* S.S. Montgomery
NSW Fisheries, Fisheries Research Institute, P.O. Box 21, Cronulla, NSW, Australia 2230 Received 7 February 2000; received in revised form 14 August 2000; accepted 6 September 2000
Abstract
The hypothesis tested was that catches of J. verreauxi recruits (numbers of pueruli to early juveniles) on sea-weed-type collectors were affected by the nearness of the collectors to vegetated reef and relative exposure of the collectors to sea-swell. Numbers of recruits on collectors set far from reef were greater than on collectors near reef. Collectors far from reef may be the first, preferred type of habitat encountered by recruits as they move across the Continental Shelf. Alternatively, fewer recruits may inhabit collectors set near reef because of the nearness of natural, vegetated reef. A cost–benefit analysis determined that the optimal sampling strategy for surveys of the abundance of recruits at one location and time, would be three replicate collectors at each of three sites. Sampling once with this strategy at one location would give a standard error of around 20% of the mean. This information was used in developing a uniform and optimal methodology for full surveys to monitor the relative abundance of J. verreauxi recruits along the coast of New South Wales. 2000 Elsevier Science B.V. All rights reserved.
Keywords: Lobster; Puerulus; Recruitment; Sampling strategy
1. Introduction
Data on the abundance of individuals in early stages of their life-history provide knowledge about recruitment to populations. One use of this information is in the assessment of the impact of various managerial options on the population. In the case of spiny lobsters, the information is also used to predict the quantity of catch by the commercial fishery in future years (Phillips, 1986).
*Tel.: 161-2-9527-8411; fax:161-2-9527-8576.
E-mail address: [email protected] (S.S. Montgomery).
Scientists have commonly used the relative abundance of the puerulus stage in the life cycle of spiny lobsters as an index of recruitment to the population (Booth and Phillips, 1994). Like studies on many other terrestrial (e.g., Childers and Brecht, 1996; Maelzer et al., 1996) and aquatic (e.g., Goldberg and Widman, 1989; Donaldson et al., 1991; Killgore and Baker, 1996) organisms, a type of device specifically designed to catch pueruli has been used in studies of the relative abundance of spiny lobster populations (Phillips and Booth, 1994). Devices designed to catch pueruli have become commonly known as ‘collectors’ (Phillips and Booth, 1994).
Surveys to measure the relative abundance of organisms should be uniform, unbiased, and optimal with respect to the quantity of the target species caught, and sufficiently replicated in space and time to account for patchiness in distributions (Kennelly et al., 1993). Pilot studies need to be done as a first step when developing a strategy for sampling a population so that information is collected about (i) the effects of external factors on catches in sampling gear and (ii) the spatial and temporal heterogeneity in distributions inherent in populations (Snedecor and Cochran, 1967; Winer et al., 1991; Underwood, 1997). This information can be used in cost–benefit analyses to determine optimal levels of replication in stratified, random surveys and allow a priori decisions to be made about the level of precision that is required around mean estimates of relative abundance. These techniques have been used successfully in studying terrestrial (Robinette et al., 1974; Caughley et al., 1976) and marine (Kennelly and Underwood, 1984; Kennelly, 1989; Kennelly et al., 1993) communities. But, despite the number of studies that have used collectors to study patterns in the relative abundance of pueruli of palinurids, there is little in the peer-reviewed literature about how external factors affect catches of spiny lobster recruits on collectors (but see Cruz et al., 1991; Herrnkind and Butler, 1994; Phillips and Booth, 1994) or about pilot studies and cost–benefit analyses done to optimise sampling design.
One factor that may affect catches of pueruli on collectors is the position of the collector in relation to its place in the water column, its nearness to reef and its exposure to sea-swells. For instance, some of the greatest catches of Panulirus argus pueruli (Latreille, 1804) have been taken from collectors set near the fringe of outer reefs, or the oceanic side of bays and inlets (Cruz et al., 1991; Herrnkind and Butler, 1994), whilst Phillips and Booth (1994) reported that catch rates of pueruli on collectors may be lower where ‘‘there is an abundance of natural settlement substrate’’.
The spiny lobster J. verreauxi (H. Milne Edwards, 1851), commonly called the eastern rock or packhorse lobster, occurs in waters off the east coast of Australia (Fig. 1) and also off New Zealand (Kensler, 1967). The species is considered in New South Wales to be a ‘botique’ seafood and supports a commercial trap fishery with a current Total Allowable Commercial Catch of 150 t, worth approximately $5 million.
