DI CAGAR ALAM PULAU DUA
DEWI ELFIDASARI
SEKOLAH PASCA SARJANA
INSTITUT PERTANIAN BOGOR
PELIMPAHAN HAK CIPTA
Dengan ini saya nyatakan bahwa disertasi berjudul Peran burung air liar dalam penyebaran virus Avian Influenza Subtipe H5N1 di Cagar Alam Pulau Dua adalah benar karya saya dengan arahan komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir disertasi ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, Juli 2013
Dewi Elfidasari
Avian Influenza Subtipe H5N1 di Cagar Alam Pulau Dua. Dibimbing oleh : DEDY DURYADI SOLIHIN, RETNO DAMAYANTI SOEJOEDONO, dan SRI MURTINI.
Unggas air seperti bebek/itik,entok dan angsa merupakan reservoir alami virus Avian Influenza (AI) subtipe H5N1. Pada reservoir alami, virus AI subtipe H5N1 bersifat low pathogenic LPAI. Selain Unggas air, burung air liar juga diduga berperan sebagai reservoir alami virus AI subtipe H5N1. Virus low pathogenic Avian Influenza (LPAI) telah berhasil diisolasi dari 105 jenis burung liar, yang sebagian besar merupakan ordo Anseriformes dan Charadiiformes. Dugaan adanya peran burung liar dan burung migran dalam proses penyebaran virus AI hingga saat ini masih belum dapat dibuktikan, sehingga perlu dilakukannya penelitian yang intensif. Tingkat kematian yang tinggi pada burung-burung air liar dilaporkan terjadi di sejumlah negara, tetapi belum jelas penyebab kematian tersebut.
Penelitian ini bertujuan untuk (1) menentukan seroprevalensi virus Avian Influenza (AI) subtipe H5N1 dari burung air liar penetap di kawasan Cagar Alam Pulau Dua (CAPD); (2) mengidentifikasi keberadaan gen Mx pada burung air liar terkait kemampuan ketahanan spesies ini dari serangan virus AI; (3) mengidentifikasi keberadaan virus AI subtipe H5N1 pada burung-burung air liar penetap di CAPD serta melihat potensi burung-burung tersebut sebagai reservoir virus AI subtipe H5N1,
Diagnosa adanya paparan atau infeksi virus AI digunakan untuk menentukan keberadaan virus AI di dalam tubuh unggas. Uji serologis dilakukan untuk melihat titer antibodi di dalam tubuh unggas. Pembentukan antibodi diawali ketika tubuh terinfeksi oleh virus. Sebagai substansi khusus yang dibentuk oleh tubuh, antibodi merupakan respon terhadap stimulasi antigen yang bersifat antigenik.
Hasil seroprevalensi pada tahun 2008, 2010 dan 2011 terhadap sampel serum burung air liar di CAPD menunjukkan telah terbentuk antibodi akibat paparan virus Avian Influenza subtipe H5N1 pada burung-burung tersebut. Nilai rataan titer antibodi pada masing-masing spesies sangat rendah.
Rendahnya titer antibodi dapat disebabkan beberapa faktor antara lain paparan virus AI terjadi secara tidak langsung, jumlah virus yang memaparkan relatif sedikit, waktu pemaparan relatif singkat dan virus yang memaparkan bersifat patogenitas rendah. Transmisi tidak langsung terjadi melalui air yang merupakan sumber air minum bagi burung-burung air liar maupun unggas domestik. Sekresi air liur dan cairan hidung, serta feses dari unggas yang terinfeksi tersebar dengan perantara air yang terdapat pada tempat interaksi antara burung-burung air liar dan unggas domestik.
dan 3 spesies unggas domestik (ayam, bebek dan entok) asal sekitar CAPD menggunakan primer NE-F2 dan NE-R2/R diperoleh produk PCR dengan panjang pita 100 bp. Ini menunjukkan bahwa keberadaan gen Mx dapat dilihat pada produk PCR tersebut. Hasil ini menjelaskan bahwa di dalam tubuh burung air liar penetap di kawasan CAPD memiliki gen Mx yang sama seperti yang dimiliki oleh ayam, bebek dan entok.
Hasil pemeriksaan Restriction Fragment Length Polymorphisms (RFLP) pada burung air liar dan unggas air dengan menggunakan enzim pemotong RsaI menunjukkan satu pita pemotongan sebesar 73 bp (hanya bergenotip GG). Ini memberikan informasi bahwa pada burung air liar dan unggas air domestik (entok & bebek) di sekitar CAPD tidak menunjukkan polimorfisme karena semua fragmen terpotong pada satu pita sebesar 73 bp. Pada ayam, pemotongan fragmen 73 bp dengan genotip GG menunjukan aktivitas antivirus yang rendah karena memiliki gen Mx yang rentan terhadap serangan virus AI subtipe H5N1. Hal ini tidak terjadi pada bebek, entok dan burung air liar, meskipun hanya memiliki satu potongan pita yang menunjukkan alel Mx-, akan tetapi burung air liar penetap di CAPD tahan terhadap paparan virus AI subtipe H5N1.
Identifikasi keberadaan virus dilakukan dengan RT-PCR menggunakan primer H5 Kha-1 dan reverse H5 Kha-3 untuk mendeteksi keberadaan gen H5. Hasil identifikasi HA subtipe H5 pada burung-burung air liar penetap di CAPD dengan menggunakan pasangan primer H5 menjelaskan bahwa subtipe H5 tidak terdeteksi pada burung-burung tersebut. Hal ini ditunjukkan dengan tidak terdapatnya pita positif yang mengindikasi tidak adanya virus AI subtipe H5 pada burung air liar tersebut.
Tidak terdapatnya hasil positif yang menunjukkan adanya virus AI subtipe H5 pada analisa molekuler terhadap RNA virus dari burung air liar penetap di CAPD semakin memperkuat dugaan bahwa burung air bukan tempat replikasi virus AI subtipe H5N1. Potensi burung air liar penetap sebagai sumber penularan virus AI subtipe H5N1 sangat kecil sekali karena 2 faktor, pertama hasil positif dari deteksi material genetik tidak mampu menjelaskan adanya virus yang hidup atau mati sehingga kemampuan untuk menyebarkan virus AI belum dapat dipastikan. Kedua, pada saat dilakukan pengambilan sampel usap kloaka bukan merupakan waktu viral shedding. Meskipun demikian tetap harus diwaspadai adanya kemungkinan penularan virus AI subtipe yang lain pada burung air liar penetap melalui unggas di sekitar kawasan CAPD.
Influenza Virus Subtype H5N1 at Pulau Dua Nature Reverse. Supervised by DEDY DURYADI SOLIHIN, RETNO DAMAYANTI SOEJOEDONO and SRI MURTINI.
Waterfowls such as ducks, muscovy, and swans are natural reservoir of influenza virus subtype H5N1. In natural reservoir, the AI virus is Low Pathogenic Avian Influenza (LPAI). Instead of domestic waterfowls, wild waterbird species could also have a role in spreading AI subtype H5N1 virus. So far LPAI have been isolated from 105 species of wild birds, most of them were from Anseriformes and Charadiiformes order. Presumption that wild birds and migratory birds have a role in spreading AI virus, has not been proven until now because the intensive research has not been done. The mortality rate of wild waterbirds that reported in several countries, had not clearly explained the cause of death.
The objectives of this research were (1) to estimate the seroprevalence of AI virus subtype H5N1 from residence wild waterbirds, in Pulau Dua Nature Reverse; (2) to identify the Mx gene as resistant anti viral gene from AI virus in wild waterbirds; (3) to identify the presence of AI virus subtype H5N1 on residence waterbirds in Pulau Dua Nature Reverse, also to observe the potential of wild waterbirds as reservoir for spreading AI virus subtype H5N1
The serological test to detect the presence of antibody antiviral AI virus subtype H5N1 was used to determine the exposure of AI virus subtype H5N1 among wild waterbirds in CAPD. Since antibody is the substance that appear in individual exposed when they are exposed by the strange antigen.
The serosurveillans from wild waterbirds in CPAD had been done in 2008, 2010 and 2011. Serological test by Haemagglutinin-inhibition (HI) showed that six from nine species of residence wild waterbirds in CPAD produced antibody AI virus H5N1. It is explain that that those species were exposed by AI virus subtype H5N1 with low antibody titer.
The low antibody titer can be caused by several factors such as AI virus exposure occurring indirectly, there was a small number of virus that exposure to wild waterbirds, the exposure time was very short, and the virus was low pathogenic. Indirect transmission occurred through the water as the source of drinking water for wild and domestic waterbirds, through saliva and nasal secretions and feces. Water as a media for transmission virus where there were interactions between wild waterbirds, waterfowls and domestic birds.
Mx protein as part of dynamin super family (GTP ase) is a part of the innate immune system. Mx proteins were known to have the ability to inhibit the replication of various viruses. Mx protein coded by Mx genes that specifically anti-viral influenza in several animals. Mx gene was the one of the several genes that regulated myxovirus resistance.
The results of PCR-RFLP in wild waterbirds and domestic waterfowls using the restriction enzyme RsaI showed band in 73 bp length (only have GG genotype). It informed that there was no polymorphism because all fragments display only one band (73bp ) in wild waterbirds and domestic waterfowls (muscovy and duck) around CAPD showed. In chickens, the 73 bp fragment (GG genotype) showed low antivirus activity because it has sensitive Mx gene (allele Mx-) and responsive to AI virus subtype H5N1 attack. This is not happened to ducks, muscovy, and wild waterbirds, although they have only one band (Mx -alleles). It was not caused death or clinical symptoms by the exposure of AI virus subtype H5N1.
The virus identification by RT-PCR with primer used in identification of HA subtypes H5. Forward primers was H5 Kha-1 and reverse H5 Kha-3primer pair. The identification results of HA subtypes H5 in residence wild waterbirds in CAPD showed that the H5 subtype was not detected in these wild waterbirds. This was showed by the absence of positive bands that indicate the presence of the AI virus subtypes H5 in wild waterbirds.
