Keragaman Genetik dan Korelasi Parameter Warna Beras dan Kandungan Antosianin Total Sebelas Kultivar Padi Beras Hitam Lokal
Genetic Variability of Rice Pericarp Color Parameters and Total Anthocyanine Content of Eleven Local Black Rice and Their Correlation
Kristamtini1, Taryono2, Panjisakti Basunanda2 dan Rudi Hari Murti2
ABSTRACT
Black rice starts to be consumed as functional food due to high anthocyanine content which functioned as an antioxidant. The different in an existing name is predicted due to the pericarp color differences which are from light to heavy black. It is therefore that morphological characteristics observation of pericarp color is required. This study aimed to identify genetic variability of 11 Indonesian local cultivar of black rice based on pericarp color parameters and total anthocyanine content, even their correlations. L*, a*, b* color parameters were observed using Chroma Meter - Konica Minolta – Minolta CM-2006, and white standard color was used for calibration, whereas total anthocyanine content was measured based on absorbent value of grinded rice grains using 535 nm wavelength of spectrophotometer. Color parameters and total anthocyanine content data were analyzed using analysis variance to estimate Genetics Variability Coefficient and cluster analysis to know the similarity among these local cultivars of black rice. The result showed that b* color variable and total anthocyanine content indicate broad genetic variability, whereas L* and a* color variables depict narrow genetic variability. There were 3 different groups of black rice based on L*, a* and b* color parameters and total anthocyanine content. These were Cempo ireng; Banjarnegara, Banjarnegara-Wonosobo, Magelang berbulu, Magelang tak berbulu, Nusa Tenggara Timur and Pari Ireng; and Sragen, Jlitheng, Bantul and Melik groups. Positive significant correlation was observed between L*, a*, b* color parameters, and there was negative significant correlation between total anthocyanine content and L*, a*, dan b* color parameters.
Key words: genetic variability, pericap colour parameters, total anthocianin content, local black rice
INTISARI
Beras hitam mulai banyak dikonsumsi sebagai pangan fungsional karena tingginya kandungan antosianin yang berfungsi sebagai antioksidan. Perbedaan nama beras hitam yang ada diduga disebabkan oleh keragaman warna berasnya, yaitu dari hitam cerah sampai hitam pekat. Oleh karena itu pengamatan sifat morfologi warna beras sangat diperlukan. Penelitian ini
1
Balai Pengkajian Teknologi Pertanian (BPTP) Yogyakarta
2
bertujuan untuk mengidentifikasi keragaman genetik sebelas kultivar padi beras hitam lokal Indonesia berdasarkan parameter warna beras dan kandungan antosianin total serta korelasinya. Pengamatan parameter warna L*, a*, b* dilakukan menggunakan Chroma Meter - Konica Minolta – Minolta CM-2006, dan standarisasi dilakukan dengan kalibrasi warna standar (putih). Adapun analisis kandungan antosianin total dilakukan berdasarkan nilai absorban ekstrak tepung beras pada panjang gelombang 535 nm dengan alat spektrofotometer. Data pengamatan parameter warna maupun kandungan antosianin total dianalisis untuk mengetahui keragaman genetiknya yang ditunjukkan oleh nilai Koefisien Keragaman Genetik (KKG) dan analisis gerombol (cluster analysis) sehinga diketahui kedekatan hubungan antar kultivar lokal padi beras hitam tersebut. Hasil penelitian menunjukkan
bahwa peubah warna b* dan kandungan antosianin total memiliki keragaman
genetik yang luas, sedangkan peubah warna L * dan a * memiliki keragaman genetik yang sempit. Terdapat 3 kelompok kultivar padi beras hitam lokal berdasarkan parameter warna L*, a*, dan b* dengan kandungan antosianin total, yaitu beras hitam Cempo ireng; kelompok beras hitam Banjarnegara, beras hitam Banjarnegara-Wonosobo, beras hitam Magelang berbulu, beras hitam Magelang tak berbulu, beras hitam NTT dan Pari ireng; dan kelompok kultivar beras hitam Sragen, beras hitam Jlitheng, beras hitam Bantul dan beras hitam Melik. Keeratan hubungan antar pengamatan menunjukkan bahwa terdapat korelasi positif antar parameter warna L*, a*, b*, dan korelasi negatif antara parameter warna (L*, a*, dan b* ) dengan kandungan antosianin total
Kata Kunci: keragaman genetik, parameter warna, antosianin total, beras hitam, dan lokal.
