PENGARUH POLA KETIADAAN INANG TERHADAP EKOLOGI REPRODUKSI Snellenius manilae ASHMEAD
(HYMENOPTERA: BRACONIDAE)
GAMATRIANI MARKHAMAH
DEPARTEMEN PROTEKSI TANAMAN
FAKULTAS PERTANIAN
INSTITUT PERTANIAN BOGOR
BOGOR
Dibimbing oleh DAMAYANTI BUCHORI, ALI NURMANSYAH.
Parasitoid Snellenius manilae memiliki potensi untuk mengendalikan hama Spodoptera litura. Penelitian ini bertujuan mengetahui pengaruh pola ketiadaan inang S. manilae terhadap ekologi reproduksinya. S. manilae yang memiliki umur sama diberikan perlakuan berupa pola ketiadaan inang. Inang yang diberikan kepada parasitoid tersebut adalah larva instar II S. litura sebanyak 30 ekor per 1 parasitoid. Pemaparan larva S. litura ke S. manilae dilakukan selama 24 jam dan penggantian inang larva S. litura ini dilakukan setiap hari sesuai dengan perlakuan ketiadaan inang yang diuji. Ketiadaan inang di awal selama empat hari cenderung dapat meningkatkan parasitisasi S. manilae pada hari ke-8, sedangkan ketiadaan inang dua hari di tengah cenderung dapat menurunkan parasitisasi
S. manilae pada hari ke-8. S. manilae yang mengalami ketiadaan inang 4 hari baik
di awal maupun di akhir hidupnya cenderung dapat meletakkan telur pada hari ke-8 lebih banyak daripada pola ketiadaan inang lainnya. S. manilae yang mengalami ketiadaan inang empat hari di awal hidupnya cenderung dapat meningkatkan terjadinya superparasitisasi pada hari ke-8. Kejadian superparasitisasi terbanyak adalah dua butir telur parasitoid per inang, sedangkan kejadian yang paling sedikit adalah enam butir telur parasitoid per inang. Parasitoid S. maniale yang mengalami ketiadaan inang dapat meletakkan telur dengan jumlah yang lebih banyak pada hari pertama setelah ketiadan inang (18 butir-31 butir). Total produksi telur dan lama hidup imago S. manilae dipengaruhi oleh lamanya ketiadaan inang, sedangkan sisa telur dalam ovari S. manilae dipengaruhi baik oleh pola ketiadaan inang maupun lama ketiadaan inang. Ketiadaan inang menyebabkan total produksi telur mengalami penurunan dan lama hidup menjadi lebih panjang. Sisa telur dalam ovari S. manilae cenderung meningkat ketika ketiadaan inang terjadi di tengah masa hidupnya dan semakin meningkat ketika lama ketiadaan inang meningkat.
PENGARUH POLA KETIADAAN INANG TERHADAP EKOLOGI REPRODUKSI Snellenius manilae ASHMEAD
(HYMENOPTERA: BRACONIDAE)
GAMATRIANI MARKHAMAH
A34070045
Skripsi
sebagai salah satu syarat untuk memperoleh gelar Sarjana Pertanian di Departemen Proteksi Tanaman, Fakultas Pertanian, Institut Pertanian Bogor
DEPARTEMEN PROTEKSI TANAMAN
FAKULTAS PERTANIAN
INSTITUT PERTANIAN BOGOR
BOGOR
NIM : A34070045
Tanggal lulus:
Disetujui, Dosen Pembimbing 2
Dr. Ir. Ali Nurmansyah, M.Si. NIP 19630212 199002 1 001 Disetujui,
Dosen Pembimbing 1
Dr. Ir. Damayanti Buchori, M.Sc. NIP 19601028 198603 2 002
Diketahui, Ketua Departemen
Dr. Ir. Abdjad Asih Nawangsih, M.Si. NIP 19650621 198910 2 001
RIWAYAT HIDUP
Penulis lahir di Tangerang pada tanggal 26 November 1989 sebagai anak kedua dari empat bersaudara pasangan Ibu Faridah Bintari dan Bapak Irfanudin Poeger Army. Penulis menyelesaikan pendidikan sekolah lanjutan atas di SMA Negeri 1 Ciputat, Tangerang (2004-2007).
Pada tahun 2007 penulis melanjutkan pendidikannya di Institut Pertanian Bogor (IPB) melalui jalur Undangan Seleksi Masuk (USMI) IPB pada kurikulum berbasis mayor-minor. Penulis diterima sebagai mahasiswa Departemen Proteksi Tanaman, Fakultas Pertanian, IPB dan mengikuti masa Tingkat Persiapan Bersama selama 1 tahun. Pada tahun berikutnya penulis melanjutkan pendidikannya dengan Mayor Proteksi Tanaman, Fakultas Pertanian.
Selama masa kuliah, penulis aktif bergabung dengan beberapa organisasi kampus seperti Koperasi Mahasiswa (KOPMA), Unit Kegiatan Mahasiswa (UKM) bola basket, Seroja Putih, Badan Eksekutif Mahasiswa (BEM) Fakultas Pertanian, Organic Farming, dan Metamorfosa, mengikuti kepanitiaan pada beberapa acara kampus, menjadi kontingen basket putri untuk Olimpiade Mahasiswa IPB (OMI), dan mengikuti beberapa pelatihan, seperti pendidikan dasar koperasi mahasiswa, pelatihan jurnalistik, dan pelatihan penulisan artikel ilmiah. Penulis menjadi asisten praktikum mata kuliah Biologi Patogen pada bulan September 2009-Januari 2010 dan memiliki pengalaman bekerja sebagai asisten di Klinik Tanaman IPB pada tanggal 14 Juni-12 Juli 2009.
Puji dan syukur penulis panjatkan ke hadirat Allah SWT karena berkat rahmat, hidayah serta kasih sayang-Nya penulis dapat menyelesaikan skripsi yang berjudul “Pengaruh Pola Ketiadaan Inang Terhadap Ekologi Reproduksi
Snellenius manilae Ashmead (Hymenoptera: Braconidae)”. Penelitian dan
penulisan skripsi ini merupakan salah satu syarat untuk memperoleh gelar Sarjana Pertanian di Departemen Proteksi Tanaman, Fakultas Pertanian, Institut Pertanian Bogor. Penelitian dilaksanakan di Laboratorium Bioekologi Parasitoid dan Predator, Departemen Proteksi Tanaman dari bulan Februari 2011 sampai September 2011.
Ucapan terima kasih penulis sampaikan kepada Dr. Ir. Damayanti Buchori, M.Sc selaku dosen pembimbing skripsi dan dosen pembimbing akademik yang telah memberikan bimbingan, ilmu, pengetahuan, saran, dan motivasi; Dr. Ir. Ali Nurmansyah, M.Si selaku dosen pembimbing skripsi yang telah memberikan banyak saran, bimbingan, ilmu dan pengetahuan; Dr. Ir. Kikin Hamzah Mutaqin, M.Si selaku dosen penguji tamu yang telah memberikan saran dan motivasi;
Rasa terima kasih yang sangat mendalam ditujukan kepada kedua orang tua, Dra. Faridah Bintari, MM dan Drs. Irfanudin Poeger Army yang selalu mendoakan penulis dan memberikan motivasi serta dukungan moril maupun materil; Zuhdi Muntaha S.Mn sebagai kakak yang sering memberikan saran yang bermanfaat serta adik-adik, Ruchdi Mutaqin dan Kahfi Mubegi yang sering membantu penulis; Teman-teman seperjuangan yang bekerja di Laboratorium Bioekologi Predator dan Parasitoid, Eldiary Akbar, Yulius Dika C, Indri A., Aisyah, Putri Syahierah, dan Amanda M; Teman-teman yang sering membantu dan memberi semangat kepada penulis, Anik N., Sherli A., Sistania A., Dwi Dinar M., Dolpina A.R., Reka P., Triyastuti P., Mey F.; Yayu S.N., Nelly N., Ikra N, Riska N. P., Doni A.S., Avanty W. M Lutfi A., Hendi I., Elyta S., Eka Intina W., Irna Rizkiana D, Dian M., Alice Mayella A,.; Mahasiswa Departemen Proteksi Tanaman, khususnya angkatan 44 atas semangat dan canda tawa yang selalu menghiasi hari-hari; Staff yang sering membantu, Adha Sari, Mba Nita, Pak Yusuf, Pak Sodik, dan Pak Karto.
Penulis menyadari bahwa dalam skripsi ini masih terdapat banyak kekurangan untuk itu penulis mengharapkan kritik dan saran yang membangun. Semoga skripsi ini dapat bermanfaat bagi perkembangan ilmu pengetahuan.
Bogor, 10 Januari 2012
DAFTAR ISI
Halaman DAFTAR TABEL ... ix DAFTAR GAMBAR ... x DAFTAR LAMPIRAN ... xi PENDAHULUAN ... 1 Latar Belakang ... 1 Tujuan ... 2 Manfaat ... 3 TINJAUAN PUSTAKA ... 4 Parasitoid S. manilae ... 4S. litura sebagai Inang Parasitoid S. manilae ... 6
Kaitan Ketersediaan Inang dengan Kebugaran Parasitoid ... 7
BAHAN DAN METODE ... 9
Waktu dan Tempat ... 9
Metode Penelitian ... 9
Perbanyakan S. litura ... 9
Perbanyakan Parasitoid S. manilae ... 10
Rancangan Pola Ketiadaan Inang ... 10
Pemberian Perlakuan ... 12
Penghitungan Jumlah Telur ... 13
Analisis Data ... 15
HASIL DAN PEMBAHASAN ... 16
Hasil ... 16
Pengaruh Pola Ketiadaan Inang pada S. manilae Terhadap
Tingkat Parasitisasi dan Produksi Telur ... 18
Pengaruh Pola Ketiadaan Inang pada S. manilae Terhadap Sisa Telur, Total Telur, dan Lama Hidup ... 26
Pembahasan ... 28
Pengaruh Ketiadaan Inang pada S. manilae Terhadap Tingkat Parasitisasi dan Produksi Telur ... 29
Pengaruh Pola Ketiadaan Inang pada S. manilae Terhadap Sisa Telur, Total Telur, dan Lama Hidup ... 32
KESIMPULAN ... 35
DAFTAR PUSTAKA ... 36
ix
DAFTAR TABEL
Nomor Halaman 1 Sebaran jumlah telur S. manilae yang ditemukan pada setiap inang
S. litura ... 17
2 Pengaruh pola ketiadaan inang pada S. manilae terhadap tingkat
parasitisasi per hari hingga hari ke-8 ... 21 3 Pengaruh pola ketiadaan inang pada S. manilae terhadap jumlah
telur yang diletakkan per hari hingga hari ke-8 ... 22 4 Pengaruh pola ketiadaan inang pada S. manilae terhadap sisa
DAFTAR GAMBAR
Nomor Halaman
1 Perlakuan ketiadaan inang pada parasitoid S. manilae ... 12 2 Rata-rata tingkat parasitisasi S. manilae per hari ... 19 3 Diagram pencar korelasi antara tingkat parasitisasi dengan
jumlah telur yang diletakkan pada hari ke-8 oleh S. manilae yang
telah diberikan perlakuan ... 20 4 Total telur yang diletakkan hingga hari ke-8 oleh S. manilae ... 23 5 Jumlah telur yang diletakkan S. manilae pada hari pertama
setelah ketiadaan inang .. ... 24 6 Superparasitisasi pada hari ke-8 oleh S. manilae yang telah
diberikan perlakuan ... 25 7 Diagram pencar korelasi antara lama hidup dengan total produksi
telur pada imago betina S. manilae yang telah diberikan
xi
DAFTAR LAMPIRAN
HalamanGambar 1 (A) Telur parasitoid S. manilae (umur 24 jam);
(B) Larva parasitoid S. manilae (umur 48 jam) ... 39
Gambar 2 Larva parasitoid S. manilae (umur 72 jam) ... 40
Gambar 3 Imago betina S. manilae (A) tampak dorsal (B) tampak ventral ... 40
Tabel 1 Rata-rata tingkat parasitisasi S. manilae per hari ... 41
Tabel 2 Pengaruh pola ketiadaan inang pada S. manilae terhadap total telur hingga hari ke-8 ... 41
Tabel 3 Pengaruh pola ketiadaan inang pada S. manilae terhadap jumlah telur yang diletakkan pada hari pertama setelah ketiadaan inang ... 42
Tabel 4 Pengaruh pola ketiadaan inang pada S. manilae terhadap superparasitisasi pada hari ke-8 ... 42
Tabel sidik ragam tingkat parasitisasi pada hari ke-8 ... 43
Tabel sidik ragam rata-rata tingkat parasitisasi per hari ... 43
Tabel sidik ragam jumlah telur pada hari ke-8 ... 43
Tabel sidik ragam jumlah telur pada hari pertama setelah ketiadaan inang ... 43
Tabel sidik ragam total telur hinga hari ke-8 ... 43
Tabel sidik ragam sisa telur dalam ovari ... 44
Tabel sidik ragam total produksi telur ... 44
Tabel sidik ragam lama hidup ... 44
Latar Belakang
Parasitoid soliter yang dapat ditemukan di daerah tropis seperti Indonesia ialah Snellenius manilae (Hymenoptera: Braconidae). Selain terdapat di daerah tropis, ternyata parasitoid ini juga ditemukan memarasit larva S. litura di Pulau Okinawa, Jepang yang merupakan daerah subtropis (Ken et al 2006). Parasitoid dari genus yang sama dapat memarasit inang yang berbeda, contohnya Microplitis
rufiventris memarasit Spodoptera littoralis (Aly 2010); Microplitis plutellae
memarasit Plutella xylostella (Gharuka & Talekar 2004); Microplitis croceipes (Cresson) memarasit Heliothis virescens dan Helicoverpa armigera (Hübner) (Cameron et al. 2006); Microplitis demolitor memarasit Heliothis virescens dan
Pseudoplusia includens (Harvey AJ et al. 2004).
