PENGARUH POLARITAS DAN KONSENTRASI EKSTRAK
ANDALIMAN TERHADAP PERTUMBUHAN Bacillus cereus
ABSTRAK
Ekstraksi andaliman secara bertingkat dengan heksana, etilasetat dan metanol, dan masing-masing menghasilkan ekstrak nonpolar, semipolar dan polar. Ekstrak etilasetat dan ekstrak metanol andaliman dapat menghambat pertumbuhan
Bacillus cereus, sedangkan ekstrak heksana tidak menunjukkan adanya aktivitas
penghambatan.
Secara umum ekstrak etilasetat menghasilkan penghambatan yang relatif lebih tinggi dibandingkan ekstrak metanol. Fase pertumbuhan eksponensial
B. cereus adalah fase yang paling peka terhadap ekstrak etilasetat dibandingkan
fase adaptasi dan stasioner. Konsentrasi ekstrak etilasetat 10 -50% (w/w) pada fase eksponensial menunjukkan diameter penghambatan terhadap B. cereus berkisar 12.42-19.22 mm. Nilai MIC dan MBC ekstrak etilasetat terhadap B. cereus sebesar 0.20 dan 1.20%. Aktivitas penghambatan ekstrak etilasetat lebih tinggi baik terhadap spora maupun sel vegetatif. Bila dibandingkan dengan antibiotik (penisilin G, streptomisin dan polimiksin) aktivitas antibakteri ekstrak andaliman terhadap B. cereus hanya ¼ kali lebih rendah.
PENDAHULUAN
Pemilihan pelarut organik yang digunakan dalam ekstraksi komponen bioaktif tanaman merupakan fak tor penting dan menentukan untuk mencapai tujuan dan sasaran ekstraksi komponen. Dengan mengetahui sifat senyawa yang akan diekstraksi dapat dipilih pelarut yang sesuai berdasarkan polaritasnya. Senyawa polar lebih mudah larut dalam pelarut polar dan senyawa nonpolar lebih mudah larut dalam pelarut nonpolar, sedangkan semipolar dapat larut diantara pelarut polar dan nonpolar. Derajat polaritas tergantung pada ketetapan dielektrik , semakin besar tetapan dielektrik, maka pelarut semakin polar (Ho ughton dan Raman 1998).
Houghton dan Raman (1998) lebih lanjut mengemukakan bahwa secara umum terdapat tiga metode ekstraksi, yaitu metode maserasi, refluks dan perkolasi. Selain itu dapat dilakukan ekstraksi bertingkat dengan menggunakan pelarut yang berbeda, secara berturut-turut mulai dengan pelarut nonpolar (n-heksana, siklo heksana, toluena dan kloroform) lalu menggunakan pelarut yang semipolar (etilasetat, diklorometan dan dietileter), kemudian dengan pelarut polar (metanol, etanol dan air). Dari proses ekstraksi bertingkat seperti ini akan
dihasilkan ekstrak awal (ekstrak kasar) yang mengandung berturut-turut senyawa nonpolar, semipolar dan polar (Hostettmann et al. 1997).
Hasil penelitian Ardiansyah (2001) yang menggunakan ekstraksi bertingkat metode maserasi dan refluks dengan pelarut heksana, etilasetat, dan metanol menunjukkan bahwa konsentrasi 5% ekstrak etilasetat andaliman mampu menghambat B. cereus sebesar 7.1 mm. Selanjutnya Yasni (2001) menyatakan bahwa ekstrak andaliman mengandung minyak atsiri yang dapat menyebabkan penghambatan terhadap bakteri patogen. Hasil pengujian ekstrak yang diperoleh dengan metode refluks menunjukkan aktivitas penghambatan lebih rendah dibandingkan dengan metode maserasi (Ardiansyah 2001). Penelitian ini bertujuan untuk mengetahui lebih mendalam pengaruh polaritas dan konsentrasi ekstrak andaliman terhadap aktivitas antibakteri pada beberapa fase pertumbuhan
B. cereus dengan menggunakan metode difusi sumur.
METODOLOGI Bahan dan alat
Bahan yang digunakan adalah buah andaliman Varietas S imanuk, yang berasal dari Medan dan diperoleh dari Pusat Pasar Senen Jakarta. Kultur Bacillus
cereus FNCC 134 diperoleh dari koleksi kultur Pusat Antar Universitas Pangan
dan Gizi, Universitas Gadjah Mada (UGM), Yogyakarta.
Analisis Proksimat (Apriyantono et al. 1989)
Pengujian proksimat yang dilakukan adalah kadar air (metode oven), kadar protein (metode Kjeldhal-mikro), kadar lemak (metode soxhlet), kadar abu total, kadar karbohidrat (by difference).
Ekstraksi dengan Metode Maserasi (Harbone 1996)
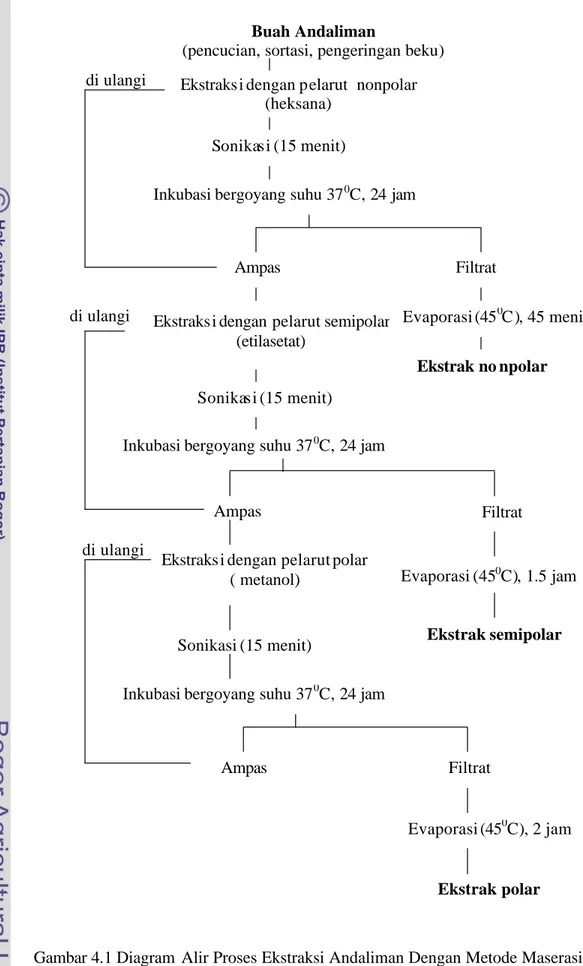
Ekstraksi dilakukan dengan perbandingan bahan dan pelarut 1:4 (w/v). Proses ekstraksi dilakukan secara bertingkat dengan metode maserasi, menggunakan pelarut heksana untuk memperoleh ekstrak nonpolar; ampas diekstraksi lebih lanjut dengan pelarut etilasetat untuk memperoleh ekstrak semipolar; dan kemudian ampas yang diperoleh diekstraksi dengan pelarut metanol untuk memperoleh ekstrak polar. Secara rinci dapat dilihat pada diagram alir proses ekstraks i menggunakan metode maserasi (Gambar 4.1).
Buah Andaliman
(pencucian, sortasi, pengeringan beku)
Gambar 4.1 Diagram Alir Proses Ekstraksi Andaliman Dengan Metode Maserasi Ekstraks i dengan pelarut nonpolar
(heksana) Sonikas i (15 menit)
Ampas
Inkubasi bergoyang suhu 370C, 24 jam
Filtrat
Ekstraks i dengan pelarut semipolar (etilasetat)
Evaporasi (450C), 45 menit
Sonikas i (15 menit)
Inkubasi bergoyang suhu 370C, 24 jam
Ekstrak no npolar
Filtrat
Evaporasi (450C), 1.5 jam Ekstraks i dengan pelarut polar
( metanol)
Sonikasi (15 menit)
Inkubasi bergoyang suhu 370C, 24 jam
Ampas Filtrat Evaporasi (450C), 2 jam Ampas Ekstrak polar Ekstrak semipolar di ulangi di ulangi di ulangi
Penentuan Fase Pertumbuhan B. cereus (Lin et al. 2000)
Sebanyak 10 µl suspensi diinokulasikan ke dalam 10 ml NB (nutrient
broth) dan diinkubasi pada suhu 370C selama 0, 1, 2, 3, 4, 5, 6, 8, 10, 12, 16, 20, 25 jam (Harrigan 1998); Lin et al. 2000). Pada setiap waktu inkubasi dilakukan penghitungan jumlah sel dengan metode pour plate menggunakan media NA (nutrient agar) (Harrigan 1998).
