METABOLISME PROTEIN
DAN ASAM AMINO
DISUSUN OLEH
KELOMPOK 5
WINNY AYUWIRA ASHARY (21)
YOHANA ROTUA SITUMORANG (22)
ANIKE SITUNGKIR (23)
ELISABET SIHOMBING (24)
ELVIANI (25)
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
UNIVERSITAS NEGERI MEDAN
BAB I
PENDAHULUAN
Dalam tubuh makhluk hidup terdapat banyak proses-proses biokimia, salah satu diantaranya ialah proses metabolisme protein dan asam amino. Protein berasal dari kata proteos yang berarti “pertama” atau utama adalah makromolekul yang paling banyak ditemukan di dalam semua sel dan semua bagian sel. Protein memiliki fungsi biologis sebagai instrumen molekuler yang menyampaikan informasi genetik.
BAB II
PEMBAHASAN
METABOLISME PROTEIN DAN ASAM AMINO
2.1. Mekanisme Reaksi Sintesa Protein
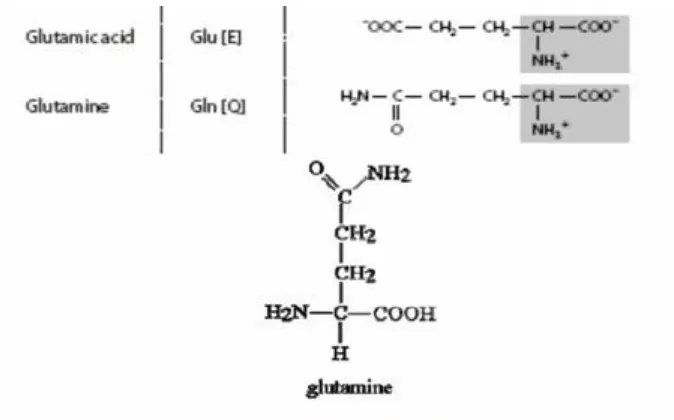
Ada dua hal yang penting pada sintesa protein, yaitu pembentukan ikatan yang membutuhkan energy dan spesifitas.Tiap ikatan peptide membutuhkan energi sebesar lebih kurang 3000 kalori/mol yang diperoleh dari ATP.Prototip sintesa protein yang dipergunakan sebagai dasar-dasar gagasan sintesa polipeptida ini adalah sintesa asam hipurat, glutamin dan glutation, dimana ATP sebagai sumber energy terlibat dalam reaksi.
Gambar 2.1. Jenis sintesa protein untuk sintesa polipeptida
Glutamat + sistein + ATP y-glutamilsistein + ADP Pan
y-glutamilsistein + glisin + ATP glution + ADP + Pan
Dari tiga reaksi yang disajikan diatas dapat diketahui, bahwa biosintesa senyawa tersebut sudah terarah pada hasil tertentu yang dilakukan oleh enzim tertentu. Pada reaksi pembentukan glutamin, aktivasi gugus oleh ATP terjasi pada karboksil y dan tidak pada yang α. Pada reaksi pertama gugus karboksil yang diaktifkan oleh ATP adalah y-karboksil dari asam glutamat dan bukan gugus karboksil kepunyaan sistein. Pada reaksi berikutnya dipeptide yang terbentuk diaktifkan pada bagian karboksil sistein yang selanjutnya ditempeli oleh glisin. Cara aktivasi berbeda dari yang telah diutarakan diatas akan terbentuk tripeptida bukan glutation yang dengan sendirinya fungsinya berbeda.
2.2. Tahapan Reaksi Biosintesa
Biosintesa polipeptida dapat dibagi menjadi tahap-tahap aktivasi permulaan, pemanjangan dan pengakhiran.
