RAGAM JENIS DAN FLUKTUASI POPULASI
SIMULIUM
(Diptera: Simuliidae) DI KABUPATEN KUPANG NUSA
TENGGARA TIMUR
JULIANTY ALMET
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA*
Dengan ini saya menyatakan bahwa tesis berjudul Ragam Jenis dan Fluktuasi Populasi Simulium (Diptera: Simuliidae) di Kabupaten Kupang Nusa Tenggara Timur adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, Agustus 2015
Julianty Almet
RINGKASAN
JULIANTY ALMET. Ragam Jenis dan Fluktuasi Populasi Simulium (Diptera: Simuliidae) di Kabupaten Kupang Nusa Tenggara Timur. Dibimbing oleh UPIK KESUMAWATI HADI, SUSI SOVIANA.
Lalat hitam (Simuliidae) adalah satu di antara kelompok paling penting dari serangga penggigit, karena kebiasan mengisap darah dan kemampuan untuk menularkan/menyebarkan parasit patogen. Informasi mengenai spesies lalat ini khususnya yang berhubungan dengan aspek taksonomi dan ekologinya di Indonesia, tidak cukup tersedia. Oleh karena itu, penelitian ini dilakukan untuk mengeksplorasi ragam jenis lalat hitam di Indonesia sehingga dapat menjadi sumber informasi yang penting. Penelitian ini bertujuan untuk mengidentifikasi jenis-jenis lalat Simulium pada lokasi air terjun Oenesu dan Oehala, mempelajari habitat larva Simulium; dan aktivitas mengisap darah lalat Simulium pada ternak besar.
Penelitian dilakukan di lokasi penelitian Oenesu Kabupaten Kupang Barat dan di Oehala Kabupaten Timor Tengah Selatan. Adapun metode yang dilakukan terdiri atas 3 tahapan yaitu tahap pertama koleksi dan identifikasi
Simulium pradewasa di sepanjang air terjun berdasarkan ketinggian tempat. Tahap kedua berupa pengukuran karakteristik habitat larva Simulium seminggu sekali yang dilakukan bersamaan dengan pengambilan Simulium pradewasa. Tahap ketiga melakukan koleksi lalat Simulium dewasa di peternakan sapi/kandang ternak, menggunakan perangkap cahaya (light trap) dari pukul 18.00-06.00 dengan frekuensi pengambilan dua minggu sekali selama 3 bulan.
Hasil penelitian menunjukkan bahwa terdapat 4 spesies Simulium yang teridentifikasi di lokasi penelitian Oenesu, adalah S. (Simulium) timorense
(75.59%), S. (Gompostilbia) sundaicum (18.76%), S. (Wallacellum) sp (3.71%),
S. (Nevermania) aureohirtum (1.93%). Sementara itu, terdapat 3 spesies larva
Simulium yang ditemukan di Oehala yaitu, S. (S.) timorense (46.5%), S. (G.) sundaicum (43.1%), S. (N.) aureohirtum (10.2%). Berdasarkan karakteristik habitat larva berupa: suhu, pH, kecepatan arus dan keberadaan naungan. Lokasi air terjun Oenesu tergolong sesuai untuk perkembangan larva Simulium.
S. (S.) timorense memiliki populasi tertinggi pada area T1 (±1200 mdpl) dan T4 (1165 mdpl) dengan jumlah masing-masing 840 dan 689 larva. Berdasarkan uji BNT populasi S. (S.) timorense dan S. (N.) aureohirtum pada setiap area pengamatan (T1-T4) tidak berbeda secara signifikan (P>0.05), sedangkan S. (G.) sundaicum dan S. (Wallacelum) sp pada area T1 dan T2 tidak berbeda signifikan ( P˃ 0.05), sedangkan pada area T3 dan T4 menunjukkan berbeda nyata (P<0.05).
Hasil koleksi lalat Simulium dewasa di kandang ternak ditemukan 2 jenis yaitu S (S.) timorense (71.5%), S (G.) sundaicum (28.8%). Aktivitas mengisap darah dari S. (S). timorense dimulai dari pukul 18.00, sedangkan S. (G.) sundaicum baru dimulai pukul 20.00 dan puncak aktivitas mengisap darah dari kedua jenis lalat ini terjadi pada pukul 00.00-02.00.
SUMMARY
JULIANTY ALMET. Species Diversity and Population Fluctuation of Simulium
(Diptera; Simuliidae) in Kupang District of East Nusa Tenggara. Supervised by
UPIK KESUMAWATI HADI and SUSI SOVIANA.
Black flies (Simuliidae) are one of the most important biting insects due to their blood-sucking habits and ability to transmit parasites. There is no sufficient information of the fly species in Indonesia, particularly in relation to taxonomical and ecological aspects. Therefore, the present study was conducted to explore black flies in Indonesia and was expected to contribute the importance information. This study aimed to identify Simulium diversity from Oenesu and Oehala Waterfalls, study the habitat of Simulium larvae, and the biting activities of Simulium on livestock.
The study was conducted at Oenesu of West Kupang District and Oehala of South Timor Tengah District. The method was divided into three steps:
Simulium weekly collection and identification every week as the first step, the measurement of Simulium larvae habitat characteristic conducted weekly at the same time as the collection of Simulium larval as the second step, and the last step was the collection of adult Simulium flies from cattle farms/livestock pens using light trap for 12 hours (18:00-06:00) once in two weeks for three months.
The study revealed there were four Simulium species from Oenesu site, namely S. (Simulium) timorense (75.59%), S. (Gompostilbia) sundaicum
(18.76%), S. (Wallacellum) sp (3.71%), and S. (Nevermania) aureohirtum
(1.93%); while from Oehala were 3 Simulium larval species, i.e. Simulium (S.) timorense (46.5%), Simulium (G.) sundaicum (43.1%), and Simulium (N.) aureohirtum (10.2%). Oenesu and Oehala study sites were suitable for Simulium
larval growth based on the habitat characteristic such as temperature, pH, current to velocity, and shade presence.
Simulium timorense had the highest population at T1 (1200 mdpl) and T4 (1165 mdpl). While based on LSD (least significant differences) test, there was no significant difference in the population of S. (S.) timorense and S. (N)
aureohirtum at each observation site population of S (G.) sundaicum and S. (Wallacellum)sp at T1 and T2 showed no significant difference (P>0.05); while at T3 and T4 showed real difference (P<0.05).
There were 2 adult fly species (52 flies) collected from livestock pens, i.e. S. (S.) timorense (71.5%), and S. (G.) sundaicum (28.8%). Blood-sucking activity of S. (S). timorense was begin at 18:00, while S. (G.) sundaicum at 20:00. The peak of blood-sucking activity of the two fly species was at 00:00-02:00.
© Hak Cipta Milik IPB, Tahun 2015
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains
pada
Program Studi Parasitologi dan Entomologi Kesehatan
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
BOGOR 2015
JULIANTY ALMET
RAGAM JENIS DAN FLUKTUASI POPULASI
SIMULIUM
PRAKATA
Puji syukur Kepada Tuhan Yang Maha Agung dan Mulia atas kasihnya sehinggga tesis dengan judul Ragam Jenis dan Fluktuasi Populasi Simulium di Kabupaten Kupang Nusa Tenggara Timur dapat disusun dan diselesaikan dengan baik.
Pada kesempatan ini penulis sampaikan ucapan terima kasih yang sangat besar kepada yang terhormat Ketua Komisi Pembimbing Prof Dr Upik Kesumawati Hadi, MS PhD, atas bimbingan arahan, motivasi, dan semangat yang diberikan kepada penulis selama pendidikan hingga penyelesaian studi. Ucapan terima kasih yang sebesar-besarnya juga penulis sampaikan Dr drh Susi Soviana, MSi atas segala bimbingan, arahan dan masukan hingga penyelesaian studi. Jasa dan kebaikan dari komisi pembimbing kepada penulis sungguh sangat berharga dan tidak akan terlupakan.
Terima kasih kepada Dekan Fakultas Kedokteran Hewan Universitas Nusa Cendana drh Max U. E. Sanam, MSc dan Pembantu Dekan Drh Diana Wuri MSi, yang telah memberikan kesempatan bagi penulis untuk mengikuti program magister dan telah mendukung hingga berlangsungnya penelitian hingga penyelesaian tesis ini. Terima kasih yang sebesar-besarnya buat Drh Nely Selan, MSc bersama para mahasiswa Ricky, Yopy, Umbu, Koly , Yeremylo, Joel yang telah membantu dalam proses pengambilan sampel hingga penelitian dapat terselesaikan.
Penulis mengucapkan terimakasih kepada rekan seperjuangan teman-teman Pascasarjana PS PEK dan Pasca Flobamora NTT atas kebersamaan dan dukungan dalam menempuh pendidikan Pascasarjana. Akhirnya, penulis sampaikan terima kasih yang tak terhingga kepada keluarga terkasih, Papa, Kakak, Adik, Suami tercinta dan anak terkasih, terima kasih banyak atas kasih sayang, pengorbanan dan kesabaran yang telah diberikan kepada penulis selama menyelesaikan studi.
Tesis ini masih banyak kekurangan, oleh karena itu kritik dan saran yang sifatnya membangun dan menyempurnakan tesis ini sangat penulis harapkan. Semoga tesis ini bermanfaat, khususnya di bidang Parasitologi dan Entomologi Kesehatan.
