REFERAT
Embriologi, Neuroanatomi dan Fisiologi
Talamus
Pembimbing:
dr. Tunggul Marpaung, Sp.BS, M.Kes
Disusun oleh:
Gabriela Enneria Sibarani
(09-61050-124)
KEPANITERAAN KLINIK ILMU BEDAH
PERIODE 18 NOVEMBER 2013 – 18 JANUARI 2014
FAKULTAS KEDOKTERAN
UNVERSITAS KRISTEN INDONESIA
KATA PENGANTAR
Puji dan syukur penulis panjatkan ke hadirat Tuhan Yang Maha Esa,
karena atas rahmat-Nya, penulis dapat menyelesaikan penyusunan referat yang
berjudul “Embriologi, Neuroanatomi, dan Fisiologi Talamus”.
Terima kasih penulis sampaikan kepada semua pihak yang telah banyak
mendukung dan membantu hingga selesainya referat ini.
1 dr. Tunggul Marpaung, Sp.BS., M.Kes selaku pembimbing referat.
2 dr. Anggi Simbolon, selaku asisten pembimbing referat yang senantiasa
meluangkan waktu untuk membimbing dalam penulisan referat.
3 Segenap staf Bagian Ilmu Kedokteran Bedah Fakultas Kedokteran
Universitas Kristen Indonesia.
4 Rekan-rekan kepaniteraan klinik Ilmu Kedokteran Bedah Fakultas
Kedokteran Universitas Kristen Indonesia yang telah memberikan bantuan
baik material maupun spiritual.
Dalam penyusunan referat ini, penulis merasa masih banyak kekurangan
baik secara teknik maupun materi penulisan, mengingat akan kemampuan yang
dimiliki penulis. Untuk itu kritik dan saran dari semua pihak sangat penulis
harapkan demi penyempurnaan pembuatan referat ini.
Jakarta, 13 Januari 2014
DAFTAR ISI
Kata Pengantar...i
Daftar Isi...ii
Daftar Gambar...iii
1. Pendahuluan………..iv
1.1 Latar Belakang………....iv
1.2 Rumusan Masalah………...…vi
2. Isi...1
2.1 Embriologi Talamus………...1
2.1.1 Rombensefalon: Otak Belakang...3
2.1.2 Mesensefalon: Otak Tengah...5
2.1.3 Prosensefalon: Otak Depan...6
2.2 Anatomi Talamus ………...12
2.2.1 Nuklei Talamus...14
2.2.2
Nuklei Thalami Spesifik dan Hubungan-hubungannya...152.2.3
Nuklei Thalami Nonspesifik dan Hubungan-hubungannya...172.3 Fisiologi Talamus ………...18
2.3.1 Sinyal Somatosensorik...19
2.3.2 Sensasi Penglihatan...28
2.3.3 Sensasi Pendengaran...30
3. Kesimpulan………...…33
4. Daftar Pustaka………..29
DAFTAR GAMBAR
Gambar 1. Pandangan dorsal mudigah pada usia (A) 18 hari; (B) 20 hari...1
Gambar 2. Pandangan dorsal mudigah pada usia (A) 22 hari; (B) 23 hari...1
Gambar 3. Potongan Sagital mudigah pada usia 27 hari...2
Gambar 4. Potongan Sagital mudigah pada usia 32 hari...2
Gambar 5. Diensefalon...12
Gambar 6. Nuklei Talamus...14
Gambar 7. Hubungan Aferen dan Eferen Kelompok Nuklear Medial (Merah), Dorsal (Ungu/Biru), dan Lateral (Biru)...17
Gambar 8. (A) Traktus Spinoserebelaris Anterior dan Posterior; (B) Kolumna Posterior; (C)
Traktus Spinotalamikus Anterior; (D) Traktus Spinitalamikus Lateralis...28
PENDAHULUAN
1.1 Latar Belakang
Sistem saraf, bersama dengan sistem endokrin, mengurus sebagian besar pengaturan
fungsi tubuh. Pada umumnya, sistem saraf mengatur aktivitas tubuh yang cepat, misalnya
kontraksi otot, perubahan viseral yang berlangsung dengan cepat, dan bahkan juga kecepatan
sekresi beberapa kelenjar endokrin. Sistem endokrin, sebaliknya, terutama mengatur fungsi
metabolik tubuh.
Sejak pembentukannya, sistem saraf mempunyai sifat-sifat mengatur yang sangat
kompleks dan khusus. Ia menerima berjuta-juta rangsangan informasi yang berasal dari
bermacam-macam organ sensorik, dan semua ini bersatu untuk dapat menentukan respon
yang akan diberikan oleh tubuh.
Sistem saraf terdiri atas lebih dari 100 juta neuron. Sinyal yang datang masuk melalui
sinaps pada dendrit-dendrit neuron atau badan sel; untuk berbagai jenis neuron, mungkin
hanya terdapat beberapa ratus atau sampai 200.000 sambungan sinaptik dari serabut yang
masuk. Selain itu, sinyal yang keluar berjalan melalui jalur akson tunggal meninggalkan
neuron, tetapi akson ini memiliki banyak cabang yang berbeda ke bagian-bagian lain sistem
saraf atau tubuh bagian perifer.
Bagian somatik sistem sensorik menghantarkan informasi sensorik dari reseptor di
seluruh permukaan tubuh dan beberapa struktur dalam. Informasi ini akan masuk ke dalam
sistem saraf pusat melalui saraf-saraf perifer dan dihantarkan ke berbagai area sensorik pada:
(a) Semua tingkat medula spinalis
(b) Substansia retikuler dari medula, pons, dan mesensefalon
(c) Serebelum
(d) Talamus
(e) Area somestetik dari korteks serebri
Namun di samping area sensorik primer ini, sinyal-sinyal terutama akan disiarkan dengan
baik ke bagian sistem saraf lainnya.
Peran yang paling penting dari sistem saraf adalah mengatur berbagai aktivitas tubuh. Hal ini
dapat dicapai melalui pengaturan:
(a) Kontraksi otot rangka seluruh tubuh
(b) Kontraksi otot polos organ dalam
(c) Sekresi kelenjar eksokrin dan endokrin di sebagian besar tubuh
Seluruh aktivitas ini disebut fungsi motorik sistem saraf, sedangkan otot dan kelenjar disebut
efektor karena otot dan kelenjar ini melaksanakan fungsinya sesuai yang diperintahkan oleh
sinyal sarafnya.
Sistem saraf manusia mempunyai sifat-sifat khas yang diturunkan pada setiap tahap
perkembangan evolusi. Dari sifat-sifat yang diwariskan ini, maka akan didapatkan tiga
tingkat utama dari sestem saraf pusat yang mempunyai sifat-sifat fungsional yang khas, yaitu:
1. Tingkat Medula Spinalis
Medula spinalis bukan hanya suatu saluran untuk menyalurkan sinyal yang
berasal dari perifer tubuh ke otak atau dari arah yang berlawanan, yakni dari otak
kembali ke tubuh. Walaupun medula spinalis telah dipotong setinggi daerah leher atas,
ternyata sebagian besar fungsi medula spinalis masih tetap ada. Contohnya, kontak
antara neuron-neuron dalam medula spinalis dapat menyebabkan timbulnya gerakan
berjalan, refleks untuk menarik bagian tubuh dari suatu objek, refleks untuk
mengeraskan kaki guna menunjang tubuh terhadap gaya berat, dan refleks yang
dipakai untuk mengatur pembuluh-pembuluh darah setempat, gerakan
gastrointestinal, dan masih banyak fungsi lainnya.
Cara kerja bagian atas sistem saraf sering tidak secara langsung mengirimkan
sinyal ke bagian perifer tubuh, melainkan dengan mengirim sinyal ke pusat-pusat
pengatur dalam medula spinalis, yang memerintah pusat-pusat medula spinalis untuk
berfungsi.
