ALIH BAHASA
Karakterisasi Molekuler dan Morfologi
Populasi Luar Biasa Meloidogyne arenaria dari Pohon Traveler, Ravenala madagascariensis
Nurul Dwi Handayani, SP. M.Si NIP. 19750621 200003 2 001
POPT Pertama/III-c
Balai Uji Terap Teknik dan Metode Karantina Pertanian Badan Karantina Pertanian
1 Journal of Nematology 40(3): 179-189, 2008. © The Society of Nematologist 2008.
Karakterisasi Molekuler dan Morfologi Populasi Luar biasa Meloidogyne arenaria dari Pohon Traveler, Ravenala madagascariensis
AM. Skantar, LK. Carta, ZA. Handoo
Ringkasan: Variasi luar biasa dari Meloidogyne arenaria ditemukan pada akar
pohon traveler (Ravenala madagascariensis) dimaksudkan untuk pameran di arboretum publik Pennsylvania. Populasi membangkitkan rasa ingin tahu karena kurangnya puru yang tampak pada akar tanaman yang terinfeksi, dan bagian vulva betina biasanya dikelilingi oleh kantung telur. Morfometri paling sesuai yang dilaporkan dalam kisaran M. arenaria, dengan bentuk mosaik yang sama dengan
M. platani atau Meloidogyne spp. dari daerah tropikal lain. Karakterisasi
molekuler termasuk analisis empat lokus. Sikuen mitokondria, memanjang dari sitokrom oksidase II (COII) ke gen 16S (IRNA), hampir sama dengan populasi M.
arenaria yang lain dan terkait erat dengan sikuen dari M. morocciensis dan M. thailandica. Pemanjangan bagian 28S D2-D3 paling mirip dengan M. arenaria, M. incognita, M. paranaensis, dan IGS-2 paling berhubungan dengan M. thailandica, M. arenaria dan M. incognita. Analisis terhadap sebagian sikuen
genom Hsp90 mengungkapkan bahwa yang paling mirip adalah M. arenaria, M.
thailandica dan haplotype Hsp90 dari M. floridensis, dan gabungan sikuen terdiri
dari EST M. arenaria. Tidak ada cara morfologi atau molekuler yang dapat dengan jelas membedakan bentuk populasi dari spesies baru, dan ketika dianggap sebagai keseluruhan, bukti ini menunjukkan identifikasi sebagai M. arenaria.
Kata kunci: puru, Hsp90, intergenic spacer, DNA mitokondria, biologi molekuler, Ravenala madagascariensis, DNA ribosom, nematoda puru akar, taksonomi, pohon traveler, variasi.
Nematoda puru akar (Meloidogyne spp.) merupakan parasit tanaman yang mempunyai arti penting secara ekonomi, berdampak terhadap berbagai tanaman inang, dan sejauh ini sudah 95 spesies yang dideskripsikan. Pada bulan Juni 2007, nematoda puru akar (NPA) ditemukan pada akar pohon traveler (Ravenala
madagascariensis) yang dimaksudkan untuk pameran di sebuah taman umum di Pennsylvania. Tumbuh dalam suatu campuran tanah buatan, seperti pohon kelapa yang terserang diperoleh arboretum dari seorang petani di Florida. Keterangan yang tepat tentang asal pohon traveler tidak tersedia. Bagian akar yang terkena
2
penyakit dikirim untuk identifikasi melalui ekstensi ahli Delaware untuk Laboratorium Nematologi USDA di Beltsville, MD.
Perkembangan metode molekuler untuk identifikasi empat spesies utama NPA (M. incognita (Kofoid dan White, 1919) Chitwood, 1949; M. arenaria (Neal, 1889); M. javanica (Treub, 1885) Chitwood, 1949; dan M. hapla Chitwood, 1949) menjadi tujuan dari banyak sekali penelitian. Sementara analisis isozim terus menjadi cara yang efektif untuk membedakan banyak spesies NPA (Esbenshade dan Triantaphyllou, 1990; Carneiro, 2000), pengurutan sikuen DNA sangat mudah dan memberikan manfaat dalam menyediakan data yang berguna untuk analisis filogenetik. DNA penanda yang membantu identifikasi spesies Meloidogyne termasuk DNA ribosom sub unit kecil (SSU) 18S (Powers, 2004), sub unit besar (LSU) perluasan bagian 28S D2-D3 (Chen dkk., 2003; Palomares Ruis dkk., 2007), intergenic spacer (IGS) (blok dkk., 1997; Wishart dkk., 2002),
internal transcribed spacer (Powers dan Harris, 1993) dan DNA mitokondria
(Powers dan Harris, 1993; Stanton dkk., 1997; Blok dkk., 2002; Xu dkk., 2004; Jeyaprakash dkk, 2006). Random Amplified Polymorphic DNA (RAPD) (Cenis, 1993; Williamson dkk, 1997; Dong dkk, 2001; Cofcewicz dkk., 2004; Adam dkk., 2007) dan penanda sequeance characterized amplified regions (SCAR) (Zijlstra dkk., 2000; Randig dkk., 2002) juga telah mengembangkannya. Satu penelitian menggabungkan IGS PCR, penanda SCAR dan analisis RAPD termasuk kunci diagnostik untuk membedakan tujuh spesies NPA (Adam dkk., 2007). Baru-baru ini bagian sikuen dari gen yang mengkode protein Hsp90 digunakan untuk konfirmasi identitas pada populasi yang luar biasa agresif yaitu M. hapla yang menyerang kopi di Maui, HI (Handoo dkk., 2005b). Lokus ini juga menunjukkan potensi pembentukan hubungan filogenik diantara spesies nematoda parasit pada tanaman (Skantar dan Carta, 2004).
