KARAKTERISTIK DOMINANSI MONYET HITAM SULAWESI (Macaca nigra) DI CAGAR ALAM TANGKOKO- BATUANGUS,
SULAWESI UTARA
SAROYO
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI DISERTASI DAN SUMBER INFORMASI
Dengan ini saya menyatakan bahwa disertasi Karakteristik Dominansi Monyet Hitam Sulawesi (Macaca nigra) di Cagar Alam Tangkoko-Batuangus, Sulawesi Utara adalah karya saya sendiri dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir disertasi ini
RINGKASAN
Penelitian ini bertujuan untuk menentukan sistem sosial monyet hitam Sulawesi (Macaca nigra) melalui kajian karakteristik dominansi di Cagar Alam Tangkoko-Batuangus. Penelitian dilaksanakan bulan Januari–Desember 2004. Kelompok monyet yang diamati adalah Kelompok Rambo II (KRII) dan Kelompok Rambo I (KRI) dengan ukuran masing-masing pada awal pengamatan sebesar 51 ekor.
Hasil pengamatan selama setahun, didapat nisbah jantan dan betina dewasa berkisar dari 1:1,9-1:7,5. Nilai nisbah ditentukan oleh filopatri betina dan faktor migrasi jantan. Daerah jelajah KRII seluas 232 ha, dan jelajah hariannya sepanjang 1,8-4,1 km dengan rata-rata 3,05 ± 0,58 km. Daerah jelajah KRI seluas 119 ha, dan jelajah hariannya sepanjang 1,7-3,3 km dengan rata-rata 2,09 ± 0,34 km.
Dominansi ditentukan melalui pengamatan interaksi agonistik pada enam jantan KRII, enam jantan KRI, dan 14 betina KRI. Interaksi agonistik intrakelompok hanya mencakup 1,8% dari total waktu harian. Interaksi agonistik meliputi agresi ringan sebesar 59,5% dan agresi berat sebesar 40,5%. Agresi lunak diikuti dengan rekonsiliasi sebesar 75,8% dan tingkah laku arah diri/TAD sebesar 24,2%, sedangkan agresi berat diikuti dengan rekonsiliasi sebesar 33,3% dan TAD sebesar 66,7%. Angka rekonsiliasi pada agresi lunak sebesar 75,8% dan pada agresi berat sebesar 33,3%. Inisiasi rekonsiliasi dilakukan oleh individu dominan sebesar 59,5% dan 40,5% dilakukan oleh individu subordinan.
Hierarki dominansi jantan bersifat linear sempurna. Jika terjadi ketidakseimbangan hubungan antarjantan, hierarki dapat bersifat tidak linear. Ketidakseimbangan hubungan antarjantan terjadi pada saat terjadi perubahan tingkah laku agresif sampai terbentuknya hierarki baru. Hierarki dapat berubah karena faktor perubahan tingkah laku agresif jantan dan faktor migrasi. Dengan demikian, interaksi sosial jantan bersifat asimetris yang ditunjukkan melalui hubungan dominansi.
Koalisi dapat menurunkan peran dominansi untuk akses terhadap pakan, tempat, dan kawin, serta dapat meningkatkan status individu dalam hierarki dominansinya. Koalisi dapat terdiri dari 2-6 ekor monyet. Koalisi dapat bertipe konservatif jika dua individu dominan melawan individu subordinan, tipe jembatan jika individu dominan bekerja sama dengan individu subordinan melawan individu peringkat menengah, dan tipe revolusioner jika individu-individu subordinan melawan individu dominan. Koalisi berbentuk konservatif sebesar 32,3%, berbentuk revolusioner sebesar 35,5%, dan berbentuk jembatan sebesar 32,3%.
Migrasi terjadi pada individu jantan dewasa dan individu jantan pada saat mencapai umur dewasa. Empat jantan KRII bermigrasi ke KRI selama bulan Oktober dan November. Pada saat migrasi terjadi, terdapat 15 ekor betina KRI sedang estrus, sedangkan pada KRII hanya terdapat satu betina estrus. Migrasi jantan tampaknya disebabkan ketertarikan secara seksual dengan betina yang reseptif pada kelompok lain.
Menelisik merupakan aktivitas yang mencakup 12,3% dari total waktu aktivitas harian monyet. Betina lebih banyak menelisik jantan daripada jantan menelisik betina. Menelisik berperan dalam: pendekatan dalam tingkah laku seksual, sarana rekonsiliasi, ikatan sosial, mencegah tertelisik pindah tempat, dan sebagai sarana TAD. Terdapat kecenderungan jantan dominan mendekati jantan subordinan, betina dominan mendekati betina subordinan, dan betina mendekati jantan. Interaksi antarkelompok dapat bersifat agonistik maupun afiliatif, bahkan terjadi perkawinan.
ABSTRACT
The social system of Sulawesi macaques is considered as ‘egalitarian’. Generally, social system patterns relate to females. The aims of this study were to investigate whether ‘egalitarian’ social relationships also apply to Sulawesi crested black macaque males and the function of dominance to social interactions. For this, the male and female dominance pattern, dominance hierarchy changes, and its function to social interactions were studied. This study has been conducted on two identified and habituated groups at Tangkoko-Batuangus Nature Reserve, North Sulawesi from January to December 2004. Data were collected by focal animal sampling of six males of Rambo II, six males of Rambo I and 14 females of Rambo II. Linearity of male hierarchy was calculated with the Landau’s index of linearity (h). The dynamic of dominance hierarchy was monitored by observation of changes in the direction of agonistic interactions. Results showed that: 1) during periods with stable male relationships, the male dominance hierarchy was linear and transitive, whereas during unstable periods, it was non-linear; 2) during one year, α -male replacements occurred two times on Rambo II and one times on Rambo I; 3) the female dominance hierarchy was imperfectly linear; 4) migration process only occurred in adult and growing adult males; 5) dominance determined priority for accessing to safety and comfortable place and food; 6) coalition might play a role in decreasing domination of dominant individual; 7) to decrease agonistic frequency and intensity, the Sulawesi black macaques used several behavioral mechanisms, such as allogrooming and postconflict affiliation; 8) females groomed males more frequently than males did, grooming among females was unrelated to their ranking; 9) there was a tendency for high rank males to approach lower rank males, for high rank females to approach lower rank females and for females to approach males; 10) intergroup interaction included agonistic and affiliative interactions between the two group members and intergroup mating; and 11) natural sexual behavior was varied; 12) mating frequency of the males was not influenced by their ranking; 13) there were mate choice factors and mating strategy of lower rank to avoid intervention by high rank. From the results, it can be concluded that contrary to females that have egalitarian society, in male Sulawesi crested black macaques, the dominance hierarchy is usually linear and male relationships are non-‘egalitarian’. When defining a social system in primates, it should thus always be clear to which sex the definition is referring to.
Key words: Dominance characteristics, Sulawesi crested black macaques (Macaca nigra), Tangkoko-Batuangus Nature Reserve
KARAKTERISTIK DOMINANSI MONYET HITAM SULAWESI (Macaca nigra) DI CAGAR ALAM TANGKOKO-BATUANGUS, SULAWESI
SAROYO
Disertasi
sebagai salah satu syarat untuk memperoleh gelar Doktor pada
Program Studi Primatologi
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
BOGOR 2005
Judul Disertasi : Karakteristik Dominansi Monyet Hitam Sulawesi (Macaca nigra) di Cagar Alam Tangkoko-Batuangus, Sulawesi Utara Nama : Saroyo
Disetujui Komisi Pembimbing
Dr. Ir. Sri Supraptini Mansjoer Ketua
Prof. Dr. Rudy C. Tarumingkeng Anggota
Dr. Ir. Dedy Duryadi Solihin, DEA Anggota
Prof. Kunio Watanabe, Ph.D. Anggota
Diketahui
Ketua Program Studi Primatologi Dekan Sekolah Pascasarjana
Dr. Ir. Sri Supraptini Mansjoer Prof. Dr. Ir. Syafrida Manuwoto, M.Sc.
Ujian Tanggal: 11-11-2005
Tanggal Ujian:
Lulus tanggal:
Tanggal Lulus:
Puji syukur penulis panjatkan ke hadirat Allah SWT yang atas berkat dan rahmat-Nya penulis dapat menyelesaikan disertasi ini. Disertasi ini berjudul Karakteristik Dominansi Monyet Hitam Sulawesi (Macaca nigra) di Cagar Alam Tangkoko-Batuangus, Sulawesi Utara, berdasarkan hasil penelitian yang dilaksanakan dari bulan Januari sampai dengan Desember 2004.
Penyelesaian disertasi ini tidak terlepas dari peran dan bantuan berbagai pihak. Oleh karena itu penulis menghaturkan terima kasih dan penghargaan kepada Komisi Pembimbing, yaitu Ibu Dr. Ir. Sri Supraptini Mansjoer, Bapak Prof. Dr. Rudy C. Tarumingkeng, Bapak Dr. Ir. Dedy Duryadi Solihin, DEA, serta Bapak Prof. Kunio Watanabe, Ph.D. yang telah meluangkan waktu, tenaga, dan pikiran selama persiapan, penelitian, dan penyusunan disertasi ini. Ucapan terima kasih juga disampaikan kepada Dekan Sekolah Pascasarjana Institut Pertanian Bogor beserta staf dan pegawai, Rektor Universitas Sam Ratulangi Manado beserta staf dan pegawai, Kepala Pusat Studi Satwa Primata LP-IPB beserta staf dan pegawai, serta PEMDA Sulawesi Utara yang telah memberi kesempatan dan bantuan untuk menunaikan tugas belajar di PS Primatologi SPS IPB. Dengan selesainya penelitian di CA. Tangkoko-Batuangus, penulis menyampaikan banyak terima kasih kepada Kepala Balai KSDA Sulawesi Utara beserta staf dan pegawai atas bantuan dan kerjasama selama penelitian. Terima kasih juga disampaikan kepada PEMKOT Bitung, Lurah, dan masyarakat Batuputih yang banyak membantu penulis selama di lokasi penelitian. Banyak pihak telah membantu pendanaan penelitian, sehingga penulis menyampaikan terima kasih kepada DIRJEN DIKTI DEPDIKNAS, Director of The Rufford Small Grant dan Primate Research Institute Kyoto University atas bantuan dana untuk penelitian. Kepada keluarga, sahabat, dan mahasiswaku penulis sampaikan terima kasih atas doa dan dorongannya.
Semoga disertasi ini bermanfaat, terutama sebagai salah satu landasan konservasi monyet hitam Sulawesi dan pengelolaan kawasan konservasi.