2.2.1. Tahap aktivasi
Reaksi aktivasi ini membutuhkan energy yang berasal dari ATP yang dikatalis oleh amino-asil-t-RNA sintesa. Reaksi jumlahnya adalah sebagai berikut:
Asam amino + ATP + t-RNA amino-asil-t-RNA +AMP +P – P
Enzim sintesa yang mengkatalisa reaksi diatas mempunyai tiga sisi aktif yang dapat mengikat tiga substrat, yaitu asam amino, ATP dan t-RNA.Senyawa pentarsfer asama amino ini sifatnya spesifik, artinya t-RNA tertentu hanya mengangkut asam amino tertentu pula.Kekhususan terletak pada sisi antikodon yang terdapat pada t-RNA tersebut.Karena da 20 jenis asam amino maka dalam sel juga terdapat sekurang-kurangnya 20 jenis t-RNA pula yang kesemuanya (pada E.coli) sudah dapat diisolasi bahkan sebagian besar dapat dikristalkan. Reaksi aktivasi itu dapat dipecahkan menjadi dua sub tahap, yaitu:
Asam amino + ATP ↼( asam aminoadenilat) + P ⎼ P
( Asam aminoadenilat) + t-RNA spesifik aminoasil-t-RNA + asam adenilat)
Gambar 2.2. Tahap Aktivasi
Walaupun enzim yang mengkatalisa reaksi ini bersifat spesifik dan selektif, namun terjadi suatu kesalahan tidaklah mustahil (taraf kesalahan ± 0,01%). Misalnya p-fluorofenilalamin,etionin atau isoleusin sempat pua masuk kedalam sisi aktif enzim, sehingga protein yang terjadi terdapat senyawa yang mirip asam amino dan yang tidak di kehendaki berdasarkan informasi yang ada.
Masuknya senyawa-senyawa di atas kedalam rantai polipeptida berlangsung seperti yang terjadi pada basa asam-asam amino alam.
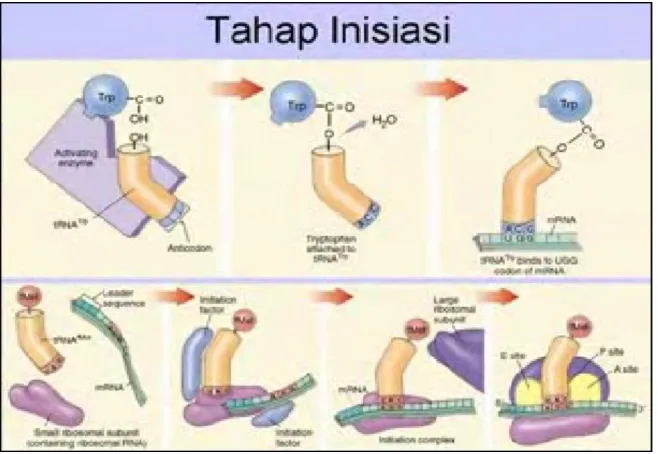
2.2.2. Tahap Permulaan (inisiasi)
Pada semua sel sintesa protein dimulai dengan metionin yang dibawa oleh t-RNA dalam bentuk formil-metionin-t-RNA pada sel prokariotik ( dan mitokondria ) dan metionil-t-RNA pada sel eukariotik. T-RNA yang mengangkut dua bentuk metioni ini sedikit berbeda ditandai dengan t-RNA dan t-RNA Met. Perubahan gugus metiolin yang ada pada t-RNA menjadi gugu formilnya oleh tetrahidrofolat.
Enzim yang mengkatalisa reksi di atas tidak mengikat gugus formil pada metionin bebas atau pada metionin-t-RNA met.
Setelah formilmetionil-t-RNAf (metionil_t-RNAmet) terbentuk, maka ia akan menuju ke permukaan endoplasmic terikulum di mana terdapat zarah-zarah ribosom. Pada waktu yang bersamaan terjadilah kegiatan yang dapat digambarkan sebagai berikut.