PENDAHULUAN
Padi merupakan serealia utama yang hasilnya dikenal sebagai beras
yang dikonsumsi sebagai makanan pokok oleh lebih dari setengah populasi
dunia (Battacharjee et al., 2002). Terdapat beragam warna padi/beras dan
warna tersebut tergantung pada pigmen warna khususnya antosianin pada
lapisan perikarp, kulit biji (seed coat) atau aleuron, seperti beras merah (red
rice) dan beras hitam (black rice), tetapi sebagian besar beras yang
dikonsumsi adalah beras putih (Chaudhary, 2003). Pada saat ini, beras
hitam mulai banyak dikonsumsi sebagai pangan fungsional yaitu pangan
yang secara alami atau melalui proses tertentu mengandung satu atau lebih
kesehatan dalam hal ini kandungan antosianin yang dapat berfungsi sebagai
antioksidan.
Beras hitam lokal memiliki sebutan yang beragam tergantung daerah
asalnya. Beras hitam di Kotamadya Surakarta, Propinsi Jawa Tengah disebut
beras Wulung, di Kabupaten Subang, Jawa Barat dikenal dengan beras
gadog. Di Propinsi Daerah Istimewa Yogyakarta, terdapat beberapa nama
beras hitam. Di Sleman Yogyakarta, dikenal dengan nama Cempo Ireng
(Kristamtini, 2008) dan beras jlitheng (Kristamtini, 2009), sedangkan di Bantul
dikenal beras Melik, di Nusa Tenggara Timur (NTT), dikenal Aen Metan dan
Hare Kwa (Suhartini dan Suardi, 2010), di Magelang dikenal 2 jenis beras
hitam yaitu cempa dan berbulu dengan sebutan nama Jawa melik.
Perbedaan nama beras hitam tersebut diduga disebabkan oleh
keragaman warna berasnya, dari hitam cerah sampai hitam pekat.
Perbedaan warna beras terjadi sebagai akibat adanya perbedaan kandungan
antosianin. Dengan kajian warna beras maka kemungkinan dengan mudah
dapat dibedakan suatu indvidu dengan individu lainnya (Kaplan, 2001).
Kajian keragaman genetik dapat dilakukan dengan penanda
morfologis, biokimia, dan molekuler (Solouki et al., 2008). Penyandian
sifat morfologi banyak dipengaruhi lingkungan, namun demikian sifat
morfologi telah banyak memberi manfaat dalam membentuk sejumlah kultivar
unggul sejak tahun 1950-an. Penyifatan secara morfologi merupakan cara
determinasi yang paling akurat untuk menilai sifat agronomi dan
mengelompokkan secara taksonomi beragam tanaman (Li et al., 2009).
Penyifatan secara morfologi dapat digunakan untuk mengenali koleksi
plasma nutfah ganda, kajian pendugaan keragaman genetik dan korelasi
antara sifat morfologi dengan sifat penting agronomi yang lain (Ciat, 1993;
Rimoldi et al., 2010; Talebi et al., 2008). Penyifatan pada tingkat morfologi
diperlukan terutama untuk keperluan mengenali fenotipe dan perubahannya
Keragaman genetik antara individu atau populasi dapat diduga
dengan menggunakan penanda morfologi (Garcia et al., 1998). Identifikasi
keragaman dengan cara penyifatan akan menghasilkan data berisi informasi
tentang sifat-sifat morfologi seperti warna bunga, dan bentuk daun dan
agronomi termasuk umur panen, tinggi tanaman, dan daya hasil. Penyifatan
secara morfologi lebih utama dilakukan dibandingkan molekuler karena
mudah dilakukan dan nampak secara jelas. Penanda morfologi yang
digunakan merupakan penanda yang didasarkan pada pewarisan Mendel
yang sederhana, seperti bentuk, warna, ukuran, dan berat. Sifat morfologi
(fenotipe) dapat digunakan sebagai petunjuk nyata untuk gen khusus dan
penanda gen dalam kromosom karena sifat-sifat yang mempengaruhi
morfologi dapat diturunkan (Sofro, 1994). Dalam jumlah besar penanda
morfologi telah dipelajari dan dipetakan pada beragam jasad hidup termasuk
manusia, tikus, lalat buah, jagung, tomat, dan ubi jalar (Liu, 1998; Karuri et
al., 2010).
