NON-INVASIVE HORMONE MONITORING: FAECAL
ANDROGEN AND GLUCOCORTICOID IN MALE CRESTED
MACAQUES (Macaca nigra) IN RELATION TO SEASONAL
AND SOCIAL FACTORS
GHOLIB
POSTGRADUATE SCHOOL
BOGOR AGRICULTURAL UNIVERSITY
BOGOR
STATEMENT OF THE ORIGINALITY OF THE THESIS AND
INFORMATION RESOURCES
Accordingly, I declare that all statement in the thesis with the title of Non-Invasive Hormone Monitoring: Faecal Androgen and Glucocorticoid in Male Crested Macaques (Macaca nigra) in Relation to Seasonal and Social Factors
are original and obtained from my own research results under supervision of the advisory committee. All data and information resources cited in the thesis are stated clearly and written in the list of references and these can be objectively assessed. This thesis is never proposed to take a degree at any other universities.
Bogor, May 2011
ABSTRACT
GHOLIB. Non-Invasive Hormone Monitoring: Faecal Androgen and Glucocorticoid in Male Crested Macaques (Macaca nigra) in Relation to Seasonal and Social Factors. Under direction of IMAN SUPRIATNA, MUHAMMAD AGIL, and ANTJE ENGELHARDT
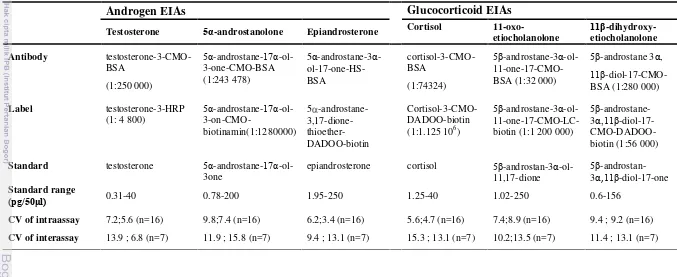
Non-invasive hormone monitoring using faeces has become an increasingly popular technique, but it is essential to validate before applying the technique to a new species. The aims of this study were to validate a method for measuring androgen and glucocorticoids noninvasively in feaces of crested macaques (Macaca nigra) and investigated the impact of season (climate) and social factors (copulation rates, male aggression, number of fertile females and dominance male) on male hormones. The study was carried out on two groups of crested macaques (Rambo 1 and Rambo 2) living in the Tangkoko-Batuangus Nature reserve, North Sulawesi, Indonesia. Three established enzymeimmunoassays (EIAs) for androgens (epiandrosterone, testosterone and
5α-androstanolone) and glucocorticoids (cortisol,11-oxo-CM and 11β -dihydroxy-CM) were tested using biological validation and immunoreactivity tests. Moreover, behavioural observations,measuring rainfall and temperature, and hormone measurements were performed. A General Linier Mix Model (GLMM) test was then conducted to investigate the effect of season and social factors on male hormones. The result shown that except 5α-androstanolone, both of androgen EIAs appeared reliable in their ability to discriminate androgen levels between juvenile and adult males, although HPLC data clarified that epiandrosterone is the most abundant. On the other hand, 11-oxo-CM and 11β -dihydroxy-CM assays discriminated better between pre-stress and stress periods than the cortisol. However, HPLC results shown that 11β-dihydroxy-CM detected the most abundant glucocorticoid metabolites. Result of GLMM test shown that androgen and glucocorticoid levels were not significantly influenced by rainfall and temperature. Androgen levels were significantly influenced by copulation rate (p=0.029) and dominance rank (p=0.002). Glucocorticoid levels were significantly influenced by number of fertile females (p=0.004) and dominance rank (p=0.02). In conclusion, biological validation can be used to validate androgen and glucocorticoid metabolites using faecal samples which epiandrosterone and 11β-dihydroxy-CM are the most realible and useful as a measure of androgen and glucocorticoid excretions respectively. Moreover, pattern of androgen and glucocorticoid levels in male crested macaques do not show a clear seasonal variation which levels of both hormones are not directly linked with enviromental factors (e.g., climate), but associated with social factors such as copulation rate, number of fertile females and dominance rank.
RINGKASAN
GHOLIB. Non-Invasive Hormone Monitoring: Faecal Androgen and Glucocorticoid in Male Crested Macaques (Macaca nigra) in Relation to Seasonal and Social Factors. Dibimbing oleh IMAN SUPRIATNA, MUHAMMAD AGIL, dan ANTJE ENGELHARDT.
Monitoring hormon secara non-invasiv menggunakan sampel feses merupakan teknik yang semakin populer. Hal ini didasarkan pada prinsip bahwa selain dari darah, hormon juga bisa diukur dari feses, urin dan saliva. Feses mengandung hormon steroid seperti progesteron, estrogen, androgen dan glukokortikoid dalam bentuk metabolit. Meskipun koleksi sampel feses relatif mudah, tetapi analisis hormon tidaklah mudah. Oleh karena itu, uji validasi untuk menentukan jenis metabolit hormon merupakan hal yang perlu dan penting untuk dilakukan, sehingga akan diperoleh hasil yang valid. Teknik non-invasiv ini memberikan kemudahan bagi kita untuk mempelajari fisiologi dari hewan liar atau hewan terancam punah (endangered) seperti monyet hitam Sulawesi (Macaca nigra) dari habitat alaminya. Monyet hitam Sulawesi adalah monyet endemik dari Sulawesi dengan status kritis (critically endangered animal). Namun, informasi tentang biologi reproduksi, fisiologi dan tingkah laku pada monyet ini masih sangat terbatas.