Penulis dilahirkan di Boyolali pada tanggal 24 Juni 1968. Pendidikan menengah ditempuh di SMA I Boyolali Program A2 dan lulus pada tahun 1987. Lulus pendidikan sarjana dari Program Studi Pendidikan Biologi pada Fakultas Keguruan dan Ilmu Pendidikan Universitas Sebelas Maret Surakarta pada tahun 1992. Melanjutkan pendidikan Pra-S2 di Program Studi Biologi Institut Teknologi Bandung pada tahun 1993 melalui program Calon Tenaga Akademik Baru (CTAB), dilanjutkan dengan pendidikan magister tahun 1994 pada program studi yang sama pada bidang Biologi Perkembangan dengan beasiswa Tim Manajemen Program Doktor (TMPD) dari DIKTI. Lulus pendidikan magister pada tahun 1996. Menempuh program doktor di Institut Pertanian Bogor pada Program Studi Primatologi pada tahun 2001 dengan nomor mahasiswa B066010011.
Pada tahun 1996, penulis diangkat sebagai staf pengajar di Universitas Sam Ratulangi, Manado di Fakultas Peternakan, Program Studi Ilmu Produksi Ternak. Pada tahun 2000 penulis dipindahkan sebagai staf pengajar di Fakultas Matematika dan Ilmu Pengetahuan Alam, Program Studi Biologi. Beberapa kursus telah diambil, antara lain Method in Microbiology, Natural Product Chemistry, Field Course in Primate Behavior and Ecology Tangkoko Nature Reserve, Kursus Singkat Biologi Molekuler, Pelatihan Penulisan Artikel Ilmiah Tingkat Nasional, Kursus Pekerti, Kursus AA, Penataran Calon Penulis Buku Ajar Perguruan Tinggi, Tinjil Island Primate Research Project Field Course: Primate Behavior and Ecology, Pelatihan Kiat-Kiat Penyusunan Proposal Penelitian Berdaya Saing Tinggi dan Penelusuran Informasi Ilmiah Mutakhir.
Mulai bekerja untuk monyet hitam Sulawesi di Cagar Alam Tangkoko-Batuangus dari tahun 1998 sebagai peneliti maupun pembimbing penelitian mahasiswa. Beberapa hasil penelitiannya telah dipublikasikan pada beberapa jurnal, yaitu Zootek, Jurnal Ilmiah Sains, Eugenia, dan Jurnal Primatologi Indonesia.
DAFTAR ISI
DAFTAR GAMBAR ... xii
MATERI DAN METODE PENELITAN Tempat dan Waktu ... 29
Materi dan Alat ... 29
Metode Penelitian ... 29
HASIL DAN PEMBAHASAN Karakteristik Kelompok ... 45
Ukuran dan Komposisi Kelompok ... 45
Pertumbuhan Kelompok ... 46
Daerah Jelajah dan Jelajah Harian ... 52
Aspek Karakteristik Dominansi ... 58
Dominansi Jantan ... 58
SIMPULAN, SARAN, DAN REKOMENDASI ... 114
DAFTAR PUSTAKA ... 115
LAMPIRAN ... 122
GLOSARIUM ... 130
Halaman 1 Perbandingan hubungan sosial primata betina keempat tipe
kompetisi ... 18
2 Tiga teori untuk menjelaskan pembentukan koalisi (Widdig 2000) .. 22
3 Komposisi KRII (Januari 2004) ... 45
4 Komposisi KRI (Juli 2004) ... 45
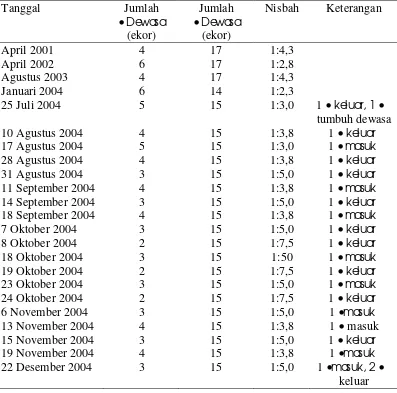
5 Rekapitulasi kelahiran, kematian, imigrasi, dan emigrasi KRII ... 46
6 Komposisi KRII pada akhir pengamatan (Desember 2004) ... 47
7 Komposisi KRI pada akhir penelitian (Desember 2004) ... 49
8 Dinamika dalam nisbah jantan dan betina dewasa KRII ... 51
9 Dinamika dalam nisbah jantan dan betina dewasa KRI ... 52
10 Jarak jelajah harian KRII ... 54
11 Jarak jelajah harian KRI ... 54
12 Karakteristik jantan dewasa KRII ... 59
13 Rekapitulasi hasil pertemuan agresif dan arah ancaman atau tunduk pada jantan dewasa KRII ... 60 14 Hasil interaksi agonistik dan arah ancaman atau tunduk pada jantan dewasa KRI ………... 60 15 Dinamika hierarki jantan KRII ... 62
16 Dinamika hierarki jantan KRI ... 63
17 Komposisi individu dewasa KRII dan KRI pada bulan Oktober awal 68 18 Perbandingan lama proses migrasi pada dua jantan KRII ... 70
19 Matriks sosiometrik hasil interaksi agresif antarbetina KRII pada awal pengamatan (Juni 2004) ... 72 20 Matriks sosiometrik hasil interaksi agresif antarbetina KRII pada akhir pengamatan (Agustus 2004) ... 72 21 Rekapitulasi pendekatan antarjantan KRII ... 76
22 Hasil uji pakan pada enam jantan KRII ... 80
23 Interaksi agonistik antarindividu KRII ... 84
24 Hasil pengamatan koalisi ... 88
25 Rekapitulasi pendekatan KRII ... 94
26 Perkawinan antarkelompok KRI dan KRII ... 100
27 Tingkah laku seksual dan variasinya ... 103
28 Rekapitulasi variasi tingkah laku kawin ... 104
29 Rekapitulasi frekuensi kawin jantan KRII ... 106
DAFTAR GAMBAR
Halaman
1 Monyet hitam Sulawesi ... 6
2 Pengaruh ukuran kelompok pada kompetisi perebutan ... 16
3 Pengaruh ukuran kelompok pada kompetisi pertandingan ... 17
4 Peta lokasi penelitian ... 30
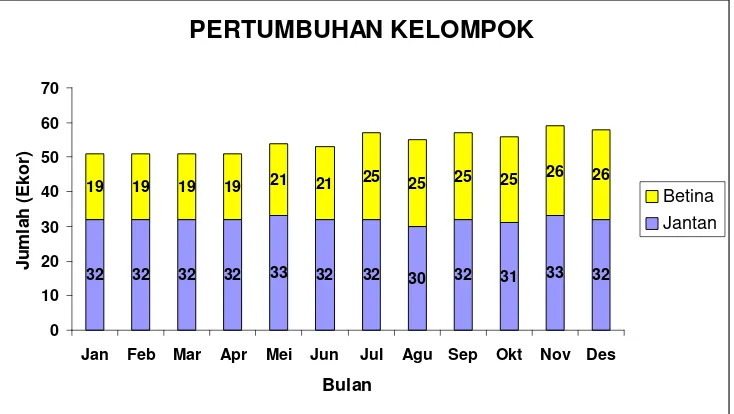
5 Pertumbuhan KRII selama tahun 2004 ... 47
6 Daerah jelajah KRII dan KRI ... 53
7 Tipe-tipe vegetasi pada daerah jelajah KRII dan KRI ... 55
8 Hubungan segitiga antarjantan KRII ... 61
9 Migrasi jantan dewasa yang teramati selama penelitian ... 65
10 Dinamika dalam hierarki dominansi betina dewasa KRII pada awal pengamatan (Juni) dan akhir pengamatan (Agustus) ... 74 11 Pergeseran daerah jelajah KRII selama tahun 2004 ... 77
DAFTAR LAMPIRAN
Halaman
1 Dinamika dalam hierarki dominansi KRII ... 122
2 Dinamika dalam hierarki dominansi KRI ... 125
3 Proses migrasi jantan KRII ... 127
PENDAHULUAN
Latar Belakang
Sulawesi termasuk salah satu pulau dalam daerah Wallacea. Daerah Wallacea adalah daerah peralihan antara Zoogeografi Oriental dan Zoogeografi Australia (Coates dan Bishop 2000). Satwa Sulawesi merupakan yang paling khas di seluruh Indonesia terutama di antara mammalia. Dari 127 spesies mammalia asli, 62% bersifat endemik (Whitten et al. 1987). Monyet (genus Macaca) adalah satwa khas Oriental yang daerah penyebarannya sampai di Sulawesi (Bynum 1999). Nenek moyang monyet Sulawesi menyeberang ke Pulau Sulawesi pada pertengahan Plestosen dari Pulau Kalimantan atau Jawa (Whitten et al. 1987). Pada saat ini terdapat enam spesies monyet endemik Sulawesi, yaitu M. nigra, M. nigrescens, M. hecki, M. tonkeana, M. maura (M. maurus), dan M. ochreata (Groves 2001), atau tujuh spesies menurut Fooden (1969 dalam Bynum 1999) dengan tambahan M. brunnescens.
Penelitian tentang aspek sosial monyet Sulawesi belum banyak dilakukan, kecuali untuk M. maurus. Beberapa penelitian tentang aspek sosial pada M. maurus, antara lain: organisasi sosial (Watanabe dan Matsumura 1996), afiliasi pascakonflik (Matsumura 1996), hubungan dominansi rileks antarbetina (Matsumura 1996), faktor-faktor yang mempengaruhi proksimitas antaranggota kelompok selama makan, bergerak, dan istirahat (Matsumura dan Okamoto 1997), serta sejarah hidup dan demografi (Okamoto et al.
2000).
Matsumura (1998) didapatkan hasil bahwa hubungan sosial antarbetina dewasa bersifat egaliter atau mempunyai pola dominansi rileks.
Hubungan sosial monyet Sulawesi betina dikategorikan sebagai dominansi rileks atau sistem sosial egaliter seperti pada monyet Barbary (M. sylvanus), monyet bonnet (M. radiata), dan monyet stumptail (M. arctoides) (Matsumura 1998). Hubungan sosial yang berbeda, yang disebut despotik ditemukan pada sistem sosial monyet Jepang (M. fuscata), monyet Rhesus (M. mullata), dan beruk (M. nemestrina) (Matsumura 1998).
Masyarakat egaliter atau despotik pada satwa primata umumnya ditentukan dari interaksi antarbetina. Sistem sosial M. nigra (monyet hitam Sulawesi) betina bilamana termasuk sistem sosial egaliter seperti pada M. maurus atau despotik belum banyak diketahui. Sistem sosial monyet hitam Sulawesi jantan bilamana mempunyai pola yang sama atau berbeda dengan sistem sosial pada betina juga belum banyak diketahui. Demikian juga jika monyet hitam Sulawesi mempunyai sistem sosial seperti pada monyet Sulawesi lainnya, karakteristik dominansi pada monyet hitam Sulawesi juga masih harus diteliti. Oleh karena itu studi yang mendalam tentang sistem sosial monyet hitam Sulawesi, terutama tentang pola dominansi menjadi sangat penting.
