1
DIKTAT KULIAH
OLEH
I MADE DIRA SWANTARA
PROGRAM STUDI MAGISTER KIMIA TERAPAN PROGRAM PASCASARJANA
UNIVERSITAS UDAYANA
DENPASAR 2015
2
PRAKATA
Pengetahuan tentang sterol yang berasal dari biota laut khususnya sterol pada ganggang laut makin dirasakan manfaatnya bagi mahasiswa yang menekuni bidang kimia organik bahan alam dan bidang kesehatan. Hal ini disebabkan sterol pada ganggang laut merupakan sumber sterol pada invertebrata yang memakan ganggang laut tersebut. Selain itu, kalau dipandang dari penggunaan sterol sebagai bahan dasar pembuatan obat-obat turunan hormon steroid, maka sterol yang bersumber dari biota laut khususnya dari ganggang laut mempunyai peranan yang sangat penting karena sumber sterol pada organisme darat semakin menipis.
Mengingat buku maupun bahan kuliah lainnya seperti diktat yang membahas tentang sterol yang berasal dari biota laut secara umum maupun yang berasal dari ganggang laut dalam bahasa Indonesia belum banyak, maka penulis memberanikan diri menyusun diktat Sterol Ganggang Laut ini. Buku ini dapat digunakan sebagai penunjang mata kuliah Kimia Bahan Alam yang diberikan di Program Magister Kimia Terapan, Universitas Udayana.
Isi diktat ini mencakup uraian umum tentang sterol serta sumber dan variasi sterol pada ganggang laut. Diktat ini tidak akan muncul bila tidak ada dorongan dari teman sejawat. Kepada mereka, penulis mengucapkan banyak terima kasih. Sudah terang, tulisan manusia tidak luput dari kekurangan. Karena itu, penulis sangat mengharapkan kritik dan saran dari sejawat dan para ahli, dalam rangka penyempurnaan diktat ini.
Denpasar, Nopember 2015. Penulis
3
DAFTAR ISI
Hlm. Sampul ……… 1 Prakata ………... 2 Daftar Isi ………... 3 Bab I STEROL ……… 4 1.1 Pendahuluan ……… 4 1.2 Klasifikasi Sterol ………. 4 1.3 Biosintesis Sterol ………. 7 1.4 Biokonversi Sterol ………... 11 1.5 Isolasi Sterol ……… 151.6 Penentuan Struktur Molekul Senyawa Sterol Secara Spektroskopi………. 19
BAB II GANGGANG LAUT ……… 27
2.1 Pendahuluan ……… 27
2.2 Tumbuhan Laut ………... 27
2.3 Tumbuhan Talus ………. 30
2.3.1 Myxophyceae (Ganggang hijau-biru) ……….. 31
2.3.2 Chlorophyceae (Ganggang hijau) ……… 32
2.3.3 Phaeophyceae (Ganggang coklat) ……… 39
2.3.4 Rhodophyceae (ganggang merah) ……… 46
2.3.5 Chrysophyceae (Ganggang hijau-kuning) ……… 53
BAB III STEROL PADA GANGGANG LAUT ……… 58
3.1 Pendahuluan ……… 58
3.2 Sterol Pada Ganggang ………. 60
3.3 Sterol Pada Ganggang Merah ……….. 62
3.4 Sterol Pada Ganggang Hijau ………... 63
3.5 Sterol Pada Ganggang Coklat ………. 65
4
BAB I
STEROL
1. 1 Pendahuluan
Kata sterol berasal dari bahasa Yunani yang terdiri atas kata stereos + ol; stereos
berarti padatan. Sterol pada umumnya terdapat pada bagian yang tidak tersabunkan dari jaringan tumbuhan dan hewan (Fieser & Fieser, 1950). Menurut Robinson (1991), sterol adalah suatu senyawa steroid yang mempunyai rantai samping delapan atom karbon dan mengandung gugus hidroksil pada atom C-3. Di dalam “Kamus Lengkap Kimia” disebutkan bahwa sterol adalah sekelompok steroid berdasar alkohol dan mempunyai rantai samping hidrokarbon yang terdiri atas 8-10 atom karbon (Daintith, 1994). William Nes (dalam Goad & Akihisa, 1997) mengemukakan bahwa sterol adalah steroid terhidroksilasi yang struktur rantai sampingnya mempertahankan beberapa atau seluruh atom karbon skualen. Definisi sterol yang lebih tepat dikemukakan oleh David Nes (dalam Goad & Akihisa, 1997) sebagai berikut ini. Sterol adalah senyawa isopentenoid tetrasiklik kiral yang terbentuk melalui proses siklisasi skualen oksida dan mempertahankan gugus polar (seperti hidroksil atau keto) pada C-3. Proses siklisasi terjadi melalui senyawa peralihan yang memiliki stereokimia dengan konfigurasi trans-syn-trans-anti-trans-anti.
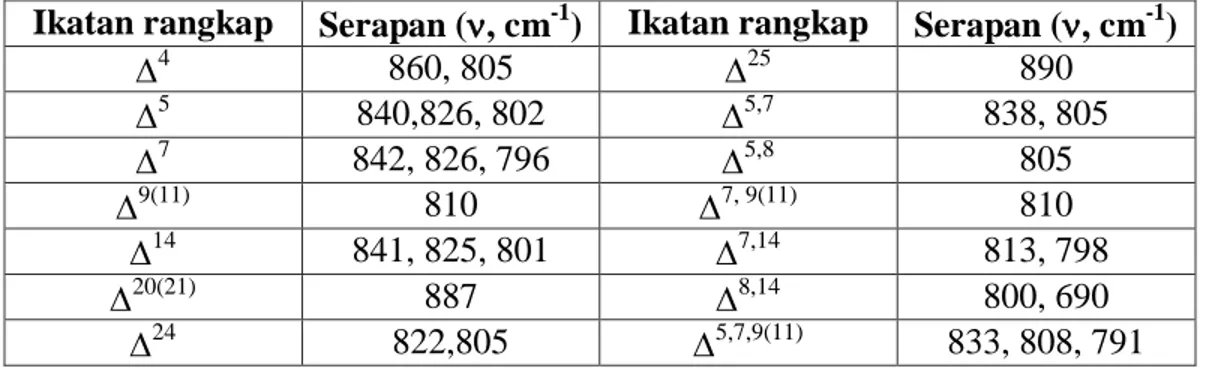
1. 2 Klasifikasi Serol
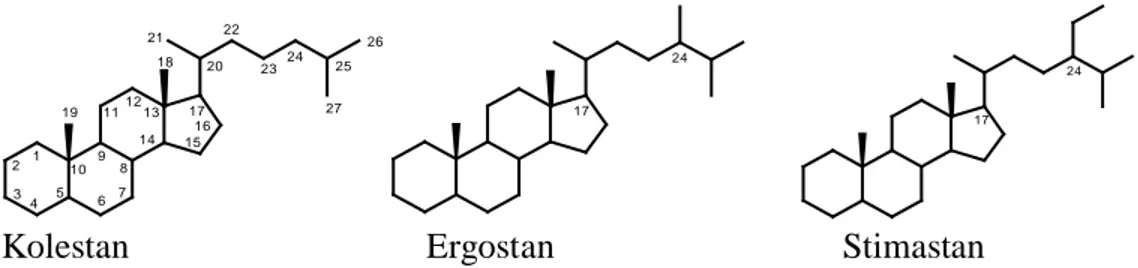
Berdasarkan kerangkanya, senyawa turunan sterol dapat dibagi menjadi tiga kelompok yaitu: kolestan, ergostan, dan stigmastan seperti terlihat pada Gambar 1.1. Ketiga kelompok ini berbeda pada atom C-24 dalam hal apakah atom C tersebut tidak
5
bercabang, bercabang metil, atau bercabang etil. Senyawa turunan sterol dengan atom C-24 tidak bercabang dikelompokkan ke dalam kolestan, yang bercabang metil ke dalam ergostan, dan bercabang etil ke dalam stigmastan (Manito, 1981).
Kolestan Ergostan Stimastan
Gambar 1.1 Struktur tiga kerangka sterol: kolestan, ergostan, stigmastan
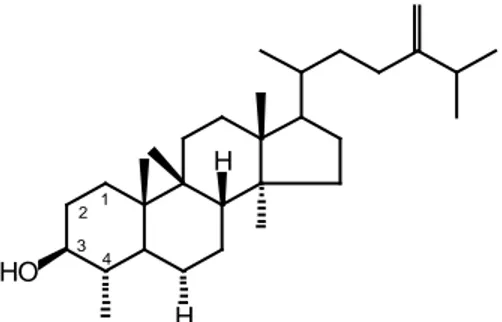
Ketiga kerangka sterol tersebut pada dasarnya mempunyai substituen hidroksil atau keto pada C-3 (Goad & Akihisa, 1997). Jika terdapat substituen hidroksil atau oksigen yang lain, maka substituen itu akan tersubstitusi pada atom karbon yang lain selain C-3, baik pada inti steroid maupun pada rantai sampingnya. Keberadaan hidroksil atau oksigen pada kerangka kolestan ditunjukkan pada dua senyawa di bawah ini (Gambar 1.2) yang merupakan hasil penelitian Sheu et al. (1996).
24,25-Epoksikolest-5-en-3β-ol 24ξ-Hidroperoksikolesta-4,25-dien-6β-ol-3-on Gambar 1.2 Kerangka kolestan yang tersubstitusi gugus hidroksil atau oksigen selain
pada atom C-3. O HO O OH OOH 17 24 17 24 17 24 1 2 3 4 5 6 7 8 9 10 111213 14 15 16 18 19 20 21 22 23 25 26 27
6
Keberadaan hidroksil atau oksigen pada kerangka ergostan ditunjukkan pada dua senyawa di bawah ini (Gambar 1.3) yang merupakan hasil penelitian Schrofder et al. (1980).
Ergost-5-en-3β,7α-diol Ergost-5-en-3β,7-on
Gambar 1.3 Kerangka ergostan yang tersubstitusi gugus hidroksil atau oksigen selain pada atom C-3.
Selanjutnya, keberadaan hidroksil atau oksigen pada kerangka stigmastan ditunjukkan pada dua senyawa di bawah ini (Gambar 1.4) (Schrofder et al.,1980).
Stigmast-5-en-3β,7β-diol Stigmast-5-en-3β-ol,7-on
Gambar 1.4 Kerangka stigmastan yang tersubstitusi gugus hidroksil atau oksigen selain pada atom C-3.
