1.
KEANEKARAGAMAN
HYPHOMYCETES
PADA PISANG
(
Musa x paradisiaca
)
DESI MARIA PANJAITAN
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
Dengan ini saya menyatakan bahwa tesis berjudul Keanekaragaman Hyphomycetes pada Pisang (Musa x paradisiaca) adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Dengan ini saya melimpahkan hak cipta karya tulis saya kepada Institut Pertanian Bogor.
Bogor, November 2016
(Musa x paradisiaca). Dibimbing oleh NAMPIAH SUKARNO dan MIEN AHMAD RIFAI.
Hyphomycetes adalah cendawan anamorf yang bentuknya beranekaragam dan terdistribusi luas di berbagai habitat. Pemanasan global menyebabkan beberapa spesies model yang digunakan dalam pembelajaran mikologi saat ini sulit ditemukan di lingkungan sekitar. Studi keanekaragaman cendawan Hyphomycetes dan ontogeni konidia masih minim dilakukan di Indonesia padahal eksplorasi harus tetap dilakukan karena diperkirakan banyak cendawan yang belum sempat diketahui. Penelitian ini bertujuan menyediakan informasi mikoflora terkini khususnya Hyphomycetes di Indonesia dengan cara mengumpulkan, mengisolasi dan mengidentifikasi keanekaragaman spesies Hyphomycetes serta menguraikan ontogeni konidia dan perkembangan konidianya. Pisang, salah satu tanaman budidaya penting dan mudah ditemukan di seluruh Indonesia, sengaja dipilih untuk memfasilitasi penyediaan salah satu sumber bahan ajar keanekaragaman cendawan Indonesia guna kepentingan perkuliahan mikologi.
Eksplorasi dilakukan menggunakan metode purposive sampling di beberapa kebun yang ditanami pisang di Bogor dan sekitarnya. Daun, tangkai daun, dan pelepah kering tanaman pisang yang ditumbuhi cendawan Hyphomycetes dikoleksi dan dilakukan pembuatan herbariumnya. Spesimen diamati di laboratorium menggunakan mikroskop dan diidentifikasi secara morfologi. Konidia diisolasi menggunakan teknik isolasi spora tunggal. Analisis molekuler dilakukan pada beberapa koleksi isolat berdasarkan daerah ITS ribosomal DNAnya.
(Musa x paradisiaca). Supervised by NAMPIAH SUKARNO and MIEN AHMAD RIFAI.
Hyphomycetes is a group of anamorphic fungi which diverse forms and wide distribution in various environment. Global warming has affected several fungal model species used in mycology class make them now difficult to find. Hyphomycetes diversity and its conidial ontogeny are not well studied yet in Indonesia. Therefore this research has been undertaken to explore, collect, isolate, identify, and describe Hyphomycetes species and to elucide their conidial ontogeny and conidial development. Banana, one of the major and widely distributed crop plant in Indonesia, has been intentionally chosen as one source to facilitate mycological education.
Exploration was carried out using purposive sampling method in some banana plantation around Bogor. Collected samples consisted of decaying and dry leaf, petiole, and pseudostem of banana. Specimens were observed under microscope and fungal identification was done based on morphological characters. Conidia were isolated using single spore isolation technique. Molecular analysis was done on some collections based on DNA ribosomal ITS region.
Based on the observation of 127 collected specimens, 31 species of Hyphomycetes were identified.Most of the species are new records for mycoflora of Indonesia. In addition, four species belonging to three genera are proposed as new to science namely Dictyosporium bogoriense, Stachybotrys atrolaevis, Stachybotrys musicola, and Oncopodium javanicum. The other species are Bipolaris sp., Curvularia lunata, Dendryphiella vinosa, Dendryphion comosum, Dictyosporium heptasporum, D. hydei, D. zeylanicum, Exochalara sp., Lacellinopsis sacchari, Periconia sp., Phaeoisaria clematidis, Phaeostalagmus cyclosporus, Pithomyces sp. 1, Pithomyces sp. 2, Pyriculariopsis parasitica, Ramichloridium musae, Spegazzinia deightonii, S. tessarthra, Stachybotrys echinata, S. levispora, S. subsimplex, Stachylidium bicolor, Tetraploa aristata, Torula herbarum, and Zygosporium oscheoides. The absence and inadequate modern references made it impossible to identify all species and two others could not be identified due to the lack of morphological character so that they were refered to Hyphomycetes sp. 1 and Hyphomycetes sp. 2. Six species were succesfully cultured namely Bipolaris sp., Hyphomycetes sp. 2, S. atrolaevis, S. echinata, P. cyclosporus, and T. herbarum. Molecular analysis of two isolates of Stachybotrys spp. showed the match identity only to S. echinata.
The species found in this study vary in morphological characters mainly of conidiophore, conidiogenous cell and conidia. Seven types of conidial ontogeny were found and described. They are terminally integrated, acropetally blastic, fragmentationlly basipetal, sympodially denticulate, phialidic, tretic, and basauxic cell. Five types of conidia were also found namely amerospore, cheroidspore, dictyospore, phragmospore and staurospore.
©
Hak Cipta Milik IPB, Tahun 2016
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB.
DESI MARIA PANJAITAN
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains
pada
Program Studi Mikrobiologi
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
Pujian dan syukur penulis ucapkan untuk semua berkat dan kebaikan yang diberikan oleh Tuhan Yang Maha Kuasa sehingga penulis dapat menyelesaikan pendidikan pascasarjana, penelitian, dan penulisan tesis yang berjudul Keanekaragaman Hyphomycetes pada Pisang (Musa x paradisiaca). Penulisan tesis ini adalah salah satu syarat yang harus disusun sebagai upaya mendapatkan gelar Magister Sains di Sekolah Pascasarjana Institut Pertanian Bogor (IPB). Pelaksanaan penelitian ini didanai oleh DIKTI melalui program Beasiswa Pendidikan Pascasarjana Dalam Negri (BPPDN) Calon Dosen tahun 2013.
Penulis menyampaikan ucapan terima kasih yang tidak terhingga kepada ketua komisi pembimbing, Ibu Dr. Ir. Nampiah Sukarno yang dengan penuh ketekunan memberikan ajaran, arahan, dan dukungan selama masa perkuliahan dan juga selama penyelesaian tugas akhir di Program Studi Mikrobiologi IPB. Penulis juga mengucapkan terima kasih kepada anggota komisi pembimbing, Bapak Prof. Mien A. Rifai, M.Sc PhD, yang dengan penuh kesabaran dan kerelaan hati memberikan waktu, pikiran dan tenaga yang luar biasa kepada penulis dalam mempelajari Hyphomycetes beserta perihal taksonominya. Beliau pula lah yang sebenarnya pertama kali memberi buah pikiran untuk melaksanakan topik penelitian ini. Penulis juga menghaturkan terima kasih kepada penguji luar komisi, Ibu Dr. Atik Retnowati, S.P. M.Sc, yang berkenan menguji dan memberi kritik serta saran yang sangat membangun demi meningkatkan kualitas tesis saya.
Penulis ucapkan pula terima kasih pada semua pengajar di Program Studi Mikrobiologi terutama Bagian Mikologi yang namanya tidak dapat saya sebutkan satu per satu. Selain itu terima kasih kepada seluruh teman-teman di Laboratorium Penelitian Mikologi dan IPB Culture Collection: Astri Ariyani S.Si, Dewi S.Si, Indriati Ramadhani S.Si, Ivan Permana M.Si, Neng Karisma, Oktan D. Nurhayat S.Si, Sepriyadi S.Si, Sophia Mujahidah S.Si, Nurul S.Si dan juga para sahabat saya Fandri S. Fastanti S.Si dan Frans G. Naibaho M.Si yang telah memberikan uluran tangan, saran, dan diskusi berharga selama penelitian. Ungkapan terima kasih penulis sampaikan juga kepada orang tua, Bapak Guntur Panjaitan, Ibu Dameria Siahaan dan Beybel Tobing, serta abang dan adik-adikku yang senantiasa mendukung dan mendoakan.
Ketidaksempurnaan dan kekurangan dalam tesis ini dan pengetahuan yang penulis miliki adalah hal yang pasti. Oleh sebab itu penulis terbuka terhadap kritik dan saran dari pihak manapun. Penulis berharap karya ilmiah ini bermanfaat bagi semua pembacanya terutama berkontribusi dalam dunia taksonomi cendawan Indonesia.