Pengujian Antibakteri Ekstrak Andaliman Menggunakan Metode Difusi Sumur (Garriga et al. 1993)
Pengujian ini menggunakan ekstrak etilasetat dan metanol masing-masing sebanyak 60 µl ekstrak dengan konsentrasi 0, 10, 20, 30, 40, dan 50% (w/w). Cara pembuatan konsentrasi masing -masing ekstrak andaliman dapat dilihat pada Lampiran 2, sedangkan kontrol menggunakan pelarutnya masing-masing. Penentuan aktivitas antibakteri berdasarkan zona penghambatan yang ditunjukkan dengan diameter areal bening.
Pengujian Kualitatif Komponen Fitokimia Ekstrak Andaliman (Harbone 1996)
Pengujian ini dilakukan untuk menentukan penggolongan ekstrak andaliman nonpolar, semipolar dan polar pada komponen alkaloid, fenol hidrokuinon, terpenoid, steroid, flavonoid dan saponin.
Penentuan MIC dan MBC Ekstrak Andaliman (Kubo et al. 1995)
Ke dalam 14 tabung ekstrak andaliman dengan konsentrasi 0.0, 0.2, 0.4, 0.8, 1.2, 1.6, 2.0, 2.4, 2.8, 3.2, 3.6, 4.0, 5.0, dan 6.0% (w/w) diinokulasikan 30 µl suspensi B. cereus, lalu diinkubasi 370C selama 24 dan 48 jam. Nilai MIC (%) yaitu konsentrasi minimum ekstrak yang dapat menghambat pertumbuhan bakteri uji sebanyak 90% selama 24 jam. Nilai MBC diperoleh dengan menentukan konsentrasi terendah dari 14 seri tabung uji yang menunjukkan penurunan pertumbuhan bakteri uji secara drastis (> 99.9%) setelah diinkubasi 48 jam dibandingkan dengan jumlah bakteri uji awal (nol jam).
Pengaruh pH terhadap Aktivitas Ekstrak Andaliman (Carson dan Riley 1995)
Sebanyak 25 ml agar yang mengandung B. cereus pada fase eksponensial (106 cfu/ml) dituangkan ke dalam cawan petri hingga ketebalan 4 mm. Setelah agar membeku, dibuat sumur dengan diameter 6 mm dan dimasukkan 60 µl ekstrak yang telah diatur pHnya masing-masing pada pH 4, 5, 6 dan 7. Penentuan pengaruh pH berdasarkan zona penghambatan yang efektif, ditunjukkan dengan diameter areal bening.
Pembandingan Potensi Bakterisidal Ekstrak Andaliman dengan Streptomisin, Polimiksin B dan Penisilin G (Lin et al. 2000)
Pengujian dilakukan dengan metode difusi sumur (Garriga et al. 1993) pada konsentrasi antibiotik masing -masing sebesar 10, 100, 1000 µg/l (Lewis
et al. 2004) untuk polimiksin B, streptomisin dan penisilin G, sedangkan
konsentrasi ekstrak andaliman 4000 µg/l (Ardiansyah 2001). Penentuan potensi bakterisidal dari ekstrak terhadap B. cereus didasarkan pada zona penghambatan yang ditunjukkan dengan diameter areal bening untuk masing-masing antibiotik dan ekstrak andaliman.
Pengujian Aktivitas Antibakteri Ekstrak Andaliman terhadap Spora (Nuraida et al. 1999)
Suspensi B. cereus mengandung 106 cfu/ml spora/sel vegetatif diuji dengan metode difusi sumur (Garriga et al. 1993) dengan ekstrak uji sebanyak 60 µl. Untuk mengetahui pembentukan spora pada B. cereus terlebih dahulu bakteri ditumbuhkan pada suhu 37OC dan diamati pembentukan sporanya setelah diinkubasi 24, 48, 72 dan 96 jam.
HASIL DAN PEMBAHASAN
Rendemen, Proksimat dan Sifat Fisiko Kimia Andaliman
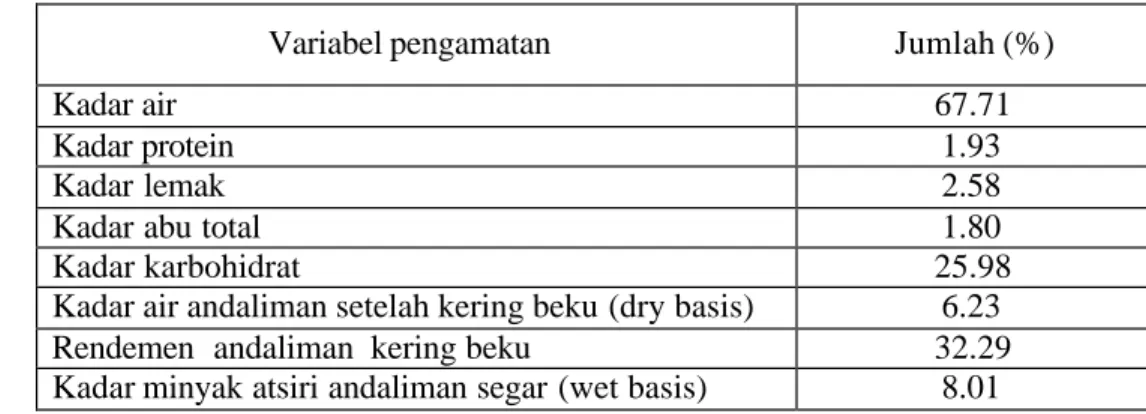
Hasil analisis proksimat andaliman dan rendemen minyak atsiri yang dihasilkan dapat disimak pada Tabel 4.1. Kadar lemak buah andaliman (2.58%) lebih tinggi daripada kadar protein (1.93%) sehingga hasil ekstraksi yang
diperoleh dengan pelarut nonpolar juga leb ih tinggi dibandingkan dengan ekstrak semipolar maupun polar. Rendemen ekstrak bubuk andaliman berdasarkan polaritas pelarut dan hasil analisis kualitatif komponen aktif dari masing-masing jenis ekstrak andaliman dapat dilihat pada Tabel 4.2.
Tabel 4.1 Analisis Proksimat dan Kandungan Minyak Atsiri Andaliman
Variabel pengamatan Jumlah (%)
Kadar air 67.71
Kadar protein 1.93
Kadar lemak 2.58
Kadar abu total 1.80
Kadar karbohidrat 25.98
Kadar air andaliman setelah kering beku (dry basis) 6.23
Rendemen andaliman kering beku 32.29
Kadar minyak atsiri andaliman segar (wet basis) 8.01
Tabel 4.2 Rendemen dan Hasil Analisis Kualitatif Komponen Aktif Ekstrak Andaliman
Komponen aktif Ekstrak Heksana Ekstrak Metanol Ekstrak Etilasetat Rendemen (%) 6.30 4.15 3.17 Alkaloid + + + Tanin - + - Fenol Hidrokuinon - + + Flavonoid + + + Triterpenoid + + + Saponin - + + Steroid + + + Keterangan: + : mengandung komponen - : tidak mengandung komponen
Kadar air bubuk andaliman cukup rendah (6.23%) dan rendemen buah setelah pengeringan beku relatif tinggi (32.29%). Demikian pula rendemen minyak atsiri andaliman segar yang diperoleh cukup tinggi, (8.01% w/w) dibandingkan dengan minyak atsiri biji atung sebanyak 1.79% (Murhadi 2002), sereh segar sebanyak 0.25 -0.50% (Oyen dan Nguyen 1999) atau minyak atsiri kunyit segar sebanyak 3 – 5% (Purseglove 1981). Rendemen yang relatif tinggi dan tingginya kandungan minyak atsiri buah andaliman ini merupakan potensi
untuk pemanfaatannya sebagai pengawet pangan alami. Profil komponen aktif minyak atsiri andaliman yang dilaporkan Yasni (2001) menyatakan terdapat 11 komponen dengan 5 komponen utama yaitu a-pinen, limonen, geraniol, sitronella dan geranil asetat. Komponen minyak atsiri tersebut dapat menghambat pertumbuhan bakteri patogen (E. coli, Pseudomonas, B. cereus, dan S. aureus) dan kapang (Fusarium sp, Penicillium sp dan Aspergillus flavus).
Pada Tabel 4.2 dapat dilihat bahwa rendemen dengan pelarut heksana, etilasetat dan metanol berturut-turut sebesar 6.30, 4.15 dan 3.17%. Menurut Houghton dan Raman (1998), ekstrak heksana (nonpolar) mengandung komponen yang bersifat nonpolar seperti lilin, lemak dan minyak atsiri, sedangkan ekstrak etilasetat (semipolar) sebagian besar mengandung senyawa-senyawa alkaloid, aglikon-aglikon, dan glikosida. Ekstrak metanol (polar) terutama mengandung kelompok senyawa gula, asam -asam amino, glikosida dan kelompok senyawa yang juga larut dalam petroleum eter, heksana, kloroform, etilasetat, etanol dan air dalam jumlah dan proporsi berbeda-beda.
