TINJAUAN PUSTAKA
Sejarah Tanaman Kelapa Sawit
Tanaman kelapa sawit (Elaeis guineensis Jacq.) pertama kali diperkenalkan di Indonesia oleh pemerintah Belanda pada tahun 1848. Pada waktu itu ada 4 bibit kelapa sawit yang dibawa dari Mauritius dan Amsterdam ditanam di kebun Raya Bogor. Pada tahun 1911, kelapa sawit mulai diusahakan dan dibudidayakan secara komersial. Perintis usaha perkebunan kelapa sawit di Indonesia adalah Adrien Hallet yang berasal dari Belgia. Perkembangan budidaya kelapa sawit dilanjutkan oleh K.Schadt yang mempelopori usaha perkebunan kelapa sawit di Indonesia. Perkebunan kelapa sawit pertama berlokasi di Pantai Timur Sumatera Utara (Deli) dan Aceh dengan luas areal 5.123 ha (Lubis, 1992).
Pada masa pendudukan Jepang, perkebunan kelapa sawit menjadi terlantar sehingga ekspor CPO sempat terhenti. Banyak kebun kelapa sawit yang diganti tanamannya menjadi tanaman pangan, sehingga banyak pabrik kelapa sawit yang menjadi tutup. Setelah merdeka, perkebunan kelapa sawit mulai bangkit dan berkembang lagi. Pada tahun 1957, luas kebun kelapa sawit berkembang menjadi 103.000 ha dengan produksi CPO 160.000 ton (Lubis, 1992).
Pada masa pemerintahan Orde Baru, pembangunan perkebunan ditujukan untuk menciptakan kesempatan kerja, meningkatkan kesejahteraan masyarakat dan sektor penghasil devisa Negara. Lahan perkebunan kelapa sawit Indonesia berkembang pesat terutama perkebunan rakyat dengan dukungan Pemerintah yang melaksanakan program Perusahaan Inti Rakyat Perkebunan (PIR – BUN) (Lubis, 1992). Selain itu, Perusahaan Besar Negara atau PTPN juga memperluas lahan kelapa sawit. Wilayah kerja PTPN tersebar mulai dari NAD sampai dengan Sulawesi. Mulai tahun 2007, perkebunan kelapa sawit mulai tumbuh di Papua. Perusahaan Swasta yang bergerak di bidang kelapa sawit antara lain adalah Socfindo, Lonsum, dan Tania Jaya. Dengan berkembangnya industri hasil olahan minyak kelapa sawit, banyak Perusahaan Swasta baru yang bergerak di sektor perkebunan kelapa sawit, terutaman perusahaan besar
seperti Perusahaan Bayer, PT Sinar Mas Group, PT Sampoerna Agro Tbk, PT Wilmar dan PT Astra Agro Lestari Tbk (Administrator, 2000a).
Berdasarkan data statistik Ditjenbun (Administrator, 2008b), luas areal kebun kelapa sawit nasional pada tahun 2003 adalah 5.283.600 ha dan meningkat menjadi 6.324.300 ha pada tahun 2007 yang meliputi 22 provinsi. Secara rinci, luas areal kelapa sawit nasional dapat dibagi menjadi 3 kelompok, yaitu: (1) Perkebunan Rakyat yang mencapai 1.854.400 ha pada tahun 2003 dan meningkat menjadi 3.058.800 ha pada tahun 2007. (2) Perusahaan Besar Negara yang luasnya 662.800 ha pada tahun 2003 dan meningkat menjadi 694.300 ha pada tahun 2007. (3) Perusahaan Besar Swasta dimana luasnya paling besar yaitu 2.766.400 ha pada tahun 2003 dan pada tahun 2007 meningkat menjadi 3.058.800 ha.
