POLIMORFISME GEN PENYANDI KARAKTER
OBESITAS (MC4R/RESEPTOR MELANOKORTIN 4)
PADA MONYET EKOR PANJANG (
Macaca
fascicularis
) ASAL BALI, JAWA TIMUR DAN
SUMATERA
I GUSTI AGUNG ARTA PUTRA
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
SURAT PERNYATAAN
Dengan ini saya menyatakan bahwa disertasi berjudul Polimorfisme Gen
Penyandi Karakter Obesitas (MC4R/Reseptor Melanokortin 4) pada Monyet Ekor Panjang (Macaca Fascicularis) Asal Bali, Jawa Timur dan Sumatera
adalah karya saya sendiri dengan arahan komisi pembimbing dan belum diajukan dalam bentuk apa pun pada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam daftar pustaka di bagian akhir disertasi ini.
Bogor, 30 November 2009
ABSTRACT
I GUSTI AGUNG ARTA PUTRA. Polymorphism of Coding Gene of Obesity
Character (MC4R/Melanocortin 4 Receptor) in Cynomolgus Macaque Originated from Bali, East Java and Sumatera. Under the direction of DONDIN SAJUTHI,
DEDY DURYADI SOLIHIN, and R.R. DYAH PERWITASARI
Melanocortin 4 receptor (MC4R) is one of G protein-coupled receptors that plays an important role in regulation of energy homeostasis. MC4R mutations constitute the most common cause of human obesity. Since cynomolgus macaque is one of animal models, the study on adult male macaque was conducted in order to investigate the polymorphism of MC4R gene, phenotype of obesity and the association of MC4R mutation with obesity phenotype. Fifty six adult male macaques from Bali (Ubud and Uluwatu), East Java (Alas Purwo and Baluran), and Sumatera island (Palembang) were used in this research. The animals had been anaesthetized using ketamine (10 mg/kg of body weight) and xylazine (2 mg/kg of body weight) before phenotype data and blood samples were collected. The phenotype parameters measured in this study were body weight, crown rump length, body mass index; circumference of triceps, chest, waist, and thigh; skin fold thickness of abdomen, thigh, back, triceps, and dorsal neck. Blood samples were used as source of DNA. To determine MC4R polymorphism, coding region of this gene was amplified and sequenced. The results showed that there were 20 polymorphism sites identified and 13 of them are non-synonymous. Among the non-synonymous mutations, five mutations were only found in obese cynomolgus macaques; two mutations were found in both obese and obese macaques; and six mutations were only found in non-obese macaques. In addition, there were significant difference (P<0.05) of phenotype parameters among the study sites.
RINGKASAN
I GUSTI AGUNG ARTA PUTRA. Polimorfisme Gen Penyandi Karakter Obesitas
(MC4R/Reseptor Melanokortin 4) pada Monyet Ekor Panjang (Macaca
fascicularis) Asal Bali, Jawa Timur dan Sumatera. Dibimbing oleh DONDIN
SAJUTHI, DEDY DURYADI SOLIHIN, dan R.R DYAH PERWITASARI.
Obesitas pada manusia merupakan predisposisi utama untuk terjadinya penyakit terutama sindroma metabolik, diabetes melitus tipe 2 dan hipertensi. Saat ini, obesitas dianggap penyakit kompleks multifaktor yang disebabkan oleh faktor genetik dan lingkungan. Gen yang berkaitan dengan kejadian obesitas pada manusia telah diinventarisasi sebanyak 127 gen kandidat. Dari 127 gen tersebut, sebelas di antaranya bersifat monogenik sehingga apabila gen tersebut bermutasi akan menyebabkan obesitas. Dari sebelas gen yang bersifat monogenik, gen reseptor melanokortin 4 (MC4R) diwariskan secara dominan.
Mutasi MC4R merupakan hal yang paling umum menyebabkan obesitas pada manusia. MC4R adalah salah satu reseptor berprotein-G yang berperan penting dalam pengaturan homeostasis energi. Reseptor ini banyak ditemukan di dalam otak dan berfungsi dalam penghantaran isyarat kekenyangan. Bila reseptor ini tidak berfungsi akan menyebabkan nafsu makan berlebihan dan dapat menyebabkan terjadinya obesitas.
Monyet ekor panjang merupakan hewan model untuk penelitian yang hasilnya dimanfaatkan untuk kepentingan manusia. Monyet ekor panjang menunjukkan gejala obesitas yang mirip dengan kejadian obesitas pada manusia. Kejadian obesitas banyak dijumpai pada monyet ekor panjang jantan yang ada di kawasan wisata di Bali. Kejadian obesitas ini merupakan salah satu alasan untuk melakukan penelitian tentang polimorfisme gen MC4R pada monyet ekor panjang. Penelitian ini bertujuan untuk 1) mengidentifikasi variasi gen MC4R pada monyet ekor panjang asal Bali, Jawa Timur dan Sumatera, 2) mengkaji variasi fenotipe obesitas pada monyet ekor panjang asal Bali, Jawa Timur dan Sumatera, 3) mendapatkan asosiasi mutasi gen MC4R dengan obesitas pada monyet ekor panjang.
Penelitian ini menggunakan monyet ekor panjang jantan dewasa sebanyak 56 ekor yang berasal dari Bali (Ubud dan Uluwatu), Jawa Timur (Alas Purwo dan Baluran) dan Sumatera (Palembang). Monyet asal Bali dan Jawa Timur ditangkap langsung di lokasi sedangkan monyet asal Sumatera diperoleh di penangkaran PSSP-IPB. Monyet dibius dengan ketamine (10 mg/kg bobot badan) dan xylazine (2 mg/kg bobot badan) sebelum dilakukan pengukuran data fenotipe dan pengambilan darah. Fenotipe yang diukur adalah bobot badan, tinggi duduk, indeks massa tubuh (IMT); lingkar lengan atas, dada, pinggang dan paha; serta tebal lipatan kulit di daerah perut, paha, punggung, trisep, dan leher bagian dorsal. IMT merupakan rasio bobot badan (kg) per tinggi duduk (m2). Ekstraksi DNA (deoxyribonucleic acid) total dari darah menggunakan QIAamp DNA Blood Kits. Amplifikasi daerah penyandi gen MC4R dilakukan dengan
primer forward (5’–AATAACTGAGACGACTCCCTGAC–3’) dan reverse (5’–
CAGAAGTAC AATATTCAGGTAGGG–3’)berdasarkan Yeoet al. (1998) dengan
teknik polymerase chain reaction (PCR). Pengurutan produk PCR menggunakan
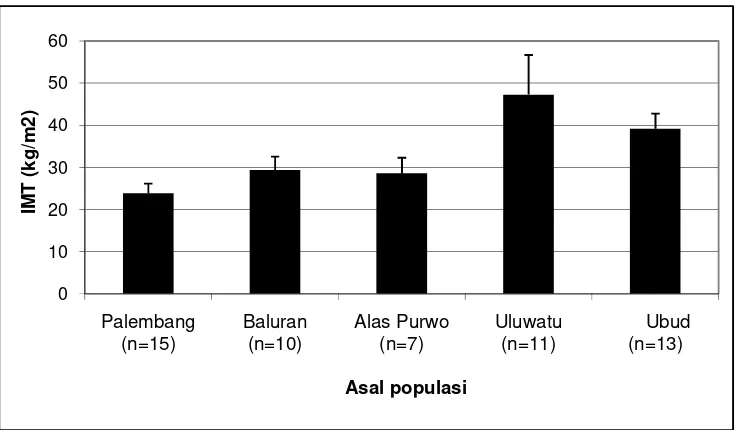
Berdasarkan data fenotipe, seluruh (100%) monyet ekor panjang jantan asal Bali menunjukkan obesitas (IMT>30kg/m2). Persentase obesitas monyet asal Baluran dan Alas Purwo masing-masing adalah 30% (3/10) dan 28,6% (2/7). Di lain pihak, monyet asal Palembang tidak ada yang menunjukkan obesitas. IMT yang tinggi disebabkan oleh bobot badan yang tinggi. Di Uluwatu, ditemukan monyet dengan bobot badan 16 dan 17 kg. Monyet yang mengalami obesitas (IMT>30kg/m2), sebagian besar disertai dengan lingkar pinggang yang lebih besar dari 39 cm dan tebal lipatan kulit perut lebih besar daripada 5 mm. Oleh karena itu, lingkar pinggang dan tebal lipatan kulit perut perlu dipertimbangkan dalam penentuan obesitas pada monyet ekor panjang disamping IMT.
Hasil analisis urutan gen MC4R (48 urutan) menunjukkan adanya 20 situs polimorfik yang terdiri dari tujuh situs bersifat sinonim dan 13 situs bersifat nonsinonim. Situs polimorfik ditemukan paling banyak pada monyet asal Alas Purwo sedangkan yang paling sedikit ditemukan pada monyet asal Uluwatu. Dari 13 situs yang bersifat nonsinonim, lima situs ditemukan hanya pada monyet yang mengalami obesitas. Kelima mutasi tersebut adalah Asn3His, Leu250Pro, Leu300Arg, Leu 328Trp, dan Ser329Ala. Dua situs mutasi ditemukan baik pada monyet gemuk maupun monyet tidak gemuk. dan enam situs mutasi ditemukan hanya pada monyet tidak gemuk.
Kejadian substitusi nukleotida yang bersifat transversi lebih besar daripada substitusi yang bersifat transisi. Hal ini diduga ada kaitannya dengan komposisi basa nukleotida dari gen MC4R. Kandungan adenina (A) dan timina (T) adalah lebih besar daripada kandungan guanina (G) dan sitosina (C). Kemungkinan substitusi transversi akan meningkat sesuai dengan peningkatan kandungan basa nukleotida A dan T.
Analisis filogeni menunjukkan bahwa keseluruhan sampel berada dalam satu kelompok dan terpisah dari simpanse. Filogeni yang dibuat dari gen MC4R ternyata tidak dapat membedakan monyet yang berasal dari daerah yang berbeda. Bila dibandingkan dengan spesies lain, monyet ekor panjang satu
kelompok dengan monyet dalam genus yang sama (Macaca mulatta).
Hasil penelitian ini juga menunjukkan bahwa mutasi nonsinonim MC4R terjadi baik pada monyet gemuk maupun yang tidak gemuk. Dari 48 sampel urutan, sebanyak 26 ekor monyet ekor panjang tergolong gemuk dan 22 ekor tidak gemuk. Sebanyak 11,5% monyet gemuk dan 22,7% monyet tidak gemuk merupakan pembawa mutan MC4R. Monyet gemuk asal Uluwatu dan Ubud, membawa mutasi sinonim dan nonsinonim pada gen MC4R, tetapi obesitas mungkin juga hasil gen non-MC4R atau perubahan pada bagian promoter genMC4R.
