EKSPLORASI MIKROBA RIZOSFER DAN HUBUNGANNYA
DENGAN SIFAT FISIK & KIMIA TANAH PADA KELAPA
SAWIT TERINFEKSI
Ganoderma boninense
Pat.
WIDYA YUNIVEN
DEPARTEMEN PROTEKSI TANAMAN
FAKULTAS PERTANIAN
INSTITUT PERTANIAN BOGOR
BOGOR
PERNYATAAN MENGENAI SKRIPSI DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA
Dengan ini saya menyatakan bahwa skripsi berjudul Eksplorasi Mikroba Rizosfer dan Hubungannya dengan Sifat Fisik & Kimia Tanah pada Tanaman Kelapa Sawit Terinfeksi Ganoderma boninense Pat. adalah benar karya saya dengan arahan dari dosen pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir skripsi ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, 2014
Widya Yuniven
ABSTRAK
WIDYA YUNIVEN. Eksplorasi Mikroba Rizosfer dan Hubungannya dengan Sifat Fisik & Kimia Tanah pada Kelapa Sawit Terinfeksi Ganoderma boninense
Pat.. Dibimbing oleh TITIEK SITI YULIANI.
Kelapa sawit (Elaeis guineensis Jacq.) merupakan salah satu tanaman perkebunan yang memiliki arti penting dalam menyumbangkan devisa bagi negara dari sektor non migas. Salah satu hambatan dalam pengembangan budidaya kelapa sawit adalah adanya penyakit busuk pangkal batang yang disebabkan oleh Ganoderma boninense. Penelitian ini bertujuan mengeksplorasi mikroba di sekitar rizosfer tanaman kelapa sawit dan hubungannya dengan kondisi sifat fisik dan kimia tanah. Penelitian dilakukan dengan mengambil contoh tanah dari tanaman kelapa sawit untuk dibuat suspensi. Suspensi dibuat seri pengenceran hingga tingkat pengenceran 10-8. Uji antagonisme in vitro antara kandidat antagonis dari golongan aktinomiset dengan G. boninense dilakukan dengan metode dual culture. Pengamatan dilakukan dengan mengukur jari-jari cendawan patogen yang menjauhi isolat aktinomiset dan jari-jari cendawan patogen yang mendekati isolat aktinomiset. Hasil isolasi mikroba diperoleh 76.9% isolat cendawan, 12.82% isolat bakteri, dan 10.26% isolat aktinomiset. Kandungan sifat kimia tanah pada lokasi penelitian yaitu pada pH 4.43 termasuk ke dalam jenis tanah agak masam dan kandungan C/N rasio 8.78% tergolong rendah. Hasil percobaan in vitro menunjukkan isolat AK1 (aktinomiset 1), AK2 (aktinomiset 2), dan AK3 (aktinomiset 3) pada hari kelima memberikan pengaruh yang nyata terhadap pertumbuhan cendawan patogen G. boninense pada media PDA.
ABSTRACT
WIDYA YUNIVEN. The Exploration of Rhizosphere Microbial and Its Relationship with Woil Physical and Chemical Characteristics of Oil Palm Infected by Ganoderma boninense Pat.. Supervised by TITIEK SITI YULIANI.
Oil palm (Elaeis guineensis Jacq.) is one of the plantation crops which has a significance in contributing foreign exchange to the country from non-oil sector. One of the fence in development oil palm cultivation is the presence of stem rot disease caused by Ganoderma boninense. This research aims to explore the microbial rhizosphere surrounding to the oil palm plants, as well as its relationship to the condition of physical and chemical soil characteristics. The study was conducted by taking a surrounding soil of oil palm plantations as a sample and made its suspension. The suspension was made to serial dilutions to 10-8 dilution rate. In vitro antagonism test between antagonist candidate of actinomycetes to G. boninense conducted by using a dual culture methods. Observations were made by measuring the radial growth of pathogenic fungi and actinomycetes isolates were away from fingers pathogenic fungi actinomycetes isolates were approaching. The results of microbial isolation consist of 76.9% fungal isolates, 12.82% bacterial isolates, and 10.26% actinomycetes isolates. The content of chemical characteristics soil at the study was sited at pH 4.43 it is included in a slightly acid soil type and content of C/N ratio 8.78 is low. The results of in vitro experiments showed that isolate AK1 (actinomycetes 1), AK2 (actinomycetes 2), and AK3 (actinomycetes 3) have a significant effect on the growth of pathogenic G. boninense fungi on PDA at the fifth day.
© Hak Cipta Milik IPB, Tahun 2014
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumberdaya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB
EKSPLORASI MIKROBA RIZOSFER DAN HUBUNGANNYA
DENGAN SIFAT FISIK & KIMIA TANAH PADA KELAPA
SAWIT TERINFEKSI
Ganoderma boninense
Pat.
WIDYA YUNIVEN
Skripsi
sebagai salah satu syarat untuk memperoleh gelar Sarjana Pertanian
pada
Departemen Proteksi Tanaman
DEPARTEMEN PROTEKSI TANAMAN
FAKULTAS PERTANIAN
INSTITUT PERTANIAN BOGOR
BOGOR
Judul Skripsi : Eksplorasi Mikroba Rizosfer dan Hubungannya dengan Sifat Fisik & Kimia Tanah pada Tanaman Kelapa Sawit Terinfeksi Ganoderma boninense Pat.
Nama Mahasiswa : Widya Yuniven
NIM : A34090038
Disetujui oleh
Dr. Ir. Titiek Siti Yuliani, SU Dosen Pembimbing
Diketahui oleh
Dr. Ir. Abdjad Asih Nawangsih, M.Si Ketua Departemen
PRAKATA
Puji syukur penulis panjatkan kehadirat Allah SWT yang telah melimpahkan rahmat-Nya, sehingga penulis dapat menyeleseikan penelitian yang berjudul “Eksplorasi Mikroba Rizosfer dan Hubungannya terhadap Sifat Fisik & Kimia Tanah pada Tanaman Kelapa Sawit Terinfeksi Ganoderma boninense Pat.”. Penelitian dilaksanakan di Laboratorium Mikologi Tumbuhan, Departemen Proteksi Tanaman, Institut Pertanian Bogor dari bulan Maret sampai Desember 2013.
Penulis menyampaikan ucapan terima kasih kepada Dr. Ir. Titiek Siti Yuliani, SU selaku dosen pembimbing yang telah banyak membimbing, memberikan ilmu dan pengarahan, serta dukungan moril yang sangat besar dalam penelitian penulis. Ucapan terima kasih juga kepada Dr. Ir. Ali Nurmansyah, Msi selaku dosen penguji tamu yang telah memberikan masukan, arahan dan saran yang bermanfaat kepada penulis, serta kepada PT Tidar Kerinci Agung, perkebunan kelapa sawit, Sumatera Barat atas izin dan kerjasamanya dalam pengambilan contoh tanah.
Penulis juga menyampaikan ucapan terima kasih kepada kedua orang tua, Guswarmen dan Yulisma yang tak henti-hentinya memberi perhatian dan bantuan moril, yang mana setiap langkah, gerak, dan ucapannya merupakan do’a bagi penulis, serta kedua adik penulis Meninda olavenia dan Fernando Septriandoni. Selain itu, ucapan terima kasih yang tulus kepada rekan kerja di Laboratorium Mikologi, Griya Pink dan Pondok Surya, atas dukungan dan semangat yang telah diberikan kepada penulis.
