J. Biol. Indon. Vol 6, No.2 (2010)
Vol. 6, No. 2, Juni 2010 Akreditasi: No 816/D/08/2009 BOGOR, INDONESIA
JURNAL
BIOLOGI
INDONESIA
ISSN 0854-4425Isolasi Bakteri Pendegradasi Phenanthrene dari Batanta-Salawati Raja Ampat Papua Rini Riffiani
153
Perubahan Tutupan Hutan Mangrove di Pantai Timur Sumatera Utara Periode 1977-2006
Onrizal
163
Keragaman Genetika Ramin [Gonystylus bancanus (Miq.) Kurz] dari Provinsi Riau Berdasarkan Profil Random Amplified Polymorphic DNA
Yulita Kusumadewi, Yuyu S. Poerba, &Tukirin Partomihardjo
173
Laju Kehilangan dan Kondisi Terkini Habitat Baning Sulawesi (Indotestudo forstenii) di Semenanjung Santigi, Sulawesi Tengah, Indonesia
Awal Riyanto, Suprayogo Soemarno dan Achmad Farajallah
185
Plant- β Diversity and Composition in Mount Nok and the Waifoi Forest of the Waigeo Raja Ampat Islands: with Special Reference to The Threatened Species
Didik Widyatmoko
195
Emisi Gas Dinitrogen Oksida dari Tanah Sawah Tadah Hujan yang diberi Jerami Padi dan Bahan Penghambat Nitrifikasi
A. Wihardjaka
211
Pengelompokan Kelelawar Pemakan Buah dan Nektar Berdasarkan Karakteristik Jenis Pakan Polen di Kebun Raya Bogor, Indonesia
Sri Soegiharto, Agus P. Kartono, & Ibnu Maryanto
225
JURNAL
BIOLOGI
INDONESIA
J. Biol. Indon. Vol 6, No. 2 (2010)
Jurnal Biologi Indonesia diterbitkan oleh Perhimpunan Biologi Indonesia.
Jurnal ini memuat hasil penelitian ataupun kajian yang berkaitan dengan masalah biologi yang diterbitkan secara berkala dua kali setahun (Juni dan Desember).
Editor Pengelola
Dr. Ibnu Maryanto Dr. I Made Sudiana Dr. Anggoro Hadi Prasetyo
Dr. Izu Andry Fijridiyanto
Dewan Editor Ilmiah
Dr. Abinawanto, F MIPA UI Dr. Achmad Farajalah, FMIPA IPB
Dr. Ambariyanto, F. Perikanan dan Kelautan UNDIP Dr. Aswin Usup F. Pertanian Universitas Palangkaraya Dr. Didik Widiyatmoko, PK Tumbuhan, Kebun Raya Cibodas-LIPI
Dr. Dwi Nugroho Wibowo, F. Biologi UNSOED Dr. Parikesit, F. MIPA UNPAD
Prof. Dr. Mohd.Tajuddin Abdullah, Universiti Malaysia Sarawak Malaysia Assoc. Prof. Monica Suleiman, Universiti Malaysia Sabah, Malaysia
Dr. Srihadi Agung priyono, F. Kedokteran Hewan IPB Y. Surjadi MSc, Pusat Penelitian ICABIOGRAD
Drs. Suharjono, Pusat Penelitian Biologi-LIPI Dr. Tri Widianto, Pusat Penelitian Limnologi-LIPI
Dr. Witjaksono Pusat Penelitian Biologi-LIPI
Alamat Redaksi Sekretariat Oscar efendi SSi MSi
d/a Pusat Penelitian Biologi - LIPI
Jl. Ir. H. Juanda No. 18, Bogor 16002 , Telp. (021) 8765056 Fax. (021) 8765068
Email : [email protected]
Website : http://biologi.or.id
Jurnal ini telah diakreditasi ulang dengan nilai A berdasarkan SK Kepala LIPI 816/ D/2009 tanggal 28 Agustus 2009.
J. Biol. Indon. Vol 6, No.2 (2010)
KATA PENGANTAR
Jurnal Biologi Indonesia edisi volume 6 nomer 2 tahun 2010 yaitu memuat 11 artikel lengkap dan sebuah artikel tulisan pendek. Penulis pada edisi ini sangat beragam yaitu dari Departemen Kementerian Kehutanan, Pertanian, Fakultas MIPA IPB, Fakultas Kehutanan IPB, Fakultas. MIPA Universitas Indonesia, Fakultas Pertanian Universitas Sumatra Utara, Pusat Konservasi Kebun Raya Bogor, Pusat Penelitian Limnologi-LIPI Bogor dan Pusat Penelitian Biologi LIPI Bogor. Topik yang dibahas pada edisi ini meliputi empat topik dalam bidang Botani, dua topik tentang mikrobiologi satu topik mengenaik hasil perombakan bakteri dan bahan organik lainnya dan lima topik dalam bidang zoologi
Beragamnya penulis pada edisi ini yang membahas tiga topik utama yaitu Zoologi, Botani dan Mikrobiologi diharapkan semakin banyak keragaman pembaca dan akhir kata yang diharapkan dari editor jurnal ini akan semakin banyak penulis yang berkeinginan membagi hasil karya penelitiannya dengan menulis ke dalam Jurnal Biologi Indonesia.