Kata kunci: monyet ekor panjang (Macaca fascicularis), obesitas, reseptor
©Hak Cipta milik IPB, tahun 2009 Hak Cipta dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan yang wajar IPB
POLIMORFISME GEN PENYANDI KARAKTER
OBESITAS (MC4R/RESEPTOR MELANOKORTIN 4)
PADA MONYET EKOR PANJANG (
Macaca
fascicularis
) ASAL BALI, JAWA TIMUR DAN
SUMATERA
I GUSTI AGUNG ARTA PUTRA
Disertasi
sebagai salah satu syarat untuk memperoleh gelar Doktor pada
Mayor Primatologi
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Penguji pada Ujian Tertutup : Prof. Dr. Ir. Hadi S. Alikodra, MS. Prof. Dr. Ir. Cece Sumantri, M.AgrSc.
PRAKATA
Puji syukur penulis panjatkan ke hadirat Tuhan Yang Maha Esa, karena berkat rahmat-Nya disertasi yang berjudul ” Polimorfisme Gen Penyandi Karakter
Obesitas (MC4R/ Reseptor Melanokortin 4) pada Monyet Ekor Panjang (Macaca
fascicularis) Asal Bali, Jawa Timur dan Sumatera” dapat diselesaikan.
Selama menyelesaikan penelitian dan penulisan disertasi, penulis mendapatkan bantuan dari berbagai pihak. Pada kesempatan ini penulis menyampaikan terimakasih yang sebesar-besarnya kepada Prof. drh. Dondin Sajuthi, MST., Ph.D, Dr. Ir. Dedy Duryadi Solihin, DEA, dan Dr. Ir. R.R. Dyah Perwitasari, M.Sc selaku komisi pembimbing yang telah meluangkan waktu untuk membimbing baik selama penelitian maupun penulisan disertasi ini. Ucapan terimakasih juga penulis tujukan kepada Dekan Fapet dan Rektor UNUD yang telah memberikan izin tugas belajar untuk mengikuti program doktor (S3) di
Program Studi/Mayor Primatologi (PRM), Sekolah Pascasarjana IPB.
Terimakasih juga kepada Rektor IPB, Dekan Pascasarjana IPB, Ketua Mayor PRM serta seluruh staf pengajar dan administrasi PRM. Terimakasih juga kepada tim manajemen Beasiswa Program Pascasarjana (BPPS) Dirjen Dikti Depdiknas yang memberikan dana pendidikan S3. Terimakasih juga kepada team hibah doktor IPB, Yayasan Damandiri dan APPERI atas batuannya.
Penulis juga menyampaikan terimakasih kepada drh. Nengah Budiarsa (APPERI), Prof. Dr. Ir. Sri Supraptini Mansjoer, Prof. Dr. IDK Harya Putra, Prof. Dr. IB. Sudana, Prof. Dr. Ir. Hadi S. Alikodra, MS., Prof. Dr. Ir. Cece Sumantri, M.AgrSc., Prof. Dr. Ir. Muladno, MSA., Prof (R). Dr. Ir. M. Bismark, MS., Prof. Dr. Agustin Fuentes, Kelly Lane, MSc. Dr. drh. I Nengah Wandia, drh. Aida LT Rompis, Dr. drh. I Kt. Suatha, drh. IGd. Soma MKes, drh. I Pt. Yasa, drh. Sri Kayati Widyastuti MSi, drh I Putu Gede Yudi Arjentinia, Dr. drh. W. Batan MSi dan staf pengelola Jurnal Veteriner, Dr. drh. N. Suarsana, MSi., Muharam Saepulloh SSi., MSc., dr. Anwar Wardi SpS, M Nasir Spt., MSi., drh. Susana Widjaya, Keni Sultan SPt., MSi., Ria Oktarina SPt., MSi., Nurjayanti Spt., Yana dan teman lainnya yang tidak bisa disebutkan satu persatu.
staf, di PSSP-IPB, Novita Anggraeni SPt., dan Ramdan di Laboratorium Biologi dan Reproduksi PSSP-IPB, Dr. Soaloon Sinaga, Dr. Fitma, Dr. Roza Elvyra, Dr. Agus Nuryanto, Dr. Agus Nasri, Dr. Jakaria, Pak Heri, Pak Pras, teman-teman Punhawacana Bali dan adik-adik di Asrama Brahmacarya Cikuray 10 Bogor.
RIWAYAT HIDUP
Penulis dilahirkan di Bindu (Bali) , 30 November 1962. Penulis merupakan anak ke empat dari delapan bersaudara dari pasangan I G A N Kembar dan I G A K Rai. Penulis menikah dengan AAA Alit Adiari, SH dan dikaruniai dua putri yaitu A A Angga Primantari (16 tahun) dan A A Anisca Primadwiyani (13 tahun).
DAFTAR ISI
Halaman
DAFTAR TABEL ... xiii
DAFTAR GAMBAR ... xiv
DAFTAR LAMPIRAN ... xv
PENDAHULUAN ... 1
Latar Belakang ... 1
Tujuan Penelitian... 4
Hipotesis ... 4
Manfaat Penelitian ... 5
Kerangka Pemikiran ... 5
TINJAUAN PUSTAKA ... 7
Monyet Ekor Panjang ... 7
Obesitas ... 9
Melanokortin ... 12
Reseptor Melanokortin 4 ... 14
Perubahan MC4R dan Akibatnya ... 15
Mutasi Gen MC4R ... 16
Klasifikasi Mutasi MC4R... 18
Mekanisme Molekuler Homeostasis Energi ... 20
Teknik PCR ... 22
MATERI DAN METODE... 24
Waktu dan Lokasi Penelitian... 24
Materi... 24
Monyet Ekor Panjang ... 24
Bahan dan Alat Pengambilan Sampel Darah dan Data Fenotipe ... 25 Bahan dan Alat Isolasi DNA... 25
Bahan dan Alat Analisis PCR... 26
Bahan dan Alat Elektroforesis... 26
Metode... 26
Pengambilan Darah dan Data Fenotipe... 26
Ekstraksi DNA dari Darah... 27
Amplifikasi Gen MC4R... 28
Pengurutan Produk PCR... 30
Analisis Data... 30
Analisis Data Fenotipe... 30
Analisis Data Molekuler... 32
HASIL DAN PEMBAHASAN... 34
Keragaman Fenotipe Obesitas Monyet Ekor Panjang... 34
Indeks Massa Tubuh, Bobot Badan dan Tinggi Duduk... 34
Lingkar Pinggang, Dada, Paha dan Lengan Atas... 38
Tebal Lipatan Kulit Perut, Trisep, Paha, Punggung dan Leher... 40 Analisis Korelasi Parameter Fenotipe... 43
Analisis Komponen Utama... 44
Obesitas dan Struktur Sosial... 46
Karakteristik Genetik Gen MC4RMonyet Ekor Panjang... 47
Amplifikasi Gen MC4R... 47
Keragaman Intra Populasi... 48
Keragaman Antar Populasi... 48
Komposisi Nukleotida dan Asam Amino... 49
Analisis Filogeni... 50
Perbandingan Gen MC4R Monyet Ekor Panjangdengan Organisme Lainnya... 55 Polimorfisme Gen MC4R pada Monyet Ekor Panjang... 55
Asosiasi Mutasi MC4R dengan Fenotipe Obesitas pada Monyet Ekor Panjang ... ... 57 PEMBAHASAN UMUM... 60
Fenotipe Obesitas... 60
Polimorfisme MC4R... 62
Hubungan Kegiatan Konservasi dengan Monyet Gemuk... 66
SIMPULAN DAN SARAN... 67
Simpulan... 67
Saran... 67
DAFTAR PUSTAKA... 68
DAFTAR TABEL
Halaman
1 Jumlah monyet ekor panjang jantan yang digunakan dalam
penelitian ini... 24
2 Indeks massa tubuh, bobot badan, dan tinggi duduk... 34
3 Lingkar pinggang, dada, paha dan lengan atas ... 38
4 Tebal lipatan kulit perut, trisep, paha, punggung dan leher ... 41
5 Nilai korelasi Pearson dan nilai P antar parameter fenotipe... 43
6 Rata-rata jumlah nukleotida dan asam amino yang berbeda pada gen MC4R monyet ekor panjangdalam masing-masing populasi ... 48
7 Matriks perbedaan basa nukleotida gen MC4R monyet ekor panjang antar populasi... 49
8 Matriks perbedaan asam amino gen MC4R monyet ekor panjang antar populasi... 49
9 Rata-rata komposisi nukleotida (%) pada gen MC4R monyet ekor panjang... 49
10 Rata-rata komposisi asam amino (%) gen MC4R pada monyet ekor panjang ... 50
11 Polimorfirme gen MC4R pada monyet ekor panjang ... 56
12 Jumlah substitusi transisi dan transversi pada gen MC4R monyet ekor panjang... 57
13 Mutasi nonsinonim daerah penyandi gen MC4R monyet ekor panjang... 57
DAFTAR GAMBAR
Halaman
1 Kerangka pemikiran... 6
2 Obesitas pada monyet ekor panjang ... 10
3 Sinyal protein G yang mengatur produksi cAMP ... 13
4 Organisasi gen MC4R ... 14
5 Kejadian mutasi MC4R pada manusia ... 17
6 Klasifikasi mutasi MC4R pada manusia ... 19
7 Hormon yang mengatur nafsu makan ... 21
8 Peta lokasi asal monyet ekor panjang yang digunakan dalam penelitian... 25
9 Lokasi pengukuran lingkar bagian tubuh dan tebal lipatan kulit... 27
10 Rata-rata indeks massa tubuh (IMT) monyet ekor panjang... 35
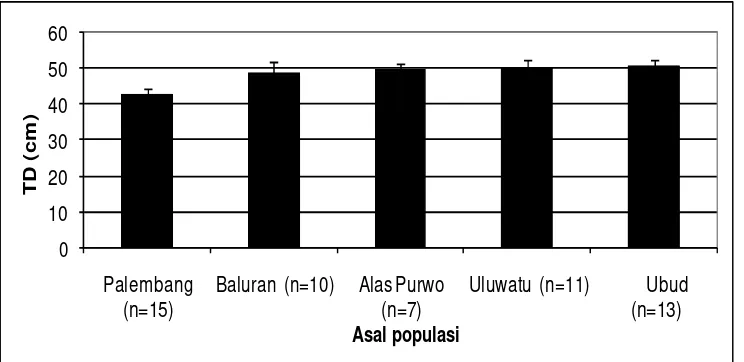
11 Persentase kejadian obesitas monyet ekor panjang... 35
12 Rata-rata tinggi duduk (TD) monyet ekor panjang... 36
13 Rata-rata bobot badan (BB) monyet ekor panjang... 37
14 Rata-rata lingkar pinggang (LPi), dada (LD), paha (LPh), dan lengan atas (LTr) ... 39
15 Rata-rata tebal lipatan kulit perut (TLkAb), leher (TLkLh), punggung (TLkPg), paha (TLkPh), dan trisep(TLkTr) ... 42
16 Plot komponen utama pertama dan komponen utama kedua... 45
17 Skema letak penempelan primer MC4R foward dan MC4R reverse untuk mengamplifikasi gen MC4R monyet ekor panjang ... 47
18 Hasil elektroforesis beberapa produk PCR gen MC4R yang diamplifikasi menggunakan pasangan primer MC4R ... 47
19 Filogram antar populasi yang dibuat dengan metode Neighbor Joining (NJ) berdasarkan perbedaan nukleotida ... 51
20 Filogram antar populasi yang dibuat dengan metode NJ berdasarkan perbedaan asam amino... 51