2. Tingkat Otak Bagian Bawah
Banyak, tetapi tidak semua. Aktivitas bawah sadar dari tubuh diatur oleh
bagian bawah otak, yaitu pada medula oblongata, pons, mesensefalon, hipotalamus,
talamus, serebelum, dan ganglia basalis. Pengaturan bawah sadar dari tekanan arteri
dan pernafasan terutama dicapai di dalam medula dan pons. Pengaturan
keseimbangan merupakan fungsi gabungan dari bagian serebelum serebelum yang
lebih dahulu terbentuk dari substansia retikuler dari medula, pons, dan mesensefalon.
Refleks untuk makan, seperti salivasi sebagai respon terhadap rasa makanan, dan
jilatan bibir diatur oleh daerah-daerah dalam medula, pons, mesensefalon, amigdala,
dan hipotalamus; sebagian besar pola emosi, seperti marah, sifat menyerang, aktivitas
seksual, reaksi terhadap nyeri, atau reaksi terhadap rasa senang, semuanya ini dapat
timbul pada binatang yang tidak mempunyai korteks serebri.
3. Tingkat Otak Bagian Atas atau Tingkat Korteks
Sesudah semua fungsi sistem saraf yang dapat terjadi pada tingkat medula spinalis
dan otak bagian bawah, korteks serebri merupakan gudang memori yang sangat besar.
Korteks itu tidak pernah berfungsi sendiri, tetapi selalu berhubungan dengan
pusat-pusat bagian bawah atau dari sistem saraf.
Tanpa adanya korteks serebri, maka fungsi pusat-pusat otak bagian bawah
sering sangat tidak teliti lagi. Tempat penyimpanan atau gudang informasi yang luas
dalam korteks biasanya akan mengubah fungsi-fungsi ini menjadi tindakan yang lebih
tepat dan tertentu.
Akhirnya, korteks serebri itu berguna untuk proses berpikir, tetapi tidak dapat
bekerja sendiri. Pada kenyataanya, pusat-pusat di bagian bawah otaklah yang
menimbulkan kesadaran dalam korteks serebri, sehingga akan membuka bank memori
menjadi mesin otak untuk berpikir.
1.2.
Rumusan Masalah
Dengan melihat latar belakang yang telah dikemukakan, maka timbullah pertanyaan
yang perlu dijawab dalam tulisan ini. Ada beberapa masalah yang dapat dirumuskan dan akan
dibahas dalam makalah ini, yaitu sebagai berikut.
i.
Apa itu talamus?
ii.
Bagaimana embriologi, anatomi dan fisologi dari talamus?
2.1. Embriologi Talamus
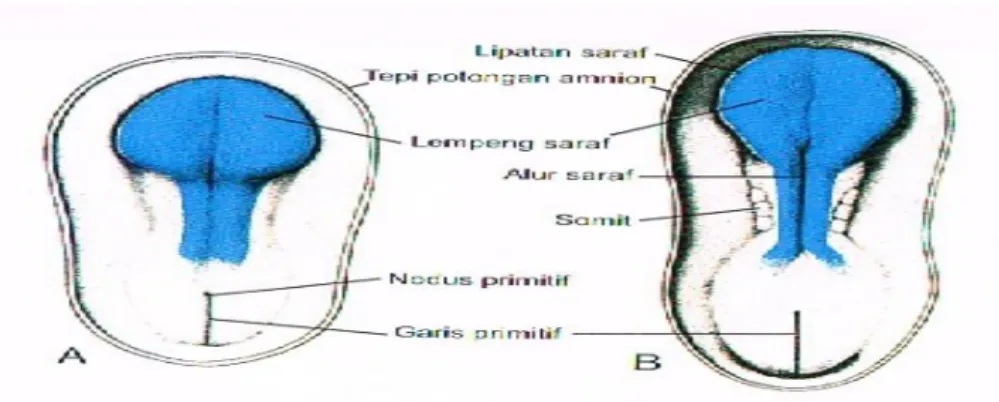
Sistem saraf pusat (SSP) muncul pada awal minggu ketiga sebagai suatu lempeng penebalan ektoderm berbentuk sandal, lempeng saraf (neural plate) di regio middorsal di depan primitive node
Gambar 1. Pandangan dorsal mudigah pada usia (A) 18 hari; (B) 20 hari
Seiring dengan perkembangan lebih lanjut, lipatan saraf tersebut terus meninggi, saling mendekati di garis tengah, dan akhirnya menyatu membentuk tabung saraf (neural tube). Penyatuan dimulai di daerah servikal dan berlanjut ke arah sefalik dan kaudal. Jika penyatuan telah dimulai, ujung-ujung bebas tabung saraf membentuk neuroporus kranialis dan kaudalis yang berhubungan dengan rongga amnion di atasnya. Penutupan neuroporus kranialis berlangsung ke arah kranial dari tempat penutupan awal di regio servikal dan dari suatu tempat di otak depan yang terbentuk belakangan. Tempat yang belakangan ini berjalan ke arah kranial, untuk menutup regio paling rostral tabung saraf, dan ke arah kaudal untuk bertemu dengan penutupan dari daerah servikal. Penutupan akhir neuroporus kranialis terjadi pada stadium 18 sampai 20 somit (hari ke 25); penutupan neuroporus kaudalis terjadi sekitar 2 hari kemudian.
Gambar 2. Pandangan dorsal mudigah pada usia (A) 22 hari; (B) 23 hari Ujung sefalik tabung saraf memperlihatkan tiga dilatasi, vesikel otak primer:
(a) Prosensefalon, atau otak depan (forebrain) (b) Mesensefalon, atau otak tengah (midbrain) (c) Rhombensefalon, atau otak belakang (hindbrain) Secara bersamaan ujung ini membentuk dua fleksura:
(a) Fleksura servikalis di taut otak belakang dan korda spinalis (b) Fleksura sefalika di regio otak tengah
Gambar 3. Potongan Sagital mudigah pada usia 27 hari Ketika mudigah berusia 5 minggu, prosensefalon terdiri dari dua bagian:
(a) Telensefalon yang dibentuk oleh bagian tengah dan dua kantong luar lateral, hemisferum serebri primitif, dan
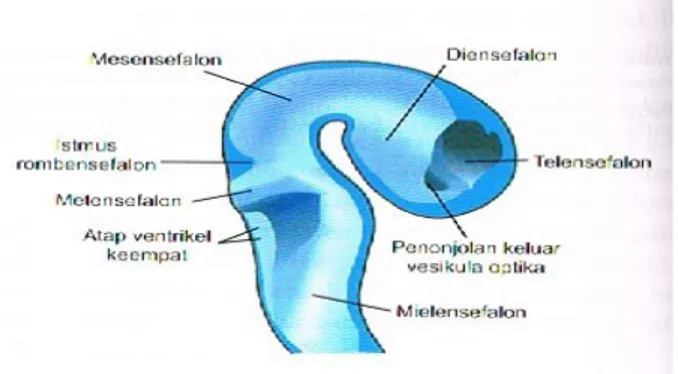
(b) Diensefalon yang ditandai oleh pertumbuhan keluar vesikel mata (vesikula optika). Suatu alur dalam, istmus rombensefalon, memisahkan mesensefalon dari rombensefalon
Rombensefalon juga terdiri dari dua bagian yang dibatasi oleh fleksura pontina: (a) Metensefalon yang kemudian membentuk pons dan serebelum, dan (b) Mielensefalon
Gambar 4. Potongan Sagital mudigah pada usia 32 hari
Lumen korda spinalis, kanalis sentralis, bersambungan dengan lumen vesikel otak. Rongga rombensefalon adalah ventrikel keempat, rongga diensefalon adalah ventrikel ketiga, dan rongga-rongga di hemisferum serebri adalah ventrikel lateral. Lumen mesensefalon menghubungkan ventrikel ketiga dan keempat. Lumen ini menjadi sangat sempit dan kemudian dikenal sebagai akueduktus Sylvius. Ventrikel lateral berhubungan dengan ventrikel ketiga melalui foramen interventrikulare Monro.
Di kedua sisi dari garis tengah di rombensefalon dan mesensefalon terdapat lempeng basal dan lempeng alar yang jelas terpisah dan masing-masing mempresentasikan daerah motorik dan sensorik. Namun, di prosensefalon, lempeng alar mengalami aksentuasi sementara lempeng basal mengalami regresi.