HO O
HO OH
7 1. 3 Biosintesis Sterol
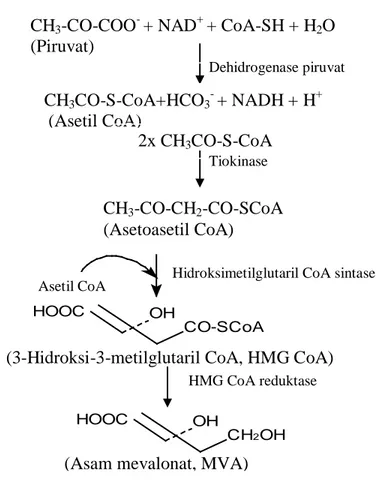
Tahap pertama pada biosintesis sterol diawali oleh piruvat sampai menghasilkan senyawa antara, yaitu asam mevalonat. Dengan adanya nikotinamid adenin dinukleotida (NAD+)dan katalis dehidrogenase piruvat, maka piruvat berubah menjadi asetil koenzim A (asetil CoA). Dua molekul asetil CoA dengan katalis tiokinase membentuk asetoasetil CoA yang selanjutnya menjadi 3-hidroksi-3-metilglutaril CoA (HMG CoA) dengan katalis hidroksimetilglutaril CoA sintase. HMG CoA dengan katalis HMG CoA reduktase diubah menjadi asam mevalonat (MVA) (Lehninger, 1982; Goad & Akihisa, 1997). Tahapan biosintesis ini ditunjukkan pada Gambar 1.5.
Gambar 1.5 Tahap pembentukan asam mevalonat pada biosintesis sterol (Lehninger, 1982; Goad & Akihisa, 1997).
CH3CO-S-CoA+HCO3- + NADH + H+
(Asetil CoA)
2x CH3CO-S-CoA
CH3-CO-CH2-CO-SCoA
(Asetoasetil CoA)
(3-Hidroksi-3-metilglutaril CoA, HMG CoA) CH3-CO-COO- + NAD+ + CoA-SH + H2O
(Piruvat)
Dehidrogenase piruvat
Tiokinase
Hidroksimetilglutaril CoA sintase Asetil CoA
HOOC OH
CO-SCoA
(Asam mevalonat, MVA)
CH2OH OH
HOOC
8
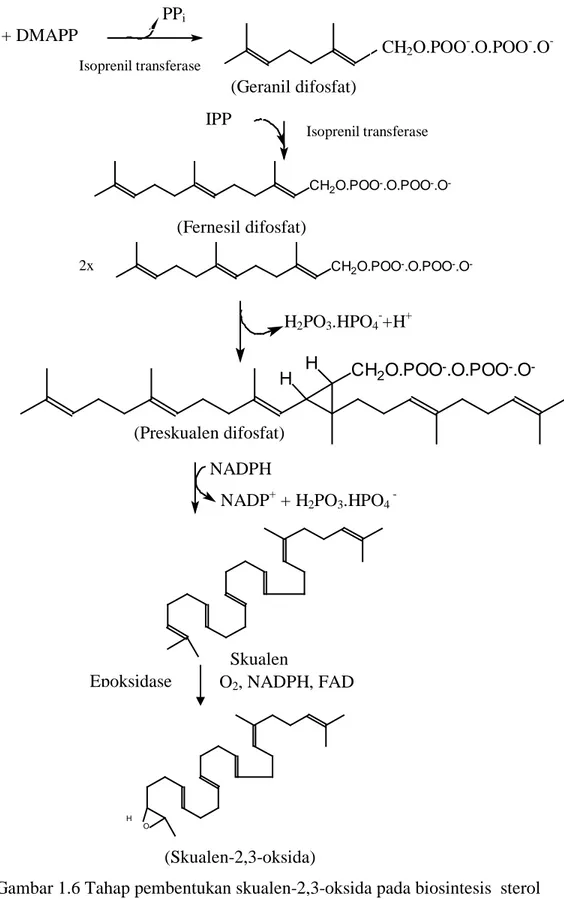
Tahapan reaksi biosintesis sterol selanjutnya adalah pengikatan dua gugus fosfat oleh mevalonat sehingga terbentuk mevalonat terfosforilasi. Mevalonat terfosforilasi yang terbentuk ini kemudian kehilangan gugus karbon dioksida dan satu molekul air, menghasilkan isopentenil difosfat (IPP) yang beresonansi dengan dimetilalil difosfat (DMAPP). Selanjutnya, molekul IPP dan DMAPP bergabung membentuk geranil difosfat yang dilanjutkan dengan pembentukan farnesil difosfat. Dua melekul farnesil difosfat membentuk preskualen difosfat. Dengan melibatkan NADPH, preskualen difosfat membentuk skualen, yang dilanjutkan dengan pembentukan skualen-2,3-oksida (Scheuer, 1973; Goad & Akihisa, 1997). Tahapan biosintesis ini ditunjukkan pada Gambar 1.6. CH2O-POO-.O.POO-O -HOOC OH CH2OH HOOC OH Mevalonat kinase Mevalonat fosfat kinase
(Mevalonat terfosforilasi)
Mevalonat difosfat anhidrodekarboksilase
CO2 + H2O
CH2O.POO-.O.POO- CH2O.POO-.O.POO-.O
9 (Skualen-2,3-oksida)
Gambar 1.6 Tahap pembentukan skualen-2,3-oksida pada biosintesis sterol (Scheuer, 1973; Goad & Akihisa, 1997).
PPi CH2O.POO-.O.POO-.O -Isoprenil transferase (Geranil difosfat) IPP + DMAPP CH2O.POO-.O.POO-.O -(Fernesil difosfat) Isoprenil transferase IPP CH2O.POO-.O.POO-.O -2x H2PO3.HPO4-+H+ NADPH NADP+ + H2PO3.HPO4 -CH2O.POO-.O.POO-.O -H H (Preskualen difosfat) O H Skualen
10
Tahap terakhir dari biosintesis sterol dalam fungi dan hewan adalah siklisasi skualen-2,3-oksida untuk membentuk lanosterol, yang dikatalisis oleh 2,3 oksidoskualen-lanosterol siklase. Selanjutnya, dari oksidoskualen-lanosterol akan terbentuk kolesterol melalui pelepasan tiga gugus metil. Sementara itu, pada organisme yang melakukan fotosintesis, siklisasi skualen-2,3-oksida akan membentuk sikloartenol yang selanjutnya membentuk fitosterol (Goad & Akihisa, 1997). Tahapan siklisasi skualen-2,3-oksida ditunjukkan pada Gambar 1.7
(Skualen-2,3-oksida)
(Ion karbonium protoskualen) (a) (b)
(Lanosterol) (Sikloartenol) Kolesterol Fitosterol Gambar 1.7 Tahap siklisasi skualen-2,3-oksida pada biosintesis sterol (a) dalam fungi dan hewan, (b) dalam organisme fotosintesis (Goad &
Akihisa, 1997). HO H H H + O H HO HO
11 1. 4 Biokonversi Sterol
Senyawa golongan steroid dalam tubuh manusia pada umumnya bersifat hormonal. Contohnya adalah hormon korteks adrenal (glukokortikoida dan mineralokortikoida), androgen, estrogen, dan hormon yang aktif selama kehamilan seperti progesteron. Androgen, estrogen, dan progesteron digunakan sebagai obat. Turunan estrogen dan progesteron selain digunakan sebagai obat juga digunakan sebagai kontraseptik (Tarigan, 1991).
Sehubungan dengan meningkatnya kebutuhan akan obat turunan steroid, maka diperlukan bahan baku steroid alam dalam jumlah yang lebih besar. Namun, persediaan bahan baku steroid seperti diosgenin, yang diperoleh dari Dioscorea composita, Coctus sp., dan Testudinaria sylvatica tidak banyak tersedia. Untuk mengatasi kekurangan diosgenin ini dilakukan studi yang intensif mengenai kemungkinan penggunaan sterol yang tersedia cukup banyak di alam untuk menggantikan diosgenin.
Kolesterol dapat diperoleh dari hewan, dan dalam jumlah besar dapat diperoleh dari hasil pencucian lemak bulu domba. Sitosterol serta stigmasterol diperoleh dari kedelai dan kampesterol dapat dihasilkan dalam jumlah besar sebagai hasil sampingan dari industri kertas (Tarigan, 1991). Karena sterol merupakan salah satu komponen lipida dalam hewan dan tumbuhan (kolesterol dalam hewan dan sitosterol serta stigmasterol dalam tumbuhan), maka senyawa turunan sterol dapat tersedia secara melimpah pada hewan dan tumbuhan.
Pencarian sumber sterol dan berbagai senyawa turunan sterol lebih intensif dilakukan sejak diketahui bahwa rantai samping berbagai sterol alam dapat diputus secara selektif oleh mikrorganisme (Amini, 1999). Kolesterol, kampesterol, sitosterol, dan stigmasterol atau campurannya dapat dijadikan bahan dasar atau bahan baku untuk
12
pembentukan beberapa senyawa antara seperti 1,androstadien-3,17-dion (1,4 ADD), 4-androsten-3,17-dion (AD), progesteron, dan senyawa antara yang lainnya. Selanjutnya, berbagai senyawa antara ini dikonversi menjadi obat-obat turunan steroid. Sebagai contoh kolesterol dapat dikonversi menjadi senyawa ∆5 pregnan-3β-ol-20-on yang dilanjutkan dengan pembentukan progesteron. Progesteron ini dapat diubah menjadi hormon korteks adrenal, testosteron, dan estradiol. Estradiol dapat diubah menjadi estron dengan bantuan Actinomyces albus atau Proactinomyces sp. (Capek et al., 1966). Hasil penelitian yang dilakukan oleh Nagazawa pada tahun 1969 menunjukkan bahwa biokonversi kolesterol menjadi 1,4 ADD dilakukan oleh Arthrobacter simplex dengan hasil 15% (Tarigan, 1991).
Mamoli dan Vercellone (1937) mengkonversi senyawa steroid secara mikrobiologi dengan cara hidrogenasi atau dehidrogenasi (Amini, 1999). Penelitian selanjutnya dilakukan oleh Turfitt pada tahun 1948, yaitu biotransformasi kolesterol menjadi asam 4-dehidroetiokolonat dengan menggunakan Nocardia sp. Kramli dan Hovath pada tahun 1948 dan 1949 membuat 7-hidroksikolesterol dari kolesterol dengan menggunakan Nocardia resens seperti yang ditampilkan pada Gambar 1.8 (Tarigan, 1991). Nocardia resens
7-hidroksokolesterol Kolesterol
Asam 4-Dehidroetiokolonat Gambar 1.8 Biokonversi kolesterol menjadi asam 4-hidroetiokolonat dan 7-
hidroksikolesterol (Tarigan, 1991).
HO
HO OH
O
13
Dengan cara biokonversi, kolesterol dapat juga diubah menjadi kolestenon dengan bantuan Azotobacter sp., Proactinomyces roseus, Proactinomyces erythropolis, atau
Flavobacterium maris. Sementara itu, pregnenolon dapat dikonversi menjadi progesteron
oleh Aspergillus atau Actinomyces (Capek et al., 1966). Progesteron sebagai senyawa antara untuk pembuatan obat-obatan turunan steroid telah mampu diproduksi dalam jumlah banyak dan harganya relatif murah. Progesteron ini merupakan hasil biokonversi stigmasterol (Tarigan, 1991).
