PEMULIAAN KETAHANAN TANAMAN KOPI
TERHADAP NEMATODA PARASIT
Breeding for Resistance of Coffee to Plant-Parasitic Nematodes Retno Hulupi
Peneliti Pemuliaan Tanaman Pusat Penelitian Kopi dan Kakao Indonesia
Jl. PB. Sudirman No. 90 Jember Ringkasan
Apabila dibandingkan dengan pemuliaan ketahanan kopi Arabika terhadap penyakit karat daun, pemuliaan ketahanan kopi terhadap nematoda parasit kurang berkembang, karena masalah ketersediaan sumber gen tahan serta terbatasnya informasi hasil penelitian yang mendukung perbaikan genetika ketahanan kopi terhadap nematoda. Sementara itu disadari bahwa penggunaan kultivar tahan dinilai merupakan metode pengendalian yang ekonomis, awet serta ramah lingkungan. Hasil kajian beberapa pustaka serta hasil penelitian pendahuluan menunjukkan bahwa uji saring pada beberapa koleksi plasma nutfah kopi untuk mendapatkan tetua sumber gen tahan nematoda parasit merupakan langkah awal kegiatan pemuliaan ketahanan. Mengingat kopi Arabika mempunyai susunan genetik berbeda dengan kopi Robusta, demikian pula dengan sifat penyerbukannya, sehingga strategi pemuliaan ketahanan kopi Arabika berbeda dengan kopi Robusta. Sebelumnya perlu dipelajari hal-hal yang berkaitan dengan perilaku nematoda P. coffeae dan R. similis, mekanisme ketahanan serta genetika ketahanannya untuk setiap jenis kopi (Arabika dan Ro-busta). Metode pemuliaan ketahanan kopi terhadap nematoda parasit yang dapat memotong daur seleksi adalah metode pemuliaan populasi yang menghasilkan varietas sintetik atau varietas komposit, sifat ketahanan bersifat awet (durable) serupa ketahanan horisontal karena disusun oleh banyak gen ketahanan dalam satuan populasi. Berdasarkan beberapa hasil penelitian, pemuliaan ketahanan kopi Arabika terhadap nematoda parasit diarahkan terhadap nematoda R. similis, sedangkan pemuliaan ketahanan kopi Robusta terhadap nematoda P. coffeae.
Summary
Breeding for resistance of coffee to plant-parasitic nematodes has not been established, due to the availability of gene source and the limited studies related with improvement that genetic resistance. Meanwhile, use of resistant cultivar is considered to be the economic operation method. Several published references and preliminary studies indicated that screening test on germplasm coffee
collec-tions to get gene source an parental is an early activity of breeding for resistance. Considering that Arabica coffee differs genetically from Robusta coffee, including their pollination, breeding efforts to obtain durable resistance to nematodes on Arabica differ with Robusta coffee. To set up strategy coffee breeding to plant-parasitic nematodes in the first step to required study behavior of nematodes and the genetic mechanism the resistance of each coffee species. Breeding method to obtain coffee resistance to plant-parasitic nematode that is able to shortcut the cycle of selections in population level is breeding to get synthetic or composite varieties. The resistance of those varieties is similar with durable character of horizontal resistance since they are composed of resistant genes in set of popula-tion. Based on some researches, for Arabica coffee, breeding for resistance to plant-parasitic nematode should be focused on R. similis, while for Robusta on
P. coffeae.
PENDAHULUAN
Pemuliaan ketahanan terhadap hama-penyakit adalah bagian dari kegiatan penelitian untuk mendapatkan bahan tanam unggul. Sebagaimana diketahui, tuntutan untuk mendapatkan bahan tanam kopi unggul tahan terhadap serangan nematoda parasit semakin mendesak untuk segera dilaksanakan. Hal ini mengingat rekomen-dasi pengendalian secara preventif dengan pemberian pupuk kandang tidak mudah diterapkan, terutama jika areal serangannya cukup luas serta bertopografi bergelom-bang, sehingga akan kesulitan dalam pengangkutan. Bahkan dalam beberapa kasus sering menimbulkan serangan hama lain sejenis uret yang telurnya terbawa dalam pupuk kandang. Sementara itu pengendalian menggunakan nematisida selain dinilai tidak ekonomis juga tidak ramah lingkungan. Berdasarkan beberapa kasus tersebut maka penggunaan bahan tanam tahan dinilai merupakan metode pengendalian yang ekonomis, awet serta ramah lingkungan.
Selama ini penelitian pemuliaan ke-tahanan terhadap nematoda pada kopi kurang banyak diminati. Apabila di-bandingkan dengan pemuliaan ketahanan kopi arabika terhadap penyakit karat daun, pemuliaan ketahanan kopi terhadap nematoda parasit kurang berkembang, karena selain masalah ketersediaan sumber gen tahan, juga terbatasnya informasi hasil penelitian yang mendukung perbaikan genetika ketahanan kopi terhadap nematoda. Apabila dilihat dari gejala kerusakan yang timbul, dampak serangan nematoda parasit memang lebih lambat dibanding serangan jamur penyebab penyakit karat daun, meskipun pada akhirnya tingkat kerusakan yang ditimbulkannya sama, sehingga wajar jika pemuliaan ketahanan terhadap jamur Hemileia vastatrix lebih maju. Penelitian mekanisme ketahanan kopi terhadap nematoda parasit juga belum banyak dilaporkan, padahal informasi tersebut diperlukan sebagai dasar pemuliaannya. Di Brazil, India, Kenya, dan Columbia pemuliaan ketahanan terhadap nematoda parasit ditekankan untuk mendapatkan
varietas tahan serangan nematoda Meloi-dogyne spp. karena jenis nematoda ini dianggap paling merusak pertanaman kopi Arabika di daerah tersebut. Strateginya selain diarahkan untuk mendapatkan batang bawah tahan dari spesies kopi lain (Arango-Bernal et al., 1982) juga mendapatkan varietas kopi Arabika tahan melalui seleksi hasil persilangan dengan tetua tahan (Anzueto et al.,1993, Bertrand et al., 1995). Sebagaimana halnya di Brazil upaya mengatasi serangan nematoda parasit pada kopi Arabika yang dilakukan di Costa Rica adalah dengan penggunaan batang bawah kopi Robusta tahan dengan nomor seleksi C 2258 yang disambung dengan varietas anjuran, Mundonovo & Catuai Vermelho (Campos et al., 1990). Demikian pula di India, meskipun persilangan buatan untuk menemukan kopi Arabika tahan Meloidogyne spp. dan P. coffeae terus diupayakan, namun metode pengendalian yang dianjurkan adalah menggunakan batang bawah kopi Robusta tahan dengan nomor seleksi S 274 kemudian disambung dengan batang atas varietas anjuran (Kumar & Samuel, 1990). Tampaknya uji saring pada beberapa koleksi plasma nutfah untuk mendapatkan batang bawah tahan merupakan cara yang relatif cepat untuk mendapatkan bahan tanam tahan dalam upaya mengatasi serangan nematoda parasit di beberapa daerah penghasil kopi di dunia. Demikian pula di Indonesia upaya mendapatkan bahan tanam tahan diawali dengan seleksi dan uji saring terhadap beberapa nomor koleksi plasma nutfah dari jenis kopi Liberica - Excelsa serta kopi
Robusta yang diketahui memiliki sistem perakanan lebih baik serta regenerasi akar lebih cepat sehingga diharapkan dapat digunakan sebagai batang bawah yang selanjutnya disambung dengan varietas/ klon anjuran.