Monyet hitam Sulawesi menempati habitat hutan hujan tropis primer dan sekunder di beberapa lokasi di semenanjung utara Pulau Sulawesi dan beberapa pulau satelitnya (Lee dan Kussoy 1999, Supriatna dan Wahyono 2000). Penelitian Reed et al. (1997) pada tahun 1994 selama enam minggu pada Kelompok Rambo (97 ekor), menunjukkan bahwa dominansi pada jantan berbentuk linear dan transitif di antara enam jantan dewasa. Frekuensi dan intensitas agresi di antara jantan berkorelasi kuat dengan jarak peringkat. Jantan dari seluruh peringkat secara signifikan menunjukkan pula tingkat agresivitas yang lebih tinggi terhadap betina, yang reseptif secara seksual dibandingkan dengan betina pada fase yang lain. Kesimpulan hasil penelitian menunjukkan bahwa monyet hitam Sulawesi jantan mempunyai organisasi sosial yang sama dengan kelompok banyak jantan-banyak betina seperti pada spesies Macaca lainnya.
Kelompok kecil yang disebut Kelompok Gila mempunyai ukuran 13 ekor. M. maurus
mempunyai ukuran kelompok yang lebih kecil, yaitu 20-30 (Matsumura 1998).
Penelitian aspek dominansi pada spesies lain, misalnya pada monyet Jepang (M. fuscata) didapatkan hasil bahwa hierarki di antara individu dewasa dan remaja dari kedua jenis kelamin bersifat linear sempurna (Chaffin et al. 1995). Hasil yang sama juga didapatkan pada monyet Rhesus (M. mullata) dan monyet stumptail (M. arctoides) (Chaffin et al. 1995).
Umumnya penelitian tentang dominansi terkonsentrasi pada jantan dewasa dibandingkan dengan pada betina dewasa (Chalmer 1980). Secara umum, pada satwa primata yang menunjukkan sifat dominansi, jantan dewasa mempunyai peringkat dominansi yang lebih tinggi dibandingkan dengan betina dewasa, dengan kekecualian pada talapoin (Miopithecus talapoin) (Chalmer 1980) dan Lemur (Rowe 1996). Dominansi pada betina lebih terkait dengan kesempatan untuk mendapatkan pakan dan keberhasilan dalam reproduksi (Matsumura 1998, Range dan Noe 2002). Betina yang lebih dominan mempunyai kesempatan untuk mendapatkan pakan yang lebih banyak daripada betina yang kurang dominan, terutama untuk sumber pakan yang terbatas (Chalmer 1980). Betina peringkat tinggi mempunyai angka kelahiran dan angka kesintasan anak yang lebih tinggi dibandingkan dengan betina peringkat yang lebih rendah (Chalmer 1980).
Perbedaan ukuran kelompok pada kondisi habitat di Cagar Alam Tangkoko-Batuangus kemungkinan akan berpengaruh terhadap karakteristik dominansi monyet hitam Sulawesi. Oleh karena itu masalah tersebut menarik untuk diteliti.
Tujuan Penelitian
1) Mengkaji karakteristik dominansi sebagai dasar dalam penentuan sistem sosial monyet hitam Sulawesi.
2) Menentukan peran dominansi dalam berbagai interaksi sosial monyet hitam Sulawesi.
Ruang Lingkup
Penelitian ini merupakan penelitian eksploratif bidang tingkah laku satwa primata di alam. Salah satu ciri penelitian ini adalah tidak dapat dibuat hipotesis dan apabila dipaksakan maka hipotesisnya akan berbetuk sangat umum (Suratmo 2002). Penelitian ini dititikberatkan pada salah satu aspek sistem sosial monyet hitam Sulawesi, yaitu aspek karakteristik dominansi. Aspek pendukung yang juga diamati meliputi karakteristik kelompok yang mencakup ukuran dan komposisi kelompok, pertumbuhan kelompok, serta daerah jelajah dan jelajah harian. Aspek peran dominansi dalam interaksi sosial monyet meliputi aspek tingkah laku sosial dan aspek reproduksi. Luaran penelitian diharapkan dapat dijadikan rekomendasi bagi otoritas manajemen kawasan konservasi dan organisasi yang bergerak dalam bidang konservasi ex-situ.
Manfaat Penelitian
1) Menambah informasi tentang karakteristik dan peran dominansi dalam berbagai interaksi sosial sebagai dasar dalam penentuan sistem sosial monyet hitam Sulawesi. 2) Dasar dalam pengelolaan populasi monyet hitam Sulawesi melalui pengelolaan
tingkah laku kelompok, terutama yang berkaitan dengan sistem sosial bagi otoritas manajemen CA Tangkoko-Batuangus dan kawasan konservasi lainnya di Sulawesi Utara dan lembaga/organisasi konservasi ex-situ dalam pengelolaan kelompok, sehingga kelestarian spesies ini dapat dipertahankan.
Permasalahan:
- Ada tidaknya perubahan sistem sosial dengan berubahnya ukuran kelompok
- Sistem sosial jantan sama atau berbeda dengan sistem sosial pada betina
- Karakteristik dominansi pada M. nigra
Analisis dan evaluasi hasil didasarkan pada kajian teoritis dan fakta di lapangan.
Koleksi Data: - Pola dominansi
- Peran dominansi dalam interaksi sosial
Jantan Betina
Penyimpulan
Sistem sosial monyet Sulawesi dikategorikan sebagai sistem sosial egaliter
- Penelitian serupa banyak dilakukan pada M. maurus
TINJAUAN PUSTAKA
Biologi Monyet Hitam Sulawesi Klasifikasi
Monyet hitam Sulawesi atau dalam bahasa lokal disebut yaki (Gambar 1) merupakan satu dari enam jenis monyet Sulawesi. Monyet Sulawesi meliputi dare (M. maura/maurus), yaki (M. nigra), dihe (M. nigrescens), dige (M. hecki), boti (M. tonkeana), hada (M. ochreata) (Groves 2001).
Gambar 1 Monyet hitam Sulawesi
Menurut Corbet dan Hill (1992) serta Collinge (1993), monyet hitam Sulawesi
dimasukkan ke dalam Bangsa Primates, Suku Cercopithecidae, Marga Macaca, dan Jenis
Macaca nigra. Pemberian nama yang salah untuk jenis ini, misalnya Celebes black ape
bahasa daerah, monyet ini disebut yaki (Tonsea, Bacan), wolai (Tondano), dan bolai
(Mongondow) (Supriatna dan Wahyono 2000).
Penyebaran
Monyet hitam Sulawesi tersebar di semenanjung utara Pulau Sulawesi di sebelah timur Sungai Onggak Dumoga dan Gunung Padang yang berbatasan dengan penyebaran
M. nigrescens (Groves 2001). Di Sulawesi Utara, monyet ini dapat dijumpai di CA Tangkoko-Batuangus, CA DuaSudara, SM Manembonembo, Kotamobagu, dan Modayak. Monyet ini juga ditemukan di CA Gunung Lokon, CA Gunung Ambang, CA Tanggale, Pulau Manado Tua, dan Pulau Talise (Lee dan Kussoy 1999, Supriatna dan Wahyono 2000). Monyet ini telah diintroduksi ke Pulau Bacan di Maluku Utara dan populasinya lebih banyak dibandingkan dengan populasi aslinya. Penelitian oleh Rosenbaum et al. (1998) didapatkan hasil bahwa kerapatan monyet hitam Sulawesi di Cagar Alam Gunung Sibela di Pulau Bacan mencapai 170,3 ekor/km2, sedangkan di hutan yang sudah terganggu mencapai 133,4 ekor/km2.
Pertelaan
Monyet hitam Sulawesi mempunyai ciri tubuh yang mudah dibedakan dengan jenis lainnya. Panjang tubuh betina dewasa 445-550 mm dan pada jantan dewasa 520-570 mm, ekor sepanjang 25 mm (Rowe 1996). Bobot tubuhnya 7-15 kg (Supriatna dan Wahyono 2000). Rambut yang menutupi seluruh tubuh berwarna hitam kelam, namun bagian belakang (punggung) dan paha berwarna lebih terang dibandingkan dengan bagian lain (Bynum 1999, Supriatna dan Wahyono 2000). Wajah berwarna hitam dan tidak ditumbuhi rambut. Moncong jauh lebih menonjol dibandingkan dengan monyet Sulawesi
melahirkan 65 bulan, interval kelahiran 18 bulan, dan lama hidup dapat mencapai 18 tahun (Rowe 1996). Bayi berambut putih pada wajah, lengan, dan bagian bawah badan. Warna ini akan berubah perlahan-lahan menjadi hitam sebelum umur empat sampai enam bulan (Kinnaird 1997). Indeks intermembran sebesar 94 dan bobot otak dewasanya mencapai 94,4 g (Rowe 1996).
Ekologi 1. Habitat
Monyet hitam Sulawesi hidup di hutan tropis primer dan sekunder (Rowe 1996) di Sulawesi Utara. Mereka dapat dijumpai pada hutan primer atau sekunder dataran rendah hingga dataran tinggi sampai 2.000 dpl (Supriatna dan Wahyono 2000). Ular python (Python reticulatus) merupakan predator alaminya (Kilner 2001).
2. Pakan
Seperti pada jenis monyet lain, monyet hitam Sulawesi termasuk omnivor. Monyet ini memakan berbagai bagian tumbuhan, mulai dari daun, pucuk daun, bunga, biji, buah, dan umbi, serta beberapa jenis serangga, moluska, dan invertebrata kecil. Terdapat lebih dari 145 jenis buah yang dimakan. Di CA Tangkoko-Batuangus, sekelompok monyet sering ditemukan di tepi laut untuk mencari moluska sebagai salah satu sumber pakannya (Supriatna dan Wahyono 2000). Pohon ara merupakan salah satu sumber pakan bagi monyet yang paling melimpah. Di CA Tangkoko-Batuangus dan CA DuaSudara terdapat 45 jenis. Buah ara merupakan 20 persen dari total pakan monyet (Kinnaird 1997). Beberapa jenis serangga yang dimakan monyet ini meliputi tawon, rayap, ulat dalam gulungan daun Pongamia sp., lebah, semut, dan belalang (Saroyo 2002b).