2.1.1 ROMBENSEFALON: OTAK BELAKANG
Rombensefalon terdiri dari mielensefalon, bagian paling kaudal vesikel otak, dan metensefalon yang berjalan dari fleksura pontina ke istmus rombensefalon
Mielensefalon
Mielensefalon adalah vesikel otak yang menghasilkan medula oblongata. Bagian ini berbeda dari korda spinalis karena dinding lateralnya mengalami eversi. Lempeng alar dan basal yang dipisahkan oleh sulkus limitans dapat dibedakan dengan jelas. Lempeng basal, serupa dengan yang terdapat di korda spinalis, mengandung nukleus-nukleus motorik. Nukleus-nukleus ini dibagi menjadi tiga kelompok:
(a) Kelompok eferen somatik medial
(b) Kelompok eferen viseral khusus intermediat, dan (c) Kelompok eferen viseral umum lateral
Kelompok pertama mengandung neuron-neuron motorik yang membentuk lanjutan sel kornu anterior ke arah sefalik. Karena berlanjut ke arah rostral ke mesensefalon, kelompok eferen somatik ini disebut kolumna motorik eferen somatik. Di mielensefalon, kelompok ini mencakup neuron-neuron nervus hipoglosus yang menyarafi otot lidah. Di metensefalon dan mesensefalon, kolumna masing-masing mengandung neuron nervus abdusens, nervus troklearis, dan nervus okulomotorius. Saraf-saraf ini menyarafi otot mata.
Kelompok eferen viseral khusus meluas ke metensefalon, membentuk kolumna motorik viseral khusus. Neuron-neuron motoriknya menyarafi otot lurik arkus faring. Di mielensefalon kolumna ini diwakili oleh neuron-neuron nervus aksesorius, nervus vagus, dan nervus glosofaringeus.
Kelompok eferen viseral umum mengandung neuron-neuron motorik yang menyarafi otot involunter saluran nafas, saluran cerna, dan jantung.
Lempeng alar mengandung tiga kelompok nukleus pemancar sensorik (sensory relay nuclei). Yang paling lateral dari ketiganya, kelompok aferen somatik (sensorik), menerima impuls dari telinga dan permukaan kepala melalui nervus vestibulokoklearis dan nervus trigeminus. Kelompok intermediat, atau aferen viseral khusus,menerima impuls dari papil pengecap lidah dan dari palatum,
orofaring, dan epiglotis. Kelompok medial, atau aferen viseral umum, menerima informasi interoseptif dari saluran cerna dan jantung.
Lempeng atap mielensefalon mengandung satu lapisan sel ependim yang ditutupi oleh mesenkim vaskular, pia mater. Kombinasi keduanya dikenal sebagai tela koroidea. Karena proliferasi aktif mesenkim vaskular, terbentuk sejumlah tonjolan invaginasi berbentuk kantong ke dalam rongga ventrikel di bawahnya. Invaginasi-invaginasi seperti umbai ini membentuk pleksus koroideus, yang menghasillkan cairan serebrospinal.
Metensefalon
Metelsefalon, serupa dengan mielensefalon, ditandai oleh lempeng basal dan alar. Dua komponen baru adalah:
(a) Serebelum, suatu pusat koordinasi untuk postur dan gerakan, dan
(b) Pons, jalur untuk serabut saraf antara korda spinalis dan korteks serebri serta korteks serebeli Masing-masing lempeng basal metensefalon mengandung tiga kelompok neuron motorik:
(a) Kelompok eferen somatik medial, yang menghasilkan nukleus abdusens
(b) Kelompok eferen viseral khusus, mengandung nukleus nervus trigeminus dan nervus fasialis, yang menyarafi otot arkus faring pertama dan kedua, dan
(c) Kelompok eferen viseral umum yang akson-aksonnya menyarafi kelenjar submandibula dan sublingual
Lapisan marginal lempeng basal metensefalon meluas sewaktu membentuk jembatan untuk serabut-serabut saraf yang menghubungkan korteks serebri dan korteks serebeli dengan korda spinalis. Karena itu, bagian metensefalon ini dikenal sebagai pons (jembatan). Selain serabut-serabut saraf, pons mengandung nukleus pons yang berasal dari lempeng alar metensefalon dan mielensefalon.
Lempeng alar metensefalon mengandung tiga kelompok nukleus sensorik:
(a) Kelompok aferen somatik lateral yang mengandung neuron nervus trigeminus dan sebagian kecil kompleks vestibulokoklear
(b) Kelompok aferen viseral khusus, dan (c) Kelompok aferen viseral umum
Serebelum
Bagian dorsolateral lempeng alar menekuk ke arah medial dan membentuk bibir rombik. Di bagian kaudal metensefalon, bibir ini terpisah jauh, tetapi tepat di bawah mesensefalon kedua bibir ini saling mendekat di garis tengah. Akibat semakin dalamnya fleksura pontina, bibir-bibir rombik menekan ke arah sefalokaudal dan membentuk lempeng serebelum. Pada mudugah 12 minggu, lempeng ini memperlihatkan bagian tengah yang kecil, vermis, dan dua bagian lateral, hemisfer.
Kemudian segera terbentuk suatu alur transversal yang memisahkan nodul dari vermis dan flokulus lateral dari hemisfer. Secara filogenetis, lobus flokulonodularis ini adalah bagian paling primitif dari serebelum.
Pada awalnya, lempeng serebelum terdiri dari lapisan neuroepitel, mantel, dan marginal. Selama perkembangan lebih lanjut, sejumlah sel yang terbentuk oleh neuroepitel bermigrasi ke permukaan serebelum untuk membentuk lapisan granular eksterna. Sel-sel lapisan ini mempertahankan kemampuannya untuk membelah dan membentuk suatu zona proliferatif di permukaan serebelum.
Pada bulan keenam perkembangan, lapisan granular eksterna menghasilkan berbagai jenis sel. Sel-sel ini bermigrasi ke arah sel Purkinje yang telah berdiferensiasi dan menghasilkan sel granular.
Sel keranjang (basket cell) dan sel stelata dihasilkan oleh sel-sel proliferatif di substansia alba serebelum. Korteks serebeli yang terdiri dari sel Purkinje, neuron Golgi II, dan neuron yang dihasilkan oleh lapisan granular eksterna, mencapai ukuran definitifnya setelah lahir. Nukleus-nukleus serebelum dalam, misalnya nukleus dentatus, mencapai posisi akhirnya sebelum kelahiran.
2.1.2 MESENSEFALON: OTAK TENGAH
Di mesensefalon, masing-masing lempeng basal mengandung dua kelompok nukleus motorik:
(a) Kelompok eferen somatik medial, yang diwakili oleh nukleus okulomotorius dan nervus troklearis yang menyarafi perototan mata, dan
(b) Kelompok eferen viseral umum kecil yang diwakili oleh nukleus Edinger-Westphal yang menyarafi m.sfingter pupilae.
Lapisan marginal masing-masing lempeng basal membesar dan membentuk krus serebri. Krusa ini berfungsi sebagai jalur untuk serabut saraf yang turun dari korteks serebri ke pusat-pusat yang lebih rendah di pons dan korda spinalis. Pada awalnya, lempeng alar mesensefalon tampak sebagai dua elevasi longitudinal yang dipisahkan oleh sebuah cekungan dangkal di garis tengah. Pada perkebangan lebih lanjut, terbentuk suatu alur transversal yang membagi masing-masing elevasi tersebut menjadi kolikulus anterior (superior) dan kolikulus posterior (inferior). Kolikulus posterior berfungsi sebagai stasiun pemancar sinaptik untuk refleks auditorik; kolikulus anterior berfungsi sebagai pusat korelasi dan refleks untuk impuls penglihatan. Kolikulus dibentuk oleh neuroblas yang bermigrasi secara bergelombang menuju zona marginal di atasnya. Di sini, sel-sel ini tersusun dalam lapisan-lapisan.
2.1.3 PROSENSEFALON: OTAK DEPAN
Prosensefalon terdiri dari telensefalon yang membentuk hemisferum serebri, dan diensefalon, yang membentuk cawan optik(optic cup) dan tangkai optik (optic stalk), hipofisis, talamus, hipotalamus, serta epifisis.
