Phytochemical screening and in silico studies of
fl
avonoids from
Chlorella pyrenoidosa
Rajasri Yadavalli
a,*, John Reddy Peasari
a, Priyadarshini Mamindla
b, Praveenkumar
c,
Sri Mounika
a, Jayasree Ganugapati
aaDepartment of Biotechnology, Sreenidhi Institute of Science and Technology (Autonomous), Yamnampet, Ghatkesar, Hyderabad, Telangana state, India bAthon LLC, Houston, TX, USA
cAgasthya International Foundation, Bengaluru, India
A R T I C L E I N F O
Keywords: Chlorella Phototrophic Heterotrophic Flavonoids Auto dock vina Schrodinger
A B S T R A C T
The recent explosion of interest in the bioactivity of theflavonoids of microalgae is due to the potential health benefits of the polyphenolic components that are major dietary constituents. The present study focuses on the phytochemical screening and in silico studies offlavonoids. Totalflavonoids content inChlorella pyrenoidosawas estimated in two modes of cultivation (Autotrophic and Heterotrophic) and its implication in anti-proliferation and anti-inflammatory activity was assessed through in silico approach. H-Ras p21(PDB-4L9S) and Lip-oxygenase (PDB-3V99) involved in proliferation pathway and inflammatory pathway were selected as the target proteins for in silico studies. Seven compounds were selected for molecular docking. Pharmacokinetic properties of these compounds were calculated using online tools and docking was performed using Auto Dock Vina. By comparing and analyzing their binding energies in Maestro Schrodinger, suite, it was observed that Epi-gallocatechin gallate exhibited least binding energy of 9.1 kcal/mol and hence has anti-inflammatory activity. Catechin has best binding affinity with H-Ras p21 and hence has anti proliferative activity.
1. Introduction
Microalgae are a key natural resource for a vast array of compounds viz. biodiesel, nutraceuticals, including proteins, vitamins, minerals, carotenoid pigments, such as xanthophylls and carotenes, Flavonoids etc.
Subspecies ofChlorellaare known to have several bioactive secondary
metabolites which can have bacteriostatic, bactericidal, antioxidant
anti-proliferative, antifungal, antiviral and antitumor activity [1,2]. Recent
exploration of anti-proliferative and anti-angiogenic properties of Chlorella pyrenoidosa,a unicellular fresh water green alga,paves way for exploring its use in treating inflammation and proliferation associated with various diseases.
Flavonoids are the largest groups of phenolic compounds are known to contain a broad spectrum of chemical and biological activities
including antioxidant and free radical scavenging properties [3].
Flavo-noids includeflavonols,flavones, catechins, proanthocyanidins,
antho-cyanidins and isoflavonoids. In the recent times,flavonoids have gained
increasing interest as they exhibit beneficial health effects due to their
potential antioxidant [4], anti-inflammatory and anti-cancer activities.
Information is scarce on the presence of secondary metabolites,
respon-sible for anti-proliferative and anti-inflammatory properties ofC.
pyr-enoidosa. Hence in this study, autotrophic and heterotrophic cultures of C.pyrenoidosawere grown and HPLC analysis was carried out for the
algal biomass for analyzing variousflavonoids. The aim of the present
study is to assess totalflavonoids content and to study the role of
flavo-noids as anti-proliferative and anti-inflammatory agents using in silico analysis.
In this study above mentioned properties offlavonoids were
evalu-ated by in silico methods with lipoxygenases-an enzyme relevalu-ated to oxidation of various fatty acids and Ras proteins which is a member of a super family of small GTPase involved in cell growth. The 5-lipoxygenase protein enzyme (5LO) and its leukotriene metabolites have long been known to be important modulators of inflammation in other disease
states [5]. The ras oncogene p21 antigen (p21) has been identified in
several epithelial malignancies, including breast, colon, bladder, and
prostate [6].
* Corresponding author. Department of Biotechnology, Sreenidhi Institute of Science and Technology (Autonomous), Yamnampet, Ghatkesar, 501301, Hyderabad, Telangana State, India.
E-mail address:[email protected](R. Yadavalli).
Contents lists available atScienceDirect
Informatics in Medicine Unlocked
journal homepage:www.elsevier.com/locate/imu
https://doi.org/10.1016/j.imu.2017.12.009
Received 11 November 2017; Received in revised form 21 December 2017; Accepted 21 December 2017 Available online 26 December 2017
2. Materials and methods
2.1. Inoculum preparation
2.1.1. Phototrophic culture
Chlorella pyrenoidosa(NCIM NO: 2738) was obtained from National
Centre for Industrial Microorganisms (NCIM), Pune, India.C. pyrenoidosa
is used for culturing in phototrophic mode by using BG11 as medium [7].
800 ml of media was taken in four different conicalflasks and 10% of
inoculum was added to eachflask. The cultures were grown for one week
at room temperature under continuous light illumination of
55μmol m 2s 1. The prevailing conditions like oxygen supply and
car-bon dioxide supply were monitored by providing air continuously at
2 l min 1and CO2for 10 min daily.