J. Biol. Indon. Vol 6, No. 2 (2010)
UCAPAN TERIMA KASIH
Jurnal Biologi Indonesia mengucapkan terima kasih dan penghargaan kepada
para pakar yang telah turut sebagai penelaah dalam Volume 6, No 2, Juni 2010: Dr. Niken TM. Pratiwi, Fakultas Perikanan dan Ilmu Kelautan-IPB
Ir. Majariana Krisanti MSi, Fakultas Perikanan dan Ilmu Kelautan-IPB Onrizal MSi, Universitas Sumatra Utara
Dr.Tike Sartika, Balitnak, Departemen Pertania, Ciawi bogor Dr. Dwi Astuti, Puslit Biologi-LIPI
Drs. Edi Mirmanto MSc, Puslit Biologi-LIPI Drs. Roemantyo, Puslit Biologi-LIPI Drs. M. Noerdjito, Puslit Biologi-LIPI
Drh. Anang S. Achmadi MSc, Puslit Biologi-LIPI Sigit Wiantoro SSi ,MSc Puslit Biologi-LIPI Ir. Dwi Agustiyani MSc, Puslit Biologi-LIPI
J. Biol. Indon. Vol 6, No.2 (2010)
DAFTAR ISI
Isolasi Bakteri Pendegradasi Phenanthrene dari Batanta-Salawati Raja Ampat Papua Rini Riffiani
153
Perubahan Tutupan Hutan Mangrove di Pantai Timur Sumatera Utara Periode 1977-2006
Onrizal
163
Keragaman Genetika Ramin [Gonystylus bancanus (Miq.) Kurz] dari Provinsi Riau Berdasarkan Profil Random Amplified Polymorphic DNA
Yulita Kusumadewi, Yuyu S. Poerba, &Tukirin Partomihardjo
173
Laju Kehilangan dan Kondisi Terkini Habitat Baning Sulawesi (Indotestudo forstenii) di Semenanjung Santigi, Sulawesi Tengah, Indonesia
Awal Riyanto, Suprayogo Soemarno dan Achmad Farajallah
185
Plant- β Diversity and Composition in Mount Nok and the Waifoi Forest of the Waigeo Raja Ampat Islands: with Special Reference to The Threatened Species
Didik Widyatmoko
195
Emisi Gas Dinitrogen Oksida dari Tanah Sawah Tadah Hujan yang diberi Jerami Padi dan Bahan Penghambat Nitrifikasi
A. Wihardjaka
211
Pengelompokan Kelelawar Pemakan Buah dan Nektar Berdasarkan Karakteristik Jenis Pakan Polen di Kebun Raya Bogor, Indonesia
Sri Soegiharto, Agus P. Kartono, & Ibnu Maryanto
225
Kemampuan Kawasan Nir-Konservasi dalam Melindungi Kelestarian Burung Endemik Dataran Rendah Pulau Jawa Studi Kasus di Kabupaten Kebumen
Eko Sulistyadi
237
Analysis of Nutrient Requirement and Feed Efficiency Ratio of Maroon Leaf Monkey (Presbytis rubicunda Mueller, 1838)
Wartika Rosa Farida
255
Oksidasi Nitrit Oleh Bakteri Heterotrofik Pada Kondisi Aerobik
Dwi Agustiyani, Ruly Marthina Kayadoe & Hartati Imamuddin 265 Pencirian Karbon Organik Air Sungai Citarum Hulu Dari Masukan Air Limbah Penduduk dan Industri
Eko Harsono dan Sulung Nomosatryo
277
TULISAN PENDEK
Arti Kebun Raya Bogor Bagi Kehidupan Kumbang Sungut Panjang (Coleoptera, Cerambicidae)
Woro Anggaraitoningsih Noerdjito
173 Jurnal Biologi Indonesia 6(2): 173-183 (2010)
Keragaman Genetika Ramin [Gonystylus bancanus (Miq.) Kurz]
dari Provinsi Riau Berdasarkan Profil Random Amplified Polymorphic
DNA
Yulita Kusumadewi, Yuyu S. Poerba, &Tukirin Partomihardjo
Laboratorium Genetika Tumbuhan, Pusat Penelitian Biologi, Cibinong Science Centre Jl. Raya Bogor Km. 46 Bogor 16911
Email: [email protected]
ABSTRACT
Genetic Diversity of Ramin [Gonystylus bancanus (Miq.) Kurz] from Riau Province Based on Random Amplified Polymorphic DNA Fingerprint. Gonystylus bancanus is a commercial
timber found only on peat swamp forests, scatteredly distributed in Sumatra and Kalimantan. Their existence is now under severe threat due to habitat conversion. One of the remaining natural populations of ramin was in Riau Province, Sumatra. This study aimed to assess genetic diversity of this species within their natural populations in Riau Province using Random Amplified Polymorphic DNA (RAPD). RAPD profiles were obtained by performing PCR amplification using five arbitrary primers. One hundred and eleven putative loci of RAPD were scored and analysed using Popgene and NTSYS software. Eleven of RAPD bands were commonly found in all populations and 16 bands were distinctively found in certain populations. These unique bands may serve as population diagnostic marker for such populations. The average genetic diversity within population (0.1606) was lower than that of among populations (0.1894). Genetic differentiation (Gst) indicated that 95.56% of total genetic diversity in ramin was attributed to the differences among populations. The highest genetic diversity was found in population 3 (He:0.1858) and 3 (I:0.2864), while the lowest genetic variation was observed in population 1 (He: 0.1438) and 2 (I: 0.2201). Total genetic diversity for all population (Ht) was 0.1982 with an average value of genetic diversity within populations (Hs) was 0.1606. The low level of genetic diversity found in ramin with high population differentiation may suggest that these remaining populations was undergoing genetic bottleneck resulted from severe habitat fragmentation.
Keywords: genetic diversity, populations, ramin, Gonystylus bancanus, RAPD.