S. manilae termasuk dalam parasitoid koinobion, yaitu parasitoid yang
membiarkan inangnya terus berkembang setelah parasitisasi (Godfray 1994). Jika diklasifikasikan berdasarkan stadia inang yang diserang, S. manilae termasuk dalam parasitoid larva. Perkembangan S. manilae dari telur sampai menjadi imago terjadi ketika S. litura dalam stadia larva. S. litura instar 2 merupakan stadia yang paling sesuai untuk perkembangan hidup larva S. manilae, namun ternyata
S. manilae lebih memilih meletakkan telurnya pada S. litura instar 3 daripada S. litura instar 2 (Ratna 2008). Ketika S. manilae masih dalam stadia larva, suplai
nutrisi untuk perkembangan didapat dari hemolymph larva S. litura. Stadia imago
S. manilae mendapatkan asupan nutrisi dari embun madu atau nektar. Asupan
nutrisi berpengaruh terhadap kebugaran imago parasitoid, diantaranya lama hidup, keperidian, dan keefektifan pencarian inang (Jervis & Kidd 1997).
Imago S. manilae yang berada di lapangan terkadang tidak bertemu inang untuk beberapa hari. Kemungkinan pertemuan S. manilae dengan inang di lapangan bisa sangat beragam, diantaranya S. manilae bertemu dengan inang pada hari pertama kemunculannya menjadi imago, kemudian akan bertemu kembali
dengan inang ketika imago berumur enam hari. Kemungkinan lain ialah
S. manilae tidak bertemu dengan inang sejak hari pertama kemunculannya
2
hari. Kondisi-kondisi tersebut kemungkinan akan berpengaruh terhadap reproduksi S. manilae. Berdasarkan penelitian yang telah dilakukan oleh Hegazi
et al. (2007) mengenai pengaruh ketiadaan inang pada Microplitis rufiventris,
yaitu M. rufiventris yang mengalami ketiadaan inang lebih lama, memiliki muatan telur yang rendah di oviduk. Hegazi dan Khafagi (2001) melaporkan ketiadaan inang sampai lima hari pada Trichogramma cacoeciae dan T. dendrolimi tidak memengaruhi parasitisasi di hari pertama oviposisi, namun pada T. dendrolimi terjadi penurunan tingkat parasitisasi yang signifikan pada hari kedua setelah oviposisi.
Parasitoid proovigenic dan synovigenic memberikan tanggapan yang berbeda ketika menghadapi ketiadaan inang di awal kemunculannya. Ketiadaan inang pada parasitoid proovigenic tidak menyebabkan resorpsi telur (Jervis & Kidd 1997), sedangkan ketiadaan inang pada parasitoid synovigenic dapat menginduksi resorpsi telur yang berdampak pada pengurangan jumlah telur pada ovari (Navasero & Elzen 1992). Hal yang sama juga dinyatakan oleh Papaj (2000), bahwa salah satu faktor penting yang memengaruhi muatan telur pada parasitoid synovigenic ialah ketersediaan inang.
Dengan adanya potensi yang dimiliki oleh S. manilae sebagai agen hayati, maka informasi lebih dalam mengenai hal-hal yang memengaruhi reproduksi parasitoid ini perlu diketahui dengan baik. Adanya informasi mengenai pengaruh ketiadaan inang pada S. manilae terhadap ekologi reproduksinya diharapkan dapat mengoptimalkan peran S. manilae sebagai pengendali S. litura di lapangan.
Tujuan
Penelitian ini bertujuan untuk mengetahui pengaruh pola ketiadaan inang terhadap ekologi reproduksi S. manilae.
Manfaat
Hasil penelitian ini diharapkan dapat memberikan informasi mengenai kemampuan S. manilae ketika tidak menemukan inang sehingga perencanaan parasitoid ini sebagai pengendali S. litura di lapangan dapat dioptimalkan dengan baik.
TINJAUAN PUSTAKA
Parasitoid S. manilae
Parasitoid merupakan serangga yang larvanya berkembang di dalam atau pada tubuh organisme inang, inang dari parasitoid biasanya berupa serangga lain (Godfray 1994). Parasitoid memiliki tingkah laku yang rumit, diantaranya strategi dalam pencarian makanan dan inang. Ada beberapa kriteria yang membedakan parasitoid dengan hewan parasit lainnya, yaitu kebiasaan memarasit hanya terjadi pada stadia larva, stadia dewasa/ imago hidup bebas, parasitoid biasanya hanya membunuh dan mengonsumsi satu inang, ukuran tubuh parasitoid tidak jauh berbeda dengan inang, siklus hidup parasitoid cenderung sederhana, parasitoid biasanya memiliki kekerabatan yang dekat dengan inang dalam taksonominya, parasitoid memiliki kapasitas reproduksi diantara parasit sejati dan bentuk hidup bebas (Dout 1959).
Braconidae merupakan salah satu famili yang sudah sangat luas digunakan dalam pengendalian hayati, terutama untuk melawan kutu-kutuan dan beberapa jenis Lepidoptera, Coleoptera, dan Diptera. Parasitoid S. manilae termasuk salah satu parasitoid dari famili Braconidae. Telur S. manilae berbentuk silindris dan sedikit membengkok pada bagian tengah dengan kedua ujung membulat serta pada salah satu ujungnya terjadi penyempitan (Prabowo 1996). Stadia larva parasitoid terdiri dari empat instar dengan ciri-ciri instar pertama manibulat-kaudat, instar kedua ekor telah menghilang digantikan dengan vesikel anal, instar tiga bertipe vesikulat, dan instar empat bertipe hymenopteriform (Prabowo 1996). Pupa bertipe ekserat berada dalam kokon dan imago berwarna hitam dengan pangkal metasoma bagian ventral berwarna lebih terang (Prabowo 1996).
Godfray (1994) menyatakan bahwa perkembangan telur hingga stadia larva tahap akhir parasitoid terjadi di dalam tubuh inang. Menurut Godfray (1994) parasitoid yang berkembang dan makan di dalam tubuh inang dikenal sebagai endoparasitoid. Berdasarkan ciri tersebut, S. manilae termasuk dalam endoparasitoid. Telur yang biasa diletakkan S. manilae di dalam tubuh inang biasanya berjumlah satu butir telur. Jika ternyata ada parasitoid dari spesies yang
sama memarasit inang yang telah terparasit sebelumnya, maka terjadilah peristiwa superparasitisasi. Walaupun dalam satu tubuh inang terdapat lebih dari satu telur, setelah telur tersebut menetas hanya satu larva saja yang dapat berkembang menjadi imago. Berdasarkan ciri tersebut, S. manilae termasuk dalam parasitoid soliter. Godfray (1994) menyatakan bahwa parasitoid yang mendapatkan makanan dari inangnya untuk dirinya sendiri (tunggal) dikenal sebagai parasitoid soliter, sedangkan parasitoid yang mendapatkan makanan dari inangnya untuk dua atau lebih larva secara bersamaan dikenal sebagai parasitoid gregarius. Larva S. litura instar awal merupakan inang yang biasa diserang oleh S. maniale. Berdasarkan dari inang yang diserang, S. manilae termasuk dalam parasitoid larva. Menurut Ratna (2008), S. manilae lebih memilih meletakkan telurnya pada S. litura instar 3, namun ternyata S. litura instar 2 merupakan stadia yang paling sesuai untuk perkembangan hidup larva S. manilae. S. litura yang terparasit oleh S. maniale tidak langsung mengalami kematian namun tetap hidup dan berkembang sampai batasan tertentu. Berdasarkan ciri tersebut, S. manilae tergolong dalam parasitoid
koinobiont. Quickie (1997) menyatakan bahwa parasitoid yang memarasit inang,
namun inang tersebut tetap terus berkembang setelah parasitisasi merupakan parasitoid koinobiont.
Masa pra oviposisi imago betina S. manilae cukup singkat, yaitu kurang dari satu hari (Prabowo 1996). S. manilae betina yang baru keluar dari pupa
kemudian berkopulasi dapat dengan segera meletakkan telur pada inang.