Keberadaan keragaman fenotipe dalam suatu populasi tanaman
sangat penting dan dapat dijadikan sebagai penduga keragaman genetik,
agar seleksi dengan maksud untuk mendapatkan sifat-sifat unggul dapat
dilakukan. Makin tinggi keragaman fenotipenya pada sifat yang tidak
dipengaruhi oleh faktor lingkungan maka peluang untuk mendapatkan
genotipe unggul semakin besar (Greech dan Reits, 1971). Keragaman
fenotipe pada sifat tersebut menunjukkan keragaman faktor genetik terhadap
sifat yang disandikan (Knight, 1979).
Berdasarkan hal-hal tersebut, penelitian ini bertujuan untuk
mengidentifikasi keragaman genetik sebelas kultivar padi beras hitam lokal
Indonesia berdasarkan parameter warna beras dan kandungan antosianin total
BAHAN DAN METODE
Bahan tanaman yang digunakan dalam penelitian ini adalah 11
kultivar padi beras hitam lokal Indonesia (Tabel 1).
Tabel 1. Materi penelitian yang dilibatkan dalam penelitian ini
No. Nama kultivar/varietas Kode Daerah Asal
1. Melik A Kedon – Ganjuran – Bantul –
Semua materi yang digunakan dalam penelitian ini ditanam dalam pot
plastik di rumah plastik hingga panen menggunakan rancangan lingkungan
Acak Lengkap yang diulang 5 kali.
Pengamatan warna beras dan kandungan antosianin total beras
dilakukan setelah padi dipanen. Parameter warna beras L* (lightnes), a*
(redness – greenness), dan b* (yellowness–blueness) diamati menggunakan
Chroma Meter - Konica Minolta – Minolta CM-2006, dan pembakuan dilakukan
dengan kalibrasi warna baku-putih. L* menunjukkan kecerahan (lightness), a*
(redness –greenness) mengindikasikan kemerahan sampai kehijauan dan b*
(yellowness-blueness) menunjukkan kekuningan-kebiruan (Bao et al, 2005).
Analisis kandungan antosianin total menggunakan metode Lees dan
gelombang 535 nm dengan alat spektrofotometer - Spectro Simadzu. UV VIS
tipe 1601 dengan pelarut metanol.
Data kuantitatif dianalisis dengan pendekatan analisis varian dan
untuk mengetahui keragaman genetiknya yang ditunjukkan oleh nilai
Koefisien Keragaman Genetik (KKG), dihitung menurut rumus Singh dan
Chaudary (1979) :
KTg -KTs σ2
G = ---
r
( σ2 G)
KKG = --- x 100
X
Menurut Moedjiono dan Mejaya (1994), koefisien keragaman genetik
yang telah diperoleh dapat dikelompokkan menjadi 4 kriteria keragaman yaitu
keragaman rendah ( 0 – 25 % dari KKG tertinggi), keragaman sedang (25 –
50 % dari KKG tertinggi), keragaman tinggi (50 -75 % dari KKG tertinggi) dan
keragaman sangat tinggi (> 75 % dari KKG tertinggi).
Data hasil pengamatan kuantifikasi warna (parameter L*, a*,
dan b*) yang diperoleh dianalis is menggunakan perangkat lunak
Multivariate Statistical Package (MVSP) ver 3.01 (Kovach, 2007) berdasarkan
koefisien DIST/rataan jarak taksonomi sehingga diperoleh dendogram dan
jarak taksonomi.