3. Tingkah Laku
Monyet hitam Sulawesi merupakan spesies diurnal, terestrial, dan arboreal yang
dan jelajah hariannya dapat mencapai 5 km (Supriatna dan Wahyono 2000). Berdasarkan penelitian pendahuluan (Saroyo 2002b, Saroyo et al. 2004b), Kelompok Rambo II di CA Tangkoko-Batuangus mempunyai daerah jelajah yang cukup sempit yaitu 59 ha dan jarak jelajah hariannya 2.839 ± 423,6 m. Monyet hitam Sulawesi aktif pada siang hari (diurnal), dan sore hari menjelang tidur mereka memilih tumbuhan yang rimbun. Tidur dilakukan pada tajuk tinggi pepohonan yang ditinggalkan menjelang matahari terbit
untuk segera turun mencari makan. Monyet ini menghabiskan setengah waktunya di tanah dan setengahnya lagi di pepohonan dengan bergelantungan dari satu pohon ke pohon lain untuk mencari makan (Kinnaird 1997).
Status Konservasi
Monyet hitam Sulawesi dilindungi oleh Pemerintah RI dengan SK Menteri Pertanian 29 Januari 1970 No. 421/Kpt/um/8/1970, SK Menteri Kehutanan 10 Juni 1991 No. 301/Kpts-II/1991 dan Undang-Undang No. 5 tahun 1990 (Supriatna dan Wahyono 2000). Dalam daftar yang dikeluarkan IUCN, spesies ini digolongkan sebagai satwa genting dan dicantumkan dalam Appendix II CITES (Supriatna dan Wahyono 2000).
Berdasarkan kecenderungan hilangnya hutan, status spesies ini perlu diubah menjadi kritis (Lee et al. 2002).
Populasi
Habitat monyet hitam Sulawesi telah banyak mengalami penyusutan akibat penebangan dan pembukaan lahan perkebunan (Lee et al. 2001). Saat ini mereka telah kehilangan 60% habitatnya dari 12.000 km2 menjadi 4.800 km2, dan hanya menempati areal seluas 2.750 km2 dalam kawasan konservasi (Supriatna dan Wahyono 2000).
Berdasarkan penelitian Lee dan Kussoy (1999), kerapatan populasi monyet hitam Sulawesi di Tangkoko sebesar 58,0 ekor/km2, di Pulau Talise 21,5 ekor/km2, di Manembonembo (22,8 ekor/km2), dan di Manado Tua 34,0 ekor/km2.
Penelitian MacKinnon dan MacKinnon pada tahun 1978, kerapatan populasi monyet di CA Tangkoko-Batuangus-DuaSudara sebesar 300 ekor/km2, dan berdasarkan penelitian Sugardjito et al. pada tahun 1988, kerapatannya menjadi 76 ekor/km2 (Rosenbaum et al. 1998). Penelitian Rosenbaum et al. pada tahun 1994 menunjukkan bahwa kerapatan populasi monyet sebesar 66,7 ekor/km2 (Rosenbaum et al. 1998). Berdasarkan hasil survei Kyes et al. (2002) pada tahun 1999-2002, kerapatannya sebesar 39,8 ekor/km2.
Cagar Alam (CA) Tangkoko-Batuangus
Sulawesi Utara mempunyai beberapa kawasan perlindungan, baik cagar alam, taman nasional, maupun suaka margasatwa. CA Tangkoko-Batuangus terletak di Kecamatan Bitung Utara, Kota Bitung, Sulawesi Utara. Kawasan ini ditetapkan sebagai cagar alam berdasarkan GB No. 6 Stbl 1919, tanggal 12 Februari 1919 (KSDA Sulawesi Utara). Luas cagar alam ini 3.196 ha, terletak pada 1o30’-1o34’N dan 125o10’-125o81’E (Sunarto et al. 1999). Sebagian dari wilayahnya digunakan sebagai daerah wisata alam, dan sampai saat ini belum diketahui pengaruh kegiatan tersebut terhadap kehidupan monyet. Beberapa kelompok monyet, seperti KRII dan KRI mempunyai daerah jelajah yang melalui daerah Taman Wisata Alam (TWA) Batuputih (Saroyo 2002a). CA Tangkoko-Batuangus menyatu dengan CA DuaSudara, TWA Batuangus, dan TWA Batuputih. Luas keempat kawasan ini 8.718 ha (KSDA Sulawesi Utara 2002).
CA Tangkoko-Batuangus meliputi beberapa tipe habitat (Whitten et al. 1987, Saroyo 2003b) sebagai berikut ini.
1) Pantai, yaitu formasi barringtonia, meliputi tumbuhan: Barringtonia asiatica
(bitung), Pandanus sp. (pandan), Callophyllum soulattri (nyamplung), Morinda citrifolia (mengkudu daun besar), Morinda bracteata (mengkudu daun kecil), Cycas rumphii (pakis haji), Terminalia catappa (ketapang), Hibiscus tiliaceus (waru laut),
Ixora sp. (soka/suwing), Erythrina sp. (dadap), Pongamia pinnata (lakehe), Leea indica (momaling biasa), Kleinhovia hospita (bintangar), dan Barringtonia acutangula (salense).
2) Hutan sekunder dengan pohon khas Cocos nucifera (kelapa), Mangifera indica
3) Semak-semak yang tersusun atas Imperata cylindrica (alang-alang), Saccharum spontaneum (glagah), Eupatorium odoratum, Lantana camara (tembelekan), Piper aduncum (sirih hutan), dengan diselingi pohon Macaranga sp. (binunga), dan Melia azedarach (mindi/bugis).
4) Semak-semak lain yang tersusun atas rumput Cyrtococcum oxyphyllum yang hampir tertutup oleh Piper aduncum (sirih hutan), Melia azedarach (mindi/bugis),
Macaranga sp. (binunga), Lantana camara (tembelekan), Pterospermum diversifolium (wolo daun besar), Alstonia sp. (kayu telur), Laportea sp. (kemaduh), dan Tectona grandis (jati).
5) Hutan primer dengan berbagai jenis pohon, yang menyolok antara lain Ficus sp.
(ara), Leea indica (momaling biasa), Palaquium amboinense (nantu), Ardisia sp.
(kayu anoa), Eugenia sp. (gora hutan), Garuga floribunda (kayu kambing),
Dracontomelum dao (rao), Livistona rotundifolia (woka), dan Baringtonia acutangula (salense).
6) Di atas ketinggian 800 m sudah merupakan vegetasi pegunungan.
Organisasi Sosial Monyet Hitam Sulawesi
Bangsa Primates merupakan salah satu Bangsa dalam Kelas Mammalia yang hidup dalam suatu kelompok sosial. Hidup bersosial memberikan beberapa keuntungan untuk akses terhadap pakan, proteksi terhadap predator, akses untuk kawin, dan mempermudah dalam pemencaran keturunan (Collinge 1993). Beberapa terminologi berikut merujuk pada Collinge (1993). Suatu Kelompok Sosial tersusun dari satwa-satwa yang berinteraksi pada suatu basis reguler. Primata mampu mengenal satu dengan yang lain dan menggunakan lebih banyak waktu dengan anggota kelompoknya. Struktur Sosial menunjukkan bentuk fisik kelompok berkaitan dengan kelompok umur dan jenis kelamin, serta hubungan interaksi satu dengan lainnya. Organisasi Sosial merupakan ekspresi
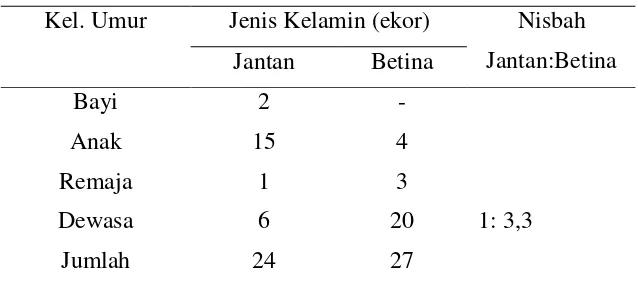
Berdasarkan sistem klasifikasi sosioseksual, struktur sosial monyet hitam Sulawesi termasuk kelompok banyak jantan-banyak betina. Di alam, monyet hitam Sulawesi hidup dalam kelompok besar, yaitu 20-70 ekor (Supriatna dan Wahyono 2000). Mereka hidup dalam kelompok dengan nisbah (rasio) jantan dan betina dewasa 1:3,4 (Rowe 1996). Nisbah jantan dan betina ini merupakan fungsi dari pola emigrasi jantan dan filopatri betina (Napier dan Napier 1985). Filopatri betina berarti bahwa betina tetap
berada dalam kelompok kelahirannya. Masyarakat monyet ini berpusat pada keluarga betina, sementara jantan keluar dari kelompok kelahirannya (Matsumura 1998).
Interaksi Sosial
Tingkah laku sosial monyet hitam Sulawesi sangat terorganisir dan kompleks. Jantan membentuk hierarki kekuasaan. Jantan paling dominan ditandai dengan ukuran tubuh besar dan paling kuat memegang prioritas dalam mendapatkan pakan dan pasangan (Kinnaird 1997). Betina dewasa menanggung sebagian besar tugas membesarkan anak, sehingga jantan sempat membersihkan segala parasit dari rambut tubuh mereka dan
membantu kaum betina memperkuat ikatan sosial dengan anggota lainnya. Kaum remaja melewatkan waktu dengan berjumpalitan dan berkejar-kejaran atau bergumul dengan sebayanya. Meringis lebar merupakan senyuman mengajak bermain-main bukan menantang berkelahi (Kinnaird 1997).
Interaksi sosial dibedakan menjadi dua tipe dasar yaitu kompetitif (antagonistik) dan kooperatif (positif atau afiliatif). Beberapa penelitian tentang interaksi sosial antaranggota kelompok pada satwa primata, antara lain: dominansi pada monyet Jepang (M. fuscata) (Chaffin et al. 1995), hubungan dominansi betina dewasa pada monyet Jepang di alam (Nakamichi et al. 1995), hubungan proksimitas pada monyet Jepang (Nakamichi 1996), dominansi pada monyet Assam (M. assamensis ) (Bernstein dan Cooper 1999), menelisik, ikatan sosial, dan agonistik pada monyet Rhesus (M. mullata) (Matheson dan Bernstein 2000), dan hubungan kekeluargaan dan dominansi betina pada
suatu kelompok besar (97 individu). Hasil penelitian menunjukkan bahwa dominansi pada jantan berbentuk linear dan transitif di antara enam jantan dewasa. Frekuensi dan intensitas agresi di antara jantan berkorelasi kuat dengan jarak peringkat. Jantan dari seluruh peringkat secara signifikan menunjukkan pula tingkat agresivitas yang lebih tinggi terhadap betina yang secara seksual reseptif daripada terhadap betina pada fase yang lain. Kesimpulan hasil penelitian menunjukkan bahwa jantan monyet hitam Sulawesi mempunyai organisasi sosial yang sama dengan pada spesies Macaca lainnya (Reed et al. 1997).