Penelitian ini menggambarkan bentuk morfologi dari populasi yang luar biasa pada M. arenaria yang ditemukan pada pohon traveler, menekankan perbedaan dari nilai yang diharapkan untuk spesies dan titik tumpang tindih dengan spesies terkait. Demikian juga, penelitian tentang hubungan NPA berasal dari daerah tropis lain, berdasarkan pada analisis sikuen mitokondria (interval dari
3
oksidasi sitokrom COII ke gen 16S rRNA, IRNA), perluasan bagian 28S D2-D3, daerah intergenic spacer ribosom (IGS) dan Hsp90.
Bahan dan Metode
Karakterisasi Morfologi : Tanah dan akar dari pohon traveler yang
terinfeksi dikirim ke Laboratorium Nematologi Departemen Pertanian Amerika Serikat (USDA) di Beltsville, MD, untuk identifikasi spesies. Massa telur disimpan dalam cawan petri dengan sedikit air pada suhu ruang. J2 diekstraksi dari tanah dengan saringan dan metode Baermann funnel. Setelah difiksasi selama semalam dalam 3% formaldehyde pada suhu ruang, nematoda betina, jantan dan J2 kemudian dibedah dari akar yang terinfeksi. J2 dan jantan difiksasi dalam 3% formaldehyde dan diproses dengan glycerin sesuai dengan metode formalin-glycerin (Hooper, 1970; Golden, 1990). Prosedur yang digunakan dalam mengukur dan menyiapkan spesimen pada dasarnya dari Golden dan Birchfield (1972), kecuali beberapa nematoda betina tetap dipotong dan diawetkan dalam larutan lactophenol. Pengukuran dari semua stadia nematoda dibuat dengan mikrometer okuler dengan ukuran dalam mikrometer, kecuali dinyatakan lain. Gambar diambil menggunakan Zeiss Ultraphot III (Carl Zeiss, Inc., Jena, MD, USA) dengan kamera Toshiba IKTU CCD (Toshiba Corp., Japan) atau Zeiss Axiophot 100 mikroskop Inverted (Zeiss, Inc., Thornwood, NY) dengan kamera Q Imaging RTV Micropublisher 5 CCD (Qimaging, Surrey, BC, Canada). Gambar tubuh sebagian digabung dengan program Adobe Photoshop CS (Adobe system Inc., San Jode, CA).
Karakterisasi Molekuler : satu ekor J2 yang terdapat pada populasi Ravenala dan yang lain (M. arenaria dari Maryland; M. floridensis, dari Florida)
dikumpulkan secara mekanik dalam 20 µl buffer ekstraksi menurut Thomas dkk. (1997) kemudian disimpan dalam tabung PCR pada suhu -80 ºC sampai saat akan digunakan. Meloidogyne thailandica dari Thailand terdiri dari sembilan ekor J2 dalam 50 µl, diperlakukan seperti di atas. Ekstrak disiapkan dengan cara menginkubasi tabung pada suhu 60 ºC selama 60 menit dilanjutkan pada suhu 90 ºC selama 15 menit untuk deaktifasi proteinase K dan disentrifus sebentar sebelum digunakan dalam PCR. Biasanya, masing-masing 2.5 µl ekstrak
4
nematoda digunakan per 25 µl reaksi PCR. Primer yang digunakan dalam penelitian ini terdapat pada Tabel 1. Reaksi juga berisi 1 unit Eppendorf MasterTaq (Brikmann, Westbury, NY) dan buffer disediakan oleh pabrik; semua komponen lainnya ditambahkan sesuai dengan panduan khusus untuk masing-masing gen. Ribosom hasil PCR diamplifikasi dari intergenic spacer (IGS-2) menurut Blok dkk. (2002) dan dari perluasan bagian 28S D2-D3, seperti yang dilakukan De Ley dkk. (2005). Bagian sikuen Hsp90 diamplifikasi sebelumnya menurut Skantar dan Carta (2004). Amplifikasi daerah mitokondria antara gen
COII dan IRNA merupakan modifikasi Powers dan Harris (1993) dan Tigano dkk.
(2005), dilanjutkan : 1 siklus pada suhu 94 ºC selama 2 menit, diikuti 45 siklus pada suhu 94 ºC selama 30 detik, 48 ºC selama 30 detik dan 68 ºC selama 2 menit, diakhiri dengan 1 siklus pada suhu 68 ºC selama 5 menit.
Hasil PCR divisualisasi dengan cahaya UV setelah diberi warna dengan ethidium bromide. DNA dikeluarkan dari gel dan dimurnikan dengan QIAquick Gel Extraction Kit (Qiagen, Valencia, CA). Hasil PCR dibersihkan dari sikuen lain secara langsung atau dikloning ke dalam vektor pCR2.1 dan dipindahkan kedalam sel Escherichia coli TOP10 menurut TOPO TA instruksi pada peralatan cloning (Invitrogen, Carlsbad, CA). DNA plasmid dipersiapkan dengan Qiagen miniprep kit (Qiagen, Valencia, CA) dan diolah dengan Eco RI untuk konfirmasi kebenaran identitas yang dimasukkan. Pengurutan sikuen DNA dilakukan di University of Maryland Center for Biosystems Research. Untuk setiap lokus, 2 atau 4 klon atau hasil PCR mewakili dua atau lebih individu yang dianalisis. Sikuen DNA baru diajukan ke GenBank dengan nomor aksesi EU364878-EU364890 dan FJ238508.
