ISSN : 2302 – 8226
12
ANTAGONISME BACILLUS TERHADAP FUSARIUM OXYSPORUM F.SP.CEPAE PENYEBAB BUSUK PANGKAL BAWANG PUTIHDwi Hermawan1), Zainal Djauhari Fatawi2), Hadiwiyono2) 1)
Undergraduate Student of Study Program of Agrotechnology, Faculty of Agriculture the University of Sebelas Maret (UNS) in Surakarta.
2)
Lecturer Staff at Study Program of Agrotechnology, Faculty of Agriculture the University of Sebelas Maret (UNS) in Surakarta
Contact Author: hadi_hpt@yahoo.com ABSTRACT
Basal rot on garlic caused by Fusarium oxysporum f.sp.cepae (FOCe) has become a disease of disadvantage and threaten planting garlic in Tawangmangu Karanganyar, Jawa Tengah which is being a new constraint since the planting season of 2000 until now. Disease control of basal rot on garlic is very difficult to control because the pathogen can hold on the soil. One alternative to control this disease is to utilize microbial biological control. Bacillus is one of microbiological control agens which have antagonist and can reduce the progression of the disease. The research aimed to study the mechanism of inhibition and physiological properties of Bacillus in vitro. there were 20 isolates of Bacillus were tested in vitro they are : antagonism, filtrate, chitinase, pectinase, and volatile tests. This research showed that Bacillus were able to anthagonist (inhibit) the growth of FOCe colony in vitro. Antagonism showed by mechanism of inhibition through antibiosis, chitinase and pectinase.
Keywords: Basal rot, FOCe, Bacillus, anthagonism. AGROTECHNOLOGY RESEARCH JOURNAL
Hermawan D, Fatawi ZD, Hadiwiyono. 2014. Bacillus antagonism toward Fusarium oxysporum f.sp.cepae caused basal rot on garlic. Agrotech Res J 3(2): 12-17.
Hermawan D, Fatawi ZD, Hadiwiyono. 2014. Antagonisme Bacillus terhadap Fusarium oxysporum f.sp.cepae penyebab busuk pangkal bawang putih. Agrotech Res J 3(2): 12-17.
PENDAHULUAN
Penggunaan bawang putih sudah cukup luas terutama sebagai bumbu masakan guna menambah citarasa makanan, bahan pelengkap untuk makanan dan obat-obatan penyakit tertentu. Karena banyaknya penggunaan, tentunya harus diimbangi dengan peningkatan total produksi. Produktivitas bawang putih di Indonesia dari 2008 sampai 2012 menurun hingga 16,50 %. Penurunan ini salah satunya disebabkan oleh Fusarium oxysporum f.sp. cepae (FOCe) penyebab busuk pangkal bawang putih (DEPTAN 2012). Jamur ini dapat menyebabkan kerugian besar terutama pada varietas yang rentan dan pada kondisi lingkungan yang sesuai untuk jamur tersebut (Agrios 2005). Menurut Hadiwiyono (2009) busuk pangkal bawang putih merupakan penyakit yang merugikan dan mengancam pertanaman bawang putih Tawangmangu, Karanganyar, Jawa Tengah. Lebih dari 92 % lahan penanaman bawang putih di daerah tersebut telah terjangkit Fusarium oxysporumf.sp.cepae.