Tujuan penelitian ini adalah: 1) melakukan uji validasi biologis hormon androgen dan glukokortikoid, 2) mengkarakterisasi hormon androgen dan glukokortikoid pada monyet hitam Sulawesi jantan untuk menentukan ada tidaknya variasi musiman pada kadar hormon tersebut, 3) untuk mengetahui pengaruh musim (curah hujan dan suhu) dan faktor sosial (tingkat kopulasi, agresi antar jantan, jumlah betina subur dan dominasi jantan) terhadap kadar hormon androgen dan glukokortikoid pada monyet hitam Sulawesi jantan.
Hasil uji validasi hormon androgen menunjukkan bahwa jantan dewasa secara nyata mempunyai kadar immunoreaktif hormon androgen lebih tinggi pada epiandrosterone (p = 0,003) dan testosterone (p = 0,02) dibandingkan pada jantan anak, tetapi kadar 5α-androstanolone antara jantan dewasa dan jantan anak tidak menunjukkan perbedaan yang nyata (p = 0,2). Ini menunjukkan bahwa epiandrosterone dan testosterone mampu membedakan kadar hormon androgen antara jantan dewasa dan jantan anak, sedangkan 5α-androstanolone tidak dapat membedakan hal tersebut. Namun hasil analisis HPLC menunjukkan bahwa jumlah kadar immunoreaktiv pada epiandrosterone lebih besar dibandingkan kadar immunoreactiv pada testosterone. Hasil uji validasi hormon glukokorticoid menunjukkan bahwa kadar hormon glukokortikoid pada semua assai yang digunakan 2 kali lebih tinggi pada hari ke 2-3 setelah cekaman. Namun, pada assai 11-oxo-CM dan 11β-dihydroxy-CM, kadar hormon glukokortikoid selama stres menunjukkan perbedaan yang nyata lebih tinggi dibandingkan sebelum stres (11-oxo-CM: p = 0,004 dan 11β-dihydroxy-CM: p = 0,007), sedangkan pada cortisol tidak menunjukkan perbedaan (p = 0,274). Hasil analisis HPLC menunjukkan bahwa kadar immunoreaktiv pada 11β-dihydroxy-CM lebih tinggi dibandingkan kadar immunoreactiv pada 11-oxo-CM.
Hasil penelitian tentang pengaruh musim (iklim) dan faktor sosial terhadap kadar hormon androgen dan glukokortikoid menunjukkan bahwa tidak ada variasi musiman pada kadar hormon androgen dan glukokortikoid pada monyet hitam Sulawesi jantan. Hasil uji GLMM menunjukkan bahwa curah hujan tidak berpengaruh nyata terhadap kadar androgen (p = 0,675) dan glukokortikoid (p = 0,533). Kadar hormon androgen nyata dipengaruhi oleh tingkat kopulasi (p = 0,029) dan dominasi jantan (p = 0,002), tetapi agresi antar jantan dan jumlah betina subur tidak berpengaruh nyata terhadap kadar hormon androgen (agresi antar jantan: p = 0,834 dan jumlah betina subur: p = 0,600). Kadar hormon glukokortikoid nyata dipengaruhi oleh jumlah betina subur (p = 0,004) dan dominasi jantan (p = 0,02), tetapi tingkat kopulasi dan agresi antar jantan tidak berpengaruh nyata terhadap kadar hormon glukokortikoid (tingkat kopulasi: p = 0,078) dan agresi antar jantan: p = 0,144).
Dari hasil penelitian dapat disimpulkan bahwa uji validasi secara biologis dapat digunakan untuk validasi hormon androgen dan glukokortikoid, yang mana epiandrosterone dan 11β-dihydroxy-CM merupakan hormon metabolit yang tepat untuk pengukuran hormon androgen dan glukokortikoid pada monyet hitam Sulawesi jantan. Kadar hormon androgen dan glukokortikoid tidak secara langsung dipengaruhi oleh faktor lingkungan seperti iklim, tetapi lebih dipengaruhi oleh faktor sosial seperti tingkat kopulasi, jumlah betina subur dan dominasi jantan.
Copyright
©
2011 by Bogor Agricultural University (IPB)
All rights reserved
1. A part or the whole publication may not be cited without addressing the publisher
a. Citation is allowed for education, research, scientific publication, academic report or review of literature purpose only
b. Any citation do not loose the provisions of the copyright holder (IPB) 2. No part of this publication may be reproduced, stored in a retrieval system or
NON-INVASIVE HORMONE MONITORING: FAECAL
ANDROGEN AND GLUCOCORTICOID IN MALE CRESTED
MACAQUES (Macaca nigra) IN RELATION TO SEASONAL
AND SOCIAL FACTORS
GHOLIB
Thesis
as partial fulfillment requirement for the Master Degree at
The Sudy Program of Biology of Reproduction
POSTGRADUATE SCHOOL
BOGOR AGRICULTURAL UNIVERSITY
BOGOR
Title of Thesis : Non-Invasive Hormone Monitoring: Faecal Androgen and Glucocorticoid in Male Crested Macaques (Macaca nigra) in Relation to Seasonal and Social Factors
Name : Gholib
Registered Number : B352080031
Study Program : Biology of Reproduction
Approved by Advisory Committee
Prof. Dr. drh. Iman Supriatna Chairman
Date of examination: 18 March 2011 Date of graduation: 10 May 2011 Dr. drh. Muhammad Agil, M.Sc.Agr
Member
Dr. Antje Engelhardt Member
Head of Study Program of Biology of Reproduction
Dr. drh. M Agus Setiadi
Dean of Postgraduate School
PREFACE
The research is proposed due to the situation of crested macaque is extremely serious threatened. Moreover, their status has recently been categorized as critically endangered (IUCN Red List 2009), the population is decreasing rapidly due to illegal hunting (poaching) and habitat destruction. However, almost nothing is known about the reproductive biology, physiology and behavioural of this species in the wild.
