama seperti medium awal bertujuan untuk mempercepat fase adaptasi dan mempersiapkan sel pada fase eksponensial. Bakteri yang berada dalam fase eskponensial atau tahap propagasi ini mensintesis enzim dan mengatur aktivitasnya sehingga mampu tumbuh lebih efisien dalam kondisi baru. Peremajaan juga memberikan nutrisi baru bagi bakteri sehingga sel-selnya dapat tumbuh sehat (Hartanti 2010). Nutrient agar (NA) merupakan medium yang umum digunakan untuk mengisolasi organisme dalam kultur murni. Medium NA dapat digunakan pada beberapa bakteri kosmopolit seperti B.subtilis dan B.thuringiensis.
Pertumbuhan B.subtilis yang diinkubasi pada suhu 310C selama 24 jam relatif cepat, hal ini dapat dibuktikan dengan tebalnya koloni B.subtilis pada medium NA (Gambar 11). Cook dan Korsten (1996) menyatakan bahwa B.subtilis memiliki waktu inkubasi optimum 24 jam. Suhu pertumbuhan B.subtilis yaitu pada kisaran 30 - 370C. Sreekumar dan Soundrajan (2010) menyatakan bahwa suhu optimum B.subtilis yaitu pada 350C. Susanti dan Bintari (2013) menyatakan bahwa B.subtilis dapat tumbuh pada range pH 6.0 - 7.0. Sarikaya dan Gurgun (1999) menyatakan bahwa pH optimum bakteri B.subtilis yaitu 7.0. Wijiyono (2009) menyatakan bahwa B.subtilis merupakan bakteri halotoleran yang dapat
tumbuh pada berbagai salinitas yaitu mulai dari salinitas 0 - >30 ppt. Pada salinitas >30 ppt B.subtilis dapat tumbuh dengan jumlah koloni 20.62 x 107 cfu/ml.
Adapun kondisi kultivasi B.thuringiensis tidak berbeda halnya dengan B.subtilis yaitu dilakukan inkubasi pada suhu 310C selama 24 jam. B.thuringiensis dapat tumbuh pada interval suhu 0 - 500C dan memiliki pertumbuhan optimum pada suhu 400C (Mawadza et.al. 2000; Li et.al. 2006; Fan et.al. 2007 dalam Lin et.al 2012). Selain itu, B.thuringiensis dapat tumbuh pada kisaran pH 6.4 - 7.5 (Kwalimwa 2012) dan mampu tumbuh optimum pada pH 7.0 (Kesharvazi et.al 2005). B.thuringiensis dapat tumbuh optimum pada kisaran salinitas 20 - 50 ppt (Phillips et.al. 2012) karena bakteri B.thuringiensis merupakan bakteri halotoleran yang dapat mentolerir berbagai tingkat salinitas (Van-Thuoc et.al 2012).
Peremajaan biakan isolat bakteri B.subtilis dan B.thuringiensis menggunakan tabung reaksi pada agar miring dan kultur padat menggunakan cawan petri. Kultur padat dilakukan menggunakan metode goresan kuadran (Streak quadrant). Metode ini hampir sama dengan goresan T, namun berpola goresan yang berbeda yaitu dibagi empat daerah. Daerah 1 merupakan goresan awal sehingga masih mengandung banyak sel mikroorganisme. Goresan selanjutnya dipotongkan atau disilangkan dari goresan pertama sehingga jumlah semakin sedikit dan akhirnya terpisah-pisah menjadi koloni tunggal (Gambar 11). Dalam kultur murni, goresan T merupakan tipe goresan yang sangat tepat. Prinsip kerjanya yaitu mengencerkan (dilusi) dan menyebarkan (dispersi) sel ketika diinkubasi, sehingga bakteri akan membentuk koloni terisolasi yang dipisahkan. Setelah inkubasi, terlihat dari setiap kuadran koloni bakteri yang terbentuk mengalami perubahan hingga mendapatkan koloni tunggal (Microbiology Faculty 2010). Koloni tunggal digunakan untuk proses uji aktivitas selulolitik karena isolatnya telah murni.
Bakteri B.subtilis yang diremajakan pada penelitian ini memiliki karakteristik morfologis diantaranya bakteri Gram-positif (Lampiran 16), uniseluler yang berbentuk batang dan hidup secara aerob. B.subtilis membentuk
tipe khusus saat dorman yang disebut endospora. Endospora terbentuk dari sel vegetatif sebagai respon terhadap lingkungan yang ekstrim (Todar 2011)
Thimann (1955) menyatakan bahwa bakteri B.subtilis mempunyai kapsul yang berisi polipeptida dari asam D-glutamat. Bakteri B.subtilis merupakan bakteri berspora. Jika ditumbuhkan pada media segar yang cocok (misalnya pada glukosa sederhana atau alanin), spora dapat segera tumbuh. Untuk pertumbuhan sporanya, B.subtilis mengalami dua tahap. Pada tahap pertama terjadi pembengkakan spora. Pembiakan vegetatif pada tahap ini dapat dilihat dengan bantuan metilen biru. Pada tahap kedua dinding spora terbagi melalui bidang ekuator, kemudian di bagian dalam terjadi pemanjangan sel vegetatif yang cepat, membalik sampai spora bermunculan. Pertumbuhan spora B.subtilis terjadi pada pada lingkungan dengan pH mendekati netral.
Gambar 11. Hasil Peremajaan Biakan dan Kultur Padat Isolat Bakteri B.subtilis
a. Agar Miring b. Agar Cawan
Isolat bakteri B.thuringiensis yang didapat dalam penelitian ini memiliki karakteristik morfologis diantaranya merupakan gram positif berbentuk batang (Lampiran 16). Bakteri B.thuringiensis merupakan salah satu anggota B.cereus grup bersama dengan B.anthraxis. Bakteri B.thuringiensis mempunyai ciri khusus yaitu kemampuannya untuk menghasilkan protein kristal protoksin intraseluler dari kelompok δ-endotoksin sehingga dapat dibedakan dengan B.cereus. Bakteri B. thuringiensis memiliki ukuran lebar 1,0 - 1,2 μm dan panjang 3 - 5 μm (Bravo et al., 1998).
Koloni B.thuringiensis jika ditumbuhkan pada media padat berbentuk bulat (Gambar 12). Koloni B.thuringiensis berbentuk oval, berwarna terang,
b a
letaknya subterminal, berukuran 1,0-1,3 μm dengan tepian berkerut, memiliki diameter 5 – 10 mm, berwarna putih, elevasi timbul dan permukaan koloni kasar (Buchner 1981 dalam Syamsu dkk 2011). Ciri khas yang terdapat pada B.thuringiensis adalah kemampuannya membentuk kristal bersamaan dengan pembentukan spora, yaitu pada waktu sel mengalami sporulasi. Kristal tersebut merupakan komplek protein yang mengandung toksin (d-endotoksin) yang terbentuk di dalam sel 2-3 jam setelah akhir fase eksponensial dan baru ke luar dari sel pada waktu sel mengalami autolisis setelah sporulasi sempurna.