2.1.2. Heterotrophic culture
100 ml of the phototropic culture is taken as the inoculum for growing
the heterotrophic culture ofChlorella pyrenoidosa.It is added to one liter
of modified BG11 medium with glucose as carbon source in absence of light. It was grown in dark for one week by providing aeration in a
controlled manner 2 l min 1. Sub culturing of the same was done again to
obtain pure heterotrophic culture ofC. pyrenoidosa.
2.2. Sample preparation for HPLC analysis from phototrophic and heterotrophic cultures
An accurately weighed 2 g of dried biomass obtained from
photo-trophic and heterophoto-trophic cultures ofC.pyrenoidosawere taken each and
the samples were extracted with 2 ml of hexane for 30 min at 20
C temperature. The tubes were centrifuged at 4500 g for 10 min and the supernatant was recovered. The extraction was repeated with 2 ml of
hexane and the supernatants were collected [8]. The remaining residue
was subsequently extracted twice with ethyl acetate of 2 ml for 30 min at
20
C temperature and the supernatants were again collected. Subse-quently, the residues were further extracted twice with water 2 ml each
time for 30 min at 80
C and the supernatants were combined. The hexane, ethyl acetate and together with aqueous extracts were all stored
at 10
C before using them for biochemical analysis and HPLC analysis.
2.3. Biochemical analysis
Both phototrophic and heterotrophic biomass of C.pyrenoidosawas
used individually for performing biochemical analysis to test for the
presence offlavonoids.
2.3.1. Test forflavonoids
5 ml of dilute ammonia solution was added to a portion of the hexane, ethyl acetate and aqueous extracts of both photo and heterotrophic
samples followed by addition of concentrated H2SO4. A yellow color in
each extract indicated the presence offlavonoids. The yellow color
dis-appears on standing. Few drops of 1% aluminum solution were added to
portion of each extractfiltrate.
2.3.2. Determination of total phenolic compounds
According to Slinkard and Singleton [9], total soluble phenolic
compounds were determined with Folin-Ciocalteu reagent using pyro-catechol as a standard phenolic compound. Briefly, 1 ml of the extract
(1 mg/ml) in a volumetricflask diluted with 46 ml distilled water. One
milliliter of Folin-Ciocalteu reagent was added and the content was thoroughly mixed. After 3 min, 3 ml of sodium carbonate (2%) was added and then was allowed to stand for 2 h with intermittent shaking. The absorbance was measured at 760 nm in UV–vis spectrophotometer [Elico SL-210]. The total concentration of phenolic compounds in the extract determined as microgram of pyrocatechol equivalent (PE) per milligram of dry extract.
2.3.3. Totalflavonoids content
Dowd method [10] was used to estimate the totalflavonoids content.
Briefly, two milliliters of 2% aluminum trichloride (AlCl3) in methanol
was mixed with the same volume of the extract solution (1 mg/ml). The mixture was incubated at room temperature for 10 min, and the
absor-bance was measured at 415 nm in spectrophotometer. The total
flavo-noids content determined as microgram of rutin equivalent (RE) per milligram of dry extract.
2.4. HPLC analysis
The hexane, ethyl acetate and aqueous extracts of both phototrophic
and heterotrophic C.pyrenoidosawere subjected to HPLC analysis
[Shi-madzu LC-10AT vp] using PDA as detector with wavelength 270 nm using RP C18 as column in an isocratic manner with HPLC grade
meth-anol: water in the ratio 90:10 as solvents at 30
C. Theflow rate was
adjusted to 1 ml/min with sample injection volume 10μl. The run time
was set for 30min. The obtained results were used to carry out in silico studies to assess the anti-oxidant and anti-proliferation capacity of C. pyrenoidosa.
2.5. In silico analysis
2.5.1. Retrieval of protein structure
The X ray crystal structure of proteins 3V99 (5-lipoxygenase) at 2.25
Aºresolution and 4L9S (Signaling protein) at 1.61 Aºresolution used in
this study were retrieved from RCSB Protein Data Bank [11]. They play a
major role in inflammation and proliferation pathways respectively. The proteins were prepared for molecular docking by removing the water molecules and other hetero molecules from the original crystal structure. Active site analysis was performed using Swiss Protein Viewer, SPDBV
[12].
2.5.2. Retrieval of ligands and ADME property prediction
3D structures offlavonoids that were identified from HPLC analysis of
Chlorella pyrenoidosawere retrieved from NCBI Pub Chem Compounds in
SDF format [13]. 2D structures were sketched using Chemspider and
Molinspiration. The names and CID numbers of the compounds are Caffeine (CID: 2519), Protocatechic acid (CID: 528594), Catechin (CID:
73160), Epicatechin (CID: 72276),Epigallocatechin –gallate (CID:
65064), Caffeoyl-D-glucose (CID: 129661118), Dihydroquercetin-7,
40
-dimethyl ether (3D structure was generated using molinspiration).
ADME [14] properties (i.e., absorption, distribution, metabolism and
excretion) of the selected compounds was predicted by Molinspiration.