Pengendalian penyakit busuk pangkal bawang putih secara konvensional menimbulkan banyak dampak negative, sehingga perlu alternatif lain untuk mengendalikan penyakit busuk pangkal bawang. Salah satunya dengan memanfaatkan mikrobia agens pengendali hayati. Pengendalian dengan cara ini cukup efektif dan belum ada yang melaporkan timbulnya
ketahanan jamur 15ampak15e terhadap agens pengendali hayati (Freeman et al. 2002). Bacillus merupakan agens pengendali hayati yang memiliki aktivitas antifungal yang tinggi dan berperan dalam menekan beberapa fungi yang bersifat 15ampak 15e, seperti Rhizoctonia, Fusarium dan Aspergillus. Selain memiliki kemampuan dalam menekan perkembangan fitopatogen, Bacillus juga diketahui dapat mendukung pertumbuhan tanaman (Zhang dan Tambong 2009). Bacillus merupakan bakteri rizosfer yang memiliki efek positif terhadap tanaman inang dan dapat menghambat patogen terutama patogen tular tanah (Jos et al. 2009).Penelitian tentang eksplorasi dan karakter Bacillus perlu dipelajari untuk mengetahui seberapa besar efektifitas dan pengaruh penghambatannya terhadap Fusarium oxysporum f.sp.cepae(FOCe). BAHAN DAN METODE
Isolasi, koleksi dan karakterisasi Bacillus
Isolasi dan koleksi Bacillus dilakukan di Laboratorium Hama Penyakit Tumbuhan pada bulan November sampai Desember 2013. Isolat Bacillus di daerah perakaran bawang putih merupakan hasil eksplorasi dari beberapa tempat di wilayah Tawangmangu, Karanganyar.
*Fak. Pertanian UNS Surakarta Jl. Ir. Sutami 36 A Surakarta
Antagonisme Bacillus terhadap Fusarium Oxysporum F.Sp.Cepae Penyebab Busuk Pangkal Bawang Putih (Dwi Hermawan,Zainal Djauhari Fatawi, Hadiwiyono)
Hermawan et al. Vol 3. No 2. 2014
13
Uji produksi kitinase dan pektinase oleh Bacillus Uji Kitinase dan pektinase dilakukan dengan cara setiap isolat Bacillus ditumbuhkan dalam King’s B cair selama 24 jam. Masing-masing isolat diletakkan pada cawan petri yang berisi agar kitin dan agar pektin (Salvador et al. 2005) sebagai perlakuan serta agar tanpa bahan aktif pektin dan kitin sebagai kontrolnya. Uji diamati secara kualitatif dilihat adanya pertumbuhan koloni diameter disekitar isolat.
Uji antagonisme Bacillus terhadap FOCe in vitro Antagonisme invitro menggunakan rancangan acak lengkap (RAL) dengan metode kultur ganda Bacillus terhadap FOCe. Persentase hambatan dihitung:
dengan r1 = jari FOCemenjauhi Bacillus, r2 = jari-jari FOCe mendekati Bacillus. Analisis data dengan uji F taraf 5% dan uji jarak berganda Duncan taraf 5%.
Uji filtrat Bacillus terhadap FOCe in vitro
Uji Filtrat Bacillus disusun menggunakan RAL dengan metode kultur ganda pada 20 isolat Bacillus yang telah dinonaktifkan melalui sterilisasi, dan penyinaran UV.
Uji produksi senyawa volatil Bacillus terhadap FOCe in vitro
Uji produksi senyawa volatil (uap biakan) dilakukan menggunakan RAL dengan bahan 20 isolat Bacillus dan FOCe sebagai perlakuan serta kontrol berupa air steril. Masing-masing isolat ditangkupkan satu sama lain pada media biakan yang berbeda, NA untuk Bacillus dan PDA untuk FOCe. Pertumbuhan diameter koloni FOCediamati setiap 2 hari sekali selama 14 hari. Analisis data dengan uji F taraf 5% dan uji jarak berganda Duncan taraf 5%.
HASIL DAN PEMBAHASAN Karakterisasi koloni Bacillus
Karakterisasi Bacillus dilakukan dengan pengamatan morfologi koloni secara langsung pada media Nutrient Agar (NA) dalam petri dan pengecatan gram. Bacillus berdasarkan pengamatan morfologi koloninya dapat dilihat dari bentuk, tepi, permukaan, elevasi dan warna isolat yang yang beragam (Tabel 1). Hasil pengamatan menunjukkan bahwa koloni Bacillus didominasi oleh bentuk Circular dengan tepi Entire, yang memiliki permukaan Smooth, Glistening dan elevasi Umbonate, serta berwarna putih, sedangkan untuk pewarnaan gram hasilnya menunjukkan bahwa
isolat yang mempunyai gram positif yang ditunjukkan dengan warna biru (Bacillus), sedangkan gram negatif ditunjukkan dengan warna merah (dengan menggunakan kontrol BDB).