PENDAHULUAN
Ramin (Gonystylus bancanus, Thymelaeaceae) merupakan salah satu jenis tumbuhan kayu komersial yang menjadi primadona dalam dunia perdagangan kayu. Negara penghasil kayu ramin yang potensial saat ini hanya
Malaysia dan Indonesia. Kayu ramin diperdagangkan dalam berbagai produk seperti papan interior, komponen perabotan, pigura, lantai, pasak kayu dan mainan anak-anak. Jenis-jenis ramin ini hanya tumbuh di hutan gambut dan tersebar secara mengelompok di Sumatra dan Kalimantan (Jayusman 2007). Salah
174
Kusumadewi, Poerba, & Partomihardjo
satu populasi alam ramin yang masih tersisa adalah di area hutan konsesi Provinsi Riau, Sumatra.
Seperti halnya dengan jenis-jenis kayu komersil lainnya, jenis ini banyak dieksploitasi untuk diambil kayunya sehingga keberadaannya terancam punah. Hal ini diperburuk dengan kenyataan bahwa ramin memiliki siklus regenerasi yang sangat sulit dan lambat, baik di habitat alaminya maupun di area percobaan. Kesulitan dalam mengem-bangkan ramin melalui kultur jaringan juga dialami oleh Laboratorium Kultur Jaringan di Bidang Botani Puslit Biologi-LIPI. Ramin dikategorikan sebagai jenis terancam dengan status rawan (VuA1cd IUCN 2008) dan dimasukkan dalam Appendix II CITES (2009).
Studi keragaman genetika dilakukan untuk mengetahui variasi genetika pada tingkat individu maupun populasi yang salah satunya untuk tujuan konservasi genetik, populasi maupun jenis. Studi ini telah dilakukan dengan menggunakan berbagai marka baik molekuler (Random Amplified Polymorphic DNA (RAPD), Amplified Fragment Length Polymor-phism (AFLP), Random Fragment Lenght Polymorphism (RFLP), Simple Sequence Repeats (SSRs) maupun protein (isozyme dan allozyme). Beberapa studi sudah dilakukan untuk memperkirakan keragaman genetika ramin, termasuk mengembangkan marka mikrosatelit untuk melacak asal usul lokasi ramin untuk mencegah perdagangan kayu illegal (Smulders et al. 2007) dan pengembangan protokol ekstraksi kayu olahan ramin (Asif & Cannon 2005). Penelitian ini bertujuan
untuk mengetahui keragaman genetika antar individu dan populasi ramin di kawasan propinsi Riau dengan menggunakan marka RAPD.
RAPD adalah marka molekuler yang telah banyak digunakan untuk identifikasi genotipe (Jimenez et al. 2002; Poerba et al. 2007) dan kultivar (Malik
et al. 2006; Shabaan et al. 2006; Jain et al. 2007) serta memperkirakan
keraga-man genetika jenis-jenis pohon kayu tropis (Siregar et al. 2008; Rath et al. 1998; Poerba et al. 2007). Keuntungan utama dari RAPD adalah menghasilkan polimorfisme yang cukup tinggi, random
sampling dalam genom total dan secara
teknis cukup cepat dan mudah dilakukan. Kekurangan dari penggunaan marka RAPD yaitu marka ini hanya mengam-plifikasi alel dominan dan memiliki tingkat keberulangan yang rendah. Hal ini dapat diatasi antara lain dengan melakukan amplifikasi PCR lebih dari satu kali untuk memastikan konsistensi hasil yang diperoleh.
BAHAN DAN CARA KERJA
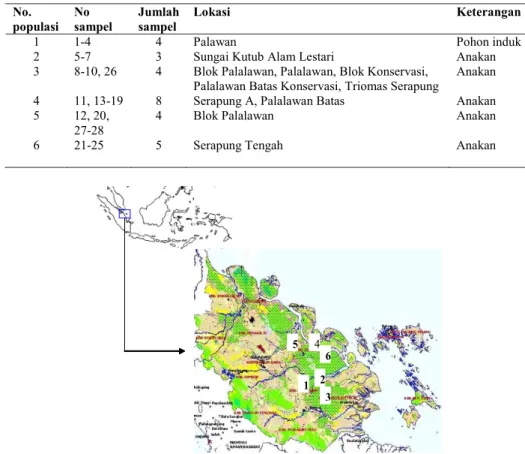
Sampel ramin yang dianalisis dalam studi ini sejumlah 28 (Tabel 1) yang berasal dari enam populasi alam di propinsi Riau (Gambar 1). Populasi 1, 2 and 3 berada di area konsesi PT Riau Andalan Pulp and Paper (RAPP), populasi 4 dan 6 berada di PT Ara Ara Abadi, dan populasi 5 berada dalam konsesi PT Bina Daya Bintara. Sampel ini dikoleksi dalam bentuk daun yang dikeringkan dalam silica gel.
175
Keragaman genetika Ramin [Gonystylus bancanus (Miq.) Kurz]
No.
populasi No sampel Jumlah sampel Lokasi Keterangan
1 1-4 4 Palawan Pohon induk
2 5-7 3 Sungai Kutub Alam Lestari Anakan
3 8-10, 26 4 Blok Palalawan, Palalawan, Blok Konservasi,
Palalawan Batas Konservasi, Triomas Serapung
Anakan
4 11, 13-19 8 Serapung A, Palalawan Batas Anakan
5 12, 20,
27-28
4 Blok Palalawan Anakan
6 21-25 5 Serapung Tengah Anakan
Tabel 1. Daftar sampel ramin dari setiap populasi.