Untuk mengatur strategi pemuliaan kopi terhadap nematoda parasit di Indone-sia, sebelumnya perlu dipelajari hal-hal yang berkaitan dengan perilaku nematoda tersebut, serta genetika ketahanannya untuk setiap jenis kopi.
JENIS DAN KULTIVAR KOPI YANG DIBUDIDAYAKAN
DI INDONESIA
Jenis kopi yang pertama kali dibudi-dayakan di Indonesia adalah Arabika varietas Typica, namun sejak masuknya penyakit karat daun ke Indonesia pada tahun 1959, varietas tersebut hanya mampu ber-tahan jika ditanam di lahan tinggi (> 1250 m dpl.). Untuk mengisi kekosongan lahan rendah diintroduksi jenis kopi lain yaitu kopi Liberika (Coffea liberica) serta kopi Ekselsa (Coffea dewevrei var. excelsa). Namun pengembangan jenis kopi ini tidak ber-langsung lama, karena selain produksinya rendah citarasanya asam, sehingga kurang disukai (Yahmadi, 1972), dan pada saat ini jenis kopi tersebut hanya dijumpai di lahan gambut (Sumatera dan Kalimantan) sebagai konsumsi lokal. Selanjutnya diintroduksi Coffea canephora var. robusta (kopi Ro-busta). Kopi jenis inilah yang hingga saat ini merupakan jenis kopi yang paling
merajai di Indonesia, yaitu mencapai lebih dari 90 persen, sedangkan 9 persen sisanya merupakan kopi Arabika dan jenis lain (AEKI, 2007).
Dalam kaitannya dengan reaksi ke-tahanan beberapa kultivar kopi yang di-budidayakan di Indonesia terhadap nematoda parasit, hasil pengamatan lapangan menunjukkan bahwa sebagian besar klon anjuran maupun varietas lokal kopi Robusta di Indonesia rentan serangan nematoda parasit. Klon-klon kopi Robusta anjuran, BP 42, BP 358, BP 409, BP 436, BP 534, BP 936 dan BP 939 rentan nematoda P. coffeae (Wiryadiputra et al., 1998), dan sampai saat ini baru ditemukan satu genotipe yang menunjukkan reaksi tahan, yaitu klon BP 308 (Mawardi et al., 2003). Hal serupa juga terjadi pada kopi Arabika, semua varietas anjuran kopi Arabika tidak tahan serangan nematoda parasit, dengan tingkat kerentanan berbeda-beda. Varietas S 795 merupakan varietas yang mampu beradap-tasi pada kondisi lahan marginal namun ternyata rentan terhadap nematoda parasit. Demikian pula halnya varietas lokal dari Sumatera Utara, Sigarar Utang dan dua nomor seleksi keturunan varietas Catimor yang diintroduksi dari Portugal (Kartika 1 dan Kartika 2) menunjukkan reaksi sangat rentan terhadap nematoda R. similis, namun Hibrido de Timor H 832 – 1 menunjukkan reaksi tahan (Hulupi, 2006 a). Pada varietas Catimor, gen ketahanan terhadap penyakit karat daun dilaporkan berasal dari tetua Hibrido de Timor (Rodrigues Jr, 1988), namun tampaknya sifat tahan terhadap nematoda R. similis pada Hibrido de Timor
tidak semuanya diwariskan pada bastar Catimor, bahkan sifat tahan penyakit karat daun tidak selalu diikuti dengan sifat tahan nematoda parasit.
NEMATODA PARASIT PADA KOPI
Nematoda parasit yang berasosiasi dengan kopi Arabika dan kopi Robusta sesungguhnya lebih dari 14 spesies, namun berdasarkan hasil survei, jenis nematoda parasit yang sering menyerang tanaman kopi di Indonesia hanya tiga, yaitu nematoda luka akar, Pratylenchus coffeae, nematoda penggali akar, Radopholus similis dan nematoda puru akar, Meloidogyne spp. (Dropkin, 1992; Wiryadiputra, et al., 1995). Dua spesies yang pertama merupakan endoparasit berpindah (migratory en-doparasitic nematodes), sedangkan yang terakhir merupakan endoparasit menetap yang membentuk gal (sedentary endopara-sitic nematodes). Oleh Brown & Colbran (1980) dan Román (1986) nematoda R. similis digolongkan sebagai nematoda parasit obligat yang hanya dapat hidup pada inangnya. Hampir seluruh daur hidupnya berlangsung dalam jaringan korteks akar, sehingga selalu mencari inang lain yang sesuai. Menurut Noe (1998), inang lain nematoda R. similis sangat banyak, termasuk di antaranya beberapa gulma, rumput Guatemela (Tripsacum laxum) serta beberapa tanaman buah-buahan, seperti pisang dan jeruk, sehingga jika akar kopi habis terserang maka nematoda segera mencari inang lain. Pada tanah bertekstur
pasiran dengan pola sebaran hujan hampir merata sepanjang tahun nematoda jenis endoparasit berpindah (R. similis dan P. coffeae) lebih cepat berkembang (Fassuliotis cit Sidhu et al.,1981). Pada tipe lahan seperti itu nematoda tersebut juga mampu mencapai kedalaman tanah lebih dari 50 cm di bawah permukaan tanah (Kumar & Samuel, 1990). Populasi R. similis dilaporkan paling banyak di-temukan pada kedalaman tanah 30–100 cm, sedangkan penetrasinya dapat mencapai akar yang terletak pada kedalaman 370 cm dari permukaan tanah. Hal inilah yang menjadi salah satu penyebab sulitnya pengendalian R. similis menggunakan nematisida ataupun perlakuan kultur teknis lainnya (Kumar & Samuel, 1990; Kusno– Amidjojo, 1995b).
Hasil pengujian sebaran populasi nematoda R. similis dan P. coffeae yang dilakukan di beberapa lahan endemik serangan nematoda yang menyerang kopi Arabika rentan (Kartika 1) menunjukkan bahwa populasi nematoda P. coffeae dalam akar yang terletak pada kedalaman kurang dari 30 cm lebih tinggi dibanding populasi R. similis, sedangkan jumlah nematoda R. similis yang berasal dari akar pohon yang sama namun pada kedalaman lebih dari 50 cm lebih banyak dibanding P. coffeae. Hal ini menguatkan dugaan adanya korelasi antara perbedaan daerah sebaran akar kopi pada kedalaman tanah berbeda dengan populasi nematoda R. similis dan P. coffeae. Nematoda R. similis lebih banyak berada pada zona kedalaman tanah lebih dari 50 cm di bawah permukaan tanah, sedangkan P. coffeae lebih dominan berada
pada zona perakaran kurang dari 30 cm di bawah permukaan tanah (Hulupi & Mulyadi, 2007).
Pada lahan kopi Robusta semula banyak dijumpai serangan nematoda puru akar, namun dewasa ini tampaknya populasinya didominasi oleh nematoda berpindah, P. coffeae dan R. similis. Perubahan populasi ini diduga berkaitan dengan perubahan pola budidaya kopi di Indonesia yang secara tidak langsung menekan perkembangan jenis nematoda tersebut atau tertekan oleh jumlah populasi nematoda R. similis dan P. coffea yang lebih cepat berkembang.
GEJALA KERUSAKAN TANAMAN KOPI AKIBAT
SERANGAN NEMATODA 1. Radopholus similis Cobb.