Botani Tanaman Kelapa Sawit
Kelapa sawit termasuk dalam Divisio : Magnoliophyta, Kelas : Monocotyledoneae, Ordo : Arecales, Famili : Arecaceae, Sub famili : Palminae, Genus : Elaeis dan Species : Elaeis guineensis Jacq. Untuk membedakan varietas kelapa sawit, terdapat dua kriteria yang digunakan yaitu berdasar tebal tipisnya cangkang (endocarp) dan warna buah. Berdasar tebal tipis cangkang, kelapa sawit digolongkan menjadi 3 varietas, yaitu: Dura, Pisifera dan Tenera. Sedangkan berdasar warna buah, kelapa sawit dibagi menjadi 3 varietas, yaitu: Nigrescens, Virescens dan Albescens (Lubis, 1992).
Kelapa sawit memiliki sifat-sifat bagian vegetatif dan bagian generatif yang khas. Bagian vegetatif kelapa sawit yang penting antara lain akar, batang dan daun. Akar kelapa sawit terdiri dari 4 macam, yaitu: akar primer, sekunder, tersier dan kuarter. Batang kelapa sawit tumbuh lurus ke atas dengan diameter sekitar 40 – 60 cm. Tinggi batang dapat mencapai 30 m. Daun kelapa sawit bersirip genap dan bertulang sejajar. Tiap pelepah daun terdapat anak daun yang jumlahnya dapat mencapai 160 pasang. Duduk daun (filotaksis) pada batang tersusun melingkar membentuk spiral (Corley & Gray, 1976; Lubis, 1992).
Bunga kelapa sawit termasuk tipe berumah satu (Monoeceous) dengan tandan bunga terletak di ketiak daun. Primordia bunga terbentuk kurang-lebih 34 bulan sebelum bunga matang yang siap melakukan penyerbukan. Diferensiasi bunga jantan dan betina terjadi pada 17-25 bulan sebelum antesis. Bunga jantan maupun betina biasanya terbuka selama dua hari dan pada musim hujan dapat mencapai 4 hari. Tepung sari dapat menyerbuk selama 2-3 hari, namun semakin lama daya hidupnya semakin menurun. Morfologi bunga kelapa sawit jantan dan betina ditunjukkan pada Gambar 1.
Gambar 1. Bunga tanaman kelapa sawit, a). Infloresensia bunga betina saat antesis, b). Infloresensia bunga betina yang telah dewasa dengan putik yang terlihat jelas, c). Infloressensia bunga jantan saat antesis, d). Infloresensia bunga jantan yang telah dewasa (Adam et al., 2005).
Tanaman kelapa sawit di lapangan mulai berbunga pada umur 12-14 bulan yang berasal dari bibit umur 11 bulan, namun buah yang dihasilkan belum ekonomis untuk dipanen. Mulai umur 2,5 tahun, tanaman kelapa sawit berproduksi secara stabil dan konsisten bila tidak terjadi gangguan lingkungan seperti perubahan iklim dan serangan hama. Oleh karena itu, pemanenan buah kelapa sawit dilakukan mulai umur 2,5 tahun (Lubis, 1992).
a b
c d
Proses pembentukan buah dimulai dari proses polinasi dan fertilisasi tepung sari ke putik sampai perkembangan buah menjadi masak. Waktu yang diperlukan selama proses pembentukan tidak sama tergantung iklim setempat. Buah yang masih muda berwarna ungu, kemudian secara berangsur berubah menjadi merah-kekuningan seiring dengan tingkat kematangan. Proses pembentukan minyak berlangsung selama 24 hari sampai buah mencapai tingkat masak. Masaknya buah dalam satu tandan berangsur-angsur mulai dari bagian atas menuju ke arah bawah atau bagian pangkal (Corley & Gray, 1976; Lubis, 1992).
Buah yang masak akan terbentuk 5-6 bulan setelah antesis tergantung iklim setempat. Tiap buah panjangnya 2-5 cm dan beratnya mencapai 30 g atau lebih. Buah kelapa sawit termasuk tipe buah batu yang tersusun atas: kulit buah (exocarp), daging buah (pulp, endocarp), cangkang (tempurung) dan inti/biji (kernel, endosperm). Exocarp dan mesocarp disebut sebagai pericarp yang merupakan bagian yang mengandung minyak terbesar (24%) sedangkan kandungan minyak dalam kernel hanya 4% (Corley & Gray, 1976; Lubis, 1992).