Gambar 12. Hasil Peremajaan Biakan dan Kultur Padat Isolat Bakteri B.thuringiensis
a. Agar Miring b. Agar Cawan
4.2 Hasil Uji Aktivitas Selulolitik
Pengujian aktivitas selulolitik pada isolat bakteri B.subtilis dan B.thuringiensis dilakukan dengan menambahkan CMC (Carboxy Methyl Celullose) 1% pada medium nutrient agar + air laut steril. Hal ini bertujuan untuk melihat respon dari isolat bakteri B.subtilis dan B.thuringiensis terhadap substrat selulosa (Tabel 5).
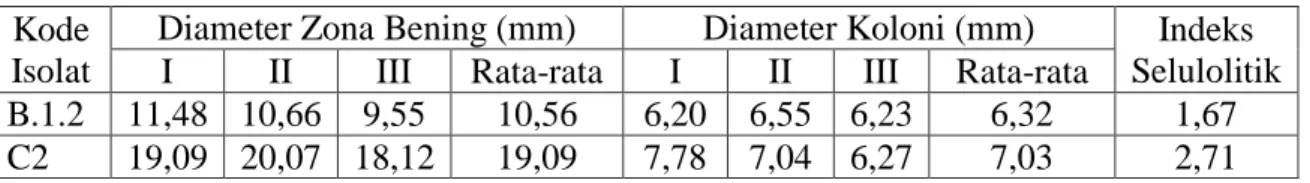
Tabel 5. Uji Aktivitas Selulolitik Pada Isolat Bakteri B.subtilis dan Bakteri B.thuringiensis
Kode Isolat
Diameter Zona Bening (mm) Diameter Koloni (mm) Indeks Selulolitik I II III Rata-rata I II III Rata-rata
B.1.2 11,48 10,66 9,55 10,56 6,20 6,55 6,23 6,32 1,67 C2 19,09 20,07 18,12 19,09 7,78 7,04 6,27 7,03 2,71
Keterangan:
B.1.2 = Isolat bakteri B.subtilis C2 = Isolat bakteri B.thuringiensis
rata-rata diameter zona bening rata-rata diameter koloni
Berdasarkan tabel 5 dapat diketahui bahwa indeks selulolitik isolat B.subtilis lebih kecil dibandingkan dengan indeks selulolitik B.thuringiensis. Hal ini menunjukkan bahwa kemampuan selulolitik B.thuringiensis lebih besar dibandingkan B.subtilis. Baik B.subtilis maupun B.thuringiensis terbukti memperlihatkan kemampuan selulolitik yang ditunjukkan dengan terbentuknya zona bening di sekitar koloni pada medium agar CMC dengan visualisasi menggunakan pewarna Red Congo 0,1% (Gambar 13 dan 14). Isolat bakteri B.subtilis memiliki nilai indeks selulolitik 1,67 mm.
Gambar 13.
Uji Aktivitas Selulolitik Hasil Pewarnaan Red congo (Isolat B.1.2)
Isolat bakteri B.thuringiensis memiliki nilai indeks selulolitik 2,71 mm.
Besarnya nilai indeks selulolitik pada B.thuringiensis menunjukkan kemungkinan produksi enzim endoglukanase lebih besar dibandingkan dengan bakteri B.subtilis, sehingga berpotensi sebagai katalis yang efektif dalam proses pembuatan bioetanol dari rumput laut.
Zona Bening
Koloni Bakteri Indeks selulolitik (IS)
Gambar 14.
Uji Aktivitas Selulolitik Hasil Pewarnaan Red congo (Isolat C2)
CMC digunakan sebagai substrat dalam penentuan aktivitas enzim selulase. CMC merupakan substrat paling murni dibandingkan substrat-substrat lain seperti kertas saring, avicel dan lain-lain (Fitriani 2003). Selain itu, CMC adalah selulosa murni yang mudah terlarut dalam medium dan mudah terhidrolisis (amorf) dibandingkan jika selulosa yang diambil dari alam yang masih berikatan dengan lignin dan hemiselulosa serta masih memiliki struktur yang tidak mudah larut (kristalin) yang tinggi (Astutik 2012). CMC diketahui merupakan substrat efektif untuk produksi enzim endoglukanase (Tae-Il et al. 2000), lebih lanjut Zhang et.al (2006) menyatakan bahwa CMC merupakan substrat yang memiliki spesifikasi untuk pengujian aktivitas enzim endoglukanase dibandingkan dengan substrat selulosa yang lain.
Pada pengujian aktivitas selulolitik dengan penambahan CMC 1% sebagai substrat selulosa pada medium kultur memberikan zona bening yang tidak tampak secara visual, sehingga perlu dilakukan pewarnaan menggunakan red congo untuk melihat zona bening yang dihasilkan (Ramadhan 2012).
Pewarna red congo memiliki interaksi sangat kuat dengan polisakarida yang mengandung unit-unit β-D-Glukan seperti selulosa (Teather and Wood 1981). Kemampuan interaksi ini dijadikan indikator degradasi selulosa dalam medium agar dengan penambahan CMC yang ditunjukan oleh adanya zona bening di sekitar koloni mikroorganisme/ bakteri selulolitik setelah diberi pewarna red congo (Ramadhan 2012). Pembentukan zona bening disekitar koloni bakteri
Koloni Bakteri Zona Bening
menunjukkan sudah tidak terdapatnya (degradasi) CMC pada daerah tersebut. Hal ini mengindikasikan produksi enzim endoglukanase oleh isolat bakteri B.subtilis dan B.thuringiensis untuk memecah CMC menjadi gula yang lebih sederhana sebagai alternatif sumber karbon.
4.3 Teknik Molekuler dengan Polymerase Chain Reaction (PCR) dan Bioinformatik
4.3.1 Isolasi DNA Genom Bakteri
Isolasi DNA genom menggunakan Wizard® Genomic DNA Purification Kit Promega. Isolasi DNA secara umum memiliki empat tahap, yaitu pemecahan sel, ekstraksi DNA, presipitasi DNA dan pencucian DNA. Penggunaan Lysozyme dan Nuclei Lysis Solution berperan dalam proses pemecahan sel dan ekstraksi DNA sedangkan dalam tahap presipitasi digunakan menggunakan Protein Precipitation Solution dan terakhir pengawetan pellet hasil isolasi DNA genom menggunakan ethanol 70% dan DNA Rehydration Solution.
Sebelum masuk ke dalam tahap isolasi, hasil kultur cair disentrifugasi pada 13.000 rpm selama 2 menit untuk mengambil pellet bakteri. Hal ini bertujuan untuk mendapatkan pellet bakteri untuk diisolasi menggunakan Wizard® Genomic DNA Purification Kit Promega. Tahap pertama yaitu dengan menambahkan EDTA yang berperan dalam pelemahan struktur lipid bilayer pada dinding sel bakteri. Selain itu, EDTA juga berperan dalam mempertahankan integritas DNA hasil isolasi dengan cara mengikat ion magnesium yang merupakan kofaktor esensial bagi enzim nuklease yang mampu mendegradasi DNA (Yagi et.al 1996). Tahap kedua yaitu penambahan Lysozyme untuk menyempurnakan proses lisis dinding sel dari bakteri. Lysozyme merupakan enzim yang berperan untuk menghidrolisis rantai polisakarida yang terdapat pada dinding sel bakteri (Sopyan 2009). Suhu inkubasi yang digunakan yaitu selama 45 menit pada suhu 370C.