21 Filogram monyet ekor panjang menggunakan metode NJ dan model p distance dari nukleotida MC4R (999pb)... 52
22 Filogram monyet ekor panjang menggunakan metode NJ dan model p distance dari asam amino MC4R (332 aa) ... 53
23 Filogram menggunakan metode NJ dan model p distance dari nukleotida gen MC4R (999pb) berbagai spesies primata... 55
24 Situs mutasi pada MC4R monyet ekor panjang... 63
DAFTAR LAMPIRAN
Halaman
1 Penjajaran nukleotida (n=999) gen MC4R monyet ekor panjang hasil penelitian dengan urutan gen yang sama yang diperoleh dari
GenBank... 76
2 Penjajaran asam amino (n=332) gen MC4R monyet ekor panjang hasil penelitian dengan urutan gen yang sama yang diperoleh dari GenBank ... 94
3 Analisis varian parameter fenotipe obesitas monyet ekor panjang... 101
4 Nilai eigen dan loading factor pada analisis komponen utama... 103
PENDAHULUAN
Latar Belakang
Saat ini, obesitas dianggap penyakit kompleks multifaktor yang disebabkan oleh interaksi faktor genetik dan lingkungan (Hill dan Peters 1998). Menurut WHO(World Health Organization 1997), obesitas merupakan salah satu dari 10
kondisi yang berisiko di seluruh dunia dan salah satu dari 5 kondisi yang berisiko di negara-negara berkembang. Obesitas merupakan predisposisi utama untuk kondisi terjadinya penyakit, terutama sindroma metabolik, diabetes melitus tipe 2 (DM tipe2) dan hipertensi. Sindroma metabolik merupakan kelainan akibat gangguan metabolik yang ditandai dengan hiperinsulinemia, resistensi insulin, hiperglikemia, dislipidemia aterogenik dan hipertensi. Semua gejala ini merupakan faktor risiko penting untuk penyakit kardiovaskuler. Di samping itu, obesitas dianggap mempunyai andil dalam terjadinya berbagai penyakit kanker seperti kanker kolon, payudara (pada wanita setelah berhenti haid), endometrium, ginjal, esofagus (adenocarcinoma), lambung, pankreas, kandung
kemih, dan hati (Calle dan Kaaks 2004).
Pada umumnya kejadian obesitas banyak ditemukan pada usia dewasa namun demikian obesitas dapat terjadi pada usia muda (anak-anak). Kejadian yang dilaporkan WHO (2005) adalah sekitar 400 juta orang dewasa dan 20 juta anak-anak mengalami obesitas. Pada anak-anak, ada yang dikenal dengan
sindroma MC4R (melanocortin 4 receptor) ditandai dengan peningkatan
pertumbuhan, peningkatan lemak dan kepadatan mineral tulang, hiperfagia dan hiberinsulinemia, yang akhirnya menyebabkan obesitas (Farooqi et al. 2000).
Hewan model (tikus dan monyet ekor panjang) juga menunjukkan gejala obesitas yang mirip dengan manusia. Pada monyet ekor panjang, obesitas terjadi pada monyet dewasa baik pada jantan maupun betina (Putra et al. 2006)
lemak ini juga dapat dilihat jelas bila monyet dalam keadaan duduk. Pada posisi ini, perut monyet kelihatan buncit sebagai akibat dari adanya timbunan lemak tersebut.
Keadaan obesitas pada monyet ekor panjang banyak dijumpai pada monyet-monyet yang hidup di kawasan wisata di Bali. Monyet-monyet ini
menunjukkan tanda-tanda obesitas dengan IMT sampai 61.57 kg/m2 pada jantan
dan 60.07 kg/m2 pada betina (Putra
et al. 2006). IMT ini jauh lebih tinggi
dibandingkan dengan IMT normal pada manusia yaitu 18.5-24.9 kg/m2 (WHO 2005). Lebih jauh, dinyatakan bahwa 100% monyet jantan yang ada di Uluwatu mengalami obesitas, sedangkan kejadian obesitas di daerah lainnya seperti Sangeh, Alas Kedaton, Ubud, Pulaki, dan Bukit Gumang adalah bervariasi antara 12-75%. Kejadian obesitas ini tidak pernah ditemukan pada monyet anakan dan remaja. Padahal obesitas pada manusia dapat terjadi baik pada bayi, anak-anak, remaja maupun dewasa.
Gen yang berkaitan langsung maupun tidak langsung dengan kejadian obesitas pada manusia telah diinventarisasi ada sebanyak 127 gen kandidat. Dari 127 gen tersebut, sebelas diantaranya bersifat monogenik sehingga apabila gen tersebut mengalami mutasi akan berakibat langsung pada timbulnya obesitas (Rankinen et al. 2006). Mutasi pada gen tersebut berupa substitusi, delesi, dan insersi. Hal ini telah dilaporkan terjadi pada gen MC4R. Lebih jauh dinyatakan bahwa kejadian obesitas pada manusia akibat dari mutasi MC4R (monogen) ini diwariskan secara dominan (Dubern et al. 2001; Challis dan Yeo
2002) walaupun prevalensi kejadiannya hanya berkisar antara 4-6% dari individu yang mengalami obesitas (Lubrano-Berthelier et al. 2003). Individu lain yang
tidak mengalami mutasi pada gen MC4R tetapi secara fenotipik individu tersebut gemuk maka diduga kejadian tersebut berasal dari pengaruh kandidat gen lain yang berkontribusi pada obesitas (Mutch dan Clément 2006). Hal tersebut pernah dilaporkan pada tikus yang menunjukkan obesitas akibat terjadinya
mutasi pada gen LEPR (leptin receptor) padahal gen MC4Rnya tidak mengalami
mutasi (Pomp 1999).
MC4R adalah satu dari lima reseptor berprotein-G yang mengikat melanokortin berperan mengatur perilaku makan dan homeostasis energi. Gen MC4R pada manusia telah diurut secara penuh oleh Kopatz et al. (2003) dan
urutan urutannya telah dilaporkan dan dicatat dalam GenBank (NCBI=National
diketahui pertamakali oleh Yeo et al. (1998) sebagai gen yang berkontribusi atau mempunyai asosiasi dengan obesitas akibat mutasi pada gen tersebut. Tao (2005) melaporkan lebih dari 70 jenis mutasi MC4R pada manusia yang menyebabkan berkurang sampai hilangnya fungsi MC4R. Bila MC4R tidak berfungsi maka tidak ada pengisyaratan kekenyangan pada otak sehingga terjadi peningkatan asupan pakan. Peningkatan nafsu makan yang tidak disertai dengan peningkatan penggunaan energi menyebabkan terjadinya penimbunan energi dan mengarah pada obesitas.
Para peneliti telah banyak menggunakan primata nonmanusia sebagai model penelitian untuk mengetahui hubungan antara obesitas dengan penyakit metabolik (Banks et al. 2003). Penelitian obesitas pada primata nonmanusia
dilakukan baik dengan memanipulasi dietnya (Astuti et al. 2007) maupun dengan
cara mengontrol aktivitas fisik dalam kandang individu (Hansen 2001). Pada penelitian model obesitas secara alami pada kelompok monyet rhesus yang hidup bebas di Pulau Cayo Santiago, didapatkan terjadinya obesitas pada populasinya sebanyak 7% (Schwartz et al. 1993). Belakangan ini, pada baboon
dewasa liar telah menunjukkan kejadian yang mirip sindroma metabolik yaitu adanya peningkatan kadar leptin dalam serum darah, obesitas, hiperlipidemia dan resistensi insulin (Banks et al. 2003).
Walaupun dalam penelitian sebelumnya lebih banyak menggunakan rodensia sebagai hewan model untuk penelitian tentang obesitas, saat ini monyet ekor panjang telah digunakan sebagai hewan model penelitian yang berkaitan
dengan obesitas (Chen et al. 2003). Banyak kesamaan fisiologi antara manusia
dan monyet ekor panjang yang menyebabkan monyet ekor panjang merupakan hewan model obesitas yang lebih realistik dibandingkan spesies lain seperti rodensia. Berdasar pada urutan gen MC4R yang telah diteliti oleh Kopatz et al.
(2003) dapat dibandingkan baik asam amino maupun nukleotida dari gen MC4R manusia dengan monyet ekor panjang. Data MC4R monyet ekor panjang yang didapatkan oleh Kusuda et al. (2002) dan tercatat di GenBank (NCBI) dengan
kode akses AB083317) menunjukkan kemiripan susunan asam amino (dari total 332 asam amino) dengan MC4R pada manusia yaitu sebesar 98.5%. Sedangkan, MC4R tikus (kode akses NC_000084) mempunyai kesamaan 93.4% dengan MC4R manusia.
serta interaksi gen-gen dan gen-lingkungan yang tak terkendali. Di samping itu, karena alasan etika, penggunaan teknik invasif pada manusia untuk mengukur ekspresi gen di dalam hipotalamus tidak dapat dilakukan. Oleh karena itu, penggunaan hewan model merupakan suatu keharusan. Penggunaan hewan model dalam penelitian penyakit manusia memberi keuntungan untuk mengungkap sifat (trait) yang heterogen dan poligen.