Kultur Jaringan Tanaman Kelapa Sawit
Penelitian mengenai kultur jaringan kelapa sawit (Elaeis guineensis Jacq.) telah dimulai lebih dari tiga dasawarsa yang lalu oleh ORSTOM-IRHO/CIRAD di Perancis (Rabechault et al., 1972) dan Unilever di Inggris (Smith & Thomas, 1973) melalui embriogenesis somatik, untuk menghasilkan bibit kelapa sawit secara massal. Sistem kultur yang dilakukan pada awalnya adalah kultur dalam medium padat. Dalam perkembangannya, telah dilakukan kultur dalam medium cair (Sumaryono et al., 1994; Ginting & Fatmawati, 1997) dan medium cair sistem perendaman sesaat (TIS, temporary immersion system) (Tahardi, 1998b; Etienne & Berthouly, 2002).
Sistem kultur cair dikembangkan terutama dengan tujuan otomatisasi dan scale-up produksi planlet serta meningkatkan pertumbuhan dan keseragaman kultur (Touchet et al., 1991; Sumaryono et al., 1994; Ginting & Fatmawati, 1997; Tahardi, 1998a, 1999). Sedangkan sistem perendaman sesaat (PS) ditujukan terutama untuk menghindari terjadinya abnormalitas. Sistem ini telah diaplikasikan untuk perbanyakan in vitro secara massal pada berbagai jenis tanaman (Etienne & Berthouly, 2002). Hasil
penelitian pada kelapa sawit (Tahardi, 1998b) menunjukkan bahwa frekuensi produksi embrio somatik dari kalus nodular sangat tinggi, di samping itu sinkronisasi perkembangan embrio juga lebih baik sehingga diperoleh embrio somatik yang lebih seragam.
Beberapa peneliti melaporkan bahwa produktivitas tanaman kelapa sawit klonal hasil kultur jaringan terbukti lebih tinggi dibandingkan dengan tanaman asal benih. Berdasarkan satuan tanaman, produktifitas tersebut meningkat 23 sampai 39% (Subronto et al., 1995; Soh et al., 2006). Namun terjadinya abnormalitas organ reproduktif dari populasi bibit asal kultur jaringan menyebabkan penurunan produksi. Tingkat abnormalitas bibit asal kultur jaringan dilaporkan rata-rata 5-10% (Jaligot et al., 2000), 10-40% (Subronto et al., 1995) bahkan sampai 100% (Soh et al., 2006). Hal ini tentu sangat merugikan karena akan menurunkan produktivitas kebun. Akibatnya, komersialisasi bibit kelapa sawit asal kultur jaringan menjadi terhambat
Abnormalitas Organ Reproduktif Tanaman Kelapa Sawit
Permasalahan yang timbul pada tanaman kelapa sawit asal kultur jaringan adalah terjadinya abnormalitas organ reproduktif. Beberapa laporan menunjukkan adanya beberapa jenis abnormalitas yang terjadi, yaitu bunga steril (jantan semua), gugur buah dan buah bersayap atau bermantel (mantled). Dibandingkan dengan tanaman kelapa sawit normal, secara fenotipik tanaman abnormal ini memperlihatkan perbedaan yang sangat mencolok. Kadang-kadang dalam satu pohon tandan bunga yang terbentuk sebagian besar berupa bunga jantan atau jantan semua. Fenotipe abnormal yang paling banyak dijumpai adalah buah mantled. Abnormalitas jenis ini ditandai dengan terbentuknya buah yang tampak seperti terbelah-belah atau berlapis-lapis sehingga terlihat seperti bermantel atau bersayap yang berasal dari organ bunga betina. Hetharie (2008) menggolongkan tingkat abnormalitas buah bermantel kelapa sawit menjadi 3 kelompok yaitu: (1) Abnormalitas ringan (AbR), dicirikan dengan terbentuknya karpel tambahan dimana batasan antara karpel tambahan tersebut hanya terlihat pada bagian ujung buah, mesokarp berdaging, dan mempunyai biji. (2) Abnormalitas berat (AbB), dicirikan dengan adanya karpel tambahan dari bagian ujung ke bagian tengah
buah yang terpisah dengan karpel utama, batasan antara karpel tambahan sangat jelas dari ujung ke bagian tengah buah dan selanjutnya manyatu dengan karpel utama, mesokarp berdaging dan mempunyuai biji. (3) Abnormalitas sangat berat (AbSB), dicirikan dengan terbentuknya karpel tambahan yang terpisah dari karpel utama mulai dari ujung sampai sepertiga dari pangkal buah demikian juga antar karpel tambahan serta tidak mempunyai biji.