2.5.3. Grid preparation and molecular docking
Molecular Docking was performed using Autodock Vina [15] and
MGL tools [16and17]. Docking Inputfiles were created using AutoDock
tools batchfile of MGL tools. Docking was performed between selected
macromolecules and ligands. Hydrogen atoms, Kollman charges were
added to protein molecules and prepared as PDBQTfiles. The ligand was
prepared in PDBQT by setting flexible torsion angles at all rotatable
bonds, while the protein was kept as a rigid structure. The Lamarckian Genetic Algorithm (LGA), a local search algorithm was utilized for li-gands conformations searching.
Configuration files were created for both the proteins by setting
suitable Cartesian coordinates to generate Grid box. For protein 4L9S
grid box parameters are X¼36.212, Y¼ 11.524,&Z¼5.085 and grid
box dimensions was set at 60*60*60 Aºwhich covers all the amino acids
in the active site. For protein 3V99 coordinates for X, Y, and Z axis were
11.446, 73.596, 24.378 and dimensions for grid box are 70*70*70 Aº.
The docked complex forming hydrogen bonds and other parameters
like intermolecular energy (Kcal/mol) and inhibition constant (μM) were
ligands were analyzed using Schrodinger Suite [18].
3. Results
3.1. Total phenolic andflavonoid contents and HPLC analysis
Flavonoids with their multiple activities viz. microbial, anti-cancer, and anti-diabetic can play a vital role in today's dietary
supple-ments. Qualitative analysis performed in the all three extracts ofChlorella
pyrenoidosain the present study, demonstrated the presence of
phyto-chemicals like phenolic compounds and flavonoids (Table 1). .When
compared to Phototrophic mode, heterotrophic mode yielded high
amount of total phenols and flavonoids content. In both the modes,
aqueous extract gave more phytochemical yield (1.21 mg PE/mg of total
phenols and 0.87 mg RE/mg dry cell weight of flavonoids) when
compared to hexane and ethyl acetate extracts. Our results are similar
with thefindings of Baviskar and Khandelwal [19] who also extracted
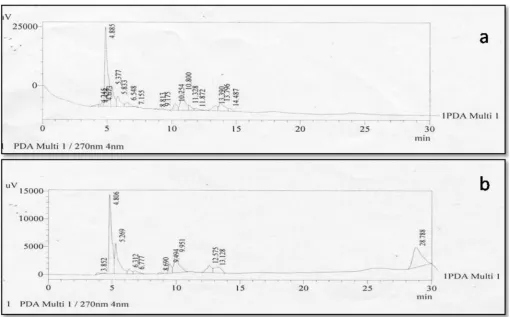
flavonoids from microalgae grown in pond water and ricefields. Fig. 1represents the HPLC of various phytochemical constituents of
aqueous extract ofChlorella pyrenoidosain both photo and heterotrophic
conditions. The number of peaks represents the different biologically active phytochemical constituents and the major peak area compounds
could belong to the polyphenols andflavonoids.
HPLC analysis carried out from biomass of both phototrophic and
heterotrophicChlorella pyrenoidosa,showed the presence offlavonoids
(Table 2) caffeine (Rt-4.320), catechin (Rt-5.269),epicatechin (Rt-6.548),
pigallocatechingallate (ECGC)(Rt-11.328),dihyroquerecetin-7,40
-dimethyl-ether (DQME)(Rt-10.800), caffeoyl-D-Glucose (Rt-12.575),
protocatechuic acid (Rt-7.155)(Table 2). Ludmila Machu et al. [20]
evaluated antioxidant capacity of Chlorella pyrenoidosa, and Spirulina
platensis. HPLC analysis of their study showed that the most abundant phenolic compound was epicatechin. In another similar study on
microalgal species, Jayshree et al. investigated phenolic andflavonoid
content inC. vulgaris and Chlamydomonas reinhardtii, followed by
anti-cancer, antioxidant and antimicrobial activities. They observed that both species exhibited free radical scavenging activity and high antioxidant
potency. Both species also revealed that flavonoids present in their
biomass exhibited potential anticancer activity with high correlation
coefficient values. They concluded thatflavonoids tend to be significant
Table 1
Phenolic and flavonoids content in various extracts of C.pyrenoidosa. Results are meanS.D. of three parallel measurements of different plant extracts.
Mode of cultivation
Phytochemical Content
Aqueous extract
Hexane extract
Ethyl acetate extract
Phototrophic Total Phenols (mg PE/mg)
0.920.08 0.730.09 0.690.07
Flavonoids (mg RE/mg)
0.60.07 0.520.09 0.500.05
Heterotrophic Total Phenols (mg PE/mg)
1.210.06 0.960.08 0.810.06
Flavonoids (mg RE/mg)
0.870.06 0.780.07 0.600.06
Fig. 1. HPLC separation of Aqueous extract ofC. pyrenoidosaa) in heterotrophic culture b) in phototrophic culture.
Table 2
HPLC chromatogram retention time of phytochemical constituents obtained in photo-trophic and heterophoto-trophic modes.
Phytochemical constituents Retention time (minutes)
Caffeine 4.245
Protocatechic acid 7.155
Catechin 5.269
Epicatechin 6.548
Epigallocatechin -gallate 11.328
Caffeoyl-D-glucose 12.575
cause for exhibiting anticancer activity than phenolic compounds in the
biomass [21]. Hence, theflavonoids identified from HPLC analysis in this
study are used as the inhibitors for carrying out the in-silico studies to assess for the anti-inflammatory and anti-proliferative activity of C. pyrenoidosa.