This thesis will provide a knowledge about the non-invasive method for assessing physiological, in particular hormonal (endocrine) in male crested macaques and the importance of a careful validation in order to generate meaningful and accurate results. Moreover, information about the impact of season and social factors on androgen and glucocorticoid hormones in male crested macaques, a non-seasonally breeding primate will also presented. The information obtained from this study will help us to understand how physiology, behaviour and ecology interact, which could be benefit for the species and habitat management.
I do hope this thesis could produce valuable informations that could support the success of reproductive management and conservation programs of the crested macaques. Finally, I do hope this thesis is usefull for people who are interested to the non-invasive techniques for assessing hormonal (endocrine) and committed to the conservation of the crested macaques and other primate species.
CURRICULUM VITEA
Author was born in Gresik 21th of March 1982. Author is the younghest son of Mr. Ngatawi and Mrs. Sumri.
In 2001, the author was graduated from SMA Muhammadiyah 1 Gresik and accepted at Bogor Agricultural University through “Undangan Seleksi Masuk IPB (USMI) at the same year. Author was accepted at Study Program of Animal Production Technology, Faculty of Animal Science and graduated as Bachelor in Animal Science in 2005. In 2008, author continue to study at Master Program and registered at the study program of Reproductive Biology, Postgraduate School, Bogor Agricultural University.
Author has been worked as research assistant and camp manager in
ACKNOWLEDGEMENT
This research is a continuation of research program of Macaca nigra Project, as part of collaboration between German Primate Centre (DPZ) and Bogor Agricultural University which is led by Dr. Antje Engelhardt from DPZ. This research is generously supported mainly by Macaca nigra Project, Department Reproductive Biology, German Primate Centre. I would like to express my gratitude to the Department of Reproductive Biology, German Primate Centre for the financial support of my research and gave me an opportunity to conduct the hormone analysis in Hormone Labolatory, Department of Reproductive Biology, German Primate Centre.
I wish to express my earnest thanks and sincere appreciation to Prof. Dr. drh. Iman Supriatna, Dr. drh. Muhammad Agil, M.Sc.Agr, and Dr. Antje Engelhardt as Chairman and Member of advisory committee. I am deeply appreciate and respected them for their guidance, knowledge, encouragement, valuable suggestion, good advice, constructive critism and technical comments in preparation this thesis.
My especially heartiest gratitude and heartfelt appreciation to Prof. Dr. J. Keith Hodges, the head of Department of Reproduvtive Biology, German Primate Centre for his financial support during studying in Master Program at IPB. I thank the director of Tanoto Foundation for the scholarship during the second year of my study in IPB. I also acknowledge all team members of the Macaca Nigra Project (Brita R, Jacklyn R, Hani T, Heri, Julie D, Christof N, Dafne K, Giyarto, Teija F) at Tangkoko Nature Reserve for supporting data and sampel collections.
Domingus and Pak Yunus, for all permits provided for conducting research in the Tangkoko-Batuangus Nature Reserve.
Finally, I am grateful to my parents (Ibu Sumri and Bpk. Ngatawi) for their support during the entire of my studies, thanks to my foster parents in Bogor
CONTENTS
Page
LIST OF TABLES ... xvi
LIST OF FIGURES ... xvii
LIST OF APPENDIXS ... xviii
GENERAL INTRODUCTION ... 1
Background ... 1
Research Objective ... 4
Outcome of the Research ... 4
Framework of the Research ... 5
References... 7
LITERATURE RIVIEW... 9
The Crested Macaques (Macaca nigra)... 9
Taxonomy... 9
Morphology... 9
Biology and ecology of crested macaques... 10
Distribution, population and conservation status of crested macaques... 12
Biosynthesis of Steroid Hormones: Androgen and Glucocorticoids... 12
Metabolism and Excretion of Steroid Hormones: Androgen and Glucocorticoids... 16 Impact of Season and Social Factors to Testosterone and Cortisol Levels in Non-Human Primates... 20 Non-Invasive Method of Assessing Hormone... 22
References ... 24
BIOLOGICAL VALIDATION OF ENZYMEIMMUNOASSAYS FOR MEASUREMENT OF FAECAL ANDROGEN AND GLUCOCORTICOID METABOLITES IN WILD CRESTED MACAQUES... 29 Abstract... 29
Introduction... 29
Material and Methods... 31
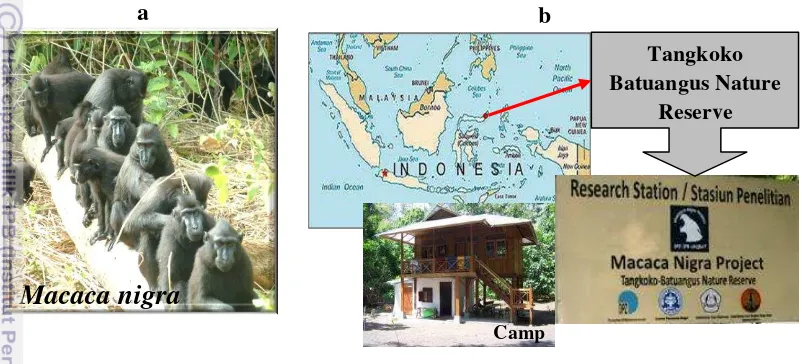
Animal and study site... 31
Sample collection... 32
Sample preparation... 33
Biological validation... 34
High-Performance Liquid Chromatography (HPLC) Analysis... 37
Hormone assay... 37
Statistical analysis... 38
Results... 39
Conclusion ... 45
References ... 46
ANDROGEN AND GLUCOCORTICOID LEVELS IN WILD MALE CRESTED MACAQUES IN RELATION TO SEASONAL AND SOCIAL FACTORS ... 49
Abstract ... 49
Introduction ... 49
Material and Methods ... 52
Study site, animal and period... 52
Enviromental seasonality... 53
Determination of the degree of male-male competition... 53
Behavioural observation and determination of male dominance rank... 53
Collection of faecal samples and hormone analysis... 55
Statistical analysis... 57
Results... 57
Discussion... 61
Conclusion... 65
References... 65
GENERAL DISCUSSION... 69
CONCLUSION AND SUGGESTION... 74
REFERENCES ... 75
LIST OF TABLES
Page 1 Major urinary and faecal metabolites of androgen and glucocorticoids
in non-human primates ...