Larkin & Scowcroft (1981) mengemukakan bahwa, abnormalitas pada tanaman kelapa sawit hasil kultur jaringan merupakan salah satu bentuk variasi somaklonal, yaitu keragaman genetik pada tanaman yang diregenerasi dari kultur jaringan. Kadangkala tanaman abnormal dapat pulih menjadi tanaman normal seiring dengan waktu, karena itu kelainan ini bersifat epigenetik, akibat terjadinya perubahan pada ekspresi gen (Tregear et al., 2002). Abnormalitas tersebut mungkin disebabkan oleh zat pengatur tumbuh (Jones, 1991; Paranjothy et al., 1993; Eeuwens et al., 2002a), kondisi kalus (Duran-Gasselin et al., 1993), lama subkultur dan umur kalus (Paranjothy et al., 1993; Eeuwens et al., 2002b), serta tekanan seleksi, jenis eksplan, level ploidi dan kecepatan proliferasi kalus (Karp, 1995). Hasil penelitian Eeuwens et al. (2002a) menunjukkan bahwa terbentuknya buah mantel diakibatkan penggunaan hormon tumbuh auksin (2,4-D dan NAA) dan sitokinin (kinetin dan BAP) dalam medium kultur. Semakin tinggi perbandingan antara sitokinin dengan auksin maka semakin tinggi tingkat abnormalitas.
Penelitian abnormalitas kelapa sawit di tingkat molekuler telah dimulai lebih dari satu dasawarsa. Jaligot et al. (2000) dan Matthes et al. (2001) melaporkan bahwa terbentuknya variasi somaklonal pada tanaman kelapa sawit disebabkan karena terjadinya perubahan pola metilasi DNA. Hasil penelitian lain menyebutkan bahwa terbentuknya buah mantel ini akibat proses feminisasi androecium bunga jantan maupun betina menjadi struktur menyerupai karpel (karpeloid) dan petal kedua bunga tersebut berkembang menjadi struktur menyerupai sepal (sepaloid) (Corley et al., 1986). Pada bunga jantan abnormal, stamen berkembang sebagai struktur karpeloid sedangkan bunga betina abnormal dan staminodes (vestigial stamen) berkembang sebagai struktur pseudocarpel. Dengan demikian, abnormalitas kelapa sawit hasil kultur jaringan mirip dengan mutan tipe B pada tanaman A. thaliana maupun A. majus (Tregear et al., 2002).