The absence of AI virus subtype H5N1 in nine species of residence wild waterbirds including Ciconiformes and Pelecaniformes order showed that these birds do not have the potential as a reservoir for AI virus subtype H5N1. It gives the information that birds are not a suitable host for the cultured AI virus subtype H5N1, so that there were no viral shedding into the environment from residence wild waterbirds.
© Hak Cipta Milik IPB, Tahun 2013
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik atau tinjauan suatu masalah dan pengutipan tersebut tidak merugikan kepentingan IPB
DI CAGAR ALAM PULAU DUA
DEWI ELFIDASARI
Disertasi
Sebagai salah satu syarat untuk memperoleh gelar Doktor Pada
Program Studi Biosains Hewan
SEKOLAH PASCA SARJANA
INSTITUT PERTANIAN BOGOR
Penguji pada Ujian Tertutup Prof. Dr. drh. I WayanTeguh Wibawan, MS Dr. Tike Sartika, MS
NIM G362080051
Disetujui oleh
Komisi Pembimbing
Dr. Ir. Dedy Duryadi Solihin, DEA Ketua
Prof Dr. drh. Retno D. Soejoedono, MS Dr. drh. Sri Murtini. M. Si
Anggota Anggota
Diketahui oleh
Ketua Program Studi
Biosains wan
Dr. Bambang Suryobroto
NIM : G362080051
Disetujui oleh Komisi Pembimbing
Dr. Ir. Dedy Duryadi Solihin, DEA Ketua
Prof. Dr. drh. Retno D. Soejoedono, MS Dr. drh. Sri Murtini, M.Si Anggota Anggota
Diketahui oleh
Ketua Program Studi Dekan Sekolah Pascasarjana Biosains Hewan
Dr. Bambang Suryobroto Dr. Ir. Dahrul Syah, M.ScAgr
PRAKATA
Bismillahirahmanirrahim
Alhamdulillah penulis panjatkan Kehadirat Allah SWT atas rahmat, karunia dan hidayah-Nya penulisan disertasi berjudul Peran Burung Air Liar dalam Penyebaran Virus Avian Influenza Subtipe H5N1 di Cagar Alam Pulau Dua
dapat diselesaikan dengan baik. Penelitian mendapat dana dari Insentif Riset Dasar KNRT 2007-2008, Hibah Kompetitif Prioritas Nasional DP2M DIKTI tahun 2010, Grant LP2M UAI 2012 dan Hibah Fundamental DP2M DIKTI 2013.
Penulis menghaturkan terimakasih dan penghargaan yang setinggi-tingginya kepada Dr. Ir. Dedy Duryadi Solihin, DEA; Prof. Dr. drh. Retno D. Soejoedono, MS dan Dr. drh. Sri Murtini, M.Si selaku pembimbing, atas bimbingan dan segala bantuan berbagai kemudahan penggunaan fasilitas penunjang penelitian dan kelancaran penyelesaian studi.
Kepada para penguji luar komisi Prof. Dr. drh. I Wayan T Wibawan, M.S dan Dr. Tike Sartika, M.Si (pada Ujian Tertutup); drh. Kamaludin Zarkassie, M.Sc., Ph.D dan Ir. Dewi Malia Prawiradilaga, M.Rur.Sc., Ph.D (pada Ujian Terbuka) penulis mengucapkan terimakasih atas perbaikan, saran dan masukan demi kesempurnaan disertasi ini.
Terimakasih kepada Staf BKSDA Serang, Jagawana CAPD (Pak Umar dan Pak Madsahi), drh. Tutur, drh. Iyas, drh. Nandi, Rahma Micho, S.Si, Dwi Putra A, S.Si, Lia M, S,Si, Edwinata, S.Si, Frisa, drh. Okti N. Poetri M.Si, Dr. Ni Luh Putu Ika, drh. Ita, Mega, drh. Lina, drh. Vivi, Pak Lukman, Mas Wahyu dan Pak Nur atas bantuan selama pengambilan sampel di CAPD dan analisa di lab. Terpadu Bag. Imunologi IPHK Fakultas Kedokteran Hewan IPB.
Kepada Bapak drs. Yus Rusila Noor (WI-IP), Prof. Dr. Ani Mardiastuti, M.Sc (KSDA Fahutan-IPB) terimakasih atas kesempatan diskusi dan berbagi ilmu terkait burung air dan konservasi. Rekan-rekan Mayor BSH SPs IPB angkatan 2008 (Ibu Wahyu Prihatini, Ibu Harini Nurcahya, Dr. Melta Rini, Dr. Islamul Hadi, Dr. Nurlisa), staf Laboran dan rekan-rekan di Lab. BioMolekuler Hewan PPSHB (Pak Heri, Dr. Dewi Indriani R, Dr. Niken Subekti, Dr. Fahma Wijaya, Dr. Suriana, Pak Hari Prayogo, Pak Tri Haryoko, Dr. Luciana) terimakasih atas
kebersamaan, saling bantu dan “support” selama 5 tahun ini.
Penulis mengucapkan terimakasih kepada sivitas akademik Universitas Al Azhar Indonesia terutama pada Fakultas Sains dan Teknologi Prodi Biologi (Ibu Dr. Nita Noriko; Risa Swandari W; Ibu Riris; Pak Syafitri Jumianto; Arief Pambudi) atas bantuan, dukungan dan kerjasama selama ini.
Kepada keluarga tersayang, Ibunda Sri Sarifah, suami Mulya Fahrizal dan empat buah hati (Annisa Nurul Hanifah, Achmad Hanif Khairullah, Arifah Husna Khairunnisa dan Amirah Nurul Husniyah), serta Lativa Savitri Elba terimakasih atas doa, motivasi, dukungan, ketulusan, keikhlasan dan kesetiaan yang diberikan.
Semoga disertasi ini bermanfaat bagi perkembangan ilmu pengetahuan khususnya bidang biologi.
1.
PENDAHULUAN
Latar Belakang
Unggas air seperti bebek/itik, entok dan angsa merupakan reservoir alami virus Avian Influenza (AI) subtipe H5N1. FAO (2008) memasukan unggas air ke dalam reservoir alami yang keberadaannya diperhitungkan sebagai sumber penularan virus AI subtipe H5N1. Pada reservoir alami, virus AI subtipe H5N1 bersifat low pathogenic Avian Influenza (LPAI). Burung air liar diduga sebagai reservoir alami virus AI subtipe H5N1, demikian pula unggas air. Sejumlah penelitian menunjukkan adanya peran burung liar sebagai reservoir virus AI subtipe H5N1 (CDC 2005; Khawaja et al. 2005; Hlinak et al. 2006; Clark dan Hall 2006; Munster et al. 2007; Wallensten et al. 2007).
Virus low pathogenic Avian Influenza (LPAI) telah berhasil diisolasi dari 105 jenis burung liar, yang sebagian besar merupakan ordo Anseriformes dan Charadriiformes (FAO 2008). Kedua ordo tersebut mencakup berbagai jenis burung air liar (penetap maupun migran). Serangkaian penelitian terdahulu memberi informasi adanya potensi dan peran burung migran dalam menyebarkan virus AI subtipe H5N1 (Ricardson 2006; Van Gils et al. 2007; Robinson 2007; Winker et al. 2007; FAO 2008). Pada burung-burung tersebut masuknya virus AI ke dalam tubuh tidak menyebabkan timbulnya gejala klinis, akan tetapi dapat menurunkan aktivitas mencari pakan dan migrasi. Sebanyak 75 jenis dari 10 ordo burung diketahui memiliki peran dalam menyebarkan virus AI subtipe H5N1, lebih dari 50% diantaranya merupakan burung air (FAO 2008).
Burung air yang merupakan salah satu indikator kondisi lingkungan adalah kelompok burung yang secara ekologis bergantung pada kawasan perairan baik sebagai habitat atau sebagai tempat mencari makan (Elfidasari 2007; FAO 2008; Lewis et al. 2008. White et al. 2009). Berdasarkan cara hidupnya, burung air dapat dibagi menjadi burung air domestik dan burung air liar. Burung air domestik sering disebut dengan unggas air, adalah burung air yang sengaja dipelihara dan dibudidayakan oleh manusia untuk memenuhi kebutuhan hidupnya, seperti bebek, itik dan entok. Burung air liar adalah burung air yang hidup dan menetap di alam liar, contohnya bangau, kuntul, camar, dan pecuk. Berdasarkan keberadaannya di suatu daerah, burung air liar dibagi menjadi dua kelompok yaitu burung air liar bermigrasi (migratory waterbird) dan burung air liar penetap (residence waterbird).
ditempat yang sama inilah yang memungkinkan terjadinya penularan virus AI subtipe H5N1 tersebut (Hulse-Post et al. 2005; Monke dan Corn 2007)
Highly Pathogenic Avian Influenza (HPAI) merupakan penyakit zoonosis yang bersifat sangat fatal dan menular. HPAI subtipe H5N1 dapat mengakibatkan gejala klinis berupa gangguan pada saluran pernapasan, gastro-intestinal dan atau syaraf. Wabah AI di Asia pertama kali dilaporkan di Cina Selatan tahun 1996-1997, selanjutnya menyebar dan menyebabkan kematian unggas di negara-negara kawasan Asia sejak awal tahun 2004 (Smith et al. 2004). Di Indonesia wabah AI pertama kali terjadi pada awal September 2003 hingga April 2004, menyebabkan kematian pada unggas akibat infeksi AI di beberapa wilayah di Indonesia meliputi Pulau Sumatera, Jawa dan Bali. Wabah ini menyerang ayam petelur dan pedaging, burung puyuh, dan ayam kampung. Tingkat hewan sakit (morbiditas) dan tingkat kematian (mortalitas) akibat infeksi AI sangat tinggi yaitu mencapai 90%. Penyebaran penyakit ini berlangsung sangat cepat terutama melalui perdagangan dan lalulintas unggas (Dharmayanti et al. 2004; Dharmayanti et al. 2006).