Tingginya rendemen ekstrak nonpolar andaliman menunjukkan bahwa komponen yang dapat larut dalam heksana lebih banyak dibandingkan komponen semipolar (etilasetat) maupun komponen polar (metanol). Selain itu rendemen ekstrak nonpolar yang tinggi dapat juga disebabkan karena pelarut yang digunakan pertama adalah heksana. Kemampuan pelarut heksana untuk melarutkan banyak kelompok senyawa organik terutama dalam bentuk senyawa nonpolar suatu campuran organik dapat menghasilkan rendemen lebih besar.
Pada penelitian ini penghilangan pelarut dilakukan dengan menggunakan vakum rotavapor pada suhu 450C dan dikeringbekukan hingga terbentuk konsentrat pekat. Penghilangan pelarut pada suhu 450C kemungkinan komponen aktif andaliman masih belum mengalami perubahan, hal ini merupakan salah satu penyebab tingginya rendemen ekstrak yang diperoleh .
Komponen-komponen alkaloid dalam jaringan tanaman dapat diekstrak melalui beberapa metode, diantaranya dengan metode maserasi menggunakan aseton (Ramsewak et al. 1999); metode homogenisasi menggunakan pelarut etanol (polar) dalam etilasetat (Holstege et al. 1995); dan metode soxhlet
menggunakan metanol yang diakhiri dengan ekstraksi menggunakan diklorometan (Brooke et al. 1996).
Pada Tabel 4.2 dapat dilihat pula bahwa komponen aktif yang terdapat pada ekstrak etilasetat dan metanol adalah alkaloid, fenol hidrokuinon, flavonoid, triterpenoid, saponin, steroid, sedangkan tanin hanya terdapat pada ekstrak metanol. Puuponen-Pimia et al. (2001) melaporkan bahwa komponen fenolik yang terkandung pada ekstrak berry mampu menghambat beberapa bakteri Gram negatif, diantaranya adalah Salmonella enterica SH-5014 dan E. coli CM871.
Dari beberapa hasil penelitian dapat diketahui bahwa senyawa-senyawa fenolik tanaman yang telah terbukti memiliki aktivitas antibakteri diantaranya adalah turunan dari p-benzekuinon sepert i: 2,3-dimetoksi-5-metil-p-benzokuinon, 2,6-difenil-p-benzekuinon, dan 2,6-dimetoksi-p-benzokuinon (Nishina et al. 1991). Ekstrak metanol dari tanaman Mitracarpus scaber yang mempunyai komponen asam galat dan asam 3,4,5-trimetoksi asam benzoat, dapat menghambat pertumbuhan S. aureus (MIC 3.90 dan 0.97 µg/ml), sedangkan senyawa 4-metoksiasetofenon dan 3,4,5-trimetoksiasetofenon sangat efektif menghambat Candida albicans (MIC 1.95 µg/ml), serta komponen kaemferol-3-o-rutinosida, rutin dan psoralen mempunyai aktivitas rendah terhadap bakteri dan kapang, hal ini ditunjukkan dengan nilai MIC 125-500 µg/ml (Bisignano et al. 2000). Selain itu ekstrak nonpolar rosemary yang termasuk dalam komponen fenolik mampu menghambat bakteri Gram positif seperti B. cereus, S. aureus dan
Streptococcus pyogenes (Campo et al. 2000).
Senyawa-senyawa tanin yang telah terbukti memiliki aktivitas antibakteri terutama yang berasal dari ekstrak teh hijau yang mempunyai kemampuan untuk menghambat pertumbuhan bakteri Streptococcus mutans. Senyawa aktif dari tanin tersebut adalah galokatekin, epigalokatekin, dan epigalokatekin galat. Diantara ketiga senyawa aktif tersebut galokatekin mempunyai aktivitas antibakteri paling tinggi dengan nilai MIC 250 µg/ml. Penghambatan oleh ketiga senyawa aktif tersebut diduga karena adanya gugus hidroksil (Sakanaka et al. 1989). Tanin bersifat dapat menggumpalkan protein, dan membentuk kompleks dengan beberapa polisakarida, asam nukleat dan alkaloid, dan juga dapat mempengaruhi warna, serta berkontribusi terhadap rasa dan aroma (Shahidi dan Naczk 1995).
Steroid merupakan subklas dari triterpenoid dengan komponen C30
(Banthorpe 1996). Ekstrak metilen diklorida tanaman R. apicula mengandung tiga komponen steroid, yaitu kampesterol (4.61%), stigmaterol (18.47%) dan sitosterol (76.92%). Keller et al. (1998) melaporkan bahwa kandungan steroid yang mempunyai aktivitas antibakteri merupakan senyawa metabolit sekunder yang dihasilkan oleh fungi Fomitopsis pinicola.
Daun kedaung mengandung saponin dan antosianin yang keduanya berhubungan dengan antibakteri dan antiparasit (Zuhud et al. 2001). Senyawa saponin dan flavonoid dilaporkan memiliki daya antibakteri terhadap beberapa spesies bakteri, sedangkan senyawa terpen merupakan senyawa antibakteri utama dalam rempah (N aidu dan Davidson 2000).
Senyawa antibakteri alkaloid tanaman, diantaranya dilaporkan oleh Bhattacharyya et al. (1993) dari kulit kayu tanaman Clausena heptaphylla, Chakraborty et al. (1995) dari daun tanaman Clausena heptaphylla, dan Ramsewak et al. (1999) dari daun tanaman Murraya koenigii.
Pola Pertumbuhan B. cereus
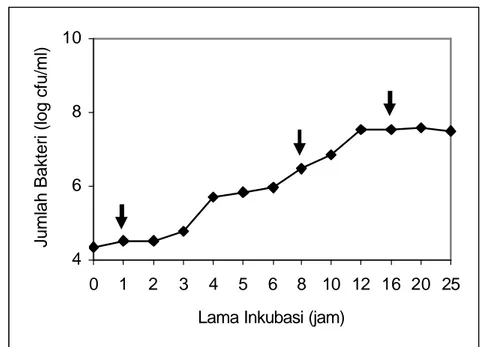
Hasil pengamatan pola pertumbuhan bakteri B. cereus dapat dilihat pada Gambar 4.2.
Gambar 4.2 Kurva Pola Pertumbuhan B. cereus
4 6 8 10
0 1 2 3 4 5 6 8 10 12 16 20 25
Lama Inkubasi (jam)
Fase adaptasi B. cereus berlangsung pada interval waktu 1-3 jam, sedangkan fase eksponensial berlangsung sekitar 9 jam mulai dari jam ke 3 sampai dengan jam ke 12, dan stasioner berlangsung mulai dari jam ke 16 sampai dengan jam ke 25.
Berdasarkan data tersebut ditetapkan bahwa untuk mewakili fas e adaptasi dipilih 1 jam, sedangkan fase eksponensial dan fase stasioner masing-masing 8 jam dan 16 jam. Jumlah sel B. cereus selama fase adaptasi belum meningkat, yaitu pada jam ke-3 sebesar 104 cfu/ml. Fase eksponensial mulai jam ke-3 sampai jam ke-12 meningkat pesat sampai 106 cfu/ml dan setelah jam ke-16 sampai jam ke-25 jumlahnya tetap (fase stasioner), yaitu 108 cfu/ml. Jumlah sel B. cereus setelah 1 jam inkubasi sebanyak 5.6x104 cfu/ml, setelah 8 jam inkubasi meningkat sampai 7.4x106 cfu/ml dan mencapai jumlah tetap yaitu 3.4x108 cfu/ml setelah 16 jam inkubasi. Data selengkapnya disajikan pada Lampiran 1. Fase adaptasi
B. cereus berlangsung selama 3 jam dan merupakan persiapan untuk fase
berikutnya. Menurut Madigan et al. (2003) fase eksponensial pertumbuhan bakteri sangat cepat, teratur, dan semua bahan dalam sel berada dalam keadaan seimbang. Ultee et al. (2000) melaporkan bahwa kisaran fase pertumbuhan B. cereus adalah fase adaptasi 1-3 jam, fase eksponensial 8-10 jam sedangkan fase stasioner berkisar 16 -20 jam.
Pengaruh Polaritas dan Konsentrasi Ekstrak Andaliman terhadap B. cereus
Hasil pengujian aktivitas antibakteri ekstrak heksana (nonpolar), etilasetat (semipolar) dan metanol (polar) terhadap B. cereus dengan metode difusi sumur dapat dilihat pada Lampiran 2, 3 dan 4. Secara umum ekstrak etilasetat (semipolar) memiliki daya penghambatan lebih tinggi dibandingkan ekstrak metanol (polar) terhadap B. cereus pada masing-masing fase pertumbuhan. Ekstrak heksana (nonpolar) andaliman tidak menunjukkan efek penghambatan terhadap sel B. cereus.