Degradasi senyawa 19-hidroksikolestenon oleh Nocardia restrictus ATCC (American Type Culture Collection) 14887 atau Nocardia sp. ATCC 19170 menghasilkan estron sebanyak 30%. Degradasi ini tidak sempurna karena estron yang dihasilkan bersifat toksik terhadap mikroorganisme pendegradasinya, sehingga metabolisme selanjutnya tidak dapat berlangsung. Senyawa 3β -asetoksi-hidroksikolestan adalah substrat yang lebih baik dibandingkan dengan 19-hidroksikolestenon, karena dapat dikonversi menjadi estron oleh Nocardia sp. ATCC 19170 dengan hasil kira-kira 79% dalam 96 jam. Estron juga dapat diperoleh dari senyawa 19-nor-kolesta-1,3,5(10)-trien-3-ol dan 19-hidroksi-4,7-kolestadien-3-on dengan bantuan Nocardia restrictus ATCC 14887. Selain Nocardia sp., beberapa spesies mikroorganisme juga dapat mengkonversi 19-nor-hidroksisterol atau 19-hidroksisterol dan beberapa senyawa turunan sterol lainnya menjadi estron. Sebagai contoh,
Mycobacterium sp. dapat mengkonversi 19-hidroksi-4,7-kolestadien-3-on menjadi estron
(20%) dan ekuilin (16%). Demikian juga Corynebacterium simplex dan Nocardia rubra
14
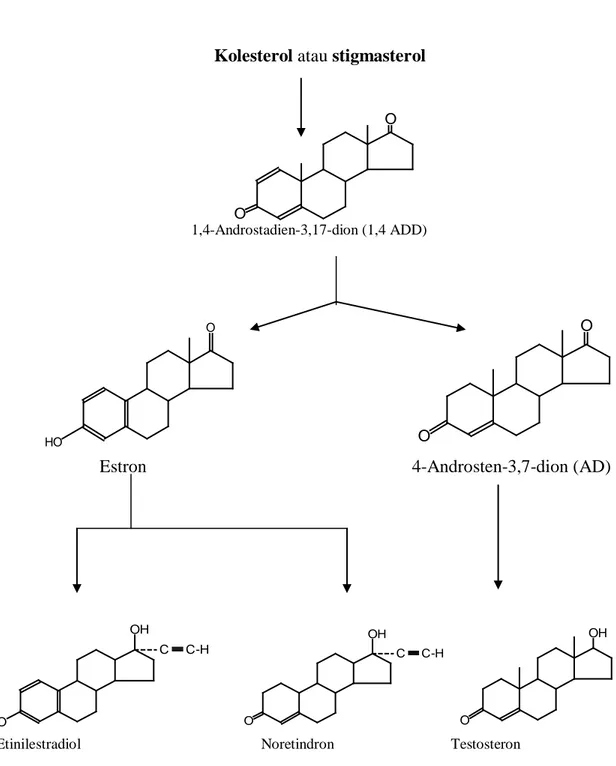
1999). Skema sintesis estron, 19-norsteroid, dan testosteron dari kolesterol atau stigmasterol ditampilkan pada Gambar 2.9 (Tarigan, 1991).
Kolesterol atau stigmasterol
1,4-Androstadien-3,17-dion (1,4 ADD)
Estron 4-Androsten-3,7-dion (AD)
Etinilestradiol Noretindron Testosteron
Gambar 1.9 Skema sintesis estron, 19-norsteroid, dan testosteron dari kolesterol atau stigmasterol (Tarigan, 1991).
O O O O HO O C-H C OH O C C-H OH HO O OH
15 1. 5 Isolasi Sterol
Pengisolasian senyawa turunan sterol merupakan pekerjaan yang memerlukan beberapa tahapan. Masing-masing tahapan tersebut mempunyai beberapa persyaratan yang harus diperhatikan. Tahapan isolasi sterol yang harus dikerjakan adalah ekstraksi, pemisahan, dan pemurnian.
1. 5. 1 Metode ekstraksi
Ektraksi merupakan suatu cara untuk menarik seluruh atau sebagian metabolit yang terdapat di dalam sampel. Ekstraksi metabolit sebaiknya dilakukan dalam keadaan segar. Akan tetapi, dalam hal tertentu seperti lokasi pengambilan sampel yang tidak memungkinkan untuk mengekstraksi langsung, maka ekstraksi dapat dilakukan dalam keadaan tidak segar. Beberapa persyaratan ekstraksi agar memberikan hasil yang maksimal antara lain, teknik preparasi sampel, jenis pelarut yang digunakanm, dan kondisi saat melakukan ekstraksi.
Menurut Goad & Akihisa (1997), preparasi sampel yang sering dilakukan untuk ekstraksi sterol ada beberapa cara.
a. Bahan segar langsung diekstraksi setelah dipotong atau dihaluskan. b. Bahan segar dihomogenkan dalam pelarut pengekstraksi dengan blender. c. Bahan segar mula-mula dibekukan lalu dihomogenkan dengan pelarut.
d. Bahan mula-mula dikeringkan pada oven atau kering angin, kemudian dihaluskan dan diekstraksi atau dihomogenkan dalam pelarut pengekstraksi.
16
Bahan yang berasal dari tumbuhan sering langsung diekstraksi tanpa preparasi seperti dipotong, dikeringkan, dan dihaluskan. Akan tetapi pada bahan yang berasal dari jaringan hewan umumnya dihomogenkan.
Pelarut yang digunakan untuk ekstraksi sterol harus mencakup rentang polaritas yang luas dari non-polar seperti heksan sampai pada kepolaran yang tinggi seperti metanol. Pelarut tersebut dapat berupa pelarut tunggal atau campuran. Pada beberapa kondisi, ekstraksi dilakukan dalam dua atau beberapa pelarut secara berurutan. Pelarut yang biasa digunakan untuk ekstraksi adalah kloroform, aseton, metanol, campuran kloroform-metanol, etanol, propanol, heksan atau petroleum eter (PE) (Goad & Akihisa, 1997).
Ekstraksi lipid secara umum menggunakan satu atau kombinasi metode berikut ini (Goad & Akihisa, 1997).
a. Bahan dikocok satu atau beberapa kali dengan pelarut pengekstraksi pada temperatur kamar atau pada pemanasan 40oC. Ekstraksi dilakukan selama 30 menit sampai satu jam atau untuk waktu yang lama (di atas tiga minggu).
b. Bahan direfluks selama 2-18 jam dengan pelarut pengekstraksi tunggal atau direfluks berturut-turut dengan pelarut yang berbeda polaritasnya.
c. Bahan diekstraksi dengan alat sokslet selama 2-48 jam dengan menggunakan pelarut tunggal atau dengan beberapa pelarut yang polaritasnya semakin tinggi.
d. Bahan dihomogenkan dalam pelarut pengekstraksi.
17
Ekstraksi dengan sokslet sering digunakan pada bahan yang berasal dari tumbuhan. Akan tetapi, jika bahan yang berasal dari jaringan hewan umumnya dihomogenkan dalam pelarut pengekstraksi. Bahan yang berasal dari organisme bersel tunggal, fungi, atau alga, ekstraksinya dilakukan di dalam pengocok ultrasonik.
Kesempurnaan ekstraksi tumbuhan yang mengandung pigmen dapat diamati melalui perubahan warna ekstrak. Akan tetapi, ekstraksi jaringan hewan (yang mengandung sedikit pigmen) dianggap sempurna apabila sudah dilakukan tiga atau empat kali pergantian pelarut (Goad & Akihisa, 1997).
Lipid total dapat diekstraksi dengan menggunakan pelarut dengan polaritas tinggi seperti metanol, etanol, aseton, atau campuran kloroform-metanol. Pelarut yang kurang polar seperti kloroform, dietil eter, atau etil asetat digunakan untuk melarutkan lipid non-polar bersama dengan sterol bebas, sterol teresterifikasi, dan senyawa kurang non-polar lainnya. Ekstrasi lipid yang berasal dari mikroorganisme, organisme lautan, dan jaringan tumbuhan pada beberapa kondisi ekstraksi harus menggunakan hidrolisis basa langsung pada bahan segar atau kering. Proses ini dikenal dengan saponifikasi (penyabunan). Proses saponifikasi pada umumnya menggunakan larutan kalium hidroksida atau natrium hidroksida dalam metanol atau etanol dengan cara direfluks. Dalam proses saponifikasi tersebut, akan terjadi hidrolisis gugus yang teresterifikasi. Hidrolisis tersebut pada umumnya terjadi pada sterol ester, triasilgliserol, dan fosfolipid sehingga menghasilkan lipid yang tidak tersabunkan (Goad & Akihisa, 1997).
Ekstraksi bahan yang berasal dari organisme lautan sangat efisien bila menggunakan teknik ekstraksi pada temperatur rendah. Teknik ini khusus untuk isolasi sterol teresterifikasi asam lemak, sebab asam lemak tak jenuh kemungkinan rentan
18
terhadap degradasi oksidasi. Pada kondisi ekstraksi ini, banyak sterol yang relatif stabil kecuali sterol yang mengandung ikatan rangkap pada C-5,7.
Kondisi pelarut yang digunakan untuk ekstraksi lipid dengan sistem campuran pelarut memperlihatkan bahwa campuran kloroform-metanol (2:1) dan heksan-dietil eter (4:1) adalah pelarut yang baik untuk ekstraksi lipid non-polar. Campuran pelarut etanol-dietil eter-air (2:2:1) merupakan pelarut yang lebih polar dibandingkan dengan kloroform-metanol, tetapi bisa juga digunakan untuk ekstraksi lipid non-polar. Metanol dan etanol adalah pelarut polar, dapat mengekstraksi semua lipid polar dan non-polar, tetapi pelarut ini efektif untuk mengekstraksi fosfolipid (Robinson, 1991; Goad & Akihisa, 1997).
1. 5. 2 Pemisahan Sterol
Ekstrak lipid sering langsung dikromatografi untuk memperoleh sterol terkonyugasi seperti sterol ester, sterol glikosida, atau sterol sulfat. Jika ingin memperoleh sterol bebas, ekstrak lipid non-polar dihidrolisis dengan cara merefluks ekstrak tersebut dengan larutan kalium hidroksida atau natrium hidroksida dalam metanol atau etanol. Selanjutnya, hidrolisat diekstraksi dengan pelarut yang kurang polar untuk melarutkan lipid yang tidak tersabunkan termasuk sterol bebas (Goad & Akihisa, 1997).
Menurut Robinson (1991) dan Goad & Akihisa (1997), fraksionasi lipid total dan lipid non-polar sering menggunakan teknik kromatografi kolom dengan fase diam silika gel atau alumina. Jika sampel yang difraksionasi dalam jumlah sedikit, maka digunakan teknik kromatografi lapis tipis. Proses kromatografi kolom dapat dilakukan beberapa kali. Kromatografi kolom pertama digunakan untuk memisahkan lipid non-polar dari
19
campurannya sebelum dilanjutkan dengan pemisahan masing-masing sterol. Dengan berkembangnya teknik kromatografi cair kinerja tinggi (KCKT), pemisahan sterol dapat dilakukan relatif lebih cepat dengan hasil yang lebih baik jika dibandingkan dengan kromatografi kolom. Biasanya KCKT digunakan untuk tahap pemisahan terakhir karena fase diamnya relatif mahal (Goad & Akihisa, 1997).