Bogor, November 2016
DAFTAR GAMBAR DAFTAR LAMPIRAN
1 PENDAHULUAN 1
Latar Belakang 1
Tujuan Penelitian 2
2 METODE 3
Waktu dan Tempat Penelitian 3
Bahan 3
Alat 3
Pengambilan sampel dan pembuatan herbarium 3
Pengamatan mikroskopis cendawan, pembuatan preparat, dan ilustrasi 3
Isolasi spora tunggal 4
Identifikasi morfologi 4
Identifikasi molekuler 5
Analisis data 6
3 HASIL DAN PEMBAHASAN 7
Variasi morfologi Hyphomycetes pada Musa x paradisiaca di Bogor 7 Keanekaragaman Hyphomycetes pada Musa x paradisiaca di Bogor 11
Kunci Identifikasi Genus 13
Taksonomi 15
4 SIMPULAN DAN SARAN 52
Simpulan 52
Saran 52
DAFTAR PUSTAKA 53
LAMPIRAN 56
DAFTAR GAMBAR
dengan Hyphomycetes pada spesies-spesies Musa lainnya 12
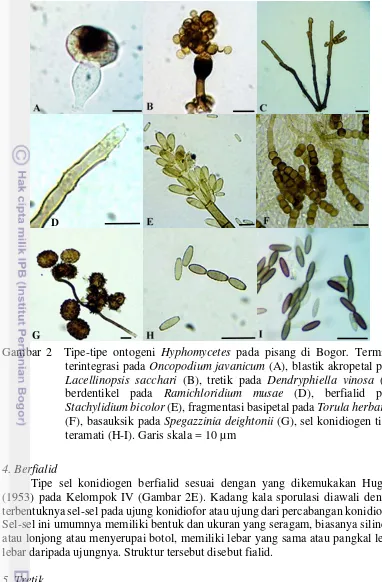
1 Variasi konidiofor Hyphomycetes pada pisang di Bogor. Makronema (A-C), konidiofor tunggal pada Stachybotrys subsimplex (A), konidiofor bercabang pada Stachylidium bicolor (B), konidiofor sinemata pada Phaeoisariaclematidis (C), konidiofor semimakronema pada Torula herbarum (D), konidiofor mikronema atau tereduksi pada Dictyosporium zeylanicum (E). Tanda panah menunjukkan konidiofor 7 2 Tipe-tipe ontogeni Hyphomycetes pada pisang di Bogor. Terminal
terintegrasi pada Oncopodium javanicum (A), blastik akropetal pada Lacellinopsis sacchari (B), tretik pada Dendryphiella vinosa (C), berdentikel pada Ramichloridium musae (D), berfialid pada Stachylidium bicolor (E), fragmentasi basipetal pada Torulaherbarum (F), basauksik pada Spegazzinia deightonii (G), sel konidiogen tidak
teramati (H-I). Garis skala = 20 µm 9
3 Lima Tipe konidia yang ditemukan pada Musa x paradisiaca. Amerospora yaitu konidia tidak bersekat (A). Fragmospora yaitu konidia bersekat lebih dari satu atau banyak (B). Keiroidspora yaitu konidia bersekat banyak menggepeng pada satu sisi dan seperti kumpulan jemari tangan (C). Diktiospora, konidia bersekat banyak dan tidak menggepeng (D). Staurospora, konidia seperti bintang, memiliki lebih dari satu sumbu karena adanya perpanjangan bagian konidia ke
beberapa arah (E) 10
4 Dictyosporium bogoriense pada substrat alami. Garis skala = 10 µm 16 5 Dictyosporium heptasporum pada substrat alami. Konidia silindris
tersusun oleh 7 baris lengan tersusun rapat. Garis skala = 10 µm 16 6 Dictyosporium hydei pada substrat alami. Kumpulan konidia (A).
Konidia memiliki tiga buah embelan membulat di bagian atas pangkal. Garis skala = 10 µm
17 7 Dictyosporium zeylanicum pada substrat alami. Konidia menggepeng
pada satu sisi dan tersusun oleh lima baris lengan. Garis skala = 10 µm 18 8 Oncopodium javanicum pada substrat alami. Konidiofor melebar
seperti kandung kemih (B). Garis skala = 10 µm 19 9 Pithomyces sp. 1 pada substrat alami. Garis skala = 10 µm 20 10 Pithomyces sp. 2 pada substrat alami. Garis skala = 10 µm 21 11 Tetraploa aristata pada substrat alami. Garis skala = 10 µm 21 12 Lacellinopsis sacchari pada substrat alami. Garis skala = 10 µm 23 13 Periconia sp pada substrat alami. Garis skala = 10 µm 24 14 Zygosporium oscheoides pada substrat alami. Garis skala = 10 µm 25 15 Bipolaris sp pada substrat alami (A-B, I). Konidia membentuk tabung
DAFTAR LAMPIRAN
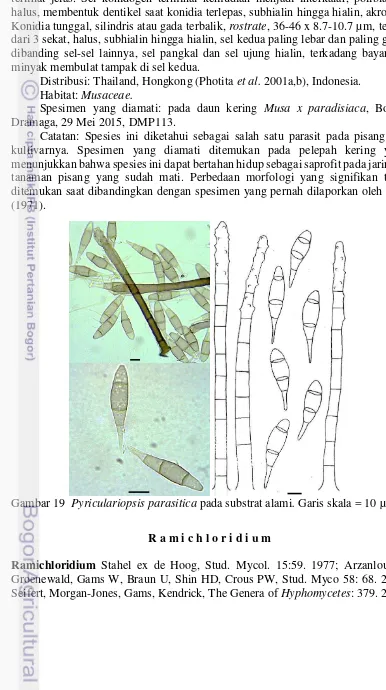
16 Curvularialunata pada substrat alami. Garis skala = 10 µm 28 17 Dendryphiella vinosa pada substrat alami. Garis skala = 10 µm 28 18 Dendryphion comosum pada substrat alami. Garis skala = 10 µm 30 19 Pyriculariopsis parasitica pada substrat alami. Garis skala = 10 µm 31 20 Ramichloridium musae pada substrat alami. Garis skala (A-D) = 10
µm, (E) = 20 µm 32
21 Phaeoisariaclematidis pada substrat alami. Garis skala = 10 µm 33 22 Exochalara sp. pada substrat alami. Garis skala= 10 µm 35 23 Stachybotrys atrolaevis pada substrat alami (A-B). Koloni
menghasilkan pigmentasi kuning kecokelatan pada media PDA inkubasi 7 hari pada suhu ruang tampak atas (C) dan tampak
sebaliknya (D). Garis skala = 10 µm 37
24 Stachybotrys echinata pada substrat alami (A-B, E). Koloni pada media PDA inkubasi 7 hari pada suhu ruang tampak atas (C) dan
tampak sebaliknya (D). Garis Skala = 10 µm 38
25 Stachybotrys levispora pada substrat alami. Garis skala = 10 µm 39 26 Stachybotrys musicola pada substrat alami. Garis skala = 10 µm 40 27 Stachybotrys subsimplex pada substrat alami. Garis skala= 10 µm 41 28 Pohon filogeni DMP41 dan DMP43 dan Stachybotrys spp. dengan
Cordyceps heteropoda IFO 33060 sebagai outgroup, menggunakan model Tamura 3 Parameter Neighbor Joining (NJ) dengan 1000
bootstrap 42
29 Phaeostalagmus cyclosporus pada substrat alami dengan pewarnaan laktofenol biru (A-C). Koloni pada media PDA inkubasi 7 hari pada suhu ruang tampak permukaan (D) dan tampak sebaliknya (E). Garis
skala = 10 µm 43
30 Stachylidium bicolor pada substrat alami. Garis skala = 10 µm 45 31 Torula herbarum pada substrat alami (A, E). T. herbarum media
Potato Dextrose Agar (PDA) (B-D). Koloni pada media PDA inkubasi 7 hari pada suhu ruang tampak permukaan (C), dan tampak ketika
dibalik (D). Garis skala = 10 µm 46
32 Spegazzinia deightonii pada substrat alami. Garis skala = 10 µm 48 33 Spegazzinia tessarthra pada substrat alami. Garis skala = 10 µm 49 34 Hyphomycetes sp. 1 pada substrat alami. Garis skala = 10 µm 50 35 Hyphomycetes sp. 2. Konidia pada substrat alami (A, E). Konidia pada
media PDA inkubasi 7 hari pada suhu ruang (B). Kultur koloni pada media PDA inkubasi 7 hari pada suhu ruang tampak atas (C), dan
tampak sebaliknya (D). Garis skala = 10 µm 50
1 Hasil elektroforesis DNA hasil amplifiksasi isolat DMP41 (1) dan DMP43 (2) menggunakan primer ITS1 dan ITS4 menunjukkan dua
1
PENDAHULUAN
Latar Belakang
Hyphomycetes merupakan kelompok cendawan bermiselium fertil dan menghasilkan spora aseksual yang disebut konidium. Miselium Hyphomycetes umumnya berdiferensiasi membentuk struktur mirip tangkai yang disebut konidiofor yang membawa penghasil konidium yang dinamakan sel konidiogen. Ragam bentuk dan proses terbentuknya sel konidiogen menjadi salah satu karakter kunci pengelompokan cendawan Hyphomycetes yang sebagian bermelanin (Ellis 1971). Beberapa penelitian telah menemukan berbagai kerugian dan manfaat yang diakibatkan oleh Hyphomycetes. Galur tertentu dari Stachybotrys chartarum yang ditemukan pada bangunan berdinding lembab menghasilkan spora toksigenik bagi manusia (Center for Disease Control 2002). Empat senyawa bioaktif yang diisolasi dari Tricladium castaneicola dapat digunakan untuk menghambat cendawan patogen tumbuhan seperti Phytopthora sp. (Han et al. 2015).
Hyphomycetes sangat beragam dan terdistribusi luas di berbagai habitat misalnya di laut, air tawar, kotoran hewan, dan berbagai habitat terestrial terutama di sisa tumbuhan (Chatmala et al. 2004; Lee et al. 2004; Jeamjit et al. 2006; Raja et al. 2007; Sudheep & Sridhar 2012). Organ tumbuhan yang umumnya banyak dikolonisasi ialah yang kaya selulosa seperti pada batang, pelepah, tangkai, dan daun. Hyphomycetes pada tumbuhan dapat hidup sebagai parasit, endofit, saprofit, atau bahkan lebih dari satu cara hidup bergantung pada keadaan lingkungan hidupnya. Beberapa penelitian menemukan adanya pergantian cara hidup cendawan endofit dan patogen bertahan menjadi saprofit saat inang mati (Zhou & Hyde 2001; Photita et al. 2004).
Pisang adalah tanaman budi daya yang tersebar luas dan merupakan salah satu inang bagi cendawan anamorf termasuk Hyphomycetes. Beberapa studi menemukan bahwa cendawan Hyphomycetes adalah kelompok yang paling beranekaragam dibandingkan dengan cendawan lainnya. Pendataan yang dilakukan oleh Photita et al. (2002) menunjukkan sebanyak 58% dari 204 spesies cendawan yang dideskripsikan dari Musaceae terdiri dari Hyphomycetes. Studi mengenai cendawan saprofit pada Musa acuminata L. di Hong Kong dan Thailand juga menunjukkan bahwa Hyphomycetes adalah kelompok yang paling beranekaragam dibandingkan dengan kelompok cendawan lainnya (Photita et al. 2003).