Penulis menyadari bahwa skripsi ini masih jauh dari kesempurnaan. Oleh karena itu, penulis mengharapkan kritik dan saran yang bersifat membangun untuk kedepannya. Semoga penelitian ini dapat memberikan manfaat bagi pembaca dan kalangan umum.
Bogor, Juni 2014
DAFTAR ISI
DAFTAR TABEL x
DAFTAR GAMBAR x
DAFTAR LAMPIRAN x
PENDAHULUAN 1
Latar Belakang 1
Tujuan Penelitian 2
Manfaat Penelitian 2
BAHAN DAN METODE 3
Waktu dan Tempat 3
Bahan dan Alat 3
Metode Penelitian 3
Pengambilan Sampel Tanah 3
Isolasi Mikroba Tanah 3
Analisis Sifat Fisik dan Kimia Tanah 4
Isolasi Ganoderma boninense 4
Uji Antagonisme in vitro 4
Rancangan Percobaan dan Analisis Data 5
HASIL DAN PEMBAHASAN 6
Gejala Tanaman Sakit di Lapangan 6
Isolasi Mikroba Rizosfer Kelapa Sawit 7
Hubungan Sifat Fisik & Kimia Tanah dengan Populasi Mikroba 9
Isolasi Patogen Ganoderma boninense 12
Uji Antagonis Aktinomiset terhadap G. boninense 13
SIMPULAN DAN SARAN 16
DAFTAR PUSTAKA 17
DAFTAR TABEL
1 Pearson Korelasi anatara jenis mikroba dengan sifat fisik & kimia 9 tanah
2 Persentase penghambatan isolat aktinomiset terhadap pertumbuhan 15 patogen G. boninense
DAFTAR GAMBAR
1 Gejala penyakit BPB : (a) Basidiokarp dewasa di bagian pangkal 6 batang, (b) 2-3 daun tombak tidak terbuka dan beberapa daun tua patah 2 Hasil isolasi mikroba 10 hsp dari rhizosfer tanaman kelapa sawit: (a) 7
cendawan. (b) bakteri. (c) aktinomiset
3 Rata-rata populasi cendawan pada lahan sehat dan terinfeksi 8 4 Rata-rata populasi bakteri pada lahan sehat dan terinfeksi 8 5 Rata-rata populasi aktinomiset pada lahan sehat dan terinfeksi 8 6 Hubungan sifat kimia tanah (pH) dengan populasi mikroba rizosfer 10 7 Hubungan sifat kimia tanah (C/N rasio) dengan populasi mikroba 10
rizosfer
8 Hubungan sifat fisik tanah (pasir) dengan populasi mikroba rizosfer 11 9 Hubungan sifat fisik tanah (debu) dengan populasi mikroba rizosfer 11 10 Hubungan sifat fisik tanah (liat) dengan populasi mikroba rizosfer 12 11 (a) Badan buah G. boninense, (b) Koloni G. boninense pada media 12
PDA
12 Isolat aktinomiset: (a) isolat AK1 berasal dari sampel berbukit sakit, 13 (b) AK2, (c) AK3, dan (d) AK4 berasal dari sampel berbukit sehat
13 Uji antagonisme dengan metode dual culture 14
DAFTAR LAMPIRAN
1 Rata-rata populasi cendawan, bakteri, dan aktinomiset pada lahan 20 berbukit
2 Rata-rata populasi cendawan, bakteri, dan aktinomiset pada lahan 20 datar
3 Rata-rata populasi cendawan, bakteri, dan aktinomiset pada 21 lahanbergelombang
4 Jari-jari pertumbuhan koloni patogen G. boniense pada media PDA 22 5 Persentase penghambatan isolat akttinomiset terhadap cendawan
PENDAHULUAN
Latar Belakang
Kelapa sawit (Elaeis guineensis Jacq.) merupakan salah satu tanaman perkebunan yang memiliki arti penting dalam menyumbangkan devisa bagi negara dari sektor non migas. Tanaman kelapa sawit mulai dibudidayakan secara komersial sejak akhir 1970an (Semangun 2000). Kebutuhan minyak dunia yang semakin meningkat menempatkan hasil bahan baku minyak kelapa sawit Crude Palm Oil (CPO) sebagai sumber energi terpenting untuk masa mendatang. Peningkatan kebutuhan tersebut tidak hanya disebabkan oleh penggunaan industri pangan, tetapi juga untuk industri non pangan seperti farmasi, kosmetik, hingga bahan bakar seperti diesel (Susanto 2002).
Perkembangan budidaya kelapa sawit di Indonesia pada beberapa tahun terakhir mengalami kemajuan, yang berkaitan dengan pertambahan luas areal maupun tingkat produksi. Salah satu faktor yang mempengaruhinya adalah peningkatan konsumsi minyak kelapa sawit yang semakin beragam. Luas areal pertanaman kelapa sawit di Indonesia pada tahun 2011 mencapai 5.34 juta ha dengan produksi minyak kelapa sawit sebesar 15.2 juta ton dan pada tahun 2012 meningkat menjadi 5.46 juta ha dengan produksi sebesar 15.42 juta ton (BPS 2012). Kelapa sawit juga berperan dalam memasok kebutuhan minyak dalam negeri dan dapat menghidupi sekitar 5 juta rakyat Indonesia (Susanto 2002). Peningkatan hasil telah diusahakan baik dengan cara intensifikasi maupun melalui perluasan areal perkebunan (Abadi 1987).
Salah satu masalah dalam pengembangan budidaya kelapa sawit adalah adanya kerusakan oleh patogen. Semangun (2000) dan True (1998) mengatakan
Ganoderma boninense, penyebab busuk pangkal batang (BPB) merupakan patogen yang paling merugikan. Sinaga et al. (2003) menjelaskan penyakit BPB merupakan ancaman bagi perkebunan kelapa sawit yang telah mengalami peremajaan, serta ancaman yang sangat besar terhadap produksi kelapa sawit. Patogen G. boninnense tidak hanya menyerang pada tanaman tua, tetapi keberadaan patogen dapat ditemukan pada tanaman belum menghasilkan (TMB). Kejadian penyakit pada tanaman belum menghasilkan (TBM) pada peremajaan satu, dua, tiga, dan empat masing-masing sebesar 0, 4, 7, dan 11%, sedangkan pada tanaman produktif dengan peremajaan satu, dua, dan tiga masing-masing sebesar 17, 18, dan 75% (Susanto et al. 2005). Tanaman kelapa sawit yang mengalami peremajaan hingga beberapa kali dapat meningkatkan persentase kejadian penyakit dan infeksi patogen dapat terlihat pada tanaman berumur 1 tahun. Serangan penyakit BPB sangat rendah pada lahan dengan peremajaan periode pertama dan meningkat pada periode kedua maupun ketiga seiring dengan seringnya peremajaan dilakukan. Menurut Yulianti (2001), kerusakan yang ditimbulkan G. boninense dapat mencapai 80% hingga 100%, bahkan menyebabkan kematian pada tanaman yang terinfeksi.
2
perkebunan lain seperti: kelapa, karet, teh, dan kakao, serta berbagai jenis tanaman berkayu (Arifin et al. 2000).
Usaha dalam mengendalikan penyakit BPB ini telah banyak dilakukan diantaranya: pengendalian kultur teknis, mekanis, dan kimiawi. Namun, belum ada yang berhasil menekan perkembangan penyakit BPB. Salah satu penyebabnya adalah G. boninense yang bersifat tular tanah (soil borne). Patogen tular tanah mempunyai kemampuan saprofitik yang tinggi dan parasit fakultatif dengan kisaran inang yang luas, memiliki beberapa struktur bertahan dalam kondisi lingkungan yang kurang mendukung perkembangannya: seperti miselium resisten, basidiospora, dan klamidiospora serta dapat bertahan lama di dalam tanah meskipun tidak ada inang (Arifin et al. 2000; Flood et al. 2000; Susanto et al. 2005).