Zarah ribosom yang aktif (70 S) terpisah menjadi 2 bagian, yaitu ribosom 50 S dan 30 S. bagian terakhir ini slanjutnya ditempeli oleh sebuah factor inisial IF-3 yang kemudian bila menerima m-RNA. Asam ribonukleat (m-RNA) ini mengandung urutan dan frekuensi basa N yang diarahkan oleh DNA.Oleh karena itu maka RNA ini membawa informasi genetic atau pesan (“message”) dari DNA. Untuk membedakan dari RNA yang lain dan fungsinya sebagai pembawa informasi, maka senyawa tersebut dinamakan m-RNA. Informasi genetic yang dijabarkan sebagai urutan basa N pada m-RNA.Dinamakan sani atau kdon yang dikelopok-kelompokkan.Yang masing-masing kelompok kodon terisi atas tiga urutan basa N.
Formilmetionil-t-RNAf yang dibentuk dalam sitosol di atas kemudian menuju ke m-RNA dan menempel dengan bantuan IF – 1, GTP – IF – 2.Bagian aminoasil-t-RNA yang menempel pada m-RNA adalah antikodonnya sedemikian rupa, sehingga terjadi ikat-mengikat antara basa N t-RNA dan N m-RNA melalui ikatan jembatan hidrogen. Pengikat itu sifatnya komplementer (A = T dan G = C) dan antiparallel. Energy terbentuknya kompleks ini berasal dari GTP.
Gambar 2.3. Tahap permulaan sintesa protein
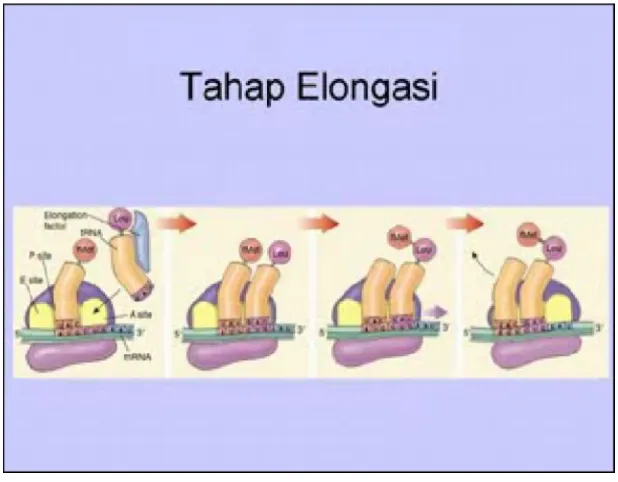
2.2.3. Tahap Pemanjangan
Dari gamabar di atas dapat diketahui bahwa celah P pada ribosom ditempati oleh met-t-RNA, sedangkan celah A masih kosong, yang pada tahap pemanjangan ini dapat di temapti oleh aminosil-t-RNA. Jenis aminoasil-t-RNA yang mana yang bias masuk kedalam celah tergantung dari kodom 3 kata basa N yang ada pada m-RNA. ( kamus yang dibuat pada halaman berikut ini dapat dipergunakan untuk mengetahui jenis aminosil-t-RNA). Sebagai contoh ialah apabila kodon m-RNA itu UUA, maka aminosil-t-RNA yang menempel pada kodon m-RNA itu dalah leusil-t-RNA Leu.
Atau dapat pula dikatakan bahwa aminosil-t-RNA yang menempel pada kodon tersebut adalah yang berantikodon t-RNA tersebut pada gugus adenosilnya mengangkut asam amino leusin.
aminoasil (A) menyerang secara nukleofilik pada gugus karbosil pada t-RNA pada celah P, yang berakibat lepasnya gugus peptil dari RNA yang ada di celah P. t-RNA yang tidak lagi mempunyai menumpang ini kemudian meninggalkan celah dan tahap berikutnya ialah telah terjadi proses translokasi (pindah tempat) yang disebabkan bergesernya zarah ribosom atau sebaliknya bergeraknya m-RNA ke arah ujung 3’ m-RNA, yang dibantuoleh EF-G-GTP. Pemindahan ribosom ini menyebabkan tergesernya peptidil-t-RNA dari celah A ke P, sehingga celah A menjadi kosong. Dengan mekanisme yang sama, maka celah A itu bias diisi oleh aminoasil-t-RNA yangdiikuti oleh pembentuka ikatan peptida dan translokasi, sehingga pada akhirnya terbentuk polipeptida yang tetap masih terikat t-RNA pada celah ribosom.