Bogor, Agustus 2015
DAFTAR ISI
DAFTAR TABEL vi
DAFTAR GAMBAR vi
DAFTAR LAMPIRAN vi
1 PENDAHULUAN 1
Latar Belakang 1
Tujuan Penelitian 2
2 TINJAUAN PUSTAKA 3
Jenis Simulium di Dunia 3 Jenis Simulium di Indonesia 3 Morfologi Simulium 4
Siklus Hidup 5
Peran Simulium sebagai Vektor 6
3 METODE PENELITIAN 7
Tempat dan Waktu Penelitian 7
Koleksi dan Identifikasi Simulium 7
Pengukuran Karakteristik Habitat larva Simulium 8
Koleksi Lalat Simulium Dewasa di Peternakan Sapi/Kandang Ternak 8
Analisis Data 8
4 HASIL DAN PEMBAHASAN 9
Ragam Jenis Simulium di Lokasi Penelitian 9 Deskripsi Morfologi Spesies Simulium 10
Kelimpahan Nisbi dalam Angka Dominasi Larva Simulium di Oenesu 16
Dinamika Populasi Larva Berdasarkan Ketinggian Tempat 17 Karateristik Habitat Simulium 18
Populasi Larva Simulium Dihubungkan dengan Indeks Curah Hujan 19 Kepadatan Lalat Simulium di Kandang Sapi 20
5 SIMPULAN 21
DAFTAR PUSTAKA 22
LAMPIRAN 25
DAFTAR TABEL
1 Ragam jenis larva Simulium di Oenusu (Kabupaten Kupang) dan Oehala
(Kabupaten Timor Tengah Selatan) 9
2 Morfologi larva, pupa, dan dewasa Simulium yang ditemukan di lokasi
penelitian 10
3 Kelimpahan nisbi dan angka dominasi spesies di lokasi Oenusu Maret-Mei
2014 16
4 Dinamika populasi larva Simulium berdasarkan ketinggian tempat dari 4x
penangkapan per bulan (Maret-Mei 2014) 17
5 Rata-rata populasi larva setiap spesies Simulium berdasarkan area pengamatan
pada ketinggian berbeda 17
6 Karakteristik habitat Simulium pada area pengamatan pada ketinggian berbeda 19
7 Rata-rata jumlah lalat yang tertangkap setiap 2 jam (lalat/2 jam/malam) di
kandang ternak 21
DAFTAR GAMBAR
1 Fluktuasi populasi larva Simulium dan indeks curah hujan selama Maret-Mei
2014 20
2 Jumlah individu lalat S. (S.) timorense dan S. (G.) sundaicum per dua jam
penangkapan 21
DAFTAR LAMPIRAN
1 Tabel indeks ragam jenis larva Simulium bulan Maret 2014 25 2 Tabel indeks ragam jenis larva Simulium bulan April 2014 25 3 Tabel indeks ragam jenis larva Simulium bulan Mei 2014 25 4 Tabel hasil uji korelasi spearman hubungan antara indeks curah hujan dengan
populasi larva Simulium 25
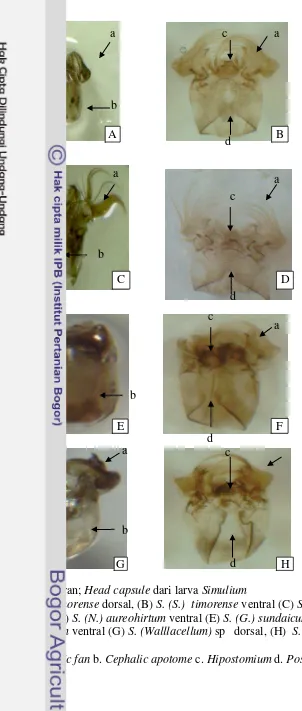
5 Gambar head capsule dari larva Simulium 26
1 PENDAHULUAN
Latar Belakang
Pengetahuan mengenai biologi serangga vektor penyakit seperti struktur anatomi, siklus hidup dan perilaku, serta hubungannya dengan patogen yang ditularkan, merupakan hal penting dalam pemahaman terhadap epidemiologi penyakit tular vektor dan keberhasilan pengendaliannya.
Simulium adalah sejenis lalat kecil (3-8mm), pengisap darah yang termasuk ke dalam Ordo Diptera, Subordo Nematocera, Famili Simuliidae. Simulium di Indonesia dikenal dengan istilah lalat punuk karena memiliki bagian toraks yang menonjol. Sebagian besar dari spesies ini menyerang hewan dan manusia melalui gigitannya. Disamping sebagai serangga pengganggu utama di beberapa negara di dunia, penyebarannya yang sangat luas menyebabkan lalat ini mudah ditemukan hampir di semua tempat yang memiliki aliran air, sungai kecil yang bersih sebagai habitat bagi stadium pradewasa (Hadi 1999).
Menurut Crosskey (1972,1981) terdapat 1270 spesies yang telah diketahui, dan dari 19 genera yang ada hanya empat genera yang berperan penting dari segi ekonomi yaitu Simulium, Austrosimulium, Prosimulium dan Cnephia.
Simulium merupakan genus terbanyak dan terpenting yang tersebar di seluruh wilayah zoogeografik. Crosskey (1981) mencatat sebanyak 43 jenis lalat
Simulium yang diketahui sebagai lalat pengganggu dan vektor penyakit pada manusia dan hewan domestik. Gangguan-gangguan utama akibat gigitannya, baik berupa iritasi dan perdarahan ptechie, terutama yang disebabkan karena serangan kelompok lalat ini pada musim semi.
Lalat hitam (Simuliidae) merupakan salah satu serangga yang penting secara medis karena semua anggota famili lalat ini merupakan vektor
Onchocerca volvulus, parasit kausatif dari onkoserkiasis manusia (“river
blindness”) di Afrika dan Amerika Latin, dan spesies-spesiesnya termasuk salah satu penyakit yang menyerang manusia, hewan, dan unggas yang terkenal di dunia. Di wilayah timur pedalaman Kanada, dan Selandia Baru Selatan, Simuliidae merupakan serangga yang paling berbahaya walaupun mereka tidak terlibat dalam penyebaran penyakit pada manusia.
Onchocerciasis merupakan infeksi mikrofilaria fatal yang dapat menyebabkan kebutaan dan penderitaan dalam jangka panjang, serta masalah sosio-ekonomis yang mengikutinya. Penyakit ini disebabkan oleh Onchocerca volvulus melalui gigitan spesies kompleks Simulium damnosum. Onchocerciasis
umumnya disebut sebagai penyakit Buta Sungai (river blindness desease) karena larva Simulium yang menyebarkannya hidup di wilayah sungai. Penyakit ini menyerang jutaan orang di dunia, terutama di Afrika selama berabad-abad dan karena itu penyakit ini dianggap sangat penting secara ekonomis. Kurang lebih 2 juta orang buta karena penyakit ini, dan sekitar 85.5 juta orang di Amerika Latin serta Timur Tengah hidup di wilayah endemik (Smith 1993).
Di antara unggas, lalat Simulium menyebarkan secara siklis protozoa darah
Leucocytozoon smithii, dan L. simondi, penyebab malaria kalkun dan parasit patogen pada bebek lokal di USA yang secara ekonomis merugikan. Simulium
Di antara mamalia, Simulium menularkan berbagai spesies cacing nematoda dari genus Dirofilaria (pada beruang), Mansonella (pada manusia), dan Onchocerca (pada mamalia ungulata dan manusia (Hadi 1996).
Kawanan besar lalat hitam seringkali menyerang manusia, serangan tersebut bisa sangat mengganggu. Pada individu yang sensitif, reaksi terhadap ludah lalat hitam yang diinjeksikan bisa menyebabkan sindrom yang disebut “demam lalat hitam”, yang diindikasikan dengan sakit kepala, demam, mual, dan seringkali radang saluran getah bening (Harwood & James, 1979). Ketersediaan sungai dan sungai yang beraliran deras merupakan faktor alami yang mendukung aktivitas perkawinan spesies Simulium di suatu area penelitian. Pengetahuan mengenai variasi spesies akan menambah formulasi strategi untuk mengontrol
onchocerciasis di suatu wilayah
Lalat Simulium diketahui mempunyai kebiasaan khas dalam memilih tempat untuk berkembang biak yaitu pada air bersih yang minim polusi dan berarus deras (Leveque 1989). Habitat ini sangat sesuai dengan kondisi air terjun Oenesu yang berada di Kecamatan Kupang Barat, Kabupaten Kupang. Oenesu merupakan salah satu obyek wisata yang cukup dikenal dan ramai dikunjungi. Selain sebagai air terjun yang alami, di wilayah Oenesu juga masih terdapat hutan alam di sekelilingnya. Di wilayah Oenesu terdapat ternak yang berpotensi sebagai sumber darah bagi lalat Simulium. Potensi alam air terjun Oenesu sebagai habitat Simulium terbukti dengan penemuan spesimen Simulium pada survei awal di lokasi ini..
Sejauh ini di Indonesia, informasi mengenai Simulium kurang tersedia. Arsip pertama mengenai monografi Simuliidae diterbitkan oleh Edwards (1934), akan tetapi informasi tersebut tidak lengkap. Hasil pemeriksaan ulang terhadap spesies Simulium yang telah diketahui di Jawa, ditemukan empat spesies baru yaitu Simulium sigiti, S. javaense, S. parahiyangum, S. upikae (Takaoka dan Davies 1996). Penelitian eksplorasi lalat Simulium umumnya dilakukan pada daerah dan musim yang terbatas, sehingga peluang ditemukannya spesies baru masih tinggi. Demikian pula halnya dengan potensi alam pada air terjun Oenesu sebagai habitat lalat Simulium dan minimnya informasi mengenai lalat ini di Indonesia, mendorong perlunya dilakukan penelitian tentang keragaman jenis dan fluktuasi kepadatan Simulium.