Pengunaan fungisida kemungkinan tidak efektif karena sifat fisik dan kimia tanah akan terdegradasi oleh mikroba di dalam tanah sebelum mencapai sasaran. Sifat-sifat fisik tanah sangat mempengaruhi pertumbuhan dan produksi tanaman. Kondisi fisik tanah menentukan penetrasi akar di dalam tanah, retensi air, drainase, aerasi, dan nutrisi tanaman. Sifat fisik tanah juga mempengaruhi sifat-sifat kimia dan biologi tanah.
Dalam pengendalian penyakit tanaman diperlukan pengendalian yang bersifat terpadu. Salah satu alternatif pencegahan dan pengendalian yang dapat dilakukan adalah dengan pemanfaatan mikroba tanah pada rizosfer. Tanah memiliki potensi mikroba yang bersifat antagonis yang mampu menekan perkembangan patogen tular tanah dan sebagian besar hidup sebagai saprofit. Populasi mikroba di rizosfer biasanya lebih banyak dan beragam dibandingkan pada tanah yang bukan rizosfer (Lynch 1983).
Mikroba yang hidup pada daerah rizosfer biasanya digunakan sebagai agens pengendalian hayati. Menurut Julyanda (2011) rizosfer merupakan daerah yang ideal bagi tumbuh dan berkembangnya mikroba tanah, termasuk di dalamnya agens pengendalian hayati. Kehadiran sejumlah populasi mikroba dapat mempengaruhi pertumbuhan tanaman karena dapat meningkatkan produktivitas tanaman dan mempertahankan kesuburan tanah. Penggunaan mikroba antagonis sebagai agens pengendali biologi dapat menekan perkembangan penyebaran
Ganoderma (Abadi 1987). Oleh sebab itu, perlu dilakukan studi untuk melihat keragaman mikroba tanah yang diduga antagonis terhadap penyakit BPB kelapa sawit, mekanisme antagonisme serta uji keefektifan agens antagonis terhadap patogen G. boninense.
Tujuan Penelitian
Penelitian ini bertujuan mengeksplorasi mikroba di sekitar rizosfer tanaman kelapa sawit sehat dan sakit, serta hubungannya dengan kondisi sifat fisik dan kimia tanah.
Manfaat Penelitian
BAHAN DAN METODE
Waktu dan Tempat
Penelitian dilakukan dari bulan Maret hingga Desember 2013 di Laboratorium Mikologi, Departemen Proteksi Tanaman, Fakultas Pertanian, Institut Pertanian Bogor. Pengambilan sampel tanah dilakukan di tanaman kelapa sawit PT. Tidar Kerinci Agung, Dharmasraya, Sumatera Barat.
Bahan dan Alat
Bahan dan alat yang digunakan dalam penelitian ini adalah sampel tanah yang berasal dari rizosfer tanaman kelapa sawit terinfeksi G. boninense dan sehat, Media Martin Agar (MA), media Potato Dextrose Agar (PDA), media Yeast Casamino-Acid Exstract Dextrose Agar (YCED), media Nutrient Agar (NA), kloramfenicol, streptomisin, alkohol 70%, khlorox, linggis, parang, cawan petri, labu erlenmeyer, tabung reaksi, Laminar air flow, dan digital camera.
Metode Penelitian
Pengambilan Sampel Tanah
Kegiatan pertama yang dilakukan adalah menentukan blok berdasarkan topografi lahan berbeda yaitu datar, berbukit, dan bergelombang. Satu blok diambil 6 titik contoh yang terdiri dari 3 titik merupakan tanaman terinfeksi: datar terinfeksi (TSD), berbukit terinfeksi (TSB), bergelombang terinfeksi (TSG) dan 3 titik tanaman sehat: datar sehat (SD), berbukit sehat (SB), bergelombang sehat (SG), sehingga dibutuhkan 18 sampel tanah dari rizosfer tanaman kelapa sawit.
Tanah diambil disekitar rizosfer kelapa sawit sebanyak 1 kg hingga kedalaman 25 cm dengan menggunakan parang. Untuk membedakan tanaman sakit dengan tanaman sehat dilihat dari penampakan gejala luar. Gejala tanaman kelapa sawit sakit oleh G. boninense menunjukkan empat atau lima daun tombak yang tidak membuka dan berwarna hijau pucat, lilit batang pada pangkal daun terlihat mengecil, dan ukuran daun yang semakin sedikit dengan ukuran buah semakin kecil.
Isolasi Mikroba Tanah
4
ulangan. Hasil biakan dari ketiga media tersebut diamati selama 7 hari setelah pencawanan (hsp) untuk golongan cendawan dan bakteri serta 10 hsp untuk golongan aktinomiset (Saraswati et al. 2007). Setiap pertumbuhan mikroba dicatat, dan koloni yang tumbuh dikelompokkan berdasarkan bentuk dan warna koloni, kemudian dimurnikan.
Analisis Sifat Fisik dan Kimia Tanah
Analisis sifat fisik dan kimia tanah bertujuan mengetahui hubungannya dengan populasi mikroba rizosfer tanaman kelapa sawit. Variabel yang diamati adalah populasi mikroba, tekstur tanah (pasir, liat, debu), pH tanah, dan nisbah C/N rasio. Untuk analisis sifat fisik tanah dilakukan menggunakan metode tekstur pipet, selanjutnya kadar masing-masing fraksi ditentukan dalam % liat, % debu, dan % pasir. Penentuan kadungan pH tanah diuji menggunakan campuran air (H2O) dan KCl, sedangkan kandungan bahan organik yaitu: kandungan C berdasarkan metode Walkley & Black dan kandungan N menggunakan metode Kjedahl (BPT 2013). Nilai C dan N yang diperoleh dapat dihitung nila C/N rasio.
Isolasi Ganoderma boninense
Isolat G. boninense yang digunakan pada penelitian ini berasal dari isolasi tubuh buah G. boninense asal perkebunan kelapa sawit PT. Tidar Kerinci Agung, Sumatera Barat. Tubuh buah didisinfeksi menggunakan alkohol 70%, kemudian dipotong-potong berukuran 5 x 5 mm dan ditumbuhkan pada media PDA. Miselium yang tumbuh dimurnikan.
Uji Antagonisme in vitro
Uji antagonisme secara in vitro hanya dilakukan pada isolat aktinomiset, karena pada penelitian sebelumnya telah banyak menggunakan kelompok cendawan dan bakteri dalam uji antagonisme terhadap patogen lain. Pengujian dilakukan pada media PDA menggunakan metode uji ganda (dual culture) antara kandidat aktinomiset dengan G. boninense. Cendawan patogen G. boninense
5
Besarnya pengaruh penghambatan agens antagonis terhadap patogen dihitung menggunakan rumus persentase (Fokkema 1973):
Rancangan Percobaan dan Analisis Data
Rancangan percobaan yang digunakan pada uji penghambatan in vitro yaitu Rancangan Acak Lengkap (RAL). dengan 5 ulangan. Data yang diperoleh dianalisis dengan Microsoft Office Excel 2007 dan dianalisis sidik ragam (ANOVA) menggunakan program Statistical Office System (SAS) versi 9.1.3. Perlakuan yang berpengaruh nyata diuji lanjut dengan uji Duncan dengan taraf
nyata α = 5% (Mattjik & Sumertajaya 2006). Untuk mengetahui hubungan antara
jumlah populasi mikroba dengan sifat fisik kimia tanah diuji dengan Korelasi Pearson. Hasil yang diperoleh ditampilkan dalam bentuk grafik dan tabel.