1. 6. Penentuan Struktur Molekul Senyawa Sterol Secara Spektroskopi
Senyawa turunan sterol hasil isolasi yang sudah merupakan senyawa tunggal dapat ditentukan strukturnya dengan cara mengambil beberapa spektrum senyawa tersebut pada beberapa spektrometer. Spektrum yang umum digunakan untuk menentukan atau mengelusidasi struktur suatu senyawa metabolit sekunder seperti sterol adalah: spektrum inframerah, spektrum ultraviolet, spektrum massa, dan spektroskopi resonansi magnet inti (satu dan dua dimensi).
1. 6. 1 Spektroskopi inframerah
Sebagian besar vibrasi regang C-H pada sterol menghasilkan serapan kuat pada bilangan gelombang (ν) 2800-3000 cm-1. Ikatan C=C olefinik menghasilkan serapan lemah pada ν 1620-1650 cm-1. Jika ikatan rangkap tersebut terkonyugasi, maka serapannya akan bergeser dari ν 1620-1650 ke ν 1575-1630 cm-1. Sterol yang memiliki tiga ikatan rangkap dua memberikan serapan tekuk C-H yang lemah pada ν 790-850 cm
-1
. Sterol dengan ikatan rangkap pada C-5 memberikan serapan pada ν 840 dan 802 cm-1 dengan intensitas yang sama dan serapan lemah pada ν 826 cm-1. Jika ikatan rangkap dua tersebut terletak pada C-7, serapan pada ν 826 cm-1 akan sama intensitasnya dengan ν 840 cm-1. Sterol dengan ikatan rangkap terkonyugasi pada C-5,7 memberikan serapan
20
pada ν 840 cm-1 yang lebih kuat dari serapan ν 802 cm-1 (Hanson, 1968; Goad & Akihisa, 1997). Serapan spektrum inframerah pada sterol yang mengandung ikatan rangkap ditampilkan dalam Tabel 1.1 (Goad & Akihisa, 1997).
Tabel 1.1 Serapan inframerah C-H tekuk untuk ikatan rangkap dua pada sterol (Hanson, 1968; Goad & Akihisa, 1997).
Ikatan rangkap Serapan (ν, cm-1) Ikatan rangkap Serapan (ν, cm-1)
∆4 860, 805 ∆25 890 ∆5 840,826, 802 ∆5,7 838, 805 ∆7 842, 826, 796 ∆5,8 805 ∆9(11) 810 ∆7, 9(11) 810 ∆14 841, 825, 801 ∆7,14 813, 798 ∆20(21) 887 ∆8,14 800, 690 ∆24 822,805 ∆5,7,9(11) 833, 808, 791
Serapan regang O-H terlihat pada ν 3600-3650 cm-1, serapan regang C-O dari 3α/β-OH khas pada ν 990-1060 cm-1. Sterol dengan cincin A/B jenuh dan tersubstitusi gugus karboksil pada C-3 memberikan serapan pada ν 1704 - 1722 cm-1.
1. 6. 2 Spektroskopi Ultraviolet
Panjang gelombang dengan serapan maksimum (λmaks) dan harga koefisien
absorpsi molar (ε) suatu sterol yang mengandung ikatan rangkap, tergantung pada posisi dan sifat ikatan rangkap tersebut. Ikatan rangkap terisolasi menghasilkan serapan maksimum pada panjang gelombang (λ) 190-220 nm (Hanson, 1968; Goad & Akihisa, 1997). Daerah panjang gelombang maksimum dan harga koefisien absorpsi molar untuk senyawa turunan sterol dengan ikatan rangkap terkonyugasi disajikan dalam Tabel 1.2.
21
Tabel 1.2 Serapan maksimum dan koefisien absorpsi molar (ε) sterol dengan ikatan rangkap terkonyugasi (Goad & Akihisa, 1997).
Posisi ikatan rangkap λmaks (nm) Koefisien absorpsi molar, ε ∆4,6 239 (232, 248)* 23500 ∆5,7 282 (271, 293) 11900 ∆5,7,9(11) 324 (311, 339) 11800 ∆5,7,14 319 16200 ∆5,7,9(11),14 304 (250, 259, 269) 21800 ∆6,8,14 245 31600 ∆7,9(11) 245 (236, 251) 11800 ∆7,14 242 10000 ∆8,14 248 18600 ∆22,24(28) 230 21400 ∆5,7,22,24(28) 281 (230, 271, 293) 12800
* Harga di dalam kurung menunjukkan λmaks bahu pada spektrum
1.6.3 Spektroskopi Massa
Pemilihan teknik ionisasi dalam spektroskopi massa tergantung pada sifat fisik dan kimia senyawa yang dianalisis. Teknik ionisasi elektron atau tumbukan elektron
(electron impact, EI) pada spektroskopi massa telah banyak digunakan untuk menentukan
berat molekul berbagai senyawa kimia.
Ion molekul senyawa sterol bebas pada spektrum massa dapat diamati secara jelas. Ion [M-18]+ dihasilkan dari pemenggalan gugus 3β-hidroksi sebagai molekul air. Pengulangan pelepasan molekul H2O mengindikasikan sterol yang terhidroksi pada posisi
lain selain pada C-3. Pemenggalan dan intensitas beberapa ion petunjuk dalam spektrum massa sterol bebas kadang-kadang sama dengan yang dihasilkan oleh turunan sterol. Penggalan khas lainnya diakibatkan oleh pelepasan gugus ROH yang menghasilkan penggalan [M-ROH]+. Khusus untuk sterol bebas 3β-hidroksi-5-en dengan rantai samping jenuh (swbagai contoh kolesterol) memberikan dua pola pemenggalan yang khas pada cincinnya yaitu, M-85 dan M-111 (Goad & Akihisa, 1997).
22
Sterol yang terderivatisasi TMS, penggalan kuat terjadi pada m/z 129 dan [M-129]+ akibat pemutusan cincin C, D, rantai samping, dan TMSOH (Goad & Akihisa, 1997). Pada umumnya, pemutusan rantai samping sebagai kelanjutan dari pemutusan 3β -hidroksi menghasilkan penggalan [M-rantai samping-42-ROH]+ dengan R = H untuk sterol bebas; R = AcO untuk sterol asetat; dan R = TMS untuk sterol TMS (Goad & Akihisa, 1997).
1. 6. 4 Spektroskopi Resonansi Magnet Inti
Spektrum resonansi magnet inti (RMI) untuk keperluan penentuan struktur tidak terdiri atas spektrum 1D (satu dimensi) saja. Apabila spektrum 1D tidak bisa menentukan letak dan jenis proton yang ada pada senyawa yang dianalisis, maka diperlukan spektrum RMI 2D untuk mengatasi keterbatasan spektrum RMI 1D tersebut.
1. 6. 4. 1 Resonansi magnet inti satu dimensi
Pengambilan spektrum RMI 1D untuk menentukan struktur senyawa kimia dapat dilakukan dalam beberapa tipe, di antaranya spektrum resonansi magnet inti proton (RMI-1H) dan resonansi magnet inti karbon (RMI-13C). Khusus untuk spektrum RMI-13C, dapat diambil dua teknik, yaitu spektrum broad band decoupled (spektrum bebas gandengan) dan spektrum distortionless enhancement by polarization transfer (DEPT). Spektrum RMI-13C bebas gandengan (broad band decoupled) memberikan informasi tentang jumlah minimal atom karbon dalam senyawa tersebut. Spektrum DEPT memberikan informasi tentang multiplisitas sinyal karbon yang teramati pada spektrum bebas gandengan, sehingga dapat ditentukan sinyal metil, metilen, dan metin (Dodrell et al., 1982).
23
Karakteristik resonansi senyawa sterol pada RMI adalah adanya resonansi metil angular pada spektrum RMI-1H dan RMI-13C serta resonansi karbon kuarterner jenuh pada spektrum RMI-13C. Kerangka sterol dalam spektrum RMI-1H memberikan dua puncak resonansi proton metil angular singlet pada geseran kimia (δ) 0,5-2,0 ppm. Demikian juga, spektrum RMI-13C memberikan dua puncak resonansi karbon metil angular (karbon primer) pada δ 10-35 ppm dan dua puncak karbon kuarterner jenuh δ 35-45 ppm (Hanson, 1968; Harlim, 1982; Ikan, 1991).
Dalam spektrum RMI-1H, kelompok metil rantai samping ketiga kerangka sterol (kolestan, ergostan, dan stigmastan) memberikan geseran kimia antara 0,5-1,0 ppm. Untuk membedakan ketiga kerangka tersebut, dapat dilihat jumlah serta multiplisitas puncaknya. Kerangka kolestan memberikan tiga puncak resonansi proton metil masing-masing doublet, ergostan empat puncak masing-masing-masing-masing doublet, dan stigmastan empat puncak dengan tiga puncak doublet dan satu puncak triplet.
1. 6. 4. 2 Resonansi magnet inti dua dimensi
Kegunaan spektrum resonansi magnet inti dua dimensi (RMI 2D) di samping untuk mengatasi keterbatasan informasi dalam spektrum RMI 1D juga diperlukan untuk mendukung penetapan struktur molekul. Secara umum, spektrum RMI 2D diperoleh berdasarkan dua tipe korelasi inti, yaitu korelasi homo dan heteronuklir. Beberapa percobaan standar untuk kedua tipe korelasi tersebut adalah spektrum H-H COSY,
heteronuclear multiple quantum correlation (HMQC), dan heteronuclear multiple bond
24 1. 6. 4. 2a Spektrum H-H COSY
Spektrum H-H COSY adalah kelompok spektrum korelasi homonuklir. Secara umum, spektrum H-H COSY memberikan informasi pasangan proton yang saling bergandengan dengan jarak dua ikatan (geminal proton) atau tiga ikatan (visinal proton). Dalam spektrum H-H COSY, jika sebuah garis horizontal yang ditarik dari sebuah puncak silang sampai pada kontur diagonal dan sebuah garis vertikal dari puncak silang yang sama sampai pada kontur diagonal yang lain, itu menunjukkan bahwa puncak silang tersebut terbentuk dari korelasi kedua kelompok proton pada kontur diagonal tersebut (Bax & Lerner, 1986). Dengan demikian, dapat dikatakan bahwa kedua kelompok proton tersebut saling bergandengan atau letaknya bertetangga. Sebagai contoh, korelasi H-H COSY dari proton-proton sikloeukalenol yang struktur molekulnya ditunjukkan pada Gambar 1.10 serta spektrum korelasinya pada Gambar 1.11. Pada korelasi tersebut ditunjukkan bahwa (a) adalah proton pada C-3; (b) proton proton pada C-4; (c) adalah proton α pada C-2; dan (c’) adalah proton β pada C-2
Gambar 1.10 Struktur molekul sikloeukalenol (Goad & Akihisa, 1997).