Gen-gen Pembungaan Tanaman
LFY dan AG sebagai Gen Penting untuk Pembungaan Tanaman. Secara fisiologi molekuler tanaman, diketahui 4 perbedaan tipe meristem yang menginisiasi perkembangan organ bunga. Dalam proses perkembangannya, organ bunga tersebut membentuk susunan lingkaran (ring) dari luar ke dalam dengan urutan sebagai berikut : sepal, petal, stamen dan karpel (Coen and Carpenter, 1993). Hasil penelitian pada tanaman Arabidopsis thaliana menunjukkan bahwa perkembangan identitas organ bunga dipengaruhi oleh ekspresi beberapa gen pembungaan yaitu APETALA1 (AP1), APETALA2 (AP2), APETALA3 (AP3), PISTILATA (PI) dan AGAMOUS (AG) (Taiz dan Zeiger, 2002). Berdasar aktifitas gen-gen tersebut, dikelompokkan menjadi 3 tipe atau kelompok gen yang berpengaruh dalam perkembangan identitas organ bunga, yang kemudian dikenal sebagai model ABC seperti terlihat pada Gambar 2 (Taiz dan Zeiger, 2002). Aktifitas tipe A disandikan oleh gen AP1 dan AP2 yang mengontrol perkembangan identitas organ pada lingkaran pertama dan kedua yaitu pembentukan sepal dan petal. Tipe B disandikan oleh gen AP3 dan PI yang mengontrol perkembangan identitas organ pada lingkaran kedua dan ketiga yaitu pembentukan petala dan stamen. Sedangkan tipe C disandikan oleh gen AG yang mengontrol perkembangan identitas organ pada lingkaran ketiga dan keempat yaitu pembentukan stamen dan karpel (Pineiro & Coupland, 1998; Taiz & Zeiger, 2002). Dari hasil penelitian tentang hubungan antara masing-masing gen ABC tersebut, diperoleh tiga bentuk fenotipik bunga: (1) Apabila gen pembungaan yang bekerja hanya gen A dan B saja, maka bunga yang terbentuk hanya memiliki sepal dan petal sehingga bunga tersebut tidak mempunyai kelamin baik jantan maupun betina. (2) Apabila gen yang bekerja hanya gen A dan C, maka bunga yang terbentuk hanya memiliki sepal dan karpel saja sehingga kehilangan petal dan stamen. (3) Sedangkan bila hanya gen B dan C saja yang bekerja, maka bunga yang terbentuk hanya memiliki stamen dan karpel saja tidak memiliki sepal dan petal (Coen & Meyerowitz, 1991; Taiz & Zeiger, 2002).
Dengan semakin pesatnya kemajuan penelitian gen pembungaan pada tanaman, maka model gen ABC berkembang menjadi model gen ABCDE (Pelaz et al., 2000; Pinyopich et al., 2003). Model tersebut melengkapi fungsi dari model gen ABC dengan
penambahan gen D yang berfungsi untuk menentukan perkembangan bakal buah (ovule) dan gen E yang berfungsi untuk mendorong perkembangan seluruh organ bunga pada tanaman.
Gambar 2. Model gen ABC untuk proses pembentukan organ bunga (Taiz & Zeiger, 2002).
Lebih jauh Ordge et al. (2005) melaporkan bahwa pada Arabidopsis thaliana, gen LEAFY (LFY) berperan sebagai pusat untuk mengintegrasikan sinyal dan meregulasi sebagian pembungaan melalui interaksi antara TERMINAL FLOWER1 (TFL1) dan AGAMOUS (AG). Homologi antara LFY, TFL1 dan AG mempunyai tingkat konservasi yang tinggi pada tingkat sekuen dan menunjukkan fungsi homologi saat terekspresi secara ektopik pada tanaman Arabidopsis transgenik. Ekspresi gen LFY dapat ditriger atau dipacu oleh gen AGAMOUS-LIKE20 (AGL20) kemudian LFY mendorong ekspresi gen AP1. Pada Arabidopsis, gen LFY dan AP1 mempunyai hubungan timbal balik yang saling berpengaruh, sehingga ekspresi gen AP1 juga menstimulasi ekspresi gen LFY atau sebaliknya (Simon et al., 1996). Di samping itu, aktifitas gen LFY dan LMI1 secara bersama-sama mempengaruhi ekspresi gen CAL (CAULIFLOWER) yang
berperan penentu identitas dan formasi meristem bunga (Kempin et al., 1995; Saddic et al., 2006).
Gen AG adalah keluarga gen MADS-Box yang diperlukan dalam pembentukan identitas organ bunga. Gen ini berperan dalam pembentukan dan perkembangan stamen (kepala dan benang sari) dan karpela (pistil/putik). Tingkat ekspresi gen AG dapat ditekan oleh gen lain seperti BELLRINGER (BELL) sehingga pengaruh gen AG dalam pembentukan meristem bunga dan infloresensia menjadi terhambat (Bao et al., 2004). Mizukami & Ma (1997), melaporkan adanya fungsi ektopik gen AG yang mempengaruhi proses transisi SAM (shoot apical meristem) dari pertumbuhan vegetatif ke generatif.