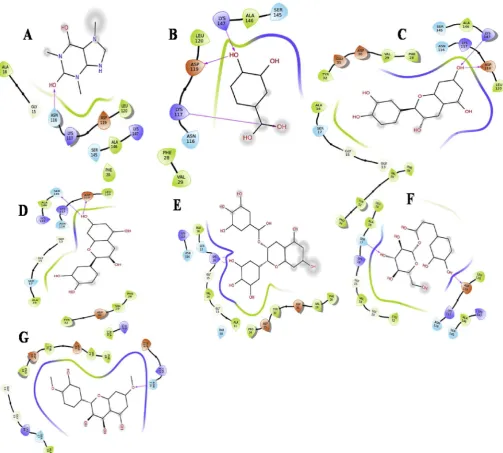
3.2. Retrieval of ligand structures
The structures of the compounds identified from the HPLC
chro-matogram (Table 2) are given in (Fig. 2). These compounds were used for
molecular docking studies to assess their proliferative and anti-inflammatory activity.
3.3. Active site analysis
Active site analysis of 3V99 and 4L9S were carried out using SPDBV.
Active site residues for protein 3V99 (Fig. 3a)were found to be SER171,
ILE406, HIS372, HIS550, ASN554, GLN557, LEU609, GLN528, PHE117
and residues for protein 4L9S (Fig. 3b) were PHE28, LYS147, VAL29,
ASP30, ALA146, SER145, ASP119, ALA18, GLU31, LEU120, TYR32, SER17, LYS117, ASN116, GLY15, LYS16, VAL14, GLY13, CYS12, ALA11.
3.4. Molinspiration
For the compound to be absorbed efficiently, logP value must be less than 5.0. Compund having logP value in reasonable range is said to possess good permeability character. Out of 7 compounds 7 have TPSA
value less than 140 Aºthat implies a good optimum drug absorption
capacity. Lipinski's 'Rule of 50
was applied to identify the best lead
compounds. ADME property values are shown in (Table 3).
All the canonical smiles of ligands were submitted online to
molins-piration to analyze ADME properties. Caffeine,
cat-echin,protocatechicacid,Caffeoyl-D
-glucose,epicatechin,dihy-droquercetin-7,40
-dimethyl ether showed 0 violations and obeyed‘Rule
of 5’, whereas compounds epigallocatechin gallate violated 2 properties. Epigallocatechin gallate violated from rule by having 11 nOH and 8 nOHNH.
3.5. Molecular docking
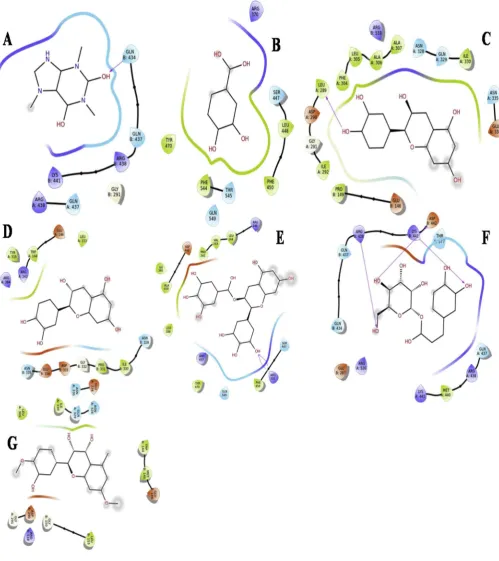
Molecular docking was performed using selected proteins and the ligands in Autodock vina. Binding energy values are indicated in (Table 4). All the docking poses showed negative binding energy indi-cating that they have good binding affinity with the target protein.
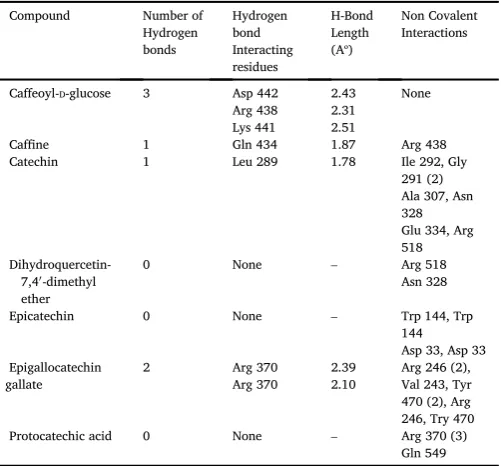
Caffeine binding energy was found to be 7.0 kcal/mol (Table 4)
with 3V99 and with 4L9S it was found to be 6.0 kcal/mol. It was found
that 3 H bonds were formed between 3V99 with 0 hydrophobic in-teractions and caffeine (Asp 442,Arg 438,Lys 441),2 H bonds and formed one non covalent interactions with Arg 438 residue against 4L9S (Ser
17,Asp 119). (Fig 4.1 (A)&4.2 (A)).