Tahap ketiga yaitu dengan menambahkan Nuclei Lysis Solution, yang memiliki peran untuk menghancurkan dinding nukleus (pada eukariot). Pada isolasi DNA prokariot, Nuclei Lysis Solution dapat berperan sebagai pengganti
senyawa kimia yang mampu merusak dinding dan membran sel. Suhu inkubasi yang digunakan yaitu selama 5 menit pada suhu 800C. Tahap keempat yaitu penambahan RNAse Solution yaitu untuk menghancurkan RNA sehingga DNA dapat diisolasi secara utuh. Suhu inkubasi yang digunakan setelah RNAse ditambahkan yaitu selama 30 menit pada suhu 370C. Setelah didapatkan DNA secara utuh, maka dilakukan penambahan Protein Precipitation Solution untuk membersihkan debris protein. Peran dari Protein Precipitation Solution yaitu untuk mengendapkan protein agar DNA yang akan diekstraksi tidak terkontaminasi oleh protein-protein sel. Cara memaksimalkan saat proses pembersihan debris protein dengan mengikubasi on-ice selama 5 menit lalu disentrifugasi pada 13.000 rpm selama 3 menit. DNA dipisahkan dari debris jaringan dan protein dengan sentrifugasi dengan landasan bahwa debris sel dan protein dengan besar molekul lebih besar akan mengendap (Angelia 2009).
Tahap ke lima yaitu supernatan yang telah dipisahkan dari debris sel dan protein lalu ditambahkan isopropanol untuk melarutkan lemak, garam alkohol dan mengendapkan DNA. Angelia (2009) menyebutkan bahwa Presipitasi DNA oleh alkohol dapat dilakukan dengan etanol absolut atau isopropanol. Kedua alkohol tersebut memiliki sifat yang berbeda, yaitu: (1) Etanol lebih volatil dibandingkan dengan isopropanol sehingga pelet dapat dikeringudarakan tanpa vakum, (2) Etanol lebih efektif dibandingkan isopropanol dalam melarutkan garam sehingga garam tidak tertinggal pada pelet DNA, (3) Etanol membutuhkan 2.5 kali volume total suspensi DNA untuk dapat mempresipitasi DNA, sedangkan isopropanol cukup dengan 1 kali volume total, dan (4) Akibat kepolarannya, etanol lebih lama mengendapkan DNA dibandingkan dengan isopropanol. Beberapa protokol menyarankan etanol atau isopropanol diberikan dalam kondisi dingin (4°C) untuk mempertahankan integritas DNA. Menurut Sambrook et.al. (1989), suhu yang dingin (4°C) atau suhu ruang (25°C) alkohol tidak berpengaruh signifikan terhadap hasil presipitasi DNA.
Setelah DNA mengendap, supernatan kemudian dibuang dan didapatkan endapan DNA yang bebas dari kontaminasi debris sel, protein, ataupun substansi lain. Pellet DNA pada sampel B.1.2 terlihat lebih banyak dibandingkan dengan
sampel C2. Pellet DNA hasil sentrifugasi ditambahkan ethanol 70% untuk mencuci benang DNA. Angelia (2009) menyatakan bahwa Etanol 70-80 % akan melarutkan garam NH3+ yang kemungkinan masih terdapat pada isolat DNA, tanpa melarutkan asam nukleat. Sentrifugasi dilakukan kembali untuk mengendapkan DNA. DNA yang telah murni lalu ditambahkan DNA Rehydration Solution untuk pengawetan DNA. Dalam membantu proses rehidrasi maka dilakukan inkubasi selama 60 menit pada suhu 650C. Setelah inkubasi, lalu DNA didinginkan selama 5 menit dalam suhu ruang. DNA hasil isolasi dielektroforesis untuk mengetahui kualitas ataupun kuantitas DNA (Gambar 15 dan 16).
Gambar. 15
Elektroforegram Hasil Isolasi DNA Genom Bakteri B.subtilis
Gambar. 16
Elektroforegram Hasil Isolasi DNA Genom Bakteri B.thuringiensis
Hasil Isolasi DNA Genom 500 bp 1000 bp 1500 bp 10.000 bp M B.1.2 Keterangan:
B.1.2 : Isolat Bakteri B.subtilis M : Marker DNA Ladder 1 kb bp : basepair
Keterangan:
C2 : Isolat Bakteri B.thuringiensis M : Marker DNA Ladder 1 kb bp : basepair M 500 bp 1000 bp 1500 bp 10.000 bp Hasil Isolasi DNA Genom
Pada elektroforegram hasil isolasi DNA genom B.subtilis (Gambar 15) nampak bahwa sampel dengan kode B.1.2 terlihat adanya pita. Sedangkan, pada elektroforegram hasil isolasi DNA genom B.thuringiensis (Gambar 16) terlihat bahwa pita DNA sangat tipis. Pita DNA yang tipis mengindikasikan bahwa kemungkinan kuantitas DNA genom yang terisolasi sedikit. Pada hasil isolasi DNA genom bakteri B.subtilis (Gambar 15) dan bakteri B.thuringiensis (Gambar 16) terlihat bahwa ukuran DNA genom bakteri berada di atas 10.000 bp. Genom bakteri diketahui berukuran 4,6 x 106 bp (Escherichia coli) (Yuwono 2005).
4.3.2 Desain Primer Gen Pengkode Endoglukanase a. Primer Spesifik Untuk Bakteri B.subtilis
Sekuen database gen endoglukanase dari bakteri Bacillus subtilis diunduh dari 3 isolat diantaranya Bacillus subtilis subsp. subtilis str. RO-NN-1, Bacillus subtilis subsp. inaquosorum KCTC 13429, dan Bacillus subtilis subsp. subtilis str. BSP1. Pemilihan ketiga isolat ini didasarkan untuk memperbesar peluang amplifikasi gen endoglukanase dari bakteri B.subtilis yang berasal dari strain yang berbeda. Dari ketiga isolat tersebut, didapatkan hasil bahwa konsensus berhasil dikonstruksi. Konstruksi konsensus merupakan kompromi sekuen diantara database yang merepresentasikan lestarinya (conserved) gen tersebut. Hasil konstruksi sekuen konsensus gen endoglukanase dari B.subtilis, maka pasangan primer dari bakteri ini didesain sebagai primer spesifik menggunakan program Primer3® yang tersedia di NCBI. Primer spesifik adalah primer yang didesain secara spesifik dari urutan nukleotida konsensus suatu gen tanpa ada pergantian basa atau (degenerate). Berikut primer gen endoglukanase bakteri B.subtilis (Tabel 6) hasil desain primer spesifik menggunakan program Primer3®.
Tabel 6. Primer Gen Endoglukanase Bakteri B.subtilis Desain Primer3 Jenis
Bakteri Sekuen Primer Tm
Product Length (bp) Bacillus subtilis F : 5’-TCAGCAGCAGGCACAAAAAC-3’ 59.90 1415 R : 5’-TTCGGTTCTGTGCCCCAAAT-3’ 60.18
Berdasarkan hasil desain primer gen endoglukanase bakteri B.subtilis didapatkan hasil bahwa primer tersebut sesuai dengan syarat primer pada umumnya, diantaranya ukuran primer 20 basa pada forward dan reverse, komposisi basa (G+C) 50%, Tm primer forward 59.900C, Tm primer reverse 60.180C. Innis and Gelfand (1990) menyatakan bahwa syarat primer dalam PCR diantaranya ukuran primer berkisar antara 17-28 basa, komposisi basa 50-60% (G+C), Tm (suhu leleh) berkisar antara 55-80oC. Selain itu, primer gen endoglukanase bakteri B.subtilis tidak mempunyai struktur sekunder yang dapat membentuk dimer (Lampiran 1).