18
2 Stress event from individuals which were used for glucocorticoid
validation test ... 33
3 Characteristics of the three different androgen and glucocorticoid EIAs, which were used to examine faecal androgen and glucocorticoid
metabolites ... 36
4 Demographic data, study males, group, dominance rank class, observation time, and number of faecal samples for hormone
analyses... 55 5 Results of GLMM with male identity and group as random factors
assessing the effect of different predictor variables on iEA and 11β
LIST OF FIGURES
Page
1 Framework of the research... 6
2 Crested Macaques (M. nigra): a) adult male (red skin colouration of scrotum), b) adult male and adult female (swelling) and, c) infant (white skin) and his mother... 10
3 Synthesis of testosterone in Leydig cells of the testis... 13
4 Synthesis of the various adrenal steroid hormones from cholesterol... 15
5 Scheme of the secretion, metabolism and excretion of glucocorticoids... 17
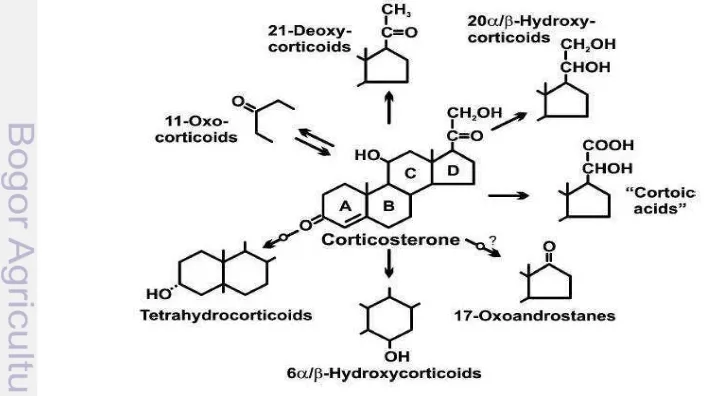
6 Possible pathways of corticosterone metabolism... 19
7 a) Wild crested macaques and b) field site and field station... 32
8 Parallelism test of 11β-dihydroxy-etiocholanolone assay... 35
9 Immunoreactive faecal epiandrosterone (iEA), testosterone (iT), and 5α-androstanolone levels in juvenile (N=5) and adult (N=5) males (mean and standard deviation) ... 39
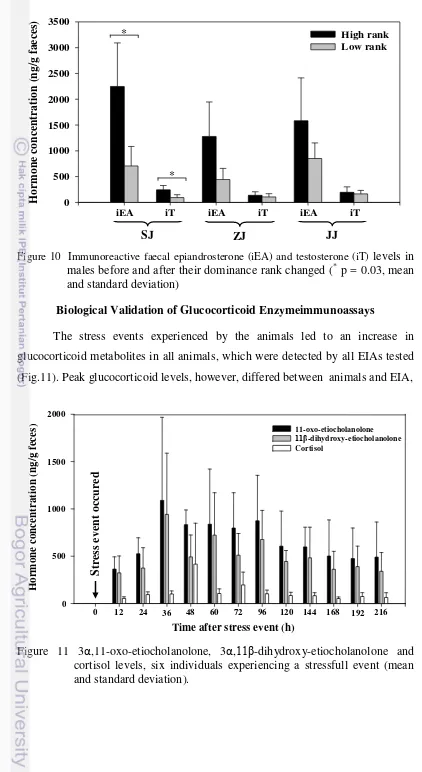
10 Immunoreactive faecal epiandrosterone (iEA) and testosterone (iT) levels in males before and after their dominance rank changed (* p = 0.03, mean and standard deviation) ... 40
11 3α,11-oxo-etiocholanolone, 3α,11β-dihydroxy-etiocholanolone and cortisol levels, six individuals experiencing a stressfull event (mean and standard deviation) ... 40
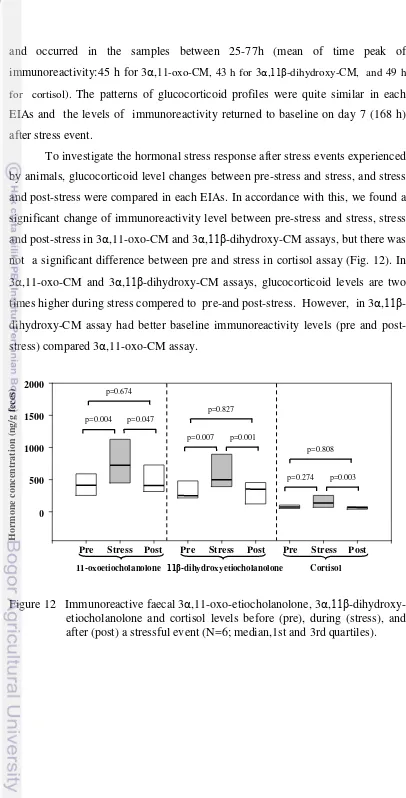
12 Immunoreactive faecal 3α,11-oxo-etiocholanolone, 3α, 11β -dihydroxy- etiocholanolone and cortisol levels before (pre), during (stress), and after (post) a stressful event (N=6; median,1st and 3rd quartiles) ... 41
13 HPLC profiles of immunoreactivity detected with the epiandrosterone, testosterone, 5α-androstanolone, 3α,11-oxo-etiocholanolone , 3α,11β-dihydroxy-etiocholanolone , and cortisol EIA in crested macaques (M. nigra). Associate elution positions of references standards: 1) cortisol (fractions 14–15), 2) corticosterone (22), 3) 3α,11β-dihydroxy-etiocholanolone (24), 4) 3α,11-oxo-etiocholanolone (29), 5) 5β-androstane-3,11,17-trione (36), 6) testosterone (43), 7) androstendione, dehydroepiandrosterone
(55-56), 8) epiandrosterone, 5b-DHT, 5b-androstane-3 β -ol-17-one (72), 9) 5β androstane-3a-ol-17-one (82), and 10) androsterone (100) (Heistermann et al. 2006) ... 42
15 Monthly variation on in a) androgen (epiandrosterone) and
glucocorticoid (11β-dihydroxy-CM) levels, b) rainfall and temperature, c) number of fertile females, and d) male copulation and
male aggression rates (mean and standard deviation)... 59 16 Dominance rank in relation to hormones levels. (a) Androgen levels
and (b) glucocorticoid levels. Box plots show the median (horizontal line), the 1st and 3rd quartiles (box) and the 10th and 90th percentiles
LIST OF APPENDIXS
Page 1 Permutation test of levels of11β-dihydroxy-CM before, during and
post stress (pre-, stress and post-stress)...
77
2 Permutation test of levels of 11-oxo-CM before, during and post stress (pre-, stress and post-stress)...
77
3 Permutation test of levels of cortisol, during and post stress (pre-, stress and post-stress)...
GENERAL INTRODUCTION
Background