(R1 – R2)
I = x 100%
R1
3 cm P A 3 cm
R1 R2
Keterangan :
P : Koloni candawan patogen G. boninense
A : Koloni aktinomiset kandidat antagonis
R1 : Jari-jari koloni G. boninense
menjauhi kandidat antagonis
R2 : Jari-jari G. boninense mendekati kandidat antagonis
Keterangan:
I : Persentase penghambatan kandidat antagonis (%)
R1 : Jari-jari koloni G. boninense
menjauhi kandidat antagonis
R2 : Jari-jari koloni G. boninense
HASIL DAN PEMBAHASAN
Gejala Tanaman Sakit di Lapangan
Gejala dini penyakit ini sukar untuk dideteksi karena perkembangan penyakit sangat lambat. Gejala akan terlihat apabila sudah gejala lanjut atau membentuk tubuh buah. Pada tanaman yang terinfeksi belum tentu ditemukan tubuh buah G. boninense dibagian pangkal batang, namun dapat dilihat pada daun tombak yang tidak terbuka sekitar 2 – 3 daun. Ukuran daun yang tidak terbuka lebih kecil daripada daun normal dan mengalami nekrotik di bagian ujung. Daun tua yang mengalami nekrotik selanjutnya patah dan akan tetap menggantung pada pohon. Pada akhirnya tanaman akan mati dan tumbang. Gejala yang tampak pada daun menunjukkan bahwa penampang pangkal batang mengalami pembusukan sebesar 50% atau lebih (Risanda 2008).
Gejala 1 Gejala penyakit BPB: (a) Basidiokarp dewasa di bagian pangkal batang, (b) 2-3 daun tombak tidak membuka dan beberapa daun tua patah
Basidiokarp yang dibentuk awalnya berukuran kecil, bulat, dan berwarna putih. Pada pertumbuhan yang cepat, basidiokarp dewasa memiliki bentuk, ukuran, dan warna yang variatif. Umumnya basidiokarp akan mengelilingi bagianpangkal batang yang sakit. Semakin besar ukuran basidiokarp menunjukkan perkembangan penyakit semakin lanjut dan akhirnya menyebabkan kematian pada tanaman (Arifin et al. 2000).
a
7
Isolasi Mikroba Rizosfer Kelapa Sawit
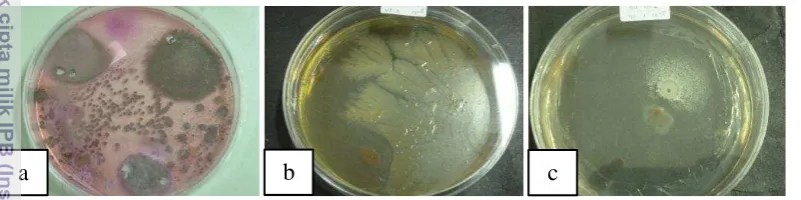
Analisis populasi mikroba pada rizosfer kelapa sawit sehat dan terinfeksi G. boninense diperoleh persentase isolat mikroba sebesar 76.9% isolat cendawan, 12.82% isolat bakteri, dan 10.26% isolat aktinomiset. Pertumbuhan masing-masing koloni mikroba berbeda. Pertumbuhan koloni cendawan mulai terlihat pada 2 hsp, namun masih sulit membedakan secara morfologi. Saat berumur lebih dari 3 hsp koloni cendawan yang tumbuh sudah dapat dibedakan antar koloni. Untuk golongan bakteri 2 hsp koloni mulai terlihat, sedangkan golongan aktinomiset pertumbuhanya mulai terlihat saat berumur di atas 10 hsp, karena mikroba golongan ini termasuk ke dalam mikroba yang pertumbuhannya lambat dibandingkan dengan mikroba lain.
Gambar 2 Hasil isolasi mikroba 10 hsp dari rhizosfer tanaman kelapa sawit: (a) cendawan, (b) bakteri, (c) aktinomiset
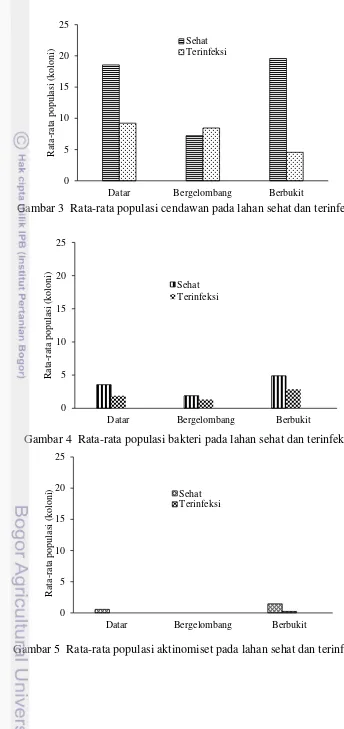
Hasil penjumlahan mikroba di ketiga lokasi menunjukkan bahwa rata-rata populasi yang diperoleh pada lahan sehat lebih tinggi dibandingkan lahan terinfeksi, namun hal ini tidak sesuai dengan analisis secara statistik. Rata-rata populasi cendawan lebih tinggi pada lahan sehat dibandingkan lahan terinfeksi hanya pada lokasi berbukit (t=6.79,P=0.000), sedangkan lokasi datar
(t=1.46,P=0.08) dan bergelombang (t=2.06,P=0.06) rata-rata populasi cendawan pada lahan terinfeksi lebih tinggi daripada lahan sehat (gambar 3). Hal ini diduga, kelimpahan populasi cendawan yang lebih mendominasi berasal dari kelompok mikroba bersifat sebagai patogen sehingga pertumbuhan mikroba nonpatogen menurun akibat adanya persaingan dalam perebutan ruang tumbuh.
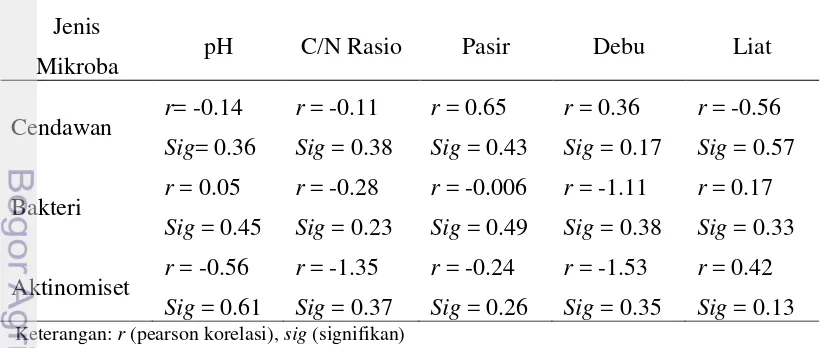
Gambar 4 menunjukkan rata-rata populasi bakteri tertinggi pada lahan berbukit sehat (SB) sebesar 4.89 koloni dan terendah sebesar 1.33 koloni pada lahan bergelombang teinfeksi (TSG). Secara statistik populasi bakteri pada lahan sehat lebih kecil daripada lahan terinfeksi berada pada lahan berbukit
(t=1.09,P=0.15) dan bergelombang (t=1.66,P=0.06). Untuk golongan aktinomiset, rata-rata populasi tertinggi terdapat pada lahan SB (berbukit sehat) diikuti SD (datar sehat). Masing-masing memiliki nilai sebesar 1.44 koloni dan 0.5 koloni, sedangkan nilai terendah sebesar 0.2 koloni pada lahan TSB dan tidak ditemukan keberadaan aktinomiset pada lahan bergelombang (gambar 5). Hal ini sesuai dengan uji statistik yang menunujukkan bahwa rata-rata populasi aktinomiset lahan sehat lebih tinggi dibandingkan lahan terinfeksi pada lokasi datar (t=1.89,P=0.04) dan berbukit (t=2.33, P=0.02).