S. manilae dapat terus memproduksi telur sepanjang hidupnya, berdasarkan ciri
ini kemungkinan besar S. manilae termasuk dalam parasitoid synovigenic. Jervis dan Kidd (1997) menyatakan bahwa parasitoid dapat dibagi menjadi jenis
pro-ovigenic yaitu parasitoid yang muncul dengan mengandung telur yang sudah
matang dan synovigenic yaitu parasitoid yang muncul dengan mengandung sebagian telur matang namun akan terus melakukan proses pematangan telur sepanjang hidupnya. Synovigenic dibagi lagi menjadi anhydropic yaitu jenis parasitoid yang mempunyai telur dalam jumlah sedikit dalam satu waktu dan
hydropic yaitu jenis parasitoid yang mempunyai banyak telur (Jervis & Kidd
6
Cukup sulit membedakan antara imago betina dan jantan S. manilae. Perbedaan yang dapat dilihat ialah pada bagian metasoma dan panjang antena. Menurut Prabowo (1996), metasoma imago jantan dari arah ventral terlihat lebih langsing daripada imago betina dan pangkal metasoma betina relatif lebih terang daripada imago jantan. Pada imago jantan antena terlihat lebih panjang daripada panjang tubuhnya, sedangkan antena imago betina cenderung lebih pendek. Ovipositor pada imago betina dapat dengan jelas terlihat dengan bantuan mikroskop stereo. Rata-rata panjang ovipositor pada imago betina adalah 0.15 mm (Prabowo 1996). Lama hidup imago S. manilae dilaporkan rata-rata 6-8 hari (Shepard 1991 et al. dalam Ratna 2008). Pemberian madu 20% dapat memperpanjang umur pada parasitoid S. manilae sampai sembilan hari dan dapat meningkatkan produksi keturunan dua kali lipat dibandingkan imago yang hanya diberi pakan air (Prabowo 1996).
S. litura sebagai Inang Parasitoid S. manilae
Pencarian inang yang sesuai merupakan hal penting bagi parasitoid agar mereka dapat meneruskan keturunannya. Kualitas inang seperti perbedaan spesies inang, stadia dan umur inang saat parasitisasi menentukan sebagian besar kebugaran dari keterununan parasitoid (Godfray 1994; Brodeur & Boivin 2004). Parasitoid menghabiskan sebagian besar hidupnya pada stadia dewasa untuk menemukan tempat inang berada (Jervis & Kidd 1997). Ada beberapa informasi yang digunakan dalam pencarian inang oleh parasitoid, yaitu stimulus dari habitat atau makanan; stimulus secara tidak langsung dari asosiasi kehadiran inang; stimulus yang dikeluarkan oleh inang itu sendiri (Godfray 1994). Kemampuan parasitoid dalam mencari inang yang sesuai untuk peletakkan telur sangat dibutuhkan dalam pengendalian hayati.
Larva S. litura merupakan inang dari S. manilae. Telur S. litura berbentuk bulat dan diletakkan secara berkelompok serta ditutupi oleh rambut halus. Kurniawati (1998) menyatakan bahwa imago S. manilae memarasit larva S.
berwarna coklat termasuk tipe obtekta, biasanya ditemukan di dalam tanah dengan diselimuti kokon. Imago ulat ini berwarna coklat muda dan aktif pada malam hari.
Kaitan Ketersediaan Inang dengan Kebugaran Parasitoid
Pada beberapa parasitoid synovigenic adanya ketiadaan inang yang cukup panjang dapat menstimulasi parasitoid untuk menyerap kembali telur-telur yang sudah diproduksi. Navasero dan Elzen (1992) menyatakan bahwa ketiadaan inang pada parasitoid synovigenic dapat menginduksi resorpsi telur yang berdampak pada pengurangan jumlah telur pada ovari. Adanya mekanisme resorpsi telur membuat imago betina dapat menggunakan energi dan material yang terkandung di dalam telur untuk mempertahankan dirinya dan meneruskan oogenesis sampai inang kembali tersedia (Jervis & Kidd 1997). Selama imago betina mengalami reabsorpsi telur, sementara itu dia tidak dapat melakukan oviposisi walaupun inang sudah tersedia (Jervis & Kidd 1997).
Heriyano (2000) menyatakan bahwa ketiadaan inang pada Eriborus
argenteopilosus dapat memengaruhi sisa telur dalam ovari dan jumlah telur di hari
ke-8 dan ke-9. Selain itu ketiadaan inang ternyata juga memengaruhi total produksi telur, dimana total produksi telur sangat tergantung terhadap frekuensi bertemu inang, jumlah inang yang diberikan, dan selang bertemu inang. Total produksi telur tergantung dari jumlah inang yang ditemui oleh parasitoid, hal ini menunjukkan bahwa parasitoid mengatur produksi telur ketika bertemu dengan inang (Drost & Carde 1992).
Ketiadaan inang dapat memengaruhi kesuburan ataupun tingkat parasitisasi parasitoid. Imago betina yang mengalami ketiadaan inang memiliki lama waktu kesuburan dan kemampuan menyerang inang lebih rendah dibandingkan imago betina yang mendapatkan inang sejak pertama kali muncul (Hougardy 2005). Berdasarkan penelitian yang telah dilakukan oleh Hegazi et al. (2007), Microplitis rufiventris yang mengalami ketiadaan inang lebih lama memiliki muatan telur di oviduk yang rendah. Selain itu Hegazi (2007) menyatakan bahwa Microplitis rufiventris yang diberikan inang larva Spodoptera
8
littoralis setelah ketiadaan inang selama 1, 3, 5, 6, 7, dan 8 mengalami puncak
parasitisasi pada saat pertama atau kedua kalinya setelah ketersediaan inang. Collin dan Dixon (1986) menyatakan bahwa, perpanjangan ketiadaan inang dapat meningkatkan superparasitisasi. Superparasitisasi dapat merugikan keturunan parasitoid yang selanjutnya, khususnya pada tingkat kebugarannya karena adanya kompetisi antar larva. Selain itu superparasitisasi juga dapat memengaruhi kesuksesan produksi keturunan (White & Andow 2008).
Ketiadaan inang ternyata dapat memengaruhi lama hidup parasitoid. Parasitoid Venturia canescens yang diberi pakan madu serta dipelihara pada suhu 15oC, 20oC, 25oC, dan 30oC memiliki lama hidup yang lebih panjang jika tidak diberi inang (Eliopoulos 2005). Hal yang sama juga terjadi pada Anaphes nitens, imago betina A. nitens yang diberi pakan madu dan tidak diberi inang memiliki lama hidup yang paling panjang dibandingkan dengan A. nitens yang diberi pakan madu dan inang (Carbone et al. 2008). Selain memengaruhi lama hidup, ketiadaan inang juga dapat memengaruhi tingkat penyebaran parasitoid. Pada Mastrus
ridibundus ketiadaan inang yang cukup lama yaitu empat hari menunjukkan
respon penyebaran yang sangat kuat (81.50 m2/h). M. ridibundus yang diberi inang secara berkesinambungan atau mengalami ketiadaan inang yang singkat (1-2 hari) menunjukkan tingkat penyebaran yang rendah ((1-2.07 m2/h) (Hougardy & Mills 2006).
Waktu dan Tempat
Penelitian dilakukan di Laboratorium Bioekologi Predator dan Parasitoid, Departemen Proteksi Tanaman, Fakultas Pertanian, Institut Pertanian Bogor. Waktu pelaksanaan penelitian dimulai sejak bulan Februari sampai September 2011.
Metode Penelitian Perbanyakan S. litura
Serangga inang S. litura diambil dari pertanaman talas di daerah Situ Gede, Dramaga, Bogor. Larva S. litura yang didapat dari lapangan selanjutnya dipelihara di laboratorium untuk perbanyakan. Larva S. litura instar 1 sampai instar 2 dipelihara dalam wadah plastik berdiameter 6.5 cm dan pada bagian atasnya dilapisi dengan tisu sebelum ditutup. Penggunaan tisu ini dilakukan untuk mencegah ulat-ulat tersebut keluar dari wadah plastik. Larva instar 3 sampai instar 4 dipelihara dalam wadah plastik berukuran 35 cm x 25 cm x 7 cm. Larva instar 5 dipelihara dalam wadah plastik yang berukuran sama, namun diberikan tambahan serbuk gergaji steril untuk tempat berpupa. Jika pupa telah terbentuk, maka pupa tersebut dipindahkan ke dalam wadah plastik bening berdiameter 11 cm yang dialasi kertas buram.
Imago yang telah keluar dari pupa diletakkan dalam wadah plastik berdiameter 25 cm yang setiap sisinya dilapisi dengan kertas buram, kecuali bagian dasar. Tujuan dari pelapisan menggunakan kertas buram ialah sebagai tempat peletakkan telur S. litura. Kertas buram yang terdapat telur kemudian digunting. Potongan kertas tersebut diletakkan dalam wadah plastik berdiameter 6.5 cm yang bagian atasnya dilapisi oleh tisu.
Larva S. litura diberi pakan buatan dalam bentuk potongan kotak berukuran sekitar 2 cm x 1 cm x 0.5 cm. Penggantian pakan dilakukan jika pakan kering atau sudah berubah warna menjadi sangat gelap. Imago S. litura diberi pakan larutan madu 20% yang diserapkan pada segumpal kapas dan diletakkan di dalam wadah plastik tempat imago S. litura dipelihara.
10
Perbanyakan Parasitoid S. manilae
Parasitoid didapat dengan cara mengambil S. litura instar 2 dari lapangan yang telah terparasit oleh S. manilae secara alamiah. Larva tersebut dipelihara dalam wadah plastik berukuran 35 cm x 25 cm x 7 cm hingga muncul pupa parasitoid yang biasanya berada di bagian posterior larva atau menempel pada tutup kasa wadah. Pupa tersebut selanjutnya diletakkan pada tabung reaksi pyrex berdiameter 1.5 cm hingga menjadi imago.
Larva S. litura instar 2 yang digunakan sebagai inang S. manilae merupakan larva hasil perbanyakan di laboratorium. Larva S. litura instar 2 dengan jumlah 30 ekor dan sehelai daun kedelai dimasukkan ke dalam tabung reaksi pyrex berdiameter 3 cm. Pada bagian dinding tabung reaksi diteteskan madu 20% sebagai pakan S. manilae. Selanjutnya imago S. manilae betina yang telah dikawinkan terlebih dahulu dimasukkan ke tabung reaksi yang telah berisi larva S. litura instar II. Pemaparan dilakukan selama 24 jam, kemudian larva yang telah dipaparkan tersebut dipelihara dalam cawan petri plastik berdiameter 9.5 cm beralaskan kertas buram dan diberi pakan buatan. Larva tersebut dipelihara sampai pupa S. manilae muncul. Larva S. litura yang tidak terparasit dikembalikan ke dalam kotak plastik pemeliharaan untuk diperbanyak kembali. Pemberian inang untuk imago S. manilae dilakukan setiap hari dengan tahapan yang sama.
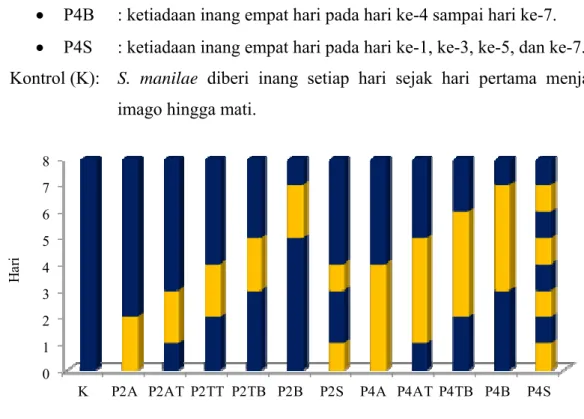
Rancangan Pola Ketiadaan Inang
Perlakuan yang digunakan pada penelitian ini ialah dua macam periode ketiadaan inang, yaitu ketiadaan inang dua hari dan ketiadaan inang empat hari. Lama ketiadaan inang dua hari diasumsikan mewakili S. manilae yang mengalami ketiadaan inang singkat di lapangan. Lama ketiadaan inang sehari tidak dipilih karena diduga kurang berpengaruh terhadap kebugaran S. manilae. Lama ketiadaan inang empat hari diasumsikan mewakili S. manilae yang mengalami ketiadaan inang cukup panjang di lapangan.