Diensefalon
Diensefalon, yang berkembang dari bgaian medial prosensefalon, diperkirakan terdiri dari suatu lempeng atap dan dua lempeng alar tetapi tidak memiliki lempeng lantai dan basal (yang menarik sonic hedgehog,suatu penanda garis tengah ventral, diekspresikan oleh lantai diensefalon yang mengisyaratkan bahwa sebenarnya terdapat suatu lempeng lantai). Lempeng atap diensefalon terdiri dari suatu lapisan sel ependim yang ependim yang ditutupi oleh mesenkim vaskular. Bersama-bersama, lapisan-lapisan ini menghasilkan pleksus koroideus ventrikel ketiga.bagian paling kaudal lempeng atap berkembang menjadi korpus pineale, atau epifisis. Korpus ini mula-mula tampak sebagai penebalan epitel di garis tengah, tetapi pada minggu ketujuh struktur ini mulai mengalami evaginasi. Akhirnya, badan ini berubah menjadi organ solid di atap mesensefalon yang berfungsi sebagai saluran penghantar gelap dan terang sehingga mempengaruhi irama endokrin dan perilaku. Pada orang dewasa, kalsium sering mengendap di epifisis dan kemudian berfungsi sebagai patokan dalam radiografi tengkorak.
Lempeng alar membentuk dinding lateral diensefalon. Suatu alur, sulkus hipotalamikus, membagi lempeng menjadi bagian dorsal dan ventral, masing-masing adalahtalamus dan hipotalamus. Akibat aktivitas proliferatif, talamus secara bertahap menonjol ke dalam lumen diensefalon. Ekspansi ini sering sedemikan hebatnya sehingga regio talamus dari sisi kanan dan kiri menyatu di garis tengah, membentuk massa intermedia atau koneksus intertalamikus.
Hipotalamus yang membentuk bagian bawah lempeng alar, berdiferensiasi menjadi sejumlah area nukleus yang mengatur fungsi alat dalam, termasuk tidur, pencernaan, suhu tubuh, dan perilaku emosi. Salah satu dari kelompok-kelompok ini, korpus mamilare, membentuk suatu tonjolan tersendiri di permukaan ventral hipotalamus di kedua sisi garis tengah.
Hipofisis terbentuk dari dua bagian yang sama sekali berbeda:
(a) Penonjolan ektoderm stomodeum tepat di depan membrana bukofaringealis yang dikenal sebagai kantong Rathke, dan
(b) Perluasan ke arah bawah dari diensefalon, infundibulum.
Ketika mudigah mencapai usia sekitar 3 minggu, kantong Rathke tampak sebagai evaginasi rongga mulut dan kemudian tumbuh ke arah dorsal menuju infundibulum. Pada akhir bulan kedua,
kantong ini kehilangan hubungannya dengan rongga mulut dan kemudian berhubungan erat dengan infundibulum.
Selama perkembangan lebih lanjut, sel-sel di dinding anterior kantong Rathke bertambah banyak dengan pesat dan membentuk lobus anterior hipofisis atau adenohipofisis. Suatu perluasan kecil dari lobus ini, pars tuberalis, tumbuh di sepanjang tangkai infundibulum dan akhirnya mengelilinginya. Dinding posterior kantong Rathke berkembang menjadi pars intermedia yang pada manusia tampaknya tidak banyak berperan.
Infundibulum menghasilkan tangkai (stalk) dan pars nervosa, atau lobus posterior hipofisis (neurohipofisis). Bagian ini terdiri dari dari sel-sel neuroglia. Selain itu, bagian ini juga mengandung sejumlah serabut saraf dari daerah hipotalamus.
Telensefalon
Telensefalon, bagian paling rostral dari vesikel otak, terdiri dari dua penonjolan lateral, hemisferum serebri, dan suatu bagian di tengah, lamina terminalis. Rongga-rongga hemisfer, ventrikel lateral, berhubungan dengan lumen diensefalon melalui foramen interventrikulare Monro.
Hemisferum serebri muncul pada awal minggu kelima perkembangan sebagai evaginasi bilateral di dinding lateran proensefalon. Pada pertengahan bulan kedua bagian basal dari hemisfer (yaitu bagian yang pada awalnya membentuk perluasan talamus ke arah depan) mulai tumbuh dan menonjol ke dalam lumen ventrikel lateral dan ke dalam lumen ventrikel lateral dan ke dalam lantai foramen Monro. Pada potongan melintang, regio yang tampak bergaris-garis dan karenanya dikenal sebagai korpus striatum.
Di regio tempat dinding hemisfer melekat ke atap diensefalon, dinding tidak membentuk neuroblas dan tetap sangat tipis. Di sini dinding hemisfer terdiri dari suatu lapisan sel ependim yang ditutupi oleh mesenkimvaskular, dan bersama-sama keduanya membentuk pleksus koroideus. Pleksus koroideus seharusnya membentuk atap hemisfer, tetapi akibat pertumbuhan tak seimbang dari berbagai bagian hemisfer, pleksus ini menonjol ke dalam ventrikel lateral di sepanjang fisura koroidea. Tepat di atas fisura koroidea, dinding hemisfer menebal , membentuk hipokampus. Struktur yang fungsi primernya adalah penciuman ini, menonjol ke dalam ventrikel lateral.
Dengan perluasan selanjutnya, hemisfer menutupi aspek lateral diensefalon, mesensefalon, dan bagian sefalik metensefalon. Korpus striatum, karena merupakan bagian dari dinding hemisfer, juga meluas ke arah posterior dan dibagi menjadi dua bagian:
(a) Bagian dorsomedial, nukleus kaudatus, dan (b) Bagian ventrolateral, nukleus lentiformis
Pembagian ini dilakukan oleh akson-akson yang berjalan dari dan ke korteks hemisfer dan menembus massa nukleus korpus striatum. Berkas serabut yang terbentuk dikenal sebagai kapsula interna. Pada saat yang sama, dinding medial hemisfer dan dinding lateral diensefalon menyatu, serta nukleus kaudatus dan talamus menjadi terletak sangat dekat.
Pertumbuhan hemisferum serebri yang terus menerus ke arah anterior, dorsal, dan inferior, masing-masing menyebabkan terbentuknya lobus frontelis, temporalis, dan oksipitalis. Namun, seiring dengan melambatnya pertumbuhan di bagian yang terletak di atas korpus striatum, daerah antara lobus frontalis dan lobus temporalis menjadi tertekan dan dikenal sebagai insula. Bagian ini kemudian ditutupi oleh pertumbuhan lobus-lobus di sekitarnya dan pada saat lahir hampir tertutup seluruhnya. Selama tahap terakhir kehidupan janin, permukaan hemisferum serebri tumbuh sedemikian pesat sehingga muncul banyak tonjolan (girus) yang dipisahkan oleh fisura dan sulkus di permukaannya.
Korteks serebri berkembang dari palium yang memiliki dua regio:
(a) Paloepalium, atau arkipalium, tepat lateral dari korpus striatum, dan (b) Neopalium, antara hipokampus dan paleopalium
Di neopalium, gelombang-gelombang neuroblas bermigrasi ke posisi subpia dan kemudian berdiferensiasi menjadi neuron matur. Saat gelombang neuroblas berikutnya tiba, sel-sel tersebut bermigrasi melalui lapisan sel yang telah terbentuk sampai mencapai posisi subpia. Karena itu neuroblas yang terbentuk awal memperoleh posisi yang lebih dalam di korteks, sedangkan yang terbentuk belakangan memperoleh posisi lebih superfisial.
Saat lahir, korteks tampak berlapis-lapis akibat diferensiasi sel dalam lapisan-lapisan. Korteks motorik mengandung sejumlah besar sel piramidalis, dan area sensorik ditandai oleh sel granular.