Korelasi antar pengamatan yang diamati dianalisis menggunakan
perangkat lunak SPSS 16.0 dengan tujuan untuk mengetahui hubungan antar
parameter warna dengan kandungan antosianin total.
σ2
G = ragam genetik
KTg = kuadrat tengah genotipe
KTs = kuadrat tengah galat
KKG = Koefisien keragaman genetik
̅ = rerata
HASIL DAN PEMBAHASAN
Survei genotipe beras dari berbagai koleksi plasma nutfah dapat
mengenali kandungan fenolat total, flavonoid dan kapasitas antioksidan
(Shen et al., 2009). Memahami dasar genetika yang mendasari keragaman
genetik kandungan fitokimia (fenolat total, flavanoid dan antioksidan)
kaitannya dengan manfaat kesehatan dapat membantu proses pemuliaan (Jin
et al., 2009). Beras hitam memiliki kandungan antosianin tinggi pada lapisan
perikarp, yang memberikan warna ungu gelap (Ryu et al., 1998; Takashi et
al., 2001). Beras hitam memiliki keragaman dalam hal warna dan dapat dikaji
dengan parameter warna.
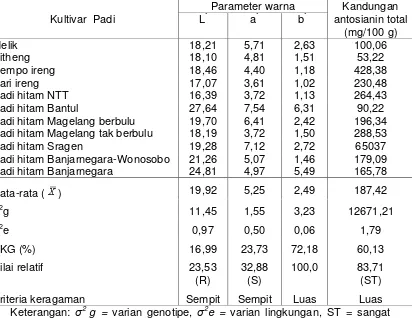
Tabel 2. Varian genetic parameter warna (L*, a*, dan b*) serta kandungan antosianin total sebelas kultivar lokal padi beras hitam
Padi hitam Magelang berbulu 19,70 6,41 2,42 196,34
Padi hitam Magelang tak berbulu 18,19 3,72 1,50 288,53
Padi hitam Sragen 19,28 7,12 2,72 65037
Padi hitam Banjarnegara-Wonosobo 21,26 5,07 1,46 179,09
Padi hitam Banjarnegara 24,81 4,97 5,49 165,78
Rata-rata ( ) 19,92 5,25 2,49 187,42
Kriteria keragaman Sempit Sempit Luas Luas
Tabel 2 tampak bahwa varian lingkungan pada semua parameter warna
L*, a*, dan b* serta kandungan antosianin total lebih rendah dari pada varian
genotipe. Hal tersebut menunjukkan bahwa keragaman parameter warna L*,
a*, dan b* serta kandungan antosianin total lebih dipengaruhi oleh faktor
genetis dari pada faktor lingkungan. Oleh karena itu parameter warna dan
kandungan antosianin total merupakan sifat yang dipengaruhi oleh sedikit gen
(sifat kualitatif). Hal ini sesuai pendapat Mangoendidjojo (2003) bahwa sifat
kualitatif diatur oleh satu atau dua gen, sedikit dipengaruhi lingkungan
sehingga cara pemilihan/seleksi dapat dilakukan secara visual.
Karena peubah warna b* mempunyai nilai Koefisien Keragaman
Genetik (KKG) tertinggi (Tabel 3) sebesar 72,18 % maka ditetapkan sebagai
nilai relatif 100%. Nilai keragaman genetik sedang terdapat pada peubah
warna a* dan warna L* memiliki nilai keragaman genetik rendah, sedangkan
kandungan antosianin total memiliki nilai keragaman genetik sangat tinggi.
Suatu sifat yang memiliki nilai keragaman genetik sangat tinggi dan tinggi
menunjukkan bahwa perbaikan melalui seleksi langsung (direct selection)
dimungkinkan. Sifat yang memiliki nilai keragaman tinggi dan sangat tinggi
dapat dikatakan mempunyai keragaman genetik yang luas, demikian
sebaliknya sifat yang memiliki nilai keragaman genetik rendah dan sedang
dapat dikatakan mempunyai keragaman genetik yang sempit, sehingga
peubah warna b*dan kandungan antosianin total memiliki keragaman genetik
yang luas, sedangkan parameter warna L* dan a* memiliki keragaman genetik
yang sempit.