Total kombinasi ketiadaan inang dua hari ialah 21 kombinasi, sedangkan total kombinasi ketiadaan inang empat hari ialah 35 kombinasi. Total kombinasi tersebut didapat dari rumus kombinasi (Walpole 1995), yaitu:
Keterangan:
݊ = jumlah hari perlakuan (7 hari) ݎ = lama ketiadaan inang (2 dan 4 hari)
Dari kombinasi ketiadaan inang dua hari, diambil enam kombinasi untuk perlakuan, sedangkan dari kombinasi ketiadaan inang empat hari, diambil lima kombinasi untuk perlakuan. Kombinasi yang dipilih tersebut kemudian dijadikan sebagai pola ketiadaan inang. Pada akhirnya terdapat sebelas perlakuan dan satu kontrol dengan sepuluh ulangan untuk setiap perlakuan. Perlakuan tersebut dipilih karena diduga dapat memberikan pengaruh yang berbeda pada kebugaran
S. manilae. Perlakuan yang dipilih yaitu:
P2 : Ketiadaan inang selama 2 hari, yang dibagi lebih lanjut ke dalam perlakuan ketiadaan inang di awal (A), di awal hingga tengah (AT), di tengah (TT), di tengah hingga akhir (TB), di akhir/ belakang (B), dan seling (S):
• P2A : ketiadaan inang dua hari pada hari ke-1 dan ke-2. • P2AT : ketiadaan inang dua hari pada hari ke-2 dan ke-3. • P2TT : ketiadaan inang dua hari pada hari ke-3 dan ke-4. • P2TB : ketiadaan inang dua hari pada hari ke-4 dan ke-5. • P2B : ketiadaan inang dua hari pada hari ke-6 dan ke-7. • P2S : ketiadaan inang dua hari pada hari ke-1 dan ke-4.
P4 : Ketiadaan inang selama 4 hari, yang dibagi lebih lanjut ke dalam perlakuan ketiadaan inang di awal (A), di awal hingga tengah (AT), di tengah hingga akhir (TB), di akhir/ belakang (B), dan seling (S):
12 0 1 2 3 4 5 6 7 8
K P2A P2AT P2TT P2TB P2B P2S P4A P4AT P4TB P4B P4S
Ha
ri
Perlakuan
Ketiadaan inang Pemberian inang
• P4AT : ketiadaan inang empat hari pada hari ke-2 sampai hari ke-5. • P4TB : ketiadaan inang empat hari pada hari ke-3 sampai hari ke-6. • P4B : ketiadaan inang empat hari pada hari ke-4 sampai hari ke-7. • P4S : ketiadaan inang empat hari pada hari ke-1, ke-3, ke-5, dan ke-7. Kontrol (K): S. manilae diberi inang setiap hari sejak hari pertama menjadi
imago hingga mati.
Gambar 1 Perlakuan ketiadaan inang pada parasitoid S. manilae.
Pemberian Perlakuan
Imago betina S. manilae yang digunakan untuk semua perlakuan dalam satu ulangan memiliki umur yang seragam, yaitu S. manilae betina yang baru muncul dari pupa (berumur 1 hari). S. manilae betina tersebut dikawinkan terlebih dahulu, kemudian dimasukkan ke dalam tabung reaksi pyrex berdiameter 3 cm yang telah ditetesi larutan madu 20% pada bagian dindingnya. Pemberian inang berupa larva S. litura instar 2 sebanyak 30 ekor per tabung untuk satu S. manilae betina disesuaikan dengan Gambar 1. Pemaparan inang oleh S. manilae dilakukan selama 24 jam. Inang yang telah terpapar dibiarkan dalam tabung reaksi, untuk selanjutnya dilakukan pembedahan. S. manilae yang telah melakukan pemaparan
dipindahkan ke tabung reaksi lain untuk terus menjalani perlakuan yang telah ditentukan. S. manilae betina yang masih hidup setelah hari ke-8 akan diberikan inang terus hingga S. manilae tersebut mati.
Penghitungan Jumlah Telur
Penghitungan jumlah telur dilakukan dengan membedah inang yang telah dipaparkan oleh S. manilae. Pembedahan dilakukan untuk melihat telur atau larva
S. manilae di dalam larva inang. Pembedahan dilakukan di bawah mikroskop
stereo perbesaran 2.5 x 10 dengan menggunakan jarum bertangkai dan lampu. Larva yang akan dibedah ditaruh di atas preparat yang telah ditetesi cairan ringer. Cairan ini berfungsi untuk menjaga agar ketika hemolymph keluar tidak menyebar, sehingga obyek yang diamati (telur) tetap berada pada cairan ringer.
Pembedahan terhadap larva inang yang telah terpapar oleh S. manilae dilakukan sesegera mungkin, tidak lebih dari tiga hari setelah pemaparan perlakuan. Hal ini dilakukan karena umur telur S. manilae di dalam inang tidak lebih dari 24 jam. Jika terjadi superparasitisasi pada inang dan telur-telur tersebut menetas, maka akan terjadi kompetisi antar larva. Larva yang tidak dapat bertahan hidup akan mati dan jasadnya tereduksi, sehingga dapat luput dari penghitungan.
Pembedahan tidak hanya dilakukan pada inang yang telah dipaparkan, namun pembedahan juga dilakukan pada imago betina S. manilae yang telah mati untuk dihitung jumlah telur yang tersisa di dalam ovarinya. Proses pembedahan ovari S. manilae tidak jauh berbeda dengan pembedahan inang. Pada proses pembedahan ovari S. manilae jarum bertangkai yang digunakan ialah jarum bertangkai mikro dan mikroskop stereo dengan perbesaran 4 x 10.
Peubah amatan dalam penelitian ini terdiri dari: (1) sebaran jumlah telur per inang, (2) tingkat parasitisasi di hari ke-8, dihitung dengan rumus:
14
(3) jumlah telur yang diletakkan di hari ke-8; dilakukan pembedahan terhadap larva yang telah dipaparkan, kemudian jumlah telur yang terdapat di dalam tubuh larva dihitung, (4) jumlah telur yang diletakkan di hari pertama setelah ketiadaan inang; dilakukan pembedahan terhadap larva yang telah dipaparkan, kemudian dihitung jumlah telur yang terdapat di dalam tubuh larva, (5) sisa telur dalam ovari S. manilae; dilakukan pembedahan ovari S. manilae yang telah mati, (6) total produksi telur; jumlah telur yang diletakkan pada setiap inang ditambah telur yang tersisa dalam ovari, (7) lama hidup imago betina, dan (8) tingkat superparasitisasi di hari ke-8, dihitung dengan rumus:
Berdasarkan uji pendahuluan diketahui bahwa rata-rata lama hidup imago S. manilae betina ialah delapan hari. Oleh karena itu perlakuan pola ketiadaan inang diberikan sampai hari ke-7, kemudian di hari ke-8 semua perlakuan diberikan inang. Hari ke-8 dijadikan patokan pengamatan karena diasumsikan sebagai hari terakhir hidup S. manilae. Sehingga pada hari ke-8 dapat dilihat apakah perlakuan yang diberikan memberikan pengaruh terhadap tingkat parasitisasi (2), jumlah telur yang diletakkan (3), dan tingkat superparasitisasi (4).
Pada peubah amatan jumlah telur yang diletakkan di hari pertama setelah ketiadaan inang (4) P2S dan P4S memiliki perbedaan dari perlakuan lainnya. P2S dan P4S merupakan pola berseling. Hal tersebut mengakibatkan pertemuan inang dengan S. manilae yang telah mengalami ketiadaan inang terjadi lebih dari satu kali. Pada P2S hari pertama setelah ketiadaan inang terjadi pada hari 2 dan 5. Pada P4S hari pertama setelah ketiadaan inang terjadi pada hari 2, 4, ke-6, dan ke-8. Hari pertama setelah ketiadaan inang yang lebih dari satu kali tersebut ditambahkan dalam pengolahan data.
Tambahan data berupa jumlah telur pada hari pertama setelah ketiadaan inang dari P2S dan P4S diberi nama tersendiri. P2S2 merupakan jumlah telur yang diletakkan di hari ke-2 pada P2S, sedangkan P2S4 merupakan jumlah telur
yang diletakkan di hari ke-5 pada P2S. P4S2 merupakan jumlah telur yang diletakkan di hari ke-2 pada P4S, P4S4 merupakan jumlah telur yang diletakkan di hari ke-4 pada P4S, P4S6 merupakan jumlah telur yang diletakkan di hari ke-6 pada P4S, dan P4S8 merupakan jumlah telur yang diletakkan di hari ke-8 pada P24S. Data dari kontrol tidak dimasukkan karena S. manilae selalu diberi inang dari awal sampai akhir hidupnya.
Analisis Data
Perbedaan pengaruh pola ketiadaan inang pada seluruh peubah pengamatan dianalisis dengan sidik ragam. Pemeriksaan terhadap perlakuan-perlakuan yang berbeda dianalisis dengan uji Kisaran Ganda Duncan pada taraf nyata 5%. Kedua analisis di atas dilakukan menggunakan program SAS. Data dalam satuan persen ditransformasi dengan ArcSin , sedangkan data dalam satuan butir dan hari ditransformasi dengan Log(y).
HASIL DAN PEMBAHASAN
Hasil
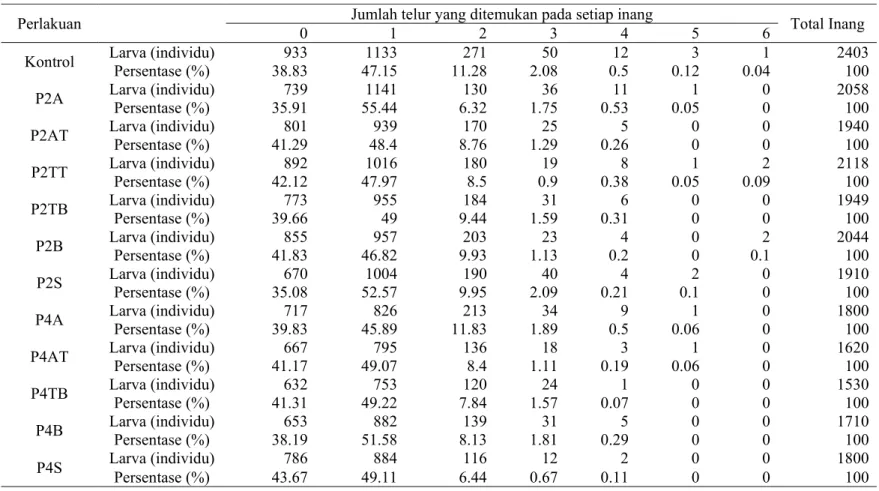
Sebaran Jumlah Telur S. manilae Per Larva Inang
Berdasarkan hasil penelitian diketahui bahwa rata-rata jumlah inang yang terparasit lebih dari 50%. Pada setiap perlakuan inang yang terparasit oleh
S. manilae berkisar 58%-63%, sedangkan inang yang tidak terparasit berkisar
37%-42% (Tabel 1). Inang yang terparasit satu butir telur memiliki persentase paling tinggi, yaitu berkisar 46%-55%, sedangkan inang yang terparasit lebih dari satu butir telur memiliki persentase yang rendah yaitu berkisar 0.04%-12%.