1 2 3
4
6 5
Gambar 1. Lokasi pengambilan sampel pada enam populasi ramin di propinsi Riau. 1) Palawan, 2) Sungai Kutub Alam Lestari, 3) Blok Palalawan, Palalawan, Blok Konservasi, Palalawan Batas Konservasi, Triomas Serapung, 4) Serapung A, Palalawan Batas, 5) Blok Palalawan, 6) Serapung Tengah.
Ekstraksi total DNA genom
Ekstraksi DNA dilakukan dengan mengikuti protokol Delaporta et al. (1983). Lima µL total DNA dielektro-foresis dalam 0.7% gel agarosa dalam larutan penyangga TEA(Tris-EDTA), kemudian diwarnai dengan ethidium bromida, lalu divisualisasi menggunakan lampu ultraviolet kemudian difoto dengan menggunakan gel documentation
system untuk memastikan kuantitas dan
kualitas DNA.
Analisis RAPD
Analisis RAPD yang dilakukan pada studi ini mengikuti protokol William et al. (1990) dengan menggunakan lima primer RAPD (Operon Technology Ltd) yang diseleksi dari 10 primer RAPD dari Operon Kit A, B, C, N, dan U. Reaksi PCR dilakukan dua kali ulangan untuk memastikan konsistensi profil yang dihasilkan. Hasil amplifikasi PCR divisualisasi pada gel agarosa 2.0% dalam larutan penyangga TEA secara
176
Kusumadewi, Poerba, & Partomihardjo
elektroforesis, lalu diwarnai dengan etidium bromida. Hasil pemisahan fragmen DNA dideteksi dengan meng-gunakan UV transluminator, kemudian difoto dengan menggunakan gel
documentation system. Sebagai standar
digunakan 100 bp plus DNA marker (Promega) untuk menetapkan ukuran pita hasil amplifikasi DNA.
Analisis data
Skor data
Setiap pita RAPD dianggap sebagai satu lokus putatif. Hanya lokus yang menunjukkan pita yang jelas yang disekor: ada (1) dan tidak ada (0). Matriks binari fenotip RAPD ini kemudian disusun untuk digunakan pada beberapa analisis.
Keragaman genetika
Parameter-parameter yang digunakan untuk mengkarakterisasi pola keragaman genetika dikalkulasi dengan menggunakan program POPGENE ver. 1.31 (Yeh et al. 1997) dibawah asumsi hukum keseimbangan Hardy-Weinberg. Parameter yang digunakan untuk mengukur keragaman genetika dalam populasi adalah 1) Prosentase Lokus Polimorfik (PLP, Nei 1973); 2) Jumlah alel putatif per lokus (na); 3) Jumlah alel efektif per lokus/gene pool (ne,, Hartl & Clark 1989); 4) keragaman genetika yang diharapkan (H/He=expected
heterozy-gosity (Nei 1973) dan keragaman
fenotipik berdasarkan Shannon’s information index (Lewontin 1972).
Sedangkan parameter yang digunakan untuk mengukur keragaman genetika antar populasi adalah 1) Nei’s
(1978) unbiased measures of genetic
distance; 2) the relative magnitude of differentiation among populations
(GST=Dst/Ht, Nei 1987); 3) Gene flow (Nm, Mc Dermott & McDonald 1993).
HASIL
Profil dan distribusi RAPD di populasi
Lima primer RAPD digunakan dalam penelitian ini dan diperoleh 111 fragmen DNA yang dihasilkan, berukuran dari 100 hingga 2800pb (pasang basa), yakni 99.11% diantaranya merupakan pita polimorfik (Tabel 2). Primer OPD-20 menghasilkan pita terbanyak (28) sedangkan primer OPU-12 menghasilkan pita tersedikit (17). Ada 11 pita umum yang dijumpai pada seluruh populasi, yang sebagian besar berasal dari primer OPD-20. Sementara itu ada 15 pita unik yang terdapat pada populasi tertentu yang umumnya dijumpai pada populasi 4 (Tabel 2).
Keragaman genetika
Keragaman genetika tertinggi dijumpai pada populasi 3 dan 4 masing-masing berdasarkan parameter He (0,1858) dan I (0,2864). Keragaman genetika terendah dijumpai pada populasi 1 dan 2 masing-masing berdasarkan parameter He (0,1438) dan I (0,2201) (Tabel 3).
Nilai total keragaman genetika pada semua populasi (Ht) adalah 0.1982+ 0,0179 dengan nilai rata-rata keragaman genetika dalam populasi (Hs) adalah 0,1606+ 0,0097. Nilai keragaman genetika antar populasi (Dst) adalah
177
Keragaman genetika Ramin [Gonystylus bancanus (Miq.) Kurz]
Tabel 2. Sebaran pita polimorfik RAPD pada enam populasi ramin di propinsi Riau.
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 M
Gambar 2. Salah satu profil RAPD yang dihasilkan dari primer OPN-14 menunjukkan pita DNA polimorfik. M: DNA marker. Angka 1-28: nomor sampel. Anak panah menunjukkan pita umum ukuran 1800 pb.