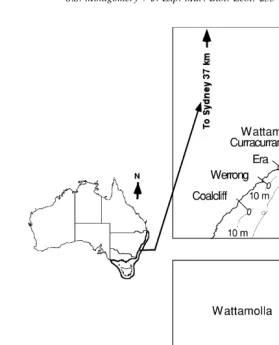
Fig. 1. Locations of sites used to investigate the effects of proximity to reef and exposure to sea-swell on catches of J. verreauxi recruits. The small map shows the distribution of J. verreauxi off Australia and the study area.
the phyllosome stage to a puerulus, probably near the outer edge of the Continental Shelf, and then actively move toward shore (Montgomery and Kittaka, 1994).
hypothesis that catches of J. verreauxi recruits will be greater on collectors set far from natural reef than on collectors set near reef. Collectors set far from reef would be the only piece of preferred habitat in the vicinity, available to the recruits. So, those animals that encounter the collector and choose to stay would have no other area of preferred habitat available to them. In contrast, collectors set near reef would be part of a greater area of habitat for recruits to settle and perhaps move within. It follows from this reasoning that the number of recruits on collectors set far from reef would be expected to be greater than on those near reef. I also present cost–benefit analyses on data from the experiment to determine the optimal numbers of collectors (replicates) and sites at each location to use in future surveys.
2. Materials and methods
2.1. Study sites
The experiment was done at two exposed (Coalcliff and Era) and two sheltered (Wattomalla and Curracurrang) sites near the New South Wales Fisheries Research Institute, the base for this study (Fig. 1). At each site, collectors were set around 5 m from vegetated reef (Near Position) or in waters around 200 m from reef and over bare sand (Far Position).
2.2. Procedure
The sea-weed-type collectors used in this study were similar to those described by Phillips (1972) for collecting pueruli of Panulirus cygnus (George, 1962). The collectors consisted of three PVC 6133530.4 mm panels supported by an aluminium frame with
four polystyrene buoys through its centre (Fig. 2). Attached to each panel were ‘tufts’ of polyethylene split rope fibre. A tuft was comprised of 80 g of 500 mm lengths of 125 text polyethylene fibre. The middle of the tuft was tied with a plastic tie, passed through a hole in the PVC panel and secured by monel wire. The collectors were moored to the substratum by attaching 20 m of 14 mm polypropylene rope to 3 m of 6 mm ‘black chain’ and a car tyre filled with cement (70 kg) laying on the substratum.
Factors in the experiment were level of Exposure (fixed), Sites (nested in Exposure) and Position (fixed). At each Position, seven replicate sea-weed-type collectors were set approximately 10 m apart and about 10 cm below the surface at each Position (i.e. a total of 56 collectors).
Fig. 2. A member of staff shows the make up of the sea-weed-type collector used in this study to catch J.
Stage 1 (puerulus): transparent;
Stage 2 (puerulus): transparent, but with digestive gland evident;
Stage 3 (puerulus): cephalothorax cream to brown in colour, digestive gland no longer visible;
Stage 4 (puerulus): whole body brown, but no spines on the cephalothorax; Stage 5 (first instar juvenile): whole body dark brown, spines on the cephalothorax
and pleopods smaller than in stages 1–4; and
Stage 6 (early juvenile): whole body green, spines on the cephalothorax and at a size less than 30 mm CL that indicated that the animal had recruited this year
For the purposes of this paper recruits are defined as lobsters in stages 1–6, inclusive.
2.3. Cost–benefit analyses
After the experiment was completed, cost–benefit analyses were done to determine the optimal numbers of sites, and collectors within sites, needed to measure the relative abundance of recruits at a location. Analyses were done on a subset of data made up of the catch-rates of recruits for collectors set at the Far Position, because it became evident during the course of this experiment that it was easier to sample collectors at the Far Position and catch-rates of recruits at Far Positions were greater than those at Near Positions. These data were analysed in a two-factor nested analysis of variance to collect information on variances. The standard cost–benefit analysis procedure was followed (Snedecor and Cochran, 1967; Underwood, 1981). The product of the variance of the estimated mean of each sampling period and the total cost of each sampling period was minimised. The variances of the means of sites and collectors were calculated by dividing the appropriate mean square by the number of readings in each mean. Variance (V ) for the mean number of recruits per location is given by
2 2
V5s 1n s /n n (3.1)
e e s e s
2 2
where n , n are the numbers and s , s , the variances among collectors and sites,
e s e s
respectively.