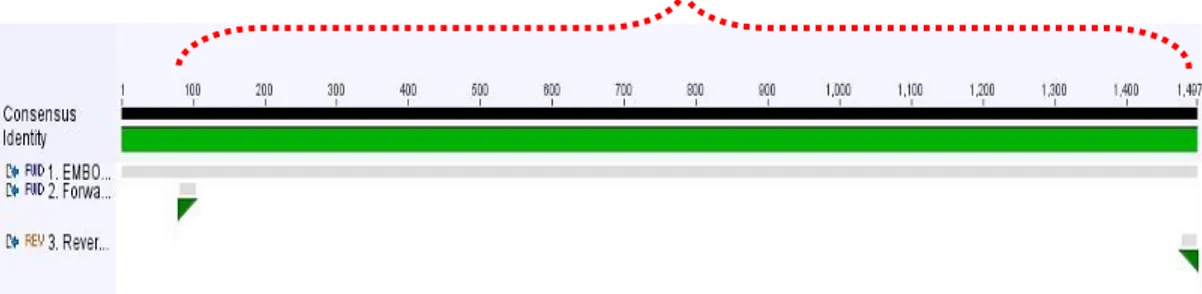
Berdasarkan primer yang tersaji dapat diketahui bahwa panjang amplikon (produk PCR) primer gen endoglukanase bakteri B.subtilis yaitu 1415 bp (472 aa). Ilustrasi penempelan pasangan primer pada sekuen konsensus gen endoglukanase bakteri B.subtilis dapat dilihat pada (Gambar 17).
Gambar 17.
Ilustrasi Penempelan Primer Spesifik Gen Endoglukanase Pada Sekuen Konsensus Bakteri B.subtilis
Pada posisi penempelan primer, dianalisis keberadaan domain fungsional dari gen tersebut. Domain fungsional yang merepresentasikan daerah sisi tapak aktif pengikatan enzim endoglukanase diharapkan mampu teramplifikasi oleh primer yang didesain. Daerah apitan primer sepanjang 1415 bp (472 aa) telah mencakup daerah domain fungsional endoglukanase (BglC). Hipotesis dari teramplifikasinya domain fungsional gen endoglukanase oleh pasangan primer ini membuktikan bahwa gen yang diisolasi merupakan gen pengkode enzim endoglukanase (Gambar 18).
Gambar 18.
Ilustrasi Penempelan Pasangan Primer pada Struktur Utama Domain Fungsional Gen Endoglukanase Pada Sekuen Konsensus Bakteri B.subtilis
(Hasil Analisis Program Conserved Domain Database (CDD)-NCBI :
http://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi)
b. Primer Degenerate Untuk Bakteri B.thuringiensis
Sekuen database gen endoglukanase dari bakteri B.thuringiensis diunduh dari 3 isolat diantaranya Bacillus thuringiensis serovar thuringiensis str. T01001, Bacillus thuringiensis serovar kurstaki str. T03a001, dan Bacillus thuringiensis serovar tochigiensis BGSC 4Y1. Pemilihan ketiga isolat ini didasarkan untuk memperbesar peluang amplifikasi gen endoglukanase dari bakteri B.thuringiensis yang berasal dari strain yang berbeda. Dari ketiga isolat tersebut, didapatkan hasil bahwa konsensus tidak berhasil dikonstruksi, yang menunjukkan kemungkinan tidak terlalu lestarinya gen endoglukanase pada bakteri B.thuringiensis. Dikarenakan tidak berhasilnya konstruksi sekuen konsensus gen endoglukanase dari B.thuringiensis, maka pasangan primer didesain sebagai primer degenerate menggunakan program CODEHOP®. Primer degenerate adalah primer yang didesain secara degenerate dari urutan nukleotida dengan kemungkinan ada peluang terjadinya pergantian basa. Primer gen endoglukanase bakteri B.thuringiensis (Tabel 7) hasil desain primer degenerate menggunakan program CODEHOP®.
472 aa
Tabel 7. Primer Gen Endoglukanase Bakteri B.thuringiensis Desain Primer Degenerate
Jenis
Bakteri Sekuen Primer Lokasi Tm
Product Length (bp) Bacillus thuringien sis F : 5'-GAATTGGATTAGCAT CTTTTTCTAATTCTws nttygcngc-3' Block x24798xblA 60.8 1250 R : 5'-TGAGTTTTCTTATCA TCTGGAACTggyttccac ca-3' Complement of Block x24798xblH 61.9
W(A/T), S(C/G), N(A/C/T/G), Y(C/T)
Berdasarkan hasil desain primer gen endoglukanase bakteri B.thuringiensis didapatkan hasil bahwa primer tersebut sesuai dengan syarat primer pada umumnya, diantaranya Tm primer forward 60.80C dan Tm primer reverse 61.90C. Innis and Gelfand (1990) menyatakan bahwa syarat primer dalam PCR diantaranya Tm (suhu leleh) berkisar antara 55-80oC. Selain itu, primer gen endoglukanase bakteri B.thuringiensis tidak mempunyai struktur sekunder yang dapat membentuk dimer (Lampiran 2).
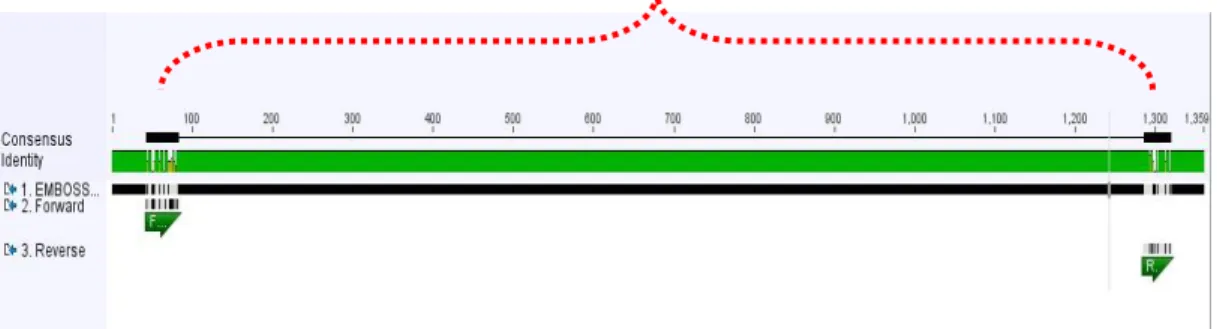
Berdasarkan primer yang tersaji diatas dapat juga diketahui bahwa panjang amplikon (produk PCR) primer gen endoglukanase bakteri B.thuringiensis yaitu 1250 bp (417 aa). Ilustrasi penempelan pasangan primer pada sekuen hasil contig ke tiga database gen endoglukanase bakteri B.thuringiensis dapat dilihat pada (Gambar 19).
Gambar 19.