1. Interaksi Kompetitif (Antagonistik)
Jika hewan hidup dalam kelompok yang stabil, pesaing superior secara konsisten
unggul terhadap pesaing inferior (Collinge 1993). Pasangan individu tersebut mempunyai hubungan dominansi. Hubungan dominansi dapat diukur melalui hasil perkelahian dalam pertemuan agresif antara dua individu atau arah sikap mengancam atau tunduk. Agresi mencakup agresi ringan (mengancam dengan membuka mulut, mengancam dengan suara, menerjang) dan agresi berat (mengusir, menendang, mencakar, menggigit) (Perry 1996).
1.1. Hierarki Dominansi
Menurut Martin dan Bateson (1999), pada banyak spesies primata, hubungan di
antara sepasang individu berbentuk asimetris. Satu individu secara konsisten akan mengusir individu lain pada saat mereka berkompetisi untuk suatu sumber yang bermanfaat, misalnya pakan, tempat, atau kawin, atau secara sederhana satu individu menghindar pada saat mereka bertemu. Jika sejumlah kejadian dicatat untuk setiap pasangan dalam kelompok, seringkali menjadi jelas bahwa satu individu cenderung untuk mengusir seluruh individu lainnya, sedangkan individu lain akan diusir yang lainnya. Keseluruhan susunan individu dominan dan subordinan dalam kelompok dikenal sebagai hierarki dominansi (Martin dan Bateson 1999). Dominansi merupakan hal penting dalam tingkah laku sosial pada spesies hewan yang hidup berkelompok dengan
suatu kelompok, sehingga harus bersaing satu dengan yang lain dari waktu ke waktu untuk akses terhadap sumber-sumber di atas. Kompetisi ini menghasilkan kemantapan dalam hierarki dominansi yang mencakup perbedaan tingkah laku individu berdasarkan jenis kelamin, ukuran, umur, status, dan kekerabatan (Swindler 1998).
Terdapat beberapa bentuk hierarki dominansi. Jika seluruh individu dalam kelompok dapat disusun dalam urutan dominansi yang pasti (misalnya C dominan
terhadap A, A dominan terhadap D, D dominan terhadap E, dan E dominan terhadap B), maka hierarki dominansinya linear (Martin dan Bateson 1999). Dalam kenyataan, hanya beberapa hierarki ini yang linear sempurna. Kadang-kadang dominansi balik dapat terjadi, jika subordinan mengalahkan individu yang secara normal lebih dominan (Martin dan Bateson 1999). Lebih jauh untuk hierarki linear sempurna, seluruh hubungan diad harus asimetris. Pada beberapa kelompok, dua atau lebih individu dapat mempunyai status yang sama. Pada hierarki linear sempurna, seluruh kemungkinan hubungan triad harus transitif (jika A dominan terhadap B dan B dominan terhadap C, maka A harus dominan terhadap C) (Martin dan Bateson 1999).
Pada saat ini kelompok monyet di CA Tangkoko-Batuangus tidak sebesar pada tahun 1994. Kelompok Rambo telah terfragmentasi menjadi dua kelompok yang lebih
kecil, yaitu KRI (±60 ekor) dan KRII (51 ekor) (Saroyo 2002a). Pola dominansi betina
pada monyet hitam Sulawesi digolongkan sebagai nepotistik-toleran (Slater 2002). Pola ini berarti bahwa betina bersifat filopatri dan terdapat kerjasama antarkerabat dalam kompetisi.
1.2. Pola Dominansi
menunjukkan sedikit toleransi terhadap subordinan, menghasilkan hierarki dominansi yang jelas dan umumnya merupakan masyarakat despotik. Sebaliknya, jika kompetisi antarkelompok tinggi dan resiko predasi rendah, maka individu dominan akan lebih toleran terhadap subordinan, untuk mencegah mereka meninggalkan kelompok dan masuk ke kelompok lain. Pola hubungan yang kurang kompetitif ini akan menghasilkan masyarakat yang lebih toleran dan hubungan dominansi lebih egaliter (Hemelrijk 1999).
Hasil penelitian yang sangat baik tentang peringkat, keberhasilan dalam reproduksi, dan dispersal telah dilakukan pada monyet Rhesus (M. mullata) di alam oleh Berard (1993). Hasil penelitian menunjukkan bahwa keberhasilan reproduksi pada jantan anggota tetap kelompok dan peringkat tinggi menurun selama periode penelitian. Di pihak lain terjadi peningkatan keberhasilan dalam perkawinan pada jantan emigran dari tahun ke tahun. Walaupun peringkat seekor jantan anggota tetap kelompok tersebut meningkat, tetapi keberhasilan dalam perkawinanya menurun. Jantan peringkat tinggi harus memutuskan untuk tetap tinggal dalam kelompok tetapi mempunyai keberhasilan dalam perkawinan rendah, atau bermigrasi ke kelompok lain menjadi peringkat rendah
tetapi keberhasilan dalam perkawinannya tinggi.
Aspek dominansi pada betina kurang mendapatkan perhatian untuk diteliti (Chalmer 1980). Secara umum, pada primata yang menunjukkan sifat dominansi, jantan dewasa mempunyai peringkat dominansi yang lebih tinggi dibandingkan dengan betina dewasa, dengan kekecualian pada talapoin (Miopithecus talapoin). Dominansi pada betina lebih terkait dengan kesempatan untuk mendapatkan pakan dan keberhasilan dalam reproduksi (Koenig 2002). Betina yang lebih dominan mempunyai kesempatan untuk mendapatkan pakan yang lebih banyak daripada betina yang yang kurang dominan, terutama untuk sumber pakan yang terbatas. Betina dengan peringkat tinggi mempunyai angka kelahiran dan angka kesintasan anak yang lebih tinggi dibandingkan dengan betina yang mempunyai peringkat lebih rendah (Chalmer 1980).
menelisik dengan betina lain dua kali lebih banyak daripada yang dilakukan dengan jantan; betina cenderung menelisik kepada yang lebih dominan. Pada langur hanuman (Semnopithecus entellus) yang mempunyai sistem sosioseksual berpusat pada jantan tunggal, hubungan hierarki betina menunjukkan ketidakstabilan, inkonsisten, individualistik, dan angka koalisi agonistik rendah (Borries 1993).
1.3. Tipe Kompetisi
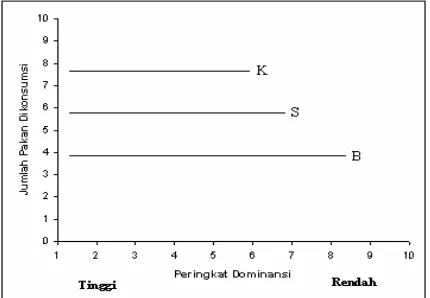
Kompetisi untuk suatu sumber dibedakan menjadi dua tipe dasar, yaitu Kompetisi perebutan atau kompetisi tidak langsung dan kompetisi pertandingan atau kompetisi langsung (http://life.bio.sunysb.edu/bio359/ 4_26_02.html). Kompetisi perebutan terjadi jika hewan harus berbagi sumber yang terbatas tetapi tidak mempunyai cara untuk membatasi akses individu lain terhadap sumber tersebut. Seluruh individu dalam kelompok bersama-sama akan mengalami pengurangan pakan. Efek kompetisi tipe ini akan meningkat dengan meningkatnya ukuran kelompok (Gambar 2). Kompetisi tipe ini untuk pakan terjadi jika ketersediaan pakan sangat sedikit dan sangat jarang.
Gambar 2 Pengaruh ukuran kelompok pada kompetisi perebutan (K = ukuran kelompok kecil; S = ukuran kelompok sedang; B = ukuran kelompok besar)
terjadi jika pakan tersebar tidak merata. Sebagai contoh, pohon kecil yang sedang berbuah merupakan sumber yang menyebabkan rejim kompetisi tipe pertandingan.
Gambar 3 Pengaruh ukuran kelompok pada kompetisi pertandingan (K = ukuran kelompok kecil; S = ukuran kelompok sedang; B = ukuran kelompok besar)
Terdapat empat tipe kombinasi yang menghasilkan tipe masyarakat yang berbeda (http://life.bio.sunysb.edu/bio359/4_26_02.html). Untuk lebih jelasnya, Tabel 1 di bawah menunjukkan perbandingan hubungan sosial primata betina pada keempat tipe kompetisi. 1) Jika kompetisi perebutan intrakelompok lebih penting daripada kompetisi pertandingan, akan menghasilkan masyarakat tipe dispersal-egaliter. Masyarakat tipe ini ditemukan pada monyet howler, monyet bajing Costa Rica, dan gorilla. 2) Jika kompetisi pertandingan intrakelompok lebih penting daripada kompetisi
perebutan intrakelompok dan kompetisi pertandingan interkelompok, akan menghasilkan masyarakat yang despotik. Pada masyarakat tipe ini, betina cenderung untuk membentuk aliansi dengan sanak dan kerabatnya. Tipe ini terutama terjadi pada primata frugivor dan mereka disebut residen-nepotistik. Sebagai contoh, misalnya pada monyet (Macaca), baboon, dan monyet capuchin.
Tabel 1 Perbandingan hubungan sosial primata petina keempat tipe kompetisi (http://life.bio.sunysb.edu/bio359/4_26_02.html)
Kategori Rejim kompetitif Respon sosial
Pertandingan intrakelompok
Pertandingan
interkelompok
Filopatri betina
Peringkat betina
Dispersal-Egaliter
Residen-Egaliter
Rendah Tinggi Ya Egaliter
Residen-Nepotistik
Tinggi Rendah Ya Nepotistik
dan despotik
Residen- Nepotistik-Toleran
Tinggi Tinggi Ya Nepotistik
dan toleran
3) Pada beberapa spesies, kompetisi pertandingan interkelompok dapat lebih penting daripada kompetisi intrakelompok. Akibatnya hubungan antaranggota kelompok menjadi lebih egaliter dan kurang nepotistik. Tipe masyarakat seperti ini disebut residen-egaliter. Sebagai contoh, misalnya guenon, monyet patas, dan langur
hanuman.
4) Pada beberapa spesies, kompetisi pertandingan interkelompok dan intrakelompok sama pentingnya, akan menghasilkan masyarakat residen-nepotistik-toleran.