1 2 3 4 H H HO
25
Gambar 1.11 Spektrum H-H COSY sikloeukalenol: (a) proton pada C-3; (b) proton pada C-4; (c) proton α pada C-2; dan (c’) proton β pada C-2 (Goad & Akihisa, 1997).
Dengan demikian, spektrum H-H COSY dapat melengkapi spektrum RMI 1D dalam menentukan letak proton yang bertetangga.
1. 6. 4. 2b Spektrum HMQC dan HMBC
Spektrum HMQC adalah bagian dari spektrum korelasi heteronuklir. Korelasi proton dan karbon yang diperoleh dari spektrum HMQC adalah korelasi dengan jarak satu ikatan (Bax & Subramanian, 1986; Lerner & Bax, 1986). Pada spektrum HMQC, sumbu vertikal adalah geseran kimia RMI-1H dan sumbu horizontal adalah geseran kimia RMI-13C. Jika sebuah garis horizontal yang ditarik dari sebuah puncak silang sampai pada sebuah puncak spektrum RMI-1H dan sebuah garis vertikal yang juga ditarik dari sebuah puncak silang yang sama sampai pada sebuah puncak spektrum RMI-13C, itu menunjukkan bahwa puncak silang tersebut terbentuk dari korelasi proton dan karbon tersebut. Dengan demikian, dapat dikatakan bahwa proton tersebut berjarak satu ikatan
26
(terikat) pada karbon tersebut. Sama seperti spektrum HMQC, spektrum HMBC juga merupakan bagian dari spektrum korelasi heteronuklir, tetapi informasi yang diperoleh dari spektrum HMBC adalah korelasi antara proton dan karbon dengan jarak lebih dua atau tiga ikatan (Bax & Summer, 1986; Bax & Marion, 1988). Jika sebuah garis horizontal yang ditarik dari sebuah puncak silang sampai pada sebuah puncak spektrum RMI-1H dan sebuah garis vertikal yang juga ditarik dari sebuah puncak silang yang sama sampai pada sebuah puncak spektrum RMI-13C, itu menunjukkan bahwa puncak silang tersebut terbentuk dari korelasi proton dan karbon tersebut. Dengan demikian dapat dikatakan bahwa proton dan karbon yang membentuk puncak silang tersebut letaknya terpisah dua sampai tiga ikatan. Dengan demikian, dapat dikatakan bahwa spektrum HMQC dan HMBC dapat melengkapi spektrum RMI 1D untuk menetapkan posisi proton dan karbon.
27
BAB II
GANGGANG LAUT
2.1 PendahuluanTerkait dengan fokus pembahasan tentang sterol pada ganggang laut, tidak ada salahnya terlebih dahulu diulas masalah sumber dari sterol tersebut, yaitu ganggang laut. Pembahasan tentang ganggang laut ini dimaksudkan untuk memberikan wawasan tentang klasifikasi ganggang dengan tujuan agar para peneliti atau pemerhati masalah biota laut dapat mengetahui tempat objek yang dibicarakan dalam dunia tumbuhan. Selain itu, disinggung juga sebaran dan perkembangbiakannya dengan maksud agar diketahui lokasi tempat tersebar serta populasi objek tersebut.
Pembahasan tentang ganggang laut tidak bisa terlepas dari tumbuhan laut secara keseluruhan. Hal ini disebabkan karena ganggang laut terdiri atas beberapa kelas dari divisi tumbuhan talus (Thallophyta). Dengan demikian, dapat diketahui hubungan satu spesies ganggang dengan spesies ganggang yang lain.
2. 2 Tumbuhan Laut
Di laut, seperti di darat, tumbuh-tumbuhan merupakan produsen yang sesungguhnya. Artinya biota ini mampu membuat zat-zat organik yang majemuk dari senyawa-senyawa anorganik yang sederhana yang larut dalam air. Tanpa tumbuh-tumbuhan laut sebagai penghasil makanan primer, perkembangan kehidupan hewan laut umumnya tidak mungkin berjalan, kecuali hewan-hewan yang terdapat di sepanjang pantai dan estuari. Di tempat tersebut zat-zat organik yang berasal dari darat dapat terbawa ke laut untuk menunjang kehidupan mereka. Pada saat ini, telah diketahui bahwa
28
di dasar laut (abisal) terdapat juga berbagai hewan yang tidak tergantung pada kemampuannya untuk berfotosintesis, tetapi dapat menghasilkan makanan melalui proses kemosintesis. Perlu dicatat bahwa sepanjang menyangkut keberadaan bentuk dan jenis tumbuh-tumbuhan laut, agaknya laut bukan dunia mereka. Ragam bentuk dan jenis tumbuh-tumbuhan yang hidup di laut jauh lebih kecil daripada di darat. Demikian juga, jenis tumbuh-tumbuhan yang memegang peranan penting sebagai produsen primer berbeda antara laut dengan darat. Di laut, tumbuh-tumbuhan pemegang peranan utama sebagai penghasil makanan primer adalah mereka yang mengapung, khususnya fitoplankton, sedangkan di darat adalah sebaliknya; tumbuh-tumbuhan yang melekat atau tumbuh-tumbuhan berakar yang memegang peranan penting.
Munculnya perbedaan ini dapat dijelaskan sebagai berikut. Ada dua faktor utama yang mempengaruhi kehidupan tumbuh-tumbuhan laut. Yang pertama adalah cahaya yang diperlukan untuk fotosintesis dan kedua, substrat untuk melekat. Laut hanya menyediakan wilayah perairan yang memenuhi persyaratan itu, yakni mintakat eulitoral
(eulittoral), yang luasnya sekitar 2% luas lingkungan seluruhnya. Kecilnya bagian
perairan yang menyediakan cahaya dan substrat yang hanya 2% dari seluruh luas lautan itu membuat tumbuh-tumbuhan yang mengapung, yang jumlahnya sangat banyak, memegang peranan utama mengalahkan peranan tumbuh-yumbuhan melekat.
Di mintakat eulitoral, tumbuh-tumbuhah yang melekat berkembang dengan baik dan merajai populasi flora serta memegang peranan sebagai produsen makanan primer. Jika kita berjalan-jalan di sepajang pantai yang dasarnya berbatu-batu dan berkarang, sering dapat dilihat dasar pantai yang luas tertutup oleh permadani hijau yang berupa daun sla laut (ganggang laut genus Ulva). Namun, tidak semua dasar pantai itu dapat
29
atau cocok ditumbuhi tumbuh-tumbuhan spesies ganggang ini dan spesies ganggang melekat lainnya. Dasar berlumpur atau dasar berpasir lepas hanya dapat ditumbuhi oleh tumbuh-tumbuhan melekat tersebut jika di dasar perairan ini terdapat batu-batu besar yang menyembul di atas lumpur atau pasir. Batu-batu ini dapat dimanfaatkan untuk melekat spesies-spesies ganggang tertentu, karena ganggang tidak mempunyai akar yang berfungsi sebagai jangkar seperti halnya tumbuh-tumbuhan tinggi. Memang ada spesies ganggang yang mampu melekat di dasar pasir walaupun tidak ada batu-batu pelekat, misalnya anggur laut, Caulerpa. Ganggang ini dapat mengikat butiran-butiran pasir yang digunakan sebagai pemberat (balast) dengan alat pelekat (holdfast) yang bercabang-cabang panjang. Sebaliknya, dasar pantai yang berbatu-batu dan berbongkah karang yang tidak stabil dan sering bergerak-gerak digeser arus atau ombak, tidak cocok untuk kehidupan ganggang-ganggang melekat itu. Banyak dari ganggang melekat sifatnya efifitik (hidup melekat pada tumbuh-tumbuhan lain), dan epizoik (hidup melekat pada hewan laut tertentu). Namun pada umumnya mereka yang melekat sifatnya litofitik (hidup melekat pada batu).
Meskipun tumbuh-tumbuhan laut tidak begitu beraneka ragam, untuk mempelajari kehidupan mereka perlu kita ketahui pembagian taksonami kelompok tumbuhan-tumbuhan laut tersebut. Perlu diketahu bahwa taksonomi tumbuh-tumbuhan tidak tergantung pada taksonomi hewan, walaupun takson-takson tingkat menengah dan bawah sama dengan takson hewan seperti ordo, famili, genus, dan spesies. Di sini dunia tumbuhan dibagi menjadi empat divisi utama, yaitu Thallophyta (tumbuhan talus),
Bryophyta (tumbuhan lumut), Pteridophyta (tumbuhan paku), dan Spermatophyta
30
Thallophyta dan Spermatophyta yang terdapat di laut. Briophyta dan Pteridophyta khusus
tumbuh-tumbuhan darat (Romimohtarto dan Juwana, 1999).
2. 3 Tumbuhan Talus
Hampir semua kelompok tumbuh-tumbuhan laut termasuk dalam divisi ini. Sifat khas divisi ini adalah primitif, artinya badannya sedikit atau tidak terbagi-bagi dalam alat vegetatif seperti akar yang sebenarnya, ranting atau cabang, dan daun. Penting untuk diketahui bahwa yang termasuk divisi ini adalah ganggang laut dan bakteri laut. Akan tetapi karena bakteri laut merupakan bidang telaah yang luas, maka bakteri laut tidak dibahas dalam buku ini.
Sebagian besar ganggang laut berwarna indah dan ada pula yang bercahaya. Pigmen dari kromofor menyadap sinar matahari untuk fotosintesis. Atas dasar warna yang dimiliki oleh ganggang laut, yang berbeda antara satu kelompok dengan kelompok yang lain, maka pembagian kelas dari divisi tumbuhan talus ini mengikuti warna yang dimiliki. Kelas-kelas tumbuhan dari tumbuhan talus adalah:
Myxophyceae (ganggang hijau-biru),
Chlorophyceae (ganggang hijau),
Phaeophyceae (ganggang coklat),
Rhodophyceae (ganggang merah), dan
Chrysophyceae (ganggang hijau-kuning, termasuk diatomae).
Sebelumnya telah disebutkan bahwa warna memberi sifat kelas tumbuh-tumbuhan laut ini. Akan tetapi sifat-sifat lain yang berkaitan dengan struktur sel dan daur hidup lebih fundamental dalam membeda-bedakan kelima kelas ganggang laut tersebut. Setiap
31
kelompok mempunyai bentuk yang sangat beragam. Keempat kelas yang pertama yaitu
Myxophyceae, Chlorophyceae, Phaeophyceae, dan Rhodophyceae kecuali beberapa
ganggang biru, terdiri atas tumbuh-tumbuhan melekat, sedangkan ganggang hijau-kuning khas tumbuh-tumbuhan mengapung atau planktonik.
2. 3. 1 Myxophyceae (ganggang hijau-biru)
Kelas tumbuh-tumbuhan ini terdiri atas tumbuh-tumbuhan kecil yang kurang terorganisasi, beberapa di antaranya terdiri atas tumbuh-tumbuhan bersel tunggal dan lainnya bersel banyak. Warna tumbuh-tumbuhan ini disebabkan karena terdapatnya pigmen tambahan terlarut dalam air yang dinamakan fikosianin.