Ekstrak Nonpolar
Hasil pengujian aktivitas antibakteri ekstrak nonpolar andaliman terhadap sel vegetatif B. cereus dapat dilihat pada Lampiran 2, 3, dan 4. Ekstrak nonpolar
andaliman tidak menun jukkan adanya aktivitas antibakteri pada fase adaptasi, fase eksponensial maupun fase stasioner. Hal ini dapat dijelaskan bahwa media NA yang digunakan bersifat polar, sedangkan ekstrak nonpolar lebih dominan mengandung komponen minyak atsiri, dimana ekstrak dalam media NA tidak mampu berdifusi secara baik sehingga tidak menunjukkan aktivitas penghambatan.
Polaritas yang rendah pada ekstrak heksana disebabkan karena mengandung minyak atsiri, bahan non minyak seperti lilin, sterol dan sedikit senyawa fenolik. Menurut Nychas (1995) komponen bioaktif yang berperan sebagai antibakteri dalam ekstrak heksana adalah minyak atsiri dan fenolik. Ekstrak heksana mengandung minyak atsiri yang relatif tinggi (Ozean dan Erkmen 2001) dan minyak atsiri ini mempunyai aktivitas antibakteri, namun keberadaan minyak atsiri dalam ekstrak tersebut tidak menunjukkan penghambatan pada B. cereus. Hasil analisis identifikasi komponen aktif ekstrak heksana pada Tabel 4.2 menunjukkan adanya kandungan alkaloid, flavonoid, triterpenoid dan steroid yang relatif rendah dibandingkan dengan ekstrak etilasetat dan ekstrak metanol. Ekstrak heksana ini ternyata tidak mengandung tanin, fenol hidrokuinon dan saponin. Kemungkinan komponen aktif yang terdapat pada ekstrak heksana tersebut pada konsentrasi 10-50% masih belum dapat menghambat pertumbuhan B. cereus, sedangkan pada ekstrak etilasetat dan metanol pada konsentrasi tersebut sudah menunjukkan penghambatan.
Hasil penelitian Kanazawa et al. (1995) melaporkan bahwa ekstrak heksana (senyawa minyak atsiri dan lipida lainnya) yang mempunyai ukuran molekul besar tidak dapat masuk berpenetrasi dalam dinding sel. Ukuran molekul besar tersebut akan menjadi penghalang masuknya komponen minyak atsiri maupun senyawa fenolik ke dalam sel akibatnya sel tetap akan tumbuh.
Ekstrak Semipolar
Senyawa yang bertanggungjawab sebagai antibakteri dalam ekstrak etilasetat adalah senyawa semipolar yang merupakan senyawa fenolik (Hiserodt, 1998), steroid, alkaloid, triterpenoid, flavonoid dan saponin (Tabel 4.2). Aktivitas ekstrak etilasetat andaliman pada setiap fase pertumbuhan (fase adaptasi, fase
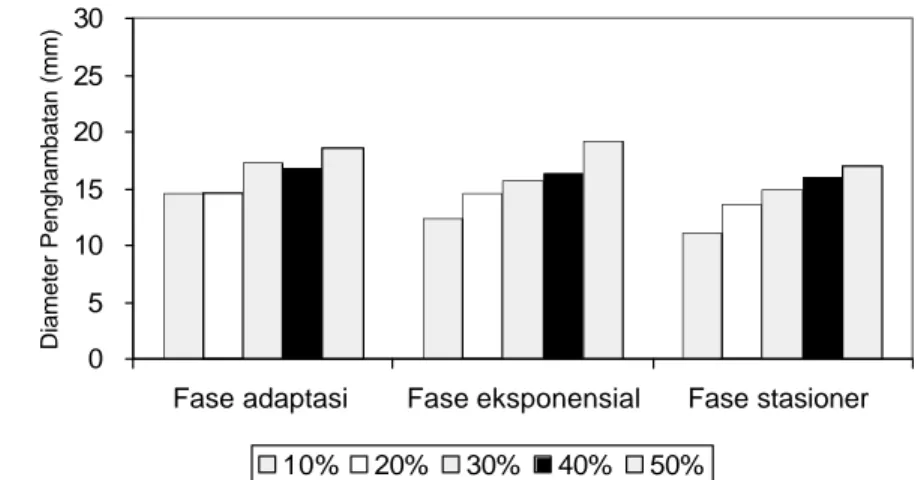
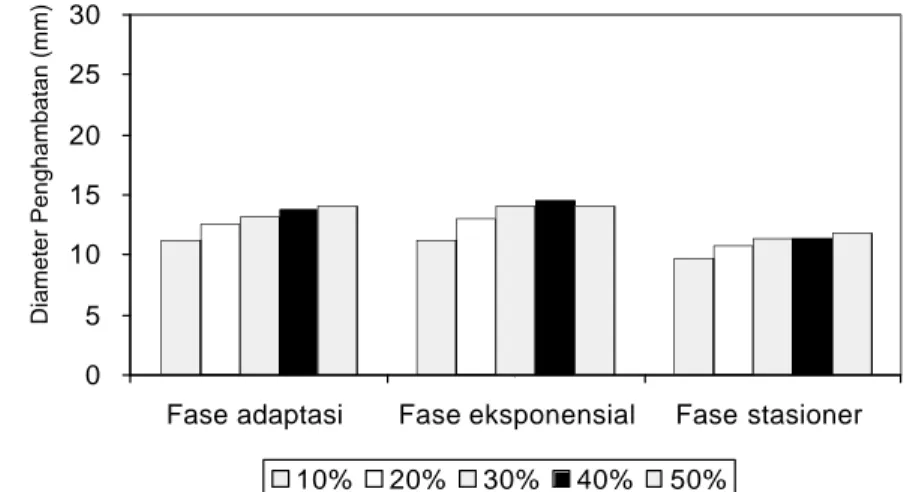
eksponensial dan fase stasioner) berpengaruh sangat nyata (p<0.01) dan menunjukkan penghambatan tertinggi (Lampiran 11, 12 dan13). Gambar diameter penghambatan masing-mas ing bakteri patogen dapat dilihat pada Lampiran 10. Semakin tinggi konsentrasi ekstrak etilasetat maka diameter penghambatan juga semakin tinggi. Aktivitas antibakteri ekstrak etilasetat terhadap B. cereus (Lampiran.2, 3, 4) paling tinggi pada fase eksponensial dibandingkan fase adaptasi dan stasioner. Konsentrasi ekstrak etilasetat 10-50% pada fase eksponensial menghasilkan penghambatan sebesar 12.42-19.22 mm (Gambar 4.3).
Gambar 4.3 Pengaruh Konsentrasi Ekstrak Etilasetat Andaliman pada Fase Pertumbuhan B. cereus
Tingginya aktivitas ekstrak etilasetat andaliman terhadap B. cereus dengan metode maserasi bertingkat telah dilaporkan oleh Yasni et al. (2001) sebesar 10.62 mm, sedangkan dengan metode mas erasi ekstrak metanol memiliki aktivitas antimikroba paling tinggi, yaitu sebesar 15.72 mm terhadap A. flavus. Pada Gambar 4.3 terlihat bahwa ekstrak etilasetat pada konsentrasi 10-50% memiliki penghambatan yang berbeda pada setiap fase pertumbuhan B. cereus. Fase pertumbuhan eksponensial B. cereus memiliki diameter penghambatan tertinggi yaitu sebesar 19.22 mm pada konsentrasi 50%. Fase eksponensial merupakan fase pertumbuhan dimana pembelahan sel terjadi sangat cepat, dan semua bahan dalam sel berada dalam keadaan pertumbuhan seimbang (balan ced growth). Maka dengan adanya ekstrak andaliman dapat mengganggu aktivitas pertumbuhannya.
0 5 10 15 20 25 30
Fase adaptasi Fase eksponensial Fase stasioner
Diameter Penghambatan (mm)
Terutama dengan adanya ekstrak etilasetat yang banyak mengandung komponen antibakteri seperti alkaloid, fenol hidrokuinon, steroid, flavonoid, triterpenoid dan saponin akan dapat mengganggu proses pertumbuhan dan pembelahan sel, menyebabkan terjadinya kerusakan sel.