Ilustrasi Penempelan Primer Degenerate Gen Endoglukanase Pada Sekuen Hasil Contig Ketiga Database Bakteri B.thuringiensis
Pada posisi penempelan primer juga dianalisis keberadaan domain fungsional dari gen tersebut. Daerah apitan primer sepanjang 1250 bp (417 aa) telah mencakup daerah superfamili Glikosida Hidrolase (Glyco_hydro_8_superfamily) dengan salah satu anggotanya yaitu endoglukanase. Hipotesis dari teramplifikasinya superfamili Glikosida Hidrolase menunjukkan peluang teramplifikasinya gen endoglukanase oleh pasangan primer ini (Gambar 20).
Gambar 20.
Ilustrasi Penempelan Pasangan Primer pada Struktur Utama Domain Fungsional Gen Endoglukanase Pada Sekuen Hasil Contig Bakteri
B.thuringiensis
(Hasil Analisis Program Conserved Domain Database (CDD)-NCBI :
http://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi)
4.3.3 Penapisan Gen Pengkode Endoglukanase Menggunakan Metode PCR
Pada tahapan ini, sampel hasil isolasi DNA genom bakteri B.subtilis dan B.thuringiensis diambil untuk dilakukan proses penapisan gen pengkode endoglukanase menggunakan metode PCR. Pada proses ini digunakan komponen reaksi PCR (Tabel 3) dan siklus PCR (Tabel 4) hasil optimasi dari penelitian Rachim (2008).
Optimasi PCR dilakukan sebanyak 4 kali pada penapisan gen endoglukanase bakteri B.subtilis dan B.thuringiensis. Optimasi sampel bakteri B.subtilis dilakukan pada suhu annealing 500C, 550C, dan 600C. Sedangkan
417 bp
optimasi sampel bakteri B.thuringiensis dilakukan pada suhu annealing 450C, 500C, dan 550C. Hal ini dilakukan karena Temperatur Melting (TM) primer gen endoglukanase dari bakteri B.subtilis maupun B.thuringiensis relatif dekat dengan suhu optimasi yang dilakukan. TM primer gen endoglukanase bakteri B.subtilis yaitu forward 65,20C dan reverse 66,20C (Lampiran 3), sedangkan TM primer gen endoglukanase bakteri B.thuringiensis yaitu forward 65,50C dan reverse 67,60C (Lampiran 3). Rybicki (1996) menyatakan bahwa optimasi PCR juga diperlukan untuk menghasilkan karakter yang diinginkan. Optimasi ini menyangkut suhu denaturasi dan annealing DNA dalam mesin PCR. Suhu denaturasi yang rendah dapat menyebabkan belum terbukanya DNA utas ganda sehingga tidak dimungkinkan terjadinya polimerisasi DNA baru. Proses penempelan primer pada utas DNA yang sudah terbuka memerlukan suhu optimum, sebab suhu yang terlalu tinggi dapat menyebabkan amplifikasi tidak terjadi atau sebaliknya suhu yang terlalu rendah menyebabkan primer menempel pada sisi genom lain yang bukan sisi homolognya; akibatnya dapat teramplifikasi banyak daerah tidak spesifik dalam genom tersebut. Suhu penempelan (annealing) ditentukan berdasarkan primer yang digunakan, dipengaruhi oleh panjang dan komposisi primer. Suhu penempelan ini sebaiknya sekitar 50C di bawah suhu leleh. Secara umum suhu leleh (Tm) dihitung dengan rumus Tm = 4(G+C) + 2(A+T)0C.
Hasil optimasi siklus PCR menunjukkan bahwa primer gen endoglukanase bakteri B.subtilis dapat teramplifikasi pada suhu annealing 500C, 550C, dan 600C, sedangkan primer gen endoglukanase bakteri B.thuringiensis dapat teramplifikasi pada suhu annealing 450C, 500C, dan 550C. Suhu annealing semakin dekat dengan TM primer, maka semakin spesifik daerah yang teramplifikasi oleh primer gen tersebut. Sekuen primer gen endoglukanase yang digunakan untuk bakteri B.subtilis yaitu primer spesifik yang didesain menggunakan program Primer3® (Tabel 6), sedangkan sekuen primer gen endoglukanse yang digunakan untuk bakteri B.thuringiensis yaitu primer degenerate yang didesain menggunakan program CODEHOP® (Tabel 7). Primer gen endoglukanase bakteri B.subtilis memiliki target amplikon 1.415 bp, sedangkan primer gen endoglukanase bakteri B.thuringiensis memiliki target amplikon 1.250 bp. Primer merupakan komponen
paling penting dalam reaksi PCR karena berperan sebagai pemula pada proses sintesis (William et.al 1990 dalam Ain 2011) dan menentukan daerah genom yang akan diamplifikasi. Hasil amplifikasi dengan PCR dipisahkan dengan elektroforesis gel agarosa dan secara langsung divisualisasikan setelah pewarnaan dengan EtBr (Gambar 21 dan 23).
Gambar. 21
Elektroforegram Amplikon Gen Endoglukanase Bakteri B.subtilis Suhu Annealing 600C (B.1.2)
Keterangan:
B.1.2 : Amplikon Gen Endoglukanase Bakteri B.subtilis M : Marker DNA Ladder 1 kb
- : Kontrol Negatif bp : basepair
Pada Gambar 21, menunjukan bahwa pita dari produk amplikon berada pada ukuran diantara 1000 dan 1500 bp sesuai dengan target saat desain primer spesifik menggunakan program Primer3® yaitu 1415 bp. Sehingga sampel hasil amplifikasi tersebut dapat dilanjutkan ke tahap selanjutnya yaitu sekuensing. Sebelum dilakukan sekuensing, terlebih dahulu dilakukan purifikasi produk amplikon oleh 1st BASE (Gambar 22).
1500 bp Amplikon Gen Endoglukanase Bakteri B.subtilis 10.000 bp 2000 bp 1000 bp 500 bp --- B.1.2 M
Gambar. 22
Hasil Purifikasi Amplikon Gen Endoglukanase Bakteri B.subtilis (B.1.2) Oleh 1st BASE
Pada Gambar 22 menunjukkan hasil purifikasi pita produk amplikon yang akan disekuensing berada pada ukuran ~1500 bp. Ukuran tersebut berada pada posisi yang sama dengan posisi pita pada elektroforegram hasil PCR.
Gambar. 23
Elektroforegram Amplikon Gen Endoglukanase Bakteri B.thuringiensis Suhu Annealing 550C (C2)
Keterangan:
1 : Kode Bakteri B.1.2 1kb : Marker DNA Ladder 1 kb bp : basepair
Keterangan:
C2 : Amplikon Gen Endoglukanase Bakteri B.thuringiensis M : Marker DNA Ladder 1 kb - : Kontrol Negatif bp : basepair 1500 bp 2000 bp 10.000 bp Amplikon Gen Endoglukanase Bakteri B.thuringiensis M C2 -- 1000 bp 500 bp 1.500 bp 1.000 bp 750 bp 250 bp 500 bp 2.000 bp
Pada Gambar 23, menunjukan bahwa pita dari produk amplikon berada pada ukuran diantara 1000 dan 1500 bp sesuai dengan target saat desain primer degenerate menggunakan program CODEHOP® yaitu mendekati ukuran 1250 bp, sehingga sampel hasil amplifikasi tersebut dapat dilanjutkan ke tahap selanjutnya yaitu sekuensing. Sebelum dilakukan sekuensing, terlebih dahulu dilakukan purifikasi produk amplikon oleh 1st BASE (Gambar 24).