Ancaman nyata akibat aktivitas antropogenik dan pemanasan global yang ditandai dengan peningkatan suhu dan kekeringan menyebabkan perubahan mikoflora baik kelompok parasit hingga saprofit. Perubahan ini dapat diamati di berbagai daerah termasuk di Bogor. Beberapa spesies model yang umumnya digunakan dalam pembelajaran mikologi perlahan mulai sulit ditemukan di sekitar kita (Rifai 2012b). Fenomena ini menuntut dilakukannya eksplorasi guna menemukan habitat atau inang lain yang relatif mudah ditemukan di lingkungan sekitar kita.
keanekaragaman Hyphomycetes pada Musa x paradisiaca di sekitar Bogor serta mendeskripsikan setiap spesies yang ditemukan dan menguraikan ontogeni konidia dan perkembangan konidianya. Inang pisang yang mudah ditemukan di seluruh Indonesia sengaja dipilih untuk memfasilitasi penyediaan salah satu sumber bahan ajar keanekaragaman cendawan Indonesia guna kepentingan perkuliahan.
Tujuan Penelitian
2 METODE
Waktu dan Tempat Penelitian
Penelitian dilakukan di bulan Juli 2014 hingga Desember 2015. Pengambilan sampel dilakukan pada beberapa kebun yang ditanami pisang yang ada di kota Bogor dan daerah sekitarnya. Pengamatan dilakukan di lapangan dan di Laboratorium Mikologi di Fakultas Matematika dan Ilmu Pengetahuan Alam, Institut Pertanian Bogor.
Bahan
Bahan-bahan yang digunakan dalam penelitian di antaranya adalah koleksi spesimen berupa daun, tangkai dan pelepah kering Musa x paradisiaca, amplop herbarium, larutan shears, laktofenol biru, akuades, cat kuku transparan, kertas kalkir, media PDA (Potato Dextrose Agar), media WA (Water Agar), membran selofan, akuades, PCR Mix dengan primer ITS1 dan ITS4 (White et al. 1990). DNA template, etanol absolut, etanol 70%, dan bahan-bahan untuk melakukan elektroforesis diantaranya seperti Ethidium bromide, gel agarosa 1%, larutan penyangga TAE.
Alat
Alat-alat yang digunakan dalam penelitian ini adalah gunting tanaman, kantong sampel, lensa tangan akromatik, mikroskop stereo, mikroskop cahaya, gelas objek, gelas penutup, jarum sampel, pena gambar, cawan Petri, kamera lusida, Optilab viewer, autoklaf, inkubator, LAFC (Laminar Air Flow Cabinet), sentrifugator, mesin PCR, shaker water bath, alat elektroforesis dan alat dokumentasi gel.
Prosedur Penelitian
Pengambilan sampel dan pembuatan herbarium
Pengambilan sampel cendawan dilakukan secara purposive sampling dengan menjelajah kebun pisang. Pohon pisang yang dijumpai diamati batang, pelepah dan daunnya yang sudah mengering atau mati menggunakan lensa tangan akromatik. Organ teramati yang dikolonisasi oleh cendawan Hyphomycetes kemudian dipotong lalu dimasukkan ke dalam amplop koleksi berukuran 10 x 10 cm dan diberi catatan koleksi seperti nomor, waktu, tempat, nama kolektor dan keterangan organ pisang yang diambil. Sampel kemudian dibawa ke laboratorium untuk diamati lebih lanjut. Spesimen herbarium yang telah diidentifikasi disimpan di Herbarium Bogoriense (BO).
mengambil sedikit bagian koloni pada spesimen menggunakan jarum sampel kemudian diletakkan pada permukaan gelas objek yang sebelumnya telah ditetesi larutan shears atau laktofenol biru. Selanjutnya, tetesan tersebut ditutup dengan cover glass dan dilakukan pemanasan menggunakan bunsen pijar untuk menghilangkan gelembung udara. Pinggiran dari cover glass direkatkan menggunakan cat kuku transparan kemudian dibiarkan hingga mengering. Preparat kemudian diamati menggunakan mikroskop binokuler. Struktur mikroskopis yang diamati mencakup miselium, konidiofor, sel konidiogen dan konidia dengan menggunakan mikroskop cahaya Olympus BX53 (Olympus, Japan) dengan perbesaran 600 dan 1000 kali menggunakan minyak imersi. Setidaknya dilakukan pengukuran 30 konidia dewasa dari masing-masing spesimen untuk mendapatkan kisaran ukuran spora. Dokumentasi dilakukan menggunakan optilab viewer yang dimodifikasi pada lensa okuler mikroskop.
Ilustrasi preparat juga dilakukan menggunakan bantuan kamera lusida yang dimodifikasi pada mikroskop cahaya. Gambar selanjutnya dipindahkan ke kertas kalkir menggunakan pena gambar dengan berbagai ukuran ketebalan pena. Gambar kemudian dipindai menjadi data digital menggunakan scanner.
Isolasi spora tunggal
Spesimen yang telah diamati kemudian diisolasi untuk mendapatkan kulturnya. Isolasi menggunakan metode suspensi spora tunggal dilakukan dan merujuk Choi et al. 1999 dengan sedikit modifikasi. Spesimen diamati menggunakan mikroskop stereo kemudian permukaan koloni diusap menggunakan jarum ose steril lalu disuspensikan dengan dua tetes akuades steril pada gelas objek. Selanjutnya satu tetes dari suspensi spora tersebut diencerkan kembali dengan dua tetes akuades steril. Suspensi spora yang telah diencerkan kemudian digoreskan pada permukaan Water Agar (WA) yang telah ditambahkan antibiotik kloramfenikol 0.5 g/L. Inkubasi dilakukan selama 24 jam pada suhu ruang (27 ºC). Pengamatan dilakukan 1x12 jam menggunakan mikroskop binokuler pada perbesaran 400 kali. Konidia tunggal yang teramati telah membentuk tabung germinasi dipindahkan ke media PDA kemudian diinkubasi selama 7 hari pada suhu ruang. Pertumbuhan isolat diamati dan diukur diameter koloni setiap hari. Kultur kerja disimpan pada agar miring PDA masing-masing sebanyak 4 ulangan.
Isolat murni yang telah didapatkan diamati dan dilakukan pembuatan preparat berumur 7 hari yang selanjutnya akan dibandingkan dengan preparat yang berasal dari spesimen herbarium. Isolat cendawan yang telah teridentifikasi disimpan di Institut Pertanian Bogor Culture Collection (IPBCC).
Identifikasi morfologi
Glosarium Biologi (Rifai & Puryadi 2008). Kunci identifikasi disusun bagi genus-genus Hyphomycetes yang ditemukan.
Identifikasi molekuler
Isolasi DNA
Isolasi DNA dilakukan terhadap isolat Stachybotrys spp. Koloni isolat ditumbuhkan pada media PDA yang sebelumnya permukaannya ditutupi dengan membran selofan kemudian diinkubasi pada suhu ruang selama 7 hari. Miselium yang tumbuh di atas membran tersebut selanjutnya dikerik secukupnya dan digunakan untuk ekstraksi DNA. Metode ekstraksi DNA merujuk pada Raeder & Broda (1985) dengan modifikasi. Miselium dimasukkan ke dalam tabung ependorf yang berisi 100 µL SDS kemudian dihancurkan menggunakan pastel steril sampai homogen. Sebanyak 500 µL SDS ditambahkan kembali ke dalam tabung ependorf kemudian diinkubasi pada waterbath selama 30 menit pada kecepatan 300 round per menit (rpm) suhu 65 ºC dan dibolak-balik setiap interval 10 menit. Selanjutnya dikejutkan pada es selama 5 menit. Sebanyak 600 µL CI ditambahkan kemudian dihomogenkan. Sampel kemudian disentrifuse pada selama 10 menit pada kecepatan 10.000 rpm pada suhu 4 ºC selanjutnya 600 µL supernatan dipindahkan ke dalam tabung ependorf steril lainnya. Sebanyak 600 µL PCI (25:24:1) ditambahkan kemudian dihomogenkan. Sampel kemudian disentrifuse selama 5 menit pada kecepatan 10.000 rpm pada suhu 4 ºC. Sebanyak 400-500 µL supernatan yang bening dipindahkan ke dalam tabung Ependorf steril yang baru. Sampel kemudian ditambahkan dengan 40 hingga 50 µL 2 M NaOC pH 5.2 dan 600 µL Etanol 100%. Sampel kemudian diinkubasi pada freezer selama 24 jam. Sampel kemudian disentrifuse selama 30 menit pada kecepatan 10.000 rpm dan suhu 4 ºC, kemudian supernatan dibuang sehingga tersisa pelet saja. Sampel kemudian ditambahkan 500 µL etanol 70% kemudian dihomogenkan. Sampel selanjutnya disentrifuse selama 10 menit pada kecepatan 10.000 ppm pada suhu 4 ºC kemudian disisakan peletnya saja. Sampel kemudian dikeringkan pada mesin vakum selama 30 menit. Sebanyak 5 µL RNAse dan 50 µL Nuclease Free Water (NFW) ditambahkan. Kemudian sampel diinkubasi selama 10 menit pada suhu 37 ºC pada kemudian diikuti inaktifasi selama 10 menit pada suhu 70 ºC.
Amplifikasi dan sekuensing DNA
Analisis molekuler
Karakter molekuler yaitu berupa urutan basa nukleotida yang diperoleh digunakan untuk analisa homologi menggunakan program BLAST-N di situs www.ncbi.nlm.nih.go. Analisa filogenetik dilakukan menggunakan MEGA versi 5 (Tamura et al. 2011).