Konsentrasi fenolik, alkaloid dan steroid dalam ekstrak etilasetat cukup tinggi untuk berdifusi dan menghambat pertumbuhan bakteri. Kemampuan senyawa semipolar untuk menghambat pertumbuhan bakteri berkaitan dengan komponen dinding sel bakteri yang tidak bersifat absolut hidrofobik maupun absolut hidrofilik. Kanazawa et al. (1995) menyatakan bahwa suatu senyawa semipolar mempunyai aktivitas antibakteri yang maksimum, karena untuk interaksi suatu senyawa antibakteri dengan bakteri diperlukan keseimbangan hidrofilik-lipofilik (HLB: hydrophilic-lipophilic balance). Kemungkinan senyawa semipolar mempunyai afinitas lebih tinggi untuk berinteraksi dengan dindin g sel, sehingga ekstrak semipolar lebih efektif menghambat pertumbuhan B. cereus daripada ekstrak metanol (polar) dan heksana (nonpolar).
Ekstrak Polar
Ekstrak polar andaliman memberikan pengaruh sangat nyata (p<0.01) terhadap pertumbuhan B. cereus (Lampiran 14, 15 dan 16). Pola penghambatan relatif sama dengan ekstrak etilasetat, tetapi ekstrak metanol memiliki diameter penghambatan lebih rendah (14.05 mm). Aktivitas antibakteri ekstrak polar menunjukkan diameter penghambatan yang berbeda dengan aktivitas ekstrak semipolar (Lampiran 2, 3, 4). Perbedaan utama adalah pada konsentrasi yang sama diameter penghambatan ekstrak etilasetat lebih tinggi daripada ekstrak metanol pada setiap fase pertumbuhan B. cereus (Gambar 4.4). B. cereus mempunyai ketahanan lebih tinggi terhadap ekstrak polar dibandingkan dengan ekstrak semipolar.
Campo et al. (2000) melaporkan bahwa ekstrak etanol rosemary pada konsentrasi 100 mg/ml dapat menghambat pertumbuhan bakteri Gram negatif
E. coli, Salmonella enteritidis dan E. caro tovora, sedangkan bakteri Gram positif
yaitu B. cereus CIP51.27, B. cereus Z4234, S. aureus, L. monocytogenes dan
sifat antibakteri yaitu yang termasuk dalam 3 grup fenolik yaitu asam fenolik, flavonoid dan diterpenoid fenolik.
Gambar 4.4 Pengaruh Konsentrasi Ekstrak Metanol Andaliman pada Fase Pertumbuhan B. cereus
Komponen yang banyak terdapat pada tumbuh-tumbuhan dan bersifat polar antara lain senyawa dari golongan fenolik. Mekanisme komponen antibakteri fenolik pada umumnya akan berinteraksi dengan protein yang ada pada dinding sel atau sitoplasma melalui ikatan hidrogen dan interaksi hidrofobik (Naidu dan Davidson 2000). Mekanisme lain dari ekstrak polar andaliman dalam menghambat pertumbuhan bakteri kemungkinan adalah dengan mengganggu aktivitas enzim dalam sel. Penelitian yang dilakukan oleh Wendaken dan Sakaguchi (1995) menunjukkan bahwa ekstrak air dan ekstrak etanol dari cengkeh, kayu manis dan saga yang bersifat polar akan menghambat enzim dekarboksilase khususnya histidin, lisin dan ornitin dekarboksilase dari bakteri
E. aerogenes.
Selanjutnya Hiserodt et al. (1998) melaporkan bahwa polaritas suatu senyawa mempengaruhi aktivitas antibakteri, seperti 6-gingerol yang mempunyai rantai alkil lebih polar daripada 10-gingerol yang memberikan penghambatan yang lebih rendah terhadap Mycobacterium avium. Fenomena ini menunjukkan bahwa senyawa polar cenderung mempunyai aktivitas antibakteri yang lebih rendah. Pada pengujian tahap selanjutnya digunakan hanya ekstrak etilasetat dan
0 5 10 15 20 25 30
Fase adaptasi Fase eksponensial Fase stasioner
Diameter Penghambatan (mm)
10% 20% 30% 40% 50% `
ekstrak metanol, karena hasil pengamatan aktivitas ekstrak heksana terhadap
B. cereus dengan metode difusi sumur tidak menunjukkan adanya penghambatan.
Ekstrak metanol memiliki aktivit as lebih rendah daripada ekstrak etilasetat. Moshi dan Mbwambo (2005) menyatakan bahwa ekstrak semipolar (etilasetat) mampu menghambat bakteri E. coli dan B. anthracis dengan diamater hambat lebih besar daripada ekstrak polar (etanol). Demikian juga penelitian Springfield et al. (2003) ekstrak etilasetat tanaman Carpobrotus muirii dan C.
quadrifidus lebih berpotensi dalam menghambat S. aureus dan Mycobacterium smegmatis daripada ekstrak air.
Potensi Aktivitas Ekstrak Andaliman Dibandingkan dengan Antibiotik
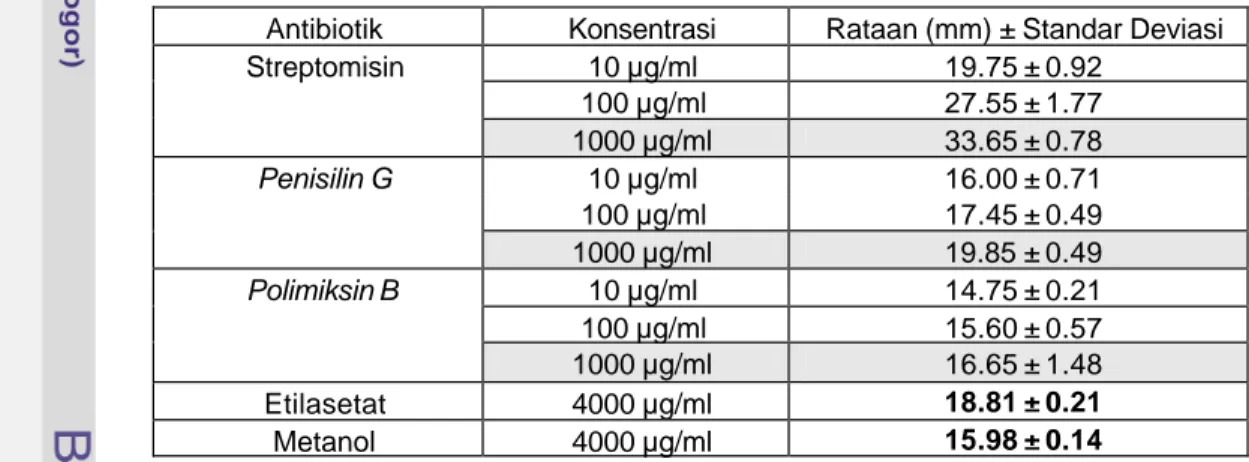
Hasil pengamatan diameter penghambatan ekstrak etilasetat, ekstrak metanol dan beberapa jenis antibiotik pada beberapa konsentrasi terhadap bakteri
B. cereus dapat dilihat pada Tabel 4.3.
Tabel 4.3 Diameter Penghambatan Beberapa Antibiotik dan Ekstrak Andaliman terhadap B. cereus
Antibiotik Konsentrasi Rataan (mm) ± Standar Deviasi
Streptomisin 10 µg/ml 19.75 ± 0.92 100 µg/ml 27.55 ± 1.77 1000 µg/ml 33.65 ± 0.78 Penisilin G 10 µg/ml 16.00 ± 0.71 100 µg/ml 17.45 ± 0.49 1000 µg/ml 19.85 ± 0.49 Polimiksin B 10 µg/ml 14.75 ± 0.21 100 µg/ml 15.60 ± 0.57 1000 µg/ml 16.65 ± 1.48 Etilasetat 4000 µg/ml 18.81 ± 0.21 Metanol 4000 µg/ml 15.98 ± 0.14
Antibiotik streptomisin menunjukkan penghambatan tertinggi (33.65 ± 0.78 mm), selanjutnya diikuti oleh penisilin G (19.85 ± 0.49 mm) dan polimiksin B (16.65 ± 1.48 mm) masing-masing pada konsentrasi 1000 µg/ml. Konsentrasi yang digunakan untuk melarutkan ekstrak etilasetat dan metanol adalah 4000 µg/ml (Ardiansyah 2001). Pada konsentrasi 4000 µg/ml pengaruh ekstrak etilasetat dan ekstrak metanol terhadap B. cereus masing-masing sebesar 18.81 ± 0.21 mm dan 15.98 ± 0.14 mm. Hal ini menunjukkan bahwa ekstrak
etilasetat memiliki potensi penghambatan yang setara dengan penisilin G, dan ekstrak metanol pada konsentrasi 4000 µg/ml memiliki potensi penghambatan yang setara dengan polimiksin B pada konsentrasi 1000 µg/ml. Potensi penghambatan ekstrak etilasetat dan ekstrak metanol pada konsentrasi 4000 µg/ml sekitar 1/8 dari kekuatan penghambatan streptomisin, dan sekitar 1/4 dari kekuatan penghambatan antibiotik penisilin G dan polimiksin B. Hal ini menunjukkan bahwa kekuatan ekstrak etilasetat dan metanol juga efektif menghambat bakteri Gram positif dan Gram negatif terutama S. aureus dan dapat di duga mekanisme aktivitas antibakteri serupa dengan polimiksin B, yaitu mengganggu keutuhan membran sel bakteri
Penisilin G merupakan antibiotik yang berspektrum luas dan biasanya efektif terhadap kebanyakan bakteri Gram positif dan Gram negatif terutama strain Staphylococcus aureus dan enterokokus yang biasanya menimbulkan penyakit infeksi pencernaan, kardiovaskular, kulit, dan lain sebagainya. Streptomisin dapat menghambat sintesis protein pada bakteri Gram positif dan Gram negatif seperti enterokokus pada penyakit endokarditis, tuberkulosis terutama pada sub unit ribosom 30S. Polimiksin bersifat bakterisidal terhadap bakteri Gram negatif berbentuk basil biasanya digunakan untuk mengobati infeksi pada permukaan kulit dan mata.