Wabah AI tersebut belum diketahui pengaruhnya terhadap burung liar 5hingga tahun 2004. Virus AI subtipe H5N1 diketahui telah membunuh 6.000 ekor burung air di Suaka Margasatwa Danau Qinghai, Barat laut China pada bulan Mei 2005. Jenis burung-burung air yang mati meliputi Anser indicus,
Phalacrocorax carbo, Larus sp dan Tadorna ferruginea. Pada tahun yang sama bulan Juli-Agustus, di Mongolia juga dilaporkan terjadi kasus kematian pada beberapa ekor burung migran Anser indicus dan Cygnus cygnus di dua danau yang berbeda di wilayah utara Mongolia (FAO 2008).
Pada tahun 2007, 38 negara yang berada di tiga benua (Asia, Eropa dan Afrika) melaporkan adanya virus AI subtipe H5N1 pada burung-burung liar. Di Asia, virus tersebut telah terdeteksi di 10 negara, sedangkan di Eropa virus tersebut telah terdeteksi di 25 negara, di Afrika virus AI subtipe H5N1 pada burung liar dijumpai pada 3 negara (FAO 2008).
Virus HPAI subtipe H5N1 dinyatakan endemik di 31 dari 33 propinsi di Indonesia pada tahun 2008. Transmisi zoonotik dari unggas ke manusia terjadi sejak pertengahan tahun 2005. Kematian manusia akibat H5N1 di Indonesia tercatat paling tinggi di dunia dengan jumlah kematian 159 orang dari 191 orang yang positif terinfeksi hingga Maret 2013. (SEARO 2013; WHO 2013). Kematian manusia paling banyak terjadi di DKI Jakarta (44 orang), Jawa Barat (38 orang), dan Banten (29 orang) (KNPZRI 2012).
Meningkatnya penderita Avian influenza Virus (AIV) di rumah sakit-rumah sakit di Propinsi Banten dan DKI Jakarta setiap bulan pada tahun 2005-2009 menunjukan bahwa tingkat penyebaran virus ini terjadi dengan sangat cepat. Keberadaan unggas peliharaan (seperti ayam, bebek dan burung) yang berada di sekitar kawasan perumahan penduduk selama ini diduga turut berperan dalam proses penyebaran virus AI subtipe H5N1 tersebut. Namun informasi mekanisme penularan virus AI pada hewan-hewan peliharaan belum jelas.
Kemungkinan 1.
Kemungkinan 2
Kemungkinan 3
Gambar 1. Alur penularan virus AI subtipe H5N1 (Mulyadi dan Prihatini 2005)
Berdasarkan kemungkinan yang dapat terjadi tersebut, maka diperlukan informasi terkait peran burung air liar dalam proses penyebaran virus AI. Sumber inforrmasi diperoleh baik melalui uji serologi maupun uji molekuler lengkap yang dapat mengidentifikasi dan mengetahui karakteristik virus AI yang terdapat pada burung air liar tersebut. Berbagai dugaan termasuk peran keberadaan burung liar dan burung migran dalam proses penyebaran virus ini masih belum dapat dibuktikan, karena belum dilakukannya penelitian yang intensif. Tingkat kematian yang dilaporkan terjadi pada burung-burung air liar yang dijumpai pada sejumlah negara belum juga menjelaskan penyebab kematian tersebut.
Cagar Alam Pulau Dua (CAPD) merupakan daerah lahan basah yang telah ditetapkan sebagai wilayah utama bagi konservasi burung-burung air pada tahun
Unggas liar Unggas domestik
Babi terinfeksi virus influenza-burung dan virus influenza manusia Manusia
Menular ke manusia lain
Unggas liar Unggas domestik
Babi terinfeksi virus influenza-burung dan virus influenza manusia manusia lain
Unggas liar Unggas domestik
1937, berdasarkan keputusan Gubenur Jenderal Hindia Belanda tanggal 30 Juli 1937 No. 21 Stbl 474 (Milton dan Marhadi 1985; Partomihardjo 1986). Terdapat 12 spesies burung air yang menjadi penetap pada kawasan tersebut, Anhinga melanogaster,Threskiornis melanocephalus, Plegadis falcinellus, Casmerodius albus, Egretta intermedia, Egretta garzetta, Bubulcus ibis, Nycticorax nycticorax, Ardeola speciosa, Phalacrocorax sp, Ardea cinerea, Ardea purpurea (Rusila-Noor 1996).
Beberapa jenis burung migran juga tercatat dijumpai di kawasan CAPD pada saat musim migrasi. Burung-burung tersebut meliputi bambangan kuning (Ixobrychus sinensis), cerek kenyut (Pluvialis fulva), cerek kalung kecil (Charadrius dubius), trinil pantai (Actitis hypoleucos), dan kirik-kirik laut (Merops phillipinus) (Prawiradilaga et.al. 2009). Pada musim migrasi ini terjadi interaksi antara burung migran dan penetap di kawasan CAPD. Interaksi yang terjadi baik secara langsung maupun tidak langsung inilah yang memungkinkan terjadinya penyebaran virus AI subtipe H5N1 (Robinson 2007; FAO 2008).
Van Gils et al. (2007) menyatakan bahwa penyebaran virus AI subtipe H5N1 diduga terjadi pada saat burung migran singgah di suatu kawasan. Burung migran tersebut akan berinteraksi dan melakukan kontak dengan burung air penetap atau unggas air yang terdapat di kawasan tersebut. Kontak langsung terjadi pada saat mencari pakan di lokasi yang sama. Penularan virus AI subtipe H5N1 dapat terjadi melalui makanan maupun air yang telah tercemar virus tersebut. Cemaran virus AI subtipe H5N1 dapat berasal dari air liur dan feses yang berasal dari burung migran dan dikeluarkan pada kawasan tersebut saat mencari pakan. Secara umum, virus AI subtipe H5N1 di permukaan air memiliki kemampuan untuk bertahan selama 4 hari pada suhu 22oC, dan lebih dari 30 hari pada suhu 0oC. Suspensi virus tersebut menunjukkan kemampuan yang baik dalam mempertahankan strukturnya di alam dan mampu mempertahankan daya penularannya selama lebih dari 100 hari pada suhu 17oC, terutama di permukaan air. Di bawah -50oC virus AI subtipe H5N1 dapat bertahan lebih lama lagi (CIDRAP 2004; Kamps et al. 2006).
Kemampuan adaptasi virus AI subtipe H5N1 yang cukup baik ini diduga menjadi salah satu faktor yang berperan dalam proses penyebaran virus AI dari burung air liar ke unggas domestik atau sebaliknya, dari unggas domestik ke burung air liar. Survei dan pengambilan sampel telah dilakukan terhadap ratusan ribu burung liar yang sehat baik migran maupun penetap, namun hingga saat ini sumber virus AI subtipe H5N1 tersebut belum teridentifikasi dengan pasti (FAO 2008).
Perumusan Masalah
burung air liar tidak dapat dihindari. Kondisi ini diduga menjadi salah satu faktor yang berperan dalam proses penularan virus AI dari burung air liar ke unggas domestik atau sebaliknya, dari unggas domestik ke burung air liar.
Adanya paparan virus AI sub tipe H5N1 pada burung air liar di CAPD tanpa memperlihatkan gejala-gejala terinfeksi penyakit diduga berkaitan dengan daya tahan (resistensi) burung air liar. Paparan virus AI tanpa gejala-gejala dapat juga diakibatkan infeksi oleh virus AI subtipe H5N1 dalam jumlah sedikit demi sedikit sehingga tubuh mampu membentuk kekebalan tubuh (antibodi) dengan titer antibodi yang rendah.
Resistensi merupakan kemampuan yang ditunjukan oleh suatu individu dalam mempertahankan tubuh terhadap infeksi penyakit yang berasal dari mikroorganisme baik virus, bakteri parasit maupun jamur. Mekanisme resisten berkaitan erat dengan sistem kekebalan tubuh (sistem imun) dalam memberi respon terhadap masuknya mikroorganisme asing ke dalam tubuh suatu individu. Sistem ini akan segera memberi reaksi yang tepat untuk mencegah timbulnya penyakit yang disebabkan masuknya antigen atau patogen asing tersebut.
Sistem imun yang memberi respon pertama terhadap masuknya mikroorganisme ke dalam tubuh hewan merupakan sistem imun nonspesifik. Sistim imun ini didapat sejak lahir dan bekerja pada saat masuknya mikroorganisme ke dalam tubuh. Mekanisme awal yang dilakukan sistem imun nonspesifik adalah mendeteksi mikroorganisme asing yang masuk ke dalam tubuh dan selanjutnya akan melindungi tubuh dari kerusakan yang diakibatkan mikroorganisme tersebut. Sistem imun nonspesifik merupakan “barier” pertama pertahanan tubuh masuknya mikroorganisme meskipun baru pertama kali dikenali (Radji 2010; Mayer 2011).
Salah satu faktor penting dalam sistem imun yang menghasilkan sifat resistensi terhadap masuknya virus ke dalam tubuh adalah interferon. Interferon merupakan golongan protein tertentu yang mampu menghasilkan kekebalan nonspesifik terhadap infeksi virus. Interferon memiliki efek antivirus baik secara langsung maupun tidak langsung. Interferon dapat bekerja dengan cara menginduksi sel-sel pembentukan enzim yang dapat merusak RNA virus dan menghambat sintesis protein sehingga mampu menghambat replikasi virus. Pada saat virus menginfeksi sebuah sel, asam nukleat virus akan menginduksi perangkat genetik sel untuk membentuk interferon. Interferon akan dikeluarkan ke dalam cairan ektraseluler dan berikatan dengan reseptor pada membran plasma sel-sel di sekitar invasi virus. Ikatan antara reseptor dengan membran plasma sel tersebut akan merangsang sel inang membentuk enzim-enzim atau protein antivirus yang akan menghambat perbanyakan virus, salah satunya adalah protein Mx (Lee dan Vidal 2003; Ewald et al. 2010; Mayer 2011).