Tujuan
Penelitian ini bertujuan untuk: (1) mengidentifikasi jenis-jenis lalat
2 TINJAUAN PUSTAKA
Jenis Simulium di Dunia
Menurut Croskey (1987) distribusi geografis Simulium sp berdasarkan karakter morfologi dan kromosom di dunia terdiri atas 1554 spesies. Jenis ini tersebar di daerah Afrotropika sebanyak 194 spesies, di daerah Australasian 120 spesies, Nearktik 163 spesies, Neotropikal 355 spesies, Oriental 178 spesies dan Palaearktik 571 spesies. Jenis-jenis tersebut tergolong kedalam Parasimuliinae (Parasimulium dan Astonemyia), Simuliinae (Prosimuliini, Araucnephia,
Araucnephioides, Cnephia, Cnesia, Cnesiamima, Crozetia, Ectemnia, Gigantodax, Greniera, Gymnopais, Levitinia, Lutzsimulium, Mayacnephia, Metacnephia, Paraustrosimulium), Simuliini (Austrosimulium, Simulium).
Jenis Simulium di Indonesia
Di Indonesia, jumlah spesies Simulium sebanyak 76 spesies yang secara geografik tersebar di Jawa Barat (17 spesies ), Jawa Tengah (6 spesies), Jawa Timur (16 spesies), Bali (4 spesies), Sumatra Selatan (9 spesies), Sumatra Barat (6 spesies ) Sumatra Utara (5 spesies), Aceh (10 spesies), Sulawesi Utara (10 spesies), Sulawesi Selatan (8 spesies), Halmahera (5 spesies), Seram (13 spesies), Ambon (3 spesies), Papua (23 spesies), Flores dan Timor (8 spesies).Dari sejumlah spesies tersebut terdapat 4 jenis baru yaitu Simulium sigiti, S. javaense, S. parahiyangum, S. upikae (Takaoka dan Davies 1996).
Di Irian Jaya, Simulium tercatat ada 21 spesies di Jayapura, Manokwari, Sorong, Fak Fak, Wamena dari lembah Baliem, dan daerah sekitar Danau Anggi serta Pulau Biak. Fauna Simulium Irian Jaya sama pentingnya dengan yang ada di Papua Nugini. Di Sumatra terdapat 13 spesies yang tercatat, diantaranya S. (N.) feuerborni, S. (G.) gyorkosae S. (G.) parahiangum, S. (S.) nebulicola, S. (G.) friederichisi dan S. (S.) eximium yang juga ditemukan di Sumatra. Di Flores, ditemukan 8 spesies di Timor ditemukan satu spesies, S. (S.) nobile
(Takaoka, 1994).
Kajian mengenai bioekologi lalat hitam telah dilaksanakan di perkebunan teh di daerah Puncak Bogor, selama 10 bulan pengamatan kejadian musiman lalat, ditemukan 12 ragam spesies lalat hitam pada 3 ketinggian yang berbeda. Pada elevasi paling tinggi (1200 mdpl), jumlah spesies paling banyak ditemukan 11 spesies yaitu: S. (N.) feuerborni, S. (G.) sundaicum, S. (G.) gyorkosae, S. (G.) atratoides, S. (S.) argyrocinctum, S. (S.) eximium, S. (S.) upikae, S. (S.) sigiti, S. (S.) iridescens, S. (S.) nebulicola, dan S. (S.) celsum). Di sekitar sungai dan di perkebunan teh dengan ketinggian menengah (900-1200 mdpl), ditemukan 7 spesies seperti S. (N.) feuerborni, S. (G.) sundaicum, S. (G.) gyorkosae, S. (G.) atratoides, S. (S.) argyrocinctum, S. (S.) iridescens, dan S. (S.) nebulicola.
Morfologi Simulium
Dalam perkembangannya lalat Simuliidae mengalami metamorfosis sempurna, yang diawali dengan empat tahap perkembangan yaitu telur, larva, pupa dan dewasa.
Telur. Telur Simuliidae mempunyai bentuk agak ovoid, biasanya sub triangular bila dilihat dari beberapa sudut pandang, dan permukaanya terlihat cembung. Telur Simulium ini mempunyai panjang berkisar antara 0.1-0.5 mm (Crosskey, 1990). Telur Simulium dapat dikenali dari bentuknya yang ovoid atau elipsoidal, dengan panjang berkisar antara 0.15-0.39 mm (Hadi, 1996). Telur mampu bertahan kurang dari 2 hari (sungai di daerah tropis) dan maksimal 1 tahun pada musim kemarau yang panjang (Crosskey, 1990).
Larva. Menurut Crosskey (1990) bentuk tubuh larva Simulium memanjang seperti cacing, bagian abdomen ramping dan membesar diposterior. Pada tubuhnya mempunyai kutikula pada abdomen, terdapat dua pseudopods (proleg), setiap proleg mempunyai mahkota yang komplek (circlet) yang dibentuk oleh beberapa baris rangkaian kait paralel, dan memiliki toraks yang dilengkapi dengan atau tanpa sisik (scales).
Kepala horisontal (pragnathous), dibentuk oleh kapsul kepala (head capsule) yang kuat yang terdiri atas hipostomium, celah postgena (postgenal cleft), dan sepasang kipas kepala (caphalic fan) (kadang-kadang tidak ada). Bagian mulut terdiri atas sepasang mandibula dengan susunan gigi dan sikat yang kompleks, sepasang maksila dengan sikat dan batang pendek seperti palp, dan labio hipofaringeal (labio hypopharyngeal) yang kompleks dan meluas. Sepasang antena serta noktah mata (eye spot) terdapat pada tiap sisi kepala. Toraks pada instar matur mempunyai tanda yang jelas yang kelak berkembang menjadi insang pupa (pupal gill), kaki, sayap dan halter. Anus berbentuk huruf X, dan kisaran panjang tubuh larva dewasa adalah 3.5-12 mm.
Pupa. Panjang pupa Simuliidae kira-kira 2-7 mm. Kepala terletak di bawah toraks dan permukaanya horisontal, antena mempunyai segmen yang jelas. Pupa dapat diketahui dari bentuk cocoon dan sepasang spiracular gills pada toraks, bentuk kepala panjang dan pipih. Toraks cembung dibagian dorsal dan rata pada bagian ventral dengan sepasang spiracular gill di bagian anterodorsal dan biasanya mempunyai cabang. Abdomen terdiri atas 9 segmen, dan lengan (proleg) memiliki sepasang kait atau tuberkel Croskey (1990).
Siklus Hidup
Simulium termasuk serangga yang mengalami metamorfosis sempurna yaitu mulai dari telur, larva, pupa dan dewasa. Stadium larva terdapat beberapa tahap perkembangan yang disebut instar, yang jumlahnya berbeda pada tiap spesies.
Lalat betina dewasa meletakkan beberapa ratus butir telur diatas batu atau tumbuhan dibawah permukaan air pada aliran sungai kecil. Telur – telur akan menetas dalam waktu 4-12 hari, tergantung pada temperatur air (Soulsby, 1982) menjadi larva. Larva berbentuk silinder dan melekatkan diri dengan menggunakan posterior sucker yaitu organ yang menyerupai lengan dengan kait-kait kecil, yang dapat juga digunakan untuk berpindah tempat. Larva mengalami pergantian kulit (moulting) sebanyak 6 kali. Larva dewasa memintal cocoon dalam bentuk segitiga pada permukaan air, dan pada saat itu stadium pupa dimulai.
Menurut Crosskey (1990), telur Simulium menetas dalam waktu dua hari, sedangkan perkembangan larva memakan waktu empat hari atau beberapa bulan terutama pada musim dingin yang panjang. Perkembangan pupa terjadi selama dua hari atau maksimum dua atau tiga minggu, setelah itu menjadi lalat dewasa.
Faktor-faktor lingkungan utama yang mempengaruhi habitat tahap pradewasa Simulium adalah: (1) suhu air, (2) jenis arus, (3) tingkatan, (4) vegetasi, (5) karakter aliran air, (6) bahan makanan dan oksigen, (7) ukuran pupa
Simulium tergantung pada ketersediaan bahan makanan selama keberadaan larva. Ketika plankton kurang melimpah (misalnya selama kekeringan) pupa lebih kecil, (8) tidak ada hubungan langsung antara kecepatan arus dan derajat kejenuhan oksigen, dan dapat disimpulkan bahwa hubungan Simulium tidak ditentukan melalui faktor oksigen. Kondisi fisik substrat memiliki efek dalam membatasi distribusi larva. Akumulasi lumpur dan pertumbuhan berlebih oleh lumut memaksa migrasi larva, (9) sulit untuk menentukan kepadatan populasi secara akurat di sungai yang berbeda. Perbandingan perkiraan hanya dicapai oleh pengamatan di lokasi yang dipilih, (10) perkembangan larva di musim dingin dipercepat dengan kenaikan suhu dan tahap pupation terjadi saat musim dingin namun durasi tahap pupa sangat panjang selama suhu rendah.
Lalat memperoleh cacing pada tingkat larva pertama ketika menggigit hewan terinfeksi, dan parasit tersebut melewati usus, dan menembus dinding usus untuk bermigrasi ke otot terbang di toraks melalui hemosol abdomen dan menjadi larva tingkat tiga atau larva infektif. Larva tingkat ini kemudian pindah ke kepala lalat, siap untuk ditularkan ketika lalat menggigit inang lain beberapa hari kemudian. Beberapa spesies lalat hitam bertindak sebagai inang intermediet yang sangat kompatibel bagi parasit, dan kesempatan keberhasilan penularan antar sapi sangat tinggi karena hewan-hewan tersebut sering dikurung dalam peternakan yang besar yang kemudian memungkinkan bagi banyak lalat untuk menemukan darah dengan mudah kapanpun mereka butuh.