Penisilin G merupakan antibiotik yang menghambat sintesis dinding sel bakteri. Dinding sel bakteri, terdiri dari polipeptidoglikan yaitu suatu kompleks polimer mukopeptida (glikopeptida). Penisilin G menghambat reaksi yang paling dini dalam proses sintesis dinding sel. Oleh karena tekanan osmotik dalam sel bakteri lebih tinggi daripada di luar sel, maka kerusakan dinding sel bakteri akan menyebabkan terjadinya lisis, yang merupakan dasar efek bakterisidal pada bakteri yang peka.
Mekanisme penghambatan antibiotik penisilin G deng an cara penggabungan gugus amino bebas asam penisilanat pada gugus-gugus karboksil bebas dari berbagai jenis radikal. Berbagai radikal (R) yang terikat pada asam amino penisilanat menentukan sifat-sifat farmakologik. Penisilin memiliki daya kerja tertinggi terhadap bakteri Gram positif tetapi tidak aktif terhadap bakteri Gram negatif (Jawetz et al. 1996).
Polimiksin B merupakan antibiotik yang mengganggu keutuhan membran sel bakteri. Polimiksin B sebagai senyawa amonium kuaterner dapat merusak membran sel setelah bereaksi dengan fosfat pada fosfolipid membran sel bakteri. Polimiksin B dapat mengubah tegangan permukaan (surface active agents) sehingga merusak permeabilitas selektif dari membran sel bakteri. Kerusakan membran sel menyebabkan keluarnya berbagai komponen penting dari dalam sel seperti protein, asam nukleat, nukleotida dan lain -lain (Setiabudy dan Gan 1999).
MIC dan MBC Ekstrak Andaliman
Hasil pengujian nilai MIC dan MBC ekstrak andaliman terhadap bakteri
B. cerus berbeda-beda. Ekstrak etilasetat dan metanol mengandung komponen
aktif yang dapat menghambat dan membunuh B. cereus. B. cereus lebih peka terhadap ekstrak etilasetat dengan nilai MIC sebesar 0.20%, sedangkan ekstrak metanol memiliki nilai MIC sebesar 0.80% (w/w). Nilai MBC ekstrak etilasetat terhadap B. cereus adalah 1.20% dibandingkan ekstrak metanol sebesar 1.60% (w/w) (Lampiran 29).
Tingkat kepolaran mempengaruhi penghambatan terhadap sel. Meurut Davidson dan Branen (1993), semakin menurun polaritas (mendekati nonpolar) akan semakin efektif menghambat bakteri Gram positif dibandingkan dengan bakteri Gram negatif (). Hal ini sejalan pula dengan hasil–hasil penelitian dari Farag et al. (1989) dan Kim et al. (1995) yang membuktikan bahwa komponen – komponen minyak atsiri yang bersifat semipolar sampai nonpolar, lebih kuat daya antibakterinya terhadap kelompok bakteri Gram positif dibandingkan kelompok bakteri Gram negatif (Friedman et al. 2004a). Bakteri Gram positif mempunyai kecenderungan lebih peka terhadap senyawa semipolar dibandingkan dengan bakteri Gram negatif. Hal ini disebabkan karena perbedaan struktur dinding sel bakteri. Pada bakteri Gram positif sebagian besar dinding selnya terdiri dari lapisan peptidoglikan dan asam teikoat sehingga mudah dilewati komponen ekstrak yang bersifat hidrofilik.
Ekstrak etilasetat memberi pengaruh peka B. cereus d ibandingkan dengan ekstrak metanol. Setiap zat yang dapat menghambat salah satu langkah dalam biosintesis peptidoglikan akan menyebabkan dinding sel bakteri yang tumbuh
menjadi lemah dan sel akan mengalami lisis (Jawetz et al. 1996). Komponen -komponen ekstrak etilasetat dan metanol diduga berikatan dengan a-karboksil residu alanin ujung dari satu rantai peptidoglikan yang menghambat sintesis dinding sel sehingga sel akan mengalami kerusakan dan lisis.
Pengaruh pH terhadap Aktivitas Antibakteri Ekstrak Andaliman
Ekstrak etilasetat dan ekstrak metanol memiliki daya hambat yang berpengaruh tidak nyata (p>0.01) pada tingkat keasaman atau pH ekstrak. Kisaran diameter penghambatan ekstrak etilasetat terhadap B. cereus secara berurutan adalah (18.23-17.15 mm) (Lampiran 32). Diameter penghambatan ekstrak etilasetat andaliman pada pH 4 paling tinggi terhadap sel B. cereus (sebesar 18.23 mm), sedangkan rataan diameter penghambatan ekstrak metanol pada pH 4 - pH 7 adalah sebesar 11.75-10.55 mm (Lampiran 33). Hasil ini menunjukkan bahwa pada kisaran pH 4 sampai dengan pH 7 ekstrak metanol memiliki daya penghambatan relatif sama, sedangkan penghambatan ekstrak metanol lebih rendah dibandingkan ekstrak etilasetat.
Perbedaan kemampuan penghambatan ekstrak etilasetat dibandingkan ekstrak metanol diduga bahwa kandungan komponen senyawa antibakteri lebih banyak terdapat pada ekstrak etilasetat dibandingkan dengan ekstrak metanol. Pada ekstrak etilasetat komponen hidrofobik dan hidrofilik masing-masing terdapat dalam ekstrak dan memiliki aktivitas antibakteri yang tinggi. Akibatnya daya hambat tertinggi diperoleh pada ekstrak etilasetat karena komponen yang bersifat hidrofobik dan hidrofilik mampu masuk dalam membran sel dan menghambat metabolisme sel.
Mekanisme penghambatan pada pH rendah disebabkan oleh kondisi asam sel yang bereaksi untuk mempertahankan pH konstan dalam sel. Jika pH diturunkan maka proton yang terdapat dalam jumlah tinggi dalam medium akan masuk ke dalam sel. Proton (ion H+) dari asam masuk dalam sel melalui gradien proton transmembran (Ray 2001), hal ini menyebabkan pH sitoplasma menurun. Penurunan pH sitoplasma menyebabkan enzim-enzim bekerja untuk mengembalikan pH intern al sel menjadi pH normal (Booth 1985). Proton ini harus dikeluarkan untuk mencegah terjadinya pengasaman dan denaturasi komponen
-komponen sel. Aktivitas mengembalikan pH internal sel menjadi pH normal memerlukan banyak energi. Bila energi yang dibutuhkan dalam jumlah tinggi, maka metabolisme sel akan mengganggu sehingga lama kelamaan sel akan mengalami kematian (Fardiaz 1992).