The total cost (C ) of sampling at one time in one location is given by
C5C n 1C n n (3.2)
s s e e s
where n and n are as above and C and C are the costs of each collector or site,e s e s respectively.
The most efficient sampling is that which reduces V and C to minima. The minimum value of the product, VC, for different sample sizes can be found by differentiating VC with respect to n and setting the differential to zero to find a minimum. This results ine the following solution:
2 2
n 5œ(Cs ) /(C s ) (3.3)
e s e e s
P3S(E) 2 0.01 ns,E 0.004 ns,E 0.003 ns
Residual 29 0.01 0.01 0.00
Total 55
a
The degrees of freedom for the residual have been adjusted for missing replicates. Higher-order effects have been pooled (P.0.25). ns, non-significant (P.0.05); *significant (P,0.05); **significant (P,0.01), ns,E, term was non-significant in original model but pooled with residual; *,ns, term was significant in original model but non-significant after pooling procedures; ns,*, term was non-significant in original model but significant after pooling procedures. Cochran’s test, C.
collectors at sea on 1 day (300 min). It cost 40 min to do each site. This was determined by measuring the time it took to prepare and travel to each site during this sampling. Similarly, I determined that it cost 10 min to sample each collector.
3. Results
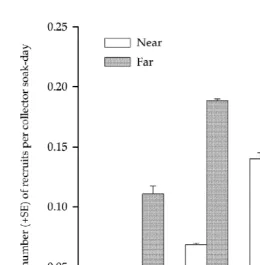
A total of 512 recruits of J. verreauxi were caught during the experiment. Mean catch-rates of recruits from collectors set at the Position far from reef were greater than those from collectors at the Position near reef (Tables 1 and 2, Fig. 3). When data were analysed separately for pueruli and for the sum of first instar and early stage juveniles (referred to below as ‘juvenile stages’), there were differences in the spatial patterns between these two categories of development. Catch-rates of pueruli were greater on collectors set at the Position far from reef, whereas there were no significant differences in catch-rates of juvenile stages between Positions.
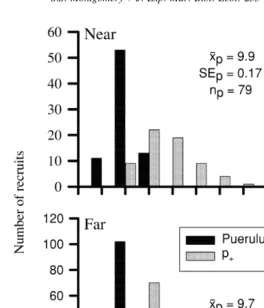
Lengths of pueruli ranged between 9 and 11 mm CL, whilst those of juvenile stages were between 9 and 19 mm CL (Fig. 4). Most individuals within the juvenile stages
Table 2
a SNK comparisons of mean numbers (S.E.) for Position (P50.05)
Near Far
Recruits 0.08 (0.01) , 0.16 (0.01) Pueruli 0.04 (0.01) , 0.09 (0.02)
a
Fig. 3. Mean catch-rates (1S.E.) of J. verreauxi recruits sampled from collectors at locations exposed (E) or sheltered (S) from sea-swell and at Positions Near or Far (200 m) from reef.
category were shorter than 13 mm CL and those longer than 15 mm CL occurred only on collectors at the Position near reef.
3.1. Cost–benefit analyses
The variance among sites and collectors and the derived optimal number of collectors and sites to be sampled are shown in Table 3. Having determined the appropriate numbers of replicate sites and collectors, the standard error of the mean of the data for one sample period may be estimated as the square root of the variance calculated in Eq. (3.1). Results suggested that, for one location, it was optimal to sample three collectors at each of three sites. This provided an estimate of mean relative abundance with a standard error of around 20% of the mean, for one sampling time at one location.
4. Discussion
Fig. 4. Distributions of lengths of J. verreauxi recruits grouped into the categories of pueruli (p) and juvenile ]
stages (p ), sampled from collectors set at Positions Near or Far from reef. The mean (x), standard error (S.E.)1
and number sampled (n) for each category of recruits are shown.
Table 3
Summary of cost–benefit analyses to determine the optimal number of sites at the far Position per location and collectors per site. Shown also are the standard error (S.E.), mean rate of catch of recruits and the level of precision (S.E. / mean) associated with the optima. Data are the subset for collectors set far from reef Summary of cost–benefit
analyses
Variance among sites 0.001 Variance among collectors 0.005 Optimal number of sites 3 Optimal number of collectors 3
Estimated S.E. 0.028
the nearness of the collectors to vegetated reefs, but not by sea-swell. The relative abundance of pueruli on collectors set further offshore from reef was greater than on collectors near reef.