In recent years, non-invasive methods for assessing physiological, in particular endocrine status have revolutionized almost in all areas of primatology, including behavioural ecology, reproductive biology, stress research, conservation and last but not least management of primates where the technology plays an integral role in assisting the husbandry, breeding and welfare of many species (Heistermann 2010). The non-invasive endocrine procedure gives several advantages over traditional invasive methods. Samples can be easily obtained without disturbing the study animals and do not put the animal in danger during capture (Möstl & Palme 2002; Wasser et al. 2000). The non-invasive techniques may also provide an accurate assessment of stress without the bias of capture-induced increases in glucocorticoids due to they do not disturb the animal (Harper & Austad 2001; Millspaugh et al. 2001; Touma et al. 2003).
Non-invasive approach is a more suitable option for field reseacher included primatologist. Using the technique, physiological data from free-ranging primates living in their natural habitat can be generated which has nowadays found widespread application in studies on relation between behaviour, ecology and conservation of many species (Hodges & Heistermann 2011). Therefore, non-invasive hormone monitoring has become an increasingly popular technique for studies of the physiology of free ranging animals and/or endangered species especially using faecal samples. However, although faecal sample collection is usually relatively easy, analysing hormones from faecal samples is not as easy as many people think, particularly not when dealing with a new species (Heistermann 2010). In this respect, the importance of a careful validation of each technique is essential in order to generate meaningful and accurate results (Heistermann et al. 2006, Tauma & Palme 2005; Heistermann 2010).
studies to the field has simultaneously permitted researchers to make direct comparisons between field and captive results, and broadened the scope of the research to include considerations of ecological variables such as seasonality, which affects diet and energetics, and reproduction, and may therefore mediate many types of hormone-behaviour interactions that are difficult to detect in captivity (Whitten et al. 1998; Gesquiere et al. 2010). Field studies can also provide a perspective on the relationships between social and ecological variables, and their interacting effects on hormones (Girard-Buttoz et al. 2009; Setchell et al. 2008; 2010).
It is a well-established fact that environmental/season (e.g., photoperiod, climate and availability of food and social factors (male-male aggression, sexual activity and number of receptive females) may influence the regulation of androgen and glucocorticoid hormones in males (Gesquiere et al. 2008;2010; Gould & Ziegler 2007). However, the response of behaviour and physiology to seasonal changes in environment differs between male primate species and depending on the degree of seasonality in breeding (Girard-Buttoz et al. 2009). Many studies on the impact of season and/or social factors on endocrine patterns have been already conducted in male seasonally breeding primates such as in rhesus macaques Macaca mulatta (Gordon et al. 1976), Japanese macaques M. fuscata (Barrett et al. 2002), tufted capuchin monkeys Cebus apella nigritus
(Lynch et al. 2002), golden lion tamarins Leontopithecus rosalia (Bales et al. 2006), ring-tailed lemurs Lemur catta (Gould & Ziegler 2007), Verreaux's sifakas Propithecus verreauxi (Fitchel et al. 2007), red fronted lemurs Eulemur fulvus rufus (Ostner et al. 2008), male Maddrills Mandrillus sphinx (Setchell et al. 2008; 2010), and assamese macaques M. assamensis (Ostner et al. 2011). The results show a clear raising in male androgen and/or glucocorticoid levels which one or both hormones increased during the mating season in association with hightened male-male aggression and sexual activity.