Hasil penelitian yang dilakukan oleh para peneliti terdahulu memberi informasi status ayam dengan tingkat resisten tinggi dan tidak menunjukkan gejala terinfeksi AI memiliki frekuensi alel A (alel Mx+) yang tinggi yaitu lebih dari 50% dibandingkan yang memiliki alel G (alel Mx-) (Ko et al. 2002; Seyama
et al. 2006; li et al. 2006; Quan et al. 2010). Menurut Sulandari et al. (2009) dan Sartika et al. (2011), ayam lokal di Indonesia mempunyai frekuensi alel A lebih dari 60% dan frekuensi alel G kurang dari 40%, hal ini menunjukan bahwa ayam lokal bersifat resisten terhadap virus AI subtipe H5N1.
Peran gen Mx sebagai gen pengatur mekanisme resistensi terhadap virus tertentu telah banyak dipelajari pada berbagai vertebrata seperti tikus, mencit, ayam dan bebek. Gen MxA pada manusia memproduksi protein yang bersifat antivirus spesifik terhadap virus yang berasal dari famili Orthomyxoviridae, Rhabdoviridae, Paramyxoviridae, Bunyaviridae, Picoviridae dan Togaviridae. Gen Mx1 dan Mx2 pada tikus dan mencit memproduksi protein antivirus spesifik terhadap Orthomyxoviridae, Rhabdoviridae dan Bunyaviridae. Gen Mx pada ayam menghasilkan protein antivirus spesifik terhadap Orthomyxoviridae dan Rhabdoviridae (Lee dan Vidal 2002).
Belum adanya data dan informasi terkait potensi dan peran gen Mx pada burung air liar menyebabkan belum diketahui bagaimana keberadaan gen Mx di dalam tubuh burung air liar. Isolasi dan identifikasi gen Mx pada burung air liar diharapkan mampu menjelaskan potensi gen tersebut sebagai gen yang menentukan tingkat resistensi burung air liar di kawasan Cagar Alam Pulau Dua terhadap infeksi virus AI.
Isolasi dan identifikasi virus AI dari burung air liar penetap akan membantu mengidentifikasi peran dan potensi burung air liar tersebut dalam penyebaran virus AI subtipe H5N1.
Tujuan Penelitian
Penelitian ini bertujuan :
1. Menentukan seroprevalensi virus Avian Influenza (AI) subtipe H5N1 dari burung air liar penetap pada kawasan Cagar Alam Pulau Dua
2. Identifikasi dan genotiping gen Mx pada burung air liar penetap di Cagar Alam Pulau Dua.
3. Identifikasi keberadaan virus AI subtipe H5N1 pada burung air liar penetap di Cagar Alam Pulau Dua dan menganalisa peranan burung-burung tersebut dalam penyebaran virus AI subtipe H5N1di daerah Serang dan sekitarnya
Manfaat Penelitian
kebijakan yang berkaitan dengan kesehatan lingkungan dan konservasi kawasan tersebut
Pengetahuan tentang peran gen Mx+ dapat memberikan informasi mengenai potensi gen tersebut dalam mengendalikan daya tahan tubuh pada burung air liar sehingga mampu bersifat resisten terhadap infeksi virus AI.
Hipotesis
1. Terjadi pembentukan antibodi dalam tubuh burung air liar di kawasan Cagar Alam Pulau Dua akibat paparan virus AI subtipe H5N1
2. Gen Mx berperan dalam mengendalikan sifat resisten pada tubuh burung air liar di kawasan Cagar Alam Pulau Dua terhadap infeksi virus AI sub tipe H5N1
Alur Penelitian
PERAN BURUNG AIR LIAR
DALAM PENYEBARAN VIRUS AI SUBTIPE H5N1 DI CAGAR ALAM PULAU DUA
- Apakah burung air liar penetap di CAPD terinfeksi virus AI subtipe H5N1 - Apakah burung air liar memiliki resistensi terhadap virus AI
- Apakah burung air liar memiliki peran menyebarkan virus AI subtipe H5N1
Hanya Koloni Burung Air Liar penetap
Peran koloni burung air liar penetap di CAPD dalam penyebaran virus AI subtipe H5N1 di sekitar CAPD Serosurveilans virus
AI subtype H5N1
Uji HI Serum
Seroprevalensi Virus AI subtipe H5N1 burung air liar di CAPD
Analisis RNA
- Propagasi virus pada
TAB-SPF
- Isolasi RNA
- RT-PCR virus AI (HA)
-Usap kloaka
Identifikasi virus AI subtipe H5
Analisis DNA
- Isolasi DNA
- PCR-RFLP gen Mx
Sel darah merah (SDM)
1.
SEROSURVEILANS VIRUS AVIAN INFLUENZA SUB-TIPE
H5N1 PADA BURUNG-BURUNG AIR LIAR PENETAP
DI CAGAR ALAM PULAU DUA
ABSTRAK
Dugaan keterlibatan burung-burung liar sebagai vektor Avian Influenza yang menjangkiti beberapa Negara di Asia perlu dibuktikan lebih dalam. Sejak bulan Pebruari hingga Juni 2007 dilakukan penelitian berupa pengambilan sampel terhadap burung-burung liar yang menghuni kawasan Cagar Alam Pulau Dua (CAPD), Serang. Serosurveilans ini bertujuan untuk mengetahui adanya infeksi virus Avian Influenza (AI) sub-tipe H5N1 pada burung-burung air liar tersebut. Metode yang digunakan adalah penangkapan pada burung-burung yang sedang berbiak, dilanjutkan pengambilan sampel darah burung-burung air liar tersebut yang kemudian dilanjutkan dengan Uji HI (Hemagglutination-inhibition). Dari hasil pengambilan sampel yang dilakukan antara Bulan Pebruari hingga Juni 2007, diperoleh sebanyak 183 sampel darah dari 183 burung yang mewakili 7 jenis burung air yang menghuni CAPD yaitu (Bubulcus ibis, Ardeola speciosa, Egretta garzetta, Egretta intermedia, Nycticorax nycticorax, Casmerodius albus
dan Ardea cinerea). Hasil serosurvelians mengungkapkan bahwa terdapat 41 sampel (23,43%) yang menunjukkan terbentuknya antibodi virus AI subtipe H5N1 yang dinyatakan sebagai hasil positif. Berdasarkan jenis burung, sebanyak 29,27% Bubulcus ibis positif, Egretta garzetta 29,27%, Egretta intermedia
4,88%, Ardeola speciosa 7,32%, dan Nycticorax nycticorax 29,27% Berdasarkan usia burung maka hasil uji dapat dibagi menjadi 2 kategori, dewasa dan anakan. Hasil uji HI menunjukkan bahwa 41,46% individu positif dijumpai pada burung dewasa, sedangkan 58,54% adalah individu anakan.
Kata kunci : Virus AI subtipe H5N1, koloni burung air liar, Cagar Alam Pulau Dua, serosurveilans
SEROSURVEILLANCE OF AVIAN INFLUENZA VIRUS
SUBTYPE H5N1 ON WILD WATERBIRDS IN PULAU DUA
SERANG NATURAL PRESERVES BANTEN PROVINCE
ABSTRACT
Haemagglutination-inhibition) test. A total of 183 samples from 7 water bird species were collection i.e Cattle egret (Bubulcus ibis), Javan pond-heron (Ardeola speciosa), Little egret (Egretta garzetta), Intermediate egret (Egretta intermedia), Black-crowned night heron (Nycticorax nycticorax), Great egret (Casmerodius albus) and Grey heron (Ardea cinerea). The result revealed that 41 (23,27%) samples showed the present of AIV antibodies serotype H5N1 which is identified as positive. The five positive-test species, included Bubulcus ibis (29,27%), Egretta garzetta
(29,27%), Egretta intermedia (4,88%), Ardeola speciosa (7,32%), and
Nycticorax nycticorax (29,27%). A total of 41.46% infected individuals were adults whereas 58,54% were juveniles.
Keywords : Avian Influenza Virus, colonial water birds, Pulau Dua Nature Reserve, serosurveillans
PENDAHULUAN
Sejak ditemukan kasus pertama, Juli 2005, jumlah kasus Flu Burung pada manusia di Indonesia hingga tanggal 28 Januari 2008 mencapai 124 orang, dimana 100 orang diantaranya meninggal dunia. Dengan demikian, angka kematian atau Case Fatality Rate (CFR) mencapai 80.6%. Avian influenza Virus
(AIV) adalah penyakit menular yang disebabkan oleh virus influenza yang ditularkan oleh unggas yang dapat menyerang manusia (FAO 2008)
Secara sistematika, virus influenza termasuk dalam famili Orthomyxoviridae
dari genus Influenza. Virus ini memiliki ukuran diamater virions 80 hingga 120 nm yang berbentuk filamen, dan terdiri dari delapan segmen berbeda dari
negative- stranded RNA (Whittaker 2001). Virus ini termasuk dalam jenis virus
host-spesific, artinya mempunyai inang tertentu yang khusus. Inang tertentu tersebut merupakan tempat virus berkembang biak dan memiliki potensi untuk menyebar melalui aktivitas tertentu, jenis hewan yang menjadi inang bagi AIV (H5N1) adalah unggas (Hulse-Post et al. 2005).