Tanaman kopi yang telah terinfeksi nematoda penggali akar R. similis dalam beberapa bulan telah menunjukkan gejala kerusakannya. Gejala awal yang muncul dapat dilihat pada saat musim kemarau, daun menguning mirip gejala defisiensi hara. Gejala kerusakan pada bibit akibat terserang nematoda ini adalah timbulnya bercak nekrosis pada ujung daun berwarna coklat kehitaman mirip gejala serangan jamur Cercospora sp. Gejala bercak nekrosis pada bibit ini serupa dengan gejala serangan nematoda P. coffea. Pada tanaman dewasa kulit akar terdapat bercak berwarna coklat kemerahan, apabila telah membusuk berwarna hitam. Tanaman tidak segera mati
tetapi tumbuh merana, daun sedikit demi sedikit rontok, jumlah buah tidak sebanding dengan jumlah daun yang mendukungnya (Gonzaga & Lordello,1986; Sijmons,1993). Pada varietas kopi Arabika yang potensial mengalami pembuahan berlebih (over bear-ing), seperti Kartika 1, Kartika 2 dan Andungsari 1, setelah berakhirnya masa pembuahan, daun-daun yang terletak di bagian tengah dekat batang pokok akan menguning dan akhirnya gugur bersamaan dengan serangan penyakit karat daun. Akar tanaman sebagai organ pencari makanan rusak dan membusuk, sehingga tanaman tumbuh meranggas tinggal ranting kering bersama buah muda yang kering karena tidak dapat masak sempurna, dan akhirnya mati (Kusno-Amidjojo, 1995; Wirya-diputra, 1997). Mekanisme merusaknya nematoda R. similis pada tanaman kopi secara umum dijelaskan oleh Dropkin (1992) sebagai berikut : larva nematoda masuk ke dalam bagian akar yang masih lunak dengan cara mengeluarkan sekresi yang berasal dari kelenjar usopagus, dan pada saat yang sama larva merusak sel akar. Setelah sel akar rusak, nematoda berpindah ke bagian korteks akar, menetap untuk berkembang biak.
2. Pratylenchus coffeae
Akar tanaman kopi yang terserang nematoda luka akar, P. coffeae warnanya tidak putih tetapi kuning, kemudian berubah menjadi coklat, sedangkan akar lateralnya busuk. Luka pada akar tersebut berakibat merusak seluruh sistem perakaran kopi sehingga menghambat penyerapan hara dari dalam tanah. Daun menunjukkan gejala
klorosis (menguning) dimulai dari daun yang terletak dekat batang, kemudian cabang-cabang utama tumbuh sedikit, dan batang pohon menjadi mudah digoyang karena akarnya habis, akhirnya tanaman mati (Campos, et al., 1992). Agak sulit dibedakan gejala serangan nematoda P. coffeae dengan R. similis. Pada bibit dan tanaman muda pertumbuhan tanaman menjadi lambat, tanaman kurus dan kerdil, daun mengecil serta klorosis. Pada daun timbul bercak nekrosis berwarna coklat tua seperti terbakar, dimulai dari ujung daun. Tanaman muda kopi Arabika varietas Kartika 1 dan Kartika 2 yang terserang berat akan mati sebelum dewasa, atau pal-ing lama setelah berbuah pertama, diawali dengan gejala menguningnya daun. Hal ini berkaitan dengan sebaran populasi nematoda tersebut mulai permukaan tanah hingga kedalaman 30 cm, dimana akar kopi muda sebagian besar berada pada zona kedalaman yang sama. Dalam hamparan pertanaman kopi dewasa, gejala menguning tampak dalam blok-blok tertentu mengikuti jalannya aliran air. Gejala kerusakan di-perparah jika tanaman kopi tidak ber-penaung, serta kurang hara (tidak dipupuk) sehingga tanaman menunjukkan gejala kerusakan lebih parah.
3. Meloidogyne spp.
Nematoda puru akar genus Meloido-gyne jika menyerang tanaman kopi akan menyebabkan timbulnya puru berbentuk bulat pada akar-akar, yang biasanya ter-bentuk setelah datangnya awal musim penghujan. Pada bagian akar nekrotik akan terlihat dengan mudah timbulnya akar
berpuru, semula berwarna kuning kecoklatan dan sering terjadi infeksi sekunder yang menyebabkan sebagian akar mati (Dropkin, 1992). Beberapa klon anjuran kopi Robusta yang terserang nematoda jenis ini sering diikuti dengan gugurnya daun setelah sebelumnya mengalami klorosis, dan akhirnya tanaman mati. Namun dewasa ini nematoda puru yang menyerang kopi di Indonesia jarang dilaporkan. Diduga serangan nematoda R. similis dan P. coffeae lebih dominan merusak dan perbanyakannya lebih cepat, sehingga populasi Meloidogyne spp. di lahan pertanaman kopi tergeser oleh kedua jenis nematoda tersebut.
KETAHANAN KOPI TERHADAP NEMATODA PARASIT 1. Terminologi Ketahanan
Hubungan parasitisme antara inang dengan patogen diekspresikan dalam bentuk ketahanan atau kerentanan. Reaksi ke-tahanan tanaman tersebut dapat terjadi sebelum infeksi (pre infection) ataupun sesudah infeksi (post infection) berupa kandungan senyawa kimia atau struktur jaringan yang dimiliki tanaman untuk menghambat reproduksi patogen (Sijmons, 1993; Williamson & Hussey, 1996; Collingborn et al. 2000). Menurut Kumar (1991) ketahanan ataupun kerentanan inang terhadap parasit erat kaitannya dengan sifat kesesuaian dan ketidaksesuaian hubungan di antara keduanya. Beberapa nematolog menguraikan istilah tanaman tahan serangan nematoda parasit sebagai bentuk interaksi
antara tanaman dengan parasit yang mampu menekan perkembangbiakan atau tingkat reproduksi nematoda karena adanya mekanisme berupa hambatan, benteng, atau sesuatu yang dapat memperlambat per-kembangbiakan nematoda. Tanaman ter-sebut mampu menekan perkembangan nematoda sehingga tetap sehat. Sementara itu tanaman disebut kebal (immune) terhadap nematoda jika nematoda tidak tertarik untuk hidup dan berkembang pada tanaman tersebut karena tidak adanya ketertarikan untuk berinteraksi dengan tanaman tersebut. Tanaman disebut toleran jika nematoda dapat berkembang biak di dalam tanaman kopi namun hanya menyebabkan sedikit kerusakan (Trudgil, 1991, Dalmasso et al., 1992). Berdasarkan gen penyusunnya, Van der Plank (1968) memilah ketahanan menjadi 3, yaitu : 1). ketahanan monogenik, 2). ketahanan oligogenik, dan 3). ketahanan poligenik. Sementara itu Horber (1980) memilah ketahanan suatu tanaman terhadap hama-penyakit secara umum menjadi 2, yaitu ketahanan sejati (true resistance) dan ketahanan semu (pseudo resistance). Suatu tanaman memiliki ketahanan sejati akibat karakteristik anatomi, fisiologi atau bio-kimiawi yang secara genetik diwariskan, sedangkan tanaman memiliki ketahanan semu jika ketahanan yang timbul sebagai akibat fenologi tanaman sehingga terhindar dari serangan patogen. Sementara itu ketahanan tersebut dapat berarti luas, yaitu apabila tanaman tahan terhadap beberapa jenis nematoda, sedangkan ketahanan dalam arti sempit, yaitu apabila tanaman hanya tahan terhadap jenis/biotipe/ras tertentu. Untuk mengetahui tingkat ketahanan suatu
tanaman terhadap serangan nematoda, Williamson & Hussey (1996) memilah menjadi beberapa tingkat, yaitu : 1). kebal (immune), 2). sangat tahan (highly resis-tant), 3). agak tahan (moderately resisresis-tant), 4). rentan (susceptible), dan 5). sangat rentan (highly susceptible). Untuk melakukan seleksi ketahanan kopi terhadap nematoda parasit, penerapan istilah-istilah tersebut tampaknya cukup sesuai sebagai dasar penilaian ketahanan.