Diferensiasi sistem olfaktorius bergantung pada interaksi epitel-mesenkim. Interaksi ini terjadi antara sel krista neuralis dan ektoderm prominensia frontonasalis untuk membentuk plakoda olfaktoria dan antara sel krista neuralis yang sama dengan lantai telensefalon untuk membentuk bulbus olfaktorius. Sel-sel di plakoda nasalis berdiferensiasi menjadi neuron sensorik primer epitel nasal yang akson-aksonnya tumbuh dan berkontak dengan neuron sekunder dalam pembentukan bulbus olfaktorius. Pada minggu ketujuh, kontak ini terbentuk sempurna. Seiring dengan berlanjutnya pertumbuhan otak, bulbus olfaktorius dan traktus olfaktorius neuron sekunder memanjang, dan bersama-sama keduanya membentuk nervus olfaktorius.
Pada orang dewasa, terdapat sejumlah berkas serabut, komisura yang menyilang garis tengah, menghubungkan paruh kanan dan kiri hemisfer. Berkas-berkas serabut terpenting menggunakan lamina terminalis. Berkas menyilang yang pertama kali tampak adalah komisura anterior. Berkas ini
terdiri dari serabut-serabut yang menghubungkan bulbus olfaktorius dan area otak terkait di satu hemisfer sisi lainnya.
Komisura kedua yang muncul adalah komisura hipokampus atau komisura forniks. Seabut-serabutnya berasal dari hipokampus dan berkumpul di lamina terminalis dekat dengan lempeng atap diensefalon. Dari sini serabut-serabut tersebut berlanjut, membentuk suatu sistem arkus tepat di luar fisura koroidea, ke korpus mamilare dan hipotalamus.
Komisura yang terpenting adalah korpus kalosum. Struktur ini muncul pada minggu ke-10 perkembangan dan menghubungkan area non-olfaktorius korteks serebri kiri dan kanan. Pada awalnya,, korpus kalosum membentuk suatu berkas kecil di lamina terminalis. Namun, akibat ekspansi neopalium yang terus-menerus, struktur ini meluas mula-mula ke anterior lalu ke posterior, membentuk arkus di atas atap tipis diensefalon.
Selain ketiga komisura yang terbentuk di lamina terminalis, tiga lainnya juga muncul. Dua di antaranya, komisura posterior dan komisura habenularum, tepat di sebelah bawah dan rostral tangkai kelenjar pineal. Yang ketiga, kiasma optikum, yang muncul di dinding rostral diensefalon, mengandung serabut-serabut dari separuh medial retina.
REGULASI MOLEKULAR PEMBENTUKAN OTAK
Pembentukan pola anteroposterior (kraniokaudal) sistem saraf pusat dimulai pada awal perkembangan, saat gastrulasi dan induksi saraf. Setelah lempeng saraf tebentuk, sinyal-sinyal untuk oemisahan otak menjadi regio otak depan, otak tengah, dan otak belakang datang dari gen-gen homeobox yang diekspresikanoleh notokord, lempeng prekordal, dan lempeng saraf. Otak belakang memilike delapan segmen, rombomber yang memiliki variasi pola ekspresi kelas Antennapedia gen homeobox, gen-gen HOX. Gen-gen ini diekspresikan dalam pola yang tumpang tindih (nested), dengan gen-gen di ujung paling 3’ dari kelompok yang memiliki batas lebih anterior dan gen-gen paralog memiliki ranah (domain) ekspresi yang identik. Gen-gen di ujung 3’ juga diekspresikan lebih awal daripada yang terletak di ujung 5’ sehingga terbentuk hubungan temporal dengan pola ekspresi. Karena itu, gen-gen ini memberikan nilai posisional di sepanjang sumbu antroposterior otak belakang, menentukan identitas rombomer, dan menentukan turunan-turunannya. Bagaimana regulasi ini terjadi masih belum diketahui, meskiipun retinoid (asam retinoat) berperan penting dalam mengatur ekspresi
HOX. Sebagai contoh, asam retinoat yang berlebihan menggeser ekspresi gen HOX ke arah anterior dan menyebabkan rombomer yang terletak lebih kranial berdiferensiasi menjadi tipe yang lebih kaudal. Defisiensi asam retinoat menyebabkan otak belakang mengecil. Juga terdapat respon diferensial terhadap asam retinoat oleh gen-gen HOX; gen-gen yang terletak di ujung 3’ kelompok yang lebih oeka daripada yang berada di ujung 5’.
Spesifikasi area otak depan dan otak tengah juga diatur oleh gen-gen yang mengandung suatu hoeodomain. Namun, gen-gen ini bukan dari kelas Antennapedia yang batas ekspresi paling anteriornya berhenti di rombomer 3’. Karena itu, diperkirakan terdapat gen-gen baru yang memiliki peran menentukan pola untuk regio otak ini yang secara evolusi merupakan “kepala baru”. Pada tahap lempeng saraf, LIM1 yang diekspresikan di lempeng prekordal, dan OTX2 yang diekspresikan di lempeng saraf, penting untuk menetukan area otak depan dan otak tengah, dengan LIM1 menunjang ekspresi OTX2. Gen-gen ini juga diekspresikan pada tahap-tahap awal gastrulasi, dan gen-gen ini membantu menentukan keseluruhan regio kranial epiblas. Setelah lipatan saraf dan arkus faring muncul, gen-gen homoebox lain, termasuk OTX1, EMX1, dan EMX2 diekspresikan dalam pola spesifik dan tumpan tindih (nested) di regio otak depan dan otak tengah serta untuk menspesifikasi identitas are-area ini. Jika batas-batas ini sudah dipastikan, dua pusat pengatur lain mucul: anterior neural ridge (ANR) di taut antara batas kranial lempeng saraf dan ektoderm non-saraf dan istmus antara otak tengah dan otak belakang. Di kedua lokasi, faktor pertumbuhan fibroblas 8 (FGF8) adalah molekul pembawa sinyal kunci, memicu ekspresi faktor transkripsi FOXG1. FOXG1 kemudian mengatur perkembangan telensefalon (hemisferum serebri) dan spesifikasi regional di dalam otak depan, termasuk telensefalon basal dan retina.
Di istmus di taut antara daerah otak tengah dan otak belakang, FGF8 diekspresi kan dalam bentuk cincin mengelilingi lokasi ini. FGF8 menginduksi ekspresi engrailed 1 dan 2 (EN1 dan EN2), dua gen yang mengandung homeobox yang diekspresikan secara gradien memancar ke arah anterior dan posterior dari istmus. EN1 mengatur perkembangan di seluruh ranah yang mengekspresikannya, termasuk otak tengah dorsal (tektum) dan oak belakang anterior (serebelum)., sedangkan EN2 hanya berperan dalam pembentukan serbelum. FGF8 juga menginduksi ekspresi WNT1 dalam bentuk pita melingkar di anterior dari regio ekspresi FGF8. WNT1 berinteraksi dengan EN1 dan EN2 untuk mengatur perkembangan regio ini, termasuk serebelum. Pada kenyataannya, WNT1 mungkin membantu spesifikasi awal daerah otak tengah karena gen ini diekspresikan di regio ini pada tahap lempeng saraf. FGF8 juga diekspresikan pada awal perkembangan di memsoderm yang terletak di bawah taut antara otak tengah dan belakang sehingga mungkin mengatur ekspresi WNT1 dan pembentukan pola awal di daerah ini. Konstriksi istmus terletak sedikit lebih posterior daripada taut otak tengah-belakang yang sebenarnya yang berada di batas kaudal ekspresi OTX2.
Pembentukan pola dorsoventral (mediolateral) juga terjadi di daerah otak depan dan otak tengah. Pembentukan pola ventral dikendalikan oleh SHH seperti di seluruh sistem saraf pusat sisanya. SHH yang disekresikan oleh lempeng prekordal, memicu ekspresi NKX2.1, suatu gen yang mengandung homeodomain yang mengatur perkembangan hipothalamus. Yang utama, sinyal SHH memerlukan pemecahan protein dan bagian terminal karboksil melaksanakan proses ini. Setelah protein SHH dibelah, kolesterol berikatan secara kovalen dengan terminal karboksil produk terminal amino. Bagian terminal amino mempertahankan semua sifat sinyal SHH, dan keterkaitannya dengan kolesterol membantu pendistribusiannya.