Virus Avian Influenza atau flu burung hidup di dalam saluran pencernaan unggas. Burung yang terinfeksi virus ini akan mengeluarkan virus melalui air liur (saliva), cairan hidung dan kotorannya. Penularan virus ini terjadi dari unggas ke unggas dan dari unggas ke manusia, sejauh ini belum ada bukti yang menyatakan bahwa virus ini dapat menular dari manusia ke manusia (pandemi). Proses penularan virus ini terjadi melalui air liur serta melalui lendir dari hidung dan feses. Virus ini dapat menular melalui udara dan air yang tercemar virus AI subtipe H5N1 yang berasal dari kotoran atau cairan unggas yang terinfeksi virus (Hulse-Post et al. 2005; Whittaker 2001).
Di Indonesia sendiri belum ada data yang menjelaskan apakah migrasi burung yang terjadi setiap tahun melalui Indonesia juga berpotensi dalam penyebaran virus AI sub-tipe H5N1 pada unggas dan manusia. Seperti telah diketahui, beberapa lokasi di Indonesia merupakan tempat persinggahan bagi burung-burung migran pada saat musim migrasi. Daerah-daerah tersebut juga biasanya merupakan tempat berbiak burung-burung penetap (residen) yang menjadi penghuni tetap kawasan tersebut. Beberapa kawasan termasuk dalam daerah yang dilindungi oleh Pemerintah sebagai Kawasan Konservasi keanekaragaman hayati (Rusila-Noor et al. 2000).
Salah satu kawasan konservasi yang merupakan daerah penting berbiak bagi burung air yang dilindungi oleh pemerintah adalah Cagar Alam Pulau Dua (CAPD) yang terletak di Teluk Banten, Kabupaten Serang Propinsi Banten. Daerah di sekitar CAPD berupa hamparan lumpur adalah habitat mencari makan burung air migran. Beberapa daerah di sekitar CAPD yang dipergunakan oleh burung air untuk mencari makan meliputi tambak, sawah, tegalan dan dataran lumpur „mudflat‟ (Elfidasari 2006a). Pada saat mencari makan itulah sering terjadi interaksi antara burung-burung air liar (penetap maupun migran) dengan unggas domestik atau dengan manusia.
Secara serologis terdapat berbagai cara untuk mendeteksi keberadaan virus AI pada unggas (serosurveilans), antara lain dengan melakukan Uji
Hemagglutination Inhibition (HI). Uji HI merupakan uji yang dapat digunakan untuk mengidentifikasi keberadaan antibodi di dalam darah. Pada uji ini digunakan antigen yang homolog sehingga akan terjadi ikatan antigen-antibodi yang menyebabkan virus tidak mampu melekat atau berikatan dengan receptor membrane sel darah merah dan aglutinasi tidak akan terjadi. Uji HI mempunyai fungsi antara lain sebagi sarana untuk mengidentifikasi jenis antibodi tertentu dengan melihat reaksinya terhadap antigen homolog yang telah diketahui, dan untuk mengetahui titer antibodi dengan mereaksikan serum yang ingin diketahui antibodinya dengan antigen standar yang telah diketahui (OIE 2005; Soejoedono 2006).
Uji HI memiliki dua metode yaitu metode α dan metode β. Metode α
digunakan untuk menguji jenis antigen, dengan melakukan pengenceran pada antigen tetapi jumlah antibodi yang telah diketahui tetap. Kelebihan dari metode ini adalah dapat langsung mengidentifikasi antigen tanpa melakukan uji HA terlebih dahulu. Sedangkan metode β digunakan untuk mengidentifikasi antibodi dan menghitung titer antibodinya. Pengujian dilakukan dengan melakukan pengenceran pada antibodi tetapi jumlah antigen tetap. Kelebihan dari metode ini ialah penggunaan jumlah antibodi yang lebih sedikit dan diketahui titer antibodinya (Soejoedono 2006).
Uji HI dapat dilakukan secara makro atau mikro titrasi, tergantung reagen-reagen yang digunakan. Hal yang membedakan dari kedua cara tersebut hanya volume reagen dan virus standar yang digunakan. Pada makrotitrasi, virus standar yang digunakan yaitu 8 HAU (Hemagglutination Unit) atau 10 HAU, sedangkan pada mikrotitrasi virus standar yang digunakan 4 HAU(Soejoedono 2006).
ini dan apakah kawasan ini merupakan salah satu lokasi yang berpotensi dalam menyebarkan virus AI sub-tipe H5N1 di sekitar Propinsi Banten dan DKI Jakarta.
Penelitian ini bertujuan untuk mengetahui keberadaan antibodi terhadap virus AI sub tipe H5N1 secara serologis (serosurveilans), serta memperoleh informasi yang jelas apakah burung-burung air liar penghuni kawasan Cagar Alam Pulau Dua Serang merupakan salah satu carrier dalam proses penyebaran virus AI sub-tipe H5N1 di sekitar Propinsi Banten dan Jakarta.
METODE
Penelitian ini terdiri dari penelitian lapangan dan penelitian laboratorium. Penelitian lapangan dilakukan pada bulan Pebruari-Juni 2008 di kawasan Cagar Alam Pulau Dua, Serang (Gambar 2). Pengambilan sampel darah dilakukan pada burung-burung air yang menghuni Kawasan Cagar Alam Pulau Dua, Serang, Propinsi Banten. Sampel yang diambil berupa sampel darah dan usapan kloaka (Lampiran 1). Pengujian sampel dilakukan di laboratorium Mikrobiologi Medis Fakultas Kedokteran Hewan Institut Pertanian Bogor. Uji serologis yang dilakukan adalah Uji HI (Hemagglutination-inhibition) (Lampiran 2).
Gambar 2. Cagar Alam Pulau Dua Serang, Propinsi Banten (sumber : google map)
Teluk banten
HASIL DAN PEMBAHASAN
Dari penelitian lapangan yang dilakukan pada bulan Pebruari-Juni 2008, jumlah total sampel yang diperoleh adalah 183 sampel darah yang dibedakan, berdasarkan usia individu burung dalam dua kategori yaitu dewasa dan anakan (Juvenil). Sampel dewasa sebanyak 98 dan sampel anakan sebanyak 85 (Tabel 1).
Hasil Uji HI terhadap seluruh sampel darah burung air liar yang diperoleh menunjukkan bahwa sebanyak 41 sampel (23,43%) memberi hasil uji positif yang ditandai dengan tidak terjadinya aglutinasi sel darah merah. Aglutinasi sel darah merah yang tidak terbentuk disebabkan karena telah terbentuknya antibodi pada tubuh burung air liar akibat paparan virus Avian Influenza (AI) tipe H5N1.
Tabel 1. Jumlah sampel dari kawasan CAPD No. Jenis Burung Nama
Indonesia Dewasa Anakan Total 1. Bubulcus ibis Kuntul kerbau 40 20 60 2. Ardeola speciosa Blekok sawah 17 14 31 3. Nycticorax nycticorax Kowak maling 15 18 22 4. Egretta intermedia Kuntul sedang 4 16 20 5. Egretta garzetta Kuntul kecil 22 14 35
6. Phalacrocorax sp Pecuk 0 1 1
7 Casmerodius albus Kuntul besar 0 1 1
8. Ardea cinerea Cangak abu 0 1 1
TOTAL 98 85 183
Menurut Stanley (2002) pada Uji HI hasil positif akan ditandai dengan tidak terjadinya aglutinasi sel darah merah. Aglutinasi sel darah merah yang tidak terbentuk disebabkan karena terdapat antibodi terhadap virus AI di dalam sampel serum. Antigen akan diikat oleh antibodi, hal ini dapat menyebabkan antigen tidak dapat berikatan dengan sel darah merah, sehingga antigen virus tidak dapat mengaglutinasi sel darah merah. Sementara itu, hasil negatif akan ditandai dengan terjadinya aglutinasi pada sel darah merah, aglutinasi terjadi karena sampel serum tidak mengandung antibodi terhadap virus AI. Sehingga antigen virus akan berikatan dengan sel darah merah yang akan menyebabkan aglutinasi pada sel darah merah.
Adanya antibodi dalam serum menunjukkan bahwa virus mungkin masih berada di dalam tubuh sehingga keberadaan antibodi berfungsi untuk melawan infeksi, atau virus sudah tidak ada lagi di dalam tubuh karena sudah terliminasi oleh antibodi (Akoso 2006). Hasil positif dari Uji HI menunjukkan bahwa burung-burung air liar di kawasan Cagar Alam Pulau Dua pernah terpapar virus AI subtipe H5N1 (Gambar 3). Kemungkinan paparan ini disebabkan karena secara geografis kawasan tersebut merupakan salah satu lokasi yang disinggahi oleh burung-burung migran pada waktu migrasi, dan kemudian berinteraksi dengan burung-burung yang berbiak di Pulau Dua. Kondisi ini memudahkan terjadinya transmisi virus AI.
Hasil ini memberi informasi yang sangat berguna mengingat bahwa burung-burung air liar yang menghuni kawasan Cagar Alam Pulau Dua Serang pada saat mencari makan sebagian besar berinteraksi, baik dengan burung bermigrasi maupun unggas-unggas domestik, seperti bebek, ayam, itik dan entok yang menjadi peliharaan masyarakat. Adanya pemanfaatan sumber bahan makanan dan lokasi mencari makan yang sama pada unggas air liar dan unggas air domestik memungkinkan terjadinya interaksi antara keduanya pada lokasi makan yang berada di sekitar kawasan CAPD seperti tambak, sawah, dataran berlumpur (Elfidasari 2007). Hal ini dapat menyebabkan terjadinya penularan virus AI subtipe H5N1.
Hasil penelitian yang dilakukan Fang et al. (2008) menunjukan bahwa interaksi yang terjadi antara unggas air liar dengan unggas air domestik dapat menyebabkan cross-infection baik dari unggas domestik ke unggas liar maupun sebaliknya. Interaksi ini dapat terjadi di daerah berair seperti kubangan, sawah, tambak, danau dan dataran berlumpur (CMS 2006). Interaksi langsung antara burung air liar bermigrasi dan burung air liar penetap terjadi pada saat musim migrasi. Pada musim tersebut burung air liar bermigrasi akan melakukan perjalan panjang ke lokasi tujuan dengan melalui wilayah pesisir Indonesia. Sementara
-5 10 15 20 25 30 35
Sampel darah
[image:30.595.97.512.82.438.2]Sampel positif
kontak langsung antara burung air liar dan unggas domestik (bebek, itik, entok) terjadi pada saat mencari makan di lokasi yang sama (van Gills et al. 2007).