Analisis data
3 HASIL DAN PEMBAHASAN
Variasi Morfologi Hyphomycetes pada Musa x paradisiaca di Bogor
Tipe konidiofor
Konidiofor adalah struktur yang menopang struktur sporulasi yaitu tempat terdapatnya sel konidiogen. Morfologi konidiofor Hyphomycetes yang ditemukan pada penelitian ini bervariasi mulai dari makronema, semimakronema hingga mikronema. Makronema terbentuk pada spesies yang memiliki struktur konidiofor memanjang dan secara morfologi mudah dibedakan dengan hifa vegetatif (Kirk et al. 2008). Semimakronema ditemukan ketika konidiofor dapat dibedakan dari hifa vegetatif akan tetapi pertumbuhannya terbatas karena terbentuknya sel konidiogen contohnya pada Torula herbarum. Beberapa cendawan, membentuk mikronema karena konidiofornya sangat pendek bahkan seringkali tereduksi sehingga terlihat seperti patahan pada pangkal konidia contohnya pada Dictyosporium zeylanicum (Gambar 1).
Beberapa spesies memiliki konidiofor tunggal seperti pada Stachybotrys subsimplex, bercabang mulai dari pangkal atau pertengahan seperti pada Stachylidium bicolor, atau berupa kumpulan beberapa konidiofor yang disebut sinema seperti pada Phaeoisaria clematidis (Gambar 1). Struktur yang mirip konidiofor dan dapat membingungkan adalah seta. Pada penelitian ini terdapat dua spesies yang memiliki seta, salah satunya yaitu Lacellinopsis sacchari yang berseta sangat panjang bahkan dapat 10 kali lebih panjang dibandingkan dengan konidiofornya. Seta adalah struktur steril seperti batang atau rambut kaku. Perbedaan antara seta dan konidiofor adalah ditemukannya sel konidiogen dan konidia pada konidiofor.
Tipe sel konidiogen
Plastisitas karakter konidiofor dan konidia yang mencakup ukuran dan warna dinilai kurang stabil karena seringkali dipengaruhi oleh kondisi lingkungan. Pada Gambar 1 Variasi konidiofor Hyphomycetes pada pisang di Bogor. Makronema (A-C), konidiofor tunggal pada Stachybotrys subsimplex (A), bercabang pada Stachylidium bicolor (B), sinemata pada Phaeoisaria clematidis (C), semimakronema pada Torula herbarum (D), mikronema atau tereduksi pada Dictyosporium zeylanicum (E). Tanda panah menunjukkan konidiofor. Garis skala = 20 µm
pertengahan abad ke-20, Hughes (1953) mengusulkan delapan seksi atau kelompok berdasarkan ciri konidiofor dan ontogeni konidia. Berbagai pengembangan variasi ontogeni konidia dalam enam dekade terakhir telah digunakan dalam taksonomi cendawan Hyphomycetes (Kendrick 1971; Hennebert & Sutton 1994; Seifert et al. 2011).
Seluruh spesies yang ditemukan dalam penelitian ini memiliki tipe konidiasi blastik dan terdiri atas tujuh kelompok utama ontogeni konidia yaitu terintegrasi terminal, blastik akropetal, fragmentasi basipetal, dentikel bersimpodial, berfialid, tretik, dan basauksik (Tabel 1). Tipe sel konidiogen yang paling banyak ditemukan adalah terminal terintegrasi (8 spesies) dan berfialid (8 spesies) dan yang paling sedikit adalah tipe fragmentasi basipetal (1 spesies). Sementara itu, dua spesies yaitu Hyphomycetes sp. 1 dan Hyphomycetes sp. 2 tidak dapat ditentukan jenisnya karena sel konidiogen tidak ditemukan pada saat pengamatan mikroskopis.
Blastik adalah pembentukan konidia yang diawali dengan membesarnya bakal calon konidia pada ujung atau bagian tertentu pada konidiofor. Istilah yang digunakan untuk sel konidiogen yang hanya menghasilkan satu lokus penghasil konidia ialah mono- (monoblastik, monofialid dan monotretik) dan ketika terdapat lebih dari satu atau banyak lokus disebut poli- (poliblastik, polifialid dan politretik). 1. Terminal terintegrasi
Sebanyak delapan spesies ditemukan memiliki tipe sel konidiogen terminal terintegrasi. Tipe ini umumnya menghasilkan konidia tunggal dari satu lokus atau monoblastik dengan ciri penting sel konidiogen berada pada ujung konidiofor, terintegrasi atau masih menempel dengan konidia dan tanpa diikuti pelepasan konidia (Gambar 2A). Oleh sebab itu, seringkali struktur dari sel konidiogen kurang dapat teramati dengan baik jika menggunakan pengamatan mikroskopis saja. Spesies-spesies dari genus Dictyosporium spp. dan Oncopodium memiliki tipe sel konidiogen ini.
2. Blastik akropetal
Terdapat tiga spesies ditemukan memiliki tipe sel konidiogen blastik akropetal atau mirip dengan Kelompok I B menurut Hughes (1953). Tipe ini memiliki ciri sel konidiogen monoblastik atau poliblastik dan menghasilkan konidia yang membentuk rangkaian rantai akropetal (Gambar 2B). Konidia yang paling muda biasanya lebih pucat dan lebih kecil berada pada ujung rantai sedangkan konidia yang paling tua berada pada pangkal rangkaian.
3. Berdentikel
Gambar 2 Tipe-tipe ontogeni Hyphomycetes pada pisang di Bogor. Terminal terintegrasi pada Oncopodium javanicum (A), blastik akropetal pada Lacellinopsis sacchari (B), tretik pada Dendryphiella vinosa (C), berdentikel pada Ramichloridium musae (D), berfialid pada Stachylidium bicolor (E), fragmentasi basipetal pada Torulaherbarum (F), basauksik pada Spegazzinia deightonii (G), sel konidiogen tidak teramati (H-I). Garis skala = 10 µm
4. Berfialid
Tipe sel konidiogen berfialid sesuai dengan yang dikemukakan Hughes (1953) pada Kelompok IV (Gambar 2E). Kadang kala sporulasi diawali dengan terbentuknya sel-sel pada ujung konidiofor atau ujung dari percabangan konidiofor. Sel-sel ini umumnya memiliki bentuk dan ukuran yang seragam, biasanya silindris atau lonjong atau menyerupai botol, memiliki lebar yang sama atau pangkal lebih lebar daripada ujungnya. Struktur tersebut disebut fialid.
5. Tretik
Sebanyak empat spesies ditemukan memiliki tipe sel konidiogen tretik yaitu dihasilkannya konidia melalui sebuah lubang renik atau pori pada dinding konidiofor. Konidia yang dihasilkan disebut atau dikenal sebagai porospora atau porokonidia. Tipe ini seperti yang dikemukakan pada seksi VI oleh Hughes (1953). Jika konidiofor hanya memiliki satu pori maka disebut monotretik misalnya pori tunggal pada ujung konidiofor Bipolaris sp. dan lebih dari satu atau banyak pori
D
disebut politretik misalnya pori baik pada terminal ataupun interkalar pada Dendryphiella vinosa (Gambar 2C).
6. Fragmentasi basipetal
Hanya terdapat satu spesies yang memiliki tipe sel konidiogen ini yaitu Torula herbarum (Gambar 2F). Tipe ini seperti yang dikemukakan oleh Hughes (1953) pada seksi VII. Sel konidiogen ditandai dengan perubahan konidia pada pangkal yang akan termodifikasi menjadi sel konidiogen kembali dan terfragmentasi dari konidiofornya.
7. Basauksik
Spesies tertentu tipe ontogeninya sangat khas dan sangat memudahkan ketika mengelompokkannya. Tipe basauksik ini seperti yang dikemukakan oleh Hughes (1953) pada seksi yang terakhir yaitu seksi VIII. Terdapat dua spesies yang ditemukan memiliki tipe sel konidiogen ini. Salah satu contoh adalah Spegazzinia deightonii (Gambar 2G). Pertumbuhan diawali dengan tumbuh keluarnya hifa konidiogen fertil dari pangkal sel ibu, kemudian konidia diproduksi secara bebas dari pelepasan hifa fertil yang memanjang.
Gambar 3 Lima Tipe konidia yang ditemukan pada Musa x paradisiaca. Amerospora yaitu konidia tidak bersekat (A). Fragmospora yaitu konidia bersekat lebih dari dua atau banyak (B). Keiroidspora yaitu konidia bersekat banyak menggepeng pada satu sisi dan seperti kumpulan jemari tangan (C). Diktiospora, konidia bersekat banyak dan tidak menggepeng (D). Staurospora, konidia seperti bintang, memiliki lebih dari satu sumbu karena adanya perpanjangan bagian konidia ke beberapa arah (E)
Tipe konidia
Konidia adalah propagul yang dihasilkan oleh sel konidiogen dan akan tumbuh dan berkembang menjadi koloni baru ketika lingkungan hidup mendukung.
A
B
C
Karakter konidia merupakan salah satu karakter penting dalam taksonomi cendawan, yang mencakup bentuk, ornamentasi, pigmentasi, dan ukuran. Secara umum, berdasarkan bentuk dan jumlah sekatnya konidia dibedakan menjadi: 7 macam yaitu amerospora, keiroidspora, diktiospora, didimospora, helikospora, fragmosporadanstaurospora.
Pada penelitian ini seluruh tipe konidia ditemukan kecuali didimospora (konidia bersekat satu) dan helikospora(konidia heliks) (Gambar 3). Amerospora adalah konidia tidak bersekat contohnya konidia pada genus Stachybotrys. Fragmospora adalah konidia bersekat lebih dari satu atau banyak contohnya yaitu konidia bersekat tiga pada Dendryphiella vinosa dan Pyriculariopsis parasitica. Spesies pada genus Dictyosporium memiliki dua tipe konidia: keiroidspora yaitu konidia bersekat banyak, dan menggepeng pada satu sisi sehingga seperti kumpulan jemari tangan, dan diktiospora yaitu konidia bersekat banyak, tidak menggepeng melainkan terdiri dari beberapa lapisan sel. Beberapa spesies memiliki konidia yang sangat unik sehingga dengan mudah dibedakan dari kelompok cendawan lainnya misalnya pada genus Tetraploa yang memiliki tipe konidia staurospora yaitu konidia seperti bintang, memiliki lebih dari satu sumbu karena adanya perpanjangan bagian konidia ke beberapa arah.