Protocatechic acid binding energy was found to be 6.4 kcal/mol
against 3V99,with 4L9S it was found to be 6.3 kcal/mol (Table 4)
which possessed almost equal energy with 3V99. It was found that no H bonds were formed between 3V99 and protocatechic acid but non co-valent interactions were formed (Arg 370 (3), Gln 549),3 H bonds against
4L9S (Lys 18,Asp 119,Lys 147) along with non-covalent interactions. (Fig
4.1 (B)&4.2 (B)).
Catechin binding energy was observed to be 8.3 kcal/mol (Table 4)
against 3V99,with 4L9S it was found to be 9.1 kcal/mol. It was found that only one H bond was formed between 3V99 and catechin (Leu 289),2 H bonds against 4L9S (Lys 147,Asp 119) with no non covalent
interactions. (Fig 4.1 (C)&4.2 (C)).
Epicatechin binding energy was found to be 8.0 kcal/mol (Table 4)
with 3V99 and with 4L9S it was found to be 8.8 kcal/mol. It was found
that no H bond was formed between 3V99 and epicatechin but non co-valent bonding was observed (Try 144 (2), Asp 33 (2)), 2 H bonds against
4L9S (Asp 119, Ser 145). (Fig 4.1 (D)&4.2 (D)).
Dihydroquercetin-7,40
-dimethyl ether binding energy was found to be
7.8 kcal/mol against 3V99, with 4L9S it was observed to be 8.6 kcal/
mol (Table 4). It was found that no H bond was formed between 3V99
and dihydroquercetin-7,40
-dimethyl ether but non covalent bonding was observed between residues (Arg 518, Asn 328),1 H bond found against
4L9S (Asn 116). (Fig 4.1 (E)&4.2 (E)).
Caffeoyl-D-glucose binding energy was found to be 8.0 kcal/mol
(Table 4) against 3V99, with 4L9S it was found to be 8.6 kcal/mol. It was observed that 3 H bonds were and formed between 3V99 and
Caf-feoyl-D-glucose (Asp 442, Arg 438, Lys 441),2 H bonds (Ser 17, 119) and
three hydrophobic interactions (Ala 18 (2), Gly 15)against 4L9S. (Fig 4.1
(F)&4.2 (F)).
Epigallocatechin gallate binding energy was found to be 9.1 kcal/
mol (Table 4) against 3V99, with 4L9S it was found to be 8.6 kcal/mol.
It was observed that 2 H bonds and 7 non covalent bonds (Arg 246 (2),Val243, Tyr 470 (2), Arg 246,Try470)were formed between 3v99 and epigallocatechin gallate with same active site residue (Arg 370, Arg 370),2 H bonds and 7 hydrophobic interactions (Gly 13, Ala 18, Ser 17 (2), Gly 15, Ser 17, Gly 15) against 4L9S (Lys 16, Lys 16) with same
amino acid in the active site region of the protein. (Fig 4.1 (G)&4.2 (G)).
It was found that most of the active site residues interacted with the
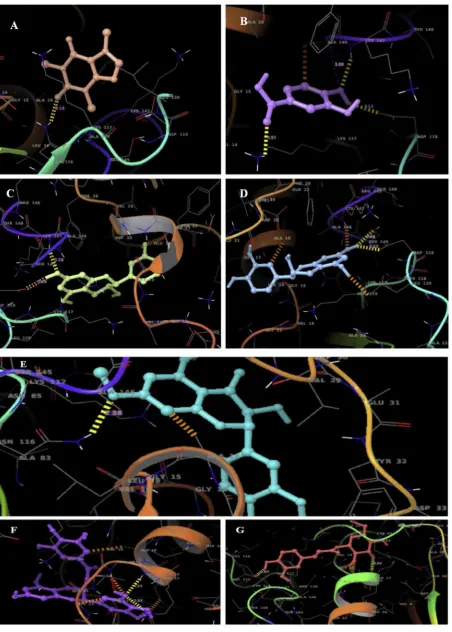
ligands through covalent and non-covalent bonding. 2D (Fig 4.1, 4.2) and
Table 3
Molinspiration property values of Compound.
Compound miLogP1 Natoms3 M.Wt4 nON5 nOHNH6 nVio7 Nrotb8 Volume6
Caffeine 0.60 14 194.19 6 0 0 0 167.63
Protocatechic acid 0.88 11 154.12 4 3 0 1 127.08
Catechin 1.37 21 290.27 6 5 0 1 244.14
Epicatechin 1.37 21 290.27 6 5 0 1 244.14
Epigallocatechin gallate
2.25 33 458.38 11 8 2 4 367.57
Caffeoyl-D-glucose 0.77 24 342.30 9 6 1 5 286.62
Dihydroquercetin-7,40-dimethyl ether 1.55 24 332.31 7 3 0 3 281.38
Where (1)Mol Log P(Partition coefficient) for octanol/water ( 2.0 to 6.5) (2)Number of atoms in a compound. (3)Molecular weight of the molecule should be in range between 160 and 500.(4)Estimated number of H-bond acceptors should not be more than 10. (5)Estimated number of H-bonds donors should not be more than 5. (6)Number of violations. (7)Molecular volume.