Tabel 3. Keragaman genetika setiap populasi ramin. Nama
primer Urutan DNA primer Jumlah dan ukuran pita terpendek-terpanjang
Ukuran pita umum
Ukuran pita unik pada setiap populasi
1 2 3 4 5 6 OPB-15 GGAGGGTGTT 19 (100-1600 pb) 100 350 1300 500 1500 450 950 1600 OPC-16 CACACTCCAG 22 (300-1600 pb) 650 700 900 - - - 450 750 - 400 OPD-20 ACCCGGTCAC 28 (300-1800 pb) 800 850 1400 1800 - - - - 750 - - - - - - - - - - - - - - - - - - - OPN-14 TCGTGCGGGT 25 (400-1800 pb) 1800 - - - - OPU-12 TCACCAGCCA 17 (300-1200 pb) 650 1500 450 - 650 400 350 Jumlah 111 11 2 3 1 4 2 3 Populasi Jumlah sampel Jumlah lokus polimorfik PLP (%) na ne He I 1 4 50 44.64 1,4464 1,2334 0,1438 0,2214 2 3 44 39,29 1,3929 1,2525 0,1480 0,2201 3 4 62 55,36 1,5536 1,3013 0,1859 0,2838 4 8 80 71,43 1,7143 1,2679 0,1769 0,2864 5 4 55 49,11 1,4911 1,2541 0,1574 0,2427 6 5 61 54,46 1,5446 1,2353 0,1520 0,2415
178
Kusumadewi, Poerba, & Partomihardjo
Tabel 4. Nilai Ht, Hs, Dst, GST dan Nm 0,1894, jauh lebih rendah Ht dan Hs. Differensiasi genetika (Gst) yang ada antar populasi adalah 95.56% dan gene flow (Nm) yang terjadi antar populasi 2,1393 (Tabel 4). Sedangkan nilai Gst untuk individual marka RAPD berkisar antara 0.0045 (OPB15-18)- 0.3097 (OPD20-23).
Jarak genetika dan analisis kluster
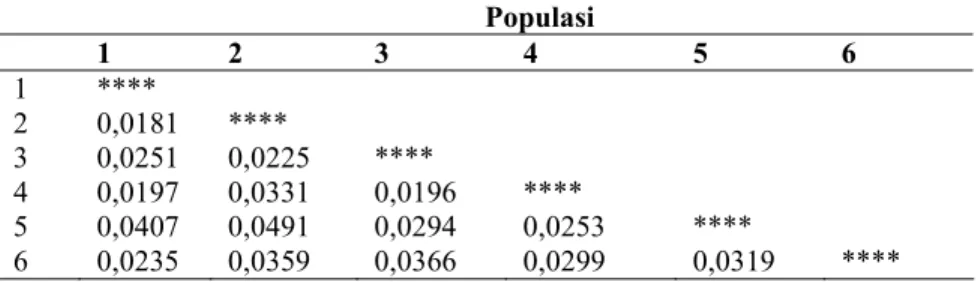
Jarak genetika tertinggi (0.0491) tercatat antara populasi 2 dan 5, sedangkan jarak genetik terendah (0.0181) tercatat antara populasi 1 dan 2 (Tabel 5).
Dendrogram UPGMA (Gambar 3) menyajikan gambaran kesamaan genetika keenam populasi berdasarkan matriks jarak genetika Nei (1978) (Tabel 5). Populasi 3 dan 4 serta 1 dan 2 memiliki kesamaan yang terbanyak, sesuai dengan jarak genetik yang cukup kecil (Tabel 5) sehingga membentuk dua kelompok di terminal cabang dendrogram. Berikutnya populasi 6 yang terdekat dengan kedua kelompok ini dan
Tabel 5. Jarak genetika antar populasi ramin berdasarkan Nei’s Unbiased Measures.
populasi 5 terletak di dasar dendrogram.
PEMBAHASAN
Profil dan distribusi RAPD di populasi
Jumlah pita yang dihasilkan setelah amplifikasi DNA dengan PCR sangat bergantung pada bagaimana primer mengenal urutan DNA komplementernya pada cetakan DNA (DNA template) yang digunakan (Tingey et al. 1994). Polimorfisme pita-pita RAPD yang dihasilkan dari penelitian ini sangat tinggi (99,11% polimorfik). Duplikasi reaksi PCR juga menghasilkan keberulangan yang memuaskan sehingga hal ini menunjukkan bahwa primer yang diseleksi menghasilkan profil RAPD yang cukup konsisten.
Variasi genetika yang ditemui pada penelitian ini berdasarkan perbedaan pola pita RAPD yang dijumpai pada individu ramin. Secara umum, 111 pita RAPD ini menyebar rata di seluruh individu ramin. Namun ada pita-pita tertentu
Populasi 1 2 3 4 5 6 1 **** 2 0,0181 **** 3 0,0251 0,0225 **** 4 0,0197 0,0331 0,0196 **** 5 0,0407 0,0491 0,0294 0,0253 **** 6 0,0235 0,0359 0,0366 0,0299 0,0319 **** Ht Hs Gst Dst Nm Rerata 0,1982+ 0,0179 0,1606+ 0,0097 0,9956 0,1894 2,1393
179
Keragaman genetika Ramin [Gonystylus bancanus (Miq.) Kurz]
yang dijumpai di seluruh individu dan pita unik yang hanya dijumpai di populasi tertentu atau bahkan di individu tertentu. Terdapatnya pita-pita tertentu yang hanya dijumpai pada populasi tertentu maka pita-pita unik tersebut berpotensi untuk dijadikan marka untuk identifikasi
provenance. Namun demikian, untuk
menjadikan RAPD sebagai marka diagnostik tingkat populasi masih perlu ditunjang studi lanjutan, karena jumlah sampel yang terdapat pada populasi 2 khususnya (3 individu) masih sangat minim. Apabila jumlah sampel diperbanyak akan ada kemungkinan ditemukan pita-pita unik lainnya atau sebaliknya pita unik menjadi pita umum. Namun mengingat densitas ramin yang cukup kecil (~1 pohon/hektar) maka jumlah individu sampel yang didapatkan juga cukup kecil.