1.4. Pemilihan Pasangan Kawin
Pemilihan pasangan kawin didefinisikan sebagai pola tingkah laku pada satu jenis kelamin yang dapat meningkatkan probabilitas perkawinan fertil dengan individu tertentu pada jenis kelamin yang berbeda (Soltis et al. 1997). Hasil penelitian Soltis et al. (1997) menunjukkan terdapat pemilihan betina oleh jantan dan pemaksaan seksual jantan oleh betina pada M. fuscata. Proksimitas betina terhadap jantan dan agresi jantan terhadap betina berkorelasi dengan peningkatan keberhasilan perkawinan. Kebanyakan agresi jantan dihasilkan dari peningkatan lama waktu dalam proksimitas oleh betina, dan sebagian disebabkan oleh pemaksaan seksual (Soltis et al. 1997).
2. Interaksi Kooperatif (Positif, Afiliatif) 2.1. Rekonsiliasi (Reuni)
pihak ketiga pascakonflik yang didefinisikan sebagai kontak afiliatif pascakonflik antara individu-individu yang bertengkar dan individu-individu di sekitarnya (Call et al. 2002).
Rekonsiliasi pada satwa primata, suatu interaksi afiliatif pascakonflik di antara individu yang bertengkar mempunyai dua fungsi: (1) untuk memperbaiki kerusakan hubungan karena agresi, dan (2) untuk mereduksi ketidakpastian pascakonflik dan tekanan pada individu tersebut (Kutsukake dan Castles 2001). Hipotesis terintegrasi
untuk rekonsiliasi terkait dengan fungsi tersebut melalui argumentasi bahwa kerusakan hubungan karena agresi akan menyebabkan tekanan yang tinggi, sehingga menimbulkan usaha untuk berekonsiliasi untuk menurunkan tekanan tersebut (Kutsukake dan Castles 2001).
Beberapa hipotesis telah diajukan oleh beberapa ahli tingkah laku. Beberapa studi mengindikasikan bahwa rekonsiliasi berfungsi untuk memperbaiki kerusakan hubungan sosial karena agresi (hipotesis perbaikan hubungan) (Kutsukake dan Castles 2000). Sebagai contoh, rekonsiliasi akan mengurangi kemungkinan korban agresi mengalami serangan berikutnya yang dilakukan oleh penyerang pertama atau individu lain, serta
meningkatkan toleransi untuk sumber pakan (Kutsukake dan Castles 2000).
Pada kenyataannya agresi tidak selalu diikuti oleh rekonsiliasi. Beberapa peneliti berusaha menjelaskan faktor-faktor yang mempengaruhi terjadinya rekonsiliasi. Mereka mengemukakan bahwa kualitas hubungan individu yang bertengkar merupakan faktor penting untuk menentukan terjadinya rekonsiliasi. Rekonsiliasi sering terjadi mengikuti agresi di antara individu yang bertengkar. Individu-individu tersebut mempunyai hubungan dengan nilai biologis yang tinggi, suatu fungsi keuntungan kebugaran yang dapat dihasilkan dari hubungan tersebut (hipotesis nilai hubungan) (Kutsukake dan
Fungsi rekonsiliasi yang lain berperan untuk mereduksi tekanan pascakonflik. Pada satwa primata terdapat tingkah laku arah diri/TAD seperti menggaruk, menelisik diri, dan menguap berkaitan dengan situasi tertekan (Cord dan Thurnheer 1993). Sebagai contoh, proksimitas oleh individu yang dominan dapat meningkatkan laju TAD pada monyet ekor panjang dan olive baboon (Papio anubis), sedangkan menelisik silang dapat mereduksi TAD dan detak jantung pada tertelisik. Oleh karena itu, proksimitas dalam
rekonsiliasi menyebabkan reduksi ketidakpastian dan menurunkan kondisi tertekan (Cord dan Thurnheer 1993).
Aureli (1997) mengajukan hipotesis terintegrasi untuk rekonsiliasi. Beberapa studi menunjukkan bahwa agresivitas tidak hanya berpengaruh pada korban, pada penyerang juga akan meningkatkan TAD. Ini membuktikan bahwa kondisi tertekan pascakonflik tidak terbatas pada korban, tetapi juga dialami oleh penyerang. Sebagaimana terjadi pada korban, pada penyerang juga terjadi penurunan laju TAD pada rekonsiliasi (Aureli 1997).
Tidak semua spesies satwa primata menunjukkan terjadinya afiliasi pascakonflik.
Pada sebagian besar spesies Cercopithecine yang hidup dalam kelompok sosial kompleks, mereka membentuk afiliasi (Call et al. 2002). Pada red-bellied tamarin
(Saguinus labiatus), agresi yang terjadi tidak merusak hubungan antaranggota kelompok, sehingga rekonsiliasi tidak diperlukan (Schaffner dan Caine 2002).
2.2. Aliansi atau Koalisi
Konflik di antara dua individu hewan sering kali diinterpretasikan sebagai kompetisi terhadap sumber yang terbatas (Widdig et al. 2000). Hasil interaksi agresif
Terdapat beberapa tipe aliasi tergantung pada hasilnya. Aliansi spesifik sumber ditemukan pada baboon savana di alam bebas (Collinge 1993). Jantan tua dan jantan peringkat rendah sering membentuk koalisi untuk mengusir hewan dominan yang sedang kopulasi dengan betina estrus. Koalisi ini bersifat resiprok terhadap kesempatan untuk kawin. Pada masyarakat banyak jantan-banyak betina, misalnya pada monyet Jepang dan monyet Rhesus, betina membentuk matrilineal yang besar bekerja sama dalam
interaksi agonistik. Pada spesies dengan jantan tetap berada dalam kelompok kelahirannya (filopatri), seperti chimpanzee dan monyet colobus merah, jantan-jantan yang berkerabat bekerja sama melawan individu dominan atau untuk berkompetisi dalam akses terhadap betina estrus. Aliansi antara jantan dan betina terjadi terutama pada saat musim kawin dalam konteks pasangan kawin. Istilah aliansi xenofobik menunjukkan bahwa seluruh anggota kelompok bersama-sama mempertahankan teritorialnya dari kelompok lain (Collinge 1993).
Berdasarkan fungsinya, terdapat dua penjelasan: (1) intervensi dapat bersifat altruistik atau (2) menguntungkan salah satu pihak saja (Widdig et al. 2000). Tingkah
laku altruistik berkaitan dengan pengeluaran oleh pemberi/altruist (misalnya waktu, energi, resiko luka, dan pembalasan) dan keuntungan bagi penerima/resipien (misalnya akses terhadap sumber yang terbatas dan kurangnya perlukaan dalam perkelahian) (Widdig et al. 2000).
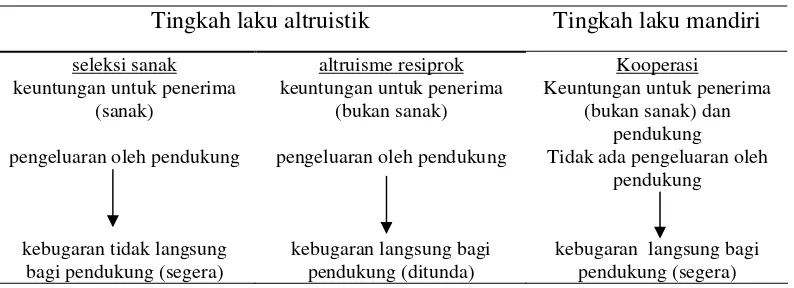
Pembentukan koalisi dapat dijelaskan melalui tiga teori, yaitu seleksi sanak, altruisme resiprok, dan kooperasi (Widdig et al. 2000). Teori seleksi sanak memprediksi bahwa individu-individu yang mendukung kerabat akan meningkatkan kebugaran secara tidak langsung karena mereka membagi gennya dengan resipien.
bergabung dengan jantan saudaranya yang lebih tua dan membentuk koalisi (Widdig et al. 2000).
Pada altruisme resiprok, pemberi berperan memberikan pengeluaran dan tanpa keuntungan segera, tetapi pemberi menerima keuntungan pada masa mendatang dari penerima. Jika keuntungan untuk penerima lebih besar daripada pengeluaran oleh pemberi, kedua pihak memperoleh kebugaran langsung untuk periode waktu yang lama
meskipun tidak berhubungan lagi (Widdig 2000). Sebagai contoh, Pasangan baboon
jantan yang tidak berkerabat memperoleh kesempatan untuk mengawini betina estrus.
Tabel 2 Tiga teori untuk menjelaskan pembentukan koalisi (Widdig 2000)
Tingkah laku altruistik Tingkah laku mandiri
seleksi sanak
Pada teori kooperasi, pendukung bukanlah pemberi karena menerima keuntungan langsung dari ikatan dengan individu yang tidak berkerabat (Widdig 2000). Chimpanzee
jantan mendapatkan keuntungan saat membantu individu yang tidak berkerabat melawan individu lain, sehingga dapat menaikkan peringkat individu tersebut (Widdig et al. 2000). Perbandingan tiga teori untuk menjelaskan pembentukan koalisi tersebut dapat dilihat pada Tabel 2.
Berdasarkan pada hierarki dominansi, aliansi dapat dibedakan menjadi tiga, yaitu (1) tipe konservatif, (2) tipe jembatan, dan (3) tipe revolusioner
menengah. Pada aliansi tipe revolusioner, individu-individu subordinan melawan individu dominan.
2.3. Menelisik
Menelisik didefinisikan sebagai tindakan mengambil kotoran atau lainnya dari rambut dan kulit dengan menggunakan jari atau gigi (Rasmussen 1993). Menelisik
merupakan bentuk komunikasi sentuhan yang secara intensif sudah dipelajari karena mempunyai peranan yang menonjol dalam kehidupan harian kebanyakan spesies primata (Collinge 1993). Di samping berfungsi untuk membersihkan kulit dan rambut dari kotoran dan parasit, menelisik juga berperan dalam interaksi sosial dalam berbagai konteks, misalnya dalam hal induk menenangkan bayinya, pasangan kawin untuk sinyal memulai kawin, persaudaraan, dan untuk rekonsiliasi. Menelisik oportunistik terhadap hewan dominan merupakan strategi yang sering digunakan oleh hewan subordinan sebagai sarana untuk membagi keuntungan aspek dominansi (Collinge 1993).