Aktivitas Antibakteri Ekstrak Andaliman terhadap Spora B. cereus
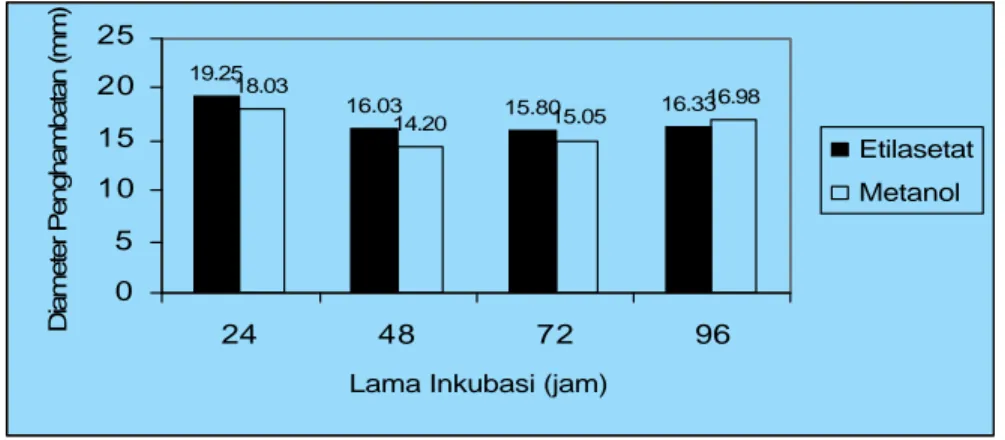
Pengujian aktivitas antibakteri spora bakteri B. cereus dilakukan untuk mengetahui aktivitas antibakteri ekstrak andaliman dibandingkan dengan sel vegetatif. Pengujian ini dilakukan pada kultur cair yang berumur 24, 48, 72 dan 96 jam, karena pada interval umur tersebut hampir semua sel vegetatif telah membentuk spora. Pengujian aktivitas antibakteri ekstrak etilasetat dan ekstrak metanol terhadap spora dan sel vegetatif B. cereus disajikan pada Gambar 4.5 dan Gambar 4.6. Selanjutnya dapat dilihat perbedaan aktivitas antibakteri ekstrak etilasetat dan ekstrak metanol terhadap spora bakteri, dan bila dibandingkan dengan sel vegetatifnya. Pada Gambar 4.5 diameter penghambatan meningkat dengan bertambahnya lama inkubasi hingga 96 jam sedangkan pada sel vegetatif sebaliknya (Gambar 4.6). Diameter penghambatan terhadap sel vegetatif umumnya lebih tinggi bila dibandingkan penghambatan terhadap spora B. cereus (Lampiran 36 dan 37) dengan perbedaan sekitar 7.83 mm selama 24 jam.
12.43 13.73 14.63 16.05 11.43 12.05 12.15 12.60 0 5 10 15 20 25 24 48 72 96
Lama Inkubasi (jam)
Diameter Penghambatan (mm)
Etilasetat Metanol
Gambar 4.5 Aktivitas Antibakteri Ekstrak Andaliman terhadap Spora B. cereus
Hal ini menunjukkan bahwa spora mempunyai ketahanan yang lebih tinggi dibandingkan dengan sel vegetatifnya. Lama inkubasi B. cereus hingga 24-96 jam
akan menyebabkan terbentuknya spora yang semakin banyak untuk mempertahankan diri, sehingga dengan adanya ekstrak andaliman pertumbuhan spora akan terhambat.
19.25 16.03 15.80 16.33 18.03 14.20 15.05 16.98 0 5 10 15 20 25 24 48 72 96
Lama Inkubasi (jam)
Diameter Penghambatan (mm)
Etilasetat Metanol
Gambar 4.6 Aktivitas Antibakteri Ekstrak Andaliman terhadap Sel Vegetatif B. cereus
Pengaruh ekstrak etilasetat pada spora B. cereus yang diinkubasi 96 jam dapat menghambat sebesar 16.05 mm, sedangkan diameter penghambatan ekstrak metanol lebih rendah, yaitu sebesar 12.60 mm. Berbeda dengan diameter penghambatan pada sel vegetatif B. cereus, lama inkubasi 24 jam pertama sudah menunjukkan penghambatan tertinggi dibandingkan dengan lama inkubasi 96 jam. Diameter penghambatan ekstrak etilasetat andaliman pada sel vegetatif dengan lama inkubasi 24 jam menunjukkan penghambatan lebih tinggi (19.25 mm), sedangkan pada ekstrak metanol diameter penghambatannya lebih rendah (18.03 mm).
Ultee et al. (1998) melaporkan bahwa spora bakteri lebih tahan terhadap aktivitas senyawa antibakteri alami dibandingkan dengan sel vegetatif. Hal ini ditunjukkan pada karvakrol yang mempunyai aktivitas sporosidal terhadap spora
B. cereus, baik pada konsentrasi 1.75 maupun 2.0 mmol/l. Spora lebih tahan
terhadap ekstrak metanol andaliman dibandingkan dengan sel vegetatif. Pada konsentrasi 2 mmol/l diperlukan waktu inkubasi 37 menit untuk membentuk spora sebanyak 2 log (cfu/ml), sedangkan untuk sel vegetatif diperlukan waktu inkubasi selama 20 menit. Nuraida et al. (1999) melaporkan bahwa ekstrak biji picung mampu menghambat spora B. cereus dengan areal penghambatan sebesar 1.0 mm, sedangkan terhadap sel vegetatif ekstrak tersebut mampu menghasilkan areal
penghambatan yang lebih besar (4.2 mm). Perbedaan ketahanan ini diduga karena perbedaan struktur fisik dan kimia antara spora dengan sel vegetatifnya (Cano dan Calome 1986; Friedman et al. 2004b).
Pada spora terdapat lapisan terluar yang tipis dan lembut yang disebut eksosporium. Di bawah lapisan eksosporium terdapat suatu lapisan lagi yang disebut dengan bungkus spora (coat spore ) yang terdiri dari satu lapisan atau berlapis-lapis yang membentuk struktur yang mirip dengan dinding sel. Adanya struktur yang berlapis-lapis pada spora akan mengakibatkan terhambatnya penetrasi ekstrak etilasetat maupun ekstrak metanol andaliman, disamping kandungan air yang lebih rendah (15%) bila dibandingkan dengan sel vegetatif (75%) juga akan menyebabkan spora lebih tahan dibandingkan dengan sel vegetatifnya (Cano dan Colome 1986). Selain itu kandungan asam dipikolinat dan tingginya kandungan ion kalsium yang hanya dimiliki oleh spora ju ga merupakan faktor yang dapat menyebabkan spora tahan terhadap perlakuan kimia (Fardiaz 1992).
SIMPULAN
Konsentrasi dan polaritas ekstrak andaliman mempengaruhi aktivitas antibakteri. Ekstrak etilasetat (semipolar) menunjukkan aktivitas yang lebih tinggi daripada ekstrak metanol (polar) terhadap B. cereus. Aktivitas antibakteri ekstrak andaliman dipengaruhi oleh fase pertumbuhan B. cereus. Ekstrak etilasetat lebih menghambat pada fase eksponensial (19.22 mm), sedangkan ekstrak metanol lebih efektif pada fase adaptasi (14.15 mm). Nilai MIC ekstrak etilasetat terhadap
B. cereus adalah 0.2% dan nilai MBC 1.20%.
Ekstrak etilasetat dan metanol berpotensi sebagai antibiotik, yaitu pada konsentrasi 4000 µg/ml sekitar 1/8 dari streptomisin, dan sekitar 1/4 dari penisilin G dan polimiksin B masing-masing pada konsentrasi 1000 µg/ml. Dengan demikian ekstrak andaliman dapat menghambat spora dan sel vegetatif B. cereus.
Ekstrak etilasetat dan metanol pada pH 4-7 menunjukkan tidak berpengaruh nyata (p>0.01) terhadap B. cereus. Kemampuan ekstrak etilasetat dalam menghambat spora B. cereus cukup baik, dimana diameter penghambatannya
setelah 96 jam lebih besar (16.05 mm) dibandingkan ekstrak metanol (12.60 mm). Demikian pula pada sel vegetatif, penghambatan ekstrak etilasetat andaliman dengan lama inkubasi 24 jam menunjukkan penghambatan lebih tinggi (19.25 mm) dibandingkan ekstrak metanol (18.03 mm).
DAFTAR PUSTAKA
Apriyantono A, Fardiaz D, Puspitasari NL, Yasni S, dan Budiyanto S. 1989.
Petunjuk Laboratorium Analisis Pangan. PAU Pangan dan Gizi IPB.
Bogor.
Ardiansyah. 2001. Teknik ekstraksi komponen antimikroba buah andaliman (Zanthoxylum acanthopodium DC) dan antarasa (Litsea cubeba). [skripsi]. Bogor: Fakultas Teknologi Pertanian, Institut Pertanian Bogor.
Banthorpe DV. 1996. Terpenoids. Di dalam: Mann J, Davidson RS, Hobbs JB, Banthorpe DV, Harbone JB, editor. Natural Product Their Chemistry and Biological Significance. Harlow: Longman. Hlm 289-359.
Bhattaacharyya P, Biswas GK, Barua AK, Saha C, Poy IB, dan Chowdhury BK. 1993. Clausenalene, a carbazole alkaloid from Clausena heptaphylla.
Phytochemistry. 33(1):248-250.
Bisignano G, Sanogo R, Marino A, Aquino R, D’Angelo V, Germano MP, De Pasquale R dan Pizza C. 2000. Antimicrobial activityof Mitracarpus
scaber extract and isolated constituents. Letter Appl Microb 30: 105 -108.