Gambar. 24
Hasil Purifikasi Amplikon Gen Endoglukanase Bakteri B.thuringiensis (C2) oleh 1st BASE
Keterangan:
1 : Kode Bakteri C2
1kb : Marker DNA Ladder 1 kb bp : basepair
Pada Gambar 24 menyatakan bahwa hasil purifikasi pita produk amplikon yang akan disekuensing berada pada ukuran ~1.500 bp. Ukuran tersebut berada pada posisi yang sama dengan posisi pita pada elektroforegram hasil PCR.
4.3.4 Analisis Hasil Sekuensing Gen Pengkode Endoglukanase
Hasil amplifikasi PCR (amplikon) gen endoglukanase bakteri B.subtilis dan B.thuringiensis kemudian disekuensing. Pada tahapan ini, sekuensing dilakukan menggunakan 1st BASE. Pada prosesnya, sekuensing dilakukan dengan
750 bp 250 bp 500 bp 1.000 bp 1.500 bp 2.000 bp
menggunakan primer forward dan reverse gen endoglukanase dari masing-masing template.
Hasil sekuensing (Lampiran 22 dan 24) berupa urutan basa-basa nukleotida penyusun DNA gen dari sampel bakteri yang dikarakterisasi secara molekuler. Sekuensing merupakan proses pembacaan urutan basa (nukleotida) dari suatu molekul DNA. Secara umum proses sekuensing mendasarkan metodenya pada prinsip sekuensing Sanger. Molekul DNA pada awalnya diamplifikasi sesuai dengan target menggunakan suatu pasangan primer. Pada proses pembacaan urutan basa, digunakan molekul dideoxynucleoside triphosphate (ddNTP) yang dilabel dengan suatu unsur radio aktif (pada umumnya 32
P) atau molekul fluorescen sebagai terminator penghubung untuk mendeteksi basa yang sesuai. Pada tahap akhir proses sekuensing, dilakukan elektroforesis masing-masing bagian template molekul DNA dan hasilnya didokumentasi dalam bentuk autoradiografi atau langsung terhubung dengan detektor fluorescen pada sekuenser DNA otomatis (Primrose and Twyman 2006).
Jenis data hasil sekuensing yang diperoleh pada penelitian ini berupa data kromatogram dan urutan nukleotida sekuen forward dan reverse yang harus diolah menggunakan perangkat/program BioEdit® (Lampiran 21). Data yang diperoleh dari hasil penggunaan program BioEdit® digunakan sebagai data dasar untuk diolah kembali pada multiple aligment (pensejajaran berganda) dengan database sekuen yang ada di GeneBank dengan NCBI BLAST pada level nukleotida dan dapat diakses di website http://blast.ncbi.nlm.nih.gov/blast.cgi.
Penggunaan pensejajaran berganda bertujuan untuk mensejajarkan dan mencocokan hasil sekuensing yang diperoleh dari hasil penelitian dengan data yang telah ada di GeneBank. Analisis ini dilakukan untuk memastikan bahwa hasil sekuensing dari penelitian ini benar-benar merupakan sekuen dari gen endoglukanase bakteri B.subtilis dan B.thuringiensis. Setelah didapatkan konsensus dari sekuen primer forward dan reverse, maka setelah itu dilakukan analisis BLASTX untuk konfirmasi awal terhadap masing-masing hasil sekuensing sehingga dapat diketahui apakah gen target merupakan gen endoglukanase atau bukan. Analisis awal BLASTX (Tabel 8) ini menunjukkan
bahwa hasil sekuensing menunjukkan tingkat homologi yang cukup tinggi dengan gen endoglukanase bakteri B.subtilis pada basis data GeneBank.
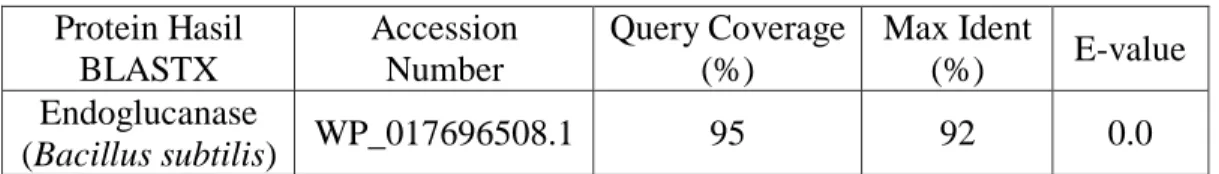
Tabel 8. Hasil Analisis Pensejajaran Berganda (BLASTX) Urutan Sekuen Hasil Sekuensing Template Bakteri B.subtilis dengan Data GeneBank
Protein Hasil BLASTX Accession Number Query Coverage (%) Max Ident (%) E-value Endoglucanase (Bacillus subtilis) WP_017696508.1 95 92 0.0 Data hasil yang diperoleh menunjukan sekuen hasil sekuensing memiliki tingkat kesesuaian/homologi yang tinggi dengan gen endoglukanase dari bakteri B.subtilis dengan Query Coverage 95% dan Max Ident 92% dengan data di GeneBank (Lampiran 23). Hasil BLASTX dari sekuen hasil sekuensing dari template amplikon bakteri B.thuringiensis disajikan pada Tabel 9 dibawah ini.
Tabel 9. Hasil Analisis Pensejajaran Berganda (BLASTX) Urutan Sekuen Hasil Sekuensing Template Bakteri B.thuringiensis dengan Data GeneBank Protein Hasil BLASTX Accession Number Query Coverage (%) Max Ident (%) E-value Endoglucanase (Bacillus thuringiensis serovar pakistani str. T13001) EEM47662 92 98 0.0
Pada Tabel 9 terlihat bahwa sekuen hasil sekuensing memiliki tingkat kesesuaian/homologi yang tinggi dengan gen endoglukanase dari bakteri B.thuringiensis. Hal ini ditunjukan dengan Query Coverage 92% dan Max Ident 98% dengan data di GeneBank (Lampiran 25).
4.3.5 Analisis Hasil Karakterisasi In Silico Sekuen Gen Pengkode Endoglukanase
a. Domain Fungsional
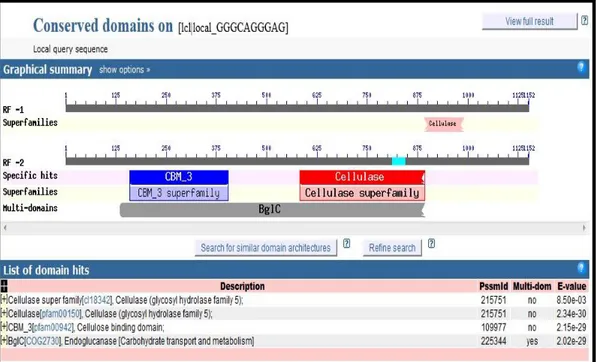
Domain fungsional merupakan sekuen lestari yang mencirikan suatu kelompok gen tertentu yang memiliki karakteristik khas secara struktural. Domain biasanya lebih panjang dari motif. Sebuah domain itu terdiri lebih dari 40 residu dan sampai 700 residu, dengan rata-rata panjang 100 residu. Sebuah domain dapat memiliki suatu bagian motif didalamnya dapat pula tidak mengandung motif (Xiong 2006). Hasil analisis karakter domain fungsional dari sekuen gen pengkode endoglukanase dari bakteri B.subtilis menunjukkan bahwa pada sekuen gen tersebut ditemukan multi-domain BglC yang menunjukkan karakter khas gen pengkode enzim endoglukanase pada bakteri (Gambar 25).