Pembentukan pola dorsal (lateral) tabung saraf dikendalikan oleh protein morfogenetik tulang 4 dan 7 (BMP4 dan BMP7) yang diekspresikan di ektoderm non-sarafdi samping lempeng saraf. Protein-protein ini memicu ekspresi MSX1 di garis tengah dan menekan ekspresi FOXG1. Setelah tabung saraf tertutup, terjadi ekspresi BMP2 dan 4 di lempeng atap dan protein-protein ini mengatur ekspresi faktor transkripsi LHX2 di korteks. Ekspresi ini kemudian memicu suatu jenjang gen yang mengatur pola regio ini.
Pola ekspresi gen-gen yang mengatur pola anterior-posterior (kraniokaudal) dan dorsoventral (mediolateral) otak bertumpang tindih dan berinteraksi di batas daerah-daerah tersebut. Selain itu, berbagai regio otak mampu berespon terhadap sinyal spesifik dan tidak terhadap yang lain. Sebagai contoh, hanya bagian kranial dari lempeng saraf yang mengekspresikan NKX2.1 sebagai respons terhadap SHH. Demikian juga, hanya lempeng saraf anterior menghasilkan FOXG1 sebagai respons terhadap FGF8; otak tengah mengekspresikan EN2 sebagai respons terhadap sinyal FGF8 yang sama. Karena itu, kompetensi untuk berespons juga membantu menentukan perbedaan regional.
2.2 Anatomi Talamus
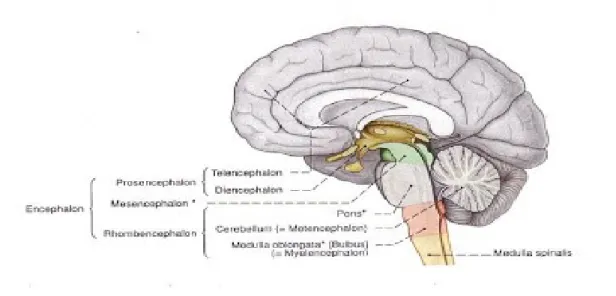
Diensefalon terletak di antara batang otak dan telensefalon. Diensefalon memiliki empat komponen:
Epithalamus, yang terdiri dari habenula dan nukleus habenularis, komissura habenularum, epifisis, dan komissura epithalamika (posterior)
Thalamus, kompleks neuron yang besar yang meliputi sekitar empat perlima volume diensefalon
Subthalamus, yang terutama terdiri dari nukleus subthalami (korpus Luysii) dan terletak di bawah thalamus dan di dorsolateral korpus mamilare
Hipothalamus, yang dipisahkan dari thalamus oleh sulkus hipothalamikus, dan mengandung berbagai kelompok neuron yang berbeda fungsi. Struktur ini merupakan pusat hierarki tertinggi (“ganglion kepala”) sistem saraf otonom pada masing-masing sisi, kolumna fornisis berjalan turun melalui dinding lateral hipothalamus dan berakhir di korpus mamilaris
Thalamus ditemukan pada kedua sisi ventrikel ketiga dan terdiri dari berbagai nukleus dengan fungsi berbeda. Struktur ini merupakan stasiun relay untuk sebagian besar jaras aferen yang naik ke korteks serebri. Beberapa jenis impuls (misalnya, impuls nosiseptif) mungkin telah diterima, diintegrasikan, dan diberikan warna afektif, dengan cara yang tidak tepat, di thalamus, tetapi pengalaman kesadaran sesungguhnya tidak dibentuk hingga impuls sensorik mencapai korteks serebri. Selain itu, thalamus memiliki hubungan yang luas dengan ganglia basalia batang otak, serebelum, dan area kortikal motorik serebri dan dengan demikian merupakan komponen utama sistem regulasi motorik.
Nukleus yang terpenting pada subthalamus adalah nukleus subthalamikus, yang fungsinya berkaitan erat dengan ganglia basalis.
Epithalamus terutama terdiri dari epifisis (glandula pinealis/korpus pineale) dan nukleus habenularis; berperan dalam regulasi irama sirkardian.
Bagian paling basal diensefalon adalah hipothalamus, yang mengkoordinasikan fungsi tubuh yang vital seperti respirasi, sirkulasi, keseimbangan cairan, suhu, dan asupan nutrisi dan dengan demikian berada pada hierarki teratas organ regulasi sitem saraf otonom. Struktur ini juga mempengaruhi aktivitas kelenjar endokrin melalui aksis hipothalamus – hipofisis.
Sistem saraf otonom berperan untuk suplai persarafan organ internal, pembuluh darah, kelenjar keringat, kelenjar saliva, dan kelenjar lakrimalis. Disebut “otonom” karena fungsinya sebagian besar tidak bergantung pada kesadaran, selain itu sistem saraf otonom juga (lebih jarang) disebut sistem saraf vegetatif. Lengan aferennya di perifer terdiri dari dua bagian yang secara anatomi dan fungsional berbeda, sistem saraf simpatis dan parasimpatis. Lengan aferennya tidak terbagi dengan cara seperti ini.
Karena fungsi diensefalon yang beragam, lesi diensefalon dapat memiliki efek yang sangat bervariasi, tergantung pada lokasi dan luasnya lesi. Lesi thalamus menyebabkan hemiparesis dan defisit hemisensorik, gangguan pergerakan, gangguan kesadaran, dan sindrom nyeri, sedangkan lesi hipothalamus mengganggu berbagai fungsi vital secara tunggal atau kombinasi, dan menyebabkan disfungsi endokrin.
Posisi diensefalon tepat di bagian oral mesensefalon; diensefalon tidak berlanjut di sepanjang aksis batang otak, tetapi menekuk ke arah rostral, sehingga terletak di dekat aksis longitudinal serebri. Diensefalon terletak di tengah otak, di bagisn ventral dan kaudal lobus frontalis, dan terletak di dekat bagian terbawah ventrikel ketiga dari kedua sisi.
Thalamus membentuk bagian teratas dinding ventrikel ketiga, dan hipothalamus bagian terbawahnya. Di dorsal, diensefalon berdekatan dengan korpus kalosum, ventrikel lateral,dan hemisfer serebri. Atap ventrikel ketiga dibentuk oleh tela khoroidea yang tipis dan pleksus khoroideus yang melekat. Perluasan diensefalon ke arah rostral dibatasi oleh lamina terminalis dan komisura anterior, perluasan ke arah kaudalnya oleh komisura posterior, komisura habenularum, dan korpus pinealis (epifisis). Foramen interventrikularis Monro, yang menghubungkan ventrikel lateral dengan ventrikel ketiga, terdapat di kedua sisi bagian anterior hingga bagian rostral thalamus, tepat di bawah krus fornicis. Bagian basal diensefalon merupakan satu-satunya bagian yang terlihat dari luar: dapat terlihat pada permukaan bawah otak di antara khiasma optikum, traktus optikus, dan pedunkulus serebri. Struktur diensefalon yang terlihat di area ini adalah korpus mamilare dan tuber sinereum, bersamaan dengan infundibulumnya (tangkai hipofisis), yang mengarah ke bawak menuju kelenjar hipofisis.
Kedua separuh bagian hipothalamus yang berhadapan satu sama lain menyeberangi ventrikel ketiga berhubungan pada 70-80% kasus dengan adhesi interthalamik (massa intermedia), yang bukan merupakan jaras serabut tetapi merupakan adhesi sekunder substansia grisea yang datang dari masing-masing sisi. Di laterar, diensefalon berbatasan dengan kapsula interna.
Globus palidus secara embriologis merupakan bagian diensefalon, meskipun dipisahkan oleh kapsula interna dan dengan demikian terletak di ganglia basalia.