Peran Simulium sebagai Vektor
Crosskey (1981) mencatat sebanyak 43 spesies Simulium yang diketahui sebagai pengganggu atau sebagai vektor penyakit pada manusia dan hewan. Sekawanan S. columbaense di tepi sungai Danube pada tahun 1923 dilaporkan telah menyebabkan kematian 20.000 ekor hewan yaitu pada kuda, sapi, domba, kambing, babi, rusa, kancil, kelinci, dan hewan lainnya. Kematian ini diduga akibat toksin yang dikeluarkan oleh Simulium saat menghisap darah. Di Kanada pertengahan 1940, S. articum menyebabkan penurunan produksi susu pada sapi perah serta produksi daging, sehingga dilakukan upaya pengendalian lalat dengan menggunakan DDT. Penurunan produksi susu mungkin disebabkan oleh gigitan sekawanan Simulium yang menimbulkan terbentuknya gelembung atau papul- papul yang sangat mengganggu pada puting ambing (Soulsby, 1982). Berdasarkan Penelitian yang dilakukan oleh Hirai (1994) Simulium ochraceum
menjadi vektor utama Onchocerciasis manusia di Guatemala, di Thailand Utara teridentifikasi 2 jenis Simulium yaitu: S. nodosum dan S. asakoe di provinsi Chiang Mai di Thailand Utara yang beresiko menginfeksi manusia sepanjang tahun Ishii et al. ( 2013).
Di Amerika Utara S. venustum berperan sebagai penular Leucocytozoon
pada itik, kalkun dan unggas, sedangkan S. rugglesi diduga sebagai penular
Haemoproteus pada itik dan angsa (Soulsby, 1982). Di Kanada, S. ornatum
merupakan inang antara cacing filarial Onchocerca gutturosa pada sapi, sedangkan S. damnosum merupakan inang antara O. vulvulus pada manusia (Crosskey, 1973). Di Indonesia S. nobile dilaporkan menghisap darah sapi perah dikawasan usaha peternakan sapi perah Cibungbulan Bogor (Hadi, 1999).
Simulium damnosium sensulato adalah kelompok yang kompleks terdiri lebih dari 40 spesies lalat hitam yang menyebar di daerah sub-Sahara Afrika dan semenanjung Arab (Cupp, 1996). Simulium ini mengakibatkan 90% kasus
Onchocerciasis di seluruh dunia dan 95% kasus di Afrika (WHO, 2003). Sekitar 54 spesies dari Simulium dilaporkan telah menggigit manusia, menularkan
Onchocerciasis (Adler et al. 2004).
implikasi pada sosial ekonomi kesejahteraan manusia dan akibatnya memuncak pada penurunan produktivitas. Keberadaan sungai dan sungai yang beraliran deras merupakan faktor alami yang mendukung aktivitas perkawinan spesies Simulium
di area penelitian. Pengetahuan mengenai variasi spesies dan binomialnya serta informasi tentang kegiatan menggigit sangat penting dalam menambah formulasi strategi untuk merancang strategi pencegahan dalam mengendalikan
onchocerciasis di suatu wilayah.
Di beberapa wilayah Amerika Selatan, Mexico dan Afrika, lalat ini telah diketahui sebagai inang antara cacing Filaria dari Onchocerca volvulus, yakni cacing parasit yang menyebabkan Onchocerciasis (riverblindness) pada manusia (Harwood dan James 1979; Adler et al. (2004), Di India terdapat 52 spesies, lalat Simulium juga melimpah di negara bagian timur laut India dan dikenal sebagai hama serius yang menyebabkan gangguan menggigit dan reaksi alergi lokal pada manusia (Singh dan Tripathi, 2003). Pada hewan, Simulium
menularkan beberapa parasit seperti Leucocytozoon pada burung dan unggas, serta Onchocerca pada sapi perah (Chema dan Ivoghli 1978; Trees et al. 2000). Peran lalat ini juga menyebabkan gangguan (gatal, kebengkakan) akibat gigitannya (Butler dan Hogsette 1998).
3 METODE PENELITIAN
Tempat dan Waktu Penelitian
Pengambilan sampel lalat dan survei lapangan (pengukuran kepadatan dan pengamatan bioekologi Simulium) dilakukan di wilayah air terjun Oenesu (Kabupaten Kupang Kecamatan Kupang Barat) dan Oehala (Kabupaten Timor Tengah Selatan). Proses identifikasi Simulium dilakukan di Laboratorium Entomologi Kesehatan, Fakultas Kedokteran Hewan Institut Pertanian Bogor. Penelitian lapangan dilaksanakan pada Maret sampai Mei 2014.
Koleksi dan Identifikasi Simulium
Koleksi dilakukan pada 2 lokasi yaitu di Oenesu (Kabupaten Kupang) dan di Oehala (Kabupaten Timor Tengah Selatan). Simulium stadium pradewasa dikumpulkan mengikuti metode Takaoka (1983). Larva dan pupa Simulium
Pengukuran Karakteristik Habitat Larva Simulium
Observasi habitat Simulium dilakukan pada 4 area di Oenesu, berdasarkan perbedaan ketinggian yaitu T1 (1200 mdpl), T2 (1183 mdpl), T3 (1171 mdpl) dan T4 (1165 mdpl). Pengukuran karakteristik habitat berupa:
Suhu air. Pengukuran terhadap suhu air dilakukan menggunakan termometer, dengan cara memasukkan termometer dalam air sungai kemudian dicatat suhunya.
pH air. Pengukuran pH air dilakukan dengan menggunakan pH meter. Caranya adalah dengan memasukkan pH meter ke dalam air yang akan diukur pada kedalaman 5 cm dan secara otomatis alat tersebut bekerja mengukur sekitar 2 sampai 3 menit sampai angka digital stabil lalu dicatat.
Kekeruhan air. Pengukurang kekeruhan air dilakukan dengan menggunakan alat turbidimeter. Sampel air dimasukkan dalam wadah turbidimeter, dicatat angka yang muncul setelah simbol lampu padam.
Kedalaman air. Pengukuran kedalaman air menggunakan meteran.
Kecepatan arus. Pengukuran menggunakan pelampung dilakukan dengan cara pelampung dilepas dan dihitung waktunya menggunakan stopwatch. Selanjutnya tali diulur agar mudah terurai, dan dicatat waktunya saat tali pelampung terbentang lurus.
Keberadaaan naungan. Diamati dari ada tidak naungan tumbuhan/pohon di pinggir/badan air.
Pengukuran karakteristik habitat Simulium dilakukan dengan frekuensi seminggu sekali bersamaan dengan pengambilan sampel Simulium pradewasa.
Koleksi Lalat Simulium Dewasa di Peternakan Sapi/Kandang Sapi Koleksi lalat Simulium dewasa dilakukan di peternakan sapi dan kambing yang berada 1.5 km dari habitat perkembangan Simulium di Oenesu. Peternakan tersebut terdiri atas 35 ekor sapi dan 20 ekor kambing. Koleksi lalat dilakukan dengan menggunakan light trap (perangkap cahaya). Light trap ditempatkan di sekitar kandang dari pukul 18.00 sampai pukul 06.00 dengan frekuensi 2 kali dalam sebulan selama 3 bulan. Simulium yang tertangkap dikoleksi dengan cara mengganti perangkap kelambu secara berkala setiap 2 jam. Lalat Simulium yang terkumpul dimasukkan ke dalam alkohol 70%, diberi label kemudian dibawa ke laboratorium untuk diidentifikasi. Identifikasi Simulium dilakukan dengan menggunakan kunci identifikasi Takaoka et al. (2006).
Analisis Data
Data yang diperoleh dianalisis dengan menghitung kelimpahan nisbi, frekuensi dan dominasi spesies dan indeks keragaman spesies Shannon - Wiener (Odum 1993) dengan perhitungan:
Kelimpahan Nisbi = ∑ Individu spesies tertentu yang tertangkap ∑ Total induvidu seluruh spesies yang tertangkap Frekuensi Spesies = ∑ lalat spesies tertentu yang tertangkap setiap minggu ∑ minggu penangkapan
Dominasi Spesies = (Kelimpahan nisbi x Frekwensi spesies) Indeks Keragaman (H) = -∑ Pi ln Pi; dengan Pi = Ni/N
dimana,
Pi : perbandingan jumlah individu suatu jenis dengan keseluruhan jenis Ni : Jumlah individu ke-i
N : Jumlah total individu
Kriteria indeks keanekaragaman adalah:
Tinggi (H>3); Sedang (1≤H≤3); Rendah (H<1) (Krebs (1987)
Kepadatan populasi larva Simulium disajikan dalam bentuk tabel dan grafik. Indeks Curah Hujan (ICH) dihitung berdasarkan rumus matematika:
ICH= ∑ curah hujan (mm) perbulan x ∑ hari hujan per bulan ∑ hari (dalam satu bulan)
Hubungan indeks curah hujan, kepadatan populasi larva Simulium dianalisis dengan korelasi Spearman dan disajikan dalam bentuk tabel dan grafik.
4 HASIL DAN PEMBAHASAN
Ragam Jenis Simulium yang Ditemukan di Lokasi Penelitian Ragam jenis Simulium yang ditemukan di Oenesu dan Oehala selama penelitian ditunjukkan pada Tabel 1.
Tabel 1 Ragam jenis larva Simulium di Oenesu (Kabupaten Kupang) dan Oehala (Kabupaten Timor Tengah Selatan)
Jenis
Oenesu Oehala* Jumlah (%) Jumlah (%)
S. (S.) timorense 2235 (75.58) 41 (46,5)
S. (G.) sundaicum 575 (18.76) 38 (43.1)
S. (Wallacellum) sp 112 (3.71) 0 0
S. (N.) aureohirtum 49 (1.93) 9 (10.2)
* Koleksi di lokasi ini dilakukan hanya 1 kali
Berdasarkan Tabel 1 jenis larva Simulium di Oenesu dengan populasi terbanyak adalah S. (Simulium) timorense (75.58%), diikuti ole S. (Gompostilbia) Sundaicum (18.76%), S. (Wallacellum) sp (3.72%), S. (Nevermania) aureohirtum (1.93%). Sementara itu, terdapat 3 spesies larva
Deskripsi Morfologi Spesies Simulium
Deskripsi morfologi spesies Simulium yang ditemukan di Oenesu dan Oehala NTT secara ringkas disajikan pada Tabel 2.