Ketiadaan inang selama dua hari di awal (P2A), di tengah (P2TB) dan seling (P2S) cenderung menyebabkan peningkatan persentase parasitisasi, sedangkan ketiadaan inang di tengah (P2AT dan P2TT) dan akhir (P2B) cenderung menyebabkan penurunan persentase parasitisasi (Tabel 1). Persentase parasitisasi pada perlakuan-perlakuan ketiadaan inang selama dua hari di tengah dan akhir jumlahnya lebih rendah dari kontrol. Ketiadaan inang selama empat hari di awal (P4A) dan di akhir (P4B) menyebabkan peningkatan persentase parasitisasi, sedangkan ketiadaan inang di tengah (P4AT dan P4TB) dan seling (P4S) justru menyebabkan penurunan persentase parasitisasi (Tabel 1).
S. manilae melakukan superparasitisasi dalam persentase yang lebih
rendah dibandingkan dengan parasitisasi soliter. Superparasitisasi terjadi ketika
S. manilae meletakkan lebih dari satu telur pada inang yang sama. Dalam
penelitian ini data superparasitisasi menunjukkan bahwa betina S. manilae bisa meletakkan dua hingga enam butir telur per larva inang. Kejadian superparasitisasi terbanyak ditemukan pada peletakkan dua butir telur per inang, sedangkan kejadian yang paling sedikit adalah enam butir telur per inang. Pada keadaan ideal seharusnya S. manilae tidak melakukan superparasitisasi bila inang yang tersedia mencukupi, namun berdasarkan Tabel 1 terlihat bahwa S. manilae tetap melakukan superparasitisasi meskipun jumlah inang yang dipaparkan mencukupi. Hal ini dapat dilihat pada kontrol, dari 30 inang yang dipaparkan setiap hari ternyata ada sekitar 39% inang yang tidak terparasit dan ditemukan kejadian superparasitisasi sekitar 14%.
Tabel 1 Sebaran jumlah telur S. manilae yang ditemukan pada setiap inang S. litura
Perlakuan Jumlah telur yang ditemukan pada setiap inang Total Inang
0 1 2 3 4 5 6
Kontrol Larva (individu) Persentase (%) 38.83933 47.151133 11.28 271 2.0850 0.512 0.123 0.041 2403100
P2A Larva (individu) Persentase (%) 35.91739 55.441141 6.32 130 1.7536 0.5311 0.051 00 2058100
P2AT Larva (individu) Persentase (%) 41.29801 48.4939 8.76 170 1.2925 0.265 00 00 1940100
P2TT Larva (individu) Persentase (%) 42.12892 47.971016 180 8.5 0.919 0.388 0.051 0.092 2118100
P2TB Larva (individu) Persentase (%) 39.66773 95549 9.44 184 1.5931 0.316 00 00 1949100
P2B Larva (individu) Persentase (%) 41.83855 46.82957 9.93 203 1.1323 0.24 00 0.12 2044100
P2S Larva (individu) 670 1004 190 40 4 2 0 1910
Persentase (%) 35.08 52.57 9.95 2.09 0.21 0.1 0 100
P4A Larva (individu) 717 826 213 34 9 1 0 1800
Persentase (%) 39.83 45.89 11.83 1.89 0.5 0.06 0 100
P4AT Larva (individu) Persentase (%) 41.17667 49.07795 136 8.4 1.1118 0.193 0.061 00 1620100
P4TB Larva (individu) Persentase (%) 41.31632 49.22753 7.84 120 1.5724 0.071 00 00 1530100
P4B Larva (individu) Persentase (%) 38.19653 51.58882 8.13 139 1.8131 0.295 00 00 1710100
P4S Larva (individu) 786 884 116 12 2 0 0 1800
Persentase (%) 43.67 49.11 6.44 0.67 0.11 0 0 100
18
Pengaruh Pola Ketiadaan Inang pada S. manilae Terhadap Tingkat Parasitisasi dan Produksi Telur
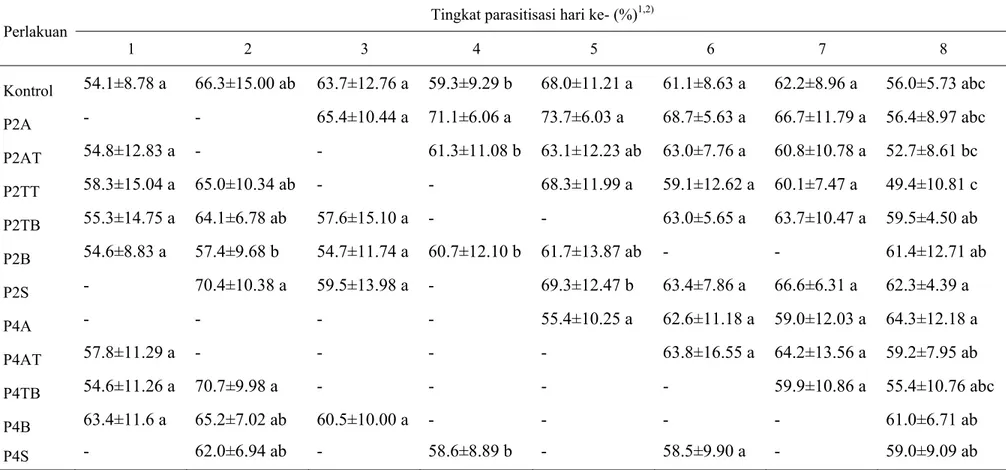
Berdasarkan hasil analisis sidik ragam ketiadaan inang di awal cenderung meningkatkan tingkat parasitisasi S. manilae pada hari ke-8, sedangkan ketiadaan inang dua hari di tengah cenderung menurunkan tingkat parasitisasi S. manilae pada hari ke-8 (Tabel 2). S. manilae yang mengalami ketiadaan inang cukup lama (4 hari) di awal atau di akhir hidupnya cenderung meletakkan telur lebih banyak pada hari ke-8 (Tabel 3). Pola ketiadaan inang tidak berpengaruh terhadap total telur hingga hari ke-8 (Gambar 4). Total telur sampai hari ke-8 menurun seiring bertambahnya lama ketiadaan inang.
Tingkat parasitisasi S. manilae per hari, dari hari pertama hingga hari ke-8 cenderung tidak berbeda nyata dengan kontrol pada semua pola ketiadaan inang, namun terlihat sedikit perbedaan pada tingkat parasitisasi S. manilae pada hari ke-8 (Tabel 2). Kisaran tingkat parasitisasi pada hari ke-8 lebih beragam dibandingkan dengan hari lainnya (Tabel 2). Ketiadaan inang selama dua hari di tengah (P2AT dan P2TT) cenderung menyebabkan penurunan tingkat parasitisasi, sedangkan tingkat parasitisasi pada hari ke-8 cenderung mengalami peningkatan pada ketiadaan inang di awal (P2A), di tengah menuju akhir (P2TB), di akhir (P2B), dan seling (P2S). Perbedaan persentase tingkat parasitisasi yang sangat mencolok terlihat pada P2S dengan P2TT (Tabel 2). Ketiadaan inang selama empat hari di awal (P4A) dan di akhir (P4B) cenderung menyebabkan peningkatan parasitisasi pada hari ke-8, sedangkan ketiadaan inang di tengah (P4AT dan P4TB) dan seling (P4S) cenderung menyebabkan penurunan tingkat parasitisasi pada hari ke-8 (Tabel 2). Persentase parasitisasi pada semua perlakuan ketiadaan inang tidak berbeda nyata dengan kontrol.
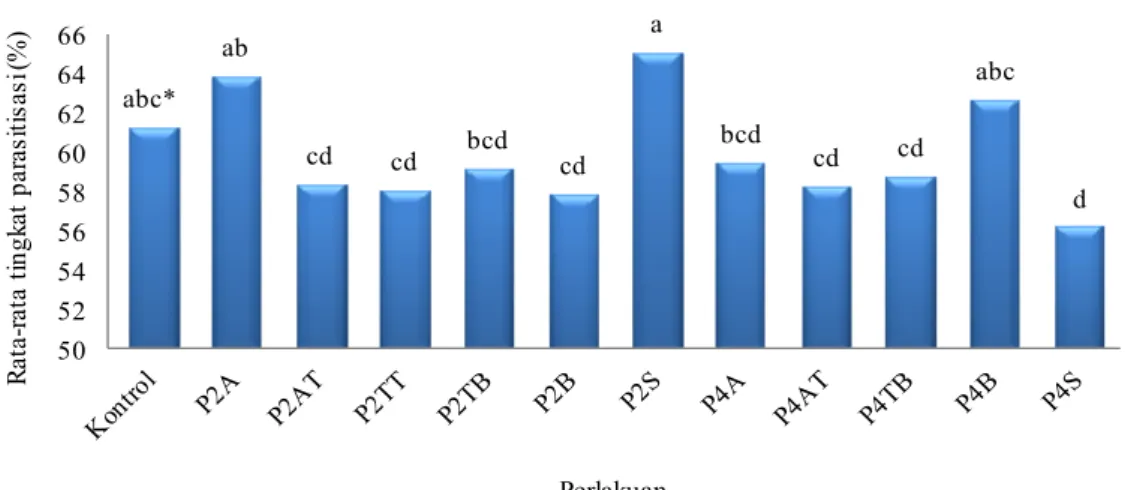
Kemampuan parasitisasi harian S. manilae yang diberi perlakuan tidak berbeda nyata dengan kontrol (61.4%), kecuali P4S (56.3%). S. manilae pada P4S memiliki kemampuan parasitisasi harian terendah dibandingkan dengan perlakuan lainnya (Gambar 2). Lamanya ketiadaan inang ternyata berpengaruh terhadap rata-rata tingkat parasitisasi harian, karena walaupun keduanya merupakan pola berseling namun P2S berbeda nyata dengan P4S. Ketiadaan inang selama 2 hari menyebabkan peningkatan persentase parasitisasi harian pada perlakuan pola awal
abc* ab cd cd bcd cd a bcd cd cd abc d 50 52 54 56 58 60 62 64 66 R ata -r ata t in gk at pa ra si tis as i ( % ) Perlakuan
(P2A) dan seling (P2S), sedangkan pada perlakuan dengan pola tengah (P2AT, P2TT, P2TB) dan belakang (P2B) persentase parasitisasi harian mengalami penurunan (Gambar 2). Perlakuan ketiadaan inang selama 4 hari cenderung menyebabkan penurunan tingkat parasitisasi harian untuk pola tengah (P4AT dan P4TB), dan seling (P4S). Peningkatan persentase parasitisasi harian hanya terjadi pada pola belakang (P4B).
Gambar 2 Rata-rata tingkat parasitisasi S. manilae per hari.
*Huruf yang sama di atas kelompok diagram tidak berbeda nyata menurut uji Duncan pada α = 5%. A: ketiadaan inang di awal; AT: ketiadaan inang di awal hingga tengah; TT: ketiadaan inang di tengah; TB: ketiadaan inang di tengah hingga akhir; B: ketiadaan inang di akhir; S: ketiadaan inang berseling; Angka: jumlah hari ketiadaan inang.