Virus AI dapat menyebar dengan cepat diantara populasi unggas, terutama virus AI subtipe H5N1. Penyebaran virus atau transmisi virus dapat terjadi melalui kontak langsung maupun tidak langsung. Transmisi langsung virus AI subtipe H5N1 dapat terjadi apabila virus AI tersebut menginfeksi unggas air atau hewan lain tanpa melalui suatu media. Sedangkan transmisi tidak langsung bisa terjadi melalui media seperti air yang menjadi sumber minum bagi burung air liar maupun unggas domestik dan telah tercemar virus AI subtipe H5N1 (Nazaruddin 2008). Transmisi tidak langsung juga dapat terjadi melalui sekresi air liur dan cairan hidung serta feses dengan perantara air yang terdapat pada tempat berinteraksi baik unggas liar maupun unggas domestik (CMS 2006).
Tercemarnya air oleh virus AI subtipe H5N1 dapat terjadi karena sekresi dari air liur atau feses unggas yang positif mengandung virus AI subtipe H5N1 tersebut masuk ke lingkungan perairan yang biasanya menjadi tempat mencari makan bagi burung-burung air dan unggas air domestik di kawasan perairan di sekitar CAPD. Air merupakan media yang baik bagi virus AI subtipe H5N1. Suspensi virus AI tersebut dalam air mampu mempertahankan daya penularannya selama lebih dari 100 hari pada suhu 17oC. Di bawah suhu -15oC virus AI dapat bertahan lebih dari 30 hari (CIDRAP 2004; FAO 2007; Si et al. 2010)).
Berdasarkan perbedaan usia objek yang berhasil ditangkap, hasil uji HI menunjukkan 17 (41,46%) sampel positif adalah individu dewasa dan 24 (58,54%) sampel positif adalah anakan (juvenile). Jumlah individu anakan yang menunjukkan hasil uji HI positif tertinggi dijumpai pada kuntul kecil (Egretta garzetta), yaitu sebanyak 9 ekor dari 12 ekor yang menunjukkan hasil positif (Gambar 4).
Pada kowak maling (Nycticorax nycticorax) dari 12 sampel yang menunjukkan hasil positif, 8 ekor merupakan anakan. Sedangkan pada kuntul kerbau (Bubulcus ibis) jumlah individu positif Uji HI terbanyak dijumpai pada
-5.00 10.00 15.00 20.00 25.00
Hasil Uji Positif Dewasa
Hasil Uji Positif Anakan
dewasa, yaitu sebanyak 9 ekor. Pada kuntul sedang Egretta intermedia dan blekok sawah Ardeola speciosa, jumlah individu anakan yang menunjukkan hasil positif Uji HI masing-masing sebanyak 2 ekor (Gambar 4).
Adanya antibodi terhadap virus AI pada burung-burung air liar dewasa kemungkinan dapat disebabkan karena burung-burung tersebut lebih banyak berinteraksi dengan lingkungan dan unggas-unggas lain seperti unggas domestik. Terbentuknya antibodi pada burung-burung anakan (juvenile) dapat disebabkan oleh beberapa faktor seperti paparan langsung virus AI yang berasal dari interaksi dengan induk, dari antibodi asal induk yang diturunkan ke anak (Maternal antibodi) atau dari makanan yang diberikan induk kepada anak (Fenner et al.
1993). Hal ini membuktikan bahwa virus AI tidak mengenal rentang umur dalam menginfeksi hewan targetnya.
Interaksi antara induk burung yang telah terpapar virus AI subtipe H5N1 sangat memberi peluang yang cukup besar bagi penularan virus tersebut pada anak burung. Demikian juga dengan pada saat pemberian makan kepada anak. Induk burung memberi makan anakan berupa mangsa yang diperoleh dari kawasan perairan di sekitar CAPD. Bila kawasan tersebut memiliki perairan yang mengandung virus AI subtipe H5N1, maka kemungkinan besar anak juga akan mengandung antibodi terhadap virus AI (Si et al. 2010). Berdasarkan hasil pengamatan pada saat pemberian makan kepada anak burung, induk akan memberi makan berupa ikan, cacing, udang dan hewan mangsa lain yang berasal dari kawasan perairan di sekitar CAPD
Penurunan antibodi dari induk kepada anak burung (maternal antibody) dapat terjadi pada populasi burung air liar di kawasan CAPD ini, akan tetapi perlu dilakukan penelitian lebih lanjut mengenai mekanisme tersebut.
SIMPULAN
3. IDENTIFIKASI DAN ANALISA POLIMORFISME GEN MX
PADA BURUNG AIR LIAR DI CAPD TERHADAP PAPARAN
VIRUS AVIAN INFLUENZA SUBTIPE H5N1
ABSTRAK
Protein Mx merupakan protein yang berperan dalam aktivitas antiviral. Protein ini dapat menentukan sifat resisten atau rentan terhadap berbagai jenis virus, termasuk virus AI subtipe H5N1. Protein Mx disandikan oleh gen Mx. Respon infeksi virus AI subtipe H5N1 pada ayam diperlihatkan oleh adanya polimorfisme gen Mx. Alel Mx+ menunjukan sifat resisten sedangkan alel Mx- menunjukan sifat rentan terhadap virus AI subtipe H5N1. Tujuan penelitian ini adalah untuk membuktikan bahwa gen Mx terdapat pada burung air liar penetap di CAPD, serta menunjukan adanya polimorfisme gen Mx sebagai respon terhadap paparan virus AI subtipe H5N1 pada burung air liar tersebut. Metode yang digunakan adalah PCR-RFLP menggunakan sepasang primer NE-F2 dan NE-R2/R dan enzim pemotongan RsaI. Sampel DNA burung air liar berasal dari 8 jenis burung air liar yang yang positif terhadap paparan virus AI subtipe H5N1 berdasarkan uji serologi. Hasil amplifikasi DNA menunjukan bahwa gen Mx pada burung air liar dijumpai pada panjang fragmen 100 bp, sama seperti gen Mx yang terdapat ayam, bebek dan entok. Akan tetapi gen Mx pada burung air liar tidak menunjukan adanya polimorfisme seperti yang dijumpai pada ayam. Hal ini membuktikan bahwa keberadaan gen Mx pada burung air liar dalam mempengaruhi mekanisme resistensi terhadap paparan virus AI subtipe H5N1 berbeda dengan yang terdapat pada ayam.
Kata kunci : gen Mx, burung air liar, PCR-RFLP, virus AI subtipe H5N1, resistensi
IDENTIFICATION AND ANALYSIS POLYMORPHISM OF
MX GENE IN WILD WATER BIRDS FROM
“
PULAU DUA
” IN
EXPOSURE OF AVIAN INFLUENZA VIRUS SUBTYPE H5N1.
ABSTRACT
subtype H5N1 virus affected in wild waterbirds using the Polymerase Chain Reaction-Restriction Fragment Length Polymorphism (PCR-RFLP) method with the primer pairs F2 and NE-R2/R and the RsaI restriction enzyme. DNA samples were obtained from eight species of wild waterbirds with positive exposure to antibody anti AI virus subtype H5N1 based on serologis test. DNA amplification results showed that the Mx gene in wild waterbirds was found in a 100 bp fragment, which is the same as the Mx gene found in chickens, duck and muscovy. However, unlike chickens, the Mx gene in wild waterbirds did not show any polymorphism. This study proves that Mx-based resistance to AI virus subtype H5N1 were different between wild waterbirds and chickens.
Keywords: Mx gene, wild waterbirds, PCR-RFLP, AI virus subtype H5N1, resistance
PENDAHULUAN
Secara alami setiap individu hewan memiliki kemampuan untuk mempertahankan diri dari paparan mikroorganisme yang masuk ke dalam tubuhnya. Mekanisme pertahanan diri berkaitan dengan sistem imun nonspesifik dan sistem imun spesifik dalam memberi respon terhadap masuknya organisme patogen ke dalam tubuh individu hewan. Respon imun nonspesifik merupakan respon imun yang dapat mendeteksi adanya benda asing dan melindungi tubuh dari kerusakan yang diakibatkannya. Respon imun ini ada sejak lahir, tidak memiliki target tertentu dan bekerja dalam waktu yang relatif singkat sesaat setelah masuknya organisme patogen. Respon imun spesifik adalah sistem kekebalan yang dapat menghancurkan patogen yang lolos dari sistem kekebalan non-spesifik (Radji 2010; Mayer 2011).
Respon imun nonspesifik mencakup reaksi inflamasi/peradangan, protein antivirus (interferon), sel Natural Killer (NK), dan sistem komplemen. Respon imun spesifik mencakup, kekebalan humoral yang dibentuk melalui produksi antibodi oleh limfosit B (sel plasma) dan kekebalan selular yang dibentuk melalui produksi limfosit T yang teraktivasi. Kedua respon imun tersebut mampu menyebabkan organisme bersifat resisten atau rentan terhadap mikroorganisme yang telah berhasil masuk ke dalam tubuhnya (Dodds 2002; Radji 2010; Mayer 2011).