Tabel 2 Morfologi larva, pupa dan dewasa Simulium yang ditemukan di lokasi Penelitian
S.(Wallacellum) sp S.(Nevermania) aureohirtum
Larva :
Panjang tubuh 3.7- 4.2 mm 4.5-5.0 mm 4.5-5.2 mm 3.4- 5.9mm
Warna tubuh abu-abu gelap
hingga coklat
coklat kehitaman coklat kemerahan Coklat
Cephalic apotome bintik kepala
Postgenal cleft postgenal sangat dalam, apeksnya
Jumlah filament 6 pasang filament 5 pasang filament 4 pasang 3 pasang
Bentuk kokon shoe shaped wall-pocket shaped no information slippers shaped
Dewasa:
Ukuran 2.0- 2.3 mm 2.0 – 2.2 mm no information 2.6- 2.8 mm
Warna coklat kehitaman coklat kehitaman no information coklat kehitaman
Antena 11 segmen 11 segmen no information 11 segmen warna
kuning pucat sampai
Sub genus Simulium Gomphostilbia Wallacellum Nevermania
dengan hasil koleksi di Oenesu sebanyak 12 kali. Keempat jenis Simulium
yang ditemukan di Oenesu masing-masing tergolong kedalam sub genus yang berbeda yaitu Simulium, Gompostilbia, Wallacellum dan Nevermania. Hal ini berbeda dengan yang ditemukan di Jawa, sub genus Wallacellum tidak ditemukan (Takaoka 1996).
Survey mengenai family Simuliidae di Flores telah dilakukan oleh Takaoka et al. (2006) menemukan 9 spesies yang terdiri dari 4 spesies baru, 4 telah dinamai dan 1 belum teridentifikasi. Seluruhnya termasuk ke dalam genus
Simulium letreille dan 4 sub genus yaitu, Gompostilbia enderlein (4 spp.), hingga coklat. Corak warna kapsul kepala bervariasi cephalic apotome bintik kepala tidak jelas (sebagian gelap), ada juga yang berwarna putih kekuningan dengan bintik-bintik kepala tidak jelas. Berdasarkan deskripsi morfologi Takaoka et al. (2006) antena terdiri atas 3 segmen dan sensillum apikal lebih panjang daripada batang kipas labral; proporsi panjang segmen pertama, kedua, dan ketiga adalah 1.0:1.1:0.6-0.7. Mandibula dengan gigi sisir yang ukurannya semakin mengecil dari gigi pertama hingga ketiga;; gigi besar di sebelah kanan mandibula di sisi apikal; supernumerary serration tidak ada. Hipostoma dengan 9 gigi apikal; gigi tengah hampir sama panjangnya dengan setiap gigi pojok, dan sedikit lebih panjang dari pada gigi intermediet di setiap sisi; garis lateral bergerigi di bagian apikal; 6 atau 7 bulu hipostomal per sisi.
Postgenal sangat dalam, apeksnya mencapai batas posterior hypostomium. Kutikula torakik telanjang. Insang pupa pharate dengan 6 filamen kurus. Kutikula abdominal telanjang kecuali di beberapa segmen posterior yang tertutup tipis oleh seta spatula dan seta kecil-kecil sederhana di permukaan dorsal dan dorsolateral, dan kedua sisi anal sclerite tertutup tipis oleh seta pendek tak berwarna. Sisik rectal tidak jelas. Organ rectal terdiri dari 3 lobus majemuk, masing-masing dengan 4-6 lobulus sekunder mirip jari. Anal sklerit berbentuk X dengan lengan anterior yang melebar dan panjangnya 0.7x panjang yang ada di posterior; 4-7 sensilla hanya ada di lengan posterior ke lengan posterior lainnya, tetapi tidak ada sensilla di basal juncture area; sclerite aksesori tidak ada. Segmen abdominal terakhir menggembung di bagian lateralnya tetapi tidak memiliki papila ventral. Posterior circlet dengan 88-96 baris hooklet dengan jumlah hooklet per baris hingga mencapai 18.
Pupa.
ventrolateral) di setiap sisi. Insang dengan 6 filamen kurus seperti benang yang berpasangan menjadi 3 pasang; pasangan filamen di bagian dorsal dan ventral mempunyai batang pendek dan pasangan di bagian tengah memiliki filamen yang langsung menempel atau dengan batang yang sangat pendek.
Semua filamen meruncing di di bagian apikal, panjang dan ketebalannya bertambah dari pasangan filamen dorsal menuju ventral: yaitu, filamen sebelah dalam (panjangnya sekitar 1.2 mm) dari pasangan filamen ventral merupakan filamen yang paling panjang, sedikit lebih panjang dari filamen sebelah luarnya (panjangnya sekitar 1.1 mm), filamen sebelah dalam (panjangnya sekitar 1.0 mm) dari pasangan filamen tengah sedikit lebih panjang dari filamen sebelah luarnya (panjangnya sekitar 0.9 mm), dan 2 filamen (panjangnya sekitar 0.8 mm) dari pasangan filamen dorsal panjangnya sama dan merupakan filamen yang paling pendek; jika dibandingkan secara basal, filamen sebelah dalam dari pasangan filamen ventral merupakan yang paling tebal, 2 kali tebalnya filamen sebelah luarnya, 2 filamen dari pasangan filamen tengah tebalnya sama satu sama lain, sedikit lebih tipis dari pada filamen sebelah luar dari pasangan filamen ventral namun sedikit lebih tebal dari pada filamen sebelah luar dari pasangan filamen dorsal yang mempunyai ketebalan 1.2 kali ketebalan filamen sebelah dalam dari pasangan filamen dorsal yang merupakan filamen paling tipis, yaitu sekitar 0.7 kali ketebalan filamen sebelah dalam yang paling tebal dari pasangan filamen ventral; cuticle filamen mempunyai banyak ridge dan alur melintang yang tepinya runcing di sebagian besar area namun kurang meruncing di bagian basal, tertutup lebat oleh tubercle kecil-kecil. Abdomen. Di bagian dorsal, segmen 1 hanya sedikit tersklerotisasi, kekuningan, halus, dengan 1 seta kurus sederhana yang panjang medium di setiap sisinya; segmen 2 transparan, halus, dengan 1 seta medium panjang dan 5 seta pendek sederhana berduri di setiap sisinya; segmen 3 dan 4 masing-masing dengan 4 duri berkait dan 2 seta kurus pendek sederhana di setiap sisinya; segmen 6 dan 7 tidak memiliki sisir duri; segmen 8 dengan sisir duri dan kelompok duri-duri kecil yang mirip sisir di baris transversal dan 2 seta kurus pendek sederhana di setiap sisinya; segmen 9 tidak memiliki sisir duri dan kait-kait terminal. Di bagian ventral, segmen 4 dengan 1 seta sederhana pendek berduri dan beberapa seta kurus pendek sederhana di setiap sisinya; segmen 5 dengan pasangan kait bercabang 2 yang berdekatan di bagian submedial di setiap sisinya; segmen 6 dan 7 masing-masing dilengkapi pasangan kait bercabang 2 atau kait sederhana di bagian dalam dan luar yang jaraknya berjauhan di setiap sisinya; segmen 4-8 masing-masing dengan kelompok duri kecil-kecil yang mirip sisir. Hooklets yang mirip jangkar tidak ada. Cocoon Berbentuk seperti sepatu, berwarna terang hingga coklat gelap, teranyam kuat, memanjang sedikit hingga sedang di bagian ventrolateral; lubang opening mengrah ke bagian atas dan depan, dengan garis tebal; benang individu tidak terlihat; panjang 2.5-3.2 mm dan lebar 1.0-1.4 mm.
Dewasa.
disinari, tertutup rambut-rambut kaku gelap dengan jumlah sedang kecuali di bagian paling atas dan bagian mediolongitudinal sempit dan tidak tertutup rambut (telanjang). Labrum 0.60-0.63 kali panjang klypeus. Antena terdiri dari 2+9 segmen, cokelat atau cokelat kekuningan dengan apikal 2 segmen flagellar kehitam-hitaman; flagellar segmen 1-7 seringkali kecoklatan di bagian dorsal, dengan bertambah gelap di bagian apikal, dengan 2 atau 3 segmen apikal yang kehitaman. Palp maksilar kecoklatan, terdiri dari 5 segmen, panjang proporsional segmen ketiga, empat, dan lima adalah 1.0:1.0:2.5; segmen ketiga (Gambar 11B) tidak membesar; vesikel sensori berukuran sedang, elipsoid, 0.25 kali panjang segmen ketiga, dengan lubang berukuran sedang di bagian apikal. Lasinia maksilar dengan gigi dalam 9-12 dan gigi luar 12. Cibarium dengan garis posterodorsal gelap yang sangat ter-sklerotisasi, dan dilengkapi dengan banyak tuberkel yang berkembang dengan baik.
Toraks. Skutum berwarna hitam kecoklatan sampai hitam, berkilau, pruinose keabuan pudar, tidak berpola, tertutup oleh rambut-rambut pendek coklat gelap yang posisinya rebah dengan intensitas sedang. Scutellum hitam kecoklatan, dengan rambut-rambut panjang gelap. Postnotum hitam kecoklatan, berkilau, berwarna keperakan ketika disinari, tanpa rambut. Membran pleural telanjang.