Keragaman genetika
Jumlah individu yang dicuplik untuk sampel bervariasi antara 3 hingga 8 (Tabel 3). Rendahnya jumlah sampel disebabkan oleh rendahnya densitas ramin. Hal ini kemungkinan disebabkan terjadinya fragmentasi habitat ramin yang cukup parah sehingga populasi-populasi ramin terpisah menjadi sub-populasi yang hanya berisi beberapa individu dalam jumlah kecil. Menurut hasil crusing PT Diamond Raya Timber, sejak tahun 1999 telah terjadi penurunan potensi pohon sebesar 80% dan saat ini ini densitas ramin di area tersebut adalah 0.7 pohon/ hektar (Jayusman 2007).
Penurunan ukuran populasi dapat menyebabkan terisolasinya populasi-populasi menjadi populasi-populasi kecil, seringkali hanya terdiri kurang dari 50 individu. 0.0360 0.0180 0.0000 0.1 pop5 pop6 pop1 pop2 pop3 pop4 Populasi 1 Populasi 2 Populasi 3 Populasi 4 Populasi 5 Populasi 6
Gambar 3. Dendrogram pengelompokkan enam populasi ramin berdasarkan jarak genetika Nei (1978).
180
Kusumadewi, Poerba, & Partomihardjo
Rendahnya jumlah individu dalam setiap populasi dapat menjadi ancaman tersendiri bagi kelangsungan hidup ramin karena dapat menyebabkan terjadinya erosi genetik dan tingginya diferensiasi antar populasi. Hasil penelitian kami menunjukkan bahwa terdapat differen-siasi genetika yang sangat tinggi (99,56%) diantara populasi ramin. Sementara itu, nilai total keragaman genetika populasi ramin (Ht: 0,1982) termasuk rendah dibanding jenis tumbuhan hutan tropis terancam lainnya, Metasequoia
glyptostroboides (H: 0,318, Li et. al.
2005), tetapi lebih tinggi dibanding
Dipterocarpus littoralis (H: 0,1958).
Rendahnya nilai keragaman genetika dalam populasi ini tidak sesuai dengan kenyataan yang umumnya terjadi pada jenis-jenis tumbuhan menyerbuk silang. Ramin, seperti halnya jenis-jenis tumbuhan tropis lainnya adalah jenis tumbuhan dominan di hutan rawa gambut,
late successional dan menyerbuk
silang. Pada jenis-jenis menyerbuk silang seperti ini biasanya nilai keragaman genetika dalam populasi lebih tinggi daripada antar populasi (Loveless & Hamrick 1984; Hamrick & Godt 1989; Pither et al., 2003). Sebaliknya, telah terjadi pada ramin yang diteliti. Rendahnya keragaman genetika ramin kemungkinan disebabkan oleh terjadinya
inbreeding antar individu dalam populasi
yang jumlahnya sedikit sehingga variasi genetik yang ada juga rendah atau bahkan selfers pada individu. Terjadinya
inbreeding dan/atau self fertilisation,
memang sulit untuk dibuktikan terutama pada ramin yang memiliki jangka hidup
panjang. Namun hal ini merupakan salah satu penjelasan yang bisa dipaparkan.
Ancaman lain bagi kelangsungan hidup ramin adalah rendahnya tingkat regenerasi. Dari informasi yang diperoleh di lapangan, tingkat predasi ramin oleh mamalia kecil (tupai) dan burung (rangkong) cukup tinggi. Masa berbuah ramin rata-rata setiap 3-5 tahun sekali yang seringkali merupakan panen raya. Tingginya predasi mengakibatkan berkurangnya jumlah biji yang dapat berkecambah menjadi semai. Kemam-puan semai untuk berkembang juga sangat rendah karena berbagai faktor, antara lain serangan herbivor dan mati kekeringan, sehingga seringkali sulit menjumpai anakan ramin di habitat alaminya. Teknik budidaya ramin juga belum sepenuhnya dikuasai hingga saat ini. Sementara itu ramin merupakan jenis kayu favorit dalam dunia perdagangan kayu yang terus diminati. Seluruh kondisi ini cukup memprihatinkan, dan apabila dibiarkan berlanjut, dikhawatirkan ramin akan segera mengalami kepunahan.
Jarak genetika dan analisis kluster
Populasi 5 dan 6 tidak mengelompok bersama dengan populasi-populasi lain. Hal ini kemungkinan karena individu-individu ramin yang berada di kedua populasi ini memiliki alel yang paling berbeda yang tercermin dari profil RAPD mereka.
Tingginya jarak genetik antara populasi 2 dan 5 yang juga tercermin dari pola pengelompokkan pada dendrogram, kemungkinan disebakan oleh rendahnya jumlah individu yang tercuplik dimana kesemua individu dalam kedua populasi
181
Keragaman genetika Ramin [Gonystylus bancanus (Miq.) Kurz]
ini juga memiliki alel yang sangat berbeda. Kedua populasi ini terletak pada dua hutan konsesi yang berbeda (Gambar 1) dalam jarak yang cukup jauh sehingga kemungkinan terjadinya percampuran materi genetik baik melalui pollen maupun pencaran biji antar kedua populasi ini juga sangat kecil.
Sedangkan mengelompoknya populasi 3 dan 4 –yang secara geografis jaraknya cukup jauh (Gambar 1)-kemungkinan disebabkan oleh dua hal. Yang pertama, adanya kesamaan profil RAPD yang dihasilkan dari amplifikasi PCR. RAPD menggunakan primer universal yang mengamplifikasi daerah genom DNA secara acak, sehingga pita-pita DNA yang dihasilkan juga bersifat acak. Kedua, terdapatnya kemungkinan bahwa beberapa individu pada kedua populasi berasal dari provenance yang sama- apabila terjadi introduksi pada kedua populasi tersebut. Namun hal yang kedua kemungkinan kecil terjadi, karena semua ramin yang berada dalam area konsesi ini merupakan hutan alam.