Possibly more pueruli were caught on collectors set further offshore because these collectors were the first piece of preferred habitat (Herrnkind et al., 1994) encountered by the pueruli as they moved shorewards from the edge of the Continental Shelf and because the collectors further offshore were an ‘island’ of preferred habitat, away from natural reef habitat. Some of the J. verreauxi pueruli moving across the Continental Shelf towards shore would encounter the collectors further offshore before those near reef and as a consequence would stay on this first piece of preferred habitat. This hypothesis is supported by the results from studies on other marine species show that animals settle in the first piece of preferred habitat they encounter (Bell and Westoby, 1986; Sogard, 1989; Kenyon et al., 1999). Pueruli choosing to stay on collectors further offshore would not have any other preferred habitat in the vicinity to move to and therefore would be more likely to stay on the collectors than those encountering collectors near reef, where the collector possibly became part of a greater area of preferred habitat. Hence, in the latter case the pueruli would have a greater space in which to settle and perhaps move between the collector and the natural reef. Whether the pueruli are actually attracted to the collector or whether it is a random encounter remains unclear.
Lengths of J. verreauxi recruits found in this study were longer than those reported by Booth and Phillips (1994) as being common for shallow water Palinurids. In light of these larger sizes and considering that J. verraeuxi has been reported to be the largest shallow water Palinurid (Phillips et al., 1980), it is possible that this species has a faster growth rate than other shallow water spiny lobsters. Several factors or combinations of factors may account for the fact that some longer recruits were found on collectors near reef, including: that (i) the recruits move off the collectors far from reef at shorter lengths than those on collectors near reef; (ii) there is a higher natural mortality amongst recruits on collectors far from reef; and (iii) recruits move between natural reef and collectors nearby.
Exposure to oceanic swell did not affect the catch-rates of recruits on collectors. This is supported by the personal observation that recruits were not easily dislodged from collectors, as they burrow deep into the tuft of fibre and grasp it tightly.
NSW Fisheries Research Institute. [AU]
References
Bell, J.D., Westoby, M., 1986. Variation in seagrass height and density over a wide spatial scale: effects on common fish and decapods. J. Exp. Mar. Biol. Ecol. 104, 275–295.
Booth, J.D., 1986. Recruitment of packhorse lobster Jasus verreauxi in New Zealand. Can. J. Fish. Aquat. Sci. 43, 2212–2220.
Booth, J.D., Phillips, B.F., 1994. Early life history of spiny lobster. Crustaceana 66, 271–294.
Caughley, G.C., Sinclair, R., Scott-Kemmis, D., 1976. Experiments in aerial survey. J. Wildl. Man. 40, 290–300.
Childers, C.C., Brecht, J.K., 1996. Coloured sticky traps for monitoring Frankliniella bispinosa (Morgan) (Thysanoptera thripidae) during flowering cycles in citrus. J. Econ. Entomol. 89, 1240–1249.
Cruz, R., de Leon, M.E., Diaz, E., Brito, R., Puga, R., 1991. Reclutamiento de puerulus de langosta (Panulirus
argus) a la plataforma Cubana. Rev. Invest. Mar. 12, 66–75.
Donaldson, W.E., Byersdorfer, S., Blau, F., 1991. Use of artificial collectors to study growth of small red king crab. J. Shell. Res. 10, 305.
Goldberg, R., Widman, J.C., 1989. Settlement and recruitment of Mercenaria mercenaria in Long Island Sound, Connecticut. J. Shell. Res. 8, 456.
Herrnkind, W.F., Butler, IV M.J., 1994. Settlement of spiny lobster, Panulirus argus (Latreille, 1804) in Florida: pattern without predictability? Crustaceana 67, 46–64.
Herrnkind, W.F., Jernakoff, P., Butler, IV M.J., 1994. Puerulus and post-puerulus ecology. In: Phillips, B.F., Cobb, J.S., Kittaka, J. (Eds.), Spiny Lobster Management. Fishing News Books, Oxford, pp. 213–219. Kennelly, S.J., 1989. Effects of soak-time and spatial heterogeneity on sampling populations of spanner crabs
Ranina ranina. Mar. Ecol. Prog. Ser. 55, 141–147.
Kennelly, S.J., Underwood, A.J., 1984. Underwater microscopic sampling of a sublittoral kelp community. J. Exp. Mar. Biol. Ecol. 76, 67–78.