Penjelasan tentang gen yang menyebabkan obesitas pada hewan model memberikan pengertian pada mekanisme molekuler yang mengatur proses fisiologi seperti keseimbangan energi. Seperti telah diungkapkan di atas, mutasi MC4R pada manusia menunjukkan asosiasi dengan obesitas. Pada hewan, seperti babi, adanya mutasi gen MC4R menyebabkan peningkatan ketebalan lemak punggung (Kim et al. 2000). Demikian juga pada tikus, bila gen MC4R ini
ditiadakan (knock out) maka tikus akan mengalami obesitas (Huszar et al. 1997).
Penelitian mengenai mutasi gen yang terkait dengan obesitas (MC4R) pada monyet ekor panjang belum pernah dilakukan walaupun monyet ekor panjang menunjukkan pola obesitas yang mirip dengan manusia. Untuk itu, perlu dilakukan penelitian polimorfisme gen MC4R pada monyet ekor panjang untuk menunjang pengembangan monyet ekor panjang sebagai hewan model obesitas pada manusia. Polimorfisme gen yang berkaitan dengan obesitas pada monyet ekor panjang dapat memperjelas mekanisme obesitas di tingkat molekuler yang dapat dimanfaatkan sebagai hewan model bagi manusia.
Tujuan Penelitian
1. Mengidentifikasi variasi gen (MC4R) yang berhubungan dengan obesitas pada monyet ekor panjang di Bali, Jawa Timur dan Sumatera.
2. Mengkaji variasi fenotipe obesitas pada monyet ekor panjang di Bali, Jawa Timur dan Sumatera.
3. Mendapatkan asosiasi mutasi gen MC4R dengan obesitas monyet ekor panjang
Hipotesis
Manfaat Penelitian
1. Memberikan informasi tentang mutasi gen MC4R pada monyet ekor panjang 2. Sebagai data dasar untuk penelitian obesitas pada manusia.
3. Memberikan gambaran gen yang ikut terlibat dalam obesitas hewan model. 4. Pengembangan penanda genetik obesitas
Kerangka Pemikiran
Saat ini, obesitas pada manusia dianggap sebagai penyakit multi kompleks dan merupakan predisposisi penyakit diabetes, kardiovaskuler dan kanker. Obesitas disebabkan oleh faktor genetik dan lingkungan atau interkasi kedua faktor tersebut. Adanya mutasi pada gen MC4R dianggap berperan dalam terjadinya obesitas. Sementara itu pada hewan model, misalnya tikus, yang dihilangkan (knock out) gen MC4Rnya menunjukkan gejala obesitas. Sebagai
TINJAUAN PUSTAKA
Monyet Ekor Panjang
Monyet ekor panjang termasuk kelompok monyet dunia lama (Old World monkey) dan diklasifikasikan sebagai berikut (Wilson dan Reeder 2005):
Kelas : Mammalia
Ordo : Primates
Subordo : Haplorrhini
Infraordo : Simiiformis
Superfamili : Cercopithecoidea
Famili : Cercopithecidae
Subfamili : Cercopithecinae
Genus : Macaca
Spesies : Macaca fascicularis (Raffles, 1821).
Monyet ekor panjang sering disebut juga crab-eating macaque atau
cynomolgus macaque. Lebih jauh, dinyatakan bahwa monyet ekor panjang terdiri
atas 10 subspesies yaitu: M. f. fascicularis, M. f. aurea, M. f. umbrosa, M. f. atriceps, M. f. condorensis, M. f. fusca, M. f. lasiae, M. f. tua, M. f. karimondjawae, dan M. f. philippinensis. Di lain pihak, Supriatna dan Wahyono
(2000) melaporkan ada empat subspesies monyet ekor panjang di Indonesia, yaitu M. f. fascicularis, (Pulau Sumatera, Kalimantan, Jawa, Bali, Lombok, Sumbawa, Flores, Sumba dan Timor), M. f. fusca (Pulau Simaleu, Sumatera), M. f. karimondjawae (Pulau Karimunjawa, Jawa Tengah), dan M. f. lasiae (Pulau
Lasia).
dan hutan sekunder. Hutan tropis Indonesia mempunyai kerapatan primata yang tinggi. Alikodra (2002) menyatakan bahwa daya hidup, natalitas dan mortalitas sangat berkaitan dengan kondisi habitat primata. Tingkat ketersediaan pakan juga mempengaruhi penyebaran, kerapatan, dan gerakan-gerakan musiman.
Kegiatan konservasi yang berkaitan dengan monyet ekor panjang telah diungkapkan oleh Ong dan Richardson (2008). Spesies ini dimasukkan ke dalam apendiks II CITES (Convention on International Trade in Endangered Species).
Di samping itu, spesies ini terdaftar pada Schedule I, Part I, Indian Wildlife
(Protection) Act, 1972-2002, dan Schedule III, Bangladesh Wildlife (Protection) Act A 1974. Monyet ekor panjang merupakan spesies yang dilindungi di Myanmar. Di samping itu, monyet ini juga dilindungi dengan Dekrit 32, Apendiks 2B di Viet Nam.
Monyet ekor panjang berjalan dengan empat kaki (quadrupedalism),
memiliki ekor yang lebih panjang dari panjang kepala dan badan, serta memiliki bantalan duduk (ischial callosity) yang melekat pada tulang duduk. Monyet ini
menunjukkan perbedaaan antara jantan dan betina (sexual dimorphism). Bobot
badan, panjang kepala dan badan, dan panjang ekor pada yang jantan masing-masing adalah 4.7-8.3 kg, 412-648 mm, dan 435-655 mm, sedangkan pada yang betina masing-masing adalah 2.5-5.7 kg, 385-503 mm, dan 400-550 mm (Rowe 1996). Putra et al. (2006) mendapatkan bahwa bobot badan monyet ekor
panjang jantan bervariasi antara 3-12 kg, sedangkan yang betina berkisar antara 3-10 kg. Warna tubuhnya bervariasi mulai dari abu-abu sampai kecoklatan dengan bagian ventral putih, sedangkan anak yang baru lahir warna rambutnya hitam (Supriatna dan Wahyono 2000).
Monyet ekor panjang hidup dalam kelompok yang terdiri dari banyak jantan dan banyak betina. Jumlah anggota kelompoknya berkisar antara 10 sampai lebih dari 100 ekor, namun ukuran kelompok biasanya antara 20 dan 50 ekor (Fuentes 2007). Di dalam kelompok, betina dewasa biasanya lebih banyak daripada jantan dewasa dan aktivitas sosialnya berputar di antara kelompok betina yang berhubungan (Berkovitch dan Huffman 1999).
Hindu di samping makanan yang diberikan oleh pengelola, dan pengunjung (Putra et al. 2001). Makanan ini sering berlebih jumlahnya sehingga tidak jarang
monyet yang ada di daerah tersebut mengalami obesitas.
Obesitas
Obesitas merupakan kejadian kelebihan timbunan lemak sebagai akibat dari ketidakseimbangan pemasukan dan penggunaan energi. Asupan makanan semakin meningkat karena ketersediaan makanan yang semakin banyak, dan mudah didapat, sedangkan aktivitas fisik semakin berkurang. Hal ini mengakibatkan energi yang masuk lebih besar daripada energi yang digunakan,
yang pada giliranya menyebabkan obesitas (Chen et al. 2000). Secara
sederhana, obesitas dapat dilihat dari penampilan luar seperti perut yang membesar akibat dari timbunan lemak subkutan. Di samping itu, obesitas dapat ditentukan dengan menghitung IMT.
Pada manusia, IMT ditentukan dengan perbandingan bobot badan (kg) dan tinggi badan kuadrat (m2) sedangkan pada monyet, IMT dihitung berdasarkan bobot badan dan tinggi duduk (panjang dari bagian tertinggi kepala sampai pangkal ekor/crown rump length). IMT yang sama atau lebih besar dari 30
digolongkan ke dalam obesitas (WHO 2005).
Obesitas ditentukan oleh faktor gen dan lingkungan (Hill dan Peters 1998). Faktor genetik sangat terlibat dalam akumulasi jaringan lemak perut (Bouchard et al. 1990). Lebih lanjut dinyatakan bahwa obesitas melibatkan multi gen,
lingkungan, dan interaksi antara kedua faktor tersebut. Ada 244 gen, bila mengalami mutasi atau diekspresikan sebagai transgen pada tikus, yang berpengaruh pada bobot badan dan adiposit (Rankinen et al. 2006).
Gambar 2 Obesitas pada monyet ekor panjang
Hotta et al. (1996) melakukan penelitian lebih lanjut pada monyet rhesus.
Mereka meneliti kaitan antara gen gemuk (ob gen) dengan bobot badan. Mereka
menemukan bahwa ada hubungan antara kadar obmRNA dengan bobot badan.
Mereka juga menemukan hubungan yang signifikan antara ob mRNA dengan
insulin puasa, walaupun stimulasi insulin selama 100-140 menit, euglikemia/
hiperinsulinemia tidak menyebabkan perubahan pada kadar ob mRNA. Kadar ob
gen yang menghasilkan leptin juga berkaitan secara signifikan dengan bobot badan. Jadi, ekspresi gen ob berkaitan dengan bobot badan dan tidak secara
akut diatur oleh insulin.
Eisner et al. (2003) menemukan bahwa depot lemak intra-abdominal dan abdominal total meningkat pada monyet rhesus betina dewasa yang terkena kelebihan androgen saat prenatal. Hasil penelitian ini menunjukkan perubahan pada adipositas abdominal sebagai konsekuensi lain kelebihan androgen prenatal terhadap gangguan sekresi insulin yang diamati pada hewan ini pada saat dewasa.
Selanjutnya, Angeloni et al. (2004) melakukan penelitian tentang
dihasilkan oleh saluran pencernaan. Mereka mengatakan bahwa ghrelin
merangsang penglepasan growth hormone (GH) dari hipofisa, merangsang nafsu
makan, dan mempengaruhi proses metabolik pada jaringan lain yang mempunyai reseptor GH. Dengan demikian ghrelin dapat mempengaruhi tingkah laku dan jalur endokrin yang berperan dalam pertambahan bobot badan. Kadar grelin dipengaruhi oleh faktor umur dan adipositas pada monyet rhesus.