Perbedaan koloni isolat Bacillus dikarenakan perbedaan tempat di wilayah Tawangmangu, Karanganyar. Bakteri yang nampak, dapat memiliki morfologi yang sama, namun keperluan nutrisi dan persyaratan ekologinya. Menurut Irianto (2007) menjelaskan juga, untuk pengamatan morfologi bakteri dengan jelas, tubuhnya perlu diisi dengan cat warna, pewarnaan ini disebut pengecatan gram pada bakteri. Tabel 1 Karakterisasi morfologi koloni isolat Bacillus dari tanaman bawang
Isolat Asal Karakter Koloni
Bentuk Tepi Elevasi Permukaan Warna
B1 Pancot Circular Entire Umbonate Smooth, Glistening Putih
B2 Pancot Punctiform Entire Umbonate Smooth, Glistening Putih
B3 Pancot Irregular Undulate Umbonate Smooth, Glistening Putih
B4 Pancot Punctiform Undulate Umbonate Smooth, Glistening Putih
B5 Pancot Circular Entire Umbonate Smooth, Glistening Putih
B6 Kalisoro Irregular Undulate Umbonate Smooth, Glistening Putih
B7 Kalisoro Circular Entire Raised Smooth, Glistening Putih
B8 Kalisoro Circular Entire Umbonate Smooth, Glistening Putih
B9 Kalisoro Circular Entire Umbonate Smooth, Glistening Putih
B10 Kalisoro Filamentous Filamentous Umbonate Rough Putih
B11 Gondosuli Filamentous Filamentous Umbonate Wringkled Putih
B12 Gondosuli Rhizoid Lobate Raised Smooth, Glistening Putih
B13 Gondosuli Circular Entire Raised Smooth, Glistening Putih
B16 Gondosuli Irregular Lobate Umbonate Smooth, Glistening Putih
B17 Gondosuli Irregular Undulate Raised Smooth, Glistening Putih
B18 Gondosuli Circular Entire Umbonate Smooth, Glistening Putih
B19 Blumbang Irregular Undulate Raised Smooth, Glistening Putih
B20 B21 Blumbang Blumbang Filamentous Circular Filamentous Entire Umbonate Umbonate Rough Smooth, Glistening Putih Putih
14
Uji produksi kitinase dan pektinase oleh BacillusUji produksi kitinase dan pektinase merupakan karakterisasi secara fisiologi pada isolat Bacillus secara kulitatif. Hasil uji menunjukan bahwa semua isolat Bacillus positif menghasilkan enzim kitinase dan pektinase (Tabel 2) dengan ditandai terbentuknya koloni disekitar isolat dengan media kitin dan pektin. Aktivitas kitinase dapat mendegradasi dinding sel jamur, karena jamur memiliki kitin pada dinding selnya. Hal ini sesuai dengan pernyataan Cabib (1987) dalam
Malik (2000) dan hasil penelitian Chen et al. (2004) yang menyebutkan bahwa kitinase merupakan enzim yang dapat mendegradasi kitin yang mana di alam terdapat sangat melimpah dan juga merupakan komponen utama penyusun dinding sel jamur. Kamil et al. (2007) juga melaporkan bahwa Bacillus efektif dalam menekan Fusarium karena memproduksi enzim kitinase yang mana dapat merombak kitin dari jamur tersebut.