Protein Mx diketahui memiliki aktivitas antiviral pada sejumlah hewan, khususnya terhadap virus influenza. Fungsi khusus protein Mx sebagai pertahanan terhadap virus terjadi dari hasil interaksi langsung antar bagian terminus karboksil protein Mx yang dimiliki organisme dengan virus patogen yang spesifik spesies (Haller et al. 1995; Lee dan Vidal 2002). Protein Mx yang disandikan oleh gen Mx diaktivasi oleh interferon yang terbentuk akibat masuknya virus ke dalam tubuh organism (Haller et al. 1998; Ko et al. 2002; Haller dan Kochs 2002)
Gen Mx merupakan salah satu gen yang mengatur resistensi terhadap myxovirus (Ding et al. 2006; Yin et al. 2010; Schusser et al. 2011). Gen Mx pada beberapa organisme berperan dalam mengendalikan kemampuan hewan tersebut dalam membentuk mekanisme pertahanan terhadap virus AI. Keberadaan gen ini juga akan menentukan sifat resisten atau rentan pada organisme yang mengalami invasi virus. Sifat resisten akan ditentukan oleh kemampuan protein Mx menghambat replikasi virus (Haller et al. 1998; Ko et al. 2002; Haller dan Kochs 2002; Li dan Lu 2011).
Sejumlah peneliti telah melakukan eksplorasi untuk memperoleh informasi peran dan fungsi gen Mx pada berbagai jenis hewan dan jenis virus yang dihadapinya. Staeheli et al. (1988) melaporkan gen Mx1 pada mencit dikontrol oleh dua alel yaitu Mx1+ dan Mx1-. Alel Mx1+ sangat resisten terhadap influenza dan ditemukan umumnya pada mencit liar, sebaliknya alel Mx1- (sangat rentan terhadap virus influenza) umum dijumpai pada mencit laboratorium. Gen Mx1+ yang menyandikan protein Mx1 diaktivasi oleh interferon (IFN)-alpha dan beta, dan protein Mx1 yang dihasilkan berfungsi menghambat replikasi virus di dalam inti sel (Ewald et al. 2011; Schusser et al. 2011; Zhou et al. 2013).
Protein Mx pada berbagai organisme terletak pada jaringan yang berbeda, terlokalisasi di dalam sel dan memainkan aktivitas antiviral yang berbeda. Protein Mx1 pada nukleus sel mencit terutama akan menghambat replikasi Orthomyxovirus (termasuk virus influenza), sedangkan protein Mx2 pada sitoplasma mencit akan menghambat virus somatitis vesikuler (VSV). Pada manusia, MxA memperlihatkan aktivitas melawan virus pada spektrum yang sangat luas meliputi Orthomyxovirus (termasuk influenza), Rhabdovirus, Bunyavirus dan Paramyxovirus (Lee dan Vidal 2002; Saint-Jean dan Perez-Prieto 2007; Yin et al. 2010)
Protein Mx pada sapi dan anjing disandikan oleh Gen Mx1 dan Mx2 yang berperan sebagai antivirus spesifik terhadap VSV (Ellinwood et al. 1998; Nakamura et al. 2005). Protein Mx pada mencit dibentuk oleh gen Mx1 yang memberi respon terhadap virus influenza (Chang et al. 1990). Pada babi protein Mx1 yang disandikan oleh gen Mx1 berperan melawan infeksi VSV (Asano et al. 2002). Hasil penelitian Saint-Jean dan Perez-Prieto (2007) menjelaskan, gen Mx pada salmon yang menyandikan protein Mx akan berperan terhadap masuknya virus infeksi pankreatik nekrosis (IPNV) dan virus infeksi salmon anemia virus (ISAV).
Gen Mx pada unggas pertama kali diketahui sejak tahun 1980. Haller et al.
dan terdiri dari 705 asam amino. Sifat resistensi terhadap virus AI ditemukan pada exon 13 nukleotida nomor 1.892 (asam amino 631) yang mengalami mutasi satu basa tunggal (single nukleotide mutation) (Haller et al. 1998; Livant et al. 2007).
Mutasi titik pada asam amino 631 gen Mx merupakan mutasi substitusi yang bersifat transisi. Mutasi tersebut terjadi dari basa nukleotida Adenin (A) menjadi basa nukleotida Guanin (G). Mutasi pada gen Mx tersebut menyebabkan perubahan triple kodon ke-631 yaitu dari menyandikan asam amino asparagin (AAT) berubah menjadi serin (AGT). Adanya asam amino asparagin pada nukleotida ke-1.892 tersebut menunjukan sifat resistensi (daya tahan) ayam terhadap AI yang dikenal sebagai alel Mx+. Mutasi asam amino dari asparagin menjadi serin akan menyebabkan ayam bersifat rentan terhadap infeksi virus AI, dikenal sebagai alel Mx-. (Haller et al. 1998; Li et al. 2007; Li dan Lu 2011). Adanya polimorfisme panjang fragmen pada gen Mx ayam, memberikan respon berbeda terhadap infeksi virus AI subtipe H5N1.
Penelitian yang telah dilakukan oleh Maeda (2005) terhadap sejumlah ayam-ayam lokal yang berasal dari 12 negara di Asia menunjukkan bahwa pada semua populasi ayam lokal memiliki alel Mx+ dan alel Mx- yang akan mempengaruhi kemampuan setiap unggas terhadap infeksi AI. Unggas tersebut menunjukkan dua jenis mekanisme pertahanan tubuh yaitu resisten atau rentan terhadap virus AI. Unggas-unggas lokal pada sejumlah negara menunjukkan frekuensi alel Mx+ yang lebih besar dibandingkan alel Mx- termasuk di Indonesia. Ayam lokal asli Indonesia memiliki frekuensi alel Mx+ lebih besar (63%) daripada alel Mx- (37%). Populasi ayam yang rentan terhadap flu burung memiliki frekuensi alel Mx- yang lebih besar dari 50%, populasi ayam tersebut dijumpai di negara Korea, Taiwan dan Srilangka (Sartika et al. 2011).
Hasil penelitian terhadap 485 ekor ayam dari 15 galur ayam lokal Indonesia yang mampu bertahan hidup dari wabah AI dengan menggunakan PCR-RFLP menunjukkan, ayam jenis cemani memiliki resistensi terhadap virus flu burung yang paling tinggi dengan frekuensi alel Mx+ (alel A) sebesar 89 %, dibandingkan galur ayam lainnya. Ayam kapas memiliki resistensi terendah dengan frekuensi alel Mx+ (alel A) hanya 35 %. Hasil penelitian menunjukkan 89% dari populasi cemani yang diteliti memiliki alel Mx+ sehingga memiliki sifat resisten terhadap invasi virus AI subtipe H5N1 yang lebih tinggi dibandingkan ayam dari galur yang lain (Sulandari et al. 2009).
Pada unggas air/burung air, penelitian Gen Mx baru terbatas dilakukan pada unggas air domestik seperti bebek/itik (Berlin et al. 2008; Dillon dan Runstadler 2010; Zhu et al. 2010; Li dan Lu 2011; Purwanto 2012). Hasil identifikasi Gen Mx yang dilakukan Purwanto (2012) dengan menggunakan primer yang didisain oleh Sironi et al. (2010) berdasarkan urutan genom Mx (kode akses No. DQ788615) membuktikan adanya gen Mx pada bebek/itik Pitala asal Sumatera Barat dengan panjang pita 50 bp. Meskipun demikian belum pernah dilaporkan hasil penelitian terkait keberadaan gen Mx pada burung air liar, khususnya yang berasal dari Indonesia.
METODE
Ekstraksi dan Purifikasi DNA total dari Sampel Darah
Ekstraksi DNA dari sampel darah burung air dilakukan dengan modifikasi metode yang dikembangkan oleh Sambrook et al. (1989). Sampel darah segar sebanyak 250-500 µl ditempatkan pada tabung 1,5 ml. Larutan penyangga pelisis A (0,32 sukrosa, 1% v/v Triton X-100, 5 mM MgCl2 dan 10 mM Tris HCl)
ditambahkan pada sampel dengan volume yang sama dan disentrifugasi satu menit dengan kecepatan 3.000 g Supernatan dibuang dan endapan ditambah dengan 200 µl larutan penyangga pencuci B atau Rinse Buffer (75 mM NaCl, 50 mM EDTA) dan dikocok sampai endapan larut. Larutan penyangga C atau Digestion Buffer
(200mM NaCl, 50 mM Tris HCl, 100 mM EDTA, 1% SDS) ditambahkan sebanyak 500 µl, 15 µl Proteinase K (10 mg/ml), dan 5 µl RNAse (10 mg/ml). Inkubasi dilakukan pada suhu 55oC selama 1 malam (16 Jam).
Sampel diambil dari inkubator, ditambahkan fenol 500 µl dan dihomogenisasi selama 30 menit sehingga tercampur semua. Sentrifugasi dilakukan 2 menit dengan kecepatan 6.000 g, sehingga larutan terpisah menjadi dua. Supernatan yang diperoleh dipindahkan ke tabung 1,5 ml yang baru. Larutan fenol dan kloroform ditambahkan dengan volume sama ke dalam supernatan, dihomogenisasi selama 30 menit dan disentrifugasi pada 6.000 rpm selama 2 menit. Supernatan diambil dan dipindahkan ke tabung 1,5 ml yang baru dan ditambahkan larutan Chloroform Iso Amil Alkohol (CIAA) µl 24:1, dihomogenisasi selama 30 menit. Sentrifugasi dilakukan 2 menit dengan kecepatan 13.000 rpm. Supernatan diambil dan ditambahkan larutan etanol absolut 2 kali volumenya. Agar larutan tercampur merata dilakukan homogenisasi dengan cara membolakbalikan tabung membentuk angka 8 selama 10 menit atau sampai gumpalan putih berupa benang-benang DNA terbentuk dan disimpan dalam freezer (30 menit).
Langkah berikutnya adalah sentrifugasi pada 13.000 rpm selama 2 menit. Larutan etanol absolut dibuang dan diganti larutan etanol 70% (600 µl), disentrifugasi pada 6.000 g selama 2 menit. Selanjutnya larutan etanol 70% dibuang dengan menggunakan pipet, pellet dikeringanginkan selama 30 menit. Larutan D atau TE (10 mM Tris HCl, 1 mM EDTA dan 0,02 mg/ml RNAse) ditambahkan sebanyak 50 µl, dilakukan homogenisasi sebentar, dan diinkubasi pada suhu 37oC selama 15 menit.