Gambar 2.4. Tahap pemanjangan rantai
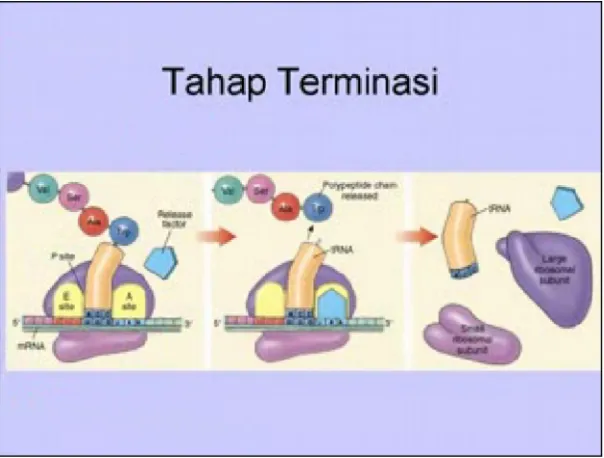
2.2.4. Tahap Pengakhiran
Tahap ini biasa terjadi oleh karena adanya kodon pada m-RNA yang tidak punya arti apa-apa. Kodon tersebut tidak dapat ditempeli oleh aminoasil-t-RNA yang manapun. Oleh karena itu maka setelah terjadi proses translokasi yang terakhir, celah A tetap kosong. Lepasnya gugus polipeptidil dari t-RNA yang terakhir dilakukan oleh faktor pelepasan yang diberi simbol R1,R2 dan R3. Ketiga
lepas dari t-RNA menjadi polipeptida bebas. Tahap akhir ini ialah lepasnya t-RNA dari celah peptidil dan terpisahnya bagian ribosom menjadi bagian-bagiannya.
Dari uraian di atas dapatlah disimpulkan, bahwa biosintesa protein tidak lain adalah suatu proses penerjemahan dari bahasa polinukleotida ke bahasa protein. Penerjamhan ini terutama dilakukan oleh m-RNA, ribosom dan t-RNA. Senyawa terakhir ni lenih dikenal sebagai adaptor. Sebagaiman adaptor pada umumnya benda tadi mempunyai dua bagian. Yang pertama adalah bagian basa N yang akan menempel pada basa N pada m-RNA dan bagian kedua ialah yang membawa gugus aminoasil yang pada waktunya akan direaksikan dengan gugus peptidil, sehingga terbentuk ikatan peptide baru. T-RNA sebagai adaptor unutk pertama kalinya dikemukakan sebagai postulat oleh FHC. Crick dan MB. Hoagland.
Gambar 2.5. Tahap Pengakhiran
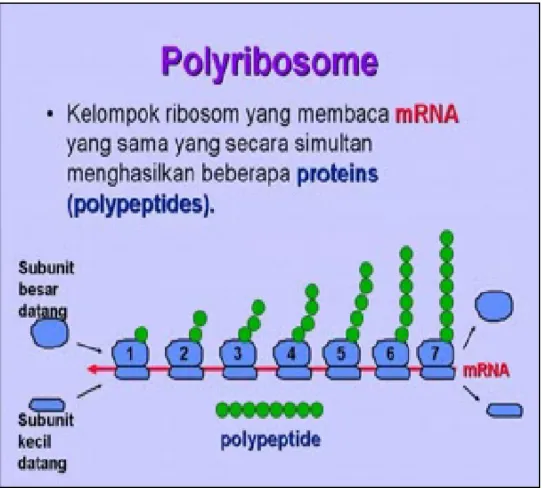
2.3. Biosintesa Protein Poliribosomal
Pada keadaan yang demikian tadi, maka tiap ribosom mempunyai tugas membentuk polipeptida. Oleh karena pola m-RNA yang dipergunakan untuk sintesa itu anya satu untuk semuanya, maka polipeptida yang dibentuk pun mempunyai komposisi, urutan dan frekuensi asam amino yang sama pula (identik).