Tabel 2 Hasil uji aktifitas kitinase dan pektinase pada Bacillus
No Perlakuan Aktivitas Kitinase *) Aktivitas Pektinase**) Kontrol ***)
1 B1 + + - 2 B2 + + - 3 B3 + + - 4 B4 + + - 5 B5 + + - 6 B6 + + - 7 B7 + + - 8 B8 + + - 9 B9 + + - 10 B10 + + - 11 B11 + + - 12 B12 + + - 13 B13 + + - 14 B16 + + - 15 B17 + + - 16 B18 + + - 17 B19 + + - 18 B20 + + - 19 B21 + + - 20 B22 + + -
Keterangan: tanda (+) menunjukan adanya aktivitas enzim kitinase dan atau pektinase, tanda (-) menunjukan tidak ada aktivitas enzim kitinase dan atau pektinase, *) media dengan penambahan zat aktif kitin, **) media dengan penambahan zat aktif pektin (citrus pektin), ***) Tanpa penambahan zat aktif kitin maupun pektin.
Uji antagonisme Bacillus terhadap FOCe secara In Vitro
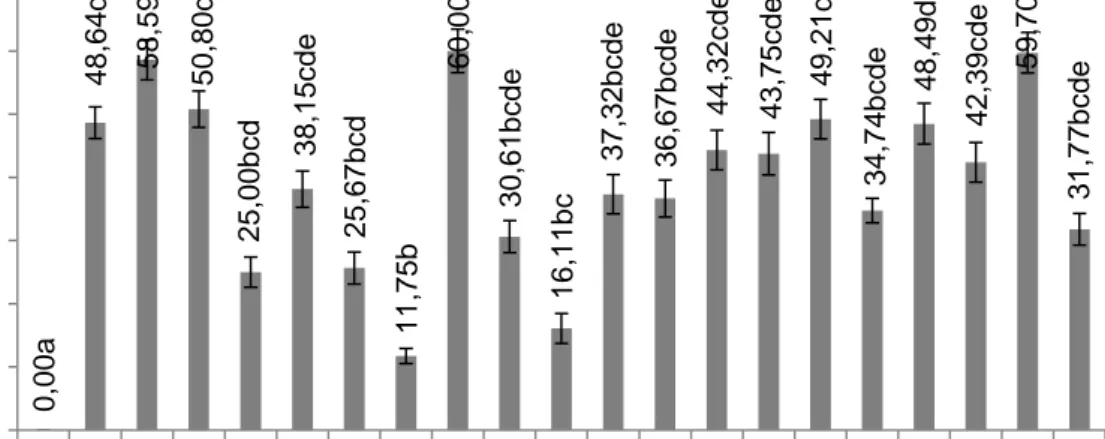
Hasil antagonisme in vitro menunjukan bahwa semua isolat Bacillus mampu menghambat diameter koloni Fusarium oxysporum f.sp.cepae (FOCe). Persentase hambatan tertinggi dihasilkan oleh isolat B8 (60.00 %) disusul oleh B21 (59,70 %) dan B2 (58,59 %) sedangkan terendah dihasilkan oleh isolat B7 (11,75%). Salah satu mekanisme uji antagonisme ini yaitu dengan adanya senyawa antifungal yang dihasilkan oleh bakteri secara umum mengakibatkan terjadinya pertumbuhan yang abnormal pada hifa
(malformasi), yang ditunjukkan dengan pembengkak-an dan pemendekan hifa yang mengakibatkan hifa tidak dapat berkembang dengan sempurna (Eliza et al. 2007).
Chen dan Michailidaes (2004) melaporkan bahwa pada uji kultur ganda, penghambatan ditunjuKkan dengan adanya zona bening. Zona bening ini terjadi karena adanya senyawa antifungal yang dihasilkan oleh
Bacillus. Penghambatan pertumbuhan miselium
disebabkan karena adanya aktivitas antibiosis oleh bakteri antagonis (Bacillus).