Tabel 1. Daftar Primer yang digunakan untuk analisis DNA
Lokus Primer Sikuen 5’ sampai 3’ Ref. PCR
Mitochondrial COII - 1RNA C2F3 1180 GGTCAATGTTCAGAAATTTGTGG TACCTTTGACCAATCACGCT
Power dan Harris, 1993 Tigano dkk., 2005 28S D2-D3 D2A D3B ACAAGTACCGTGAGGAAAGTT TCGGAAGGAACCAGCTACTA Ye dkk., 2007 IGS-2 194 195 TTAACTTGCCAGATTCGGACG TCTAATGAGCCGTACGC Blok dkk., 1997 Hsp90 U288 L1110 GAYACVGGVATYGGNATGACYAA TCRCARTTVTCCATGATRAAVAC
5
Sikuen DNA dirakit menggunakan Sequencer 4.7 (Genecodes, Ann Arbor, MI) dan dianalisis menggunakan megablast BLASTN optimasi program untuk sikuen yang sangat mirip (http:// www.ncbi.nlm.nih.gov/blast/Blast.cgi). Sikuen yang serupa diunduh dari GenBank, sesuai dengan ClustalW2 dengan parameter standar (Larkin dkk., 2007), dan jika diperlukan kesesuaian dipotong dalam GeneDoc (Nicholas dkk., 1997). Untuk setiap penyesuaian lokus, berdasarkan jarak neighbor-joining (NJ) metode ini digunakan untuk menyimpulkan hubungan filogenetik antar spesies (Saitou dan Nei, 1987).
6
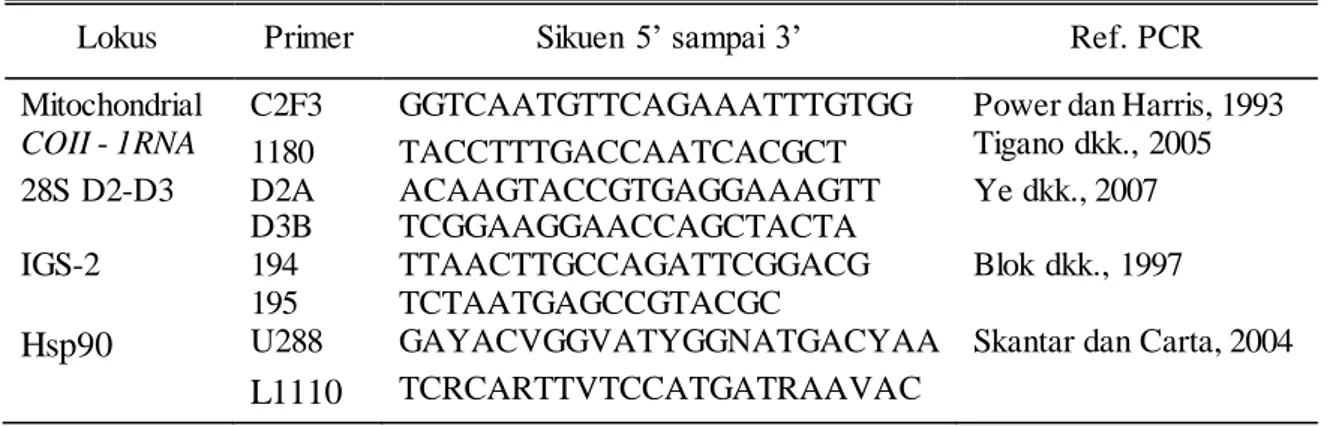
Gambar 1. Meloidogyne arenaria betina dengan mikroskop cahaya. (A-F) Pola perineal dari populasi pohon traveler. (G) Contoh tipe pola perineal, gambar slide Meloidogyne arenaria G9957, Koleksi Nematoda USDA. Spesimen dari Blacksburg, VA; tanaman inang = tomat. (H-I) Berturut-turut tubuh betina, populasi pohon traveler, tinggi dan rendah kekuatan gambar. (J-M) Populasi pohon traveler, anterior dengan lebar stilet.
7 Hasil dan Pembahasan
Analisis Morfologi: Populasi tidak menghasilkan puru pada pohon traveler,
dan daerah vulva nematoda betina biasanya dikelilingi oleh kantung telur. Perbedaan morfologi membuat populasi ini khususnya sulit untuk diidentifikasi, dan awalnya diduga spesies baru yang terkait erat dengan M. arenaria, M.
incognita dan M. javanica. Sementara populasi ini terdiri dari sebuah mosaik dari
kesamaan morfologi untuk beberapa spesies, pengukuran yang paling sesuai dalam kisaran yang dilaporkan sebelumnya untuk M. arenaria. Pola perineal nematoda betina cukup beragam (Gambar 1A-F), dengan putaran tinggi menunjukkan pola bersayap sesekali atau bahu dengan sedikit garis lateral. Dalam beberapa perineal bagian tengah dikelilingi oleh garis-garis kasar dan halus bergantian, lebih banyak, gelombang lebih terputus daripada dilihat dalam ciri khas M. arenaria (Gambar 1G). Pola perineal tidak memiliki bagian perivulval memanjang lateral yang biasanya ditemukan pada M. arenaria (Cliff dan Hirschmann, 1985; Hirschmann, 1985; Eisenback dan Triantaphyllou, 1991), melainkan suatu putaran memanjang ke arah dorsoventral. Celah vulva pada populasi ini menunjukkan lipatan lateral yang menonjol seperti tertekan.
Stilet berbentuk kerucut pada nematoda betina sedikit melengkung (Gambar 1J-M). Knob stilet betina bulat meluas, miring ke arah posterior, dan tidak secara konsisten mempunyai poros miring bertahap seperti yang ditemukan pada ciri khas M. arenaria (Eisenback dkk., 1980, 1981; Eisenback dan Triantaphyllou, 1991), melainkan menunjukkan aspek lateral lebih seperti M. incognita. Panjang stilet betina (Tabel 2) konsisten dengan M. arenaria dan berbeda dengan populasi
M. thailandica Handoo, Skantar, Carta dan Erbe, 2005, spesies yang ditemukan
pada jahe (Zingiber spp.) dari Thailand (Handoo dkk., 2005a), dan M.
morocciensis Rammah dan Horschmann, 1990 dari peach (Prunus persica L.