Menelisik mencakup manipulasi oral dan manual pada kulit dan/atau rambut
(Cooper dan Berstein 2000).Menelisik dapat dilakukan untuk diri sendiri (menelisik diri) dan dapat dilakukan untuk pasangan sosialnya (menelisik silang) (Chalmer 1980). Jika dilakukan untuk diri sendiri, menelisik berfungsi untuk membuang ektoparasit atau untuk membersihkan dan mempertahankan permukaan tubuh. Menelisik untuk fungsi ini sudah diamati pada monyet ekor singa (M. silenus), lemur ekor cincin (Lemur catta), monyet hitam Sulawesi (M. nigra), dan monyet bonnet (M. radiata) (Chalmer 1980). Menelisik sosial, di samping berfungsi seperti menelisik diri, juga untuk mempererat ikatan sosial. Menelisik sosial dapat berfungsi sebagai pembayaran oleh individu subordinan sebagai
Berdasarkan hipotesis ikatan sosial (Cooper dan Berstein 2000), betina sebagai anggota tetap dalam kelompok matrifokal saling menelisik satu sama lain dan dengan anak-anak lebih sering daripada antarjantan dan antara jantan dan anak-anak. Sebaliknya, jantan menelisik betina lebih sering dan lebih lama daripada betina menelisik jantan. Jantan dan betina lebih sering menelisik anak-anak daripada sebaliknya. Anak-anak lebih lama menelisik yang lebih tua daripada sebaliknya. Penelitian pada monyet Assam (M.
assamensis), menelisik lebih berfungsi dalam memantapkan dan memelihara ikatan sosial afiliatif daripada sebagai mekanisme spesifik untuk kawin dan fungsi resiprok (Cooper dan Berstein 2000).
2.4. Pendekatan
Pendekatan mencakup frekuensi pendekatan dan arah pendekatan (Chaffin et al 1995). Pendekatan yang dimaksud adalah pendekatan non-agonistik per jam observasi fokus. Pendekatan didefinisikan sebagai pergerakan satu individu untuk jarak yang tercapai tangan terhadap individu lain dari berbagai arah.
Hasil pendekatan dibuat skor sebagai positif/netral jika salah satu dari diad melakukan kontak tubuh positif (menelisik, berimpitian, atau kontak non-agonistik lainnya), atau duduk dalam jangkauan tangan selama 10 detik (pendekatan netral). Tanpa pendekatan jika penuju meninggalkan proksimitas dalam 10 detik dan tidak membuat kontak. Pendekatan negatif jika tertuju menjauh, ancaman gigi oleh salah satu partisipan, dan atau ancaman oleh tertuju. Jika penuju mengancam, perjumpaan diberi skor sebagai ancaman. Arah pendekatan adalah tendensi pendekatan oleh dominan vs. subordinan yang ditunjukkan dengan indeks atas/bawah (Chaffin et al. 1995).
Definisi Istilah
1. Aspek karakteristik kelompok
1) Ukuran kelompok: jumlah individu dalam suatu kelompok (Chalmer 1980).
3) Pertumbuhan kelompok: perubahan ukuran kelompok selama waktu tertentu karena faktor natalitas, mortalitas, dan migrasi (Alikodra 2002).
4) Daerah jelajah: luas area yang dijelajahi monyet (kelompok monyet) selama hidup kelompok tersebut (Collinge 1993).
5) Jelajah harian: jarak yang ditempuh monyet (kelompok monyet) dalam satu hari (Collinge 1993).
2. Aspek Karakteristik Dominansi
1) Dominansi: kemampuan untuk mengintimidasi individu lain dalam suatu konflik dan kemampuan untuk mendapatkan prioritas yang lebih untuk akses terhadap berbagai sumber, seperti pakan, ruang, dan kawin (Collinge 1993, Bramblett 1994). Individu yang memperoleh akses terhadap sumber lebih banyak dibandingkan dengan individu lain disebut dominan, sedangkan yang memperoleh akses lebih sedikit disebut subordinan (Collinge 1993).
2) Sistem sosial despotik: sistem sosial dengan keuntungan dalam memanfaatkan sumber secara kuat dimiliki oleh individu peringkat tinggi, dengan interaksi sosial
bersifat asimetris (Matsumura 1998, Hemelrijk 1999).
3) Sistem sosial egaliter: sistem sosial dengan keuntungan dalam memanfaatkan sumber tersebar merata pada semua peringkat dan interaksi sosialnya bersifat simetris (Matsumura 1998, Hemelrijk 1999).
4) Hierarki dominansi: keseluruhan susunan individu dominan dan subordinan dalam kelompok (Martin dan Bateson 1999).
3. Aspek tingkah laku sosial
1) Tingkah laku agonistik: tingkah laku yang berkaitan dengan agresi (Collinge
1993). Dalam tingkah laku agresif, individu dibedakan menjadi dua, yaitu penyerang dan korban. Agresi mencakup agresi ringan (ancaman dengan membuka mulut, ancaman dengan suara, serangan mendadak) dan agresi berat (mengusir, memukul, mencakar, menggigit) (perry 1996, Matsumura 1998).
menyertainya, seperti membuka mulut, merendahkan kepala, telinga diarahkan ke belakang, vokalisasi deheman. Terjangan kurang dari dua meter dimasukkan sebagai mengancam Menyerang mencakup mengusir, menampar atau menggigit, atau menggigit dengan ganas. Menggigit dengan ganas didefinisikan sebagai menggigit non-ritual dan intensitasnya tinggi, diberi skor jika tingkah laku agresif ini dilakukan dengan gerakan kepala yang berulang menghasilkan luka pada
lawan.
3) Rekonsiliasi: afiliasi yang dilakukan oleh penyerang dan korban segera setelah terjadinya konflik (Kutsukake dan Castles 2001).
4) Tendensi berekonsiliasi: tendensi pihak pertama untuk berasosiasi dengan pihak kedua, diukur sebagai persentase dari pasangan yang berasosiasi (Chaffin et al.
1995).
5) Aliansi/koalisi: kerjasama dalam agresi oleh dua individu atau lebih dalam menghadapi pihak ketiga (Perry 1996). Koalisi kadang-kadang terjadi pada dua individu yang mengusir atau menunjukkan muka mengancam dengan mulut
terbuka terhadap individu ketiga. Secara umum, partner koalisi berada dalam kontak fisik satu sama lain mengancam pihak ketiga. Mereka berdampingan, berangkulan, atau melakukan suatu punjian. Punjian didefinisikan sebagai satu atau lebih individu berdiri di atas kepala yang lain, menatap/mengancam individu lain. Anggukan didefinisikan sebagai seekor monyet memajukan kepalanya ke arah partner koalisi dan kemudian menatap atau mengancam lawan; anggukan merupakan tingkah laku umum yang digunakan untuk merekrut partner.
6) Pada aliansi tipe konservatif: dua individu dominan melawan individu subordinan
(http://life.bio.sunysb.edu/bio359/4_26_02. html).
7) Aliansi tipe jembatan: individu dominan bekerja sama dengan individu subordinan melawan individu peringkat menengah (http://life.bio.sunysb.edu /bio359/4_26_02.html).
9) Menelisik: tindakan mengambil kotoran atau lainnya dari rambut dan kulit dengan menggunakan jari atau gigi (Rasmussen 1993). Menelisik merupakan bentuk komunikasi sentuhan yang secara intensif sudah dipelajari karena mempunyai peranan yang menonjol dalam kehidupan harian kebanyakan spesies primata (Collinge 1993). Individu yang menelisik individu lain disebut penelisik, sedangkan individu yang ditelisik (resipien) disebut tertelisik (Chalmer 1980).
Penelisikan yang dilakukan sendiri disebut menelisik diri, sedangkan yang dilakukan untuk individu lain disebut menelisik silang (Chalmer 1980).
10)Pendekatan: pergerakan satu individu untuk jarak yang tercapai tangan terhadap individu lain dari berbagai arah (Martin dan Bateson 1999). Individu yang mendekati disebut penuju, sedangkan individu yang didekati disebut tertuju. Pendekatan disebut positif jika salah satu dari diad melakukan kontak tubuh positif (penelisikan, berimpitian, atau kontak non-agonistik lainnya). Pendekatan disebut netral jika diad duduk dalam jangkauan tangan selama 10 detik. Pendekatan disebut negatif jika tertuju menjauh, ancaman gigi oleh salah satu partisipan, dan
atau ancaman oleh tertuju. Tanpa pendekatan terjadi jika penuju meninggalkan proksimitas dalam 10 detik dan tidak membuat kontak (Martin dan Bateson 1999). 11)Arah pendekatan: tendensi untuk mendekati individu dominan versus subordinan,
ditunjukkan sebagai indeks atas/bawah (Chaffin et al. 1995). 4. Aspek reproduksi
1) Pemilihan pasangan kawin: pola tingkah laku pada satu jenis kelamin yang dapat meningkatkan probabilitas perkawinan fertil dengan individu tertentu pada jenis kelamin yang berbeda (Soltis et al. 1997).
MATERI DAN METODE PENELITIAN
Tempat dan Waktu
Penelitian dilaksanakan di Cagar Alam Tangkoko-Batuangus (Gambar 4), Kecamatan Bitung Utara, Kota Bitung, Sulawesi Utara, dari bulan Januari-Desember 2004.
Materi dan Alat Materi Penelitian
Materi penelitian adalah Monyet Hitam Sulawesi (Macaca nigra) Kelompok Rambo II (KRII) sebesar 51 ekor dan Kelompok Rambo I (KRI) sebesar 51 ekor di CA Tangkoko-Batuangus.
Alat Penelitian
Alat untuk penelitian meliputi: binokuler, pita, jam, stop watch, lembar
pengamatan, senter, penghitung (counter) tangan, kamera, alat ukur, kompas, dan alat perekam.
Metode Penelitian
Prinsip pengamatan interaksi adalah who does what to whom. Dengan demikian langkah berikutnya setelah habituasi, identifikasi setiap individu-target berdasarkan ciri-ciri khasnya dan pemberian nama berdasarkan ciri-ciri khas tersebut, sehingga mudah diingat. Identifikasi jantan dewasa KRII dilakukan pada awal bulan Januari, sedangkan jantan dewasa KRI dilakukan pada bulan Juli.
Karakteristik Kelompok 1. Ukuran dan Komposisi Kelompok
Ukuran kelompok diperoleh dengan menghitung seluruh anggota KRII dan KRI. Penghitungan dilakukan secara reguler setiap bulan, sehingga diperoleh juga gambaran pertumbuhan kelompok. Penghitungan dilakukan dengan menggunakan penghitung tangan pada saat kelompok monyet menyeberang jalan atau pada saat turun dari pohon tidur. Penghitungan dilakukan selama minimal 10 kali dan jumlah tertinggi ditetapkan sebagai ukuran kelompok yang bersangkutan.