Uji filtrat Bacillus terhadap FOCe secara In Vitro Hasil uji filtrat menunjukan bahwa semua isolat mampu menekan pertumbuhan FOCe, persentase penghambatan paling tinggi adalah isolat B18 (67,16 %) disusul dengan B3 dan B4 (64.38 %), sedangkan persentase terendah yaitu B1 (22.70 %). Hasil mengindikasikan semua isolat Bacillus dapat
memproduksi senyawa antibiotik yang mampu menekan pertumbuhan koloni FOCe. Hal ini didukung dengan pernyataan Eliza et al. (2007) bahwa penghambatan pertumbuhan miselium disebabkan adanya aktivitas antibiosis bakteri antagonis. Bacillus dapat menghasilkan antibiotik lebih dari 66 jenis yang bersifat racun terhadap mikroba lain salah satunya yaitu basitrasin yang dapat menghambat pembentukan
Hermawan et al. Vol 3. No 2. 2014
13
dinding sel pathogen, serta berbagai jenis bakteriosin diantaranya subtilin, ericin, mersacidin, sublancin, dan
subtilosin A (Mochizuki et al. 2005, Soesanto et al. 2008, Stein et al. 2004, Stein 2005).
Keterangan : Angka yang diikuti huruf sama tidak berbeda nyata pada uji jarak berganda Duncan (P<0,05),tanpa = Air steril, Bx = FOCe dengan Bacillus ke-x (x = nomor isolat).
Gambar 1 Histogram antagonisme isolat Bacillus terhadap pertumbuhan koloni FOCe. Pengaruh senyawa volatil pada Bacillus terhadap
FOce secara in vitro
Uji volatil FOCe pada media biakan PDA yang ditangkupkan di atas biakan isolat Bacillus pada media biakan NA menunjukkan diameter koloni patogen yang bervariasi. Isolat FOCe tanpa Bacillus mampu tumbuh secara maksimal pada media biakan dengan diameter rata-rata 9 cm atau penuh pada petri dengan inkubasi 14 hari, namun perlakuan yang ditangkupkan dengan Bacillus menunjukan diameter yang berbeda-beda. Hal ini menunjukkan isolat Bacillus mampu
menghenghambat pertumbuhan FOCe yang ditandai dengan nilai diameter koloni yang bervariasi.
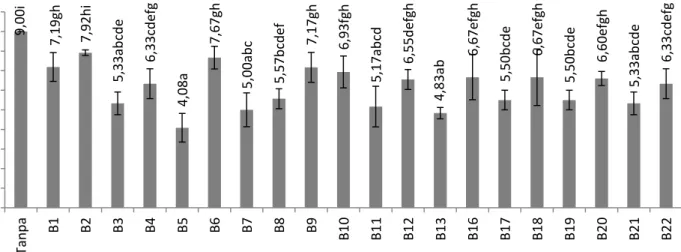
Hasil uji volatil menunjukan Isolat B5 mampu menekan kecepatan pertumbuhan FOCe secara maksimal dengan diameter koloni 4,08 cm. Hal tersebut menandakan ada zat penghambat berupa gas yang perannya menghambat pembentukan koloni FOCe. Sudhanta et al. (2011) menambahkan, terhambatnya koloni jamur FOCe diduga karena agens antagonis baik dari kelompok bakteri atau jamur mampu mengeluarkan antibiotik yang mudah menguap.
Keterangan: Angka yang diikuti huruf sama tidak berbeda nyata pada uji jarak berganda Duncan (P<0,05),tanpa = Air steril, Bx = FOCe dengan Bacillus ke-x (x = nomor isolat).
Gambar 2 Histogram pengaruh filtrat Bacillus terhadap pertumbuhan koloni FOCe.