2. Seleksi – Pengujian Ketahanan
Uji saring untuk mendapatkan tetua sumber gen tahan merupakan langkah awal kegiatan pemuliaan ketahanan. Pengujian ketahanan untuk tujuan seleksi genotipe pada generasi segregasi akan memerlukan jumlah cuplikan sangat banyak, sehingga sebelum menjadi genotipe harapan pengujian dilakukan pada aras bibit dengan cara inokulasi buatan. Dalam melakukan pe-ngujian ketahanan terhadap nematoda parasit pada aras bibit secara inokulasi buatan diperlukan inokulum dalam jumlah banyak, sehingga perbanyakan inokulum (rearing) merupakan kegiatan awal sebelum pengujian untuk mendapatkan genotipe tahan (Young, 1998). Pada jenis nematoda berpindah (migratory) seperti nematoda R. similis dan P. coffeae penyimpanan dan perbanyakan inokulum dalam bentuk telur maupun larva dalam jaringan akar sehat merupakan cara yang banyak dilakukan oleh seleksionis, sehingga kebutuhan inokulum potensial dalam jumlah banyak akan dapat dipenuhi dan selalu terbarui untuk pengujian tersebut. Cara pengujian ketahanan seperti
itu dapat menghindari kesalahan interpretasi tahan sebagai akibat terhindar (escape). Selanjutnya genotipe-genotipe tahan terpilih dilakukan pengujian di lahan endemik serangan nematoda.
Hasil seleksi dan uji saring (screening test) terhadap P. coffeae pada beberapa genotipe koleksi plasma nutfah kopi Robusta di Indonesia semula diperoleh dua nomor seleksi dengan tingkat ketahanan moderat, yaitu Qouillou 121—09 serta BP 961 (Wiryadiputra et al., 1998). Sementara itu pada hamparan populasi kopi Robusta yang ditanam di lahan endemik P. coffeae ditemukan klon BP 308 yang sehat dan tidak terserang nematoda. Hasil pengujian dengan cara inokulasi buatan juga menunjukkan bahwa klon tersebut tahan, sehingga akhirnya dilepas sebagai klon anjuran batang bawah tahan nematoda P. coffeae (Mawardi et al., 2003).
3.Evaluasi ketahanan beberapa genotipe kopi
Teknik evaluasi ketahanan untuk uji saring setiap spesies tanaman sudah cukup banyak dipublikasikan. Dalam penerapan-nya, teknik evaluasi ini diperlukan adaptasi dan optimasi untuk membedakan antara genotipe yang dikategorikan tahan dengan rentan, terutama untuk mengeliminasi perbedaan kondisi lingkungan serta inokulum yang digunakan dalam pengujian (Young, 1998).
Sebagaimana diuraikan di atas, setelah dilakukan perbanyakan inokulum (rearing) untuk mendapatkan larva yang paling agresif (potensial) perlu dilakukan beberapa
penelitian pendahuluan untuk menetapkan suatu genotipe kopi tahan atau rentan, yaitu: Optimasi volume media dalam kaitannya dengan penetapan jumlah inokulum untuk setiap tanaman yang diuji, serta masa inkubasi. Berdasarkan uji pen-dahuluan tersebut maka dapat ditetapkan metode evaluasi ketahanan.
Evaluasi ketahanan kopi terhadap nematoda parasit pada fase bibit diamati setelah 3 bulan inokulasi atau sebelumnya, yaitu setelah timbulnya gejala nekrosis daun berupa bercak berwarna coklat di bagian ujung daun. Sebagai tolok ukur ketahanan diamati beberapa peubah komponen ketahanan, yaitu: 1). selisih berat biomasa bibit, 2). persentase selisih berat akar, 3). jumlah nematoda dalam akar, 4). jumlah nematoda dalam tanah, 5). reproduksi serta 6). persentase akar nekrosis. Berdasarkan enam peubah tersebut dilakukan analisis untuk memilah genotipe yag diuji menjadi lima tingkat ketahanan, yaitu tahan, agak tahan, agak rentan, rentan dan sangat rentan (Hulupi, 2006 a).
Pada pengujian ketahanan fase tanaman muda sampai berbuah di lahan endemik, evaluasi ketahanan dilakukan setelah 1 tahun penanaman di lapangan, atau sebelumnya, yaitu jika telah menunjukkan gejala daun menguning dan pertumbuhan kerdil. Pengamatan dilakukan dua kali setahun, yaitu pada saat awal musim kemarau (bulan April) dan akhir musim kemarau (bulan September). Sebagai tolok ukur ketahanan tanaman di lahan endemik serangan nematoda diamati beberapa peubah komponen ketahanan yang meliputi: 1). banyaknya nematoda dalam 10 g akar/
pohon, 2). banyaknya nematoda dalam 100 ml tanah/pohon, 3). persentase nekrosis akar dan 4). persentase tanaman mati untuk setiap petak uji. Untuk tiga peubah pertama, setiap petak dicuplik 20 % dari jumlah pohon yang diuji (Hulupi et al., 2007).
4. Mekanisme ketahanan kopi terhadap nematoda parasit
Dalam pemuliaan ketahanan, penge-tahuan tentang mekanisme ketahanan terhadap patogen akan memudahkan dalam menetapkan variabel penilaian komponen ketahanan. Padahal sebagaimana diketahui, mekanisme ketahanan kopi terhadap nematoda parasit belum banyak dilaporkan. Secara biokimiawi Giebel (1974) membagi mekanisme ketahanan menjadi beberapa tipe, yaitu 1). Tanaman menghasilkan eksudat yang bersifat toksin yang mampu membunuh nematoda. Beberapa senyawa kimia yang berfungsi menghambat adalah senyawa fitoaleksin, senyawa prapem-bentukan molekul beracun dan enzim peng-hambat (Dropkin, 1992). Sebagai contoh adalah hasil penelitian Trudgill (1991), bahwa akar Tagetes patula dan T. erecta mengeluarkan á–tertienil dan keturunan bitienil yang efektif melawan nematoda Meloidogyne spp. dan P. coffeae yang menyerang kopi robusta. Hasil penelitian tersebut kemudian diterapkan sebagai salah satu metode pengendalian di areal perkebunan kopi Robusta di Jawa Timur (Wiryadiputra et al., 1998), meskipun akhirnya difahami penerapan dalam skala luas dipandang kurang menarik karena T. patula tidak dapat dimanfaatkan
sebagai tanaman sela yang produktif. 2). Ketahanan terbentuk karena tanaman tidak mengandung senyawa yang di-perlukan untuk pertumbuhan dan reproduksi nematoda. Tanaman mengeluarkan khloroflorenol, yaitu suatu senyawa penghambat pada kelompok morfaktin yang menghambat pembentukan sel raksasa pada puru akar tomat yang terinfeksi Meloidogyne spp. sehingga menyebabkan kehilangan nutrisi untuk pertumbuhan seksual nematoda. Hal ini ditandai dengan meningkatnya persentase nematoda jantan (Kaplan, 1994). 3). Tanaman tidak mengandung senyawa pemikat bagi nematoda. 4). Ketahanan terjadi karena reaksi hipersensitivitas jaringan tanaman terhadap nematoda yang menghambat perkembangan nematoda.