c
8
Gambar 3 Rata-rata populasi cendawan pada lahan sehat dan terinfeksi
Gambar 4 Rata-rata populasi bakteri pada lahan sehat dan terinfeksi
9
Hal ini berhubungan dengan ketersediaan nutrisi atau eksudat akar pada tanaman yang mempengaruhi kehidupan mikroba di sekitar rizosfer kelapa sawit. Eksudat tanaman dapat merangsang aktivitas mikroba dan komposisi utama eksudat akar adalah gula dan asam amino digunakan oleh mikroba untuk melaksanakan aktivitas biologinya (Yuliani 1988). Selain itu, kehadiran sejumlah populasi mikroba baik yang bersifat antagonis maupun saprofit dapat menambah keragaman spesies di dalam komunitas alami tanaman (Jeger 2001)
Berdasarkan hasil analisis kimia tanah menunjukkan bahwa rendahnya populasi bakteri dan aktinomiset disebabkan kondisi tanah pada keadaan asam, sedangkan bakteri dan aktinomiset dapat tumbuh pada kondisi basa. Menurut Handayanto dan Hairah (2007) mengatakan, pada pH kurang dari 5 populasi aktinomiset kurang dari 1% dari populasi mikroba lainnya, sehingga aktinomiset tidak toleran pada kondisi asam, tetapi toleran pada kondisi basa.
Hubungan Sifat Fisik & Kimia Tanah dengan Populasi Mikroba
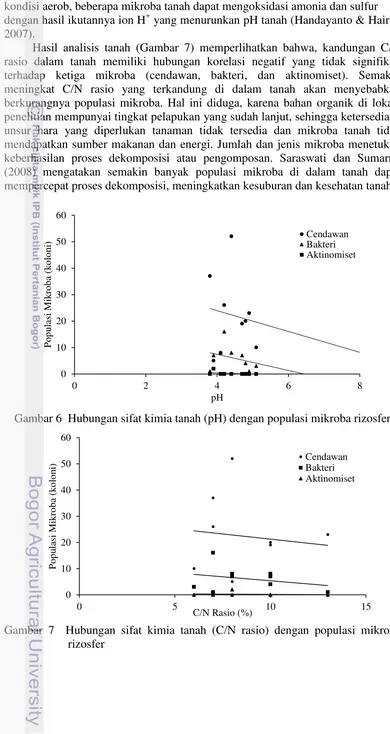
Hasil analisis kimia tanah menunjukkan pH tanah di lokasi penelitian sebesar 4.43 termasuk ke dalam jenis tanah agak masam. Pada gambar 6 menunjukkan keeratan hubungan antara populasi mikroba terhadap sifat kimia tanah. pH tanah memiliki hubungan korelasi negatif dan derajat hubungan yang cukup kuat dengan populasi bakteri dan aktinomiset dengan masing-masing koefisien sebesar -0.3, dan -0.43 (Tabel 1), namun tidak memiliki hubungan yang signifikan. Semakin meningkat pH tanah maka akan terjadi penurunan populasi mikorba rizosfer. Untuk golongan cendawan tidak memiliki hubungan korelasi terhadap pH tanah. Hal ini diduga bahwa, cendawan antagonis banyak terdapat di tanah dalam kondisi yang memungkinkan untuk tumbuh. Baker dan Cook (1974) melaporkan, aktivitas cendawan antagonis hanya terpacu pada kondisi masam, sedangkan pH tanah berada pada konsidi agak masam.
Tabel 1 Pearson Korelasi antara jenis mikroba dengan sifat fisik & kimia tanah
Jenis Keterangan: r (pearson korelasi), sig (signifikan)
10
kondisi anerob, beberapa mikroba tanah menghasilkan asam-asam organik, pada kondisi aerob, beberapa mikroba tanah dapat mengoksidasi amonia dan sulfur dengan hasil ikutannya ion H+ yang menurunkan pH tanah (Handayanto & Hairah 2007).
Hasil analisis tanah (Gambar 7) memperlihatkan bahwa, kandungan C/N rasio dalam tanah memiliki hubungan korelasi negatif yang tidak signifikan terhadap ketiga mikroba (cendawan, bakteri, dan aktinomiset). Semakin meningkat C/N rasio yang terkandung di dalam tanah akan menyebabkan berkurangnya populasi mikroba. Hal ini diduga, karena bahan organik di lokasi penelitian mempunyai tingkat pelapukan yang sudah lanjut, sehingga ketersediaan unsur hara yang diperlukan tanaman tidak tersedia dan mikroba tanah tidak mendapatkan sumber makanan dan energi. Jumlah dan jenis mikroba menetukan keberhasilan proses dekomposisi atau pengomposan. Saraswati dan Sumarno (2008) mengatakan semakin banyak populasi mikroba di dalam tanah dapat mempercepat proses dekomposisi, meningkatkan kesuburan dan kesehatan tanah.
Gambar 6 Hubungan sifat kimia tanah (pH) dengan populasi mikroba rizosfer
11
Berdasarkan analisis tanah, kandungan C/N rasio di lokasi termasuk dalam kategori rendah yaitu sebesar 8.8%. Sesuai dengan jenis sampel tanah yang digunakan berasal dari tanah podsolik merah kuning dengan ciri-ciri: tekstur lempung, pH rendah, KTK rendah, dan kandungan bahan organik rendah (Indrayatie 2009).
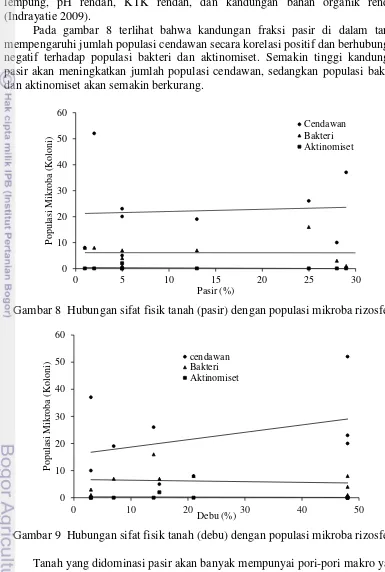
Pada gambar 8 terlihat bahwa kandungan fraksi pasir di dalam tanah mempengaruhi jumlah populasi cendawan secara korelasi positif dan berhubungan negatif terhadap populasi bakteri dan aktinomiset. Semakin tinggi kandungan pasir akan meningkatkan jumlah populasi cendawan, sedangkan populasi bakteri dan aktinomiset akan semakin berkurang.