0,00a
48,64de 58,59e 50,80de
25 ,00 bc d 38 ,15 c de 25 ,67 bc d 11,75b 60,00e 30 ,61 bc de 16,11bc 3 7 ,3 2 b c d e 36 ,67 bc de 44 ,32 c de 43 ,75 c de 49,21de 34 ,74 bc de 48,4 9 d e 42 ,39 c de 59,70e 31 ,77 bc de 0.00 10.00 20.00 30.00 40.00 50.00 60.00 70.00 ta np a B1 B2 B3 B4 B5 B6 B7 B8 B9 B 1 0 B 11 B 12 B 13 B 16 B 17 B 18 B 19 B 20 B 21 B 22 p er se n tas e Hamb atan ( % ) isolat 0,00a 22,70b 44,28b c 64,38c 64,38c 38,93b c 48,63b c 37,37b c 26,08b c 23 ,54bc 35,45b c 58,08b c 57,95b c 55,60b c 63,89b c 40,91b c 67,16c 39,03b c 38,93b c 63,66c 25,14b c 0.00 10.00 20.00 30.00 40.00 50.00 60.00 70.00 80.00 T an … B1 B2 B3 B4 B5 B6 B7 B8 B9 B 10 B 11 B 12 B 13 B 16 B 17 B 18 B 19 B 20 B 21 B 22 P res ent as i Hamb atan ( % ) Isolat 15
14
Keterangan : Angka yang diikuti huruf sama tidak berbeda nyata pada uji jarak berganda Duncan (P<0,05),K= Air steril, Bx = FOCe dengan Bacillus ke-x (x = nomor isolat).Gambar 3 Histogram pengaruh senyawa uap biakan (volatil) Bacillus terhadap diameter koloni FOCe. KESIMPULAN DAN SARAN
Semua isolatBacillus asal beberapa di Tawangmangu antagonis terhadap FOCe in vitro. Antagonisme Bacillus terhadap FOCe ditunjukan dengan adanya mekanisme penghambatan melalui
antibiosis, produksi senyawa kitinase dan pektinase. Sarannya, perlu penelitian lanjutan untuk mempelajari sifat fisiologi Bacillus yang lain dan asal dari daerah lain, serta aplikasi Bacillus di lapangan.
DAFTAR PUSTAKA
Agrios GN. 2005. Plant pathology 5th ed. New York (NY): Academic Press.
Cabib E. 1987. The synthesis and degradation of chitin. Di dalam: Malik A. 2000. Pengklonan gen kitinase bacterial menggunakan teknik mutagenesis transposon dan DNA pelacak heterologus [disertasi].
Bogor (ID): Program Pascasarjana, Institut Pertanian Bogor.
Chen CY, Wang YH, Huang CJ. 2004. Enhancement of the antifungal activity of Bacillus subtilis f29-3 by the chitinase encoded by Bacillus circulans chi a gene. Can J Microbiol 50:451-454. DOI : 10.1139/w04-027 Chen WQ, Michailides TJ. 2004. Collection and trials of
biocontrol agents against botryosphaeria panicle and shoot blight of pistachio. Postdoctoral Research Associate.
Departemen Pertanian [DEPTAN]. 2012. Isi data 5
tahun hortikultura.
http://www.deptan.go.id/infoeksekutif/horti/isi_dt 5thn_horti.php. diakses tanggal 17 Juni 2013. Eliza, Munif A, Djatnika, Widodo. 2007. Karakter
fisiologis dan peranan antibiosis bakteri perakaran Graminae terhadap Fusarium dan pemacu pertumbuhan tanaman pisang. J Hort. 17(2): 150-160. http://ejurnal.litbang.deptan.go.id/index.php/ jhort/article/view/957/825#.
Freeman S, Zveibel A, Vintal H, Maymon M. 2002. Isolation of nonpathogenic mutants of fusarium oxysporum f.sp. melonis for biological control of Fusarium wilts in cucurbits. Phytopathology 92:164-168. DOI :10.1094/PHYTO.2002.92.2.164.
Hadiwiyono, Wuspada RD, Widono S, Poromarto SH, Fatawi ZD. 2009. Kesupresifan tanah terhadap
busuk pangkal (Fusarium oxysporum f.sp.cepae) bawang putih di Tawangmangu, Karanganyar. Sains Tanah 6(1):1-6.