Tungkai. Tungkai depan: koksa dan throkanter kuning pucat; femur kuning gelap atau kuning kecoklatan, berangsur-angsur menjadi lebih gelap menuju ujung apikal; tibia hitam kecoklatan; dengan sebagian besar permukaan luarnya berkilau keputih-putihan; tarsus hitam kecoklatan, dengan sisir rambut dorsal berjumlah sedang, basitarsus sangat luas, 4.1 kali panjang dari lebar terbesarnya. Tungkai tengah: koksa hitam; trokhanter, femur, dan tibia hitam kecoklatan, tibia dengan sebagian permukaan posteriornya berkilau keputih-putihan ketika disinari; tarsus hampir putih kekuningan kecuali ujung apikal basitarsus, apikal ½ dari segmen kedua, dan segmen ketiga hingga kelima yang berwarna cokelat terang. Tungkai belakang: koksa hitam kecoklatan; trokanter putih kekuningan; femur hitam kecoklatan kecuali bagian paling dasarnya berwarna putih kekuningan; tibia hitam kecoklatan, dengan sebagian besar permukaan posteriornya berkilau keputih-putihan ketika disinari; tarsus berwarna keputih-keputih-putihan kecuali apikal 1/3 dari basitarsus, ujung apikal dari segmen kedua dan ketiga, serta segmen keempat dan kelima berwarna coklat sedang hingga gelap; basitarsus bersisian secara paralel, 5.7 kali lipat dari lebarnya, 0.65 dan 0.56 kali lipat dari lebar terbesar tibia dan femur belakang secara berturut-turut; kalsipala berkembang dengan sempurna, hampir sepanjang lebarnya dan 0.45 kali lipat dari lebar basitarsus; pedisulcus berkembang dengan sempurna. Cakar dengan gigi sub-basal kecil. Permukaan luar femora dan tibia dari tungkai tengah dan belakang sangat tertutupi rambut-rambut yang mirip sisik serta rambut-rambut sederhana biasa.
Sayap. Panjang 1.6-1.7 mm; kosta dengan spinula dan rambut yang gelap. Sub costa telanjang. Bagian basal dari urat R telanjang. R1 dengan spinula gelap serta beberapa rambut gelap; R2 hanya mempunyai rambut-rambut gelap. Sel basal tidak ada.
bintik-bintik dorsolateral berwarna keputih-putihan yang sangat terhubung satu sama lain di bagian tengahnya; tergit 6-8 besar dan mengkilap. Permukaan ventral segmen 7 tidak mempunyai pelat sternal.
Genitalia. Sterniteter-sklerotisasi dengan baik, telanjang di bagian tengah, dengan banyak rambut-rambut kaku panjang serta rambut-rambut ramping pendek secara lateral di masing-masing sisi; sternite 8 terlihat jelas dibuat secara posteromedial dan juga seolah olah secara ventral, membentuk lobus submedian segitiga dengan puncaknya membulat, masing-masing lobe tertutupi rambut-rambut kaku panjang dan pendek, terlipat secara dorsal di sepanjang garis-garis inner dan posterolateral serta mempunyai tonjolan telanjang hampir transparan yang sempit dan mengarah ke bagian belakang dengan tajam sehingga memanjang melewati ujung posterior lobe; garis inner lobus sedikit cembung, sangat terpisah satu sama lain. Ovipositor valva sangat tereduksi dan dapat terlihat karena area bermembran sempit yang terhubung ke masing-masing garis posterolateral dari sternite 8.
Percabangan genital berbentuk Y terbalik, dengan batang yang ter-sklerotisasi dengan baik; lengan-lengannya ramping, masing-masing dengan batas distal sangat tersklerotisasi yang mempunyai tonjolan pendek yang mengarah ke anterodorsal. Parapoct tersklerotisasi dengan baik, lebih banyak dihasilkan di ventroposterior sedikit di atas permukaan garis anterior cercus, dengan banyak rambut-rambut panjang di bagian ventral dan posterior; permukaan ventral sempit, tidak seperti pelat namun membulat; permukaan antero-medial dengan sensilla 5-9; parapoct di penampang lateral subtriangular, 0.74 kali dari panjang lebarnya. Cercus di penampang lateral pendek, hampir persegi atau membulat di bagian posterior, tertutup dengan beberapa rambut pendek. Spermateka berbentuk seperti telur, ter-sklerotisasi dengan baik kecuali saluran (duct) dan area besar dekat titik pertemuan (juncture) hingga ke saluran yang tidak tersklerotisasi, dengan pola-pola permukaan yang sulit dan tidak selalu dapat ditentukan serta seta internal kecil; kedua saluran aksesori hampir sama diameternya satu sama lain dan juga hampir sama dengan saluran utama berdasarkan deskripsi morfologi Takaoka et al. (2006).
S. (Gompostilbia) sundaicum Larva mature
Panjang tubuh 4.5-5.0 mm (rata-rata 4,5 mm). Cephalic apotome pucat atau agak gelap dengan bintik-bintik kecoklatan di kepala. Berdasarkan deskripsi morfologi Takaoka dan Davies (2006) sensilium apikal lebih panjang dari pada batang fan labral, panjang proporsional dari batang 3 segmen dari dasar ke ujung fan labral 1.00:0.78:0.76 dengan gigi sisir menurun dalam ukuran gigi ke 3. Gigi mandibula terdiri atas 1 besar dan 1 gigi kecil tanpa gigi supernuumerary. Sepanjang sudut gigi terdapat gigi median di setiap sisi,dan di bagian tengah lebih dari 3 disetiap sisi.
Setiap sisi terdapat gigi median dan gigi tengah lebih dari 3 disetiap sisi. Batas lateral: hypostomal setae 4 atau 5 sub parallel margin lateral. Post genal
S. (Wallacellum) sp Larva mature
Panjang tubuh 4.5- 5.2 mm (rata-rata 4.8 mm). Berdasarkan deskripsi morfologi Takaoka dan Davies (2006) bentuk badan normal, agak sedikit menggembung di segmen torak, runcing di segmen 1-4 abdominal (walaupun sedikit lebih besar di bagian posterior), menggembung dari segmen 5 hingga 6, meruncing menuju ujung posterior; warna badan keabuan, dengan pita transverse lebar berwarna coklat kemerahan gelap (tidak menyambung di bagian dorsomedial dan ventral), masing-masing di segmen 1-4 abdominal. Cephalic apotome kuning dengan bintik kepala jelas. Cervikal sklerit terdiri dari 2 bagian yang mirip batang, tidak bergabung di bagian occiput, terpisah satu sama lain di bagian medial. Antena terdiri dari 3 segmen, dan sensillum apikal lebih panjang daripada batang kipas labral; rasio panjang segmen pertama, kedua, dan ketiga adalah 1.9:2.1:1.
Mandibula dengan gigi sisir yang ukurannya semakin mengecil dari gigi pertama hingga ke 3; mandibular serration terdiri atas dari 2 gigi (1 besar, 1 kecil); gigi besar di sebelah kanan mandibula di sisi apikal; supernumerary serration tidak ada. Hipostoma dengan 9 gigi apikal salah satu baris; gigi tengah sedikit lebih panjang daripada setiap gigi pojok; gigi tengah dari 3 gigi intermedia di setiap sisinya merupakan yang paling pendek; garis lateral bergerigi; 8 bulu hipostomal per sisi, dengan posisi hampir paralel dengan garis lateral (atau sedikit menyimpang di bagian posterior dari garis lateral). Cekungan postgenal memiliki panjang medium, berbentuk lonceng menggantung, dengan perpanjangan anteromedian yang sempit. Kutikula torakik telanjang. Kutikula abdominal hampir telanjang, walaupun terdapat beberapa seta kecil-kecil sederhana tidak berwarna di permukaan dorsal masing-masing segmen, dan terdapat banyak seta sederhana tidak berwarna di setiap sisi anal sclerite segmen terakhir.
Terdapat l. organ rectal terdiri dari 3 lobus sederhana tanpa lobulus sekunder. Anal sclerite berbentuk X dengan lengan anterior yang melebar dan bercabang dua di bagian apikal ketika dilihat dari samping, dan panjangnya 0.7x posterior; basal juncture area di bagian posterior tengah memiliki sebagian kecil area yang tidak tersklerotisasi; 4 sensilla di basal juncture area dan 1 sensillum di belakang masing-masing lengan posterior; sclerite aksesori tidak ada. Segmen abdominal terakhir melebar di bagian ventrolateralnya sehingga membentuk tonjolan ventrolateral besar dan tonjolan ventral yang lebih kecil di setiap sisinya, tonjolan ventral dapat terlihat sebagai sebuah papila ventral kecil ketika dilihat dari samping. Posterior cirklet dengan 108 baris hooklet dengan jumlah hooklet per baris mencapai 18.
S. (Nevermania) aureohirtum Larva mature
tiga, 55 baris pada instar 4, 61 baris pada instar lima, 70 baris pada instar enam dan 81 baris pada instar tujuh.
Rata-rata panjang postgena larva S aureohirtum. adalah 0.26-0.45 mm, insang anus terbagi atas tiga lobus, mempunyai papila ventral dan rata- rata jumlah kait pada lingkaran anus adalah: 24 baris pada instar dua, 34 baris pada instar tiga, 55 baris pada instar 4, 61 baris pada instar lima, 70 baris pada instar enam dan 81 baris pada instar tujuh.
Pupa. Simulium aureohirtum mempunyai panjang sekitar 2.2-2.6 mm (rata-rata 2.6). Kepala dan toraks sama dengan S. taulingense. Organ insang terdiri atas 6 filamen dalam bentuk berpasangan serta mempunyai tangkai yang pendek. Seluruh filamen ini melebar secara divergent dan membentuk pita pada bagian distalnya. Pasangan ventral vilamen agak lebih panjang dan tebal daripada filamen lainnya dan panjangnya hampir sama dengan panjang tubuh pupa. Permukaan filamen mempunyai beberapa kerutan transversal yang membentuk pola retikula, dan ditutup rapat oleh tuberkel-tuberkel kecil.