As comparison, the impact of season on male physiology and the
relationship between male behavioural patterns and endocrine status are less clear
in non-strictly seasonal/non-seasonal breeding primate species (Girard-Buttoz et
testosterone levels or testicular parameters such as testis size, seminiferous tubule
diameter, nuclear diameter of Sertoli cells and Leydig cells, and the cholesterol,
glycogen and sialic acid contents of testis (stumptailed macaque M. arctoides,
Nieuwenhuijsen et al. [1987]; Hanuman langur Presbytis entellus entellus, Lohiya et al. [1998]). However, information on seasonal endocrine patterns in primate
males of non-strictly seasonally/non-seasonally primates living in the wild under
natural ecological and social conditions is extremely limited. The only study
already carried out in the wild so far was carried out on long tailed macaques (M.
fascicularis) and showed that males of this non-strictly seasonally breeding primate species show endocrine patterns generally similar to those found in
strictly seasonal one (Girard-Buttoz et al. 2009). However, it is still unclear
whether this is generally true for all non-strictly seasonally/non-seasonally primates. Therefore, more studies are needed.
In order to obtain further our understanding on the impact of season and
Research Objective
The present research include 1) a biological validation study to examine the reliability of different androgen and glucocorticoid enzymeimmunoassays (EIAs) as an useful tool for measuring male gonadal status and adrenocortical activities in male crested macaques, and 2) a study to investigate the impact of season and social factors on male physiology and the relationship between male endocrine status and reproductive behaviour in male of a non-seasonally breeding primate, crested macaques.
The specific aim of biological validation study are 1) to examine the reliability of different androgen EIAs in discriminating gonadal status by comparing androgen levels from different reproductive stages (juvenile and adult males) and 2) to examine reliability of different glucocorticoid EIAs to the response of a stress event by comparing glucocorticoid levels before (pre), during (stress), and after (post) the stress event. The specific aim of study on the the impact of season and social factors on male physiology are 1) to characterize male endocrine status via faecal androgen and glucocorticoid levels throughout a complete annual reproductive cycle from free ranging males crested macaques to find out whether there is a seasonal variation in male endocrine patterns, and 2) to examine the relationship of the androgen and glucocorticoid levels on (a) climate, (b) levels of male-male aggression, (c) copulation rate, (d) number of fertile females and (e) dominance rank.
Outcome of the Research
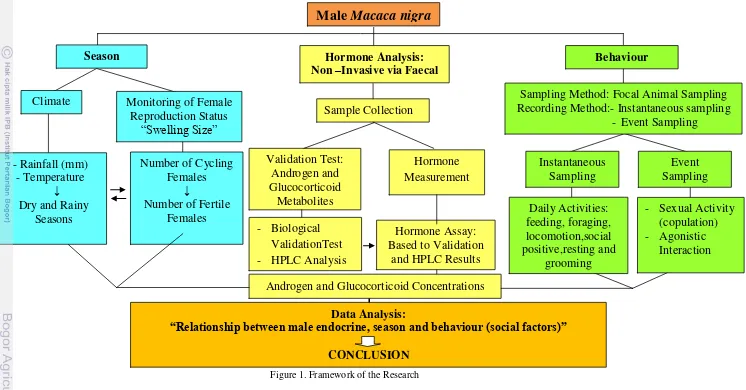
Framework of the Research
Environmental factors (such as photoperiod, climate and food) as well as social factors (such as male-male aggression, sexual activity and number of fertile females) are known to modulate androgen and glucocorticoid secretion in males. Many studies on the impact of season on endocrine patterns in male primates have been confirmed largely for seasonally breeding primate species. However, study on non-seasonally breeding primates is extremely limited and unclear.
Therefore, observation on season (climate), behaviour (male-male aggression and
sexual activity), number of fertile females and hormone measurements (androgen
and glucocorticoid) will be performed to investigate the impact of season on male
physiology and the relationship between male endocrine status and reproductive
behaviour in crested macaques. Since for crested macaques non of these
hormones thorough validation has taken place so far, first a biological validation test will be conducted to find out the realible and useful assays as a measure of androgen and glucocorticoid excretions in crested macaques. The framework of research as mentioned previously is detailed in Fig. 1 to clearly determine the
structure and step to study a relationship between seasonal and social factors with
Figure 1. Framework of the Research Data Analysis:
“Relationship between male endocrine, season and behaviour (social factors)”
CONCLUSION
Male Macaca nigra
Sample Collection
Hormone Analysis: Non –Invasive via Faecal
Hormone Assay: Based to Validation
and HPLC Results Hormone Measurement Validation Test: Androgen and Glucocorticoid Metabolites
Androgen and Glucocorticoid Concentrations
Sampling Method: Focal Animal Sampling Recording Method:- Instantaneous sampling
- Event Sampling
Behaviour
- Sexual Activity (copulation) - Agonistic Interaction Daily Activities: feeding, foraging, locomotion,social positive,resting and grooming Event Sampling Monitoring of Female
Reproduction Status
“Swelling Size”
Climate
Season
-Rainfall (mm) -Temperature ↓
Dry and Rainy Seasons
Number of Cycling Females
↓
Number of Fertile Females
Instantaneous Sampling
References
Bales KL, French JA, McWilliams J, Lake RA, Dietz JM. 2006. Effect of social status, age, and season on androgen and cortisol levels in wild male golden lion tamarins (Leontopithecus rosalia). Horm Behav 49:88-95.