Mekanisme ketahanan kopi terhadap serangan nematoda secara fisik oleh Dropkin (1992) dipilah menjadi beberapa tipe, yaitu: 1). akar kopi memiliki sel epi-dermis yang dilapisi oleh lignin, 2). akar mengandung bulu-bulu akar, 3). akar kopi dilapisi oleh lilin. Hasil pengujian awal pada irisan melintang akar kopi Arabika yang me-nunjukkan reaksi tahan meme-nunjukkan adanya indikasi mekanisme ketahanan fisik berupa lapisan lignin pada dinding selnya yang diduga berperan sebagai benteng (Hulupi, 2006a), namun pada tanaman kopi Robusta tahan (BP 308) tampaknya tidak demikian. Diduga mekanisme ketahanannya berupa senyawa kimia yang menyebabkan nematoda tidak suka mendekati jaringan akar klon tahan tersebut. Untuk mengetahui senyawa eksudat akar yang berperan sebagai penghambat tersebut diperlukan
pengujian lebih mendalam. Sambil menunggu hasil penelitian mekanisme ketahanan kopi terhadap nematoda parasit, pada saat ini identifikasi genotipe tahan maupun rentan dapat dilakukan dengan cara mengamati gejala kerusakan yang ditimbulkan setelah masa inkubasi.
5. Sumber gen ketahanan kopi terhadap nematoda parasit
Informasi sumber gen ketahanan tanaman terhadap nematoda parasit penting diketahui sebagai langkah awal untuk menentukan strategi pemuliaannya. Adanya gen-gen ketahanan terhadap nematoda parasit pada tanaman yang telah berhasil diidentifikasi sampai saat ini merupakan gen ketahanan pada tanaman semusim, yaitu kentang, barley, kedelai dan tomat (Sidhu et al., 1981; Williamson & Hussey, 1996; Liharska, 1998). Pada tanaman tomat gen ketahanan terhadap Meloidogyne spp. dikenal dengan sebutan gen R (resistance) Mi, sedangkan pada tanaman kentang dikenal dengan gen HI dan pada sejenis kacang-kacangan (Cowpea = Vigna ungui-culata L.) gen ketahanan yang berhasil diidentifikasi disebut sebagai gen Rk. Penelitian gen-gen ketahanan kopi terhadap nematoda parasit sampai sekarang belum banyak dilakukan. Di India dan Brazil penelitian gen ketahanan terhadap nematoda ditekankan pada nematoda Meloidogyne spp. yang menyerang kopi Arabika maupun Robusta, dan hal ini pun tidak banyak dipublikasikan. Di antara yang dilaporkan adalah ketahanan beberapa bastar dan keturunan F2 hasil persilangan kopi Arabika
yang telah dibuat. Meskipun demikian hasilnya tidak seperti yang telah dicapai pada tanaman semusim (Kumar, 1980; Campos et al., 1990; Anzueto et al., 1993; Bertrand et al., 1995).
6. Genetika ketahanan kopi terhadap nematoda parasit
Penelitian genetika ketahanan tanaman terhadap nematoda parasit sebagian besar dilakukan pada tanaman semusim, yaitu terhadap nematoda Meloidogyne incognita, M. javaniva dan M. hapla. Hasil penelitian menyebutkan bahwa sifat ketahanan diatur oleh beberapa gen minor (polygenic) dengan aksi gen aditif yang mempengaruhi sifat ketahanan tersebut (Roberts et al., 1998). Hasil penelitian tindak gen ketahanan terhadap nematoda Meloidogyne spp. yang menyerang kopi Arabika Catimor pernah dilaporkan oleh Bertrand et al. (1995) sebagai tindak gen dominan sebagian, khususnya untuk peubah jumlah massa telur dan laju pembentukan puru. Hasil penelitian serupa juga diperoleh Roberts et al. (1998) yang menyebutkan adanya peran gen dominan sebagian yang mengatur ketahanan vatietas tomat terhadap Meloidogyne spp., sehingga gen ketahanan pada kondisi homosigot akan melebihi ketahanan pada keadaan heterosigot.
Klon kopi Robusta BP 308 meskipun diketahui tahan terhadap serangan P. coffeae dan R. similis (Mawardi et al., 2003), tetapi belum diketahui genetika ketahanannya. Berdasarkan hasil penelitian genetika ketahanan pada kopi Arabika
terhadap serangan R. similis diketahui bahwa sifat ketahanan BP 542 A di-kendalikan oleh gen tunggal (monogenic) yang bersifat dominan sempurna, sehingga keturunannya akan menghasilkan individu tanaman tahan sebanyak 75% dari populasi, sedangkan 25% sisanya rentan. Pada persilangan antara BP 542 A dengan beberapa genotipe rentan (Kartika 1 dan Andungsari 1) gen tahan menunjukkan dominansi lebih, sedangkan aksi gen aditif dan gen dominan tampaknya mempengaruhi nilai ketahanan pada keturunannya. Pola segregasi generasi F1 pada sebagian besar komponen ketahanan mengikuti pola pewarisan digenik, dengan nisbah segregasi tahan : rentan = 3 : 1, sedangkan pada generasi F2, pola segregasinya tidak sesuai mengikuti pola pewarisan monogenik maupun digenik. Ketidak-sesuaian pola pewarisan tersebut diduga karena adanya interaksi gen non alelik (epistasis) yang turut berperan dalam menentukan ekspresi ketahanan (Hulupi, 2006a).
STRATEGI PEMULIAAN KETAHANAN KOPI TERHADAP
NEMATODA PARASIT
Strategi pemuliaan untuk mendapatkan genotipe kopi tahan didasarkan pada hasil penelitian identifikasi sumber gen tahan serta genetika ketahanan tetua tahan, sedangkan informasi mekanisme ketahanan bermanfaat dalam menetapkan peubah (variable) komponen ketahanan untuk setiap tahap pemuliaan. Berdasarkan hasil pengujian-pengujian yang telah dilakukan tersebut kemudian diatur strategi pemuliaannya.
Prosedur pemuliaan ketahanan yang digunakan untuk mendapatkan kultivar kopi tahan nematoda tergantung pada tipe penyerbukannya, dayawarisnya, jumlah gen yang berperan mengatur sifat tahan, serta sifat adaptabilitas tetua donor ketahanan (Young, 1998). Kopi Arabika mempunyai susunan genetik berbeda dengan kopi Robusta, demikian pula dengan sifat penyerbukannya, sehingga strategi pemuliaan ketahanan masing-masing jenis kopi tersebut diatur berbeda.