UCAPAN TERIMA KASIH
Penulis mengucapkan terima kasih kepada PT Riau Andalan Pulp and Paper atas fasilitas dan ijin pengumpulan material di area konservasi kawasan HTI RAPP. Terima kasih juga kepada Sdri. Agustina dan Herlina yang telah memberi bantuan teknis terhadap penelitian ini.
DAFTAR PUSTAKA
Asif, M.J. & C.H. Cannon. 2005. DNA extraction from processes wood: a
case study for the identification of endangared timber species (Gonystylus bancanus). Plant
Mol. Biol. Reporter 23: 185-192.
Convention on International trade in endangered species of wild flora and fauna. 2009. Appendices I, II and III. Valid from 22 May 2009. P. 40.
Delaporta, SL., J. Wood, & JB. Hicks. 1983. A plant DNA minipre-paration. Version II. Plant Mol.
Biol. Reporter 4:19–21.
Hamrick, JL., & MJW. Godt, 1989. Allozyme diversity in plant species. Dalam: Brown, AHD, MJ. Clegg, AL. Kahler, & BS. Weir (eds.).
Plant population genetics, breeding and genetic resources.
Sinauer Associates, Sunderland, Massachusetts.
Hartl, DL. & AG. Clark. 1989.
Principles of population
gene-tics. 2nd ed. Sinauer Associates,
Sunderland, Massachusetts. IUCN. 2008. Red List of Endangered
Species. International Union for the Conservation of Nature and Natural Resources, Gland. Switzerland. Available from URL: http://www.iucnredlist.org/ Jayusman. 2007. Degradasi sumberdaya
genetik jenis ramin dan upaya penyelamatannya. Apforgen
newsletter 4:1-4.
Jain, PK, L. Saini, MH. Pathak & VK. Gupta. 2007. Analysis of genetic variation in different banana (Musa species) variety using random amplified polymorphic
182
Kusumadewi, Poerba, & Partomihardjo
DNAs (RAPDs). African J.
Biotech. 6 (17): 1987-1989.
Jiminez , JF., P. Sanchez-Gomez, J. Guemes, O. Werner & JA. Rossello. 2002. Genetic variability in a narrow endemic snapdragon (Antirrhinum subaeticum, Schrophulariaceae) using RAPD markers. Heredity 89(5): 387-393. Lewontin, R.C. 1972. The apportionment of human diversity. J. Evol. Biol. 6: 381-398.
Li, YY., XY. Chen, X. Zhang, TY. Wu, HP. Lu, & YW Chai. 2005. Genetic differences between wild and artificial populations of
Metasequoia glyptostroboides
Hu et Cheng (Taxodiaceae): Implication for species recovery.
Conser. Biol. 19: 224-231.
Loveless, MD. & JL. Hamrick. 1984. Ecological determinants of genetic structure in plant populations. Ann.
Rev. Ecol.& Syst. 15: 65-95.
Malik, SK., R. Chaudhury, OP. Dhariwal & R.K. Kalia. 2006. Collection and characterisation of Citrus indica Tanaka and C. macroptera Montr.: wild endange-red species of north eastern India. Gen. Res. & Crop
Evol. 53: 1485-1493.
McDermott, JM. & BA. McDonald. 1993. Gene flow in plant pathosys-tems. Ann.Rev. Phytopathology 31: 353-373.
Nei, M. 1978. Estimation of average heterozygosity and genetic distance from a small number of individuals.
Genetics 89: 583-590.
Nei, M. 1973. Analysis of gene diversity in subdivided populations.
Proceedings of the National Academy Sciences USA 70:
3321-3323.
Pither, R., JS. Shore, & M. Kellman. 2003. Genetic diversity of the tropical tree Terminalia amazonia (Combretaceae) in naturally fragmented populations. Heredity. 91(3): 307-313.
Poerba, YS., A. Wawo, & KS. Yulita. 2007. Keragaman Fenotipe RAPD
Santalum album L. di Pulau Timor
bagian timur. Berita Biologi 8(6): 537- 546.
Shaaban, EA., S. Abd-El-Aal SKH., NS. Zaied & AA. Rizkalla. 2006. Assessment of genetic variability on some orange accessions using RAPD DNA markers. Res. J.
Agri.& Biol.Sci 2(6): 564-570.
Siregar, IZ., T. Yunanto, & P Pamoengkas. 2008. Implikasi genetik metode pembiakan tanaman Shorea johorensis Foxw. Pada sistem silvikultur tebang pilih tanam jalur (TPTJ).
Biodiversitas 9(4): 250-254.
Smulders, MJM., WPC. Van T Westende, B. Diway, G.D. Esselink, P.J. Van Dr Meer, & J.M. Koopman. 2007. Development of microsattelites markers in
Gonystylus bancanus (Ramin)
useful for tracing and tracking wood of this protected species.
Molecular Ecology Notes.
(Proof. read. version)
Tingey, SV., JA. Rafalski, & MK. Hanafey. 1994. Genetic analysis with RAPD markers: In Plant
183
Keragaman genetika Ramin [Gonystylus bancanus (Miq.) Kurz]
Molecular Biology. C. Coruzzi & P. Puidormenech (Eds.) p.491-498. Williams, JG., AR. Kubelik, KJ. Livak, J.A. Rafalsky, & S.V. Tingev. 1990. DNA polymorphism amplified by arbitrary primers are useful as genetic markers. Nucleic
Acids Research 18(22):
6531-6535.