Gambar 8 Hubungan sifat fisik tanah (pasir) dengan populasi mikroba rizosfer
Gambar 9 Hubungan sifat fisik tanah (debu) dengan populasi mikroba rizosfer
Tanah yang didominasi pasir akan banyak mempunyai pori-pori makro yang disebut lebih poreus. Semakin poreus tanah, maka akar akan mudah untuk berpenetrasi, serta air dan udara akan semakin mudah untuk bersirkulasi sehingga kandungan oksigennya tinggi dan mikroba akan lebih tinggi populasinya karena kondisinya yang aerob. Kandungan fraksi debu memiliki hubungan korelasi
12
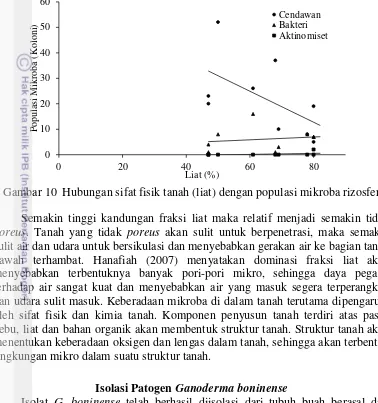
positif dengan populasi cendawan dan berkorelasi negatif dengan populasi aktinomiset. Kandungan fraksi liat memiliki hubungan korelasi negatif yang tidak signifikan dengan populasi cendawan, berkorelasi positif dengan populasi aktinomiset dan bakteri (Tabel 1, Gambar 9).
Gambar 10 Hubungan sifat fisik tanah (liat) dengan populasi mikroba rizosfer
Semakin tinggi kandungan fraksi liat maka relatif menjadi semakin tidak
poreus. Tanah yang tidak poreus akan sulit untuk berpenetrasi, maka semakin sulit air dan udara untuk bersikulasi dan menyebabkan gerakan air ke bagian tanah bawah terhambat. Hanafiah (2007) menyatakan dominasi fraksi liat akan menyebabkan terbentuknya banyak pori-pori mikro, sehingga daya pegang terhadap air sangat kuat dan menyebabkan air yang masuk segera terperangkap dan udara sulit masuk. Keberadaan mikroba di dalam tanah terutama dipengaruhi oleh sifat fisik dan kimia tanah. Komponen penyusun tanah terdiri atas pasir, debu, liat dan bahan organik akan membentuk struktur tanah. Struktur tanah akan menentukan keberadaan oksigen dan lengas dalam tanah, sehingga akan terbentuk lingkungan mikro dalam suatu struktur tanah.
13
G. boninense memiliki morfologi dan ukuran basidiokarp yang variatif tergantung kondisi lingkungan selama pembentukan basidiokarp (Seo & Kirk 2000). Basidiokarp yang diisolasi (Gambar 11a) berbentuk seperti piringan menyerupai kipas tebal mempunyai bentuk yang konsentris. Bagian atas permukaannya berwarna coklat tua, tampak mengkilap, dan kurang rata. Permukaan bawah berwarna putih pucat, memiliki lapisan pori yang merupakan tempat pembentukan basidium dan bisidiospora.
Pengamatan makroskopis G. boninense pada media agar hasil isolasi dari badan buah (Gambar 11b) memiliki miselium berwarna putih seperti kapas. Pertumbuhan miselium termasuk lambat karena pada umur 3 hsi telihat miselium seperti benang-benang tipis yang tumbuh di atas permukaan media agar. Pada umur 7 hsi miselium dapat tumbuh menutupi seluruh permukaan media. Karakteristik G. boninense dilihat secara mikroskopis berupa hifa bersekat, konidia tunggal, dan memiliki clamp connection.
Uji Antagonis Aktinomiset terhadap G. boninense
Aktinomiset yang berhasil diisolasi dari rizosfer kelapa sawit sebanyak 4 isolat. Keempat isolat digunakan dalam pengujian in vitro. Setiap isolat memiliki morfologi warna yang bervariasi (Gambar 12). Sehingga dalam membedakan antar isolat dilihat dari perbedaan warna koloni. Pengamatan warna koloni berdasarkan warna miselium aerial di media YCED. Isolat AK1 berwarna abu-abu berasal dari sampel terinfeksi, isolat AK2 berwarna putih bagian tengah koloni sedangkan tepian berwarna abu-abu, isolat AK3 berwarna cokelat dan menghasilkan pigmen kuning pada media agar, isolat AK4 berwarna putih pekat.
Menurut Susilowati et al. (2007) hifa awalnya putih, ketika pembentukan spora berubah warna. Selanjutnya koloni tampak memiliki serbuk di permukaannya. Hal ini yang membedakan koloni bakteri pada umumnya.
14
Aktinomiset merupakan bakteri yang berfilamen sehingga tampak menyerupai cendawan. Bakteri ini memiliki dua macam miselium yaitu miselium substrat dan aerial. Ciri khas dari bakteri ini berupa penampakan koloni yang berdebu (kumpulan spora), tekstur seperti beludru, dan kering. Aktinomiset dapat dibedakan dengan bakteri lain apabila dilihat dari bentuk koloni di medium padat. Koloni aktinomiset keras dan seperti ada tumbuh akar di dalam media agar, sedangkan koloni bakteri lain lunak di atas media agar (Miyado 2003). Selain itu, pertumbuhan aktinomiset termasuk lambat. Miselium aktinomiset mulai terbentuk pada permukaan media agar setelah 24 jam (Holt et al. 1994).
Koloni aktinomiset yang diperoleh dari hasil isolasi menghasilkan aroma tanah yang khas. Aroma tanah tersebut merupakan senyawa goesmin pada koloni aktinomiset (Wilkins & Scholler 2009). Beberapa genus aktinomiset yang dapat ditemukan dalam tanah perakaran yaitu Micromonospora, Nocardia, Streptomyces, Streptosporangium, dan Thermoactinomycetes (Premono et al.
1992).
Kriteria pemilihan agens antagonis berdasarkan besarnya daya penghambatan terhadap G. boninense (Susanto 2000). Pengujian dilakukan selama lima hari dengan mengukur jari-jari pertumbuhan patogen, dimana R1 merupakan jari-jari patogen menjauhi agens antagonis dan R2 merupakan jari-jari patogen mendekati agens antagonis (Gambar 13).
Gambar 13 Uji antagonisme dengan metode dual culture
Pengamatan persentase daya penghambatan calon antagonis menunjukkan zona hambat seperti yang diungkapkan oleh Maria (2002) bahwa kriteria keefektifan uji antagonisme secara in vitro dilihat dari terbentuknya atau tidak zona hambatan yaitu zona bening diantara patogen dan calon agens antagonis.
Hasil pengamatan dalam uji in vitro (Tabel 2) menunjukkan isolat AK1, AK2, AK3 dan AK4 mempunyai persentase daya hambat pada hari kelima sebesar 42.17%, 33.56%, 25.9%, dan 17.01%. Keempat isolat tersebut mampu menekan pertumbuhan hifa G. boninense. Hal ini ditunjukkan dengan adanya zona hambatan antara aktinomiset dengan patogen. Zona hambat terjadi karena aktinomiset mempunyai kemampuan untuk mengeluarkan zat antimikrobial atau yang lebih dikenal sebagai antibiotik (Sariyanto 2006).
15
Tabel 2 Persentase penghambatan isolat aktinomiset terhadap pertumbuhan patogen G. boninense
1
Hari setelah inokulasi. aAngka yang diikuti huruf yang sama pada baris yang sama menunjukkan hasil yang tidak berbeda nyata berdasarkan uji selang Duncan pada taraf α=5%
Pengamatan hari kelima menunjukkan bahwa isolat AK1, AK2, dan AK3 memberikan pengaruh yang berbeda nyata terhadap kontrol, sedangkan isolat AK4 tidak memberikan pengaruh terhadap kontrol. Isolat AK1 memiliki persentase penghambatan yang semakin meningkat dari hari pertama sampai kelima dan memberikan pengaruh terhadap pertumbuhan G. boninense, sedangkan isolat AK2, AK3, dan AK4 persentase penghambatan mengalami penurunan. Ketiga isolat tersebut mampu menghasilkan antibiotik pada hari pertama, sedangkan hari berikutnya antibiotik yang dihasilkan mulai berkurang. Hal ini disebabkan media tumbuh dalam pengujian kaya akan nutrisi, sedangkan aktinomiset dapat tumbuh dengan baik pada media miskin nutrisi. Sariyanto (2006) mengatakan, kelemahan dari aktinomiset adalah pertumbuhan dan produksi dari antibiotiknya sangat lambat pada media agar. Hal ini diduga akan mempengaruhi kemampuan aktinomiset dalam memproduksi antibiotik.