Sebagaimana diuraikan dalam bab sebelumnya, bahwa sebaran populasi nema-toda R. similis paling banyak berada pada kedalaman tanah lebih dari 50 cm di bawah permukaan tanah, sedangkan P. coffeae lebih banyak berada pada zona perakaran kurang dari 30 cm. Sementara itu kopi Arabika memiliki sebaran akar mencapai kedalaman lebih dari 60 cm di bawah permukaan tanah, sedangkan sebaran akar kopi Robusta sebagian besar terletak pada kedalaman kurang dari 30 cm (Pujiyanto et al., 1993 dan Nur et al., 2000). Oleh sebab itu perbedaan sebaran akar kedua jenis kopi tersebut berdampak pada patogenisitas kedua jenis nematoda tersebut terhadap masing-masing jenis kopi. Artinya, jika pada lahan ketinggian menengah (700– 900 m dpl.) ditanam 2 jenis kopi (Arabika dan Robusta), meskipun dua jenis nematoda, R. similis dan P. coffeae dapat menimbulkan kerusakan pada 2 jenis kopi tersebut, namun R. similis lebih merusak kopi Arabika dibanding terhadap kopi Robusta. Berdasarkan hasil penelitian tersebut maka pemuliaan ketahanan kopi Arabika terhadap nematoda parasit lebih efektif jika diarahkan
terhadap nematoda R. similis, karena genotipe kopi Arabika yang tahan R, similis sekaligus juga tahan terhadap P coffeae, namun sebaliknya jika genotipe kopi Arabika tahan terhadap P. coffeae belum tentu tahan terhadap R. similis. Selanjutnya pemuliaan ketahanan kopi Robusta lebih difokuskan terhadap nematoda P. coffeae.
Bentuk bahan tanam kopi tahan nema-toda parasit yang akan dikembangkan dapat dirancang dalam bentuk benih ataupun klonal (van der Vossen, 1985). Bahan tanaman kopi dalam bentuk benih menurut pengalaman selama ini lebih mudah diadopsi oleh pekebun dibanding bahan tanaman klonal, namun sebagai tanaman tahunan pengembangan dalam bentuk benih memiliki risiko mengalami segregasi sifat ketahanan. Di samping itu untuk menghasilkan bahan tanam kopi tahan nematoda berupa benih memerlukan waktu lama, karena menyangkut persilangan balik (back cross) serta pengujian keturunan berulang-ulang.Sementara itu pembentuk-an biotipe/ras baru sebagai akibat tekpembentuk-anpembentuk-an seleksi sangat cepat, sehingga timbul anggapan bahwa untuk dapat menghasilkan varietas kopi tahan seolah-olah pemulia dipaksa berpacu melawan pembentukan biotipe/ras baru tersebut. Salah satu metode pemuliaan ketahanan tanaman tahunan yang dapat memotong daur seleksi adalah dengan mengembangkan hibrida F1 bersari bebas (open pollinated) dari dua tetua yang mewariskan sifat tahan. Dalam hal ini tetua tahan harus memiliki sifat daya gabung khusus terhadap genotipe tetua lain untuk menghasilkan keturunan tahan. Metode pemuliaan ketahanan lain yang dapat
memotong daur seleksi adalah metode pemuliaan populasi yang menghasilkan varietas sintetik atau varietas komposit. Dalam pelaksanaannya, varietas tahan dirakit dalam satuan populasi tanaman yang terdiri dari beberapa genotipe dengan gen ketahanan berbeda, sehingga ketahanan varietas tersebut bersifat awet (durable) serupa varietas yang memiliki ketahanan horisontal, yang disusun oleh banyak gen ketahanan (Arunachalam & Katiyar, 1982).
Pemuliaan ketahanan kopi Arabika terhadap nematoda parasit
Kopi Arabika bersifat menyerbuk sendiri, sehingga bahan tanam anjuran tahan dapat berupa varietas ataupun klon. Mengingat sebagian besar genotipe kopi sumber gen tahan maupun sumber gen dayahasil tinggi bersifat heterosigot, maka bastar F1 dapat dipastikan juga heterosigot, dan sifat ketahanan serta dayahasil akan memisah pada keturunannya (Hulupi et al., 2007). Untuk menyingkat proses pemuliaan ketahanan kopi Arabika terhadap nematoda R. similis, setelah diperoleh genotipe sebagai sumber gen tahan maka kegiatan selanjutnya adalah membuat persilangan dialel yang terdiri atas tetua tahan serta tetua produksi tinggi dengan keragaman genetik cukup luas. Tetua yang memiliki daya gabung khusus tinggi untuk sifat tahan dengan dayahasil yang tinggi pula dapat dirancang untuk menghasilkan bastar F1. Selain itu beberapa bastar F1 dan resiprok tahan serta beberapa bastar silang balik tahan jika dikumpulkan menjadi genepooled potensial dapat dirakit menjadi varietas komposit yang memiliki ketahanan awet,
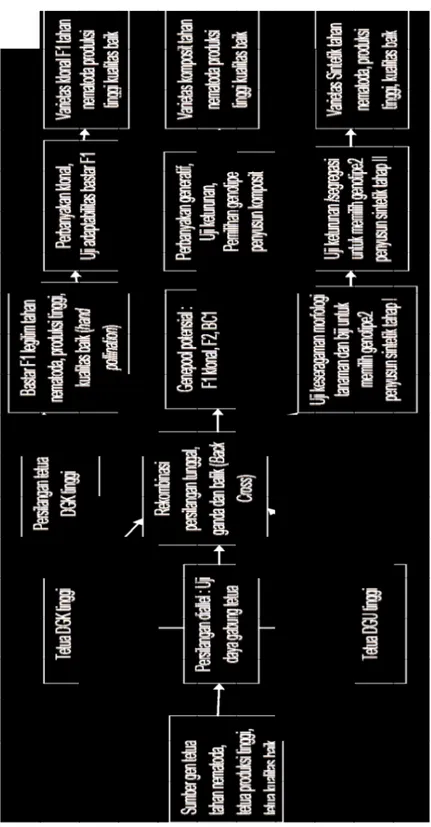
karena seolah-olah merupakan varietas yang disusun banyak gen (polygenic) serupa varietas yang memiliki ketahanan horisontal. Genotipe-genotipe tahan yang terkumpul menjadi genepooled potensial selanjutnya diuji di lahan endemik untuk dipilih beberapa genotipe yang memiliki sumber gen ketahanan berbeda, namun mempunyai keragaan morfologi serupa dan ukuran biji seragam, sebagai penyusun varietas komposit (Gambar 1).
Jika varietas dibangun dalam satuan populasi, maka bentuk varietas komposit merupakan cara yang dapat menyingkat waktu seleksi dibanding seleksi pedigree untuk mendapatkan lini homosigot. Akan tetapi untuk membangun pemuliaan populasi seperti ini diperlukan keragaman genetik yang luas untuk sifat tahan nematoda parasit sebagai modal dasar membentuk genepool potensial. Jika bahan tanam yang akan dikembangkan berupa klon maka berdasar-kan hasil uji saring dilakuberdasar-kan pengujian vegetatif di lahan endemik, sekaligus untuk mengetahui adaptabilitasnya pada kondisi lingkungan penanaman. Mengingat sebaran populasi nematoda R. similis mencapai kedalaman lebih dari 50 cm di bawah permukaan tanah, sedangkan pola sebaran perakaran kopi Arabika juga mencapai kedalaman lebih dari 50 cm, maka pemuliaan ketahanan kopi Arabika diarahkan terhadap nematoda R. similis.