Table 4
Docking scores of 3V99, 4L9S with ligands.
Compound Binding energy (kcal/
mol) 3V99
Binding energy (kcal/ mol) 4L9S
Caffeine 7.0 6.0
Protocatechic acid 6.4 6.3
Catechin 8.3 9.1
Epicatechin 8.0 8.8
Epigallocatechin gallate 9.1 8.5
Dihydroquercetin-7,40 -dimethyl ether
7.8 8.6
Caffeoyl-D-glucose 8.0 8.6
3D overlays of the docked complexes were obtained in XP Visualizer of
Schrodinger Suite (Fig 5.1 and 5.2).
4. Discussion
4.1. Flavonoids from microalgae
Many studies have focused on the biological activities of plant derived
polyphenolicflavonoids but very few were about microalgal species [22].
Hamed Safafar et al. investigated the potential of microalgae species grown on industrial waste water as a source of antioxidants. They found
from the study thatDesmodesmussp. represented a potentially rich source
of antioxidants, containing Lutein, tocopherols, and phenolic compounds
[23]. Flavonoids are potent water soluble antioxidants which prevent
oxidative cell damage along with strong anticancer activity and
anti-inflammatory activity [3,24]. In general, total phenolic and
flavonoid contents of different extracts of microalgae are solvent
dependent. In our study, aqueous extracts of C. pyrenoidosa showed
higher amount of phenolics and flavonoids, while their counterparts
showed lower concentration.
Bioactive compounds formation follows a complex pathway in uni-cellular microorganisms like microalgae. Synthesis of secondary
metab-olites like fatty acids, carotenoids, phenolics and flavonoids, initiates
through the formation of acetyl CoA by the ACCase gene through acetyl CoA carboxylation. This forms the key step at which carbon is assigned
for the secondary metabolite. In the present study, flavonoids were
observed inChlorella pyrenoidosain both autotrophic and heterotrophic
mode of nutrition. However, the production offlavonoids was observed
more in heterotrophic mode than autotrophic mode. This might be accounted for external carbon source supplemented, which in turn form
more acetyl CoA and lead to the formation of moreflavonoids. Formation
of theseflavonoids may also be attributed to the dark condition, a stress
factor maintained in heterotrophic mode. In dark condition, pigments
formation is inhibited and leads to enhanced production offlavonoids by
bypassing the pathway.
4.2. Binding mode analysis of ligands with macromolecules
In the present study it was observed that Epigallocatechin gallate had the best binding energy (BE) indicating the best possible pose with a BE
of 9.1 kcal/mol against the target molecule 3V99 (Table 5). Catechin
had the BE of 9.1 kcal/mol with the protein 4L9 (Table 6). Interactions
of docked complex of each ligand with functional residues were analyzed
and inspected in Schrodinger Suite [18].
4.2.1. Anti-inflammatory activity offlavonoids
Polyphenols are a large class of compounds synthesized by fruits, vegetables, teas, cocoa and other plants that possess certain health ben-efits. Polyphenols are divided into several groups, one of which is
rep-resented byflavonoids. Flavonoids are a group of natural compounds that
are categorized into flavonols, flavones, catechins, flavanones, antho-cyanidins, and isoflavonoids.
They exhibit anti-inflammatory activity by inhibition of
phospholipase A2, COX, and LOX. Quercetin is reported to be a strong
inhibitor of both COX-2 and 5-LOX [25,26].
due to inhibition of cyclooxygenase (COX), lipoxygenase (LOX) and
tumor necrosis factor (TNF), and nuclear factor kappa B (NF-κB [27].
Similar docking studies were conducted by Rina Herowati and Gunawan
Pamudji Widodo [28] and they reported that some flavonoids and
phenolic compounds, i.e., amentoflavone, apigenin, bilobetin, diosmine, epicatechin gallate, ginkgetin, hesperidin, luteolin, morelloflavon, and quercetin, showed lower binding energy than that of tolfenamic acid, the
selective COX-2 inhibitor. Our findings are in correlation with these
studies. In the present study it was observed that Epigallo catechin gallate had the best binding energy (BE) indicating the best possible pose with a
BE of 9.1 kcal/mol against the target protein Lipoxygenase.
4.2.2. Anti proliferative activity offlavonoids
Compounds from plant sources have been reported to possess anti proliferative properties. In vitro study using cancer cell lines and the zebrafish embryo developmental assay evaluated the anti-proliferative
activity of bark extracts ofGeissospermum reticulatum. The effects of the
extracts were examined on proliferation of T-cells. The results showed that ethanolic extracts of barks effectively exhibit cytotoxicity upon
malignant cultured cells and inhibit proliferation of healthy CD3þcells.
The anti-proliferative activity was related to the total amount of
phe-nolics andflavonoids in the ethanolic extracts [29].
Polyphenols from industrial apple waste have shown efficacy against the proliferation of several human cancers cells, such as human cervical (HeLa), human hepatoma (HepG2), and human colon cancer cells
(HT-29) [30,31].
Several Molecular docking studies reported that Flavopiridol, a
syn-theticflavonoid, was best bound to DNA topoisomerase I, Green tea
catechin was best docked with topoisomerase II and VEGFR-2 and quercetin showed very good binding interaction with telomere:
G-quadruplex [32].