Laut merah mendapat nama demikian karena terdapatnya fitoplankton dari spesies
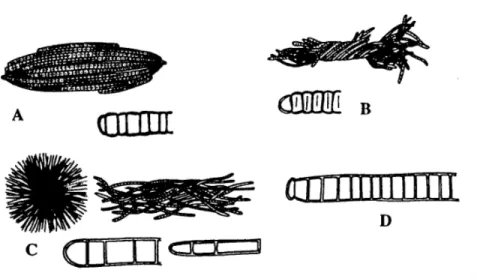
Trychodesmium erythraeum (Gambar 2.1) yang mempunyai pigmen tambahan berwarna
merah yang kadang-kadang menyebabkan permukaan air berwarna merah. Jadi, ganggang biru-hijau juga dapat berwarna merah.
32
Gambar 2.1 Beberapa spesies Trichodesmium ( Romimohtarto dan Juwana, 1999) A = Trichodesmium erythraeum
B = Trichodesmium contortum
C = Trichodesmium thiebauti
D = Trichodesmium hildebrantii
Dinding sel dari kelompok tumbuh-tumbuhan ini biasanya terdiri atas bahan kitin, bukan selulosa seperti yang dimiliki oleh tumbuh-tumbuhan lain. Karena itu, mereka menambah persediaan kitin yang dihasilkan di laut, khususnya oleh Crustecea, yakni kelas hewan yang antaranya terdiri atas udang dan kepiting. Beberapa Myxophyceae
sifatnya endophytic, yaitu mereka hidup di dalam tubuh tumbuh-tumbuhan lain dalam suatu asosiasi yang dinamakan simbiosis. Misalnya, dalam sel diatomae, Rhizosolenia, mungkin hidup ganggang spesies Richelia intracellularis.
2. 3. 2 Chlorophyceae (ganggang Hijau)
Sesuai dengan namanya, kelompok ganggang ini berwarna hijau. Pigmen dari kloroplas, yakni bentuk sel yang mengandung pigmen untuk fotosintesis, mencakup dua jenis klorofil, yaitu klorofil-a dan klorofil-b dan berbagai karotenoid. Warna kuning dan
33
oranye dari pigmen karotenoid tertutup oleh berlimpahnya klorofil yang berwarna hijau (Smith, 1955; Tjitrosoepomo, 1991). Berbeda dengan ganggang hijau-biru yang dinding selnya terbuat dari kitin, ganggang hijau dinding selnya terdiri atas karbohidrat berselulosa, lain dengan produk nitrogen berupa kitin. Beberapa ganggang hijau dari laut, seperti Halimedia (Gambar 2.2) menghasilkan kerak kapur (CaCO3). Karena itu,
organisme ini dapat memberikan sumbangan yang sangat berarti bagi terbentuknya endapan kapur di beberapa bagian perairan laut, terutama di daerah tropis. Sendi-sendi dari spesies Halimedia ini tidak berkapur, yang menyebabkan lentur dan ganggang ini dapat bergerak-gerak dalam air jika air bergerak. Kelas ganggang ini mempunyai bentuk sangat beragam, tetapi bentuk umum yang dijumpai adalah bentuk filamen (seperti benang) dengan septa (sekat) atau tanpa sekat, dan berbentuk lembaran (Tjitrosoepomo, 1991; Kadi, 1996).
Perkembangbiakan seksual kelas ganggang ini dilakukan seperti berikut. Isi dari suatu sel biasa dari tumbuh-tumbuhan yang pipih dan berlapis dua membentuk sel kelamin yang disebut gamet berbulu-getar dua. Setelah gamet ini lepas ke dalam air, mereka bersatu berpasangan dan melalui pembelahan sel berkembang menjadi tumbuh-tumbuhan baru yang dikenal sebagai sporofit, tetapi biasanya melalui fase benang dulu.
34
Gambar 2.2 Beberapa spesies Halimedia (Romimohtarto dan Juwana, 1999) A = Halimedia incrassata.
B = Halimedia monile.
C = Halimedia opuntia.
D = Halimedia fragilis.
E = Halimedia scabra.
Perkembangbiakan dapat juga dilakukan secara aseksual. Setiap sel biasa dari tumbuh-tumbuhan sporofit dapat membentuk zoospora yang berbulu-getar empat (spora adalah sel perkembangbiakan yang berbeda dengan biji, terutama karena sel ini tidak berisi embrio tumbuh-tumbuhan yang siap berkembang). Zoospora ini setelah dilepas tumbuh langsung menjadi gametofit, yakni tumbuh-tumbuhan yang menghasilkan gamet.
35
Prosesnya dikenal sebagai pergantian generasi dan terkait dengan ini adalah perubahan sitologi yang penting. Akan tetapi dalam hal Ulva, bentuk mikroskopik (sporofit dan gametofit) tidak dapat dibedakan, yakni mereka isomorfit (sporofit dan gametofit sama) atau homolog. Perkembangbiakan aseksual dapat pula terjadi dengan fragmentasi yang membentuk tumbuh-tumbuhan tak melekat.
Selama masa perkembangbiakan, sejumlah besar gamet dan zoospora dapat dilepaskan, meninggalkan tumbuh-tumbuhan induk tak berwarna dan membentuk perebakan warna hijau dalam air teluk yang tenang. Bagi banyak hewan penyaring makanan, gamet dan spora tumbuhan ini dan tumbuh-tumbuhan spesies lain menjadi sumber makanan yang tak boleh diabaikan dalam kita mempelajari makanan dari hewan-hewan litoral. Di teluk-teluk, fase renang dari ganggang dan lendir ganggang ikut membentuk lapisan tipis utama dan menjadi tumbuh-tumbuhan pengotor pada kapal dan benda-benda terendam lainnya.
Ganggang hijau terdapat terutama di mintakat litoral bagian atas, khususnya di belahan bawah dari mintakat pasut, dan tetap di daerah bawah-pasut sampai ke jelukan 10 meter atau lebih. Jadi, di habitat yang mendapat penyinaran matahari bagus, ganggang dari kelas ini terdapat melimpah di perairan hangat (tropis). Di laut Kutub Utara, ganggang hijau ini lebih jarang ditemukan dan bentuknya kerdil.
Di Indonesia, tercatat sedikitnya 12 genus ganggang hijau, yang banyak di antaranya sering dijumpai di perairan pantai. Berikut ini disampaikan beberapa genus ganggang hijau tersebut (Romimohtarto dan Juwana, 1999).
1. Caulerpa yang dikenal oleh penduduk sebagai anggur laut, terdiri atas 15 spesies dan
36
Gambar 2.3 Beberapa spesies Caulerpa (Romimohtarto dan Juwana, 1999). A = Caulerpa prolifera.
B = Caulerpa sertularioides. C = Caulerpa racemosa. D = Caulerpa floridana.
2. Ulva mempunyai talus berbentuk lembaran tipis seperti sla; karena itu dinamakan sla laut. Ada tiga spesies yang tercatat, satu di antaranya adalah Ulva reticulata. Ganggang ini biasanya melekat dengan menggunakan alat pelekat berbentuk cakram pada batu atau substrat lain atau pada tabung dari cacing beruas. Tangkainya pendek yang menghubungkan dengan daunnya yang tipis dan lebar. Bentuk dan ukuran daunnya tidak teratur. Daun yang lebar dapat mencapai 400 cm2. Daunnya mempunyai sejumlah perforasi tak teratur dan tebalnya hanya dua sel. Ganggang ini dapat tumbuh mulai dari daerah air pasang rata-rata pada pasut bulan-setengah sampai air surut terendah pada bulan purnama. Tumbuhan ini dapat terlepas dari pegangannya dan tersebar di sekitar mintakat pasut. Ganggang ini tumbuh bagus di selat dan perairan teluk yang tenang.
3. Valonia (V. ventricosa) mempunyai talus yang membentuk gelembung berisi cairan
berwarna ungu atau hijau mengkilat, menempel pada karang mati atau batu karang. Di perairan pantai Malaysia, terdapat spesies Valonia fastigiata. Ganggang ini
37
berbenang hijau bercabang dan beruas, garis tengahnya kira-kira 1 mm, tumbuh ke atas membentuk sebuah talus yang permukaan atasnya berbentuk kubah. Ujung bawah dari benang melekat kuat pada permukaan batu. Talus yang lebar dapat berukuran 4 cm garis tengahnya dan 1,5 cm tingginya.
4. Dictyospaera (D. cavernosa) dan spesies-spesies lain dari genus ini di Nusa Tenggara
Barat dinamai bulung dan dimanfaatkan untuk sayuran.
5. Halimedia terdiri atas 18 jenis. Seperti telah dikemukakan di atas genus ini berkapur
dan menjadi salah satu penyumbang endapan kapur di laut. Halimedia tuna terdiri atas rantai bercabang dari potongan tipis berbentuk kipas. Potongan-potongan ini berkapur, masing-masing bergaris tengah 2 cm. Yang terbesar dihubungkan satu dengan lainnya oleh sendi-sendi tak berkapur. Mereka terdapat di bawah air surut rata-rata pada pasut bulan-setengah, pada pantai berbatu dan pada paparan terumbu. Akan tetapi, potongan-potongannya dapat tersapu ke bagian atas pantai setelah terjadi badai. Halimedia Opintia berbeda dengan H. tuna karena spesies ini mempunyai potongan bentuk kipas lebih kecil, berwarna hijau muda, mempunyai panjang 1 cm, dan mempunyai bentuk pinggiran yang kurang teratur. Spesies ini terdapat di bawah air surut rata-rata pada pasut bulan-setengah pada pantai berbatu dan paparan terumbu.
6. Chaetomorpha mempunyai talus atau daunnya berbentuk benang yang menggumpal.
Spesies yang diketahui adalah C. crassa yang sering menjadi gulma bagi budidaya rumput laut.
7. Codium hidup menempel pada batu atau batu karang, tercatat ada enam spesies
38
Gambar 2.4 Codium tomentosum (Romimohtarto dan Juwana, 1999). A = Tumbuh-tumbuhan Codium.
B = Irisan membujur talus. C = Irisan melintag talus.
Gambar 2.5 Codium decordicatum (Romimohtarto dan Juwana, 1999).
8. Dari genus Udotea tercatat dua spesies dan banyak terdapat di perairan Sulawesi seperti di Kepulauan Spermonde dan Selat Makassar. Ganggang ini tumbuh di dasar pasir dan terumbu karang.
9. Tydemania (T. expeditionis) tumbuh di paparan terumbu karang yang dangkal dan di
39
10. Bernetella (B. nitida) menempel pada karang mati dan pecahan karang di paparan
terumbu.
11. Burgesenia (B. forbesii) mempunyai talus berbentuk kantung silendris berisi cairan
berwarna hijau tua atau hijau kekuning-kuningan, menempel di batu karang atau tumbuh-tumbuhan lain.
12. Neomeris (N. annulata) tumbuh menempel pada substrat dari karang mati di dasar
laut. N. annulata hidup di daerah pasut di seluruh perairan Indonesia.