2.2.1 Nuklei Talamus
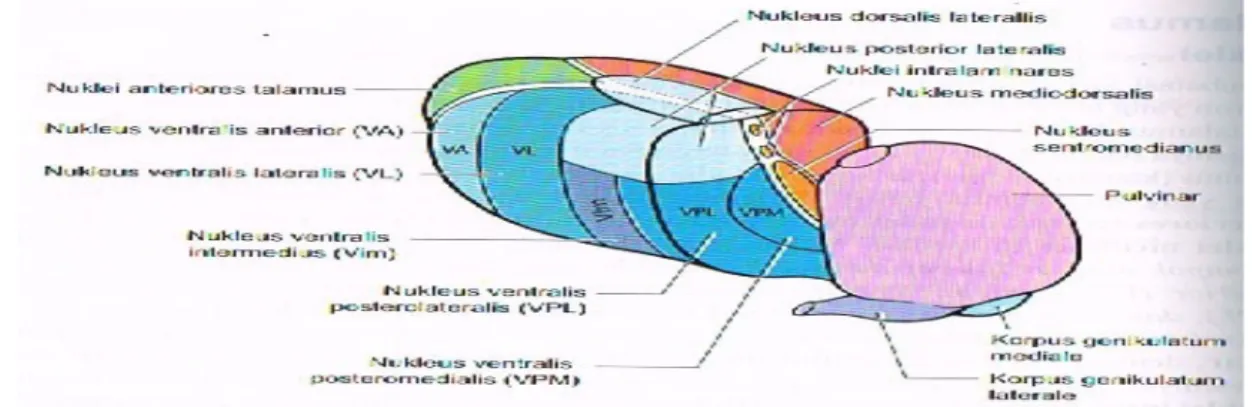
Membatasi sisi ventrikel ketiga, pada masing-masing sisi otak, terdapat kompleks neuron yang besar dan berbentuk oval dengan diameter sekitar 3 x 1.5 cm. Kompleks ini, thalamus, bukan merupakan kumpulan sel yang sama, tetapi yang berbeda, tetapi yang berbeda, dengan fungsinya dan hubungan eferen dan aferennya masing-masing. Setiap separuh bagian talamus (kanan dan kiri)
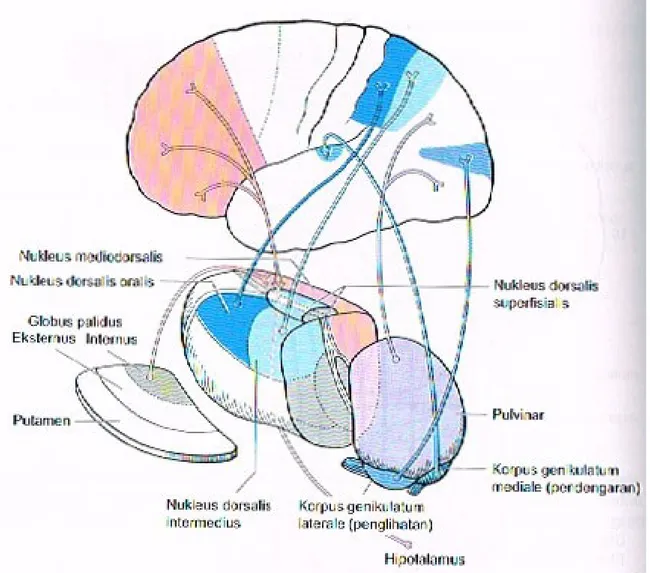
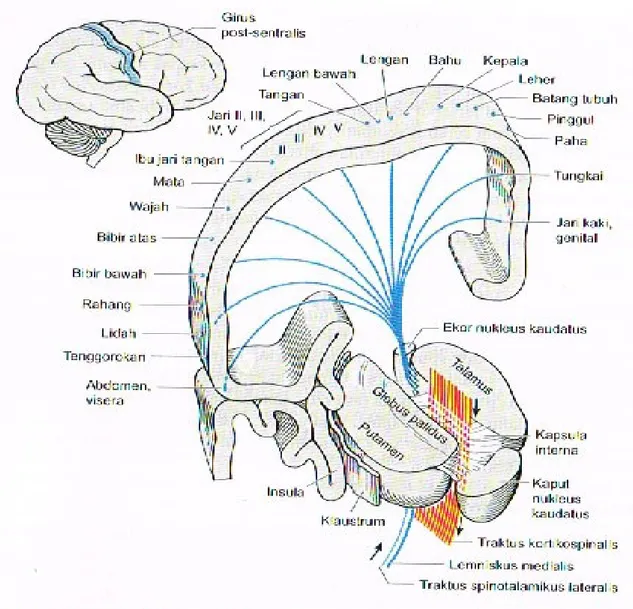
terbagi menjadi tiga regio utama oleh lapisan tipis substansia alba yang membentuk huruf Y (lamina medularis interna). Nuklei anteriores terletak di sudut huruf Y tersebut, nuklei ventrolaterales di lateral, dan nuklei mediales di medial. Nuklei ventrolaterales kemudian terbagi lagi menjadi kelompok nuklear lateral dan medial. Nukleus ventralis meliputi nukleus ventralis anterior (VA), dan nukleus ventralis lateralis (VL), nukleus ventralis posterolateralis (VPL), dan nukleus posteromedialis (VPM). Nukleus lateralis meliputi nukleus lateralis dorsalis dan nukleus lateralis posterior. Lebih ke kaudal, ditemukan pulvinar, dengan korpus genikulatum laterale dan mediale yang melekat pada sisi bawahnya. Ada beberapa kelompok kecil neuron di dalam lamina medularis interna (nuklei interlaminares), serta sebuah kompleks sel yang lebih besar, yang terletak di sentral, nukleus sentromedianus (atau centre median). Di lateral, lamina medularis eksterna memisahkan thalamus dari kapsula interna; nukleus retikularis thalami merupakan lapisan sel tipis yang terletak bedekatan dengan lamina medularis eksterna. Tiga kelompok nuklear utama (anterior, ventrolateral dan medial) secara sitologis dan fungsional terbagi lagi menjadi sekitar 120 nukleus yang lebih kecil.
Gambar 6. Nuklei Talamus
Talamus merupakan stasiun relay utama terakhir untuk semua impuls asendens (kecuali impuls olfaktorius) sebelum melanjutkan ke korteks, melalui serabut talamokortikalis. Berbagai jaras aferen pada setiap nukleus thalami memiliki terminasi yang berbeda, yang kemudian berproyeksi ke area kortikal yang sesuai. Seperti medula spinalis dan batang otak (misalnya, lemniskus medialis), nuklei thalami dan proyeksi thalamokortikalis mempertahankan organisasi somatotropik titik ke titik yang tepat.
Nuklei thalami yang menerima input dari area tubuh perifer tertentu dan menghantarkan impuls ke area kortikal yang bersesuaian (lapangan proyektif primer) disebut nuklei thalami spesifik (atau nuklei thalami primer). Nuklei thalami yang berproyeksi ke arah kortikal asosiasi unimodal dan multimodal (nuklei thalami sekunder dan tersier) juga termasuk nuklei spesifik. Sehingga perbedaan sifat nuklei spesifik adalah proyeksi langsung ke korteks serebri.
Sebaliknya, nuklei thalami nonspesifik meneri input aferennya dari berbagai organ sensorik yang berbagai, biasanya setelah sinaps penghubung di formasio retikularis dan / atau salah satu nuklei thalami primer. Serabut ini hanya berproyeksi secara tidak langsung ke korteks serebri (misalnya, melalui ganglia basalia), termasuk area asosiasi.