Polipeptida yang terbentuk baik dengan cara monoribosomal maupun yang polibosomal telah menunjukkan struktur tersier alamiah. Gugus formil yang menempel pada metionin sebagai asam amino pertama pada polipeptida yang terbentuk, dipisahkan oleh enzim deformilase. Pada umumnya juga sisa asam amino metioninnya pun hilang, karena kegiatan enzim metioninaminopeptidase. Proses penempelan gugus metal, asetil, dan lain-lain pada salah satu sisa asam amino pada protein terjadi setelah polipeptida tersebut dipisahkan dari ribosomnya.
Gambar 2.6. Sintesa protein poliribosomal
Senyawa penghambat biosintesa protein antara lain adalah tetrasiklin, puromisin, khloramfinekol, sikloheksinimida (aktidion), streptomisin, racun difteria, abrin risin, dan asam fusidat. Tetrasiklin adalah sebuah antibiotika yang menutup dan mencegah masuknya aminoasil-t-RNA ke dalam celah A. Puromisin juga termasuk dalam golongan antibiotic yang mempunyai struktur yang hampir sama dengan bagian terminal dari aminoasil-t-RNA. Hanya pada antibiotik tersebut terdapat ikatan amida antara bagian ribosil dan turunan metal tirosin. Puromisin ini mampu masuk ke dalam celah A, sehingga terjadi reaksi antar gugus petidil-t-RNA yang ada pada celah P dan NH2 bebas yang terdapat pada
puromisin, maka terbentuklah peptidilpuromisin. Senyawa ini mudah meninggalkan celah. Oleh karena itu maka peptidil yang dibentuk Selama itu ikut lepas bersama-sama puromisin.
Khloroamfinekol ini menghambat reaksi transfer peptidil sehingga aminoasil-t-RNA yang sudah menempati celah A tidak bisa memperpanjang gugus tersebut. Streptomisin dan lain-lain antibiotika yang sejenis (neomisin, kanamisin) mampu mengubah konfirmasi ribosom sedemikian rupa, sehingga aminoasil-t-RNA yang terikat pada m-aminoasil-t-RNA tidak mantap, kendur. Sehingga kemungkinan reaksi transfer tidak berjalan sempurna. Anibiotik yang disebutkan diatas pada umumnya menghambat sintesa protein pada sel prokariotik, sebaliknya alkaloid seperti emetin, racun difteria, abrin dan ricin (dua yang terakhir termasuk alkaloi pada tanaman) mengganggu sintesa protein pada sel eukariot.
2.5. Sandi (Kode) Genetik
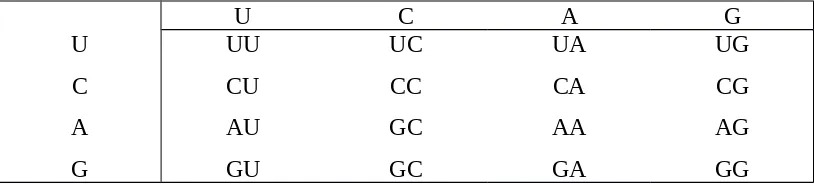
Berat molekul m-RNA berkisar antara 25.000 sampai 1.000.000 tergantung dari luas jenis protein yang dibentuk. Jumlah nukleotida yang terdapat pada m-RNA adalah sebanyak 75 sampai 3.000. apabila dipergunakan istilah sandi genetik maka “bahasa” RNA adalah basa N yang ada pada senyawa tersebut. Sebaliknya bahasa protein yang dihasilkan oleh ribosom adalah asam amino yang menyusun rantai polipeptida tersebut. Oleh karena itu maka harus ada cara terjemahan (translasi) dari bahasa RNA ke bahasa protein. Seperti diketahui bahasa RNA ada sejumlah empat (basa N) dan bahasa protein ada 20 jenis asam amino yang umum, maka hal ini tidak mungkin oleh karena dengan demikian hanya ada 4 asam amino yang dialihkan. Padahal jumlah asam amino yang perlu diterjemahkan ada 20. Bila sandi itu bukan terdiri dari satu kata melainkan dua kata (singkatan), maka dari 4 kata basa N itu dapat disusun 16 kombinan (42).