In our present study Catechin exhibited good interaction with the protein 4L9S indicating anti proliferative activity. A similar study by
Seeram et al. reported that [33] eleven catechins consisting of
(þ)-catechin, ( )-catechin, ()-catechin, (þ)-epicatechin, (
)-epi-catechin, ( )-epigallocatechin, ( )-gallocatechin, ( )-epicatechin
gallate, ( )-catechin gallate, ( )-epigallocatechin gallate, and ( )-gal-locatechin gallate were tested for their anti-proliferation activity. Their study reported that the galloyl derivatives of catechins inhibited the proliferation of the cancer cell lines.
5. Conclusion
The extraction offlavonoids fromChlorella pyrenoidosawere carried
out using HPLC analysis and the anti-oxidative and anti-proliferative
properties of C. pyrenoiodsa were assessed by performing
computa-tional studies. The extraction offlavonoids from Chlorella pyrenoidosa
were carried out using HPLC analysis and the oxidative and
anti-proliferative properties ofC. pyrenoiodsawere assessed by performing
in silico studies. Computational studies were carried out using the
selectedflavonoids with the proteins, 5-lipooxygenase (3V99) and H-ras
p21 (4L9S) using Autodock Vina. Molecular docking resulted in the best binding conformations of the ligands. From the docking it is evident that the best pose was obtained with least energy value. Epigallocatechin gallate had good binding affinity with 3V99 and Catechin exhibited good interaction with the protein 4L9S. The interaction of ligands with active site indicate that, these potential ligands play an important role as anti-inflammatory and anti-proliferative agents. Hence in silico analysis
gives scope for usingC. pyrenoidosaas a target for preparation of drugs for
the treatment of various diseases associated with inflammation and proliferation such as cancers. Further studies have to be carried out to prove their efficacy.
Acknowledgement
Authors would like to thank Technical Quality Improvement pro-gramme (TEQIP-II),World Bank funded project for procuring Schro-dinger Software used in this study. We also would like to thank our management Sreenidhi Institute of Science and Technology for providing necessary facilities in completing our work.
References
[1] Saeidnia S, Gohari AR, Shahverdi AR, Permeh P, Nasiri M, Mollazadeh K. Biological activity of two red algae, Gracilaria salicornia and Hypneaflagelliformis from Persian gulf. Pharmacogn Res 2009;1:428–30.
[2] Uma R, Sivasubramanian V, Niranjali Devaraj S. Preliminary phycochemical analysis and in vitro antibacterial screening of green micro algae,Desmococcus olivaceous, Chlorococcum humicola and Chlorella vulgaris. J Algal Biomass Utln 2011; 2:74–81.
[3] Dai J, Mumper RJ. Plant phenolics: extraction, analysis and their antioxidant and anticancer properties. Molecules 2010;15:7313–52.
[4] Thilakarathna SH, Rupasinghe HPV. Flavonoid bioavailability and attempts for bioavailability enhancement. Nutrients 2013;5(9):3367–87.
[5] Joshi YB, Pratico D. The 5-lipoxygenase pathway: oxidative and in flammatory contributions to the Alzheimer's disease phenotype. Front Cell Neurosci 2015;8: 436.
[6] Johnson TL, Lloyd RV, Thor A. Expression of ras oncogene p21 antigen in normal and proliferative thyroid tissues. Am J Pathol 1987;127(1):60–5.
[7] Rajasri Yadavalli, Rao Ramgopal, Rao CS. Lipid accumulation studies inChlorella pyrenoidosausing customized photobioreactor-effect of nitrogen source, light intensity and mode of operation. Int J Eng Res Afr 2012;2(3):2446–53.
Table 5
Comparison of interactions of selected ligands with the active site residues of 3V99.
Compound Number of
Caffeoyl-D-glucose 3 Asp 442
Arg 438
Catechin 1 Leu 289 1.78 Ile 292, Gly
291 (2)
Epicatechin 0 None – Trp 144, Trp
144
Protocatechic acid 0 None – Arg 370 (3)
Gln 549
Table 6
Comparison of interactions of selected ligands with the active site residues of 4L9S.
Compound Number of
Caffeoyl-D-glucose 2 Ser 17
Asp 119
Catechin 2 Lys 147
Asp 119
Epicatechin 2 Asp 119
Ser 145
Protocatechic acid 3 Lys 18
[8] Deyab MD, Elkatony Taha, Ward Fatma. Qualitative and quantitative analysis of phytochemical studies on brown Seaweed. Dictyota dichotoma. IJEDR 2016:674–8. [9] Slinkard K, Singleton VL. Total phenolic analyses: automation and comparison with
manual method. Am J Enol Vitic 1997;28:49–55.
[10] Meda A, Lamien CE, Romito M, Millogo J, Nacoulma OG. Determination of the total phenolic,flavonoid and proline contents in burkina fasan honey, as well as their radical scavenging activity. Food Chem 2005;91:571–7.
[11] Berman HM, Battistuz T, Bhat TN, Bluhm WF, Bourne PE, Burkhardt K, et al. The protein data bank. Acta Crystallogr D Biol Crystallogr 2002;58:899.