Gambar 25. Struktur Domain Fungsional Sekuen Gen Pengkode Endoglukanase Bakteri B.subtilis
(Hasil Analisis Program Conserved Domain Database (CDD)-NCBI :
http://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi)
Pada sekuen gen endoglukanase bakteri B.subtilis juga ditemukan daerah domain katalitik selulase (cellulase) pada residu asam amino 1-70 dan superfamilinya yang menunjukkan karakter khas kelompok enzim selulase
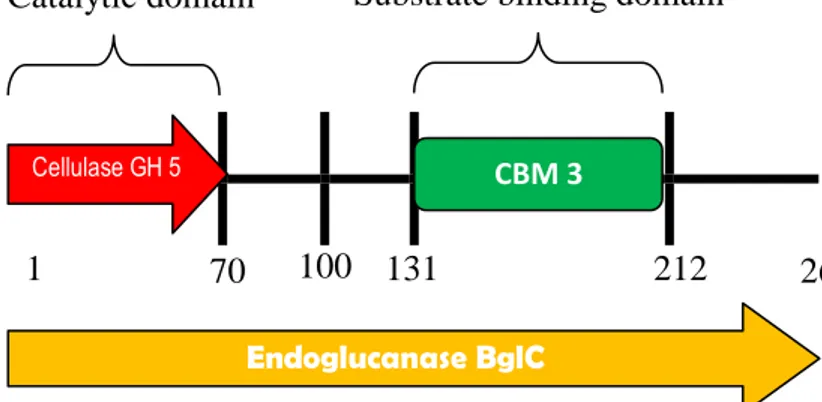
(kelompok hidrolase 5). Daerah domain CBM3 (Carbohydrate Binding Module) juga ditemukan pada sekuen tersebut yang menunjukkan daerah pengikatan enzim terhadap substrat selulase (bagian dari karbohidrat) pada residu 131-212 (Gambar 26). Satu dari 6 residu glutamat (E atau Glu) yang teridentifikasi dalam sekuen yaitu Glu211 diduga memiliki peran penting dalam analisi topologi ini. Residu asam amino ini menjadi penanda aktifitas katalitik endoglukanase pada proses hidrolisis selulosa (Rachim 2008; Armstrong et.al 1998 ).
10 20 30 40 50 60
MYALHFYAGT HGQFLRDKAN YALSKGAPIF VTEWGTSDAS GNGGVFLDQS REWLKYLDSK
70 80 90 100 110 120
TISWVNWNLS DKQESSSALK PGASKTGGWR LSDLSASGTF VRENILGTKD STKDIPETPA 130 140 150 160 170 180 KDKPTQENGI SVQYRAGDGS MNSNQIRPQL QIKNNGNTTV DLKDVTARYW YNAKNKGQNV
190 200 210 220 230 240
DCDYAQLGCG NVTYKFVTLH KPKQGADTYL ELGFKNGTLA PGASTGNIQL RLHNDDWSNY 250 260
AQSGDYSFFK SNTLKQRKKS HMTAPC
Gambar 26. Sekuen Asam Amino Gen Pengkode Endoglukanase Bakteri B.subtilis (a) Topologi Sekuen, (b) Struktur Domain Fungsional : Domain Katalitik (Cellulase GH5) Residu 1-70 ; Domain Pengikatan Substrat (CBM3) Residu 131-212, E (Glu) : Residu Glutamat (a) (b) Catalytic domain 70 1 100 131 212 266 Cellulase GH 5 CBM 3
Substrate binding domain
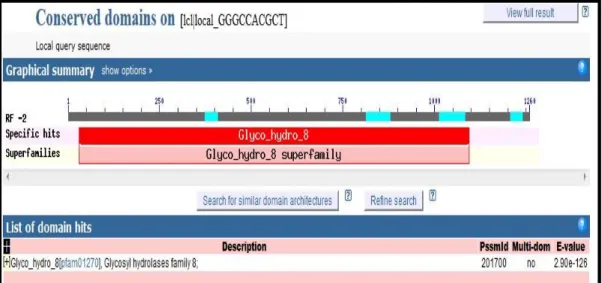
Hasil analisis karakter domain fungsional dari sekuen gen pengkode endoglukanase dari bakteri B.thuringiensis menunjukkan bahwa pada sekuen gen tersebut ditemukan daerah domain katalitik yang merupakan daerah spesifik Glyco Hydro 8 pada residu asam amino 30-370. Hal ini menunjukkan karakter khas enzim pengkode kelompok hidrolase famili 8, termasuk didalamnya adalah endoglukanase (Gambar 27).
Gambar 27. Struktur Domain Fungsional Sekuen Gen Pengkode Endoglukanase Bakteri B.thuringiensis
(Hasil Analisis Program Conserved Domain Database (CDD)-NCBI :
http://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi)
Domain katalitik Glyco Hydro 8 (GH 8) merupakan daerah yang menunjukkan karakteristik daerah fungsional kelompok enzim pengkode enzim-enzim hidrolase famili 8, termasuk didalamnya adalah selulase (Gambar 28).
Gambar 28. Kelompok Domain Glyco Hydro 8 (GH 8)
Topologi sekuen asam amino gen pengkode endoglukanase bakteri B.thuringiensis dan struktur domain fungsionalnya disajikan pada Gambar 29. Sebanyak 8 residu glutamat juga ditemukan pada sekuen ini
10 20 30 40 50 60 MKPFPQQVNY AGVIKPNHVT QESLNASVRS YYDNWKKKYL KNDLSSLPGG YYVKGEITGD
70 80 90 100 110 120
ADGFKPLGTS EGQGYGMIIT VLMAGYDSNA QKIYDGLFKT ARTFKSSQNP NLMGWVVADS 130 140 150 160 170 180
KKAQGHFDSA TDGDLDIAYS LLLAHKQWGS NGTVNYLKEA QDMITKGIKA SNVTNNNRLN
190 200 210 220 230 240
LGDWDSKSSL DTRPSDWMMS HLRAFYEFTG DKTWLTVINN LYDVYTQFSN KYSPNTGLIS 250 260 270 280 290 300
DFVVKNPPQP APKDFLDESE YTNAYYYNAS RVPLRIVMDY AMYGEKRSKV ISDKVSSWIQ
310 320 330 340 350 360
NKTNGNPSKI VDGYQLNGSN IGSYPTAVFV SPFIAASITS SNNQKWVNSG WDWMKNKRES
370
YFSDSYNLLT MLLYRLQLW
Gambar 29. Sekuen Asam Amino Gen Pengkode Endoglukanase Bakteri B.thuringiensis (a) Topologi Sekuen, (b) Struktur Domain Fungsional : Domain Katalitik (Glyco Hydro 8) Residu 30-370, E: Residu Glutamat (Glu).