Keragaman yang luas merupakan salah satu syarat terhadap seleksi
pada sifat yang diinginkan karena proses seleksi terhadap sifat tersebut akan
lebih efisien. Sesuai dengan Bahar dan Zein (1993) bahwa apabila
keragaman genetik dalam suatu populasi besar, hal ini menunjukkan individu
dalam populasi beragam, peluang untuk memperoleh genotipe yang
diharapkan akan besar. Semakin luas keragaman genetik yang dimiliki akan
keragaman yang luas juga dapat meningkatkan tanggapan seleksi (selection
response) karena tanggapan seleksi berbanding lurus dengan keragaman
genetik (Fehr, 1987; Hallauer dan Miranda, 1988; Simmonds, 1986).
Pengelompokan berdasarkan parameter warna L*, a*, dan b* serta
kandungan antosianin total selain bermanfaat untuk mengenali kultivar juga
bermanfaat untuk membantu dalam pemilihan tetua persilangan. Hal ini dapat
dimengerti karena dengan dikenalinya kultivar dan hasil pengelompokan
tersebut maka kultivar yang berada dalam satu kelompok memiliki sifat yang
sama atau hampir sama sehingga akan memudahkan dalam pemilihan tetua
persilangan.
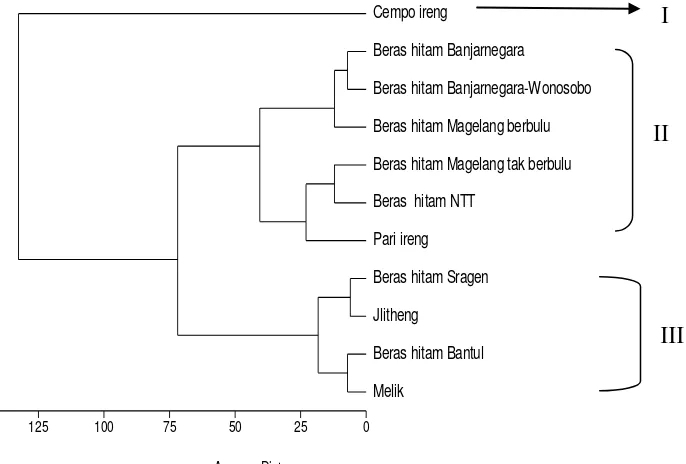
Gambar 1. Dendogram hasil pengelompokan berdasarkan parameter warna L*, a*, dan b* serta kandungan antosianin total pada 11 kultivar padi beras hitam lokal
Pada dendogram gambar 1 nampak bahwa pada jarak taksonomi 75
diperoleh 3 kelompok yaitu kelompok I terdiri dari kultivar Cempo ireng,
kelompok II terdiri dari dua kelompok yaitu kelompok beras hitam
Average Distance
Melik
Beras hitam Bantul Jlitheng
Beras hitam Sragen Pari ireng Beras hitam NTT
Beras hitam Magelang tak berbulu Beras hitam Magelang berbulu Beras hitam Banjarnegara-Wonosobo Beras hitam Banjarnegara Cempo ireng
150 125 100 75 50 25 0
I
II
Banjarnegara, beras hitam Banjarnegara-Wonosobo dan beras hitam
Magelang berbulu; dan kelompok beras hitam Magelang tak berbulu, beras
hitam NTT dan pari ireng. Adapun kelompok III terdiri dari dua kelompok yaitu
beras hitam Sragen dan beras hitam Jlitheng; dan kelompok beras hitam
Bantul dan Melik. Dengan dikenalinya kultivar dan berdasarkan hasil
pengelompokan tersebut maka kultivar yang berada dalam satu kelompok
memiliki parameter warna dan kandungan antosianin yang sama atau hampir
sama atau memiliki hubungan kekerabatan yang dekat sehingga untuk
persilangan tinggal memilih tetua persilangan yang mewakili kelompoknya.