Pemuliaan ketahanan kopi Robusta terhadap nematoda parasit
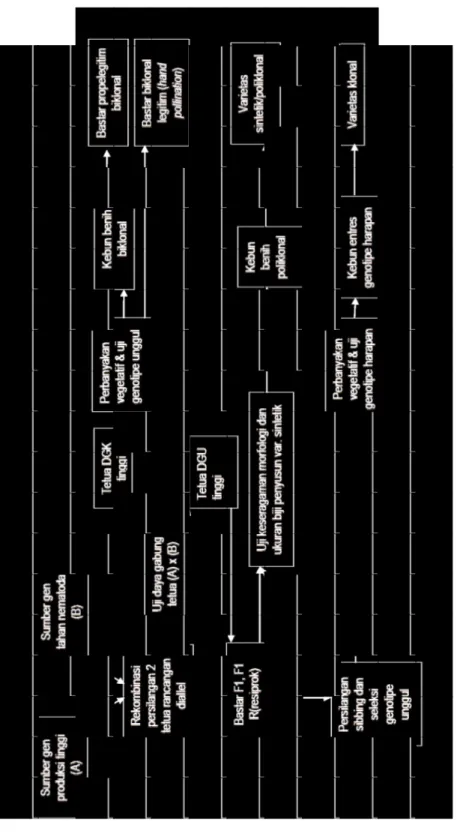
Pada kopi Robusta yang bersifat menyerbuk silang, bahan tanam tahan dirancang untuk menghasilkan bastar
Ga m ba r 1. Ba ga n s ke m at is pe m uli aa n k et ah an an k op i A ra bik a t er ha da p ne m at od a p ar as it. Fi gu re 1 . Sc he m e o f b re ed in g f or re sis ta nc e o f A ra bic a c of fe e t o p ar as itic n em at od es .
Ga m ba r 2. Sk em a p em ulia an k et ah an an k op i Ro bu sta te rh ad ap n em at od a p ar as it. Fi gu re 2 . Sc he m e o f b re ed in g f or re sis ta nc e o f Ro bu sta co ffe e t o p ar as itic n em at od es .
biklonal yang tetuanya mempunyai nilai daya gabung khusus cukup tinggi untuk sifat tahan terhadap nematoda parasit. Bastar biklonal dapat dilepas dalam bentuk benih F1 atau varietas klonal dari bastar F1 maupun F2 sibling unggul. Apabila tetua tahan memiliki sifat daya gabung umum tinggi terhadap beberapa genotipe tetua lain maka bentuk varietas yang dirancang adalah varietas sintetik. Serupa dalam mendapat-kan varietas sintetik pada jagung, varietas sintetik pada kopi Robusta satuannya adalah populasi yang disusun oleh beberapa genotipe F1 dan F2 sibling yang memiliki sifat tahan sama namun berbeda sumber gen ketahanannya, kemudian dicampur (bulked) membentuk satu varietas yang memiliki beberapa gen ketahanan berbeda sehingga menghasilkan ketahanan yang awet serupa dengan ketahanan horisontal pada satu genotipe. Pengujian ketahanan cukup hanya dilakukan satu generasi keturunannya saja, yaitu untuk mengetahui besarnya segregasi sifat ketahanan apabila varietas tersebut di-kembangkan secara meluas.
Pemuliaan ketahanan kopi terhadap nematoda puru akar, Meloidogyne sp. yang dilakukan di beberapa negara lain diawali dengan cara uji saring beberapa genotipe kopi koleksi yang menunjukkan kurangnya jumlah puru jika ditanam di lahan endemik serangan maupun jika dilakukan uji inokulasi buatan pada fase bibit. Pada nematoda berpindah seperti P. coffeae dan R. similis belum diketahui secara pasti mekanisme ketahanannya, namun beberapa kemungkinan yang dapat terjadi pada sistem ketahanan kopi Robusta klon BP 308 adalah memiliki sebaran akar yang sangat banyak,
sehingga ketahanan semacam ini dapat dikatakan sebagai ketahanan semu, karena bersifat terhindar (escape). Namun hasil pengamatan pada kopi excelsa yang juga memiliki sebaran akar yang meluas hal ini membuktikan bahwa pendapat tersebut belum tentu benar dan perlu dilakukan penelitian lebih mendalam.
K E S I MP U L A N
1. Uji saring pada beberapa koleksi plasma nutfah kopi untuk mendapatkan tetua sumber gen tahan serangan nematoda parasit merupakan langkah awal kegiatan pemuliaan ketahanan.
2. Untuk melakukan seleksi ketahanan kopi terhadap nematoda parasit, pemilahan tingkat ketahanan menjadi : 1). kebal (im-mune), 2). sangat tahan (highly tant), 3). agak tahan (moderately resis-tant), rentan (susceptible), dan 5). sangat rentan (highly susceptible) cukup sesuai sebagai dasar penilaian ketahanan. 3. Kopi Arabika mempunyai susunan
genetik berbeda dengan kopi Robusta, demikian pula dengan sifat penyer-bukannya, sehingga strategi pemuliaan ketahanan masing-masing jenis kopi tersebut diatur berbeda. Untuk mengatur strategi pemuliaan kopi terhadap nematoda parasit di Indonesia, sebelum-nya perlu dipelajari hal-hal yang berkaitan dengan perilaku nematoda P. coffeae dan R. similis, mekanisme ketahanan serta genetika ketahanannya untuk setiap jenis kopi (Arabika dan Robusta).
4. Metode pemuliaan ketahanan kopi terhadap nematoda parasit yang dapat memotong daur seleksi adalah metode pemuliaan populasi yang menghasilkan varietas sintetik atau varietas komposit, sifat ketahanan bersifat awet (durable) serupa ketahanan horisontal, yang di-susun oleh banyak gen ketahanan dalam satuan populasi.
5. Berdasarkan beberapa hasil penelitian, pemuliaan ketahanan kopi Arabika terhadap nematoda parasit diarahkan terhadap nematoda R. similis, sedangkan pemuliaan ketahanan kopi Robusta terhadap nematoda P. coffeae.
DAFTAR PUSTAKA
AEKI (2007). Statistik Kopi 2005–2007. Asosiasi Eksportir Kopi Indonesia, 82 hal.
Anzueto, F. (1993). Étude de la resistance du Caféier (Coffea sp.) a Meloydogyne sp. et. Pratylenchus sp. Thése de Docteur, IRFA & CIRAD-CP, Montpellier, 123 p.
—————; A.B. Eskes; J.L. Sarah; B. Decazy (1993). Résistance de Quelqes des-cendances de Coffea arabica et Coffea canephora vis-à-vis de deux populations de Meloydogyne spp., originaires du Guatemala et du Brésil. ASIC, 15e Colloque, Montpellier,
338—349.
Arango-Bernal, L.G.; C.A. Baeza-Aragony; J.E. Lequizamon-Caicedo (1982). Pruebas de resistencia a species de Meloy-dogyne en plantulas de Coffeae spp. ASIC 15e Colloque, Montpellier,
338—349.
Arunachalam, V. & R. K. Katiyar (1982). A Viable short-term strategy for breed-ing composite populations. Indian J. Genet., 42, 32—37.
Bertrand, B.; F. Anzueto; M.X. Pena; F. An-thony & A.B. Eskes (1995). Genetic improvement of coffee for resistance to root-knot nematodes (Meloydogyne spp.) in Central America. ASIC, 16 e Colloque, Kyoto, p. 630—636. Brown, J.F. & R.C. Colbran (1980).
Ex-amples of diseases caused by nema-todes. p. 57—70. In: J.F. Brown (ed.). Plant Protection, AAUCS. Campos, V.P.; P. Sivapalan & N.C.
Gnanap-ragasam (1990). Nematodes parasites of coffee, cocoa and tea. p. 387—460. In: M. Luc, R.A. Sikora & J. Bridge (eds.). Plant parasitic nematodes in subtropical and tropical agriculture. CAB Int, Wallingford, UK.