Susunan rambut, duri (spina) dan kait pada permukaan bagian dorsal dan ventral abdomen menyerupai S. yushangense kecuali pada terga 5 dan 6 tidak mempunyai sisir duri. Kepompong berbentuk sederhana seperti selop, tersusun oleh anyaman yang rapat dan mempunyai penonjolan antero dorsal seperti pada
S. chowi, tetapi pinggir anterior tidak menebal Takaoka dan Davies (2006). Kelimpahan Nisbi dan Angka Dominasi Larva Simulium di Oenesu Kelimpahan nisbi dan angka dominasi larva Simulium yang terdapat di Oenesu menunjukkan hasil yang bervariasi sebagaimana terlihat pada Tabel 3. Tabel 3 Kelimpahan nisbi, dan angka dominasi spesies di lokasi Oenesu dari aureohirtum (1.44%). Indeks keragaman lalat Simulium di lokasi penelitian Oenesu menurut Krebs (1987) tergolong rendah yaitu pada kisaran H < 1, dengan indeks keragaman 0.72.
terjadi lebih tinggi. Hal ini berkaitan dengan populasi larva yang tinggi yang berati populasi lalat juga tinggi.
Dinamika Populasi Larva Berdasarkan Ketinggian Tempat
Dinamika populasi larva Simulium berdasarkan ketinggian tempat pada lokasi penelitian dan rata-rata Populasi Simulium disajikan pada Tabel 4 dan Tabel 5.
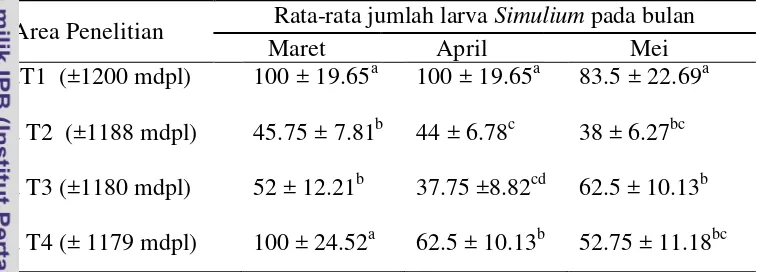
Tabel 4 Dinamika populasi larva Simulium berdasarkan ketinggian tempat dari 4x penangkapan/bulan (Maret-Mei 2014)
Area Penelitian Rata-rata jumlah larva Simulium pada bulan
Maret April Mei
Keterangan: Huruf superkrip yang berbeda pada kolom yang sama menunjukkan berbeda nyata (P < 0.05)
Keterangan: Huruf superkrip yang berbeda pada baris yang sama menunjukkan berbeda nyata (P < 0.05)
menunjukkan, rata-rata jumlah larva selama 3 bulan, pada area tertinggi T1 (± 1200mdpl) dan terendah T4 (±1165mdpl) terdapat jumlah populasi yang lebih tinggi dibandingkan dengan populasi Simulium pada ketinggian T2 (±1183mdpl), T3 (±1171mdpl). Populasi yang tinggi pada area T1 dan T4 dikarenakan adanya banyak pepohonan yang menjadi sumber naungan sehingga tersedia banyak dedaunan yang menjadi tempat perkembangbiakan larva
Simulium dan tidak adanya aktivitas mandi pengunjung. Pada area T2 dan area T3 populsai Simulium rendah dikarenakan kurang tersedianya naungan dan juga area ini sering digunakan sebagai tempat permandian bagi pengunjung, sehingga air sungai di sekitar area ini mengalami polusi yang menyebabkan larva
Simulium tidak bertahan hidup.
Hasil penelitian ini sesuai dengan yang dikemukakan Hadi (1999) yang melakukan penelitian ekologi Simulium di daerah perkebunan teh di daerah puncak Bogor yang melaporkan populasi lalat Simulium berbeda pada setiap bulan dan pada ketinggian yang berbeda. Demikian juga menurut Boakey (1993) yang melakukan penelitian Onchocerciasis dan distribusi komplek spesies
Simulium damnosum Wilayah Afrika Barat, berpendapat jumlah setiap spesies dipengaruhi oleh ketinggian air sungai. Populasi larva lalat Simulium
berdasarkan pembagian titik ketinggian di lokasi penelitiann Oenesu, secara keseluruhan bahwa ketinggian permukaan aliran sungai dan titik yang bebas polusi mengakibatkan jumlah populasi yang jauh lebih banyak.
Berdasarkan uji BNT (beda nyata terkecil) Tabel 4, menunjukkan dinamika populasi larva Simulium berdasarkan ketinggian tempat perbulan yaitu bulan Maret, April dan Mei berbeda signifikan (P<0.05). Pada area T1 selama 3 bulan populasi Simulium tidak berbeda nyata (P>0.05), akan tetapi pada area T2, T3, dan T4, yang masing-masing area perbulan berbeda signifikan (P<0.05). Rata-rata jumlah larva Simulium tertinggi terdapat pada area T1 dengan ketinggian ± 1200 mdpl, diikuti area T4 dengan ketinggian ± 1165 mdpl, area T3 dengan ketinngian ±1171 mdpl dan Area T2 dengan ketinggian ± 1183 mdpl.
Populasi larva setiap spesies berdasarkan area pengamatan Tabel 5, S. (S.) timorense memiliki populasi tertinggi pada area T1 dengan total 840 larva dan Area T4 689 larva Simulium jika dibandingkan dengan S. (G.) sundaicum, S. (Wallacellum) sp, S. (N.) aureohirtum. Berdasarkan uji BNT Populasi S. (S.) timorense dan S. (N) aureohirtum menunjukkan pada setiap area pengamatan tidak berbeda signifikan (P>0.05), sedangkan S. (G.) sundaicum dan S. (WallacelLum) sp pada area T1 dan T2 tidak berbeda signifikan tetapi pada area T3 dan T4 menunjukkan berbeda signifikan (P<0.05).
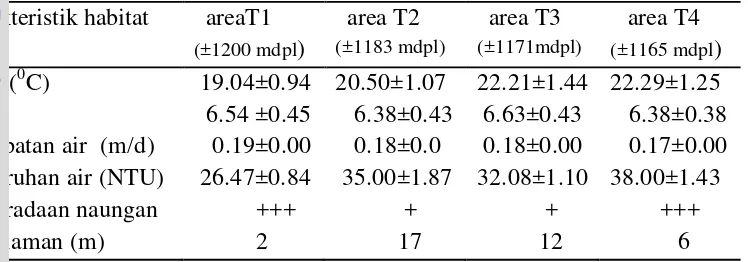
Karakteristik Habitat Simulium
Karakteristik habitat larva Simulium di Oenesu berdasarkan ketinggian tempat disajikan pada Tabel 6.
habitat larva Simulium damnosum yang optimum di Nigeria terjadi pada suhu 18-31oC.
Kecepatan aliran air pada lokasi penelitian rata-rata 0.18 m/detik dan semakin melambat kehilir. Sangat sedikit informasi mengenai pengaruh kecepatan air terhadap perkembangan pradewasa Simulium. Leveque (1989) melaporkan larva Simulium damnosum di Afrika Barat banyak ditemukan pada sungai yang beraliran deras dengan kecepatan 50 cm per detik. Nwobi dan Eneanya (2013) di Nigeria melaporkan larva Simulium lebih banyak ditemukan berkembang biak di perairan dengan kecepatan aliran 0.55-2.2 m/detik.
Kedalaman air masing – masing titik bervariasi yaitu pada area tertinggi (T1) 2 meter, (T2) 17 meter, (T3) 12 meter dan (T4) yang merupakan area terendah memiliki kedalaman air 6 meter. Larva Simulium hidup secara opivosit
pada tanaman yang berada di permukaan air atau pun di pinggiran sungai, sehingga faktor kedalaman air tidak mempengaruhi perkembangan larva
Simulium (Akpan et al. 2012). Hadi (1999) melaporkan pada ketinggian 900-1200 mdpl dengan suhu rata-rata 16-23oC, jumlah populasi larva Simulium
berbeda setiap tempat berdasarkan ketinggian dan suhu air. Boakey et al. (1993) melaporkan distribusi spesies kompleks Simulium damnosum di wilayah Afrika Barat, dipengaruhi oleh perubahan iklim yang mempengaruhi gerakan angin dan kedalaman air sungai.
Tempat perkembangbiakan lalat Simulium dipengaruhi oleh faktor fisika kimiawi seperti kekeruhan, pH dan oksigen terlarut. Kecepatan air serta temperatur lingkungan juga berperan dalam oviposisi lalat ini (Dhiman et al.
2014). Banyaknya rerumputan dan objek lain di pinggir sungai menyediakan habitat bagi Simulium, tempat larva menempel.
Tabel 6 Karakteristik habitat Simulium pada area pengamatan pada ketinggian berbeda
Keterangan : +++: banyak naungan.; + : sedikit naungan
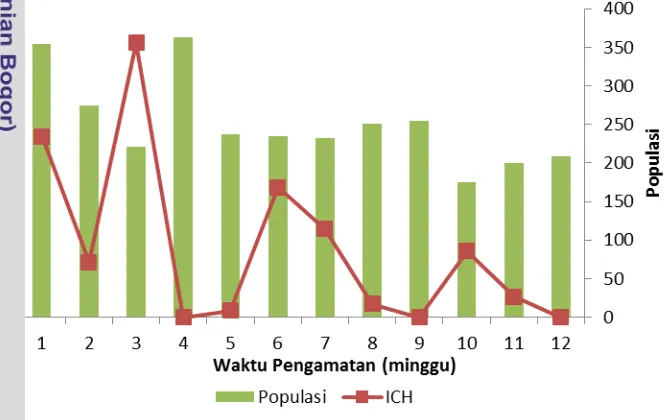
Populasi Larva Simulium Dihubungkan dengan Indeks Curah Hujan Hasil Uji Korelasi Spearman menunjukkan koefisien korelasi yang nyata (korelasi sedang) adalah antara indeks curah hujan dengan populasi larva
Simulium (r = 0.57). Persebaran bulanan dari titik penangkapan lalat menunjukkan bahwa kelimpahan S. (S.) timorense, S. (G.) sundaicum, S.
tinggi dan mulai mengalami penurunan pada bulan April dan diikuti bulan Mei, demikian juga populasi larva meningkat seiring dengan pola musim hujan dan disaat curah hujan berkurang populasi larva meningkat dikarenakan daun-daun yang menjadi habitat dari larva Simulium tidak terbawa arus. Dengan demikian dapat disimpulkan bahwa curah hujan yang tinggi mengakibatkan populasi larva Simulium semakin berkurang, sebaliknya semakin rendah curah hujan maka populasi larva Simulium semakin tinggi pula. Hal ini sesuai dengan hasil penelitian menurut Adeleke et al. (2011) yang meneliti tentang bioekologi dari kompleks Simulium damnosum di sepanjang sungai Osun Nigeria Barat Daya, melaporkan kelimpahan lalat meningkat pada saat curah hujan tinggi.