Pendugaan hubungan genetik dalam pemuliaan tanaman sangat
berguna untuk mengelola plasma nutfah, mengenali kultivar, membantu
seleksi tetua persilangan serta mengurangi jumlah individu yang dibutuhkan
untuk mengambil contoh dengan kisaran keragaman yang luas (Julisaniah et
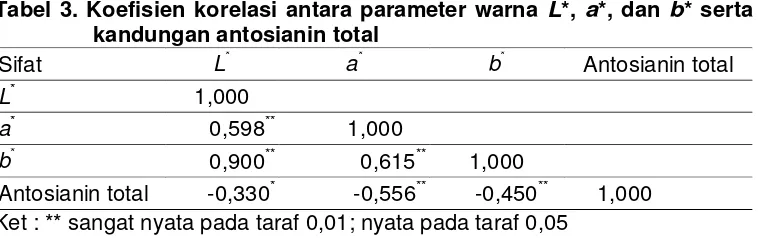
al., 2008). Terdapat korelasi yang nyata pada parameter warna L*, a*, dan b*
serta kandungan antosianin total (Tabel 3). Korelasi positif nyata teramati
antar parameter warna L*, a*, dan b*, adapun antara parameter warna L*,
a*, dan b* dengan kandungan antosianin total terdapat korelasi negatif
nyata.
Tabel 3. Koefisien korelasi antara parameter warna L*, a*, dan b* serta kandungan antosianin total
Sifat L* a* b* Antosianin total
L* 1,000
a* 0,598** 1,000
b* 0,900** 0,615** 1,000
Antosianin total -0,330* -0,556** -0,450** 1,000 Ket : ** sangat nyata pada taraf 0,01; nyata pada taraf 0,05
Korelasi negatif tersebut mengandung makna bahwa semakin tinggi
nilai parameter warna L*, a*, dan b* maka kandungan antosianin total akan
semakin rendah, demikian sebaliknya semakin rendah nilai parameter warna
L*, a*, dan b* akan semakin tinggi kandungan antosianin totalnya. Hal ini
akan semakin cerah. Hubungannya dengan hubungan antara peubah L dan
kandungan antosianin, hal ini sesuai dengan hasil penelitian Yafang et al
(2011) yang menyimpulkan bahwa terdapat korelasi negatif antara parameter
warna L* dengan kandungan antosianin total. Yodmanee et al. (2011)
mengatakan bahwa terdapat korelasi negatif antara parameter warna L*, a*,
dan b* dengan kandungan polifenol total dan kapasitas antioksidan. Shen et
al. (2009) juga menemukan bahwa parameter warna L*, a*, dan b* memiliki
hubungan dengan polifenol dan aktivitas antioksidan.
Korelasi negatif pada parameter warna a*, dan b* dengan antosianin
total dapat terjadi karena nilai peubah a* bernilai negatif menunjukkan warna
hijau dan mengarah ke warna merah apabila nilai menuju ke positif,
sedangkan parameter b* akan menunjukkan nilai negatif apabila berwarna
dekat dengan biru dan menuju ke positif apabila warna mengarah ke kuning.
Oleh karena itu maka semakin rendah nilai a* dan b* maka semakin tinggi
kandungan antosianinnya. Penjelasan tentang korelasi negatif antar
parameter warna L*, a*, dan b* dengan kandungan antosianin total tersebut
sesuai dengan Ryu et al., (1998) dan Takashi et al. (2001) bahwa beras
hitam memiliki kandungan antosianin tinggi pada lapisan perikarp, yang
memberikan warna ungu gelap. Hasil penelitian ini dapat digunakan sebagai
petunjuk (guide) di dalam seleksi dalam pengembangan varietas beras
dengan kandungan antosianin tinggi dan penggunaan parameter warna
sebagai indikator praktis dalam seleksi.
KESIMPULAN
1. Peubah warna b * dan kandungan antosianin total memiliki keragaman
genetik yang luas, sedangkan peubah warna L * dan a * memiliki
keragaman genetik yang sempit.