(a) (b) Catalytic domain Glyco Hydro 8 1 30 370 379 Endoglucanase
Pada sekuen gen pengkode endoglukanase bakteri B.thuringiensis tidak ditemukan secara spesifik multi-domain BglC (endoglukanase) seperti yang ditemukan pada endoglukanase dari bakteri B.subtilis. Hal ini dikarenakan sekuen gen tersebut memiliki kemiripan sekuen dengan domain kelompok enzim hidrolase yang lain seperti chitosanase (Lampiran 25) .
b. Sisi Aktif
Sisi aktif merupakan daerah/situs yang berkaitan dengan aktifitas enzim dalam kondisi fungsional, pada umumnya terkait juga dengan pengikatan enzim terhadap substrat. Daerah sisi aktif juga diketahui ditandai oleh keberadaan residu asam amino tertentu. Pada endoglukanase, residu glutamat (E atau Glu) dan residu asam aspartat (D atau Asp) merupakan residu yang menjadi penanda aktifitas enzim tersebut (Kawaminami et.al 1999). Pada penelusuran keberadaan sisi aktif dari sekuen protein gen pengkode endoglukanase B.subtilis diketahui terdapat daerah sisi aktif CBM3 yang diidentifikasi sebagai daerah pengikatan enzim terhadap substrat selulase (Gambar 30).
Gambar 30. Daerah Sisi Aktif Gen Pengkode Endoglukanase Bakteri B.subtilis
(Sumber :
Pada penelusuran sisi aktif gen pengkode endoglukanase bakteri B.thuringiensis juga ditemukan keberadaan sisi aktif. Sisi aktif tersebut menunjukkan karakter sisi aktif kelompok enzim gliko-hidrolase famili 8 (GH-8) dengan urutan sekuen asam amino ATDGDLDIAYSLLLAHKQW (Gambar 31).
Gambar 31. Daerah Sisi Aktif Gen Pengkode Endoglukanase Bakteri B.thuringiensis
(Sumber :
http://prosite.expasy.org/cgi-bin/prosite/ScanView.cgi?scanfile=346154428423.scan.gz)
c. Motif
Motif adalah suatu pola sekuen lestari yang berukuran pendek yang berkaitan dengan perbedaan fungsi dari suatu protein. Pada penelusuran motif dari sekuen protein gen pengkode endoglukanase bakteri B.subtilis, diketahui terdapat motif dominan yaitu CBM3, seperti yang telah dijelaskan sebelumnya bahwa motif protein ini menunjukkan aktifitas pengikatan substrat karbohidrat (selulosa) oleh enzim endoglukanase. Motif-motif pendek yang lain juga ditemukan di sepanjang sekuen protein, diantaranya ASN_GLYCOSYLATION, CAMP_PHOSPHO_SITE, CK2_PHOSPHO_SITE, MYRISTYL, dan PKC_PHOSPHO_SITE (Gambar 32).
Motif ASN_GLYCOSYLATION merupakan daerah atau situs modifikasi pasca translasi berupa proses penambahan glikan (disebut glikosilasi) pada daerah
residu aspargin (Asn), berukuran 3 asam amino (Medzihradszky 2008). Motif CAMP_PHOSPHO_SITE menunjukkan daerah terjadinya fosforilasi menggunakan cAMP atau cGMP, motif ini memiliki konsensus [RK](2)-x-[ST] dan pada sekuen gen ini ditemukan dalam urutan RKKS (Deihimi 2012). Motif CK2_PHOSPHO_SITE menunjukkan daerah tempat terjadinya fosforilasi kasein kinase II, motif ini memiliki konsensus [ST]-x(2)-[DE] dan pada sekuen gen ini ditemukan dalam urutan STKD, TTVD, dan TYLE (Gao and Wang 2006). Motif MYRISTYL menunjukkan situs tempat proses miristilasi yaitu penambahan gugus myristyl pada suatu protein diakhir proses translasi. Motif ini memiliki konsensus G-{EDRKHPFYW}-x(2)-[STAGCN]-{P} dan pada sekuen gen ini ditemukan dalam urutan GTSDAS, GASKTG, GTKDST, GSMNSN dan
GASTGN (Stroh et.al 2002). Motif PKC_PHOSPHO_SITE menunjukkan daerah tempat terjadinya fosforilasi protein kinase C, motif ini memiliki konsensus
[ST]-x-[RK] dan pada sekuen gen ini ditemukan dalam urutan SDK, STK, TAR, TYK, TLK (Leonard et.al 2011).
Gambar 32. Motif Sekuen Protein Gen Pengkode Endoglukanase Bakteri B.subtilis
Penelusuran motif pada sekuen protein gen pengkode endoglukanase bakteri B.thuringiensis menunjukkan motif protein glikohidro 8 (Glyco Hydro 8). Motif ini diidentifikasi sebagai motif yang menjadi ciri khas kelompok enzim golongan glikohidrolase, termasuk endoglukanase. Motif-motif pendek yang lain juga ditemukan di sepanjang sekuen protein, diantaranya Motif ASN_GLYCOSYLATION, motif CAMP_PHOSPHO_SITE dalam urutan sekuen
KRES, motif CK2_PHOSPHO_SITE dalam urutan sekuen SYYD, SATD, SSLD
dan SGWD, motif MYRISTYL dalam urutan sekuen GTSEGQ, GMIITV,
GLFKTA, GSNGTV, GIKASN dan GSNIGS serta motif PKC_PHOSPHO_SITE dalam urutan sekuen SVR, TAR, TFK, SKK, SNK, dan
SDK(Gambar 33).
Gambar 33. Motif Sekuen Protein Gen Pengkode Endoglukanase Bakteri B.thuringiensis
Berdasarkan hasil karakterisasi secara In Silico dari sekuen protein gen pengkode endoglukanase pada bakteri B.subtilis dan B.thuringiensis dapat diketahui bahwa kedua sekuen protein tersebut memiliki aktifitas fungsional yang menunjukkan peluang terekspresinya enzim endoglukanase dari kedua isolat bakteri tersebut bila dilakukan proses ekspresi gen. Dari kedua sekuen protein tersebut, juga diketahui memiliki kemiripan pada beberapa aspek, yang dapat mengindikasikan lestarinya (tidak terlalu banyak terjadi mutasi) sekuen gen endoglukanase pada genus yang sama. Matriks perbandingan karakterisasi In Silico dari kedua gen pengkode endoglukanase tersebut disajikan pada Tabel 10.
Tabel 10. Matriks Perbandingan Sekuen Protein Gen Pengkode Endoglukanase Dari Bakteri B.subtilis dan B.thuringiensis Isolat Ukuran Sekuen Domain Fungsional Sisi Aktif Motif B.subtilis 1151 bp/ 383 aa -Cellulase super family -Cellulase -CBM_3 -BglC CBM3 -ASN_GLYCOSYLATION -CAMP_PHOSPHO_SITE -CK2_PHOSPHO_SITE -MYRISTYL -PKC_PHOSPHO_SITE B.thuringiensis 1258 bp/ 418 aa Glyco_hydro _8 Glyco Hydro 8 -ASN_GLYCOSYLATION -CAMP_PHOSPHO_SITE -CK2_PHOSPHO_SITE -MYRISTYL -PKC_PHOSPHO_SITE Berdasarkan Tabel 10 diatas dapat diketahui bahwa kedua sekuen protein pengkode gen endoglukanase dari kedua bakteri dalam satu genus Bacillus yaitu B.subtilis dan B.thuringiensis masih memiliki karakter yang hampir mirip. Pada aspek motif protein, motif-motif pendek yang ditemukan merepresentasikan kemiripan yang sangat tinggi, hal ini mengindikasikan kekerabatan gen yang dekat diantara kedua sekuen protein tersebut.