Kennelly, S.J., Graham, K.J., Montgomery, S.S., Andrew, N.L., Brett, P.A., 1993. Variance and cost–benefit analyses to determine optimal duration of tows and levels of replication for sampling relative abundances of species using demersal trawling. Fish. Res. 16, 51–67.
Kensler, C.B., 1967. The distribution of spiny lobsters in New Zealand waters (Crustacea: Decapoda: Palinuridae). N.Z. J. Mar. Freshwater Res. 1, 412–420.
Kenyon, R.A., Haywood, M.D.E., Heales, D.S., Loneragan, N.R., Pendrey, R.C., Vance, D.J., 1999. Abundance of fish and crustacean postlarvae on portable artificial seagrass units: daily sampling provides quantitative estimates of the settlement of new recruits. J. Exp. Mar. Biol. Ecol. 232, 197–216.
Killgore, K.J., Baker, J.A., 1996. Patterns of larval fish abundance in a bottomland hardwood wetland. Wetlands 16, 288–295.
Kittaka, J., 1994b. Culture of phyllosomas of spiny lobster and its application to studies of larval recruitment and aquaculture. Crustaceana 66, 258–270.
MacWilliam, P.S., Phillips, B.F., 1987. Distinguishing the phyllosoma larvae of rock lobster species of the genus Jasus (Decapoda, Palinuridae) in the waters of Australia and New Zealand. Crustaceana 52, 1–24. Maelzer, D., Zalucki, M.P., Laughlin, R., 1996. Analysis and interpretation of long-term light trap data for
Helicoverpa punctigera (Lepidoptera: noctuidae) in Australia: population changes and forecasting pest
pressure. Bull. Entomol. Res. 86, 547–557.
Montgomery, S.S., 1992. Sizes at first maturity and at onset of breeding in female Jasus verreauxi (Decapoda: Palinuridae) from New South Wales, Australia. Aust. J. Mar. Freshwater Res. 3, 1373–1379.
Montgomery, S.S., 1998. Relative abundance of the eastern rock lobster, Jasus verreauxi. PhD thesis, University of Sydney, Sydney.
Montgomery, S.S., Kittaka, J., 1994. Occurrence of pueruli of Jasus verreauxi (H. Milne Edwards, 1851) (Decapoda, Palinuridae) in waters off Cronulla, New South Wales, Australia. Crustaceana 67, 65–70. Montgomery, S.S., Craig, J., 1997. A strategy for measuring the relative abundance of pueruli of the spiny
lobster Jasus verreauxi. In: Hancock, D.A., Smith, D.C., Grant, A., Beumer, J.P. (Eds.), Developing and Sustaining World Fisheries Resources: the State of Science and Management. CSIRO, Collingwood, Victoria, Australia, pp. 574–578.
Phillips, B.F., 1972. A semi-quantitative collector of the puerulus larvae of the western rock lobster Panulirus
longipes cygnus George (Decapoda, Palinuridae). Crustaceana 22, 147–154.
Phillips, B.F., 1986. Prediction of commercial catches of the western rock lobster Panulirus cygnus. Can. J. Fish. Aquat. Sci. 43, 2126–2130.
Phillips, B.F., Cobb, J.S., George, R.W., 1980. General biology. In: Cobb, J.S., Phillips, B.F. (Eds.). The Biology and Management of Lobsters, Vol. I. Academic Press, New York, pp. 1–82.
Phillips, B.F., Booth, J.D., 1994. Design, use and effectiveness of collectors for catching the puerulus stage of spiny lobsters. Rev. Fish. Sci. 2, 255–289.
Robinette, W.L., Loveless, C.M., Jones, D.A., 1974. Field test of strip census methods. J. Wildl. Man. 38, 81–96.
Snedecor, G.W., Cochran, W.G., 1967. Statistical Methods. Iowa State University Press, Ames, IA. Sogard, S.M., 1989. Colonization of artificial seagrass by fishes and decapod crustaceans: importance of
proximity to natural eelgrass. J. Exp. Mar. Biol. Ecol. 133, 15–37.
Underwood, A.J., 1981. Techniques of analysis of variance in experimental marine biology and ecology. Oceanogr. Mar. Biol. Annu. Rev. 19, 513–605.
Underwood, A.J., 1997. Experiments in Ecology: Their Logical Design and Interpretation Using Analysis of Variance. Cambridge University Press, Cambridge.