2. Terdapat 3 kelompok kultivar padi beras hitam lokal berdasarkan
parameter warna L*, a*, dan b* dengan kandungan antosianin total,
beras hitam Banjarnegara-Wonosobo, beras hitam Magelang berbulu,
beras hitam Magelang tak berbulu, beras hitam NTT dan Pari ireng; dan
kelompok kultivar beras hitam Sragen, beras hitam Jlitheng, beras hitam
Bantul dan beras hitam Melik
3. Terdapat korelasi positif antar parameter warna L*, a*, b*, dan terdapat
korelasi negatif antara parameter warna L*, a*, dan b* dengan
kandungan antosianin total.
UCAPAN TERIMA KASIH
Pada kesempatan ini penulis sampaikan terima kasih kepada
Kerjasama Kemitraan Penelitian Pertanian dengan Perguruan Tinggi (KKP3T)
tahun 2011 yang telah memberikan dana untuk penelitian ini dan terimakasih
kepada Laboratorium Pemuliaan dan Genetika Fak. Pertanian UGM yang
telah memberikan ijin untuk menggunakan alat Chroma Meter - Konica Minolta
– Minolta CM-2006.
DAFTAR PUSTAKA
Bao, J.S., Y. Cai, M. Sun, G. Wang, H. Corke. 2005. Anthocyanins, flavonols, and free radical scavenging activity of Chinese bayberry (Myrica rubra) extracts and their color properties and stability. Journal of Agricultural Food Chemistry 53 (6): 2327-2332
Battacharjee, P., R.S. Shinghal, and P.R. Kulkkarno. 2002. Basmati rice : A review. Int. J. Food Sci. Tech. 37 (1): 1-12.
Bahar , H., dan S. Zen. 1993. Parameter genetik pertumbuhan tanaman, hasil dan komponen hasil jagung. Zuriat 4 (1): 4-7.
Chaudhary, R.C. 2003. Speciality rices of the world : Effect of WTIO and IPR on its production trend and marketing. J. Food Agric. Env. 1 (2) : 34 -41.
CIAT. 1993. Biotechnology Research Unit. Annual Report, Cali, Colombia International Potato Centre (CIP), Asian Vegetable Research and Development Centre (AVRDC), International Board for Plant Genetic Resources (IBPGR), 1991. In: Z. Huaman (ed.), Descriptors for Sweet Potato, pp: 43–130. IBPGR, Rome, Italy
Garcia, E., M. Jamilena, J.I. Alvarest, T. Arnedo, J.L.Oliver, and R. Lozano. 1998. Genetic relationships among melon breeding lines revealed by RAPD marker and agronomic traits. Theor. Appl. Genet. 96: 878-887. Greech, J.L. and P.L. Reits. 1971. Plant Germplasm Now and Tomorrow. In:
N.C. Brady (ed). Advance in Agronomy. Academy Press.
Hallauer, A.R., and J.B. Miranda. 1988. Quantitative Genetics in Maize Breeding. Iowa State University Press. Pp. 468
Jin., L., P. Xiao, Y. Lu, Y.F. Shao, Y. Shen, J.S.Bao. 2009. Quantitative trait loci for brown rice color, phenolics, flavonoid contents, and antioxidant capacity in rice grain. Cereal Chem. 86: 609-615.
Julisaniah, N. I., L. Sulistyowati, dan A. N. Sugiharto. 2008. Analisis kekerabatan mentimun (Cucumis sativus L.) menggunakan Metode RAPD-PCR dan isozim. Biodiversitas. 9 (2): 99-102.
Karuri, H.W., E.M. Ateka, R. Amata, A.B. Nyende, A.W.T. Muigae, E. Mwasame, and S.T. Gichuki. 2010. Evaluating diversity among Kenyan sweet potato genotypes using morphological and SSR markers. Int. J. Agric. Biol. 12: 33-38.
Knight, R. 1979. Quantitative genetic statistics and plant breeding. In: R. Knight (ed). Plant Breeding, Brisbane. 41-76p.
Kaplan, D.R. 2001. The science of plant morphology : definition, history and role in modern biology (On line). American Journal of Botany 88 (10) :
1711 – 1741. http://www. American Journal of
Botany.com/journal/morphology/v88.