Batsch) di Morocco (Rammah dan Hirschmann, 1990). Dorsal Gland Outlet (DGO), posisi lubang saluran pembuangan dan vulva dengan panjang anus yang konsisten dengan rentang untuk M. arenaria (Tabel 2). Bentuk betina khas M.
arenaria tetapi banyak yang bengkok seperti huruf L (Gambar 1H, I).
8
terdapat pada M. thailandica. Kisaran panjang dan lebar tubuh betina jauh lebih besar daripada populasi M. arenaria, M. thailandica dan M. morocciensis.
Panjang stilet jantan dan panjang spikula sesuai dengan ciri khas M.
arenaria, tetapi panjang tubuh dan gubernakulum lebih pendek (Tabel 3). Rerata
panjang stilet jantan berbeda dengan populasi saat ini (22 µm) dan yang sebelumnya dilaporkan ukuran M. arenaria (23 µm) dari M. thailandica (18.7 µm) dan M. morrociensis (25 µm). Demikian juga, rerata panjang tubuh (1.493 µm) lebih panjang daripada M. thailandica (1.240 µm) dan lebih pendek daripada
M. morocciensis (1.621 µm). Populasi Ravenala juga tidak memliliki gerigi pada
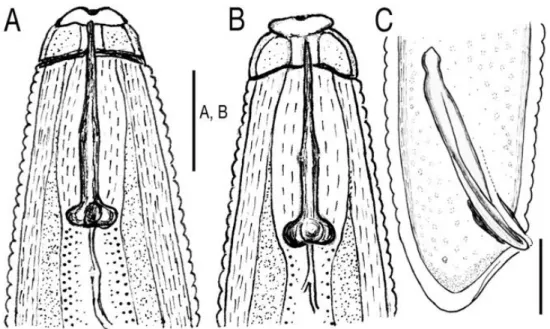
spikula terminus (gambar 2C) seperti pada M. thailandica.
Stilet pada juvenil stadia 2 (Gambar 3A) sangat mirip dengan M. arenaria, dengan rerata panjang (11.1 µm) berdeda dengan stilet M. thailandica yang lebih pendek (10.2 µm) dan lebih panjang daripada M. morocciensis (12.3 µm) (Tabel 4). Ekor juvenil panjang, meruncing dengan ujungnya membulat lebih halus dengan 1 sampai 4 badan yang ditarik masuk secara konsisten tampak pada bagian yang tidak berwarna (Gambar 3B). Panjang ekor juvenil tertinggi diantara kisaran populasi M. arenaria dan hampir sebesar M. thailandica (Tabel 4).
Gambar 2. Gambar Meloidogyne arenaria jantan dari pohon traveler. (A) Bagian lateral pada ujung anterior. (B) Bagian dorso-vnetral, ujung anterior. (C) Ekor dengan spikula dan gubernakulum.
9
Gambar 3. Juvenil stadia dua Meloidogyne arenaria dari pohon traveler dengan mikroskop cahaya. (A) Bagian ujung anterior dengan stilet dan kerongkongan.(B) Ekor. (C) Spesimen keseluruhan.
10
Tabel 2. Perbandingan morfometrika betina dari isolat Ravenala dibandingkan dengan populasi lain M. arenaria, M. platani, M. thailandica dan M.
morocciensis
Karakter (µm) M. arenaria (isolat
Ravenala) n = 6-12 M. arenaria M. platani d
n = 30 M. thailandicae n = 25 M. morocciensisf n = 35
Panjang tubuh hingga leher 893 ± 258 (718–1424)n=11 741±115b (601-985) 678±80 (540-860) 762±115.8 (570-955) 755±90.3 (5551-923) Lebar tubuh 566 ± 179 (290-906) n = 11 448±89h (334-626) 453±71 (320-585) 459±109.1 (272-690) 490±72.8 (219-608) Panjang stylet 15.5 ± 0.8 (15-17.5) n=12 15.1±0.05c(13-17) n=150 16.5±0.5 (15.8-17.3) 13.8±1.2 (12.0-15.5) 16.5±0.8 (15-18) Tinggi knob stylet 2.5 ± 0.4 (2-2.9) n = 6 2.5±0.2c(2.2-3.1) n = 150 2.5±0.2 (2.2-3.1) 2.4±0.5 (1.5-3) 2.3±0.2(2-2.6) DGO dari dasar stylet 4.5 ± 0.05 (4-5) n = 12 4.8±0.06c(3.1-6.6) n = 150 3.7±0.46 (2.8-4.6) 4.0±0.9 (3-5) 3.9±0.6(2.4-5.2) Lubang ekskretori dari ujung anterior 37.3 ± 10.5 (17.5-50) n= 12 42.2±0.94c(18-80) n=150 26.8±6.7 (16-40) 30.6±8.4 (15-47) 60.3±21.7 (24-126) Panjang celah vulva 25.8 ± 2.9 (20-29) n = 8 29.3±4.1 (24-37) n = 9b 27±2.9 (22-33) 29.1±2.6(25-35.1) 25.2±2.2 (22-30) Vulva-panjang anus 22.5 ± 1.7 (21-24) n = 8 20.7±2.6 (18-21) n = 9b 16.4±2.6(10-23) 18.0±2.4(15-22) 19.5±2.7(15-27)
a
Rerata ± standart deviasi (range): n = jumlah spesimen
b Jepson (1987); Penulis komposit beberapa populasi; tidak diberikan n jika tidak dicatat c
Cliff dan Hirschmann (1985); Penulis komposit beberapa populasi
d
Hirschmann (1982) e
Handoo dkk., (2005b)
f
11
Tabel 3. Perbandingana Morfometri nematoda jantan dari isolat Ravenala dibandingkan dengan populasi lain M. platani, M. thailandica, dan M.