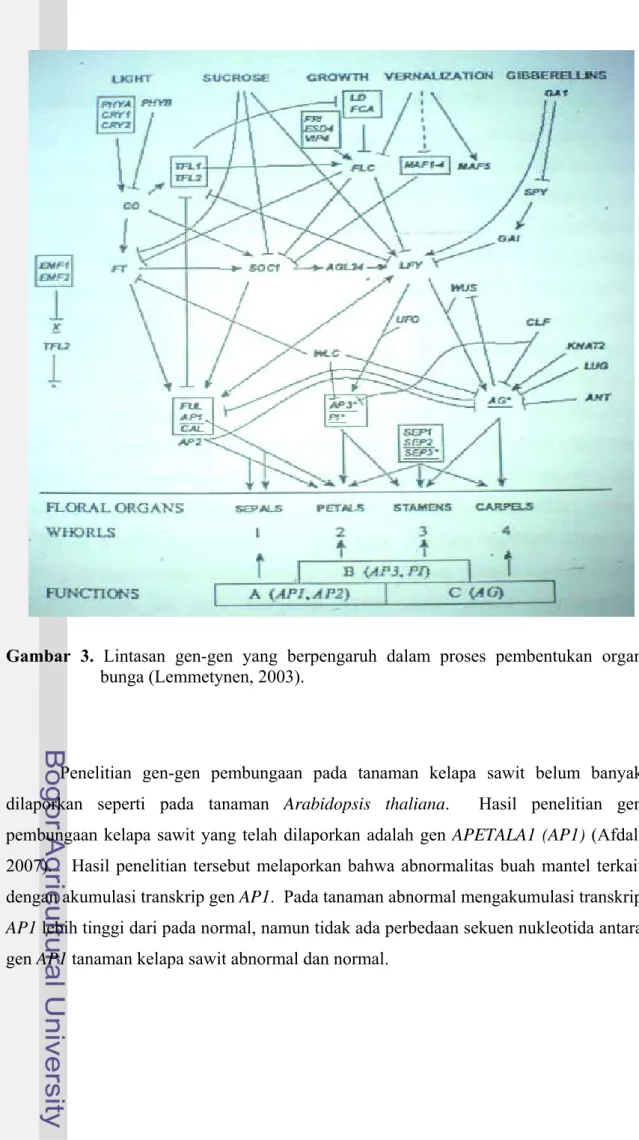
Dalam proses pembentukan organ bunga, terdapat beberapa gen yang saling berinteraksi sehingga berpengaruh pada tingkat ekspresi. Lintasan atau pathway gen-gen pembungaan dan interaksinya dengan gen-gen-gen-gen lain terlihat pada Gambar 3 (Lemmetynen, 2003). Berdasar lintasan tersebut, gen LFY secara langsung dipengaruhi GA1 dan GA24. Selanjutnya ekspresi gen LFY menginduksi ekspresi gen AP3 dan PI yang akan menginduksi pembentukan organ petal dan stamen. Disamping itu, gen LFY juga menstimulasi ekspresi gen AG yang akan menginduksi organ stamen dan karpel (organ reproduktif tanaman).
Gambar 3. Lintasan gen-gen yang berpengaruh dalam proses pembentukan organ bunga (Lemmetynen, 2003).
Penelitian gen-gen pembungaan pada tanaman kelapa sawit belum banyak dilaporkan seperti pada tanaman Arabidopsis thaliana. Hasil penelitian gen pembungaan kelapa sawit yang telah dilaporkan adalah gen APETALA1 (AP1) (Afdal, 2007). Hasil penelitian tersebut melaporkan bahwa abnormalitas buah mantel terkait dengan akumulasi transkrip gen AP1. Pada tanaman abnormal mengakumulasi transkrip AP1 lebih tinggi dari pada normal, namun tidak ada perbedaan sekuen nukleotida antara gen AP1 tanaman kelapa sawit abnormal dan normal.