Tigno et al. (2004) mengamati hubungan perubahan parameter metabolik
dengan umur pada monyet rhesus. Mereka menemukan bahwa laju
penghilangan glukosa, laju sensitivitas insulin dan kadar HDL (high density lipoprotein) berubah secara signifikan pada kelompok yang tidak menderita diabetes dengan umur yang berbeda. Pada kelompok monyet rhesus yang menderita diabetes, penurunan toleransi glukosa terlihat pada umur pertengahan (14th), dan ditemukan ada peningkatan insulin puasa sebelum menderita diabetes, dan kemudian menurun dengan bertambahnya umur.
Peneliti lainnya, seperti Takahashi et al. (2006) melakukan penelitian pada
monyet Jepang (M. fuscata). M. fuscata gemuk di Wakasa tidak mengalami
kelainan/penyakit yang berkaitan dengan obesitas seperti diabetes. Dalam kelompok Wakasa, frekuensi gemuk tinggi dalam garis maternal. Hal ini menunjukkan bahwa faktor genetik obesitas dapat diwariskan melalui garis ini.
Penelitian genetika juga dilakukan oleh Comuzzie et al. (2003) pada
baboon (genus Papio). Mereka menemukan bahwa pola akumulasi jaringan
lemak mirip dengan yang terjadi pada manusia. Lebih jauh, dikatakan bahwa ada pewarisan genetik pada fenotipe yang berkaitan dengan obesitas.
Chen et al. (2002) melakukan penelitian tentang hubungan antara bobot
badan dengan parameter biokimia serum dan hematologi pada monyet ekor panjang betina. Mereka mengatakan bahwa obesitas adalah faktor risiko yang memicu berbagai penyakit metabolik. Monyet ekor panjang (M. fascicularis) menunjukkan obesitas yang spontan pada saat dewasa dan hal ini mirip dengan manusia. Untuk menjelaskan karakter yang berkaitan dengan obesitas pada monyet ekor panjang betina, mereka menggunakan analisis regresi sederhana dan berganda untuk menentukan hubungan antara bobot badan dan parameter biokimia serum dan hematologi seperti hormon yang berkaitan dengan obesitas (leptin dan insulin). Analisis regresi sederhana menunjukkan bahwa bobot badan
secara nyata berhubungan dengan kadar leptin, insulin, konsentrasi
glukosa dan konsentrasi trigliserida. Di samping itu, model regresi berganda yang mengandung kadar leptin, insulin, MCV, dan eritrosit menjelaskan 66.9%
keragaman dalam bobot badan. Oleh karena itu, monyet ekor panjang betina menunjukkan karakter obesitas yang mirip dengan manusia, yaitu obesitas berkaitan dengan meningkatnya sintesis dan ekskresi leptin oleh adiposit, berisiko tinggi menderita diabetes melitus, dan kadar hematosit yang tinggi.
Lebih lanjut, Chen et al. (2003) mengatakan bahwa leptin berkorelasi positif
dengan insulin dan persentase lemak sedangkan adiponektin berkorelasi negatif
baik pada M. fascicularis jantan dan betina. Mereka mencoba untuk
menunjukkan bahwa rasio leptin dan adiponektin (L/A) merupakan indeks yang potensial untuk menentukan obesitas pada monyet ini. Mereka menemukan bahwa rasio leptin dan adiponektin secara nyata meningkat pada monyet yang persentase lemaknya melebihi 40%.
Melanokortin
Melanokortin (hormon adrenokortikotropik dan a, ß, ?-MSH) berasal dari prekursor protein pro-opiomelanokortin (POMC) Hormon ini bekerja melalui pasangan protein G dengan merangsang adenilat siklase (Mountjoy et al. 1992).
Melanokortin memiliki aksi fisiologi yang luas termasuk penglepasan agen neurohormonal seperti hormon prolaktin dan hormon pelutein/luteinizing hormone
(Ellerkmann et al. 1992), mengatur sistem kardiovaskuler (Li et al. 1996), obesitas (Fan et al. 1997), pewarnaan pigmen melanosit, termoregulasi, memori,
pembelajaran, tingkah laku, rasa sakit/analgesia, dan amina biogen, efek imunomodulator, sifat neurotropik dan terlibat pada proses kelahiran (Blondet et
al. 2005)
Peptida yang dihasilkan oleh gen POMC berperan dalam proses fisiologi seperti steroidogenesis adrenal, pigmentasi kulit, analgesia dan inflamasi. Pada saat ini, ditemukannya bukti pada tikus dan manusia bahwa sinyal melanokortin berperan pada neuron hipotalamus yang mengekspresikan POMC dalam mengatur nafsu makan dan bobot badan (Yeo et al. 2000)
Gen POMC diekspresikan dalam nukleus arcuata hipotalamus dan nucleus
tractus solitarius (Yeo et al. 2000). Di samping itu, gen POMC diekspresikan
adrenokortikotropin (ACTH), ß-endorfin dan a, ß, ?-MSH. Kumpulan produk yang berasal dari POMC ditentukan oleh jaringan dengan kekhasan endoprotease (konvertase) yang diekspresikan dalam jalur sekresi. Dengan demikian, kortikotrof hipofisa anterior mengekspresikan prohormon konvertase-1 (PC1) dan memecah POMC menjadi ACTH, sementara melanotrof dalam lobus intermedia (pada hewan yang lebih rendah) mengekspresikan prohormon konvertase-2
(PC2) dan memecah ACTH menjadi a-MSH. Jadi, ACTH yang disekresikan
oleh hipofisa anterior merupakan pengendali utama dari steroidogenesis adrenal. Pada amfibia dan rodensia, a-MSH dari lobus intermedia hipofisa terlibat dalam pengaturan warna kulit dan rambut. Mamalia juga mengekspresikan POMC dalam kulit, dan secara lokal menskresi a, ß, ?-MSH, ACTH dan ß-endorfin, walaupun fungsi fisiologinya tidak diketahui. Kepentingan peptida ini pada pigmentasi manusia ditunjukkan dengan hubungan varian reseptor melanokortin-1 manusia dengan rambut merah dan kulit putih (Yeo et al. 2000).
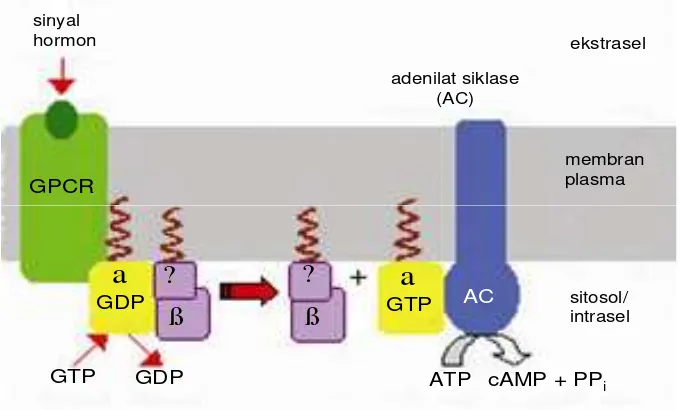
Mekanisme kerja melanokortin adalah dengan cara mengaktifkan reseptor melanokortin (MCR) yang selanjutnya mengaktifkan protein G. Sub unit a protein G mengikat guanosin trifosfat (GTP) dan mengalami pemisahan yang kemudian berikatan dengan enzim adenilat siklase. Adenilat siklase aktif akan mengkatalisis perubahan ATP menjadi cAMP dan fosfat. Dengan demikian,
proses ini menyebabkan peningkatan produksi cAMP (cyclic adenosin
monophosphate) di dalam sel. Selanjutnya, cAMP ini akan mengaktifkan molekul
[image:30.612.143.483.475.680.2]lain seperti protein kinase A (Gambar 3).
Gambar 3 Sinyal protein G yang mengatur produksi cAMP (Diwan 2007)
a
GTP sinyalhormon
adenilat siklase (AC)
ekstrasel
membran plasma
sitosol/ intrasel
AC
ß
?
a
GTP
a
GDP
?
ß
GTP GDP ATP cAMP + PPi
Reseptor Melanokortin 4
Reseptor melanokortin 4 (MC4R) disandi oleh gen MC4R. MC4R merupakan gen berekson tunggal dengan 999 pasang basa yang menyandi 332 asam amino. Organisasi gen MC4R disajikan dalam Gambar 4. Pada manusia, gen ini terletak di dalam kromosom 18q22 (Gantz et al. 1993), demikian juga
halnya pada tikus dan primata. Namun pada anjing, burung dan sapi, gen ini masing-masing terletak pada kromosom 1, 2, dan 24.
Gen yang sama pada tikus mempunyai kesamaan (93%) dengan gen
manusia(Alvaro et al. 1996). Sedangkan kesamaan gen MC4R antara manusia
dan monyet ekor panjang adalah 98.5%. Hal ini menunjukkan bahwa gen ini sangat kekal (conserved) pada mamalia.
Gambar 4 Organisasi gen MC4R
MC4R merupakan gen yang paling erat hubungannya dengan bobot badan
(Yeo et al. 2000). Peran utama MC4R dalam mengatur bobot badan ditunjukkan
pertamakali pada hewan pengerat (rodensia). Kenyataan bahwa reseptor ini mengatur asupan pakan dan penggunaan energi didasarkan pada gen target pada mencit yang menyebabkan obesitas dan disertai hiperfagia (makan berlebihan), hiperinsulinemia (kadar insulin darah meningkat), dan hiperglikemia (gula darah meningkat) (Huszar et al. 1997).
MC4R adalah pasangan protein G, reseptor transmembran-7 yang banyak diekspresikan di hipotalamus yaitu bagian otak yang terlibat dalam pengaturan nafsu makan (Mountjoy et al. 1994). Ekspresi gen ini pada manusia ternyata
dikontrol oleh beberapa anggota faktor transkripsi Sp/specific protein (Blondet et
al. 2005). Pengisyaratan MC4R diatur oleh agonis a-MSH endogen dan
antagonis AGRP (Schwartz et al. 2000). Perkembangan selanjutnya
menunjukkan bahwa selain a-MSH, ada peptida turunan POMC yang berfungsi sebagai agonis MC4R (Nickolls et al. 2003).
MC4R ditemukan terutama di dalam otak dengan analisis Northern blot dan
teknik hibridisasi in situ (Gantz et al. 1993). Sebaran reseptor ini di dalam otak
3’ 5’
lebih luas dari sebaran MC3R dan meliputi korteks, talamus, hipotalamus, batang otak, dan medula spinalis. Namun, MC4R tidak ditemukan pada sel perifer. Sebaran MC4R ini konsisten dengan keterlibatannya dalam fungsi otonom dan neuroendokrin.