morocciensis
Karakter (µm) M. arenaria (isolat
Ravenala) n = 6 M. arenaria
b
n = 150 M. platanic n = 30 M. thailandicad n = 25 M. morocciensise n = 35 Panjang tubuh 1.493±442 (1.100-1900) 1.720±287 (979-2.279) 1.355±276 (960-2.010) 1.240±205.2 (950-1.510) 1.621±155 (1.296-1863)
Panjang stylet 22±1.5 (20-23.5) 23±1.46 (22-28) 22±1.1 (19.4-24.3) 18.7±1.1 (17-20) 25±0.7(23-26)
Tinggi knob stilet 2.6±0.4 n = 6 (2.2-3.1) 4±0.4 (3-5) n = 100 3.3±0.3 (2.8-3.8) 2.6±0.2 (2.5-3.0) 3.1±0.2(2.5-3.6)
DGO dari stilet 4.2±0.8 (3.7-5.0) n = 3 6±1 (4-8) 3.7±0.6 (2.3-4.5) 3.7±0.9(2.5-5.0) 4.7±0.7(3.5-6.2)
Panjang spikula 31.4±0.9 (30-35) 32± 2 (27-39) n = 66 27.5±1.41 (25-32) 31.2±3.8 (25-38) 34.8±2.1(31-39)
Gubernaculum 8.1±1.2 (7.5-10) 9±0.8 (7-10) n = 25 8.5±0.6 (8-10) 8.9±1.2 (7.5-11.0) 9.4±0.8(8.1-10.7)
a
Rerata ± standar deviasi (range); n = jumlah spesimen b
Cliff dan Hirschmann (1985); Penulis komposit beberapa populasi
c
Hirschmann (1982) d
Handoo dkk., (2005b) e
12
Tabel 4. Perbandingana Morfometri stadia Juvenil-2 dari isolat Ravenala dibandingkan dengan populasi lain M. platani, M. thailandica, dan M.
morocciensis
Karakter (µm) M. arenaria (isolat
Ravenala) n = 6 M. arenaria
b
n = 150 M. platanic n = 30 M. thailandicad n = 25 M. morocciensise n = 35
Panjang tubuh 475±54 (390-570) 504±4.3 (392-605) 443±22.5 (395-497) 484±25.5 (450-540) 400.8±21.7 (374-454)
Lebar tubuh 14.1±1.4 (12.7-16)n=5 15.3±0.1 (13-18) 17±1.2 (15-20) 14.3±0.6 (13-15) 15.2±0.5(15-16)
Panjang stilet 11.1±0.8 (10-12.5) 11.1±0.03 (10-12) 12.2±0.04 (11.6-12.6) 10.2±0.3 (10-11) 12.3±0.5(11-13)
DGO dari stilet 3.7±0.2 (3.6-4.1) n = 5 3.7±0.04 (3-5) 3.5±0.04 (2.7-4) 2.9±0.3 (2.5-3.5) 3.8±0.4(3-4.4)
Panjang ekor 59±6.6 (50-67) 56.0±0.43 (44-59) 54.2±0.47 (50-65) 61.2±3.0 (55-65) 52.6±2.7(47-58)
Panjang ekor hyaline 14±3.3 (10-21) 12.4±0.2 (9.3-15.3) 18.3±1.9 (15-20) -
a
Rerata ± standar deviasi (range); n = jumlah spesimen b
Cliff dan Hirschmann (1985); Penulis komposit beberapa populasi
c
Hirschmann (1982) d
Handoo dkk., (2005b) e
13
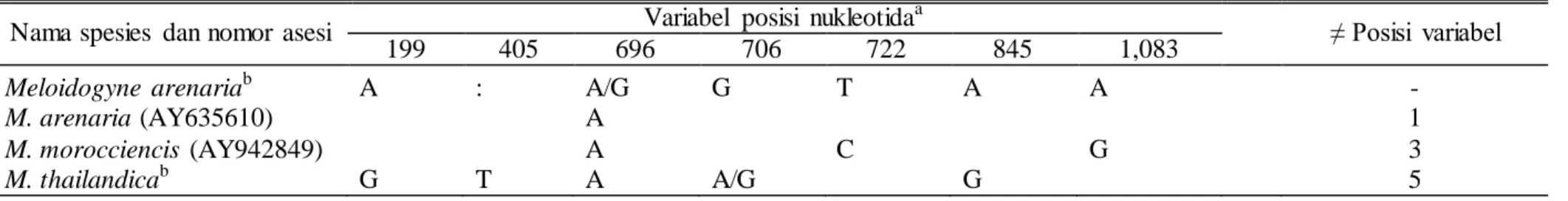
Tabel 5. Variabel posisi nukleotida dalam DNA mitokondria
Nama spesies dan nomor asesi Variabel posisi nukleotida
a
≠ Posisi variabel
199 405 696 706 722 845 1,083
Meloidogyne arenariab A : A/G G T A A -
M. arenaria (AY635610) A 1
M. morocciencis (AY942849) A C G 3
M. thailandicab G T A A/G G 5
a
Jumlah nukleotida menurut sikuen kesesuaian, dimulai dari basa pertama pada sikuen primer 5’. b
Penelitian ini.