Mekanisme penghambatan Streptomyces spp. terhadap fungi dapat terjadi karena kemampuannya dalam menghasilkan antibiotik dan senyawa hidrolitik seperti glukanase, kitinase yang mampu mendegradasi dinding sel fungi. Senyawa antibiotik ini sangat baik sebagai agens pengendali hayati penyakit tanaman tular tanah (soil borne) yang dapat dikendalikan dengan Sreptomyces scabies dan S. ipomoaea (Alexander 1978).
Aktivitas penghambatan senyawa antimikroba secara umum dapat dilakukan dengan berbagai mekanisme, diantaranya: (1) merusak dinding sel dengan cara menghambat pembentukan maupun merubah setelah terbentuk; (2) perubahan permeabilitas sel yang mengakibatkan terhambatnya pertumbuhan sel atau matinya sel; (3) perubahan molekul protein dan asam nukleat; (4) menghambat kerjanya enzim yang mengakibatkan terganggunya metabolisme sel; (5) penghambatan sintesa asam nukleat dan protein yang berakibat terganggunya aktivitas metabolisme karena DNA, RNA, dan protein memegang peranan penting dalam mekanisme sel secara normal (Pelczar & Chain 1986).
SIMPULAN DAN SARAN
Jenis mikroba yang mendominasi di sekitar rizosfer kelapa sawit adalah cendawan (76.9%), bakteri (12.82%), dan aktinomiset (10.26%). pH tanah di lokasi penelitian sebesar 4.43 termasuk ke dalam jenis tanah agak masam dan kandungan C/N rasio di lokasi termasuk dalam kategori rendah yaitu sebesar 8.8%. Hasil analisis tanah menunjukkan bahwa, sifat fisik dan kimia tidak memiliki korelasi yang signifikan dengan populasi mikroba rizosfer. Hasil percobaan in vitro menunjukkan isolat AK1, AK2, dan AK3 pada hari kelima memberikan pengaruh yang nyata terhadap pertumbuhan cendawan patogen G. boninense pada media PDA.
DAFTAR PUSTAKA
Abadi AL. 1987. Biologi Ganoderma boninense Pat. pada kelapa sawit (Elaeis guineensis Jacq.) dan pengaruh beberapa mikroba tanah antagonistik terhadap pertumbuhannya. [disertasi]. Bogor (ID): Institut Pertanian Bogor. Alexander M. 1978. Introduction to Soil Microbiology. Second Edition. New
Delhi (IN): Willey Estern Limited.
Ariffin D, Idris AS, Singh G. 2000. Status of Ganoderma in oil palm. Di dalam: Flood J, Bridge PD, Holderners M. editor, Ganoderma Disease of Perenial Crops. London (GB): CABI Publishing. hlm 49-65.
Baker KF, Cook RJ. 1974. Biological Control of Plant Pathogens. San Diego (US). W.H. Freeman and Company.
[BPS] Badan Pusat Statistika. 2012. Luas tanaman peekebunan besar menurut jenis tanaman [Internet]. Jakarta (ID): BPS; [diunduh 2013 April 12]. Tersedia pada: http://bps.go.id.
[Balittanah] Balai Penelitian Tanah, Kementrian Pertanian. 2013. Laporan hasil pengujian analisis contoh tanah. Bogor (ID): Badan Penelitian dan Pengembangan Pertanian Pertanian.
Crawford DL, Lynch JM, Whipps JM, Ousley MA. 1993. Isolation and characterization of actinomycete antagonists of a fungal root pathogen. Appl Environ Microbiol. 59(11): 3899-3905.
Fokkema NJ. 1973. The role of saprophytic fungi in antagonism againts
Drechslera sorokiniana (Helminthosporium sativum) on agar plates and on rye leaves with pollen. Physiological Plant Pathology. 3(2): 195-205.
Flood J, Bridge PD, Holdernes M. 2000. Ganoderma Disease of Perennial Crops. London (GB): CABI Publishing.
Handayanto E, Hairah K. 2007. Biologi Tanah Landasan Pengelolaan Tanah Sehat. Yogyakarta (ID): Pustaka Adipura.
Hanafiah KA. 2007. Dasar-Dasar Ilmu Tanah. Jakarta (ID): PT. Raja Grafindo Persada.
Holt JG et al. 1994. Bergeys Manual of Determinative Bacteriology. Ed ke-9. Philadelphia (BB): A Wolters Kluwers Company.
Jeger MJ. 2001. Biotic Interaction and Plant-pathogen Association. New York (US): CABI Publishing.
Lynch JM. 1983. Soil Biotechnology: Microbiological Factors in Crop Productivity. London (GB): Blackwell Scientific Publication.
Indriyatie ER. 2009. Distribusi pori tanah podsolik merah kuning pada berbagai kepadatan tanah dan pemberian bahan organik. Jurnal Hutan Tropis Borneo. 10(27): 230-236.
Julyanda M. 2011. Keragaman dan kelimpahan cendawan pada rizosfer kelapa sawit sehat dan terserang Ganoderma boninense [skripsi]. Bogor (ID): Institut Pertanian Bogor.
Maria PD. 2002. Eksplorasi dan uji antagonisme bakteri rhizosfer tanah dan endofit akar untuk pengendalian penyakit layu (Fusarium oxysporum
18
Mattjik AA, Sumertajaya M. 2006. Perancangan Percobaan dengan Aplikasi SAS dan Minitab. Bogor (ID): Institut Pertanian Bogor Press.
Miyado. 2003. Prosedur karakterisasi dan identifikasi. Di dalam: Puspita L, editor. Training Course on Identificatoin of Bacteria. Bogor (ID): hlm 1-5.
Pelczar MJ, Chain ECS. 2005. Dasar-Dasar Mikrobiologi Volume ke-1. Di dalam: Hadioetomo RS, Imas T, Tjitrosomo SS, Angka SL, editor. Elements of Microbiology. Jakarta (ID): UI-Press.
Paterson RRM. 2007. Ganoderma disease of oil palm-a white rot perspective necessary for integrated control. Braga (PT): Crop Protection. 26(1): 1369-1376.
Risanda D. 2008. Pengembangan teknik inokulasi buatan Ganoderma boninense Pat. Pada bibit kelapa sawit [skripsi]. Bogor (ID): Institut Pertanian Bogor. Saraswati R, Husen E, Simanungkalit RDM. 2007. Metode Analisis Biologi
Tanah. Bogor (ID): Balai Besar Penelitian dan Pengembangan Sumberdaya Lahan Pertanian.
Saraswati R, Sumarno. 2008. Pemanfaatan mikroba penyubur tanah. Iptek Tanaman Pangan 3(1): 41-48.
Sariyanto N. 2006. Eksplorasi agens antagonis yang berpotensi menekan penyakit layu fusarium pada pisang [sripsi]. Bogor (ID): Institut Pertanian Bogor. Semangun H. 2000. Penyakit-Penyakit Tanaman Perkebunan di Indonesia.