Kovach, W.L. 2007. Kovach computing services. MVSP Plus Version 3.1
User’s Manual. Publish by Kovach Computing Services. Pentracth,
Wales. U.K. Printed. Sept 2007. p. 137.
Kristamtini. 2009. Keragaan Beras Hitam sebagai Sumberdaya Genetik
Lokal. Prosiding Risalah Aplikasi Paket Teknologi “Mendukung Hari
Pangan Sedunia”. BPTP Yogyakarta.
Kristamtini. 2008. Penampilan Cempo Ireng sebagai Sumberdaya Genetik Lokal Beras Hitam. Prosiding Seminar Nasional. Pengembangan Produk Berbasis Sumber Pangan lokal untuk Mendukung Kedaulatan Pangan. Program Studi Teknologi Hasil Pertanian Fak. Agroindustri.
Universitas Mercu Buana Yogyakarta Bekerjasama dengan
Perhimpunan Ahli Teknologi Pangan (PATPI) Yogyakarta dan Lembaga Ilmu Penegtahuan Indonesia (LIPI). Yogyakarta. 18 Desember 2008.
Li, P., Y. Yumwen, X. Sum, and J. Han. 2009. Using microsatellite (SSR) and morphological markers to assess the genetic diversity of 12 alfalfa (Medicago sativa spp. falcata) population from Eurasia. Afr. J. Biotechnol. 8(10): 2102-2108.
Marzuki, I., M.R. Uluputty, A.A. Sandra, dan S. Memen. 2008. Karakterisasi morfoekotipe dan proksimat pala Banda (Myristica fragrans Houtt). Bul. Agron. 36(2): 145-151.
Mangoendidjojo, W. 2003. Dasar-dasar pemuliaan tanaman. Yogyakarta : Kanisius.
Moedjiono dan M.J. Mejaya. 1994. Variabilitas genetik beberapa karakter plasma nutfah jagung koleksi Balittan Malang. Zuriat: 5 (2): 27-32. Rimoldi, F., P.D.V. Filho, M.V. Kvitschal M.C. Gonzalvesvidigal, A.J. Pioli,
S.M.A.P. Prioli, and T.R. dA Costa. 2010. Genetic divergence in sweet cassava cultivars using morphological agronomic traits and RAPD molecular markers. Brazilian Archives Biology and Technology 53(6): 1447-1487.
Ryu,S.N., S.Z. Park and C.T. Ho. 1998. High performances liquid chroomatographic determination of anthocyanin pigments in some varieties of black rice. Journal of food and Drug Analysis 6 : 1710-1715.
Shen, Y., L. Jin, P. Xiao, Y. Lu, J.S. Bao. 2009. Total phenolics, flavonoids, antioxidant capacity in rice grain and their relations to grain color, size and weight. Journal of Cereal Science 49: 106-111.
Simmonds, N.W. 1986. Evaluation of Crops Plant. Longman Scientific & Technical. England. 339 pp
Singh, R.K. and B.D. Chaudhary. 1979. Biometrical Method in Quantitative Genetics Analysis. New Delhi : Kalyani Publ.
Sofro, A.S.M. 1994. Keanekaragaman Genetik. Yogyakarta: Andi Offset. Solouki, M.,H., Mehdikhani, H. Zeinali dan A.A. Emamjomeh. 2008. Study of
genetic diversity in Chamomile (Matricaria chamomilla) based on m o r p h o l o g i c a l t r a i t s a n d molecular markers. Sci. Hortic. 117: 281-287.
Suhartini, T dan D. Suardi. 2010. Potensi beras hitam lokal Indonesia. Warta Penelitian dan Pengembangan Pertanian 32(1):9-10. RAPD and agronomic markers. International Journal of Agricultural Biology 10(3): 301-305.
Yafang, S., Z. Gan and B. Jinsong. 2011. Total phenolic content and antioxidant capacity of rice grains with extremely small size. African Journal of Agricultural Research 6 (10) : 2289-2293.