Keanekaragaman Hyphomycetes pada Musa x paradisiaca di Bogor
Hasil identifikasi dan inventarisasi
Berdasarkan hasil eksplorasi lapangan, 31 spesies Hyphomycetes teridentifikasi dari 127 koleksi spesimen yang diperoleh di sekitar Bogor pada Musa x paradisiaca (Tabel 1). Spesies yang berhasil diidentifikasi, dideskripsikan dan penguraian taksonomi masing-masing diurutkan berdasarkan spesies pembentukan dan perkembangan konidia. Dua puluh sembilan spesies teridentifikasi setidaknya hingga ke tingkat genus dan termasuk dalam 20 genus.
Genus-genus yang ditemukan pada penelitian ini adalah Bipolaris (1 spesies), Curvularia (1 spesies), Dendryphiella (1 spesies), Dendryphion (1 spesies), Dictyosporium (4 spesies), Exochalara (1 spesies), Lacellinopsis (1 spesies), Oncopodium (1 spesies), Periconia (1 spesies), Phaeoisaria (1 spesies), Phaeostalagmus (1 spesies), Pithomyces (2 spesies), Pyriculariopsis (1 spesies), Ramichloridium (1 spesies), Spegazzinia (2 spesies), Stachybotrys (5 spesies), Stachylidium (1 spesies), Tetraploa (1 spesies), Torula (1 spesies), dan Zygosporium (1 spesies). Secara keseluruhan, Stachybotrys adalah genus yang paling banyak ditemukan karena terdiri atas 5 spesies, kemudian diikuti genus Dictyosporium yang terdiri atas 4 spesies.
Tabel 1 memperlihatkan rekaman hasil eksplorasi dibandingkan dengan dua penelitian sebelumnya yang juga merekam Hyphomycetes pada spesies-spesies Musa (Ellis 1971, 1976; Photita et al. 2003). Sebagian besar spesies-spesies yang ditemukan adalah catatan baru pada pisang di Indonesia. Empat di antaranya adalah spesies baru yang berturut-turut diusulkan diberi nama Dictyosporium bogoriense Rifai & Panjaitan, Stachybotrys musicola Rifai, Stachybotrys atrolaevis Panjaitan & Sukarno, dan Oncopodium javanicum Panjaitan & Rifai.
spesies yang ditemukan pada penelitian ini maupun kedua penelitian yang dibandingkan (Ellis 1971, 1976; Photita et al. 2003). Keempat spesies tersebut adalah Stachybotrys subsimplex, Stachylidium bicolor, Pyriculariopsis parasitica, dan Tetraploa aristata. Rendahnya tumpang tindih ini menunjukkan bahwa Hyphomycetes pada pisang di Bogor, Indonesia beranekaragam dan berbeda dengan daerah dan negara lainnya. Hal tersebut kemungkinan dapat dipengaruhi oleh iklim dan distribusi geografi, spesifitas inang dan kemunculan cendawan kembali pada inang (Zhou & Hyde 2001).
Tabel 1 Hyphomycetes pada Musa x paradisiaca beserta perbandingannya dengan Hyphomycetes pada spesies-spesies Musa lainnya
Tipe Sel Konidiogen Spesies B E P
Terintegrasi terminal Dictyosporium bogoriense sp.nov X
Dictyosporium heptasporum X X
Dictyosporium hydei X
Dictyosporiumzeylanicum X
Oncopodium javanicum sp.nov X
Pithomyces sp. 1 X
Pithomyces sp. 2 X
Tetraploa aristata X X X
Blastik akropetal Lacellinopsis sacchari X
Periconia sp. X
Zygosporium oscheoides X X
Berfialid Exochalara sp. X
Phaeostalagmus cyclosporus* X
Stachybotrys atrolaevis sp.nov* X
Stachybotrys echinata* X
Stachybotrys levispora X
Stachybotrysmusicola sp.nov X
Stachybotryssubsimplex X X X
Stachylidium bicolor X X X
Berdentikel Phaeoisaria clematidis X
Pyriculariopsis parasitica X X X
Ramichloridium musae X
Tretik Bipolaris sp.* X
Curvularia lunata X
Dendryphiella vinosa X
Dendryphion comosum X
Fragmentasi basipetal Torula herbarum* X X
Basauxic Spegazzinia deightonii X
Spegazzinia tessarthra X X
? Hyphomycetes sp. 1 X
? Hyphomycetes sp. 2* X
Spesimen yang diamati meliputi batang, pelepah, dan daun pisang yang telah mengering. Pisang adalah tanaman tidak berkayu dan cukup cepat terdekomposisi. Oleh sebab itu, cendawan yang mengkolonisasi jaringannya harus cepat melakukan sporulasi dengan tujuan menyebarkan spora sebelum jaringannya terurai secara sempurna. Cendawan anamorf termasuk Hyphomycetes lebih cepat bersporulasi dibandingkan cendawan yang membentuk fase teleomorf. Sehingga, Hyphomycetes paling banyak ditemukan termasuk pada tanaman monokotil lainnya contohnya seperti pada nipah (Hyde & Alias 2000), bambu (Hyde et al. 2001), rumput (Wong dan Hyde 2001), dan palem (Yanna et al. 2001).
Spesies-spesies yang ditemukan termasuk ke dalam kelompok cendawan saprofit dan beberapa di antaranya diketahui berpenyebaran kosmopolit. Dua spesies yang diketahui merupakan patogen pada pisang ditemukan dan dapat bertahan hidup sebagai saprofit yaitu Pyriculariopsis parasitica dan Ramichloridiummusae. Kedua spesies tersebut dikenal sebagai penyebab penyakit bercak daun pada pisang dan kultivarnya (Shivas et al. 2010). Hal serupa juga ditemukan oleh peneliti lainnya yang juga menemukan beberapa spesies patogen yang dapat hidup sebagai saprofit (Photita et al. 2003). Hasil penelitian Photita et al. (2001 a) yang menggali cendawan endofit pada pisang liar di Thailand, menduga bahwa P. parasitica adalah patogen laten karena dapat diisolasi dari daun sehat Musa acuminata. Hal ini menunjukkan bahwa spesies ini memiliki persistensi dan adaptasi yang tinggi terkait mode perolehan nutrisi di dalam tubuh inangnya.
Hasil isolasi spora tunggal
Dari seluruh spesimen yang didapatkan, terdapat 7 spesies yang dapat dikulturkan yaitu Bipolaris sp, Hyphomycetes sp.2, Phaeostalagmus cyclosporus, Pyriculariopsis parasitica, Stachybotrys atrolaevis, S. echinata, dan Torula herbarum. Walaupun demikian, isolat P. parasitica tidak dapat dipertahankan dan dimurnikan kulturnya karena kontaminasi oleh cendawan lainnya. Menurut Choi et al. (1999) terdapat beberapa faktor penyebab tidak berhasilnya isolasi spora tunggal yaitu tidak semua spora dalam keadaan viabel ketika dikoleksi, tidak semua spora dapat bergerminasi pada media buatan seperti PDA, kontaminasi oleh cendawan lain atau bakteri (Choi et al. 1999), dan keterbatasan dalam metode isolasi.
Kunci Identifikasi Genus
1. A. Konidiofor basauksik, konidia terdiri atas dua tipe ... Spegazzinia B. Konidiofor akroauksik, konidia hanya satu tipe ... 2 2. A. Konidiofor kebanyakan mikronema atau tereduksi ... 3 B. Konidiofor kebanyakan makronema atau semi-makronema ... 5 3. A. Sel konidiogen monoblastik, konidia bersekat banyak, menjari, mengge-
peng atau menyilindris, beberapa memiliki embelan ... Dictyosporium B. Sel konidiogen monoblastik atau poliblastik, ... 4 4. A. Konidia membintang, embelan berdivergen ... Tetraploa
B. Konidia mengavokad, mengavokad sungsang, sekat transversal 1-12 ... ... Pithomyces 5. A. Konidiofor sinema, sel konidiogen berdentikel, konidia tidak bersekat, hia-
B. Konidiofor mononema ... 6 6. A. Konidiofor melebar seperti kandung kemih, konidia terintegrasi tidak terle-
pas dari sel konidiogen dan konidiofor, bersekat banyak, klatrat atau muriform ... Oncopodium B. Konidiofor umumnya seperti tangkai, terjadi pelepasan konidia dari sel ko-
nidiogen atau konidiofor ... 7 7. A. Sel konidiogen dapat terbentuk kembali saat terjadi pemutusan sel pangkal
konidia dewasa (fragmentasi basipetal), konidia moniloid ... Torula B. Sel konidiogen tidak mengalami fragmentasi ... 8 8. A. Konidia bersekat ... 9 B. Konidia kebanyakan tidak bersekat ... 13 9. A. Sel konidiogen berupa dentikel, konidia bersekat 1-7, menggelendong,
mengavokad sungsang, atau gada terbalik, silindris ... Pyriculariopsis B. Sel konidiogen tretik ... 10 10. A. Konidia bersekat tidak sejati, silindris, menggada, atau gada terbalik, sekat
lebih dari 4, germinasi biasanya dari kedua ujung konidia ... Bipolaris B. Konidia bersekat sejati ... 11 11. A. Konidia biasanya melengkung, bersekat 2 atau lebih ... Curvularia
B. Konidia tidak melengkung ... 12 12. A. Konidiofor bercabang, membengkak atau melutut pada ujung atau interka-
lar, konidia bundar telur, lonjong, silindris, bersekat 0-3 ... Dendryphiella B. Konidiofor bercabang banyak pada ujung dan melebar seperti kipas.