Perubahan MC4R dan Akibatnya
Pengaruh MC4R pada obesitas manusia telah dipelajari sejak 1997. Hubungan obesitas (IMT, massa lemak, %lemak) dengan MC4R dideteksi dalam studi keluarga orang Kanada (Chagnon et al. 1997). Variasi missense V103I (perubahan asam amino valina pada posisi 103 menjadi isoleusina) pertama kali ditemukan pada laki-laki Inggris (Gotoda et al. 1997) tetapi tidak ada asosiasi
(hubungan) dengan obesitas. Penggantian asam amino isoleusina pada posisi 137 menjadi treonina (I137T) menyebabkan gangguan fungsi reseptor dan berhubungan dengan obesitas.
Perubahan (mutasi) reseptor ini berhubungan dengan obesitas dan ternyata kejadian ini dapat diwariskan. Bukti mengesankan pertama kali bahwa obesitas manusia diwariskan secara dominan dan berhubungan dengan mutasi gen MC4R dilaporkan pada tahun 1998 (Yeo et al. 1998). Pewarisan (h2)
karakter obesitas seperti IMT pada baboon dapat mencapai 0.46 sedangkan pada manusia bervariasi antara 0.26-0.54 (Cai 2004).
Mutasi gen ini pada hewan juga menyebabkan adanya perubahan fenotipe. Mutasi gen MC4R pada babi juga dinyatakan berhubungan dengan ketebalan lemak punggung (Kim et al. 2000). Demikian juga SNP (Single Nucleotide
Polymorphism) pada daerah penyandi gen MC4R sapi dilaporkan merupakan
hal yang khas terjadi pada jenis (breed) sapi tertentu (Haegeman et al. 2001).
sedangkan yang hanya kehilangan satu alel saja (heterosigot) menunjukkan gejala obesitas sedang(Huszar et al. 1997).
Kepentingan MC4R dalam pengaturan bobot badan manusia pertama kali
muncul pada tahun 1998, ketika ditemukan bahwa mutasi frameshift heterosigot
dalam MC4R sebagai penyebab dominan obesitas (Yeo et al. 1998).
Selanjutnya dilaporkan bahwa ada asosiasi antara varian mutan MC4R dan obesitas (Hinney at al. 1999). Mutasi ini merupakan kerusakan monogen yang
paling umum diketahui sebagai penyebab obesitas pada manusia (Farooqi et al.
2000) dan dilaporkan mencapai 4-6% dari individu gemuk (Lubrano-Berthelier et
al. 2003). Saat ini, mutasi MC4R dianggap memberikan pengaruh utama pada
obesitas (Hinney et al. 2003). Mutasi yang ditemukan pada MC4R adalah mutasi
frameshift (Yeo et al. 1998), nonsense (Hinney et al. 1999) dan missense
(Gotoda et al. 1997).
Mutasi Gen MC4R
Walaupun heritabilitas IMT cukup tinggi, mutasi yang menyebabkan keadaan sangat gemuk telah ditemukan hanya dalam satu gen, yaitu gen MC4R yang menyandi reseptor melanokortin 4. Reseptor ini berhubungan dengan sinyal oreksigenik (nafsu makan) dan anoreksigenik di dalam hipotalamus dan tempat lain di dalam sistem saraf pusat untuk mengatur asupan pakan dan penggunaan energi. Gen MC4R hanya terdiri dari satu ekson dan dapat diterima untuk analisis mutasi yang tersedia secara klinik. Yeo et al. (1998) pertama kali
melaporkan bahwa mutasi frameshift MC4R heterozigot berhubungan dengan
keadaan sangat gemuk. Sejak saat itu, lebih dari 70 mutasi dilaporkan pada berbagai studi populasi dan kebanyakan dengan keadaan sangat gemuk. Temuan tersebut meliputi keadaan hiperfagia (makan berlebihan), percepatan pertumbuhan pada anak-anak (walaupun tinggi badan akhir tampak normal), peningkatan kepadatan tulang yang umum dijumpai dalam keadaan obesitas. Individu dengan mutasi MC4R homozigot lebih gemuk daripada saudaranya yang heterozigot. Hal ini menunjukkan bahwa cara pewarisannya bersifat kodominan. Sampai saat ini, mutasi yang dilaporkan terjadi adalah dalam bentuk nonsinonim (n=57), nonsense (n=5), dan frameshift (n=10) yang ditemukan pada 140 dari
6134 orang yang sangat gemuk dalam berbagai sampel penelitian (Hinney et al.
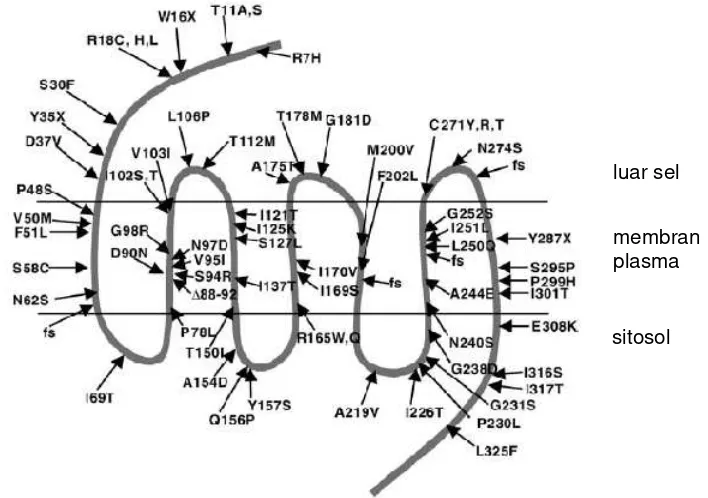
bervariasi antara 0.5-5.8% (Kublaoui dan Zinn 2006). Posisi mutasi MC4R pada manusia disajikan dalam Gambar 5.
Gambar 5 Kejadian mutasi MC4R pada manusia (Tao 2005)
Mutasi merupakan perubahan yang terjadi pada urutan DNA (deoxyribo nucleic acid). Urutan DNA biasanya disalin dengan tepat pada saat replikasi kromosom. Kesalahan kadang-kadang terjadi sehingga terjadi urutan yang baru. Kesalahan inilah yang dikenal dengan mutasi. Mutasi dapat terjadi pada sel somatik maupun sel kecambah. Karena mutasi sel somatik tidak diturunkan, hal ini dapat diabaikan dalam konteks evolusi.
Mutasi dapat digolongkan berdasarkan panjang urutan DNA yang dipengaruhi kejadian mutasi. Misalnya, mutasi dapat terjadi pada satu nukleotida (point mutation) atau beberapa nukleotida yang berdekatan. Mutasi juga dapat
digolongkan berdasarkan pada tipe perubahan yang disebabkan oleh kejadian mutasi yaitu, (1) substitusi, penggantian satu nukleotida oleh yang lain, (2) delesi, kehilangan satu atau lebih nukleotida dari DNA, (3) insersi, penambahan satu atau lebih nukleotida ke dalam urutan DNA, dan (4) inversi, rotasi 180o segmen DNA untai ganda yang terdiri atas satu atau lebih pasang basa.
membran plasma luar sel
Substitusi dibagi menjadi dua yaitu, (1) transisi dan (2) transversi. Transisi adalah substitusi antara A dan G (purin) atau antara C dan T (purimidin). Transversi adalah substitusi antara purin dan pirimidin. Substitusi nukleotida yang terjadi pada daerah penyandi protein dapat digolongkan berdasarkan pada efek pada produk proteinnya. Mutasi yang tidak menyebabkan perubahan asam amino disebut sinonim sedangkan yang menyebabkan perubahan asam amino disebut nonsinonim. Mutasi nonsinonim terdiri atas mutasi missense dan
nonsense. Mutasi missense mengubah kodon menjadi kodon yang menyandi
asam amino yang berbeda sedangkan mutasi nonsense mengubah kodon
menjadi kodon terminal (stop codon)
Delesi dan insersi dapat terjadi dengan beberapa mekanisme. Salah satu mekanismenya adalah unequal crossing over. Mekanisme ini dapat terjadi
antara dua kromosom yang menghasilkan delesi segmen DNA pada salah satu kromosom dan penambahan pada kromosom yang lain. Mekanisme lainnya adalah replication slippage atau slipped-strand mispairing. Mekanisme ini bisa
menyebabkan perubahan cetakan asam amino di daerah penyandi protein dan disebut mutasi frameshift (Li 1997).
Klasifikasi Mutasi MC4R
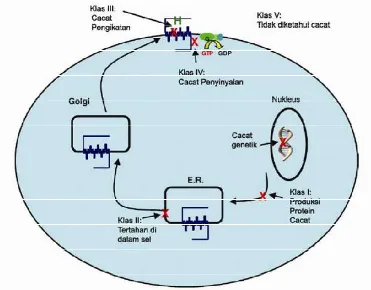
Penelitian saat ini tentang kelainan fungsi varian MC4R yang berhubungan dengan obesitas menunjukkan adanya kelainan dalam banyak hal. Tao (2005) menggolongkan mutasi MC4R menjadi lima klas (Gambar 6).
1) Klas I: Tanpa mutasi. Sintesis protein yang tidak sempurna dan/atau degradasi protein yang cepat menyebabkan tidak adanya reseptor di dalam sel. Mutan W16X, Y35X, dan L64X diduga ke dalam klas ini, walaupun penelitian tentang ekpresinya masih diperlukan untuk membuktikan dugaan ini.
2) Klas II: Mutan yang terperangkap di dalam sel. Reseptor mutan ini diproduksi tetapi tetap berada di dalam sel, kemungkinan besar di dalam retikulum endoplasma karena dideteksi sebagai lipatan yang keliru oleh sistem pengendalian mutu sel tersebut. Klas ini merupakan kumpulan mutasi MC4R yang terbanyak dilaporkan saat ini termasuk mutasi frameshift ?CTCT pada
Gambar 6 Klasifikasi mutasi MC4R pada manusia (Tao 2005)
3) Klas III: Mutan yang cacat bidang pengikatannya. Reseptor mutan ini diekspresikan di permukaan sel tetapi cacat pada ligan pengikatnya, baik yang kemampuan pengikatannya dan/atau afinitasnya menurun, sehingga menyebabkan gangguan pemberian isyarat yang dirangsang oleh hormon. Mutan ini meliputi N97D, L106P, I125K, I137T, I316S, ?88-92, I102S, dan I102T.