Secara keseluruhan, kebanyakan lalat Simulium tertangkap selama musim hujan (Maret sampai Mei). Penemuan ini sesuai dengan laporan yang telah dipublikasikan sebelumnya oleh Crosskey (1990), dan Opara et al. (2008) yang mempelajari tentang dinamika transmisi Simulium damnosum di Nigeria, serta Opuku (2006) yang meneliti tentang ekologi dan aktifitas menggigit Simulium di Ghana, kelimpahan lalat yang tinggi di musim hujan mungkin karena tingginya volume air dengan arus deras, sehingga dapat menyediakan lingkungan yang kondusif bagi berbagai tingkatan perkembangan lalat. Pada musim kemarau, tidak ada hujan menyebabkan sungai di lokasi penangkapan rusak menjadi kolam-kolam yang tidak dapat mendukung perkembangbiakan lalat hitam. Khususnya, ada pengurangan kelimpahan lalat Simulium antara bulan Juni sampai Agustus, dibuktikan dengan survei ulang pada bulan tersebut selama 2 jam pengambilan ditemukan 20 sampai 35 larva saja.
Gambar 1 Fluktuasi populasi larva Simulium dan indeks curah hujan selama Maret-Mei 2014
Dari 6 kali penangkapan (Tabel 7) sejak pukul 18.00 sampai dengan pukul 06.00 ditemukan 2 spesies lalat dengan jumlah 52 lalat yang terdiri atas 37 lalat
S. (S.) timorense dan 15 lalat S. (G.) sundaicum. Lalat S. (S.) timorense
menunjukkan aktivitas mengisap darah sudah dimulai sejak pukul 18.00 yang meningkat hingga pukul 02.00. Puncak aktivitas terjadi pada pukul 00.00-02.00. Aktivitas lalat S. (G.) sundaicum tidak berbeda dengan S. (S.) timorense, akan tetapi S. (G.) sundaicum menunjukkan aktivitas menggigit (Gambar 2) baru dimulai pada pukul 20.00. Namun demikian berdasarkan penelitian yang dilakukan oleh Dhiman et al. (2014) di India dan Akpan et al. (2012) di Nigeria, aktivitas menggigit lalat ini terjadi pada pagi hari dan meningkat di sore hari. Aktivitas menggigit lalat ini sejalan dengan aktivitas manusia sebagai inangnya.
Tabel 7 Rata-rata jumlah lalat yang tertangkap setiap 2 jam (lalat/2 jam/malam) di kandang ternak
Waktu Pengamatan S. (S.) timorense S. (G.) sundaicum
18.00 - 20.00 0.67 0.67
20.00 - 22.00 0.83 0.83
22.00 - 24.00 1.17 1.17
00.00 - 02.00 1.83 1.83
02.00 - 04.00 0.67 0.67
04.00 - 06.00 1.00 1.00
5 SIMPULAN
Ragam jenis Simulium yang teridentifikasi di lokasi penelitian Oeneseu, adalah S .(Simulium) timorense (75.59%), S. (Gompostilbia) sundaicum
(18.76%), S. (Wallacellum) sp (3.71%), S. (Nevermania) aureohirtum
(1.93%),dan di Oehala yaitu S. (Simulium) timorense (46.5%), S. (Gompostilbia) sundaicum (43.1%), S. (Nevermania) aureohirtum (10.2%). Indeks ragam jenis
Simulium pada bulan Maret 0.68%, April 0.74%, dan bulan Mei 0.72%. Berdasarkan ketinggian tempat pada area pengamatan tertinggi T1 (± 1200mdpl) dan area pengamatan terendah T4 (±1165mdpl) memiliki populasi terbanyak dibandingkan area pengamatan T2 (±1183mdpl) dan T3(±1171mdpl). Lalat Simulium dewasa di kandang ternak diperoleh 2 spesies lalat Simulium
yaitu S. (S.) timorense dan S. (G.) sundaicum, dan menunjukkan aktivitas tertinggi pada malam hari pukul 22.00-02.00.
DAFTAR PUSTAKA
Adler PH, Currie DC, Wood DM. 2004.The Black Flies (Simuliidae) of North America. New York (US): Cornell UniversityPr.
Akpan SS, Alaribe AA, Ejezie GC. 2012. The Distribution of Black Flies (Simulium species) in Ugbem and Ukwepeyiere Communities of Biase Local Government area of Cross River State, Nigeria. J. Dent Med Scien.
1(4):24-28.
Adeleke MA, Sam-woboas. 2010. Bioecology of Simulium damnosum Theobald Complex Along Osun River, Southwest Nigeria. J. Rural Trop Pub H. 10 : 39 - 43.
Atting IA, Braide EI, Ejezie GC, Opara KN, Udoidung NI. 2005. Factors affecting the transmission of human onchocerciasis by Simulium damnosum s.l. in a fringe savanna village in Cross River Sate, Nigeria. Mary Slessor J. Med. 5(2): 59-64.
Butler JF, Hogsette JA. 1998. Black Flies, Simulium sp. University of Florida (US): IFAS Extension.
Boakye DA, Back C, Fiasorgbor K. 1993. Sibling species distributions of the
Simulium damnosum complex in the West African Onchocerciasis Control Programme area during the decade 1984–93, following intensive larviciding since 1974.
Cheema AH, Ivochi B. 1978. Bovine Onchocerciasis Caused by Onchocerca urmillatu and O. gutturosa.Vet Pathol.15:495-505.
Crosskey RW. 1981. Simuliid Taxonomic-the Contemporary Scene. Dalam: Marshall Laird (ed), Blackfiles. Pp. 3-18. Academis press London.
Crosskey RW. 1972. Simuliidae. Insects and other arthropods of medical importance. The Trustees of the British Museum (Natural History), London.
Annotated World List (Eds.KimKC and Merritt RW), pp.425-520. Pennsylvania (US): Pennsylvania State University Pr.
Crosskey RW. 1990. The Natural History of Blackflies. British Museum (Natural History), London (EN): John Wiley and Sons.
Dhiman S, Hazarika S, Rabha B, Bhola RK, Singh VVL. 2014. Black fly (Simulium sp) composition, daytime biting activity and possible onchocerciasis infection in North-East, India. J. Env Biol. 26(2):64-68. Edwards FW. 1934. The Simuliidae (Diptera) of Java and Sumatra. Arch
Hydrobiol. 5:92-138.
Gallardo M.A, and J. Toja. (2002). Spatio-temporal distribution of simuliid (Diptera) and associated environmental factors in two Mediterranean basins of Southern Spain. Limnetica, 21, 47-57.
Hadi UK. 1996. Biological and Cytotaxonomical Studies of Simulium bidentatum, a Vector of Bovine Onchocerca and Related Blackfly species. PhD. Thesis.Oita Medical University, Japan 142p
Hadi UK. 1999. Taxonomical and ecological Studies Indonesian Blackflies (Diptera: Simuliidae). Final Report. URGE Project. The Young academic Program Batch II. Departemen of Parasitology and Pathology. Faculty of Veterinary Medicine. Bogor, Indonesia.
Hadi UK & Soviana S. 2010. Ektoparasit; Pengenalan, Diagnosa dan Pengendaliannya. Laboratorium Entomologi. FKH IPB.
Harwood RF, James MT. 1979. Entomology In Human And Animal Health, 7th edition. New York (US): Macmillan Publishing Company.
Hirohisa H, William S, Procunier J, Onofre Ochoa, K Uemoto. 1994.Cytogenetic analysis of the Simulium ochraceum species complex (Diptera: Simuliidae) in Central America.
Krebs CJ. 1987. Ecology The Experimental Analysis of Distribution and Abundance Third Edition. Harper and Row Publishers. New York. Leveque C. 1989. The Use of Insecticides In The Onchocerciasis Control
Programme (OCP) And Aquatic Monitoring In West Africa. Dalam: P. Bordeau, J. A. Haines, W. Klein & C. R. Krishna Murti (Eds.),
Ecotoxicology And Climate (1-20). Chichester (GB): John Wiley & Sons Limited.
Monsuro A. 2011. Bioecology of Simulium damnosum Theobald Complex Along Osun River, Southwest Nigeria. J Rural Trop Pub H. 10:39-43. Nwoke BEB. 1993. Onchocerciasis Control: A Training Guide In
Entomological Evaluation. Owerri (NG): Imo State University Pr.
Nwobi IK, Eneanya C. 2013. Ecology and Biting Activity of Simulium damnosum Complex in Nigeria Department of Parasitology and Entomology, Nnamdi Azikiwe University, Awka, Nigeria. 1:39-46 Opara, K.N., L.P. Usip and E.E. Akpabio. 2008. Transmission dynamics of
Simulium damnosum in rural communities of Akwa Ibom State, Nigeria.
J. V.Dis. 45:225-230
Opuku, AA 2006. The ecology and biting activity of blackflies (Simuliidae) and the prevalence of onchocerciasis in an agricultural community in Ghana. West Afr. J. App. Ecol., 9:1-7.