2. 3. 3 Phaeophyceae (ganggang coklat)
Ganggang coklat hampir semuanya merupakan tumbuh-tumbuhan laut; hanya sedikit yang hidup di air tawar. Ganggang yang termasuk kelas ini adalah ganggang coklat yang di antaranya berukuran sangat besar. Pigmen ganggang dari kelas ini terdiri atas klorofil yang ditutupi oleh pigmen-pigmen kuning dan coklat, santofil, karotin, dan fukosantin (Smith, 1955; Kadi, 1996).
Ganggang coklat merupakan kelas ganggang yang terbesar ukurannya di antara kelas-kelas ganggang laut. Kelas ganggang ini mempunyai ukuran dan bentuk yang sangat beraneka ragam (Gambar 2.6). Ada yang berupa tumbuh-tumbuhan bercabang berbentuk benang kecil dan halus (Ectocarpus), ada yang berbentuk rantai seperti sosis yang kopong dan kasar yang panjangnya 30 cm atau lebih (Scytosiphon), ada yang bertangkai pendek dan bertalus lebar (Laminaria, Costaria, dan Alaria beberapa di antaranya mempunyai lebar 2 m), dan ada yang bentuknya bercabang banyak (Fucus,
Agregia). Di Samudra Pasifik, terdapat ganggang berukuran raksasa dengan tangkai yang
40
Gambar 2.6. Tipe-tipe khas dari ganggang laut yang multisel. A = Trichodesmium. B = Fucus. C = Alaria D = Ulva E = Ectocarpus F = Sargassum G = Rhodymenia H = Polysiphonia I = Scytosiphon J = Lithothamnion
Ganggang coklat ada yang membentuk padang panggang (kelp bed) di laut lepas. Mereka membentuk hutan lebat dengan daun-daun dan tangkai-tangkainya yang melambai-lambai di dalam dan di permukaan laut. Di tempat ini hidup beribu-ribu ikan yang mendapatkan makanan dan berlindung di hutan ganggang ini. Ganggang ini biasa dipanen di banyak tempat untuk produk komersial yang dihasilkan. Dilihat dari bentuknya, ganggang coklat adalah yang termaju di antara semua tumbuhan-tumbuhan
41
bertalus. Untuk mengetahui secara garis besar struktur dan perkembangbiakan ganggang coklat secara umum, dapat diambil contoh ganggang spesies Nereocystis (Gambar 2.7).
Nereocystis panjangnya dapat mencapai 35 m atau lebih. Tumbuhan ini melekat
pada substrat dengan suatu alat pelekat yang bercabang banyak, tetapi tidak mempunyai akar yang sebenarnya. Dari alat pelekat ini, tumbuh tangkai (stipe) yang panjang dan berbentuk silendris. Di dalam tangkai ini terdapat rongga yang berakhir pada ujung tangkai yang berbentuk bola. Bola ini berisi gas seperti halnya tangkai, sehingga tumbuh-tumbuhan ini dapat mengapung. Pada ujung bola, terdapat daun seperti pita atau lamina.
Gambar 2.7 Nereocystis luetkeana
Bola dan tangkai berongga ini membuat bagian atas tumbuh-tumbuhan berada di dekat permukaan, menyebabkan daun-daun mendapat sinar matahari yang cukup. Seperti halnya ganggang besar lainnya, bagian ini ulet, lentur, dan licin agar mampu menghadapi pengaruh gelombang badai arus keras yang sering terjadi dengan mengeluarkan daya tahan sekecil-kecilnya.
Daur hidup ganggang coklat ini mencakup berbagai tipe pergantian generasi. Umumnya pada Laminariles, yang mencakup ganggang besar, ada pergantian generasi yang dapat digambarkan oleh daur hidup dari Nereocystis. Di sini tumbuh-tumbuhan
42
sporofit yang besar menghasilkan satu seri sori atau “fruiting areas” yang nampak sebagai bercak coklat kehitam-hitamanan, memanjang sepanjang seluruh daun. Mulai dari ujung daun, bercak ini terlepas pada saat tumbuh-tumbuhan matang, meninggalkan celah besar pada daun. Dari sori yang matang keluarlah zoospora berbulu getar yang tak terbilang jumlahnya dan jika mencapai substrat yang cocok tumbuhlah mereka menjadi tumbuh-tumbuhan berbentuk benang yang kecil, yang merupakan fase gametofit yang tak kelihatan nyata. Jadi, alih generasi seperti ini adalah heteromorfit, Beberapa ganggang coklat, seperti Dictyotales (termasuk di dalamnya Padina), menunjukkan alih generasi yang isomorfik, seperti halnya pada Ulva. Penting untuk dicatat bahwa konservasi zoospora mungkin dapat dilakukan oleh Nereocystis, yakni dengan kebiasaannya menyebarkan sorus matang yang lengkap, yang ketika tenggelam di dasar padang ganggang memungkinkan mereka menemukan substratnya yang cocok. Jadi, zoospora jika dilepaskan akan lebih banyk berkumpul di dasar atau di dekat dasar perairan ketimbang tersebar luas oleh arus seperti halnya jika mereka disebarkan melalui sporangia dari sorus di permukaan laut. Zoospora ini bersemi dalam waktu 24 jam. Tumbuh-tumbuhan gametofit yang dihasilkan berupa jantan atau betina dan setelah besar dibuahi, pertumbuhan sporofit dimulai.
Dalam kelas ganggang coklat, Fucales (termasuk di dalamnya Fucus dan
Sargassum), tumbuh-tumbuhan utamanya adalah sporofit yang berada di dalam ribuan
konseptakel (conceptacle) berbentuk cawan yang sangat kecil. Mereka bersatu setelah disebarkan bebas di air. Jadi pergantian generasi hanya nyata secara sitologik (Gambar 2.8).
43
Gambar 2.8 Fucus (Romimohtarto dan Juwana, 1999).
Ganggang coklat berkembang sangat pesat di perairan dingin. Karena itu ganggang ini khas tumbuh-tumbuhan pantai berbatu di daerah lintang tinggi. Di lain pihak, Sargassum dan ganggang lain dari ordo Fucales merupakan ganggang dari perairan troipis dan subtropis. Ganggang di laut ini berasal dari pantai. Saat mereka terpatah dari induknya, mereka hanyut ke laut lepas dan berkembang biak di sana. Mereka terus mengapung dengan bantuan kantung udara dan tumbuh secara vegetatif.
Di Indonesia, terdapat delapan genus ganggang coklat yang sering ditemukan.
1. Cystoseira sp., yang hidup menempel pada batu di daerah rataan terumbu dengan
alat pelekatnya yang berbentuk cakram kecil. Ganggang ini mengelompok bersana-sama denga komunitas Sargassum dan Turbinaria. Di perairan pantai Malaysia terdapat spesies Cystoceira prolifera yang dapat berukuran besar dan terdapat di paparan terumbu dan pantai berbatu. Ganggang ini mempunyai dua atau tiga sayap longitudinal dengan pinggiran bergigi. Lebar sayap ini dapat
44
mencapai lebih dari 0,5 cm. kantung udara kecil terdapat di sepanjang talus (Gambar 2.9).
Gambar 2.9 Cystoseiraericoides (Romimohtarto dan Juwana, 1999). A = Tumbuh-tumbuhan Cystoseira.
B = Kantung udara yang diperbesar. C = Irisan melalui reseptakel. D = Oogonium.
E =Anteridium.
2. Dicyopteris sp., hidup melekat pada batu di pinggiran luar rataan terumbu, jarang
dijumpai. Spesies ganggang ini dapat ditemukan di laut selatan Jawa, Selat Sunda, dan Bali.
3. Dictyota (D. bartayresiana) mempunyai banyak sinonim yaitu: D. crenulata, D.
cuspidana, D. dichotoma, D. patens, dan D. ciliata. Ganggang ini tumbuh menempel
pada batu karang mati di daerah rataan terumbu. Di perairan pantai Malaysia, terdapat
Dictyota beccoriana yang tumbuh di bawah paras pasut rata-rata. Warnanya coklat
tua dan mempunyai talus bercabang yang terbagi dua. Talus yang pipih, lebarnya 2 mm, tersusun oleh tiga lapis sel. Lapisan tengah yang terdiri atas sel yang besar diapit oleh dua lapisan atas dan bawah yang terdiri atas sel yang sangat kecil. Ganggang ini
45
mempunyai bagian berbentuk silendris yang merayap dan mempunyai alat pelekat dalam bentuk benang-benang seperti rambut. Talusnya menghasilkan cabang lateral yang dapat terlepas untuk membentuk ganggang baru yang bebas dalam dalam perkembangbiakan vegetatif.
4. Hormophysa (H. triquesa), hidup menempel pada batu dengan alat pelekatnya
berbentuk cakram kecil. Ganggang ini, seperti Cystoceira, hidup bercampur dengan
Sargassum dan Turbinaria hidup di rataan terumbu. Ganggang ini tersebar luas di
perairan Indonesia.
5. Hydroclathrus ( H. clatratus) sinonimnya H. cantellatus, tumbuh melekat pada batu
atau pasir di daerah rataan terumbu dan tersebar agak luas di perairan Indonesia.
6. Padina (P. australis), sinonimnya P. gymnospora, tumbuh menempel di batu pada
daerah rataan terumbu, baik di tempat terbuka di laut maupun di tempat terlindung.
Padina commersonii adalah ganggang coklat yang banyak dijumpai di bawah paras
pasut rata-rata dari perairan Singapura dan Malaysia. Alat pelekatnya yang melekat pada batu atau pasir, terdiri atas cakram pipih, biasanya terbagi menjadi beberapa cuping pipih yang lebarnya 5-8 cm. Tangkai yang pipih dan pendek menghubungkan alat pelekat ini dengan ujung meruncing dari selusin daun atau lebih membentuk kipas. Setiap daun mempunyai jari-jari 5 cm atau lebih. Pinggirannya berakhir dengan suatu meristem; di tempat itu, pertumbuhan terjadi dan khas menggulung ke dalam untuk perlindungan lebih baik. Setiap daun ditandai oleh satu seri sabuk sepusat
(concentric), yang merupakan deretan sel. Daun yang lebih lebar biasanya membelah
menjadi bagian-bagian yang lebih kecil dan pinggiran luarnya cenderung membelah ke dalam sepanjang jari-jari. Daunnya berwarna coklat kekuning-kuningan, tetapi
46
dapat kelihatan keabu-abuan yang disebabkan karena adanya kerak yang terdiri atas lapisan tipis kapur pada permukaan atasnya.