Tabel 6. Variabel posisi nukleotida pada bagian perluasan 28S rDNA D2-D3 beberapa nematoda puru akar
Nama spesies dan nomor asesi Variabel posisi nukleotida
a ≠ Posisi variabel 1 15 95 96 103 178 302 312 393 507 545 751 754 Meloidogyne arenariab A C A T G A T A C A T C A - M. arenaria (AF435803) T G/T C G 4
M. incognita (AF435794) T G/T C/T A/G G 5
M. paranaensis Mp61 (AF435798) T T G T T G 6
M. konaensis (AF435797) T G/T G/T C A/T C/T G 7
M. paranaensis Mp62 (AF435799) T T G T C/T T G 7
M. paranaensis Mp71 (AF435800) T T T G T T G G 8
a
Jumlah nukleotida menurut sikuen kesesuaian, dimulai dari basa pertama pada sikuen primer 5’. b
14
Tabel 7. Variabel posisi nukleotida pada ribosom intergenic space region IGS-2 dari beberapa nematoda puru akar
Nama spesies dan nomor asesi
Variabel posisi nukleotidaa
14 32 67 89 117 118 144 182 211 250 274 379 442 487 534 635 694 ≠ Posisi variabel
Meloidogyne arenariab T : T A C T T/C C A/C A/T T T G A/G A A G -
M. thailandica (AY858796) : T A T A 5
M. arenaria (Blok dkk. 1997) : T A/C A T A 7
M. incognita (Blok dkk. 1997) : A T A/C A T A 7
M. javanica (Blok dkk. 1997) : T A/C A T C/T A A/T AG C 10
M. floridensis (AY194853) : C G C A T A T G : A 11 a
Jumlah nukleotida menurut sikuen kesesuaian, dimulai dari basa pertama pada sikuen primer 5’. b
Penelitian ini.
Tabel 8. Variabel posisi nukleotida pada penyesuaian sebagian sikuen genom Hsp90 dari beberapa nematoda puru akar Nama spesies dan
nomor asesi
Variabel posisi nukleotidaa
11 (42) (53) (89) 108 110 150 256 288 320 361 369 443 452 (728) (738) 819 (846) (861) (869) ≠ Posisi variabel
Meloidogyne arenaria T a a c C/T A T G A A T T A A t c T : a t -
M. arenaria (MD) t T t 2
M. thailandica t C/T A/C a/t a/t 4
M. floridensis t C a 4 haplotipe A t M. javanica t T C T t 4 M. incognita : : t C T : C a : 6 M. floridensis C t T C G A A G a 7 haplotipe B a
Jumlah nukleotida menurut sikuen kesesuaian, dimulai dari basa pertama pada sikuen primer 5’. b
15
Meskipun demikian, populasi Ravenala mudah dibedakan oleh panjang ekor yang lebih pendek (14 µm) dibanding M. thailandica (18.3 µm).
Mempertimbangkan karakter yang sebelumnya ditunjukkan sebagai contoh M. arenaria, populasi Ravenala bukan dibagi beberapa karakter kualitatif dengan M. platani. Hirschmann, 1982 (Rammah dan Hirschmann, 1990). Diantara mereka yang paling menonjol adalah pola bergelombang perineal betina, bentuk knob stilet jantan dan betina kurang landai, tinggi knob betina lebih pendek Namun, populasi Ravenala berbeda sekali dari M. platani karena memiliki tubuh betina yang lebih panjang dan jarak vulva-anal lebih panjang (Tabel 2); garis pola perineal tidak seragam, dan bidang lateral dan ujung ekor umumnya berbeda. Sementara tinggi knob stilet pada keduanya yaitu M. platani dan populasi Ravenala lebih rendah dari kisaran yang dilaporkan oleh Cliff dan Hirschmann (1985), mereka konsisten dengan ukuran (kisaran 2-4 mm; rata-rata 2,8 mm) dari populasi triploid lain (Rammah dan Hirschmann, 1990).
DGO jantan juga ditemukan berguna untuk mengkarakterisasi populasi M. arenaria (Rammah dan Hirschmann, 1990). Populasi M. arenaria dari Ravenala memiliki kisaran menengah dan rata-rata DGO (Tabel 3), lebih rendah di antara M. platani (Hirschmann, 1982) dan nilai-nilainya dilaporkan untuk beberapa populasi M. arenaria. Namun, populasi Ravenala maupun M. platani kekurangan termini spicule bifida dan areolation bidang lateral jantan dan memiliki daerah kepala yang tingginya diatas rata-rata berbentuk topi (Gambar 2A,B). Bentuk ekor jantan cukup runcing pada M. platani dan populasi Ravenala (Gambar 2C) jika dibandingkan dengan ciri khas M. arenaria (Jepson 1987). Ekor panjang hialin tidak jelas adalah ciri M. arenaria (Cliff & Hirschmann 1985), tetapi jelas pada populasi M. arenaria dan M. platani. Meskipun populasi Ravenala menunjukkan penampilan peralihan antara M. arenaria dan M. platani, yang diinduksi bengkak akar besar pada pohon ara, bukan tanaman inang M. arenaria, dan gagal untuk diperbanyak pada kacang tanah, jagung atau ubi (Hirschmann 1982).
Analisis molekuler: Hasil pengujian secara molekuler mencerminkan bahwa
analisis morfologi, mengungkapkan sebuah mosaik kemiripan molekuler untuk beberapa species NPA. Sayangnya, tidak ada sikuen DNA yang tersedia untuk M.