Barret GM, Shimizu K, Bardi M, Asaba S, Mori A. 2002. Endocrine correlates of rank, reproduction, and female directed aggression in male Japanese macaques (Macaca fuscata). Horm Behav 42: 85-96.
Fichtel C, Kraus C, Ganswindt A, Heistermann M. 2007. Influence of reproductive season and rank on fecal glucocorticoid levels in free-ranging male Verreaux's sifakas (Propithecus verreauxi). Horm Behav 51:640–8. Gesquiere LR, Khan M, Shek L, Wango TL, Emmanuel O. Wango EO, Alberts
SC, Altmann J. 2008. Coping with a challenging environment: effects of seasonal variability and reproductive status on glucocorticoid concentrations of female baboons (Papio cynocephalus). Horm Behav 54:410–416.
Gesquiere LR, Onyango PO, Alberts SC, Altmann J. 2010. Endocrinology of year-round reproduction in a highly seasonal habitat: environmental variability in testosterone and glucocorticoids in Baboon males. Am J Phys Anthropol: 1-8.
Girard-Buttoz G, Heistermann M, Krummel S, Engelhardt A. 2009. Seasonal and social influences on fecal androgen and glucocorticoid excretion in wild male long tailed macaques (Macaca fascicularis). Physiol Behav 30:8-21. Gordon TP, Rose RM, Bernstein IS. 1976. Seasonal rhythm in plasma
testosterone levels in rhesus monkeys (Macaca mulatta): a 3 year study. Horm Behav 7:229-243.
Gould L, Ziegler TE. 2007. Variation in fecal testosterone levels, inter-male
aggression,dominance rank and age during mating and post-mating periods
in wild adult male ring-tailed Lemurs (Lemur catta). Am J Primatol
69:1325–1339.
Harper JM, Austad SN. 2001. Effect of capture and season on fecal glucocorticoid levels in deer mice (Peromyscus maniculatus) and red-backed voles (Clethrionomys gapperi). Gen Comp Endocrinol 123:337–344.
Heistermann M. 2010. Non-invasive monitoring of endocrine status in laboratory primates: methods, guidelines and applications. Adv Sci Res 1:1-9.
Heistermann M, Palme R, Ganswindt A. 2006. Comparison of different enzymeimmunoassays for assessment of andrenocortical activity in primates based on fecal analysis. Am J Primatol 68:257-273.
Hodges JK, Heistermann M. 2011. Field endocrinology: monitoring hormonal changes in free-ranging primates. In: Setchell JM, Curtis DJ (eds) Field and Laboratory Methods in Primatology: A Practical Guide. Cambridge: Cambridge University Press 353–370.
Lohiya NK, Sharma RS, Manivannan B, Kumar MA. 1998. Reproductive exocrine and endocrine profiles and their seasonality in male langur monkeys (Presbitis entellus entellus). J Med Primatol 27:15-20.
Lynch JW, Ziegler TE, Strier KB. 2002. Individual and seasonal variation in fecal testosterone and cortisol levels of wild male tufted capuchin monkeys (Cebus paella nigritus). Horm Behav 41:275-287.
Millspaugh JJ et al. 2001. Fecal glucocorticoidas says and the physiological stress response in elk. Wildl Soc Bul: 29(3):899-907
Möstl E, Palme R. 2002. Hormones as indicators of stress. Dom Anim Endocrinol 23: 67–74.
Nieuwenhuijsen K, Deneef KJ, Tenboschjjv, Slob AK. 1987. Testosterone, testis
size, seasonality, and behavior in group living stumptail macaques (Macaca
arctoides). Horm Behav 21:153–169.
Ostner J, Heistermann M, Schülke O. 2011. Male competition and its hormonal correlates in Assamese macaques (Macaca assamensis). Horm Behav 59:105-113.
Ostner J, Kappeler PM, Heistermann M. 2008. Androgen and glucocorticoid levels reflect seasonally occurring social challenges in male redfronted lemurs (Eulemur fulvus rufus). Behav Ecol Sociobiol 62:627–638.
Setchell JM, Smith T,Wickings EJ, Knapp LA. 2008. Social correlates of testosterone and ornamentation in male mandrills. Horm Behav 54:365-372. Setchell JM, Smith T,Wickings EJ, Knapp LA. 2010. Stress, social behaviour, and
secondary sexual traits in male primate. Horm Behav 58:720-728.
Touma C, Sachser N, Möstl E, Palme R. 2003. Effects of sex and time of day on metabolism and excretion of corticosterone in urine and feces of mice. Gen Comp Endocrinol 130: 267–278.
Touma C, Palme R. 2005. Measuring fecal glucocorticoid metabolites in mammals and birds: the importance of a biological validation. Ann NY Acad Sci 1046: 54–74.
Wasser SK et al. 2000. A Generalized fecal glucocorticoid assay for use in a diverse array of nondomestic mammalian and Avian Species. Gen Comp Endocrinol 120: 260–275
LITERATURE REVIEW
Crested Macaques
Taxonomy
The crested macaque (M. nigra) or yaki (local name) is one of the seven endemic primates of the Sulawesi Island. The sulawesi macaques are dare (M. maura), yaki (M. nigra), dihe (M. nigrescens), dige (M. hecki), bot (M. tonkeana), hada (M. ochreata), and endoke (M. brunnescens) (Fooden 1969, Albrecht 1978). The taxonomy of this species is: kingdom Animalia, phylum Chordata, class Mammalia,order Primates, family Cercopithecidae, Genus Macaca and species M. nigra (Lang 2006).