4) Klas IV: Mutan yang cacat pemberian isyaratnya. Mutan ini diekspresikan pada permukaan sel, aktivitas ligan pengikatnya normal, tetapi cacat dalam pemberian isyarat yang dirangsang oleh agonis (kemampuan dan/atau kemanjurannya menurun). Mutan yang termasuk dalam golongan ini adalah D90N, I137T, A175T, dan V253I. Mutan V253I ditemukan memiliki respon maksimum yang relatif normal, walaupun EC50 (konsentrasi hormon yang diperlukan untuk menghasilkan 50% respon maksimum) meningkat tiga kali
lipat. Namun, Marti et al. (2003) menemukan bahwa ada mutan V253I pada
5) Klas V: Mutan yang fungsinya normal. Mutan yang termasuk dalam golongan ini adalah T11A, D37V, P48S, V50M, F51L, A154D, I170V, M200V, N274S, dan S295P. Mutan ini menunjukkan ekspresi permukaan sel, pengikatan ligan dan cAMP terpacu secara normal. Mekanisme varian ini dalam menyebabkan ketidakseimbangan energi dan kegemukan masih belum jelas. Perbedaan varian ini dengan polimorfisme yang lain seperti V103I dan I251L adalah bahwa varian ini tidak ditemukan pada subjek yang normal.
Mekanisme Molekuler Homeostasis Energi
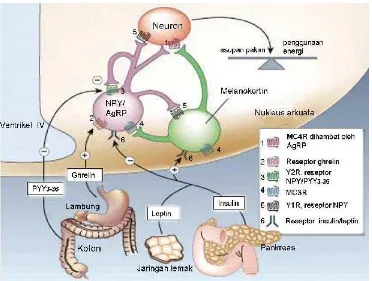
Penyimpanan lemak yang terdapat di dalam tubuh ternyata bukan merupakan hasil dari kebiasaan buruk yang bersifat pasif seperti yang diduga sebelumnya. Ternyata jaringan lemak (adiposa) berperan pada pengaturan proses homeostasis energi, yaitu suatu proses yang membutuhkan keseimbangan antara asupan energi (asupan makanan) dan pengeluaran energi (metabolisme dan gerak) serta ukuran cadangan energi (massa lemak) dalam tubuh (Woods dan Seeley 2002). Sistem yang mengatur asupan makanan, mengontrol seberapa sering dan seberapa banyak makanan yang dimakan, dan memperbaiki ketika keseimbangan jangka panjang terganggu, merupakan suatu masalah yang kompleks dan belum dipahami dengan jelas. Salah satu fungsi sistem ini antara lain mengatur bobot badan jangka pendek, seberapa sering dan berapa banyak makanan yang dimakan dalam waktu tertentu. Fungsi lainnya adalah mengatur asupan makanan dalam jangka waktu yang lebih panjang (Gambar 7). Beberapa tahun terakhir ini, para peneliti telah menemukan sejumlah komponen yang terlibat dalam sistem ini. Adanya dua hormon peptida, yaitu ghrelin dan peptida YY (PYY) yang diproduksi oleh saluran pencernaan diketahui mempengaruhi perilaku makan jangka pendek, sedangkan leptin dan insulin mengatur bobot badan dalam jangka waktu bulanan atau tahunan. Para peneliti obesitas telah menghasilkan kemajuan untuk memahami efek hormon di atas. Mereka menemukan daerah di otak, seperti arcuate nucleus (nukleus
Gambar 7 Hormon yang mengatur nafsu makan. Leptin yang diekskresikan dari jaringan lemak dan insulin dari pankreas menurunkan nafsu makan. Dalam nukleus arkuata hipotalamus, hormon ini menghambat neuron
yang menghasilkan neuropeptida Y (NPY) dan agouti-related protein
(AgRP) yang merangsang neuron penghasil melanokortin. NPY dan
AgRP adalah peptida yang merangsang makan. Hormon
perangsang melanosit-a (a-MSH), dihasilkan dengan pemrosesan pro-opiomelanokortin, adalah ligan endogen untuk reseptor melanokortin MC3R dan MC4R yang mengisyaratkan kekenyangan. Ghrelin yang dihasilkan dari lambung merangsang nafsu makan dengan mengaktifkan neuron penghasil NPY/AgRP. PYY3-36 yang dihasilkan dari kolon bertindak melalui reseptor Y2 neuropeptida dan menghambat neuron NPY/AgRP, dan akhirnya menurunkan nafsu makan (Schwartz dan Morton 2002).
Nukleus arkuata memiliki dua tipe neuron yang memiliki aksi berlawanan. Pengaktifan neuron tipe pertama, yang memproduksi neurotransmiter peptida yaitu neuropeptida Y (NPY) dan agouti-related peptide (AgRP), merangsang selera makan sambil mengurangi metabolisme. Sebaliknya, pengaktifan neuron tipe lainnya, yang dikenal sebagai neuron POMC/CART (pro-opiomelanocortin
22
Namun pada hewan yang tidak tahan, peningkatan cadangan lemak dan konsentrasi leptin dapat menghambat neuron NPY/AgRP dan mengaktifkan neuron POMC yang akhirnya menyebabkan penurunan bobot badan. Neuron NPY/AgRP dan POMC/CART menyampaikan sinyalnya melalui otak di beberapa bagian lainnya menuju nucleus tractus solitarius dari pusat otak, kemudian akan dihantarkan ke seluruh tubuh (Marx 2003).
Sejak ditemukannya leptin (hormon polipeptida yang dihasilkan oleh adiposit), banyak kemajuan telah terjadi dalam menjelaskan jalur saraf yang mengatur homeostasis energi. Sejumlah peptida, termasuk neuropeptida Y (NPY), ghrelin, peptida mirip glukagon, hormon melanin, oreksin (hipokretin) dan melanokortin, dinyatakan terlibat dalam pengaturan keseimbangan energi. Sirkuit melanocortin-leptin berperan dalam pengaturan keseimbangan energi ini (Tao 2005). Dalam sirkuit ini, leptin yang diproduksi oleh adiposit dapat menembus sawar (barrier) darah otak, berikatan dengan reseptornya pada dua neuron di nukleus arkuata hipotalamus. Neuron yang satu mengekspresikan NPY dan AGRP, sedangkan neuron lainnya mengekspresikan POMC/CART. POMC diproses dalam jaringan menjadi melanokortin, termasuk a, ß, ?-MSH dan ACTH oleh prohormon konvertase. Selanjutnya melanokortin bekerja mengikuti jalur penyinyalan protein G.
Teknik PCR
Reaksi berantai polimerase (polymerase chain reaction, PCR) adalah suatu
metode enzimatis untuk menggandakan secara eksponensial suatu urutan nukleotida tertentu dengan cara in vitro. Metode ini pertama kali dikembangkan pada tahun 1985 oleh Kary B. Mullis seorang peneliti perusahaan CETUS
Corporation. Metode ini sekarang telah banyak digunakan untuk berbagai
macam manipulasi dan analisis genetik. Pada awal perkembangannya, metode ini hanya digunakan untuk melipatgandakan molekul DNA, tetapi kemudian dikembangkan lebih lanjut sehingga dapat digunakan pula untuk melipatgandakan dan melakukan kuantitasi molekul m-RNA.
ini menunjukkan bahwa pelipatgandaan suatu fragmen DNA dapat dilakukan secara cepat. Kelebihan lain dari metode PCR adalah bahwa reaksi ini dapat dilakukan dengan menggunakan komponen dalam jumlah sedikit, misalnya DNA-cetakan yang diperlukan hanya sekitar 5µg, oligonukleotida yang diperlukan hanya sekitar 1 mM dan reaksi ini bisa dilakukan dalam volume 50-100 µl. DNA-cetakan yang digunakan juga tidak perlu dimurnikan terlebih dahulu sehingga metode PCR dapat digunakan untuk melipatgandakan suatu urutan DNA dalam genom bakteri hanya dengan mencampurkan kultur bakteri di dalam tabung PCR (Yuwono 2006).
Siklus PCR meliputi beberapa tahap yaitu 1) tahap denaturasi fragmen DNA, 2) tahap penempelan, dan 3) tahap elongasi. Pada tahap pertama terjadi pemisahan rantai ganda DNA menjadi rantai tunggal akibat suhu tinggi ( 95oC) sedangkan pada tahap kedua terjadi penempelan primer pada DNA-cetakan yang sesuai pada suhu 55oC. Tahap ketiga (elongasi) dilakukan pada suhu 72oC. Pada tahap ini Taq DNA Polymerase melakukan aktivitas polimerisasi unit DNA dengan arah 5’ ke 3’ yang komplementer dengan urutan DNA-cetakan yang diapit oleh satu set primer. Fragmen DNA hasil polimerisasi ini selanjutnya menjadi cetakan untuk siklus berikutnya sehingga produk akhir fragmen sangat banyak. Siklus PCR biasanya dilakukan sebanyak 30 kali ditambah satu siklus setelah siklus ketiga puluh dengan waktu elongasi yang lebih panjang (5 menit). Waktu elongasi yang lebih panjang memberi kesempatan untuk fragmen DNA teramplifikasi secara keseluruhan (Becker et al. 1996).
Efisiensi PCR sangat dipengaruhi oleh konsentrasi berbagai komponen. Komponen yang terlibat yaitu Taq DNA Polymerase, dNTP (deoksinukleosida trifosfat), ion Mg, DNA-cetakan dan primer. Selain itu suhu dan waktu
MATERI DAN METODE
Waktu dan Lokasi Penelitian
Pengambilan data fenotipe dan contoh darah di Uluwatu, Ubud, dan Pusat Studi Satwa Primata-Lembaga Penelitian dan Pengabdian kepada Masyarakat, Institut Pertanian Bogor (PSSP-LPPM, IPB), dilakukan pada bulan Juni sampai dengan Desember tahun 2007. Pengambilan contoh darah dan data fenotipe di Alas Purwo dan Taman Nasional Baluran (Jawa Timur) dilakukan tahun 2005. Analisis laboratorium dilakukan di Laboratorium Biologi dan Reproduksi, PSSP-LPPM, IPB, Bogor dan Laboratorium Biologi Molekuler, Pusat Penelitian Sumberdaya Hayati dan Bioteknologi IPB Bogor mulai Agustus 2007 sampai Agustus 2008. Pengurutan produk PCR menggunakan jasa Macrogen, Korea.