Brooke P, Harris DJ dan Longmore RB. 1996. Isolatio n of minor lupin alkaloids. A simple procedure for the isolation of angustifoline from lupinus
angustifollius (Cv. Fest) seeds, with application to other lupin alkaloids. J Agric Food Chem 44(8):2129-2133.
Chakraborty A, Saha C, Podder G, Chowdhury BK, dan Bhattacharyya. 1995. Carbazole alkaloid with antimicrobial activity from Clausena heptaphylla.
Phytochemistry 38(3): 787-789.
Campo JD, Amiot MJ dan Christophe NT. 2000. Antimicrobial effect of rosemary extracts. J Food Prot 10: 1359-1368
Cano RJ dan Colome JS. 1986. Microbiology. West Publishing Company. New York.
Carson CF dan Riley TV. 1995. Antimicrobial activity of the major components of the essential oil of Melaleuca alternifolia . J Appl Bacteriol 78: 264-269.
Davidson PM dan Brannen AL. 1993. Antimicrobials in Food. Marcel Dekker. New York.
Farag RS, Daw ZY, Hewedl FM dan El-Baroty GSA. 1989. Antimicrobial activity of some Egyption spice essential oils. J Food Prot 52(9):665-667.
Fardiaz S. 1992. Mikrobiologi Pengolahan Pangan Lanjut. Bogor: PAU Pangan dan Gizi Institut Pertanian Bogor.
Friedman M, Henika PR, Levin CE dan Mandreil RE. 2004a. Antibacterial activities of plant essential oils and their components against Escherichia
coli O157:H7 and Salmonella enteritica in apple juice. J Agric Food Chem
52: 6042-6048.
Friedman M, Buick R dan Elliott CT. 2004b. Antibacterial activities of naturally occuring coumpounds against antibiotic -resistant Bacillus cereus vegetative cells and spora, Escherichia coli, and Staphylococcus aureus. J
Food Prot 67:1774-1778
Garriga M, Hugas M, Aymerich T dan Monfort JM. 1993. Bacteriocinogenic activity of Lactobacilli from fermentor sausages. J Appl Bacterio 75: 142-148.
Harbone JB. 1996. Metode Fitokimia. Penuntun cara modern menganalisis
tumbuhan. Padmawinata K, Sudiro I, Penerjemah. Bandung: Penerbit ITB.
Harrigan WF. 1998. Laboratory Methods in Food Microbiology. 3rd edition. San Diego.
Hiserodt RD, Franzblau SG dan Rosen RT. 1998. Isolation of 6-,8- and 10-Gingerol from ginger rhizome by HPLC and preliminary evaluation of inhibition of Mycobacterium avium and ycobacterium tuberculosis. J
Agric Food Chem 3: 477-480.
Holstege DM, Seiber JN, dan Galey FD. 1995. Rapid multiresidue screen for alkaloids in plant material and biological samples. J Agric Food Chem 43(3): 691-699.
Hostetmann K, Wolfender JL dan Rodrigue ZS. 1997. Rapid detection and subsequent isolation of bioactive constituents of crude plant extracts.
Planta Med 63:2-10
Houghton PJ dan Raman A. 1998. Laboratory Handbook for the Fractination of
Natural Extracts. Thomson Science, London.
Jawetz E, Melnick J dan Adelberg E. 1996. Medical Microbiology. Appleton & Lange. San Fransisco.
Kanazawa A, Ikeda T dan Endo T.1995. A novel approach to mode of action of cationic biocides morphological effect on antibacterial activity. J Appl
Bacteriol 78: 55 -60.
Kim JM, Marshal MR, Cornell JA, Boston JF dan Wei CI. 1995. Antibacterial activity of carcacrol, citral and geraniols againts Salmonella typhimurium in culture medium and fish cubes. J Food Sci 60 (6): 1365-1368.
Kubo A, Lunde CS, Kubo I. 1995. Antimicrobial activity of the olive oil flavor compounds. J Agric Food Chem. 40(6):999-1003.
Lewis K, Salyers AA, Taber HW, Wax RG. 2004. Bacterial Resistance to
Antimicrobials. Marcel Dekker, Inc.
Lin CM, Preston JF III dan Wei CI. 2000. Antibacterial mechanism of allyl isothiocyanate. J Food Prot Vol 63 (6): 727-734.
Madigan MT, Martinko JM, Parker J. 2003. Brock Biology of Microorganisms. Tenth Edition. Southern Illinois University Carbondale.
Murhad i. 2002. Isolasi dan karakterisasi komponen -komponen antibakteri dari biji Atung (Parinarium glaberimum Hassk). [disertasi]. Bogor. Program Pascasarjana Institut Pertanian Bogor.
Naidu AS dan Davidson PM. 2000. Phyto-phenols. Di dalam Naidu AS, editor.
Natural Food Antimicrobial Systems. New York: CRC Press.
Nishina AK, KinaichiH, Uchibori T, Seino H dan Osawa T. 1991. 2,6-dimethoxy-p-benzequinone as an antimicrobial substance in the bark of Phyllostachys
heterocyclavar. Pubscens a species of thick-stemmed bamboo. J Agric Food Chem 39:266-269.
Nuraida L, Andarwulan N dan Kristikasari E. 1999. Aktivitas antimikroba biji picung (Pangium edule Reinw.) segar dan terfermentasi terhadap bakteri patogen dan perusak makanan. J Ilmu dan Tek Pangan 4 (2): 18-26. Nychas GJE.1995. Natural antimicrobials from plants. Di Dalam: Gould GW.
(Eds). New Methods of Food Preservation. Blackie Academic and Profesional. London.
Nychas GJE dan Tassou CC. 2000. Traditional preservatives-oils and spices. Di dalam: Robinson RK, Batt CA, Patel PD, editor. Encyclopedia of Food
Microbiology. Volume 1. :ondon: Academic Press.
Oyen LPA dan Nguyen XD. 1999. Plant resources of South-East Asia No. 19. Essential oil plants. Bogor: Prosea Bogor Indonesia.
Ozean M, dan Erkmen O. 2001. Antimicrobial activityof the essential oils of Turkish plant spices. Eur Food Res Technol 212: 658-660.
Purseglove JW.1981. Spices. Volume II. New York: Longman Inc.
Puupponen -Pimia R, Nohyyenk L. Meier C, Kahkonen M, Heihonen M. 2001. Antimicrobials properties of phenolic compound from berries. J Appl
Microbiol 90:494-507.
Ramsewak RS, Nair MG, Strasburg GM, Dewit DL dan Nitiss JL. 1999. Biologically active carbazole alkaloids from Murraya koeniglii. J. Agric
Food Chem 47 (2) 444-447.
Sakanaka S, Kim M, Tan iguchi M, Yamamoto T. 1989. Antibacterial substances in Japanese green tea extract against Streptococcus mutan , a cariogenic bacterium. Agric Biol Chem 53(9):2307 -2311.
Setiabudy R. dan Gan VHS. 1995. Antimikroba. Di Dalam: Ganiswarna SG, Setiabudy R, Suyatna FD, Purwantyastuti dan Nafrialdi. Farmakologi dan
Terapi. Edisi 4. Gaya Baru. Jakarta.
Shahidi F dan Naczk M. 1995. Foods Phenolics. Technomic Co., Inc. Lancaster. Ultee A, Gorris LGM dan Smid EJ. 1998. Bacterial activity of carvacrol towards
the fo od-borne pathogen Bacillus cereus. J App Microbiol 85: 213-218. Ultee A, Slump RA, Steging G dan Smid EJ. 2000. Antimicrobial activity of
carvacrol toward Bacillus cereus on rice. J Food Prot Vol 63. No.5:620-624.
Ultee A, Slump RA, Steging G dan Smid EJ. 2000. Antimicrobial activity of carvacrol toward Bacillus cereus on rice. J Food Prot Vol 63. No.5:620-624.Wendaken CN dan Sakaguchi M. 1995. Inhibition of amino acid decarboxylase activity of Enterobacter aerogenes by active components in spices. J Food Prot 58(3): 280.
Yasni S. 2001. Aktivitas antimikroba minyak atsiri buah andaliman (Zanthoxyllum acanthopodium DC) dan antarasa (Litsea cubeba) terhadap bakteri dan kapang serta profil deskriptif komponen aktif penyusunnya.Di dalam: Nuraida L, Dewanti-Hariyadi R, editor. Pangan Tradisional: Basis
Bagi Industri Pangan Fungsional dan Suplemen. Pusat Kajian Makanan
Tradisional IPB. Bogor. Hal 130-138.
Zuhud EAM, Rahayu WP, Wijaya CH dan Sari PP. 2001. Aktivitas antimikroba ekstrak kedawung (Parkia roxburghii G. Don) terhadap bakteri patogen.