16 platani, sehingga tidak mungkin untuk melengkapi perbandingan secara morfologi menggunakan data molekuler. Mitokondria COII-1 RNA ukuran 1.1 kb hasil PCR dari populasi Ravenala dibagi menjadi ras 1 dan ras 2 populasi M.
arenaria dari Georgia, Utara dan Selatan Carolina, dan Florida (Powers dan
Harris 1993), populasi dari Ivory Coast, Portugal dan the French West Indies (Blok et al., 2002), dan satu dari Brazil (Tigano et al., 2005). Spesies lain yang mewakili produk berukuran hampir sama meliputi M. Morocciensis (1.113 bp), dan M. floridensis (1,110 bp) (Tigano et al., 2005), diisolasi dari peach (Prunus persicus L.) di Morocco dan Florida, masing-masing; M. thailandica (1.113 bp) dari jahe (Zingiber spp.) di Thailand (Handoo et al., 2005a); dan M. paranaensis (1.255 bp) dari kopi (Coffea arabica L.) (Tigano et al., 2005). Produk PCR mitochondrial COII-1 RNA dari M. javanica dan M. incognita lebih besar (Power and Harris, 1993; Tigano et al., 2005), ini membedakan spesies tersebut dari populasi Ravenala. Produk 1.7 kb telah dilaporkan untuk beberapa populasi M. arenaria (Power dan Harris 1993). Ini kemungkinan besar sesuai dengan haplotype C/H sebelumnya, yang dapat dibedakan dari tipe A yang lebih kecil terutama dari 529 bp dari penghapusan dalam daerah intergenic (Hugall et al., 1994; Stanton et al., 1997). Sikuen mitokondria COII-1RNA (EU364879) sama dengan M. arenaria (Tabel 5), dengan satu posisi tidak jelas (A/G) ditemukan pada populasi Ravenala. Analisis hubungan kekerabatan (NJ) (tidak ditampilkan) dikelompokkan isolat Ravenala dengan populasi lain M. arenaria (AY635610) dan dengan M. morocciensis (AY942849), sesuai persetujuan dengan hasil sebelumnya (Tigano dkk. 2005), dan dengan M. thailandica (EU364879). Meskipun produk amplifikasi berukuran sama, sikuen mitokondria M. floridensis berisi indeks relatif lebih besar untuk sikuen M. arenaria dan beberapa mirip dengan salah satu dari M. incognita (Tigano dkk. 2005). Perluasan bagian D2-D3 sub unit besar (LSU) 28S rDNA adalah 754 bp untuk populasi Ravenala (EU364889). Penjajaran dengan enam dari sikuen yang paling erat kaitannya dari GenBank mengungkapkan 13 posisi variabel nukleotida (tabel 6). Sikuen yang paling mirip (AF435794) adalah populasi M. arenaria ras 2 dari kedelai (Tenente dkk. 2004), berbeda pada 4 bp, termasuk satu posisi yang tidak jelas. Sikuen lain dengan kemiripan tinggi termasuk M. incognita (AF435794) juga dari kedelai,
17
populasi M. konaensis paranaensis Mp61 (AF435797) dari kopi di Hawaii, dan Brazilian M. paranaensis Mp61(AF435798), Mp62 (AF435799) dan Mp71 (AF435800), juga diisolasi dari kopi. Karena beberapa dilaporkan sebelumnya sikuen 28S D2-D3 (Tenente dkk. 2004) berisi basa yang tidak jelas (kemungkinan besar karena sikuen hasil PCR langsung, bukan klon), ini tidak jelas bagaimana memisahkan kejadian haplotipe dengan spesies tersebut. Perbandingan pemisahan sikuen D3 termasuk beberapa dari GenBank (tidak ditampilkan) memberikan petunjuk yang sama. Analisis D3 sebelumnya menunjukkan bahwa M. arenaria, M. incognita dan M. hapla berisi tujuh haplotipe diantaranya, dengan dua (haplotipe 5 dan 7) ditemukan pada ketiga spesies tersebut (Chen dkk. 2003), dan pengujian M. thailandica diklon D3 hasil PCR menunjukkan lima haplotipe (A –
E) (Handoo dkk. 2005a). Disini ditemukan bahwa populasi M. arenaria dari
Ravenala dibagi D3 haplotipe 5 banyak kesamaan dengan populasi M. arenaria
MA, M. incognita COA dan MUL dan M. javanica JNC, J811 dan VW4 (Chen dkk. 2003), yang sama juga dengan haplotipe B dari M. thailandica (Handoo dkk. 2005a). Karena sebagian besar polimorfisme pada perbandingan kami ada di lokasi D2 (Tabel 6), tampak pada daerah ini memberikan resolusi filogenetik yang lebih baik dalam kelompok apomiksis, terutama ketika beberapa klon dari populasi yang diuji.
Penggandaan daerah IGS-2, diantara 5S dan 18S rDNA, adalah 716 bp untuk populasi Ravenala (EU364878), sesuai dengan ukuran yang dilaporkan sebelumnya untuk NPA spesies tropis dan berbeda dari hasil yang lebih besar ditemukan pada M. mayaguensis dan yang lebih kecil pada M. hapla (Blok dkk. 1997). Dari penyesuaian 717 bp, posis 17 merupakan variabel antara NPA yang dibandingkan (Tabel 7). Sikuen paling mirip adalah M. thailandica, M. arenaria,
M. incognita, M. javanica dan M. floridensis. Tanpa menentukan polimorfisme M. arenaria jelas terpisah, dan, kecuali khusus untuk sikuen M. floridensis,
nukleotida tidak jelas dalam penanda mengaburkan batas-batas antara spesies. asil P sp b a ala sikuen untuk populasi Ravenala (EU364880),
M. arenaria dari Maryland (FJ238508) dan beberapa spesies NPA yang lain
termasuk M. thailandica (EU364882) dari Thailand (Handoo dkk. 2005a), M.