Irianto K. 2007. Mikrobiologi. Yrama Widya, Bandung. Jos M, Raaijmakers, Timothy C, Paulitz, Christian S,
Claude A, Yvan M. 2009. The rhizosphere: a playground and battlefield for soilborne pathogens and benefical microorganisms. Plant Soil. 321: 341-361. DOI 10.1007/s11104-008-9568-6.
Kamil Z, Rizk M, Saleh M, Moustafa S. 2007. Isolation and identification ofrhizosphere soil chitinolytic bacteria and their potential in antifungal biocontrol.
Global J Mol Sci. 2: 57-66.
http://www.idosi.org/gjms/gjms2(2)/4.pdf.
Mochizuki M, Nishijima T, Toyota K. 2005. Predominant culturable bacillusspecies in Japanese arable soils and their potential as biocontrol agents. J Microbiol Environ. 20(1): 61-68.
Salvador S, Fontana RC, Silveira MM. 2005. Influence of pectin and glucose on growth and polygalacruronase production by Aspergillus niger
9,00i 7,19gh 7,92h i 5,33a b cd e 6,33cd ef g 4,08 a 7,67gh 5,00a b c 5,57b cd ef 7,17 gh 6,93f gh 5,17a b cd 6,55d ef gh 4,83a b 6,67e fgh 5,50b cd e 6,67e fgh 5,50b cd e 6,60e fgh 5,33a b cd e 6,33cd ef g 0.00 1.00 2.00 3.00 4.00 5.00 6.00 7.00 8.00 9.00 10.00 Tan p a B1 B2 B3 B4 B5 B6 B7 B8 B9 B10 B11 B12 B13 B16 B17 B18 B19 B20 B21 B22 Diamet er P er tu mb u h an (cm) isolat 16
Hermawan et al. Vol 3. No 2. 2014
13
in solid-state cultivation. J Microbiol Biotech. 32: 371-377.
Soesanto L, Rokhlani, Prihatiningsih N. 2008. Penekanan beberapa mikroorganisme antagonis terhadap penyakit layu gladiol. J Agrivi 30(1): 75-83. http://www.researchgate.net/publication/21126 540_Penekanan_beberapa_mikroorganisme_antago nis_terhadap_penyakit_layu_Fusarium_Gladiol. Sood A, Shivesh S, Viviek K, Ram LT. 2007.
Antagonism of dominant bacteria in tea rhizosphere of Indian Himalayan regions. J Appl Sci Envi Man 11(4):63-66. http://www.bioline.org.br/pdf?ja07096. Stein T. 2005. Bacillus subtilis antibiotics : structures.
Syntheses and specific function. Mol Microbiol 56:845-857.DOI:10.1111/j.1365-2958.2005.04587.x. Stein T, Dusterhus S, Stroh A, Entian KD. 2004. Subtilosin production by two Bacillus subtilis
subspecies and variance of the sbo-alb cluster. Appl Environ Microbiol 70:2349-2353. DOI: 10.1128/AEM.70.4.2349-2353.2004.
Sudhanta IM, Kusnarta IM, Sudana IN. 2011. Uji antagonisme beberapa jenis jamur saprofit terhadap jamur Fusarium oxysporum f.sp. cubense penyebab penyakit layu pada tanaman pisang serta potensinya sebagai agens pengurai seresah. J Agrotek 21(2): 106-119. http://fp.unram.ac.id/data/ 2012/04/21-2-
3_03-I-Made-Sudantha-_Des2011_-Rev-Wangiyana-1.pdf.
Zhang JX, Tambong JT. 2009. Evaluation of seed and soil treatments with novel Bacillus subtilis strains for control of soybean root rot caused by Fusarium oxysporum and f. graminearum. Plant Dis. 93:1317-1323.DOI: 10.1094/PDIS-93-12-1317.