Materi
Monyet Ekor Panjang
Hewan yang digunakan dalam penelitian ini adalah monyet ekor panjang (Macaca fascicularis) jantan dewasa sebanyak 56 ekor (Tabel 1). Monyet ekor
panjang jantan tidak terpengaruh oleh siklus kebuntingan sehingga bobot badan dan data fenotipe obesitas tidak dipengaruhi oleh siklus tersebut. Umur ditentukan berdasarkan gigi geligi yaitu: a) muda/remaja dengan ciri taring permanen dan molar permanen ke tiga belum tumbuh dan rambut sudah kecoklatan; b) pra dewasadengan ciri gigi geligi sudah permanen tetapi molar ke
tiga belum tumbuh dan rambut kecoklatan; dan c) dewasadengan ciri gigi taring
[image:41.612.123.513.560.648.2]sudah panjang (pada jantan) dan molar permanen berjumlah 12 buah dan rambut kecoklatan.
Tabel 1 Jumlah monyet ekor panjang jantan yang digunakan dalam penelitian ini
Pulau/Kawasan Asal Kode Jumlah
Bali Ubud
Uluwatu UW UB 13 11
Jawa Timur Alas Purwo
Baluran AP BL 10 7
Sumatera Palembang* Pal 15
Jumlah 56
*Sampel asal Palembang diambil di PSSP-IPB
Sampel asal Palembang diambil di PSSP-LPPM, IPB. Lokasi asal populasi monyet yang digunakan dalam penelitian ini ditunjukkan pada Gambar 8.
Gambar 8 Peta lokasi asal monyet ekor panjang yang digunakan dalam penelitian
Bahan dan Alat Pengambilan Sampel Darah dan Data Fenotipe
Pengambilan sampel darah monyet menggunakan bahan-bahan yaitu: alkohol 70%, EDTA (ethylene diamine tetraacetic), ketamin, xylazine dan kapas,
sedangkan alat yang digunakan adalah tulup, pistol, spuit, tabung vacutainer,
spidol permanen, kotak pendingin, dan lemari pendingin (freezer).
Alat yang digunakan untuk mendapatkan data fenotipe adalah pita ukur (skala terkecil 1 mm), timbangan (skala terkecil 0.1 kg), kaliper (skala terkecil 1 mm), alat tulis dan kertas.
Bahan dan Alat Isolasi DNA
Bahan yang digunakan dalam isolasi DNA dari darah adalah etanol absolut, larutan penyangga AW1, AW2, AL dan AE, proteinase K (produk Qiagen), sedangkan alat yang digunakan adalah mikro pipet beserta ujung
50 0 50 Kilometers
Baluran
Alas Purwo
Ubud
Uluwatu U
[image:42.612.136.504.130.421.2]pipetnya dengan ukuran 20, 200, dan 1000 µl, tabung 1.5 ml, QIAamp spin coloum,water bath, alat mikrosentrifugasi, termometer, rak dan vorteks.
Bahan dan Alat Analisis PCR
Bahan yang digunakan untuk PCR adalah ddH2O, dNTP (dATP, dCTP, dGTP, dan dTTP), primer (forward dan reverse), larutan penyangga 10X, Taq
polimerase (Invitrogen), MgCl2, DNA, dan alkohol 70%. Alat yang digunakan adalah pipet beserta ujung pipetnya ukuran 10, 20, 200 µl, rak, tabung PCR 200 µl, tabung 1.5 ml, tisu, rak pendingin sampel, alat mikrosentrifugasi (Eppendorf) tipe 5415C, penstabil voltase (Voltage Stabilizer Ferro Resonant 1500 ICA) dan
mesin PCR (Gene Amp PCR System 2400, Perkin Elmer).
Bahan dan Alat Elektroforesis
Bahan yang digunakan untuk elektroforesis adalah agarosa, loading dye,
penanda 100 pb, TBE 1X (1M Tris, 0.9 M Asam Borat dan 0.01 M EDTA pH 8.0) dan etidium bromida. Peralatan yang digunakan untuk elektroforesis adalah ujung pipet T-300 (axygen), mikropipet 10 P (Gilson), gelas ukur, timbangan
elektronik (AD HX 100), pengaduk bermagnet (magnetic stirrer MG78), pemanas
litrik, piranti Submarime Electrophoresis (Hoeffer USA), power supply
electrophoresis, alat foto UVi dan disket ukuran 3.5 inci.
Metode
Pengambilan Darah dan Data Fenotipe
Monyet dibius dengan campuran ketamin (10 mg/kg bobot badan) dan xylazine (2 mg/kg bobot badan). Obat bius disuntikkan secara intramuskuler dengan alat bantu berupa tulup atau senapan. Darah diambil dengan spuit dari vena femoralis sebanyak 5 ml dan selanjutnya dimasukkan ke dalam tabung
yang mengandung EDTA. Darah ini disimpan pada suhu -20oC yang selanjutnya
diekstrak untuk mendapatkan DNA. Tata cara pengambilan darah mengikuti prosedur yang telah disetujui Komisi Pengawasan Kesejahteraan dan Penggunaan Hewan Penelitian PSSP-IPB (ACUC No. 07-A004-IR).
(Angeloni et al. 2004; Kaufman et al. 2005). Monyet digolongkan gemuk (mengalami obesitas) bila IMT lebih besar atau sama dengan 30 kg/m2 (WHO 2005; Kaufman et al. 2005). Lokasi pengukuran lipatan kulit perut, lengan, paha,
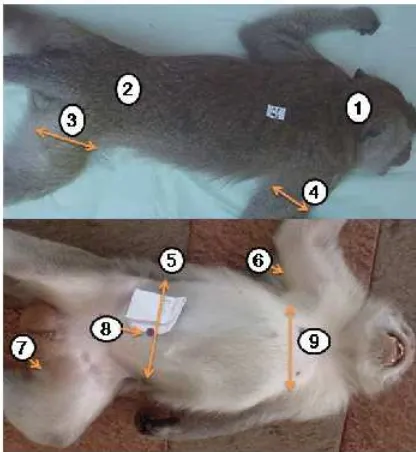
[image:44.612.183.391.276.502.2]leher dan punggung masing-masing adalah pusar, trisep, pangkal paha (selangkangan), leher bagian dorsal, dan lumbal. Tebal lipatan kulit diukur dengan skinfold lange caliper. Lingkar pinggang, dada, paha dan lengan atas (trisep) diukur dengan pita ukur. Lingkar pinggang, dada, paha dan lengan masing-masing diukur di daerah umbilikus, penonjolan bagian kaudal dari tulang sternum, pangkal paha, dan trisep (Gambar 9). Alat yang digunakan adalah timbangan (skala terkecil 0.1 kg), pita ukur dan kaliper (skala terkecil 1 mm).
Gambar 9 Lokasi pengukuran lingkar bagian tubuh dan tebal lipatan kulit
Keterangan : (1) lokasi pengukuran tebal lipatan kulit pada bagian leher dorsal, (2) lokasi pengukuran tebal lipatan kulit punggung, (3) lokasi pengukuran lingkar paha, (4) lokasi pengukuran lingkar lengan atas, (5) lokasi pengukuran lingkar pinggang, (6) lokasi pengukuran tebal lipatan kulit trisep, (7) lokasi pengukuran tebal lipatan kulit paha, (8) lokasi pengukuran tebal lipatan kulit perut, dan (9) lokasi pengukuran lingkar dada
Ekstraksi DNA dari Darah
Ekstraksi DNA menggunakan QIAamp DNA Blood Kits (Qiagen) dengan
yang selanjutnya dicampur dengan menggunakan vorteks selama 15 detik.
Campuran ini diinkubasi pada suhu 56oC selama 10 menit, kemudian
disetrifugasi beberapa saat. (2) Sebanyak 200 µl etanol (96-100%) ditambahkan pada sampel, dicampur menggunakan vorteks selama 15 detik, dan disentrifugasi beberapa saat. (3) Campuran ini dimasukkan ke dalam QiAamp spin column dan disetrifugasi dengan kecepatan 6 000 x g (8 000 rpm) selama satu menit setelah ditutup terlebih dahulu. Selanjutnya, QiAamp spin column ini
diletakkan di dalam tabung dua ml yang bersih, dan tabung yang mengandung filtrat dibuang. (4) Tutup spin column dibuka dengan hati-hati, dan 500 µl larutan
penyangga AW1 dimasukkan. QiAamp spin column ditutup kembali, dan
disentrifugasi dengan kecepata 6 000 x g (8 000 rpm) selama satu menit.
QiAamp spin column diletakkan di dalam tabung dua ml yang bersih, dan tabung
yang mengandung filtrat dibuang. (5) Sebanyak 500 µl larutan penyangga AW2
dimasukkan ke dalam QiAamp spin column, dan disentrifugasi dengan kecepatan
20 000 x g (14 000 rpm) selama tiga menit. (6) QiAamp spin column dimasukkan
ke dalam tabung 1.5 ml, ditambahkan 200 µl larutan penyangga AE, diinkubasi pada suhu ruangan (15-25oC) selama satu menit, dan disentrifugasi dengan kecepatan 6 000 x g (8 000 rpm) selama satu menit. Tabung terakhir mengandung DNA yang selanjutnya dapat digunakan untuk PCR.
Hasil ekstraksi dilihat dengan elektroforesis pada gel agarosa 1.2% dalam
larutan TBE 1x dalam piranti Submarine Eletrophoresis (Hoefer, USA). Fragmen
dimunculkan dengan pewarna etidium bromida setelah dimigrasikan selama 40 menit dengan voltase 85 V. Fragmen DNA yang terwarnai dilihat di bawah iluminasi ultraviolet. Konsentrasi DNA ditentukan dengan spektrofotometer. DNA dimasukkan ke dalam tabung 1.5 ml dan disimpan di dalam lemari pendingin dengan suhu -20oC untuk proses berikutnya.
Amplifikasi Gen MC4R
Amplifikasi daerah penyandi gen MC4R menggunakan primer forward (5’–
AATAACTGAGACGACTCCCTGAC–3’) dan reverse (5’–CAGAAGTACA
ATATTCAGGTAGGG–3’) (Yeo et al. 1998) dengan teknik PCR. Setiap unit
reaksi PCR mengandung ddH2O, larutan penyangga 1X, MgCl2 1.5 mM; dNTP
ditentukan dengan spektrofotometer dan diencerkan dengan ddH2O sehingga konsentrasi seluruh sampel sama dengan 25 ng/µl.
Setiap tabung untuk satu unit reaksi PCR (volume reaksi 50 µl) ditambahkan 5 l larutan penyangga 10x, 1.5 µl