18 M. javanica (AF201338) dari Maryland dan M. incognita (EU364881). Semua
bagian sikuen Hsp90 terdiri dari 5 ekson dan 4 intron. Populasi M. floridensis tiga dari Florida dan satu dari Georgia diwakili oleh haplotipe Hsp90 yang berbeda. Selebihnya populasi Meloidogyne menghasilkan sikuen tunggal, meskipun beberapa posisi polimorfik dideteksi antara mereka (Tabel 8). Sikuen dirapikan menjadi 972 basa untuk dihilangkan sikuen primer terakhir, yang dianggap tidak dapat diandalkan. Diantara sikuen yang dibandingkan, ada 20 posisi polimorfik, dengan 8 basa perubahan dalam intron, dua perubahan pada posisi kodon pertama, lima pada posisi kedua dan empat pada posisi ketiga. Meloidogyne arenaria dari Maryland hampir sama dengan populasi Ravenala, dengan satu peralihan C-T dan satu basa tunggal sisipan. Penyimpangan sikuen pada M. thailandica, haplotipe A dan B M. floridensis, M. javanica dan M. incognita adalah <1%, sama dengan yang ditemukan untuk lokus lain yang diteliti. Berbeda dengan rendahnya tingkat divergensi antara apomiksis spesies NPA, dua sikuen M. hapla yang ditentukan sebelumnya (Handoo dkk. 2005b) berbeda nyata (17%) dari populasi
Ravenala. Pemisahan M. hapla dari garis keturunan spesies apomiksis sesuai
dengan yang ditemukan melibatkan lokus yang lainnya (Hugall dkk. 1994, 1999; Chen dkk. 2003; Tigano dkk. 2005; Castagnone Sereno, 2006).
Sikuen yang sudah ada di perpustakaan dataset cDNA M. arenaria berisi tumpang tindih beberapa EST terkait dengan Hsp90, sehingga sikuen komposit yang dibentuk dari GenBank aksesi B1747178, B1747494 dan B1745855. Rakitan sikuen EST berisi sejumlah basa yang tidak jelas dan berbeda dari populasi Ravenala yang mengkode sikuen pada posisi enam (Gambar 4).
Beberapa ketidakjelasan muncul mengarah pada bagian ujung individu EST (tidak ditampilkan) dan mungkin disebabkan urutan basa yang diragukan yang berasal dari satu kali pembacaan atau mewakili varian sikuen Hsp90 yang asli. Pengujian klon Hsp90 dari berbagai populasi M. arenaria akan diperlukan untuk menetapkan tingkat keragaman antarspesies dalam gen, tetapi tampaknya Hsp90 tidak lebih baik atau lebih buruk untuk memisahkan spesies daripada penanda mitokondria atau ribosom yang diuji.
Sementara sebagian besar populasi M.arenaria menginduksi puru dengan kuat, beberapa menyebabkan lebih kecil, puru seperti manik-manik (Eisenback
19
dan Triantaphylou 1991). Meskipun demikian, tidak satupun dilaporkan gagal untuk menginduksi puru seperti yang satu ini. Tanaman dari ordo Zingiberales memiliki delapan famili, termasuk Steriliziaceae, Zingiberaceae dan Heliconiae. Steriliziaceae (Quénéhervé dkk. 1997) mengandung satu spesies dalam genus Ravenala (termasuk pohon traveler, R. madagascariensis), satu dari Phenakhospermum dan lima dalam Strelitzia (termasuk bird-of-paradise). Kejadian pada spesies Meloidogyne khususnya dalam Ravenala sangat sedikit yang dilaporkan. Meskipun, tiga spesies utama partenogenesis menyerang jahe (Zingiber, dalam Zingiberaceae) (Bapista dos Santos dan Lazaro Lozano 1993), dan M. thailandica (Handoo dkk 2005a) juga menyerang Heliconia spp. (bird-of-paradise palsu), sebuah genus dalam Heliconiae. Tanaman inang kesukaan mewakili fenotipe konvergen, karena tanaman inang dari ras yang sama menunjukkan variabilitas genetik yang besar (Baum dkk. 1994). Keragaman genetik paradoks dari empat spesies utama NPA partenogenesis (Castagnone-sereno 2006) adalah yang paling menonjol pada populasi M. arenaria (Powers 2004). Mungkin saat itu, tidaklah mengherankan bahwa tanggapan fenotipe tanaman inang langka harus menyertai variasi secara morfologi dan molekuler. Spesimen dari tahap yang sesuai atau jumlah diharapkan untuk ulangan analisis isozim (Esbenshade dan Triantaphyllou 1990) RAPD (Cenis 1993; Randig dkk 2002) atau pengujian kiasaran tanaman inang kurang dan sayangnya tidak dapat dimasukkan dalam diagnosa. Uji SCAR PCR menggunakan primer spesifik spesies (Zijlstra dkk 2000) tidak dilakukan, sebagai pilihan penanda molekuler diuji untuk spesimen terbatas dipengaruhi oleh kegunaan masa depan terhadap sikuen untuk penelitian filogenetik yang lebih luas. Uji ini kemungkinan akan menegaskan hasil morfologi dan molekuler yang disajikan di sini. Nilai positif untuk M. arenaria melalui isozim, RAPD atau penanda SCAR akan menambah dukungan lebih lanjut untuk diagnosis; akan tetapi, membingungkan, hasil baru atau negatif dari pengujian ini akan menegaskan kesimpulan kami bahwa populasi ini adalah sebuah varian tidak khas M. arenaria.
Penelitian ini menekankan fakta bahwa garis batas untuk spesies tampaknya memudar seperti spesies baru Meloidogyne dan ciri populasi. Diagnosis yang akurat memerlukan berbagai karakter morfologi dan molekuler seperti yang
20
disajikan di sini. Tingkat relatif variasi intraspesifik diamati dengan variasi antarspesies untuk NPA tropis lebih menyoroti pentingnya karakter tambahan dan penanda molekuler yang jelas akan membedakan spesies serta menginformasikan hubungan evolusioner mereka.