Konidia moniloid, bersekat banyak (lebih dari 4-5 sekat) ... Dendryphion 13. A. Sel konidiogen berupa dentikel, awalnya terminal menjadi simpodial, koni-
dia bersekat 0-1, umumnya membulat telur sungsang, bulat hingga jorong ... Ramichloridium B. Sel konidiogen dihasilkan dari pembengkakan ujung konidiofor, atau fialid atau dari vesikel, atau metula atau ampula, semua konidia amerospora ... 14 14. A. Seta umumnya ada, sel konidiogen berupa ampula, konidia bulat dan mem- bentuk rangkaian akropetal ... Lacellinopsis B. Seta tidak ada ... 15 15. A. Sel konidiogen bukan fialid ... 16 B. Sel konidiogen fialid ... 17 16. A. Sel konidiogen umumnya poliblastik, bulat atau agak membulat, konidia
membentuk rangkaian akropetal ... Periconia B. Sel konidiogen umumnya monoblastik, berupa vesikel yang agak membeng-
kok, konidia dihasilkan membentuk rangkaian akropetal ... Zygosporium 17. A. Fialid tunggal, berkolaret, terintegrasi di ujung konidiofor, konidia silindris,
hialin ... Exochalara B. Fialid banyak, biasanya di ujung konidiofor atau ujung percabangan ... 18 18. A. Fialid tersusun membentuk mahkota pada ujung konidiofor, konidia umum- nya berpigmentasi ... Stachybotrys B. Fialid tersusun vertisil pada konidiofor atau percabangan, konidia hialin ..
... 19 19. A. Konidiofor cokelat, fialid hialin, berkolaret ... Phaeostalagmus
Taksonomi
D i c t y o s p o r i u m
Dictyosporium Corda, Weitenweber’s Beitr. Ges. Nat. Heilk., Prag 1: 87. 1836; Ellis, Dematiaceous Hyphomycetes: 55. 1971; Seifert, Morgan-Jones, Gams, Kendrick, The Genera of Hyphomycetes: 181. 2011; Goh, Hyde, Ho & Yanna, Fungal Diversity 2: 65. 1999; Silva, Gusmao, Castaneda-Ruiz, Mycotaxon 130: 1125. 2015
= Speira Corda, Icon. Fung., 1: 9. 1837
= Cattanea Garovaglio , Rc. Ist. lomb. Sci. Lett., 2 Ser., 8: 125. 1875 Spesies tipe: Dictyosporium elegans Corda.
Koloni pada substrat menyebar atau sporodokia, cokelat gelap atau hitam. Miselium sebagian besar terbenam dalam substrat. Konidiofor mikronema, semi-mikronema, lurus hingga membengkok, sangat pendek atau tereduksi, hialin atau berpigmentasi. Sel konidiogen monoblastik, terminal, membulat, agak membulat, membotol, hialin hingga berpigmentasi. Konidia tunggal, keiroid atau seperti jemari tangan dan menggeppeng pada satu sisi, diktiospora, silindris atau tidak menggepeng, hialin atau berpigmen, bersel banyak, permukaan halus. Embelan ada atau tidak; di pangkal, pertengahan hingga ujung konidia, bulat hingga silindris, hialin hingga subhialin.
Kunci identifikasi Dictyosporium
1. A. Konidia menggepeng pada satu sisi, lengan umumnya berjumlah 5 ... ... D. zeylanicum B. Konidia tidak menggepeng, lengan umumnya berjumlah lebih dari lima .... 2 2. A. Panjang konidia kurang dari 40 µm, embelan bulat jumlah sampai tiga di atas pangkal ... D.hydei B. Panjang konidia bisa lebih dari 50 µm, tanpa embelan ... 4 3. A. Konidia memiliki 7-8 baris lengan, tiap lengan terdiri 14-16 sel ...
... D. heptasporum B. Konidia memiliki 5-6 baris lengan, tiap lengan terdiri 11-13 sel ...
... D. bogoriense
Dictyosporium bogoriense Rifai & Panjaitansp nov.(Gambar 4)
Colonies on natural substrate conspicuous, scattered and form compact sporodochia, dark brown to black. Mycelium mostly immersed on substrate. Conidiophore micronematous or reduced, hyaline. Conidiogenous cell terminal, integrated, monoblastic, hyaline. Conidia not complanate, cylindrical, 51-60 x 15-18 µm, multiseptate, smooth-walled, uniformly dark brown, consist of 5-6 rows of cells appressed closely together, each row mostly has same length and consists of 11-13 cells, apex incurved. Conidia lacking appendages.
Etimologi: —bogoriense dari daerah asal ditemukannya.
Spesimen yang diamati: pada pelepah kering Musa x paradisiaca, Bogor, Dramaga,16 Maret 2015, DMP77.
embelan memiliki konidia yang menggepeng sedangkan 11 lainnya tidak menggepeng melainkan menyilindris. Dictyosporium bogoriense memiliki morfologi yang mirip dengan D. heptasporum. Kedua spesies memiliki konidia yang tidak menggepeng. Perbedaanya adalah berdasarkan ukuran konidia, jumlah baris lengan dan jumlah sel pada setiap baris lengan. Konidia Dictyosporium bogoriense rata-rata berukuran kurang dari 60 µm, memiliki 5-6 baris lengan dan setiap lengan terdiri atas 11-13 sel sedangkan konidia D. heptasporum berukuran lebih besar yaitu 60-80 µm, secara konsisten memiliki 7 baris lengan dan setiap lengan memiliki 14-16 sel.
Gambar 4 Dictyosporium bogoriense pada substrat alami. Garis skala = 10 µm
Gambar 5 Dictyosporium heptasporum pada substrat alami. Konidia silindris tersusun oleh 7 baris lengan tersusun rapat. Garis skala = 10 µm
Dictyosporium heptasporum (Garov.) Damon, Lloydia 15:118. 1952 (Gambar 5) =Cattanea heptaspora Garov., Rc. Ist. Lomb., Milano II 8: 125. 1875
Distribusi: Amerika Utara, Eropa, (Ellis 1971), Australia, Hongkong, Thailand (Goh et al. 1999; Photita et al. 2003), Burma (Thaung 2008), Filipina (Whitton et al. 2012), Indonesia.
Habitat: mudah ditemukan misalnya pada pisang, bambu dan kelapa.
Spesimen yang diamati: pada pelepah kering Musa x paradisiaca, Bogor, Bubulak, 28 Pebruari 2015, DMP59.
Catatan: Dictyosporium heptasporum mudah dikenali karena umumnya memiliki 7 baris lengan yang ujungnya melengkung. Spesimen diamati memiliki konidia yang lebih sempit dibandingkan dengan Ellis (1971) yang disebutkan mencapai 30 µm.
Gambar 6 Dictyosporium hydei pada substrat alami. Konidia memiliki tiga buah embelan membulat di bagian atas pangkal. Garis skala= 10 µm
Dictyosporium hydei Prasher & Verma, Phytotaxa 204 (3): 196. 2015 (Gambar 6) Koloni pada substrat sporodokia, menyebar hingga 1 cm, cokelat kehitaman. Miselium sebagian besar terbenam di substrat. Konidiofor tereduksi. Sel konidiogen agak membulat dan hialin. Konidia tidak menggepeng, menyilindris, membulat telur sungsang, 31-38 x 14.7-20 µm, bersekat banyak, cokelat kehijauan hingga cokelat, terdiri dari (5)-6 baris lengan yang sama panjang dan tersusun rapat, tidak menyempit pada sekat; setiap lengan terdiri (5-6)-8 sel. Embelan supra-basal, berjumlah 1-3, hialin, agak membulat hingga membulat, diameter 6.5-10.5 µm, hialin, halus, tidak bersekat.
Distribusi: India (Prasher & Verma 2015), Indonesia. Habitat: ditemukan pada kulit kayu Tecoma stans.
Spesimen yang diamati: pada pelepah kering Musa x paradisiaca, Bogor, Dramaga, 16 Maret 2015, DMP 68; Bogor, 9 April 20, DMP 87.
Catatan: Penemuan spesies ini adalah rekaman pertama pada Musa x paradisiaca sekaligus rekaman baru di Indonesia setelah pertama kali dideskripsikan dari Tecoma stans (Prasher & Verma 2015). Konidia yang ditemukan pada penelitian ini berukuran lebih besar (31-38 x 14.7-20 µm) dibandingkan dengan holotipe (26-35) x 14-17 µm) sedangkan embelan yang ditemukan berukuran lebih kecil (6.5-10.5 x 5-8.3 µm) dibandingkan holotipe (11-17 x 8-15 µm).
Dictyosporium zeylanicum Petch, Ann. R. Bot. Gdns Peradeniya 6(3): 252. 1917 (Gambar 7)
konidiogen terminal, subhialin. Konidia menggepeng pada satu sisi, membulat telur sungsang, 24-33.6 x 17.5-22.2 µm, tebal mencapai 8 µm, bersekat banyak, menyempit pada sekat, umumnya terdiri dari 5 baris lengan yang tersusun rapat; tiga baris lengan di tengah lebih panjang, cokelat tua dan masing-masing terdiri dari 4-6 sel, sel-sel di ujung lengan lebih lebar; dua baris lengan di pengapit di pinggir lebih pendek, sedikit lebih pucat dan masing-masing terdiri (6)-7 sel, sel-sel ujung lebih kecil dan melengkung ke arah dalam. Konidia tidak memiliki embelan.
Distribusi: Argentina, Belize (Whitton et al. 2012), Sri Lanka (Goh et al. 1999), Indonesia.
Habitat: umum ditemukan pada kayu atau serasah yang membusuk.
Spesies yang diamati: pada tangkai pelepah kering Musa x paradisiaca, Bogor, Babakan Lebak, 10 Oktober 2014, DMP16; Bogor, Dramaga, 10 Juni, DMP109.