[12] Johansson MU, Zeote V, Michielin O, Guex N. Defining and searching for structural motifs using deepviews/swiss-pdb viewer. BMC Bioinf 2012;13:173.
[13] National Center for Biotechnology Information. PubChem compound database; https://pubchem.ncbi.nlm.nih.gov/summary/summary.cgi?cid¼5934766 [accessed Feb. 22, 2011].
[14] Shaw DE, Lindorff Larsen K, Piana S, Palmo K, Maragakis P, Klepeis JL, et al. Improved side-chain torsion potentials for the amber ff99SB protein forcefield. Proteins 2010;78(8):1950–8.
[15] Trott O, Olson AJ. AutoDock Vina Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J Comput Chem 2010;31:455–61.
[16] Sanner MF. Python a programming language for software integration and development. J Mol Graph Model 1999;17:57–61.
[17] Morris GM, Huey R, Lindstrom W, Sanner MF, Belew RK, Goodsell DS, et al. Autodock4 and AutoDockTools4: automated docking with selective receptor flexiblity. J Comput Chem 2009;16:2785–91.
[18] Schr€odinger release 2017-2: MS jaguar. New York, NY: Schr€odinger, LLC; 2017. [19] Baviskar JW, Khandelwal SR. Extraction, detection and identification offlavonoids
from microalgae: an emerging secondary metabolite. Int J Curr Microbiol App Sci 2015;(Special Issue2):110–7.
[20] Ludmila Machů, Ladislava Misurcova, Vavra Jarmila Ambrozova, Jana Orsavova, Jirí Mlcek, Jirí Sochor, et al. Phenolic content and antioxidant capacity in algal food products. Molecules 2015;20:1118–33.
[21] Jayshree A, Jayashree S, Thangaraju N. Chlorella vulgaris and Chlamydomonas reinhardtii: effective antioxidant, antibacterial and anticancer mediators. Indian J Pharmaceut Sci 2016;78(5):575–81.
[22] Cha KH, Koo SY, Lee DU. Antiproliferative effects of carotenoids extracted from Chlorella ellipsoidea and Chlorella vulgarison human colon cancer cells. J Agric Food Chem 2008;56(22):10521–6.
[23] Safafar Hamed, van Wagenen Jonathan, Møller Per, Jacobsen Charlotte. Carotenoids, phenolic compounds and tocopherols contribute to the antioxidative properties of some microalgae species grown on industrial wastewater. Mar Drugs 2015;13:7339–56.
[24] Kyadari M, Fatma T, Azad R, Velpandian T. Evaluation of antiangiogenic and antiproliferative potential of the organic extract of green algaeChlorella pyrenoidosa. Indian J Pharmacol 2013;45:569–74.
[25] Riberio D, Freitas M, Tome SM, Silva AM, Laufer S, Lima JL, et al. Flavonoids inhibit COX-1 and COX-2 enzymes and cytokine/chemokine production in human whole blood. Inflammation 2015;38(2):858–70.
[26] Rathee P, Chaudhary H, Rathee S, Rathee D, Kumar V, Kohli K. Mechanism of action offlavonoids as anti-inflammatory agents: a review. Inflamm Allergy - Drug Targets 2009;8(3):229–35.
[27] Skrzypczak-Jankun E, McCabe NP, Selman SH, Jankun J. Curcumin inhibits lipoxygenase by binding to its central cavity: theoretical and X-ray evidence. Int J Mol Med 2000;6:521–6.
[28] Herowati Rina, Widodo Gunawan Pamudji. In: Kandemirli Fatma, editor. Molecular docking analysis: interaction studies of natural compounds to anti-inflammatory targets, quantitative structure-activity relationship. InTech; 2017.https://doi.org/ 10.5772/intechopen.68666. Available from:https://www.intechopen.com/books/ quantitative-structure-activity-relationship/molecular-docking-analysis-interaction-studies-of-natural-compounds-to-anti-inflammatory-targets.
[29] Sajkowska-Kozielewicz Joanna J, Kozielewicz Paweł, Barnes Nicholas M, Wawer Iwona, Paradowska Katarzyna. Antioxidant, cytotoxic, and antiproliferative activities and total polyphenol contents of the extracts ofGeissospermum reticulatum bark. Oxidative Medicine and Cellular Longevity 2016 2016. 8 pages,https://www. hindawi.com/journals/omcl/2016/2573580/cta/.
[30] Tow WW, Premie R, Jing H, Ajlouni S. Antioxidant and anti proliferation effects of extractable and nonextractable polyphenols isolated from apple waste using different extraction methods. J Food Sci 2011;76:T163–72.
[31] Mari Angela, Tedesco Idolo, Nappo Annunziata, Russo Gian Luigi, Malorni Antonio, Carbone Virginia. Phenolic compound characterisation and antiproliferative activity of“Annurca”apple, a southern Italian cultivar. Food Chem 2010;123: 157–64.
[32] Phosrithong N, Ungwitayatorn J. Molecular docking study on anticancer activity of plant-derived natural products. Med Chem Res 2010;19(8):817–35.