Tabel 2.1. Jumlah sandi dan asam amino
U C A G Dengan mengambil 3 kata, maka jumlah sandi sudah sangat mencukupi yaitu 43 = 64, cocok dengan dugaan semula yang berasal dari hasil pengamatan sintesa protein tudung pada virus (TSNV). RNA yang dipergunakan untuk menyandi protein yang terdiri dari 400 asam amino ini mengandung 1200 nukleotida. Dengan demikian maka tiap asam amino disandi oleh 3 nukleotida.
Dalam penelitian berikutnya dapat diketahui bahwa sebuah asam amino ada yang disandi oleh lebih dari 1 sandi sebagai berikut.
Jumlah sandi Nama asam amino yang disandi
Alanin, glisin, prolin, treolin dan valin Isoleusin
Asparagin, aspartat, sistein, glutamine, glutamat, histidin, lisin, fenilalanin dan tirosin
Metionin dan triptofan
Sandi triplet (3 kata basa N) untuk tiap-tiap basa amino dapat dibaca dalam “kamus” di halaman berikut. Sandi yang tercantum dalam kamus sandi triplet dibaca dari kiri ke kanan yang sesuai dengan urutan pada m-RNA yang dimulai dari terminal 5’P ke terminal 3’OH bebas.
5’PPP- - - A C G A G C C G U G A G A G A G U U C U . . .
3’OH
Arah baca - - - →
Sandi- sandi yang dituliskan diatas tidak berkoma artinga pembacaan mungkin saja bergeser, apabila tidak ada tanda dimulainya pembacaan. Sandi-sandi yang dituliskan diatas adalah ACG AGC CGU GAG AGA GUU dan seterusnya. Jika pada suatu keadaan salah satu bahasa N itu hilang, maka kodon tadi berubah menjadi ACG ACC GUG AGA GAG UUC dan seterusnya.
Tabel 2.3. Sandi dan asam amino-nya
A
Kodon inisiasi selalu adalah AUG. pada kamus terlihat adanya 3 kodon yang tidak mempunyai arti (*), tidak ada satu pun t-RNA yang bisa menempel pada kodon tersebut. Sandi-sandi tersebut dipergunakan untuk mengakhiri sintesa protein. Jadi dapat dikatakan bahwa sandi-sandi itu merupakan signal agar supaya biosintesa protein tadi diakhiri.
Diantara 3 basa N yang menyusun sandi hanya dua yang pokok, sedangkan NM yan ketiga kurang mempunyai arti misalnya glisin yang hanya yang hanya disandi oleh GG dan alanin oleh GC, basa N yang ketiga oleh basa N apa saja. Jika satu asam amino disandi oleh sepasang atau lebih, maka basa yang ketiga adalah U atau pirimidin atau bisa juga A atau G (purin). Misalnya fenilalanin disandi oleh UUU dan UUC, asam aspartat disandi oleh GAU dan GAC, asam glutamate disandi oleh GAA dan GAG.
DAFTAR PUSTAKA
Restuati, M., Melva Silitonga, Uswatun Hasanah, M. Yusuf Nasution. 2015.