Elektroforesis DNA
Hasil isolasi dideteksi dengan elektroforesis DNA pada gel agarose 2%. Gel dilarutkan dalam buffer TBE, ditambah ethidium bromide 0,4 ug/ml, kemudian dicetak pada gel plat yang terdapat sisir. Gel selanjutnya diletakkan pada chamber
menit. Gel dibaca pada UV transluminator dan hasilnya didokumentasikan. Sampel DNA yang diperoleh disimpan pada suhu 4oC (Sambrook et al. 1989). Pengecekan DNA juga dilakukan dengan menggunakan 12% Polyacrylamide Gel Electroforesis (PAGE) vertikal selama 4 jam dengan tegangan listrik 160 V dan visualisasi dilakukan dengan silver stainning (Zein dan Sulandari 2008).
Pita-pita yang terbentuk menunjukkan adanya DNA pada burung-burung air liar. Hasil elektroforesis yang membentuk pita jelas dan terang selanjutnya akan digunakan untuk analisa PCR-RFLP.
Analisis PCR-RFLP
Amplifikasi untuk mendeteksi single nucleotide polymorphisme (SNP) pada posisi nukleotida ke 1892 dari sekuen DNA gen Mx yang berada pada exon ke 13 (substitusi asam amino pada posisi 631 protein Mx) dilakukan dengan menggunakan metoda PCR-RFLP (Restriction Fragment Length Polymorphysme) yang dikembangkan oleh Seyama et al. (2006). Primer yang digunakan adalah primer forward NE-F2 (5‟-AGAG GAATCTGATTGCTCAGGCGTGTA-3‟) dan primer reverse NE-R2/R (5‟-CAGAGG AATCTGATTGCTCAGGCGAATA-3‟) (Seyama et al. 2006). Amplifikasi dilakukan pada mesin PCR GeneAmp PCR System 9700, Applied Biosystem. Kondisi PCR yang digunakan, predenaturasi pada suhu 94oC selama 5 menit, denaturasi pada suhu 94oC selama1 menit, penempelan primer pada suhu 61oC selama 1 menit, elongasi pada suhu 72oC selama 1 menit dengan siklus sebanyak 35 kali, dan final ekstensi pada suhu 72oC selama 5 menit (Sulandari et al. 2009). Komposisi campuran reaksi PCR dibuat dalam 30 µl terdiri dari : 1 µl sampel DNA; 1 µl primer forward; 1 µl primer reverse; 4 µl MgCl2; 0,6 µl dNTP; 3 µl 10x PCR Buffer; dan 0,25 µl
Taq-Polymerase.
Fragmen DNA yang berhasil diamplifikasi dipotong dengan enzim restriksi RsaI, yaitu enzim yang memotong molekul DNA pada tempat yang spesifik (bersusunan basa GT↓AC). Pengecekan hasil pemotongan produk PCR-RFLP dilakukan dengan menggunakan 12% Polyacrylamide Gel Electroforesis (PAGE) vertikal selama 4 jam, tegangan listrik 160 V. Visualisasi dilakukan dengan silver stainning (Zein dan Sulandari 2008).
HASIL DAN PEMBAHASAN
Jumlah sampel DNA burung-burung air liar yang digunakan untuk PCR-RFLP sebanyak 80 sampel DNA dari 8 spesies burung air liar yang menunjukkan pita yang terang dan jelas, 10 sampel DNA bebek, 10 sampel DNA entok, dan 10 sampel DNA ayam.
Hasil PCR yang dilakukan terhadap DNA delapan spesies burung air liar, ayam, bebek dan entok asal sekitar CAPD menggunakan primer F2 dan NE-R2/R menurut Seyama et al. (2006) diperoleh produk PCR dengan panjang pita 100 bp (Gambar 5). Hasil ini menunjukan bahwa di dalam tubuh burung air liar penetap di kawasan CAPD terdapat gen Mx yang sama seperti yang dimiliki oleh ayam dan entok.
Gambar 5. Hasil PCR DNA burung air liar di CAPD dengan primer F2, NE-R2/R.
Ket. 1=ayam,2=bebek, 3=entok, 4-5=Ardeola speciosa, 6=Ardea sp, 7= Casmerodius albus, 8=Phalacrocorax sp, 9= Egretta garzetta, 10=
E. intermedia, 11=Bubulcus ibis, 12= Nycticorax nycticorax
Sejumlah penelitian untuk menentukan keberadaan gen Mx dengan menggunakan primer Seyama et al. (2006) telah dilakukan pada beberapa ayam dan bebek. Hasil penelitian menunjukan bahwa keberadaan Gen Mx dapat dilihat dari panjang produk PCR yang dihasilkan yaitu sebesar 100 bp (Zein dan Sulandari 2008; Sulandari et al. 2009; Luan et al. 2010; Zhi-Quan et al. 2010; Sartika et al. 2011). Penelitian yang dilakukan Purwanto (2012) dengan menggunakan primer menurut Sironi et al. (2010), menunjukan bahwa gen Mx pada bebek pitala teramplifikasi dengan panjang fragment 50 bp. Hasil penelitian yang dilakukan Sulandari et al. (2009) menggunakan pasangan primer menurut Ommeh et al. (2009), menunjukkan produk amplifikasi dengan panjang pita 101 bp untuk membuktikan adanya gen Mx.
Keberadaan gen Mx pada burung air liar penetap di CAPD menunjukkan bahwa gen tersebut telah teraktivasi akibat masuknya virus ke dalam tubuh burung. Kerja gen Mx sangat dipengaruhi oleh interferon yang diinduksi pada saat terjadi invasi virus ke dalam tubuh. Pada saat virus menginfeksi sebuah sel, asam nukleat virus akan menginduksi perangkat genetik sel untuk membentuk interferon. Interferon akan dikeluarkan ke dalam cairan ektraseluler dan berikatan dengan reseptor pada membran plasma sel-sel di sekitar invasi virus. Pada saat itu juga interferon akan mengaktivasi gen Mx untuk bekerja memproduksi protein
M 1 2 3 4 5 6 7 8 9 10 11 12
300 bp
200 bp
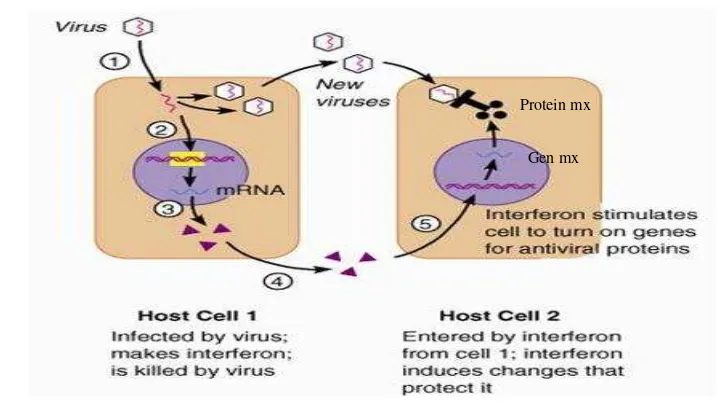
antivirus (Staehelli et al. 1985; Haller et al. 1998; Qu et al. 2009; Ewald et al. 2011; Zhou et al. 2013). Ikatan antara reseptor dengan membran plasma sel tersebut akan merangsang sel inang membentuk enzim-enzim, enzim akan mengaktivasi gen untuk menyusun protein antivirus yang akan menghambat reproduksi virus (Gambar 6).
Gambar 6. Mekanisme kerja interferon dalam tubuh organisme
(Sumber : http://immune0system.files.wordpress.com/2010/
04/mekanisme- interferon-melawan-virus.jpg)
Sejumlah penelitian mengenai virus AI yang menginvasi tubuh hewan menunjukkan peran interferon yang menghambat proses replikasi virus AI pada sel inang. Interferon bekerja dengan cara mengaktivasi gen Mx, selanjutnya gen Mx akan memproduksi protein Mx yang berperan melakukan aktivitas antivirus. Protein Mx ini akan memberi respon inang terhadap masuknya virus AI subtipe H5N1 dan akan menentukan sifat resisten atau rentan inang terhadap paparan virus AI tersebut (Haller dan Kochs 2002; Lee dan Vidal 2002; Hsiang et al 2009; Jewell et al 2010; Ewald et al. 2011).
Gen Mx pada sejumlah hewan telah dieksplorasi sejak tahun 1980-an, Staeheli et al. (1985) mendeteksi peran gen Mx sebagai pengatur mekanisme resistensi terhadap infeksi virus influenza pada mencit pada tahun 1979. Sejak saat itu penelitian terhadap gen Mx mulai berkembang dan dilakukan terhadap manusia (Zurcher et al. 1992; Haller et al. 1995; Kochs dan Haller 1999), domba (Charleston dan Stewart 1993), bebek (Bazzigher et al. 1993), mencit (Johannes et al. 1997), kerbau (Ellinwood et al. 1998), ikan salmon (Kim et al. 2000), babi (Asano et al. 2002), anjing (Nakamura et al. 2005). Pada hewan-hewan tersebut gen Mx berperan sebagai antiviral terutama pada virus-virus tertentu yang termasuk pada famili Orthomyxoviridae, Paramyxoviridae, Bunyaviridae, Rhabdoviridae, Picornaviridae, Togaviridae (Tabel 2)
Tabel 2. Lokalisasi intraseluler dan spektrum antivirus protein Mx pada hewan
Spesies Gen Kode akses Lokasi Antivirus spesifik
Jenis virus Famili
Manusia MxA P20591 Sitoplasma Influenza, thogoto Orthomyxoviridae Vesicular somatitis Rhabdoviridae