Yeh, FC., RC. Yang, TBJ. Boyle, ZH., Ye, JX. Mao 1997. POPGENE
(version 1.31), the user-friendly shareware for population genetic
analysis. Molecular Biology and
Biotechnology centre, University of Alberta, Edmonton. Alberta, Canada. Available free at http:// www.ualberta.ca/~fyeh.
Yulita, KS & T. Partomihardjo. 2010. Keragaman genetika populasi pelahlar [Dipterocarpus littoralis (Bl.) Kurz] di Pulau Nusakambangan berdasa-rkan profil Enhanced Random Amplified Polymorphic DNA. Berita
Biologi (in press.)
Memasukkan: Juni 2009 Diterima: Desember 2009
J. Biol. Indon. Vol 6, No. 2 (2010)
PANDUAN PENULIS
Naskah dapat ditulis dalam bahasa Indonesia atau bahasa Inggris. Naskah disusun dengan urutan: JUDUL (bahasa Indonesia dan Inggris), NAMA PENULIS (yang disertai dengan alamat Lembaga/ Instansi), ABSTRAK (bahasa Inggris, maksimal 250 kata), KATA KUNCI (maksimal 6 kata), PENDAHULUAN, BAHAN DAN CARA KERJA, HASIL, PEMBAHASAN, UCAPAN TERIMA KASIH (jika diperlukan) dan DAFTAR PUSTAKA.
Naskah diketik dengan spasi ganda pada kertas HVS A4 maksimum 15 halaman termasuk gambar, foto, dan tabel disertai CD. Batas dari tepi kiri 3 cm, kanan, atas, dan bawah masing-masing 2,5 cm dengan program pengolah kata Microsoft Word dan tipe huruf Times New Roman berukuran 12 point. Setiap halaman diberi nomor halaman secara berurutan. Gambar dalam bentuk grafik/diagram harus asli (bukan fotokopi) dan foto (dicetak di kertas licin atau di scan). Gambar dan Tabel di tulis dan ditempatkan di halam terpisah di akhir naskah. Penulisan simbol α, β, χ, dan lain-lain dimasukkan melalui fasilitas insert, tanpa mengubah jenis huruf. Kata dalam bahasa asing dicetak miring. Naskah dikirimkan ke alamat Redaksi sebanyak 3 eksemplar (2 eksemplar tanpa nama dan lembaga penulis).
Penggunaan nama suatu tumbuhan atau hewan dalam bahasa Indonesia/Daerah harus diikuti nama ilmiahnya (cetak miring) beserta Authornya pada pengungkapan pertama kali.
Daftar pustaka ditulis secara abjad menggunakan sistem nama-tahun. Contoh penulisan pustaka acuan sebagai berikut :
Jurnal :
Hara, T., JR. Zhang, & S. Ueda. 1983. Identification of plasmids linked with polyglutamate production in B. subtilis. J. Gen. Apll. Microbiol. 29: 345-354.
Buku :
Chaplin, MF. & C. Bucke. 1990. Enzyme Technology. Cambridge University Press. Cambridge.
Bab dalam Buku :
Gerhart, P. & SW. Drew. 1994. Liquid culture. Dalam : Gerhart, P., R.G.E. Murray, W.A. Wood, & N.R. Krieg (eds.). Methods for General and Molecular Bacteriology. ASM., Washington. 248-277.
Abstrak :
Suryajaya, D. 1982. Perkembangan tanaman polong-polongan utama di Indonesia. Abstrak Pertemuan Ilmiah Mikrobiologi. Jakarta . 15 –18 Oktober 1982. 42.
Prosiding :
Mubarik, NR., A. Suwanto, & MT. Suhartono. 2000. Isolasi dan karakterisasi protease ekstrasellular dari bakteri isolat termofilik ekstrim. Prosiding Seminar nasional Industri Enzim dan Bioteknologi II. Jakarta, 15-16 Februari 2000. 151-158.
Skripsi, Tesis, Disertasi :
Kemala, S. 1987. Pola Pertanian, Industri Perdagangan Kelapa dan Kelapa Sawit di Indonesia.[Disertasi]. Bogor : Institut Pertanian Bogor.
Informasi dari Internet :
Schulze, H. 1999. Detection and Identification of Lories and Pottos in The Wild; Information for surveys/Estimated of population density. http//www.species.net/primates/loris/ lorCp.1.html.
J. Biol. Indon. Vol 6, No.2 (2010)
Kemampuan Kawasan Nir-Konservasi dalam Melindungi Kelestarian Burung Endemik Dataran Rendah Pulau Jawa Studi Kasus di Kabupaten Kebumen
Eko Sulistyadi
237
Analysis of Nutrient Requirement and Feed Efficiency Ratio of Maroon Leaf Monkey (Presbytis rubicunda Mueller, 1838)
Wartika Rosa Farida
255
Oksidasi Nitrit Oleh Bakteri Heterotrofik Pada Kondisi Aerobik Dwi Agustiyani, Ruly Marthina Kayadoe & Hartati Imamuddin
265
Pencirian Karbon Organik Air Sungai Citarum Hulu Dari Masukan Air Limbah Penduduk dan Industri
Eko Harsono & Sulung Nomosatryo
277
TULISAN PENDEK
Arti Kebun Raya Bogor Bagi Kehidupan Kumbang Sungut Panjang (Coleoptera, Cerambicidae)
Woro Anggaraitoningsih Noerdjito