Bioinformatika
Bioinformatika adalah penelitian, pengembangan dan aplikasi teknik komputasional beserta pendekatannya yang bertujuan untuk memperkaya penggunaan data-data cabang ilmu biologi, kesehatan, dan kedokteran. Ruang lingkup dari penggunaan data ini antara lain pencarian, penyimpanan, pengorganisasian, pengarsipan, analisis dan penyajian data (Nih, 2000). Beberapa software dan data untuk pengolahan data bioinformatika telah tersedia di berbagai situs internet baik secara online seperti: www.ncbi.nlm.nih.gov; www.ebi.ac.uk; www.ddbj.nig.ac.jp dan Primer3 maupun offline seperti: program BioEdit, NJPlot, DNA Star dan Treecon.
Penyejajaran sekuen (Sequence Alignment). Penyejajaran sekuen ditujukan untuk menentukan apakah dua buah atau lebih sekuen mempunyai kemiripan dengan mempertimbangkan pengaruh secara homologi. Kemiripan adalah pengukuran kesamaan gen secara kuantitaif dengan menggunakan metode-metode yang sesuai. Sedangkan homologi adalah kesimpulan bahwa dua atau lebih sekuen mempunyai hubungan evolusi (Baxevanis, 2001).
BLAST (Basic Local Alignment Search Tool) merupakan salah satu data base dan program yang sering digunakan untuk penyejajaran sekuen. Program ini dirancang untuk mengeksplorasi semua database sekuen yang diminta baik berupa nukleotida maupun protein. Program BLAST juga dapat digunakan untuk mendeteksi hubungan antara sekuen yang hanya berbagi daerah tertentu yang memiliki kesamaan (Biotrain, 2003). BLAST menggunakan analisis statistik untuk menghasilkan skor (bits) dan E-value. Skor menunjukkan tingkat keakuratan nilai penjajaran (alignment) sekuen nukleotida atau protein yang tidak diketahui dengan sekuen nukleotida atau protein yang terdapat dalam database. Semakin tinggi skor maka semakin tinggi tingkat homologi kedua sekuen tersebut dan begitu pula sebaliknya. Skor dibawah 50 menunjukkan bahwa hasil penjajaran kedua sekuen tersebut tidak dapat dipercaya. E-value menunjukkan nilai statistik yang signifikan dari penjajaran sekuen nukleotida atau protein yang tidak diketahui dengan sekuen nukleotida atau protein yang terdapat dalam database. Semakin rendah E-value maka semakin tinggi tingkat homologi kedua sekuen dan sebaliknya. Untuk memastikan adanya homologi, E-value harus lebih rendah dari
e-04 (Claverie & Notredame, 2003). Variasi Program BLAST yang sering digunakan antara lain :
BLASTn : membandingkan sekuen nukleotida dengan sekuen nukleotida lainnya dalam database.
BLASTx : membandingkan sekuen nukleotida yang ditranslasi menjadi protein dengan protein dalam database.
BLASTp : membandingkan sekuen protein dengan protein dalam database.
Selain penyejajaran sekuen, masih banyak tersedia fasilitas lain yang terdapat pada beberapa program tersebut untuk analisis bioinformatika lainnya. Beberapa program tersebut yang sering digunakan antara lain :
Vector Screen : membersihkan kontaminan nukleotida dari vektor hasil sekuensing. BLAST2seq : membandingkan atau menyejajarakan nukelotida antara dua sekuen. Restriction map : identifikasi peta restriksi beserta daftar enzim restriksi yang dapat
digunakan pada suatu sekuen.
ClustalW : menyejajarkan susunan nukleotida pada beberapa sekuen serta tingkat similaritas atau tingkat kemiripan.
Phylogenetic tree: analisis tingkat kekerabatan spesies yang ditunjukkan dengan diagram pohon.
Reverse complement : membalik susunan nukleotida pada suatu sekuen. Find ORF : mencari titik ORF (open reading frame) pada suatu sekuen. (Claverie & Notredame, 2003).