Catatan: Secara morfologi, spesies ini sangat mirip dengan D. rhopalocystidis karena memiliki konidia yang menggepeng pada satu sisi dan ukuran konidia yang relatif sama, akan tetapi spesies tersebut memiliki konidia yang tercatat memiliki 5-6 baris lengan dan jumlah sel per barisnya adalah 5-10 (McKenzie 2010), sedangkan pada spesies ini secara konsisten ditemukan 5 baris lengan dengan jumlah sel per barisnya adalah 4-8. Spesimen yang ditemukan pada penelitian ini memiliki ukuran konidia yang sedikit lebih kecil tetapi tetap masuk dalam kisaran ukuran yang disampaikan oleh Goh et al. (1999).
O n c o p o d i u m
Oncopodium Sacc., Annls mycol. 2(1): 19. 1904; Ellis, Dematiaceous Hyphomycetes: 111. 1971; Seifert, Morgan-Jones, Gams, Kendrick, The Genera of Hyphomycetes: 320. 2011
Spesies tipe : Oncopodium antoniae Sacc. & D. Sacc.
Koloni pada substrat alami membentuk sporodokia, bergerombol, cokelat hingga hitam. Miselium terbenam dalam substrat atau sebagian di atas substrat. Konidiofor makronema, mononema, tunggal atau bercabang di pangkal, lurus atau membengkok, tidak bersekat, hialin, halus, pangkal sempit kemudian melebar seperti kantong kemih. Sel konidiogen monoblastik, terminal, membulat atau agak membulat kadang menggepeng. Konidia agak membulat hingga membulat, klatrat atau muriform, bersekat banyak, dua sisi mengerucut atau dua sisi menggepeng, bagian dorsal sedikit terbenam, hialin hingga berpigmentasi.
Gambar 8 Oncopodium javanicum pada substrat alami. Konidiofor melebar seperti kandung kemih. Garis skala = 10 µm
Oncopodium javanicum Panjaitan & Rifai sp. nov (Gambar 8)
Colony on natural substrate forms sporodochial, round, 0.1-0.2 cm in diam., black. Mycelium mostly immersed. Conidiophore macronematous, mononematous, unbranched, non-septate, hyaline, length about 15-33 µm, narrow at the base 2.8-4,2 µm, then swelling like a bladder to 9.4-12 µm and become smaller up to apex 6.8-8 µm. Conidiogenous cell terminal, monoblastic, integrated, hyaline and smooth. Conidia subglobose, 16.6-20 x 11-14 µm, multiseptate, constricted at the septate, septate not conspicious, flattened above and clathrate. Conidia consist of a dome-shaped cap of dark brown to black cells and surrounds with hyaline cells.
Etimologi: —javanicum berasal dari nama pulau ditemukannya.
Spesimen yang diamati: pada pelepah kering Musa x paradisiaca, Bogor, Dramaga, 16 Maret 2015, DMP75.
P i t h o m y c e s
Pithomyces Berk. & Broome, Botanical Journal of the Linnean Society 14: 100. 1873; Ellis, Dematiaceous Hyphomycetes: 43. 1971; Seifert, Morgan-Jones, Gams, Kendrick, The Genera of Hyphomycetes: 352. 2011
=Scheleobrachea S. Hughes, Can. J. Bot. 36: 802. 1958 Spesies tipe: Pithomyces flavus Berk. & Broome.
Koloni menyebar, hijau kecokelatan, cokelat atau hitam. Miselium semuanya atau sebagian besar terbenam dalam substrat. Konidiofor mikronema atau semi-makronema, mononema, bercabang, lurus atau sedikit membengkok, subhialin atau hijau zaitun pucat hingga cokelat, halus atau kasar. Sel konidiogen monoblastik atau poliblastik, interkalar atau terkadang terminal, lonjong, berdentikel; dentikel lonjong, pendek hingga panjang. Konidia tunggal, jorong, berbentuk gada, seperti jeruk lemon, membulat telur sungsang, lonjong dengan ujung-ujung membulat, mengavokad atau mengavokad sungsang, berwarna seperti jerami, hingga cokelat kehitaman pekat, halus, kasar, dengan sekat transversal 1-12.
Gambar 9 Pithomyces sp. 1 pada substrat alami. Garis skala = 10 µm
Pithomyces sp. 1 (Gambar 9)
Koloni pada substrat membeledu, cokelat. Miselium sebagian di atas permukaan substrat. Sel konidiogen monoblastik. Konidiofor mikronema, tunggal, tidak bercabang, lurus atau sedikit membengkok, hialin hingga subhialin, sangat pendek, 2.5-3 x 1.2-1.5 µm. Sel konidiogen terminal. Konidia tunggal, jorong, 8.3-11.4 x 4.4-5.5 µm, kasar, kecokelatan.
Spesimen yang diamati: pada pelepah kering Musa x paradisiaca di Bogor, Dramaga, 3 Desember 2015, DMP124.
Catatan:Spesimen yang diamati tidak dapat diidentifikasi hingga ke tingkat spesies.
Pithomyces sp. 2(Gambar 10)
Koloni membeledu, cokelat. Konidiofor mikronema, sangat pendek seringkali tereduksi, 2.75-3 x 1.8-1.9 µm, hialin, halus. Sel konidiogen monoblastik. Konidia tunggal, mengavokad, 16.5-24 x 9-12 µm, lurus atau kadang melengkung, menyempit pada sekat pertama, kebanyakan bersekat 3, permukaan agak kasar, hijau kecokelatan hingga cokelat gelap.
Gambar 10 Pithomyces sp. 2 pada substrat alami. Garis skala = 10 µm
T e t r a p l o a
Tetraploa Berk. & Broome, Annals and Magazine of Natural History 2, 5: 459. 1850; Ellis, Dematiaceous Hyphomycetes: 51. 1971; Rifai, Zainuddin & Cholil, Reindwardtia 10, 4: 419. 1988; Seifert, Morgan-Jones, Gams, Kendrick, The Genera of Hyphomycetes: 436. 2011
Spesies tipe: Tetraploa aristataBerk. & Broome.
Koloni menyebar, seperti butiran yang berduri, cokelat hingga cokelat kehitaman. Konidiofor mikronema atau tereduksi, bercabang dan saling terjalin membentuk jaring-jaring, hialin hingga subhialin, kasar. Sel konidiogen monoblastik atau poliblastik, interkalar, silindris. Konidia tunggal, staurospora, cokelat, sedikit kasar hingga kasar, pada konidia yang matang terdapat alur dangkal di antara 3-4 kolom sel yang berkembang atau memanjang secara bebas, dan cenderung memiliki embelan yang berdivergen.
Gambar 11 Tetraploa aristata pada substrat alami. Garis skala = 10 µm
Tetraploa aristata Berk. & Broome, Annals and Magazine of Natural History 2, 5: 459. 1850 (Gambar 11)
Koloni menyebar, cokelat gelap. Konidiofor tereduksi. Sel konidiogen monoblastik. Konidia tunggal, membintang, 30-32 x 25-27 µm, terdiri atas 8 sel yang tersusun dalam 3-4 kolom, sedikit menyempit pada sekat, cokelat, sedikit kasar, ujung konidia membentuk embelan yang berdivergen ke arah yang berbeda; embelan 3-4, di ujung konidiofor, lurus, panjang 29-36 µm, cokelat pucat.
Habitat: pada rerumputan yang mengering.
Spesimen yang diamati: pelepah kering Musa x paradisiaca, Bogor, Dramaga, 9 April 2015, DMP88; DMP89; 17 April 2015, DMP106, Bogor, Dramaga, 10 Juni 2015, DMP 110.
Catatan: Tetraploa aristata sebelumnya pernah dilaporkan dan ditemukan pada rumput kering di Kebun Raya Bogor (Rifai et al. 1988) dan tidak ada perbedaan yang signifikan dibandingkan dengan spesimen yang diamati.
L a c e l l i n o p s i s
Lacellinopsis Subramanian Proc. Indian Acad. Sci., Sect. B 37: 100-105. 1953; Ellis, Dematiaceous Hyphomycetes: 355. 1971; Seifert, Morgan-Jones, Gams, Kendrick, The Genera of Hyphomycetes : 352. 2011
Spesies tipe: Lacellinopsis sacchari Subram.
Koloni kecil, menyebar, abu-abu, hijau zaitun, cokelat atau hitam. Miselium sebagian besar terbenam dalam substrat. Stroma kecil, cokelat. Seta tegak tidak bercabang atau bercabang, tegak, lurus atau kadang membengkok, bersekat, lebar pada pangkal kemudian semakin mengecil hingga ke ujung, cokelat gelap hingga cokelat kehitaman dan semakin pucat ke ujung, halus. Konidiofor makronema, mononema, tidak bercabang, lurus atau berlekuk-lekuk, bersekat, cokelat hingga cokelat gelap atau agak memucat di ujung, halus atau kasar. Sel konidiogen terminal terkadang menjadi interkalar, poliblastik, memangkok, jorong atau jorong dengan ujung merata. Konidia membentuk rangkaian akropetal, membulat, agak bulat, tidak bersekat, halus atau kasar, cokelat hingga cokelat kemerahan.
Lacellinopsis sacchari Subram., Proc. Indian Acad. Sci., Sect B, 37: 100-105. 1953 (Gambar 12)
Koloni menyebar, berambut, seperti bulu-bulu yang merebah, lebar koloni mencapai 0.4 cm, cokelat. Seta panjangnya mencapai 400 µm, cokelat gelap dan lebih pucat di ujung. Konidiofor muncul dekat seta, terkadang berkelompok, 10-29 x 3-5.8 µm, lurus atau sedikit membengkok, cokelat. Sel konidiogen terminal atau kadang menjadi interkalar, memangkok, jorong, 3.6-11.8 x 6.6-9.2 µm, sedikit kasar, cokelat hingga cokelat kehitaman dan ujung memucat. Konidia bulat, agak membulat, diameter 4-7 µm, kasar, cokelat kemerahan.
Distribusi: Ghana, Kuba, India, Jamaika, Pakistan, Tanzania, Venezuela, Zambia (Ellis 1971), Indonesia.
Habitat: Pada dedaunan kering Oryza, Saccharum, Sorghum.
Spesimen yang diamati: Pelepah kering Musa x paradisiaca L., Bogor, Dramaga, 3 Desember 2015, DMP126.