Jumlah telur yang diletakkan oleh S. manilae per hari dari hari pertama hingga hari ke-8 tidak berbeda nyata dengan kontrol pada semua pola ketiadaan inang, namun terlihat sedikit perbedaan pada jumlah telur yang diletakkan
S. manilae pada hari ke-8 (Tabel 3). Sama seperti peubah amatan tingkat
parasitisasi pada hari ke-8, kisaran jumlah telur pada hari ke-8 terlihat lebih beragam dibandingkan dengan hari lainnya (Tabel 3). Ketiadaan inang selama dua hari di tengah (P2AT dan P2TT) cenderung menyebabkan penurunan jumlah telur di hari ke-8, sedangkan jumlah telur di hari ke-8 cenderung mengalami peningkatan pada ketiadaan inang di awal (P2A), di tengah menuju akhir (P2TB), di akhir (P2B), dan seling (P2S) (Tabel 3). Perbedaan yang nyata pada persentase
20
jumlah telur pada hari ke-8 terlihat pada P2S dengan P2TT. Ketiadaan inang selama empat hari di awal (P4A) dan di akhir (P4B) cenderung menyebabkan peningkatan jumlah telur pada hari ke-8, sedangkan ketiadaan inang di tengah (P4AT dan P4TB) dan seling (P4S) cenderung menyebabkan penurunan jumlah telur pada hari ke-8 (Tabel 3). Perbedaan nyata persentase jumlah telur hari ke-8 sangat terlihat pada P4S dan P4TB dengan P4B. Persentase jumlah telur pada P2S, P4A, P4AT, dan P4B berbeda nyata dengan kontrol.
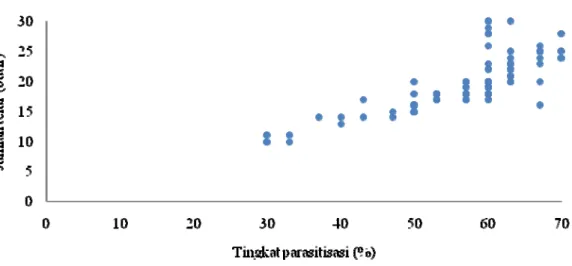
Gambar 3 menunjukkan korelasi antara tingkat parasitisasi hari ke-8 dengan jumlah telur yang diletakkan pada hari ke-8. Terlihat bahwa terdapat korelasi positif diantara keduanya. Hal ini dapat dilihat dari data berupa titik-titik bergerombol mengikuti sebuah garis lurus dengan kemiringan positif. Koefisien korelasi yang didapat ialah 0.806 menunjukkan adanya hubungan keeratan yang tinggi antara tingkat parasitisasi dengan jumlah telur yang diletakkan pada hari ke-8.
Gambar 3 Diagram pencar korelasi antara tingkat parasitisasi di hari ke-8 dengan jumlah telur yang diletakkan di hari ke-8 pada S. manilae yang telah diberikan perlakuan.
Tabel 2 Pengaruh pola ketiadaan inang pada S. manilae terhadap tingkat parasitisasi per hari hingga hari ke-8
Perlakuan Tingkat parasitisasi hari ke- (%)
1,2) 1 2 3 4 5 6 7 8 Kontrol 54.1±8.78 a 66.3±15.00 ab 63.7±12.76 a 59.3±9.29 b 68.0±11.21 a 61.1±8.63 a 62.2±8.96 a 56.0±5.73 abc P2A - - 65.4±10.44 a 71.1±6.06 a 73.7±6.03 a 68.7±5.63 a 66.7±11.79 a 56.4±8.97 abc P2AT 54.8±12.83 a - - 61.3±11.08 b 63.1±12.23 ab 63.0±7.76 a 60.8±10.78 a 52.7±8.61 bc P2TT 58.3±15.04 a 65.0±10.34 ab - - 68.3±11.99 a 59.1±12.62 a 60.1±7.47 a 49.4±10.81 c P2TB 55.3±14.75 a 64.1±6.78 ab 57.6±15.10 a - - 63.0±5.65 a 63.7±10.47 a 59.5±4.50 ab P2B 54.6±8.83 a 57.4±9.68 b 54.7±11.74 a 60.7±12.10 b 61.7±13.87 ab - - 61.4±12.71 ab P2S - 70.4±10.38 a 59.5±13.98 a - 69.3±12.47 b 63.4±7.86 a 66.6±6.31 a 62.3±4.39 a P4A - - - - 55.4±10.25 a 62.6±11.18 a 59.0±12.03 a 64.3±12.18 a P4AT 57.8±11.29 a - - - - 63.8±16.55 a 64.2±13.56 a 59.2±7.95 ab P4TB 54.6±11.26 a 70.7±9.98 a - - - - 59.9±10.86 a 55.4±10.76 abc P4B 63.4±11.6 a 65.2±7.02 ab 60.5±10.00 a - - - - 61.0±6.71 ab P4S - 62.0±6.94 ab - 58.6±8.89 b - 58.5±9.90 a - 59.0±9.09 ab
1) Angka selajur yang diikuti dengan huruf yang samatidak berbeda nyata menurut uji Duncan pada α=5% 2) Untuk keperluan sidik ragam, data ditransformasi dengan Arcsin ( )
21
Tabel 3 Pengaruh pola ketiadaan inang pada S. manilae terhadap jumlah telur yang diletakkan per hari hingga hari ke-8
Perlakuan Jumlah telur hari ke- (%)
1,2) 1 2 3 4 5 6 7 8 Kontrol 19.1±5.83 a 28.0±7.28 ab 27.9±12.02 a 23.8±3.56 ab 27.0±7.38 a 24.8±8.78 a 22.2±3.76 a 16.8±1.61 de P2A - - 27.1±4.28 a 26.8±3.76 a 25.2±7.08 a 24.0±4.12 a 20.8±1.09 a 18.1±3.60 cde P2AT 19.7±6.21 a - - 21.6±4.44 ab 23.4±9.91 a 23.4±7.02 a 20.0±4.69 a 17.4±2.01 cde P2TT 20.2±4.84 a 21.4±6.06 ab - - 24.6±8.47 a 20.0±5.29 a 19.2±2.58 a 16.3±3.56 e P2TB 21.6±9.04 a 21.4±3.28 ab 26.4±5.85 a - - 21.4±2.07 a 20.6±4.72 a 20.4± 3.71 cd P2B 19.9±4.33 a 19.6±3.91 b 22.4±6.34 a 25.8±5.35 ab 24.6±10.92 a - - 20.3±4.62 cd P2S - 26.6±5.22 a 22.1±5.66 a - 29.6±7.16 a 25.8±1.30 a 21.8±2.16 a 21.2±2.34 abc P4A - - - - 19.4±8.96 a 23.6±6.84 a 24.8±8.70 a 25.4±6.05 ab P4AT 19.3±4.37 a - - - - 22.6±7.63 a 26.0±12.46 a 20.9±3.60 bc P4TB 18.3±4.27 a 27.0±5.04 a - - - - 19.8±5.11 a 19.1±4.09 cde P4B 21.7±5.37 a 24.2±5.80 ab 24.0±5.57 a - - - - 25.6± 5.03 a P4S - 22.0±3.16 ab - 21.4±2.19 b - 20.0±2.54 a - 18.7±2.79 cde
1) Angka selajur yang diikuti dengan huruf yang samatidak berbeda nyata menurut uji Duncan pada α=5% 2) Untuk keperluan sidik ragam, data ditransformasi dengan Log (y)
a* b b b b b b c c c c c 0 20 40 60 80 100 120 140 160 180 200
Kontrol P2A P2AT P2TT P2TB P2B P2S P4A P4AT P4TB P4B P4S
Ju m la h t el ur (b ut ir) Perlakuan
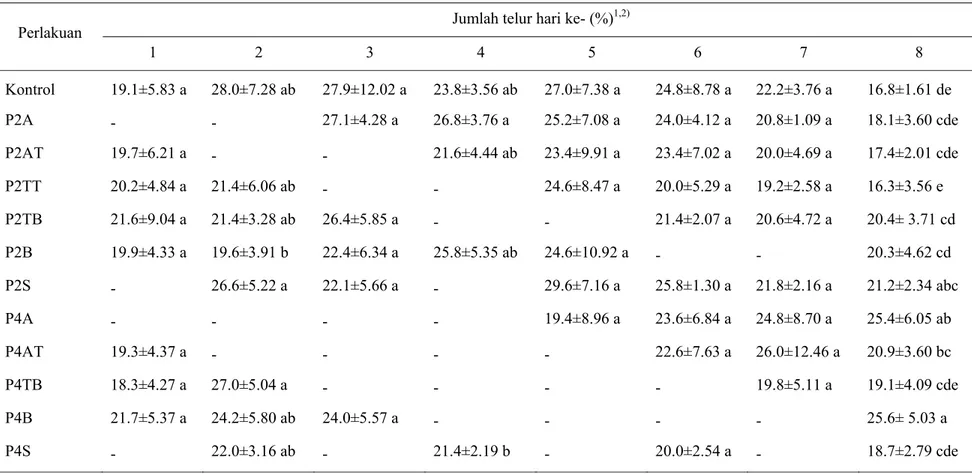
Gambar 4 menunjukkan bahwa ketiadaan inang berseling, di awal, di tengah, dan di akhir hidup S. manilae ternyata tidak berpengaruh terhadap jumlah telur yang diletakkan hingga hari ke-8. Jumlah telur yang diletakkan hingga hari ke-8 ternyata lebih dipengaruhi oleh lama ketiadaan inang. Semakin lama imago
S. manilae tidak mendapatkan inang maka jumlah telur yang diletakkan hingga
hari ke-8 semakin menurun. Dapat dilihat pada Gambar 4 bahwa jumlah telur hingga hari ke-8 tertinggi terdapat pada kontrol, diikuti oleh pola-pola ketiadaan inang dua hari, dan yang terendah ialah pola-pola ketiadaan inang empat hari. Total telur sampai hari ke-8 pada kontrol berbeda nyata dengan pola lainnya.
Gambar 4 Total telur yang diletakkan hingga hari ke-8 oleh S. manilae.
*Huruf yang sama di atas kelompok diagram tidak berbeda nyata menurut uji Duncan pada α = 5%. A: ketiadaan inang di awal; AT: ketiadaan inang di awal hingga tengah; TT: ketiadaan inang di tengah; TB: ketiadaan inang di tengah hingga akhir; B: ketiadaan inang di akhir; S: ketiadaan inang berseling; Angka: jumlah hari ketiadaan inang.
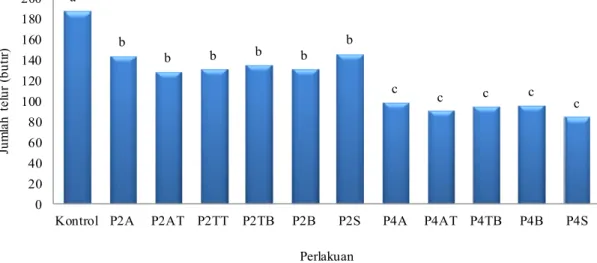
Perlakuan yang diberikan pada S. manilae dapat memengaruhi jumlah telur pada hari pertama setelah ketiadan inang. S. manilae yang mengalami ketiadaan inang selama dua hari di awal (P2A), di tengah (P2TT), dan seling (P2S) cenderung meletakkan jumlah telur yang lebih banyak pada hari pertama setelah ketiadaan inang, sedangkan ketiadaan inang selama dua hari di tengah (P2AT dan P2TB) dan di akhir (P2B) menyebabkan S. manilae cenderung
24 ab* bcd ab bcd cd ab ab bcd abc a abc bcd cd bcd d 0 5 10 15 20 25 30 35
P2A P2AT P2TT P2TB P2B P2S2 P2S5 P4A P4AT P4TB P4B P4S2 P4S4 P4S6 P4S8
Ju m la h t el ur (b ut ir) Perlakuan
meletakkan telur yang lebih sedikit pada hari pertama setelah ketiadaan inang (Gambar 5). Hanya S. manilae pada P2B yang memiliki jumlah telur pada hari pertama berbeda nyata dengan P2A. Hal yang berbeda terjadi pada S. manilae yang mengalami ketiadaan inang selama empat hari. S. manilae yang mengalami ketiadaan inang empat hari di awal (P4A), di akhir (P4B), dan seling (P4S) cenderung meletakkan telur yang lebih sedikit dibandingkan dengan S. manilae yang mengalami ketiadaan inang selama empat hari di tengah (P4AT dan P4TB) (Gambar 5).