7. Sargassum terdapat amat melimpah mulai dari air surut pada pasut bulan-setengah ke
bawah. Ganggang ini hidup melekat pada batu atau bongkahan karang dan dapat terbedol dari substratnya selama ombak besar dan hanyut ke permukaan laut atau terdampar di bagian atas pantai. Warnanya bermacam-macam dari coklat muda sampai coklat tua. Alat pelekatnya terdiri atas cakram pipih. Dari cakram ini, muncul tangkai yang pendek silendris yang tegak. Dari tangkai yang pendek ini muncul beberapa poros silendris panjang. Masing-masing poros ini panjangnya dapat mencapai 1 meter di mintakat bawah-litoral tempat Sargassum hidup. Pada poros yang silendris yang berdiameter 3 mm, terdapat bentuk-bentuk seperti daun, kantung udara, dan cabang-cabang perkembangbiakan. Di perairan Indonesia tercatat tujuh spesies, yaitu S. polycystum, S. plagiophyllum, S. duplicatum, S. ceassifolium, S.
binderi, S. echinocarpum, dan S. cinereum.
8. Turbinaria, terdiri atas tiga spesies yang tercatat, yaitu T. conoides, T. decurrens, dan
T. ornata. Mereka mempunyai cabng-cabang silendris dengan diameter 2-3 mm dan
mempunyai cabang lateral pendek yang panjangnya dari 1-1,5 cm. Ini berakhir pada sebuah reseptakel dengan pinggiran bergerigi dan garis tengahnya kira-kira 1 cm. Ganggang ini terdapat di pantai berbatu dan paparan terumbu.
2. 3. 4 Rhodophyceae (Ganggang Merah)
Hampir semua ganggang merah adalah tumbuh-tumbuhan laut. Di antara kelas ganggang laut, ganggang merah yang teramat mencolok dalam hal warna. Beberapa di
47
antaranya bercahaya. Banyak dari spesies yang kecil sekali ukurannya merupakan benda makroskopik yang indah.
Pigmen dari kromatofora terdiri atas klorofil biasa bersama-sama dengan santofil, karotin, dan sebagai tambahan fikoeritrin yang merah dan kadang-kadang fikosianin (Smith, 1955; Atmadja, 1996a). Berbagai warna tumbuh-tumbuhan terdapat dalam kelompok ganggang ini. Ada yang merah ungu, violet, coklat, atau hijau. Spesies yang tumbuh di tempat yang jeluk berwarna coklat murni. Ini mungkin berkaitan dengan kemampuannya mensintesis secara efisien pada cahaya yang redup pada perairan yang jeluk dibandingkan dengan spesies yang hidup di perairan dangkal. Meskipun biasanya berukuran kecil, bentuknya lebih beraneka ragam daripada ganggang coklat, dan jumlahnya juga lebih banyak. Semuanya bersel ganda. Yang sederhana adalah bentuk benang bercabang seperti Polysiphonia, yang bersama-sama dengan spesies ganggang benang lainnya umumnya dinamakan lumut laut. Ada bentuk berdaun lebar, seperti
Rhodymenia yang dapat berukuran panjang sekali. Namun ganggang merah yang
terpanjang kira-kira 1 sampai 2 meter.
Daur hidup beberapa spesies ganggang merah sangat majemuk. Pada bentuk yang lebih tinggi tingkatnya, terjadi pergantian generasi secara morfologis dan teratur. Dalam hal ini, dapat saja sporofit dan gametofit kelihatan sama dari luar. Untuk menggambarkan daur hidup ganggang merah, diambil contoh Polysiphonia (Gambar 2.10). Ada tiga tipe tumbuh-tumbuhan, yakni gametofit jantan dan betina dan tumbuh-tumbuhan tetrasporik yang aseksual. Yang terakhir ini timbul dari karpospora yang terjadi pada tumbuh-tumbuhan betina. Karpospora ini sebagai hasil bergabungnya gamet jantan dan betina.
48
Pada saat persemaian, tetraspora dari tumbuh-tumbuhan aseksual pada gilirannya menghasilkan tumbuh-tumbuhan seksual.
Salah satu sifat yang sangat menarik dari perkembangbiakan ganggang merah ini adalah tidak adanya samasekali spora atau gamet berenang yang berbulu getar atau bercambuk. Hal ini menyimpang dari kebiasaan yang diikuti oleh perkembangbiakan jasad hidup yang terjadi dalam media air. Hal ini membuat penyebaran dan pertemuan intim antara sel-sel perkembangbiakan tergantung pada arus. Karena itu, semuanya tergantung pada faktor kesempatan atau keberuntungan.
Gambar 2.10 Polysiphonia nigrescens (Romimohtarto dan Juwana, 1999). A = Tumbuh-tumbuhan Polysiphonia.
B = Tempat melekat dari bawah.
C = Irisan melintang batang pokok dekat pangkal. D = Ujung percabangan dengan tetraspora.
E = Sistokarpus.
49
Ganggang merah luas sebarannya, tetapi terbanyak terdapat di perairan beriklim sedang. Sebaran penegaknya menunjukkan bahwa ganggang ini menginginkan cahaya yang redup. Beberapa spesies ganggang merah hidup di mintakat pasut, tetapi pertumbuhan yang subur terdapat di mintakat bawah-pasut. Mereka dapat dijumpai dalam jumlah besar di jelukan yang kurang cocok bagi ganggang hijau dan coklat, dan di laut Mediteranean dapat dijumpai pada kejelukan 139 m. Jadi, mulai dari tempat dangkal sampai ke tempat yang jeluk, berturut-turut sebaran kelompok ganggang yang dipaparkan di sini adalah ganggang hijau, ganggang coklat, dan ganggang merah dengan areal tumpang tindih yang berbeda-beda. Di perairan tropis, ganggang merah umumnya terdapat di daerah bawah-litoral, pada daerah ini cahaya sangat kurang. Mereka umumnya berukuran kecil. Sekelompok ganggang ini ada yang disebut koralin, yang menyadap kapur dari air laut dan menjadi sangat keras seperti batu. Mereka terdapat di terumbu karang dan membentuk kerak merah muda pada batu karang dan batu cadas.
Banyak spesies ganggang merah yang mempunyai nilai ekonomis dan diperdagangkan sehingga dikelompokkan sebagai komoditi rumput laut. Di Indonesia tercatat 18 genus yang terdiri atas 42 spesies. Berikut catatan singkat dari genus ganggang merah tersebut (Romimohtarto dan Juwana, 1999).
1. Acanthophora, terdiri atas dua spesies yang tercatat yaitu: A. spicifera dan A.
muscoides. Mereka hidup menempel pada batu atau benda keras lainnya. Yang
pertama luas sebarannya di Indonesia dan yang kedua kurang meluas dan terdapat di tempat tertentu saja seperti di Kepulauan Seribu, sebelah utara Teluk Jakarta (Gambar 2.11).
50
Gambar 2.11 Acanthophora spicifera (Romimohtarto dan Juwana, 1999).
2. Actinotrichia (A. fragilis), terdapat di bawah pasut dan menempel pada karang mati.
Sebarannya luas sehingga terdapat pula di padang lamun.
3. Amansia (A. glomerata) hidup melekat pada batu di daerah terumbu karang dan dapat
hidup melimpah di padang lamun.
4. Amphiroa (A. fragilissima) tumbuh menempel pada dasar pasir di rataan pasir atau
menempel pada substrat dasar lainnya di padang lamun. Sebarannya luas.
5. Chondrococcus (C. hornemannii) tumbuh melekat pada substrat batu di ujung luar
rataan terumbu yang senantiasa terendam air.
6. Corallina belum diketahui spesiesnya. Ganggang ini tumbuh di bagian luar terumbu
yang biasa terkena ombak langsung. Sebarannya tidak begitu luas, terdapat diantaranya di pantai selatan Jawa. C. rubens, contoh spesies dari luar Indonesia.
7. Eucheuma adalah ganggang merah yang biasa ditemukan di bawah air surut rata-rata
pada pasut bulan-setengah. Ganggang ini mempunyai talus yang silendris berdaging kuat dengan bintil-bintil atau duri-duri yang mencuat ke samping pada beberapa spesies. Talusnya licin. Warna ganggangnya ada yang tidak merah, tetapi coklat kehijau-hijauan kotor atau abu-abu dengan bercak merah. Di Indonesia, tercatat
51
empat spesies yaitu E. denticulatum ( = E. spinosum), E. Edule, E. alvarezii ( =
Kapaphycus alvarezii), dan E. sera (Romimohtarto dan Juwana, 1999).
8. Galaxaura terdiri atas empat spesies, yaitu G. kjelmanii, G. subfruticulosa, G.
subverticillata, dan G. rugosa. Mereka tumbuh melakat pada substrat batu di rataan
terumbu.
9. Gelidiella (G. ecerosa) tumbuh menempel pada batu di mintakat pasut atau bawah
pasut. Ganggang ini muncul di permukaan air pada saat air surut dan mengalami kekeringan. Ganggang ini digunakan sebagai sumber agar yang diperdagangkan.
10. Gigartina (G. affinis = Corpopeltis affinis) tumbuh menempel pada batu di rataan
terumbu, terutama di tempat-tempat yang masih tergenang air pada saat air surut terendah.
11 Gelidium dikenal sebanyak 40 spesies dari berbagai negara, tetapi hanya 8 spesies
diantaranya tersebar di Indonesia, yaitu G. latifolium, G. cartilagineum, G. rigidum,
G. corneum, G. crinale, G. cologlossum, G. pusillum, dan G. pannosum. Habitat dan
sebaran Gelidium di Indonesia pada umumnya di perairan pantai berbatu dan terbuka yang kebanyakan di daerah pantai Samudera Hindia. Substrat dasar tempat melekatnya berupa batu karang mati, gamping, dan batu vulkanik. Gelidium yang tumbuh di perairann Indonesia adalah jenis-jenis yang cenderung menyukai kadar garam tinggi (sekitar 33%).
12. Gracilaria terdiri atas tujuh spesies, yaitu G. arcuata, G. coronopifolia, G. foliifera,
52
Gambar 2.12 Beberapa spesies Gracillaria (Romimohtarto dan Juwana, 1999). A = Gracillaria verrocosa
B = Tumbuh-tumbuhan dari Glacillaria foliifera.
C = Tumbuh-tumbuhan G. foliifera dengan percabangan menyirip yang sempit.
13. Halymenia terdiri atas dua spesies, yaitu H. durvillaei dan H. harveyana. Mereka
hidup melekat pada batu karang di luar rataan terumbu yang selalu tergenang.
14. Hypnea terdiri atas dua spesies, yaitu H. asperi dan H. servicornis. Mereka hidup di
habitat berpasir atau berbatu, ada pula yang bersifat epifit. Sebarannya luas.
15. Laurencia terdiri atas tiga spesies, yaitu L. intricata, L. nidifica, dan L. obtusa.
Mereka hidup melekat pada batu di daerah terumbu karang.
16. Rhodymenia (R. palmata) hidup melekat pada substrat batu atau rataan terumbu.
17. Titanophora (T. pulchra) jarang dijumpai. Spesies ini terdapat di perairan Sulawesi.
18. Porphyra terdapat mulai dari perairan subtropis sampai daerah tropis, tetapi sebaran
menegaknya sangat terbatas. Pada umumnya di alam, ganggang ini dijumpai di daerah pasut (litoral), tepatnya di atas daerah litoral. Ganggang ini hidup di atas batuan karang pada pantai yang terbuka serta bersalinitas tinggi. Meskipun demikian,