2.2.2 Nuklei Thalami Spesifik dan Hubungan-hubungannya Nuklei dengan Hubungan ke Area Kortikal Primer
(A) Nukleus ventralis posterolateralis (VPL) dan nukleus ventralis posteromedialis (VPM)
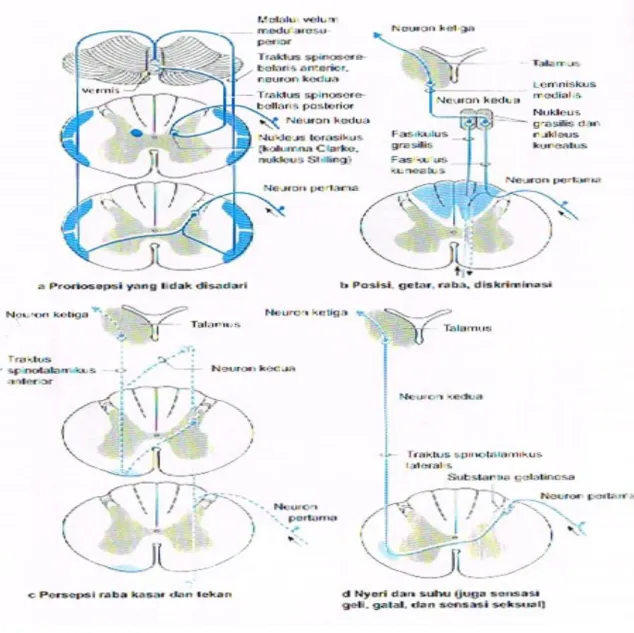
Semua serabut somatosensorik berjalan naik di dalam lemniskus medialis, traktus spinothalamikus, traktus trigeminothalamikus, dan berakhir di stasiun relay di nuklei ventrioposteriores thalami. Nukleus ventralis posterolateralis merupakan stasiun relay untuk lemniskus medialis, sedangkan untuk nukleus ventralis posteromedialis merupakan sasiun relay untuk aferen trigeminus. Nuklei ini, kemudian memproyeksikan serabut ke area korteks somatosensorik tertentu. Selain itu, serabut gustatorik dari nukleus traktus solitarius berakhir di ujung medial nukleus ventralis posteromedialis, yang kemudian berproyeksi ke regio postsentralis yang menutupi insula.
(B) Korpus genikulatum lateralis dan medialis
Korpus genikulatum medialis dan lateralis juga merupakan salah satu di antara nuklei thalami spesifik. Traktus optikus berakhir di korpus genikulatum lateralis, yang menghantarkan impuls visual secara retinotropik, melalui radiasio optika, ke korteks visual (area 17). Impuls auditorik dibawa di lemniskus lateralis ke korpus genikulatum medialis dan dihantarkan secara tonotropik, melalui radiasio akustika, ke korteks auditorik (giri temporales transversi Heschl, area41) di lobus temporalis.
(C) Nukleus ventralis oral dan nukleus ventralis anterior
Nukleus ventralis oralis posterior (V.o.p, bagian nukleus ventralis lateralis) menerima input dari nukleus dentatus dan nukleus ruber melalui traktus dentikulothalamikus dan berproyeksi ke korteks motorik (area 4), sedangkan nukleus ventralis oralis anterior (V.o.a) dan nukleus ventralis anterior (VA) yang keduanya juga merupakan anggota kelompok nukleus ventralis, yang menerima input dari globus palidus dan berproyeksi ke korteks premotorik.
Nukleus anterior, nukleus medialis, dan pulvinar merupakan nuklei thalami sekunder dan tersier, yaitu nuklei thalami spesifik yang berproyeksi ke area korteks asosiasi unimodal dan multimodal. Nuklei tersebut sebagian besar menerima inputnya tidak secara langsung dari perifer, tetapi setelah melewati relay sinaptik, yang biasanya terletak di salah satu nuklei thalami primer.
Nukleus anterior secara timbal balik berhubungan dengan korpus mamilare dan forniks melalui traktus mamilothalamikus (Vicq d’Azyr); memiliki hubungan titik-ke-titik dua arah dengan girus cinguli (area 24) dan dengan demikan merupakan bagian yang terintegrasi dengan sistem limbik. Nukleus medialis thalami memiliki hubungan titik-ke-titik dua arah dengan area asosiasi lobus frontalis dan regio premotoris. Nukleus ini menerima input aferen dari nuklei thalami lain (nukleus ventralis dan intralaminaris), dan dari hipothalamus, nukles mesensefali, dan globus palidus. Destruksi nukleus medialis oleh sebuah tumor atau proses lainnya menyebabkan sindrom lobus frontalis dengan perubahan kepribadian (hilangnya representasi diri), seperti yang ditemukan setelah leukotomi frontal. Impuls viseral yang mencapai nukleus ini melalui hipothalamus mempengaruhi keadaan afektif individu, menyebabkan rasa kesejahteraan atau kesedihan, mood baik atau buruk, dan sebagainya.
Pulvinar memiliki hubungan titik-ke-titik secara timbal balik dengan area asosiasi lobus parietalis dan lobus oksipitalis. Area asosiasi ini dikelilingi oleh korteks somatosensorik, visual, dan auditorik primer dan dengan demikian kemungkinan berperan penting pada pengumpulan berbagai jenis informasi sensorik yang datang. Pulvinar menerima input neural dari nuklei thalami lain, terutama nuklei intralaminares.
Nukleus lateralis dorsalis dan nukleus lateralis posterior tidak menerima inout neural apapun dari luar thalamus dan hanya berhubungan dengan nuklei thalami lain. Dengan demikian struktur ini disebut nuklei integratif.
2.2.3 Nuklei Thalami Nonspesifik dan Hubungan-hubungannya
Nuklei intralaminares adalah komponen sistem proyeksi thalami nonspesifik yang terpenting. Nuklei ini terletak di dalam lamina medularis interna, dan yang terbesar di antara nuklei tersebut adalah nukleus sentromedianus. Kompleks sel ini menerima input aferennya melalui serabut asendens dari formasio retikularis batang otak dan nukleus emboliformis serebeli, serta globus palidus medialis dan nuklei thalami lainnya. Serabut-serabut ini tidak berproyeksi ke korteks serebri tetapi menuju ke nukleus kaudatus, putamen, dan globus palidus. Serabut tersebut kemungkinan juga mengirimkan impuls eferen secara difus ke seluruh nuklei thalami, yang kemudian, berproyeksi ke are sekunder korteks serebri yang luas. Nukleus sentromedianus merupakan komponen yang penting pada
kompleks sel intralaminaris, yang membentuk bagian thalamik ascending reticular activating system (ARAS atau sistem kewaspadaan). Bagian lain sistem kewaspadaan ini kemungkinan melibatkan subthalamus dan hipothalamus.
Gambar 7. Hubungan Aferen dan Eferen Kelompok Nuklear Medial (Merah), Dorsal (Ungu/Biru), dan Lateral (Biru)
2.3 Neurofisiologi Talamus
Fungsi thalamus sangat rumit karena banyaknya jumlah nukleus yang dimilikinya dan hubungan aferen dan eferennya yang sangat bervariasi
Pertama-tama, thalamus merupakan titik pertemuan subkortikal terbesar untuk semua impuls sensorik propioseptif dan eksteroseptif
Selain itu, struktur ini merupakan stasiun relay untuk semua impuls yang timbul di reseptor sensorik kutaneus dan reseptor sensorik viseral, impuls auditorik dan visual, dan impuls dari hipothalamus, serebelum, dan formasio retikularis batang otak; semua impuls diproses di
thalamus sebelum ditransmisikan ke struktur lainnya. Thalamus mengirimkan komponen eferen ke striatum, tetapi sebagian besar outputnya berjalan ke korteks serebri. Semua impuls sensorik (semua impuls olfaktori) harus berjalan melewati thalamus sebelum dapat disadari. Dengan demikian, thalamus dulu disebut “pintu gerbang kesadaran”, meskipun persepsi bau yang disadari menunjukkan bahwa konsep ini tidak sempurna dan kemungkinan menyesatkan.
Namun, thalamus bukan serta merta sebagai stasiun relay, tetapi pusat integrasi dan koordinasi yang penting; keempat impuls aferen berbagai modalitas, dari regio tubuh yang berbeda, diintegrasikan dan diberikan pewarnaan afektif. Substrat neural fenomena dasar tertentu seperti nyeri, rasa tidak senang, dan rasa nyaman telah terdapat di thalamus sebelum ditransmisikan ke atas menuju korteks.
Melalui hubungan timbal-baliknya (lengkung umpan-balik) dengan korteks motorik, beberapa di antaranya melewati ganglia basalia dan serebelum, thalamus memodulasi fungsi motorik.
Beberapa nuklei thalami juga merupakan komponen ascending reticular activating system (ARAS), sistem kewaspadaan spesifik yang berasal dari otak. Impuls pengaktivasi dari ARAS dihantarkan oleh nuklei thalami tertentu (nukleus ventralis anterior, nukleus intralaminaris, terutama nukleus sentromedian, nukleus retikularis) ke seluruh neokorteks. ARAS yang intak penting untuk kesadaran yang normal.