Yogyakarta (ID): Gadjah Mada University Press.
Seo GS, Kirk PM. 2000. Ganodermataceae: nomenclature and classification. Di dalam: J Flood, Bridge PD, Holderness M, editor. Ganoderma Diseases of Perrennial Crops. London(GB): CABI Publishing. hlm 3-22.
Sinaga MS, Bonny PWS, Susanto A. 2003. Keragaman mikroorganisme rhizosfer kelapa sawit dan patogenisitas Ganoderma boninense Pat. sebagai dasar pengendalian penyakit busuk pangkal batang. [laporan akhir Hibah Bersaing IX]. Bogor (ID): Institut Pertanian Bogor.
Susanto A. 2002. Kajian pengendalian hayati Ganoderma boninense Pat. Penyebab penyakit busuk pangkal batang kelapa sawit [disertasi]. Bogor (ID): Institut Pertanian Bogor.
Susanto A, Sudharto PS, Purba RY. 2005. Enhancing biological control of basal stem root disease (Ganoderma boninense) in oil palm plantations.
Mycopathologia. 159(1): 153-157.
Tan KH. 1992. Dasar-Dasar Kimia Tanah. Yogyakarta (ID): Gajah Mada University Press.
True R. 1998. Macro Fungi in Oil Palm Plantations of South East Asia. Journal Mycologist. 12 (1): 10-14.
Wilkins K, Scholler C. 2009. Volatile organic metabolites from selected
Streptomyces strains. Actinomycotalogica. 23(2): 27-33.
Yuliani TS. 1988. Uji antagonisme mikrobia rizosfer pada tanaman lada tehadap
Phytophthora palmivora. [disertasi]. Yogyakarta (ID): Universitas Gadjah Mada.
Yulianti S. 2001. Pemanfaatan mikoriza arbuskular dan Gliocladium finbratum
20
Lampiran 1 Rata-rata populasi cendawan, bakteri, dan aktinomiset pada lahan berbukit
P Ul A B C P Ul A B C
SB1 1 1 3 13 TSB1 1 0 7 13
SB1 2 3 13 15 TSB1 2 0 5 6
SB1 3 4 0 24 TSB1 3 0 4 7
SB2 1 2 11 12 TSB2 1 0 2 1
SB2 2 2 8 26 TSB2 2 2 4 3
SB2 3 0 7 18 TSB2 3 0 1 1
SB3 1 1 0 24 TSB3 1 0 0 3
SB3 2 0 1 25 TSB3 2 0 3 5
SB3 3 0 1 19 TSB3 3 0 0 2
∑ 13 44 176 ∑ 2 26 41
R 1.44 4.89 19.56 R 0.22 2.89 4.56
Keterangan: P= perlakuan; Ul= ulangan; A= aktinomiset; B= bakteri; C= cendawan; SB= Sehat berbukit; TSB= Terinfeksi berbukit; ∑= Jumlah mikroba; R= Rata-rata mikroba
Lampiran 2 Rata-rata populasi cendawan, bakteri, dan aktinomiset pada lahan datar
P Ul A B C P Ul A B C
SD1 1 1 4 37 TSD1 1 0 1 7
SD1 2 2 4 40 TSD1 2 0 0 9
SD1 3 2 5 49 TSD1 3 0 0 7
SD2 1 0 2 5 TSD2 1 0 4 3
SD2 2 0 1 7 TSD2 2 0 2 4
SD2 3 0 0 7 TSD2 3 0 2 1
SD3 1 0 8 9 TSD3 1 0 3 18
SD3 2 0 4 10 TSD3 2 0 2 18
SD3 3 0 4 3 TSD3 3 0 3 16
∑ 5 32 167 ∑ 0 17 83
R 0.56 3.56 18.56 R 0 1.89 9.22
21
Lampiran 3 Rata-rata populasi cendawan, bakteri, dan aktinomiset pada lahan bergelombang
P Ul A B C P Ul A B C
SG1 1 0 3 8 TSG1 1 0 1 10
SG1 2 0 1 4 TSG1 2 0 2 6
SG1 3 0 1 4 TSG1 3 0 1 4
SG2 1 0 2 6 TSG2 1 0 1 10
SG2 2 0 8 2 TSG2 2 0 1 8
SG2 3 0 2 7 TSG2 3 0 5 1
SG3 1 0 0 11 TSG3 1 0 1 10
SG3 2 0 0 10 TSG3 2 0 0 14
SG3 3 0 0 13 TSG3 3 0 0 13
∑ 0 17 65 ∑ 0 12 76
R 0 1.89 7.22 R 0 1.33 8.44
22
Lampiran 4 Jari-jari pertumbuhan koloni patogen G. boniense pada media PDA
Perlakuan Ul
Jari-jari pertumbuhan patogen G. boninense (mm)
23
Lampiran 5 Persentase penghambatan isolat akttinomiset terhadap cendawan patogen G. boninense
Perlakuan Ulangan Persen Penghambatan Hari Ke-
1 2 3 4 5
Kontrol
1 0 0 0 0 0
2 0 0 0 0 0
3 0 0 0 0 0
4 0 0 0 0 0
5 0 0 0 0 0
AK1 vs Gb
1 0 16.67 11.11 30.77 37.5
2 0 0 0 40 50
3 33.33 40 41.18 28 40
4 0 50 37.50 40 33.33
5 0 0 40 50 50
AK2 vs Gb
1 20 28.57 27.27 15.38 11.76
2 33.33 33.33 45.45 33.33 44.44
3 20 28.57 30 33.33 35
4 33.33 25 16.67 20 31.58
5 33.33 50 50 40 45
AK3 vs Gb
1 20 42.86 29.63 56.67 36.67
2 33.38 8.33 12.50 0 0
3 66.67 53.846 47.82 56.67 50
4 40 30.77 13.64 33.33 43.33
5 40 10 5 0 0
AK4 vs Gb
1 33.33 -30 -17.67 -10 4.17
2 28.57 6.25 4.5 11.54 20.69
3 0 36.36 23.53 18.18 25.92
4 33.33 13.33 20 26.67 20
RIWAYAT HIDUP
Penulis dilahirkan di Alahan Panjang, Sumatera Barat pada tanggal 14 juni 1991. Penulis merupakan putri pertama dari tiga bersaudara pasangan Bapak Guswarmen dan Ibu Yulisma.
Penulis menyeleseikan pendidikan sekolah menengah atas di SMA Negeri 3 Kota Solok pada tahun 2009. Pada tahun yang sama penulis melanjutkan studinya di Departemen Proteksi Tanaman, Fakultas Pertanian, Institut Pertanian Bogor melalui jalur Undangan Seleksi Masuk Institut Pertanian Bogor (USMI).
Selama menjadi mahasiswa, penulis aktif dalam beberapa organisasi kemahasiswaan, yaitu Ikatan Pelajar Mahasiswa Minang (2009), Ikatan Keluarga Mahasiswa Solok (2009-2013), dan Badan Pengawas Anggota (BPA) sebagai Sekretaris Umum (2011-2012). Selain itu, penulis pernah menjadi asisten praktikum mata kuliah Ilmu Penyakit Tumbuhan Dasar (2011-2012), Hama dan Penyakit Tanaman Setahun (2012), dan asisten praktikum mata kuliah Dasar-Dasar Proteksi Tanaman (2013). Penulis juga aktif mengikuti Unit Kegiatan Mahasiswa (UKM) seperti UKM Tae Kwondo pada tahun 2009-2010 dan UKM Tenis Meja pada tahun 2009-2012.