Collingborn, F.M.B.; S.R. Gowen & I. Mueller-Harvey (2000). Investigations in to the Biochemical basis for Nema-tode-resistance in roots of three Musa cultivars in response to Radopholus similis infection. J. Agric.Food Chem., 48, 5297–5301.
Dalmasso, A.; P. Castagnone-Sereno & P. Abad (1992). Seminar : Tolerance and Resistance of Plants to Nema-todes- Knowledge, Needs and Pros-pects. Nematologica, 38, 466–472. Dropkin, V.H. (1992). Introduction to Plant
Nematology. Edisi Bahasa Indonesia, diterjemahkan oleh Supratoyo dan disunting oleh Mulyadi, Gadjah Mada Press, Yogyakarta, 366 p.
Giebel, J. (1974). Biochemical Mechanisms of Plant Resistance to Nematode : A Review. J. Nematol, 6, 174–184.
Gonzaga, L. & E. Lordello (1986). Plant-para-sitic nematodes that attack coffee. p. 33–41. In: J. Román (ed.), Plant Parasitic Nematodes of Bananas, Cit-rus, Grapes and Tobacco.
Horber, E. (1980). Types and Classification of Resistance. p. 15–22. In: F.G. Maxwell & P.R. Jennings (eds.). Breeding Plants Resistant to Insects, John Wiley & Sons, N.Y.
Hulupi, R. (2006 a). Identifikasi Ras Fisiologi Nematoda Radopholus similis Cobb. Yang menyerang tanaman kopi. Pelita Perkebunan, 22, 213–221.
... (2006 b). Kajian Genetika Ketahanan Kopi Arabika terhadap Nematoda Penggali Akar, Radopholus similis Cobb. Disertasi. Universitas Gadjah Mada, Yogyakarta, 137 hal. ... & Mulyadi (2007.) Sebaran populasi nematoda Radopholus similis dan Pratylenchus coffeae pada lahan per-kebunan kopi. Pelita Perkebunan, 23, 176–183.
...; Nasrullah & Soemartono. (2007). Pewarisan ketahanan kopi Arabika terhadap nematoda Rado-pholus similis Cobb. Pelita Per-kebunan, 23, 1–16.
..., Soemartono; Mulyadi & Nasrullah (2006). Ketahanan Beberapa Genotipe Kopi terhadap Nematoda Penggali Akar, Radopolus similis, Cobb. Agro-sains, 19, 337–346.
Kaplan, D.T. (1994). Molecular characteriza-tion of burrowing nematode sibling species, Radopholus citrophillus and R. similis. p. 77–83. In: F. Lamberti, C. De Georgi & David Mck. Bird (eds.). Advance in Molecular Plant
Nematology, Plenum Press, London. Kumar, A.C. (1980). Studies on nematodes in coffee soils of South India: 3. A Report on R. similis and description of R. colbrani n.sp. J. Coffee Res. 10, 43–46.
……… (1991). Host-parasitic relationship between certain plantation crops and the two races of the coffee nematode, Pratylenchus coffeae. J. Coffee Res., 21, 103–108.
……… & S.D. Samuel (1990). Nema-todes attacking coffee and its manage-ment : A review. J. Coffee Res., 20, 1–27.
Kusno-Amidjojo, M. (1995 a). Radopholus similis Cobb nematoda parasit ber-bahaya pada tanaman kopi Arabika. Warta Pusat Penelitian Kopi dan Kakao, 11, 129–133.
... (1995 b). Setaria plicata, Inang Pratylenchus coffeae dan Peranannya di Tanah Bera bekas Tanaman Kopi. Pelita Perkebunan 11, 31–37. Liharska, T. B. (1998). Genetic and
Molecu-lar Analysis of the Tomato Root-Knot Nematode Resistance Locus Mi – 1. Doctoral Thesis from Wageningen Agricultural University, 95 p. (Un-published).
Mawardi, S.; G. Suprijadji; S. Wiryadiputra; A.M. Nur; Sudarsianto; A. Soedarsan & F.F. Leupen (2003). BP 308: Klon harapan kopi robusta tahan terhadap nematoda parasit, Pratylenchus coffeae, Warta Pusat Penelitian Kopi dan Kakao Indonesia, 17, 161–171. Nur, A.M.; Zaenudin & S. Wiryadiputra
kopi Robusta klon BP 308 pada lahan endemik nematoda parasit, Praty-lenchus coffeae. Pelita Pekebunan, 16, 121–131.
Noe, J.P. (1998). Crop and Nematode-Man-agement Systems. p. 159–171. In: K.R. Baker, G.A. Pederson, G.L. Windham (Coeds.), Plant and Nema-tode Interactions, Madison, Wiscon-sin, USA Publ.
Pujiyanto, S. Mawardi & A.M. Nur (1991). Kajian sebaran akar rambut kopi Arabika tipe katai varitas BP 453 A dan BP 454 A. Pelita Perkebunan, 8, 79–84.
Roberts P.A.; W.C.Mathews & J.C. Veremis (1998). Genetic Mechanisms of Host-Plant Resistance to Nematodes. p. 209–238. In: K.R. Barker et al. (Coeds.). Plant and Nematode Inter-actions. Madison, Wisconsin, USA Publ.
Rodrigues Jr. (1988). Coffee Rust Resistance and Physiological Races. Proc. Int. Sem. On Coffee technology, Feb. 3–5, 1988, Chiang mai. 29–37. Román, J. (1986). Plant parasitic nematodes
of Bananas, Citrus, Coffee, Grapes and Tobacco. Research Triangle Park, North Carolina, USA. Union Agricultural Products Co. Inc. 71 p. Sidhu, G.M. & J.M. Webster (1981). Gene-tics of plant-nematode interactions. p. 61–85. In: B.M. Zuckerman & R.A. Rohde (eds.), Plant Parasitic Nematodes Volume III. Academic press, Inc.
Sijmons, P.C. (1993). Plant nematode inter-actions. Plant Molecular Biology, 23, 917–931.
Trudgill, D.L. (1991). Resistance to and to-lerance of plant parasitic nematodes in plants. Annu. Rev. Phytopath., 29, 167–193.
Van der Plank, J.E. (1968). Disease Resis-tance in Plant. Academic Press., Inc., New York, 206 p.
Van der Vossen, H.A.M (1985). Coffee se-lection and breeding. p. 48–96. In: M.N. Clifford & K.C. Willson (eds.). Coffee, Botany, Biochemis-try and Production of Beans and Be-verage. AVI Publ. Co. Inc.. Williams, K.J.O. & M.R. Siddiqi (1973).
Radopholus similis, Descriptions of plant parasitic nematodes, Set 2, 4p. Williamson, V.M. & R.S. Hussey (1996). Nematode pathogenesis and resistance in plants. The Plant Cell, 8, 1735– 1745.
Wiryadiputra, S. & R. Hulupi (1995). Uji ketahanan varietas kopi Arabika introduksi terhadap nematoda P. coffeae. Makalah Konggres Nasional XIII dan Seminar Ilmiah Perhimpunan Fitopatologi Indonesia. Mataram, 25–27 september 1995, 8p. Yahmadi, M. (1972). Budidaya dan Pengolahan Kopi. Balai Penelitian Perkebunan Jember, 36 p.
Young, L.D. (1998). Breeding for Nematodes Resistance and Tolerance. p. 187–208. In: K.R. Barker et al. (Coeds.). Plant and Nematode Interactions. Madison, Wisconsin, USA Publ.