Gambar 12 Lacellinopsis sacchari pada substrat alami. Garis skala = 10 µm
P e r i c o n i a
Periconia Tode, Fung. mecklenb., 2:2. 1791; Ellis, Dematiaceous Hyphomycetes: 344. 1971; Seifert, Morgan-Jones, Gams, Kendrick, The Genera of Hyphomycetes : 336. 2011
Spesies tipe: Periconia lichenoides Tode
Koloni menyebar, tegakan seperti jarum pentul atau membulu, kelabu, cokelat kehitaman hingga hitam. Miselim kebanyakan terbenam dalam substrat sebagian di atas permukaan. Stroma cokelat gelap. Seta terkadang terdapat di ujung konidiofor yang steril. Konidiofor makronema atau mikronema, mononema, tegak, lurus sedikit membengkok, bersekat, tunggal atau bercabang di bagian ujung, pangkal lebar kemudian semakin mengecil, ujung kadang membengkak, cokelat tua hingga cokelat kehitaman, halus. Sel konidiogen terminal, bulat atau agak membulat, jorong, monoblastik atau poliblastik. Konidia terbentuk dalam rangkaian akropetal, bulat, agak membulat, jorong, tidak bersekat, berbingkahan atau berduri, subhialin hingga berpigmentasi.
Periconia sp.(Gambar 13)
Spesimen yang diamati: Pelepah kering Musa x paradisiaca, Bogor, Dramaga, 29 Mei 2015, DMP115.
Catatan: Spesimen yang diamati memiliki morfologi dan ukuran konidia yang mirip dengan P. digitata akan tetapi spesies tersebut memiliki ciri khas percabangan konidiofor yang tersusun tidak terlalu rapat dan terlihat jelas terutama saat kepala konidiofor sudah terbentuk dengan matang atau dewasa (Ellis 1971). Pada spesimen yang diamati, percabangan di kepala konidiofor tampak jelas saat kepala konidiofor belum dewasa dan tersusun rapat. Saat dewasa percabangan tidak teramati dengan jelas karena tertutup massa konidia.
Gambar 13 Periconia sp. pada substrat alami. Garis skala =10 µm
Z y g o s p o r i u m
Zygosporium Mont. Ann. Sci. Nat., Sér. 2, 17: 120-121. 1842; Ellis, Dematiaceous Hyphomycetes: 322. 1971; Seifert, Morgan-Jones, Gams, Kendrick, The Genera of Hyphomycetes: 480. 2011; Whitton, McKenzie, Hyde, Fungal Diversity 12:207-222. 2003
=Pimina Grove, J. Bot., Lond., 26: 206. 1888
=Urobasidium Giesenhagen Flora, Jena, 76: 139. 1893
=Urophiala Vuillemin, Bul. Séanc. Soc. Sci. Nancy, Sér. 3, 11: 158. 1910 Spesies tipe: Zygosporium oscheoides Mont.
mononema, tidak bercabang atau bercabang, cokelat, halus atau sedikit berduri. Sel konidiogen biasanya monoblastik, ampuliform, atau jorong, tekadang berlekuk dan melancip pada satu titik, berdinding tebal, tidak berwarna atau pucat, muncul berpasangan, tiga atau empat pada vesikel yang cokelat tua. Konidia akrogen, jorong, bulat atau agak membulat, hialin hingga cokelat, harus, sedikit kasar hingga kasar dan tidak bersekat.
Gambar 14 Zygosporium oscheoides pada substrat alami. Garis skala = 10 µm
Zygosporium oscheoides Mont., Ann. Sci. Nat., Sér. 2, 77: 121. 1842; Seifert, Morgan-Jones, Gams, Kendrick, The Genera of Hyphomycetes: 480. 2011 (Gambar 14)
Konidiofor mendabus, panjangnya mencapai 75 µm, tebal pada pangkal 3-4 µm, menghasilkan vesikel tunggal pada tangkai pendek langsung dari bagian dasar; bagian atas yang steril terkadang berujung membintul. Vesikel panjangnya mencapai 8-20 µm, 7-10 µm pada bagian ketebalannya. Konidia jorong, tidak berwarna atau cokelat pucat, halus hingga sedikit kasar, 6-12 x 5-7 µm.
Distribusi: Kosmopolit. Pernah ditemukan di Brazil, Ghana, India, Jamaika, Kongo, Kuba, Papua Nugini, Sierra Leone, Tanzania, Uganda dan Venezuela (Ellis 1971), Indonesia.
Habitat: Ditemukan di berbagai spesies tumbuhan.
Spesimen yang diamati: pada daun kering Musa x paradisiaca, Bogor, Jl. Juanda, 30 September 2013, DMP6.
B i p o l a r i s
Bipolaris Shoemaker, Canad. J. Bot. 37: 882. 1959; Seifert, Morgan-Jones, Gams, Kendrick, The Genera of Hyphomycetes : 336. 2011; Manamgoda, Rossman, Castlebury, Crous, Chukeatirote, Hyde, Stud. in Myco. 79: 107. 2014
Spesies tipe: Bipolaris maydis (Y. Nisil & C. Miyake) Shoemaker
4 sekat, hialin, cokelat pucat atau gelap, cokelat kemerahan atau hijau zaitun pucat hingga pekat, bergerminasi dengan tabung germinasi pada salah satu atau kedua ujung konidia. Hilum sedikit menonjol atau terpotong.
Gambar 15 Bipolaris sp pada substrat alami (A-B, I). Konidia membentuk tabung germinasi pada kedua ujungnya (C). Konidia pada media Potato Dextrose Agar (PDA) (D). Koloni yang tumbuh pada PDA inkubasi 2 hari pada suhu ruang tampak atas (E) dan tampak ketika dibalik (F). Koloni yang tumbuh pada PDA inkubasi 7 hari tampak atas (G) dan tampak ketika dibalik (H). Garis skala = 10 µm
Bipolaris sp.(Gambar 15)
Koloni pada substrat membulu, hijau keabu-abuan. Koloni pada PDA cepat tumbuh, diameter mencapai 9 cm inkubasi 7 hari pada suhu ruang, hijau keabu-abuan pada permukaan dan hijau kehitaman ketika dibalik. Miselium sebagian besar di atas permukaan substrat. Sporulasi lambat, biasanya dimulai pada hari ke 14 dan tampak di bagian pinggir koloni, hitam. Konidiofor tunggal tidak bercabang, lurus atau sedikit bercabang, bersekat, halus, cokelat. Sel konidiogen monotretik.
I
E F G H
Konidia silindris atau terkadang seperti gada terbalik, lurus atau sedikit membengkok, 31-61 x 10.5-13.5 µm, bersekat 3-8, hilum terkadang sedikit menonjol di pangkal, germinasi terjadi pada kedua ujung konidia, halus, cokelat kehijauan, isi di dalam sekat terkadang lebih pucat dan berbercak-bercak.
Spesimen yang diamati: pelepah kering Musa x paradisiaca di Bogor, Babakan Lebak, 10 September 2015, DMP116.
Catatan: Spesies-spesies yang termasuk dalam genus Bipolaris umumnya memiliki konidiofor cokelat, konidia silindris yang umumnya memanjang dengan sekat distoseptat dan germinasi bipolar. Pada spesimen yang diamati ciri morfologi yang didapatkan dinilai belum memadai untuk dilakukan identifikasi hingga ke tingkat spesies. Hal ini dikarenakan antar spesies pada genus Bipolaris sendiri memiliki tumpang tindih karakter morfologi yang besar terutama pada karakter ukuran dan bentuk konidiofor dan konidianya (Manamgoda et al. 2014).
C u r v u l a r i a
Curvularia Boedijn, Bull. Jard. Bot. Buitenz., III 13 (1): 123. 1933; Ellis, Dematiaceous Hyphomycetes: 452. 1971; Seifert, Morgan-Jones, Gams, Kendrick, The Genera of Hyphomycetes : 163. 2011
=Malustela Bat. & J.A Lima, Publicações Inst. Micol. Recife 263: 5. 1960 =CurvisporiumCorbetta (as ‘Curvosporium’), Riso n12(3): 28, 30. 1963
Spesies tipe: Curvularia lunata (Wakker) Boedijn = Cochliobolus lunatus Nelson & Haasis.
Koloni menyebar, cokelat, kelabu atau hitam, membulu, atau membeledu. Miselium terbenam dalam substrat. Konidiofor makronema, mononema, lurus atau membengkok, atau melutut, cokelat, biasanya halus. Sel konidiogen politretik, terminal, kemudian menjadi interkalar, simpodial, lonjong atau biasanya membengkak. Konidia tidak bercabang, melengkung atau membengkok, jarang lurus dan memanjang, berbentuk ginjal, membulat telur sungsang atau mengavokad, bersekat 2 atau lebih, sel di tengah biasanya melebar, pucat atau cokelat gelap, biasanya sel yang paling ujung lebih pucat dibandingkan sel lainnya, halus atau kasar, hilum pada beberapa spesies menonjol.
Curvularia lunata (Wakker) Boedijn, Bull. Jard. Bot. Buitenz., 13 (1): 127. 1933 (Gambar 16)
=Acrothecium lunatum Wakker, De ziekten van het Suik. Jav. Die niet door dieren veeroorzaakt worden: 196. 1898
Koloni menyebar, cokelat. Konidiofor tidak teramati. Konidia membengkok, berbentuk ginjal, bersekat 3, 23-30 x 8-11 µm, sel-sel ujung lebih pucat dan lebih kecil, sel ketiga dari pangkal lebih lebar dan gelap dan merupakan titik membengkoknya konidia, halus. Hilum tidak menonjol.
Distribusi: Umum dijumpai pada daerah tropis.
Habitat: sangat umum dan terdapat pada berbagai spesies tumbuhan.
Spesimen yang diamati: pada pelepah Musa x paradisiaca, 3 Desember 2015, Bogor, Dramaga, DMP124.