Jumlah telur S. manilae pada hari pertama setelah ketiadaan inang berkisar 18 butir hingga 31 butir (Gambar 5). Jarak antara 18-31 butir terlihat cukup jauh, namun dari Gambar 5 terlihat bahwa data banyak berkumpul pada kisaran 25 butir. Telur yang diletakkan pada hari pertama setelah ketiadaan inang oleh S. manilae pada P4TB merupakan jumlah telur paling banyak (31 butir) dibandingkan dengan pola yang lain, sedangkan S. manilae pada P4S8 memiliki jumlah telur yang diletakkan pada hari pertama setelah ketiadan inang paling sedikit (18.7 butir) dibandingkan dengan pola yang lain.
Gambar 5 Jumlah telur yang diletakkan S. manilae pada hari pertama setelah ketiadaan inang. *Huruf yang sama di atas kelompok diagram tidak berbeda nyata menurut uji
Duncan pada α = 5%. A: ketiadaan inang di awal; AT: ketiadaan inang di awal hingga tengah; TT: ketiadaan inang di tengah; TB: ketiadaan inang di tengah hingga akhir; B: ketiadaan inang di akhir; S: ketiadaan inang berseling; Angka dibelakang P: jumlah hari ketiadaan inang; Angka dibelakang S: hari ke-.
bcde* e bcde cde bc bcde bcde a b bcde bcd de 0 1 2 3 4 5 6
Kontrol P2A P2AT P2TT P2TB P2B P2S P4A P4AT P4TB P4B P4S
Su pe rp ar as iti sas i ( in an g) Perlakuan
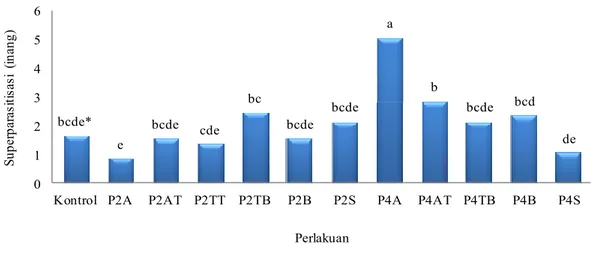
Gambar 6 menunjukkan bahwa tingkat superparasitisasi pada hari ke-8 cukup rendah, dari 30 ekor inang terdapat 1-5 inang yang mengalami superparasitisasi. S. manilae yang mengalami ketiadaan inang di awal cenderung melakukan superparasitisasi yang rendah pada hari ke-8. Hal sebaliknya terjadi pada S. manilae yang mengalami ketiadaan inang selama empat hari. Pola ketiadaan inang di awal yang cukup lama (4 hari) dapat menyebabkan S. manilae melakukan superparasitisasi yang tinggi pada hari ke-8. Terlihat hanya P4A yang memiliki superparasitisasi paling tinggi dibandingkan dengan perlakuan lainnya, kemudian diikuti oleh P4AT (Gambar 6). S. manilae yang sebelumnya telah bertemu dengan inang, kemudian mengalami ketiadaan inang selama empat hari memiliki tingkat superparasitisasi yang tidak berbeda nyata dengan kontrol. Lamanya ketiadaan inang ternyata memengaruhi superparasitisasi pada hari ke-8. Ketika S. manilae mengalami ketiadaan inang yang lama (4 hari) di awal ternyata superparasitisasi yang terjadi pada hari ke-8 jauh lebih tinggi dibandingkan dengan ketiadaan inang pada S. manilae yang mengalami ketiadaan inang dua hari di awal.
Gambar 6 Superparasitisasi pada hari ke-8 oleh S. manilae yang telah diberikan perlakuan. *Huruf yang sama di atas kelompok diagram tidak berbeda nyata menurut uji
Duncan pada α = 5%. A: ketiadaan inang di awal; AT: ketiadaan inang di awal hingga tengah; TT: ketiadaan inang di tengah; TB: ketiadaan inang di tengah hingga akhir; B: ketiadaan inang di akhir; S: ketiadaan inang berseling; Angka: jumlah hari ketiadaan inang.
26
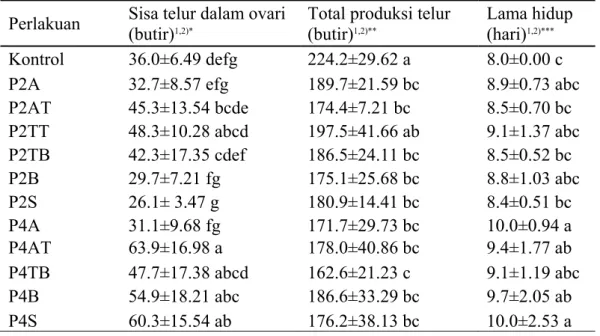
Pengaruh Pola Ketiadaan Inang pada S. manilae Terhadap Sisa Telur, Total Telur, dan Lama Hidup
Pola ketiadaan inang tidak begitu berpengaruh terhadap total produksi telur dan lama hidup imago S. manilae. S. manilae yang mengalami ketiadaan inang memiliki total telur yang lebih rendah dibandingkan dengan S. manilae yang selalu mendapatkan inang. Lama hidup imago S. manilae lebih dipengaruhi oleh lamanya ketiadaan inang, sedangkan sisa telur dalam ovari S. manilae dipengaruhi baik oleh pola ketiadaan inang maupun lama ketiadaan inang. Pola ketiadaan inang selama dua hari di tengah (P2AT dan P2TT) cenderung menyebabkan sisa telur dalam ovari S. manilae menjadi lebih banyak, sedangkan ketiadaan inang di awal (P2A), di akhir (P2TB dan P2B) dan seling (P2S) cenderung menurunkan sisa telur dalam ovari S. manilae (Tabel 4). Hal yang sama terjadi pada pola-pola ketiadaan inang selama empat hari, kecuali pada pola seling (P4S). Semakin lama S. manilae mengalami ketiadaan inang maka sisa telur dalam ovari S. manilae cenderung lebih banyak. Hal ini dapat dilihat dari ketiadaan inang dengan pola seling selama empat hari ternyata dapat menyebabkan peningkatan sisa telur dalam ovari, namun bila ketiadaan inang terjadi selama dua hari maka sisa telur dalam ovari akan menurun. Sisa telur dalam ovari S. manilae pada P2S memiliki jumlah yang paling sedikit dibandingkan dengan perlakuan lain, yaitu 26.1 butir telur.
Hasil penelitian menunjukan bahwa total telur dari semua perlakuan tidak berbeda nyata satu dengan lainnya, kecuali P2TT dengan P4TB (Tabel 4). Akan tetapi ketiadaan inang ternyata menurunkan total produksi telur S. manilae pada semua perlakuan. Secara statistik, hanya P2TT yang tidak berbeda nyata dengan kontrol. Bila dibandingkan dengan jumlah telur yang diletakkan hingga hari ke-8 terlihat bahwa lama ketiadaan inang memengaruhi jumlah telur yang diletakkan hingga hari ke-8, tetapi jumlah total produksi telur baik pada ketiadaan inang selama dua hari dan empat hari cenderung sama. Hal tersebut membuktikan bahwa lama ketiadaan inang hingga empat hari untuk semua pola tidak berpengaruh pada total produksi telur.
Tabel 4 Pengaruh pola ketiadaan inang pada S. manilae terhadap sisa telur, total produksi telur, dan lama hidup
Perlakuan Sisa telur dalam ovari (butir)1,2)*
Total produksi telur (butir)1,2)**
Lama hidup (hari)1,2)***
Kontrol 36.0±6.49 defg 224.2±29.62 a 8.0±0.00 c
P2A 32.7±8.57 efg 189.7±21.59 bc 8.9±0.73 abc
P2AT 45.3±13.54 bcde 174.4±7.21 bc 8.5±0.70 bc P2TT 48.3±10.28 abcd 197.5±41.66 ab 9.1±1.37 abc P2TB 42.3±17.35 cdef 186.5±24.11 bc 8.5±0.52 bc P2B 29.7±7.21 fg 175.1±25.68 bc 8.8±1.03 abc P2S 26.1± 3.47 g 180.9±14.41 bc 8.4±0.51 bc P4A 31.1±9.68 fg 171.7±29.73 bc 10.0±0.94 a P4AT 63.9±16.98 a 178.0±40.86 bc 9.4±1.77 ab P4TB 47.7±17.38 abcd 162.6±21.23 c 9.1±1.19 abc P4B 54.9±18.21 abc 186.6±33.29 bc 9.7±2.05 ab P4S 60.3±15.54 ab 176.2±38.13 bc 10.0±2.53 a
1) Angka selajur yang diikuti dengan huruf yang samatidak berbeda nyata menurut uji Duncan pada α=5% 2) Untuk keperluan sidik ragam, data ditransformasi dengan Log (y) *Koef. keragaman=8.84 **Koef.
keragaman=2.95 ***Koef. keragaman=5.89
Pola ketiadaan inang tidak berpengaruh terhadap lama hidup imago
S. manilae, sedangkan lama ketiadaan inang ternyata berpengaruh terhadap lama
hidup imago S. manilae. Pemberian inang secara terus menerus menyebabkan imago S. manilae memiliki umur yang paling singkat, yaitu delapan hari, sedangkan rata-rata lama hidup imago S. manilae terpanjang dalam penelitian ini adalah 10 hari (Tabel 4). Semakin lama imago S. manilae tidak mendapatkan inang maka semakin panjang lama hidupnya. Pada Tabel 4 terlihat bahwa pola ketiadaan inang selama dua hari cenderung memiliki lama hidup yang lebih singkat dibandingkan dengan pola ketiadaan inang empat hari.
Korelasi positif terjadi antara peubah amatan total produksi telur dan lama hidup. Dari Gambar 7 dapat dilihat bahwa beberapa titik begerombol mengikuti sebuah garis lurus dengan kemiringan positif, namun cukup banyak titik yang memencar atau menjauhi titik-titik dari garis lurus. Koefisien korelasi yang didapat ialah 0.494 menunjukkan adanya hubungan keeratan yang rendah antara total produksi telur dan lama hidup imago.
28
Gambar 7 Diagram pencar korelasi antara lama hidup dengan total produksi telur pada imago betina S. manilae yang telah diberikan perlakuan.