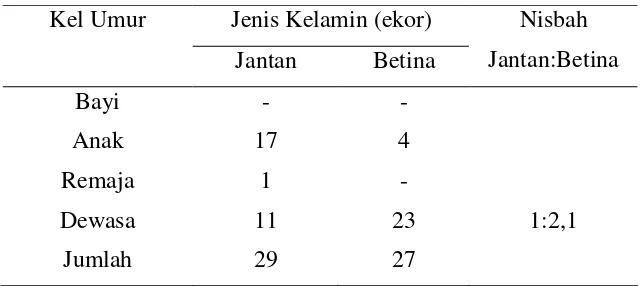
Komposisi kelompok diperoleh dengan identifikasi seluruh anggota kelompok (51 ekor untuk KRII dan 51 ekor untuk KRI) menurut umur dan jenis kelaminnya. Identifikasi dilakukan dengan mengamati ciri-ciri khas setiap individu secara seksama dengan menggunakan binokuler. Ciri-ciri yang digunakan dalam identifikasi meliputi ukuran tubuh (besar, kecil), warna rambut, bentuk bagian-bagian tubuh, kecacatan, bekas-bekas luka pada muka, tangan, kaki, telinga, dan bagian tubuh lainnya, bercak pada muka dan telinga, dan tingkah laku. Setelah identifikasi dilakukan, setiap bulan dilakukan sensus untuk seluruh anggota kelompok, sehingga dapat diketahui jika terjadi imigrasi atau emigrasi antarkelompok.
Pengamatan komposisi kelompok pada KRII dilakukan pada awal penelitian (bulan Januari) dan akhir penelitian (bulan Desember). Untuk KRI, pengamatan dilakukan pada bulan Juli dan Desember secara tidak kontinyu.
Pengelompokan umur didasarkan pada fase perkembangan individu yaitu bayi, anak, remaja, dan dewasa (Chalmers 1982). Ciri-ciri masing-masing fase didasarkan pada ciri-ciri yang dideskripsikan oleh Altman (1981) dan penelitian pendahuluan yang sudah dilakukan (Saroyo 2002a). Perkiraan umur didasarkan pada ciri-ciri fisik dan studi pustaka pada beberapa penelitian pada monyet hitam Sulawesi maupun monyet Sulawesi lainnya (Okamoto et al. 2000). Keempat kelompok umur tersebut dirinci sebagai berikut ini.
waktu lahir, diasuh oleh induknya, sampai dengan masa sapihan. Penentuan jenis kelamin dilakukan dengan mengamati organ genitalia luar. Bayi jantan mempunyai penis yang panjang bila dibandingkan dengan ukuran tubuhnya, sehingga sangat mudah diamati.
2) Kelompok anak yaitu fase yang dimulai setelah bayi sampai sebelum dewasa. Individu pada kelompok ini biasanya sudah disapih dan tidak lagi dibawa induknya, serta mempunyai fase reproduksi yang belum matang. Umumnya kelompok umur ini lebih banyak menghabiskan waktunya untuk bermain. Penentuan jenis kelamin dilakukan dengan mengamati organ genitalia luarnya.
3) Kelompok remaja mempunyai ukuran tubuh sedikit lebih besar dibandingkan dengan ukuran tubuh anak dan sedikit lebih kecil kecil dibandingkan dengan pada yang dewasa. Terdapat dimorfisme seksual pada ukuran tubuhnya, yaitu jantan lebih besar daripada betina. Pada jantan remaja, warna rambut pada bahu dan tangan mulai menunjukkan warna seperti dewasa yaitu hitam terang. Ukuran tubuhnya kira-kira di antara jantan anak besar (atau betina dewasa) dengan jantan dewasa. Karakteristik seks sekundernya belum berkembang sempurna. Skrotum mulai membesar, dan biasanya memisahkan diri dari kelompok anak, tetapi belum bergabung dengan kelompok dewasa. Pada betina remaja, ukuran tubuh mendekati ukuran dewasa tetapi puting susu masih pendek seperti pada jantan dan berwarna putih. Tanda lain adalah mulainya tanda-tanda estrus tetapi siklusnya belum teratur. Umur jantan remaja berkisar 5-7 tahun, sedangkan betina remaja berkisar 4-5 tahun.
warna puting susunya. Pada betina dewasa, puting susu panjang dan sering menggantung, serta berwarna pink.
2. Pertumbuhan Kelompok
Pertumbuhan kelompok diamati pada KRII dan KRI. Data diambil dengan menghitung dan melakukan sensus terhadap seluruh anggota kelompok setiap bulan atau pada saat-saat tertentu, sehingga dapat ditentukan natalitas, mortalitas, dan migrasi anggota kelompok tersebut. Sensus dilakukan pada saat kelompok menyeberang jalan atau pada saat turun dari pohon tidur minimal 10 kali dan jumlah terbesar ditetapkan sebagai ukuran kelompok. Data yang dikumpulkan meliputi jumlah individu pada setiap kelompok umur dan jenis kelamin. KRII diamati dari bulan Januari-Desember, sedangkan KRI dari bulan Juli-Desember. Berdasarkan penghitungan total individu setiap bulan untuk KRII dan KRI dapat ditentukan pertumbuhan kelompok tersebut. Berdasarkan pengamatan komposisi kelompok setiap bulan dapat ditentukan keberadaan setiap individu anggota kelompok. Pertumbuhan kelompok dapat ditentukan berdasarkan kelahiran, kematian, emigrasi, serta imigrasi.
3. Daerah Jelajah dan Jelajah Harian
Daerah jelajah dan jelajah harian ditentukan melalui pemetaan daerah jelajah, habituasi kelompok, dan pengukuran luas daerah jelajah dan panjang jelajah harian. Pemetaan daerah jelajah dilakukan dengan pembuatan jalur-jalur dengan arah 220o atau tegak lurus dengan jalan dari Kelurahan Batuputih menuju Pos II sampai Pantai Batu sepanjang 2.400 m. Jarak jalur satu dengan jalur berikutnya adalah 100 m. Setiap jalur dibuat sepanjang 1.300 m dan setiap selang 25 m pada jalur tersebut diberi tanda nama jalur dan jarak titik tersebut dari jalan dengan menggunakan pita (misalnya 0, 25, A-50, A-75, dan seterusnya). Penamaan jalur sebagai berikut: tepat di belakang Pos II diberi nama Jalur C; dari belakang Pos II ke arah Kelurahan Batuputih berturut-turut adalah Jalur B, A, a, b, c, d, e, f, g, dan h; sedangkan dari belakang Pos II ke arah Pantai Batu berturut-turut adalah Jalur D, E, F, G, H, I, J, K, L, M, N, O, P, dan Q.
kehadiran peneliti. Setelah kelompok terhabituasi, tahap berikutnya mengikuti pergerakan monyet selama 21 hari. Pergerakan kelompok dipetakan pada kertas milimeter sesuai dengan titik-titik atau jalur-jalur yang dilewati. Penentuan luas daerah jelajah dan jarak jelajah harian dilakukan dengan estimasi jarak pada peta dan estimasi berdasarkan pengukuran di lapangan. Luas jelajah harian ditetapkan berdasarkan jelajah terluar yang dilalui kelompok, sedangkan jarak jelajah harian didasarkan pada pergerakan anggota yang berada di tengah-tengah kelompok.
Jelajah harian yang dipetakan pada kertas milimeter setiap hari digabungkan, dan jelajah terluarnya merupakan daerah jelajah selama periode pengamatan. Pengukuran jelajah harian KRII dilaksanakan pada bulan Februari dan bulan Maret; sedangkan untuk KRI dilaksanakan pada bulan Juli, Agustus, dan Oktober. Jarak jelajah harian diestimasi di kertas milimeter dan rata-rata selama periode pengamatan merupakan rata-rata jarak jelajah harian. Luas daerah jelajah diestimasi dari gambar yang dihasilkan berdasarkan gabungan dari gambar-gambar jelajah harian dan hasil pengamatan pada waktu lainnya yang dilakukan pada saat mengikuti kelompok monyet.
Aspek Dominansi
Dominansi ditentukan melalui pengamatan interaksi agonistik (Chalmes 1982) yang mencakup sikap dan sinyal agonistik. Jumlah individu jantan yang diamati sebanyak enam ekor pada KRII dan enam ekor pada KRI. Pengamatan tingkah laku agonistik dilakukan dengan focal sampling dengan metode perekaman secara kontinyu (Matsumura 1998, Martin dan Bateson 1999). Pengamatan focal untuk setiap jantan dilakukan selama lima hari. Setelah diketahui bahwa pesaing superior selalu unggul, pengamatan focal
setiap jantan dilakukan selama satu hari. Individu yang terlibat dalam interaksi agonistik dapat dibedakan menjadi individu penyerang dan individu korban. Penyerang menunjukkan tingkah laku agresif, misalnya mengejar, menggigit, mengancam, mencakar, dan sebagainya. Korban menunjukkan tingkah laku tunduk, misalnya ekspresi takut, lari, atau diam (Matsumura 1998).
dilakukan secara kontinyu, sedangkan untuk KRI tidak secara kontinyu. Pengamatan dinamika dominansi untuk KRI dilakukan pada 3-8 Agustus, Akhir September, 7-8 Oktober, dan akhir November, dan akhir Desember.
Metode pengambilan data untuk interaksi agonistik betina sama seperti pada metode untuk dominansi jantan. Pengamatan focal untuk setiap betina dilakukan selama dua hari pada 14 betina dewasa KRII. Untuk mengamati dinamika dalam hierarkinya, pengamatan dilaksanakan dalam dua periode, yaitu bulan Juni (30 hari) dan Agustus (31 hari). Total interaksi agonistik antarbetina pada bulan Juni sebanyak 91 kali dan pada bulan Agustus sebanyak 105 kali. Data agonistik di antara betina diambil jika kedua betina tersebut berada pada jarak lebih dari satu meter dengan jantan dan betina lain untuk menghindari intervensi atau pengaruh individu lain, sehingga hasil interaksi agonistik benar-benar mencerminkan peringkat dominansinya.
Proses migrasi jantan diamati pada enam jantan dewasa dan tiga jantan remaja KRII serta enam jantan dewasa dan seekor jantan remaja KRI dengan menggunakan ad libitum sampling atau focal sampling terhadap seluruh jantan dewasa dan remaja kedua kelompok tersebut. Jika terdapat jantan yang menunjukkan tanda-tanda akan melakukan migrasi, maka jantan tersebut diikuti secara focal sampling dan pengamatan tingkah laku dilakukan secara ad libitum sampling.
Sebelum dilakukan analisis terhadap linearitas dominansi, terlebih dahulu dilakukan penghitungan angka agresi (Martin dan Bateson 1999). Persentase yang diperoleh merupakan persentase aktivitas agresif dari total aktivitas hariannya. Dari hasil pengamatan interaksi agonistik, dihitung frekuensi setiap individu sebagai pemenang dan sebagai korban dalam suatu tabel untuk menentukan peringkat setiap individu. Linearitas dominansi ditentukan dengan menggunakan Indeks linearitas dari Landau (h) dengan rumus:
12 n
h =