Pembahasan
Penelitian ini menunjukkan bahwa parasitisasi yang dilakukan oleh
S. manilae yang diberi perlakuan tidak selalu efisien. Parasitisasi soliter (satu butir
telur parasitoid per inang) memiliki persentase yang paling tinggi, namun persentase inang yang tidak terparasit juga cukup tinggi, selain itu superparasitisasi selalu terjadi pada semua perlakuan. Parasitisasi soliter pada penelitian ini tidak berbeda jauh dengan penelitian Ratna (2008), yaitu parasitisasi satu butir telur per inang S. manilae pada pemaparan inang S. litura instar II sebanyak 30 ekor yaitu sebesar 44.7%. Terlihat bahwa S. manilae yang mengalami ketiadaan inang ataupun selalu mendapatkan inang sama-sama memiliki kisaran parasitisasi soliter yang tidak berbeda jauh.
Fenlon (2009) menyatakan bahwa superparasitisasi akan menurun seiring dengan meningkatnya kepadatan inang. Ketika kepadatan inang sudah cukup memenuhi kebutuhan S. maniale untuk melakukan parasitisasi seharusnya kejadian superparsitisasi menjadi rendah atau bahkan tidak terjadi. Kepadatan inang yang diberikan untuk parasitisasi S. manilae pada penelitian ini sudah sesuai. Hal ini dapat terlihat dari cukup tingginya persentase inang yang tidak terparasit. Walaupun persentase inang yang tidak terparasit cukup tinggi namun superparasitisasi tetap terjadi dalam persentase yang rendah.
Kekeliruan S. manilae dalam membedakan antara inang yang sudah terparasit atau belum merupakan salah satu penyebab superparasitisasi serta tingginya persentase inang yang tidak terparasit. Van Giessen (dalam Ratna 2008) menyatakan bahwa diskriminasi inang oleh imago parasitoid betina sangat bergantung pada pengalaman pemilihan inang dan lingkungan. Selain itu, tempat ketika pemaparan dilakukan berupa tabung reaksi pyrex berdiameter 3 cm, terdapat kemungkinan inang melekat pada lapisan bawah daun kedelai yang berhadapan langsung dengan dinding tabung. Walaupun ukuran S. manilae kecil, namun kondisi tersebut tetap dapat membatasi ruang gerak parasitoid ini dalam melakukan parasitisasi. Hal ini membuat S. manilae mengalami kesulitan dalam menemukan inang yang berada di bagian bawah daun kedelai.
Peletakan dua butir telur per inang merupakan superparasitisasi yang paling banyak ditemukan. Kejadian superparasitisasi semakin menurun seiring bertambahnya jumlah telur yang ditemukan dalam tubuh inang. Hal ini kemungkinan merupakan strategi S. manilae untuk menghindari kompetisi antar larva yang terjadi di dalam tubuh inang. Adanya kompetisi antar larva S. manilae di dalam tubuh inang dapat menurunkan kebugaran S. manilae nantinya.
Pengaruh Ketiadaan Inang pada S. manilae Terhadap Tingkat Parasitisasi dan Produksi Telur
Secara umum tingkat parasitisasi S. manilae yang mengalami ketiadaan inang memiliki persentase yang tidak berbeda nyata dengan S. manilae yang selalu mendapatkan inang. Hal ini menunjukkan bahwa S. manilae yang mengalami ketiadaan inang sampai empat hari memiliki kemampuan memarasit yang sama dengan S. manilae yang selalu mendapatkan inang selama hidupnya. Jika dilihat dari pola ketiadaan inang, maka akan terlihat beberapa kecenderungan. Ketiadaan inang selama dua hari dan empat hari di awal menyebabkan S. manilae mengalami peningkatan parasitisasi pada hari ke-8. Ketika S. manilae mengalami ketiadaan inang di tengah menuju akhir hidupnya, tingkat parasitisasi pada hari ke-8 mengalami penurunan. Tingkat parasitisasi mengalami peningkatan kembali ketika S. manilae mengalami ketiadaan inang secara berseling dan ketiadaan inang di akhir hidupnya. Adanya keeratan/ korelasi positif yang tinggi diantara tingkat
30
parasitisasi pada hari ke-8 dan jumlah telur yang diletakkan pada hari ke-8 menyebabkan kecenderungan yang terjadi pada jumlah telur yang diletakkan pada hari ke-8 relatif sama dengan tingkat parasitisasi pada hari ke-8. Namun pada peubah amatan jumlah telur di hari ke-8 lebih terlihat jelas bahwa ketiadaan inang yang cukup lama (4 hari) di awal dan akhir menyebabkan jumlah telur yang diletakkan di hari ke-8 menjadi lebih banyak. Hal tersebut kemungkinan terjadi karena S. manilae yang sudah pernah bertemu dengan inang sebelumnya baru bertemu kembali dengan inang ketika pada hari ke-8. Perjumpaan kembali
S. manilae dengan inangnya merangsang S. manilae untuk segera meletakkan
telur yang telah di produksi. Hal ini sesuai dengan pernyataan Hegazi (2007), yaitu Microplitis rufiventris yang diberikan inang larva Spodoptera littoralis
setelah ketiadaan inang selama 1, 3, 5, 6, 7, dan 8 mengalami puncak parasitisasi pada saat pertama atau kedua kalinya setelah ketersediaan inang. S. manilae yang mengalami ketiadaan inang cukup lama di awal hidupnya membuat parasitoid ini kurang memiliki pengalaman dalam melakukan parasitisasi. Drost dan Carde (1992) menyebutkan bahwa ketiadaan inang parasitoid dapat menyebabkan rendahnya tingkat parasitisasi karena belum adanya pengalaman oviposisi. S. manilae yang mengalami ketiadaan inang cukup lama di awal hidupnya
memiliki kesempatan mendapatkan pengalaman oviposisi sebelum mencapai hari ke-8. Adanya kesempatan tersebut merangsang parasitoid ini untuk terus meletakkan telur, sehingga jumlah telur yang diletakkan pada hari ke-8 mengalami peningkatan.
S. manilae yang selalu mendapatkan inang sepanjang hidupnya maupun
yang mengalami ketiadaan inang memiliki rata-rata parasitisasi harian yang tidak terlalu berbeda. Namun adanya pola berseling yang cukup lama membuat rata-rata tingkat parasitisasi harian S. manilae menjadi rendah. Adanya pola seperti ini kemungkinan membentuk perilaku parasitisasi yang sama dari awal sampai akhir hidup imago S. manilae. Ketika S. manilae baru pertama kali menemukan inang tingkat parasitisasinya belum terlalu tinggi karena belum memiliki pengalaman oviposisi, kemudian keesokan harinya S. manilae kembali tidak bertemu inang,
hal ini terus terjadi hingga hari ke-8. Hal ini yang menyebabkan rata-rata tingkat parasitisasi jauh lebih rendah dibandingkan dengan kontrol.
Pola ketiadaan inang tidak berpengaruh terhadap total telur sampai hari ke-8, namun lamanya ketiadaan inang dapat memengaruhi total telur yang diletakkan hingga hari ke-8. S. manilae yang mengalami ketiadaan inang dua hari lebih banyak bertemu inang dibandingkan dengan S. manilae yang mengalami ketiadaan inang empat hari. Semakin lama ketiadaan inang, frekuensi S. manilae bertemu dengan inang semakin sedikit, sehingga total telur yang diletakkan hingga hari ke-8 juga lebih sedikit. Drost dan Carde (1992) menyatakan bahwa total produksi telur tergantung dari jumlah inang yang ditemui oleh parasitoid.
Jika dibandingkan dengan penelitian Ratna (2008) diketahui bahwa jumlah telur per hari yang diletakkan S. manilae pada larva S. litura ialah 20.8 butir, maka terlihat bahwa S. manilae yang mengalami ketiadaan inang cenderung meletakkan telur yang lebih banyak pada hari pertama setelah ketiadaan inang. Hal tersebut sesuai dengan yang dijabarkan oleh Hougardy (2005) bahwa puncak peletakan telur M. ridibundus terjadi pada hari pertama ketersediaan inang. Selain itu, Hegazi (2007) menyatakan bahwa Microplitis rufiventris yang diberikan inang
larva Spodoptera littoralis setelah ketiadaan inang selama 1, 3, 5, 6, 7, dan 8 mengalami puncak parasitisasi pada saat pertama atau kedua kalinya setelah ketersediaan inang.
Adanya pola berseling yang cukup panjang cenderung menurunkan peletakkan jumlah telur pada hari pertama setelah ketiadaan inang, khususnya pada hari terakhir (hari ke-8). Selain akibat adanya perubahan perilaku akibat pola ketiadaan inang yang berseling-seling, rendahnya telur yang diletakkan kemungkinan berkaitan dengan umur parasitoid yang sudah tua. Ratna (2008) menyatakan bahwa secara umum ada kecenderungan bahwa induk parasitoid berumur muda lebih banyak menghasilkan keturunan dan perlahan-lahan menurun seiring bertambahnya umur.
Superparasitisasi dapat dikatakan merugikan, karena dapat memengaruhi proporsi kesuksesan produksi keturunan (White & Andow 2008). Superparasitisasi yang terjadi pada hari ke-8 cenderung rendah. Ratna (2008)
32
melaporkan bahwa tingkat superparsitisasi parasitoid S. manilae hasil pemaparan dengan larva instar II S. litura sebanyak 30 ekor adalah 19.6% (5-6 inang). Rendahnya superparasitisasi pada hari ke-8 mungkin dikarenakan umur parasitoid yang sudah cukup tua sehingga tingkat parasitisasinya cenderung menurun dan sudah cukup banyak telur yang diletakkan pada inang sebelumnya. Selain itu pengalaman S. manilae dalam membedakan inang yang sudah terparasit sudah cukup baik.
Superparasitisasi yang cukup tinggi terjadi pada beberapa perlakuan ketiadaan inang empat hari khususnya yang mengalami ketiadaan inang di awal. Hal ini mungkin terjadi karena parasitoid mengalami ketiadaan inang dalam waktu yang cukup lama (Vinson 1985 dalam Ratna 2008). Adanya ketiadaan inang yang cukup panjang di awal hidup S. manilae kemungkinan menyebabkan parasitoid ini menjadi kurang berpengalaman dalam membedakan inang. Selain itu adanya telur yang telah diproduksi dalam ovari, dapat merangsang parasitoid ini untuk terus meletakkan telur berkali-kali pada inang yang ditemuinya. Liu dan Morton (1986 dalam Godfray 1994) menduga bahwa parasitoid lebih memilih meletakkan telur lebih dari satu di dalam tubuh inang yang ditemuinya dibandingkan mati dengan menyisakan telur di dalam ovarinya.
Pengaruh Pola Ketiadaan Inang pada S. manilae Terhadap Sisa Telur, Total Telur, dan Lama Hidup
Pada saat dilakukan pembedahan setelah mati, M. rufiventris yang diberi inang setelah ketiadaan inang selama 1, 3, 5, 6, 7, dan 8 hari memiliki jumlah telur yang berbeda-beda di dalam ovarinya (Hegazi 2007). Hal yang sama juga ditemukan pada S. manilae yang telah diberikan perlakuan ketiadaan inang. S. manilae yang mengalami ketiadaan inang di awal cenderung memiliki sisa telur dalam ovari yang rendah. Hal ini kemungkinan terjadi akibat S. manilae terus meletakkan telur yang telah diproduksinya akibat rangsangan bertemu dengan inang setelah mengalami ketiadaan inang. Lamanya ketiadaan inang terlihat cukup berpengaruh terhadap sisa telur dalam ovari S. manilae. Walaupun dengan pola yang sama (seling dan belakang) S. manilae yang mengalami ketiadaan inang selama empat hari memiliki sisa telur dalam ovari yang jauh lebih banyak