Morphology
Crested macaques have entirely black faces and bodies except for the striking bright pink colouration of their ischial callosities and white skin colour for infant (Lang 2006). The hair on their heads forms a long, backward, and upward-directed pointy crest (Rowe 1996; Groves 2001) (Fig. 2). Their tail is only about 2 cm short long (Supriatna & Wahyono 2000). Crested macaques are sexually dimorphic, with males almost twice the size of females [(male: body lenght 520 to 570 mm and weigh 9.9 kg on average (7-15 kg); female: body lenght 445 to 570 mm and weigh 5.5 kg on average (Rowe 1996)]. Sexual dimorphism in body size is significant in this species pointing to a high degree of male contest competition (Fooden 1969; Dixson 1977). In addition to a larger body size, male crested macaques have enlarged canine teeth compared to females. They use these in aggressive encounters with other males while competing for access to females and during intergroup encounters while defending food resources (Hadidian 1980).
Adult male and female crested macaques show visual and acoustic sexual signals. Males show a bright colouration of their scrotum together with a specific loud call. The scrotum‟s skin is brightly red and thus stands strikingly out of the
male‟s black fur. In addition, males utter a loud call that is species specific and
competitive interactions (Kinnaird & O‟Brien 1999). In addition to these male characteristics, females also show a big swelling size during active cycle and utter a female copulation call.
Figure 2 Crested macaques: a) adult male (red skin colouration of scrotum), b) adult male and adult female (swelling) and, c) infant (white skin) and his mother.
Biology and Ecology of Crested Macaques
Crested macaques live in multi-male multi-female social groups with group sizes of 20 till 70 individuals. The composition of the group includes infants, juveniles, subadult males/females, and adult males/females with an adult sex ratio 1:3.4 males:females (Supriatna & Wahyono 2000). The crested macaque is promiscuous, with both males and females mating multiple times with multiple
a
c
b
partners. Female receptivity is clearly indicated by tumescence (swelling) and redness of the anogenital region, which, in contrast to the black skin color, is particularly remarkable. The lenght of the menstrual cycle ranges between 23-45 days (mean=33.5) with mestruation lasting a mean of 4.6 days (Dixson 1977). Measurements of the diameter of sexual akin showed that the swelling undergoes rhythmical changes in size, being largest at mid cycle and flat during mestruation (Dixson 1977). Lang (2006) has stated that the gestation time is 174 days and female give birth usually to a single offspring. Young animals are nursed approximately one year and become fully mature in 3 to 4 years, females somewhat sooner than males. Life expectancy is estimated to be approximately 20 years.
A study of crested macaques in captive by Dixson (1977) between 1965 and 1973 revealed that there is no restricted birth season in this species. However, a study of crested macaques in the wild showed that female cycle not occur evenly distributed over the year, but culminated between September and December (Engelhardt, in prep). In accordance to female cycle activity, the majority of infants were born on February to May which almost 80 % births occurring within seven consecutive months (Dec-June) (Engelhardt,in prep).
Distribution, Population and Conservation Status of Crested Macaques
Wild crested macaques are found only on two islands in Indonesia. They are restricted to the tip of the northeastern-most peninsula of Sulawesi and on an island 345 miles from Sulawesi, Bacan island, where they were introduced by humans in 1867 (Rosenbaum et al. 1998). On Sulawesi, they are found in Tangkoko Batuangus-Duasudara, Gunung Ambang, and Gunung Manembonembo Nature Reserves as well as Bunaken Marine National Park (Supriatna & Wahyono 2000). The majority of the population is found in Tangkoko and it is here where most of the field research has been conducted.
The population of crested macaques has been reduced by at least 50% over the last ten years. MacKinnon and MacKinnon (1980) undertook extensive population surveys at the Tangkoko-Batuangus-Duasudara Nature Reserve in North Sulawesi during the late 1970s. They reported as high as 300 individuals/km2, the highest ever reported for an Asian primate. However, based on a field survey in 1987-1988, population densities of crested macaques in Tangkoko average about 76 individuals/km2, though in unprotected areas of Sulawesi they are significantly lower, with only about five individuals/km2 (Sugardjito et al. 1989). Futhermore, a survey by Rosenbaun et al. (1998) was reported that the population densities of this spesies decresed to 68 individuals/km2. Thus, this species is one of the most endangered Sulawesi macaques species. Therefore, since 2009 the International Union for Conservation of Nature (IUCN) has categorized this species as critically endangered (IUCN Red List 2009).
Biosynthesis of Steroid Hormones: Androgen and Glucocorticoids
(ACTH) with cortisol being the most important naturally occurring glucocorticoids (Senger 1999; Pineda 2003).
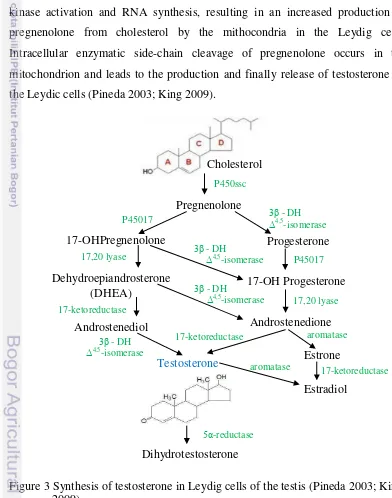
[image:32.595.104.496.246.744.2]Synthesis of testosterone is shown in Figure 3. The interstitial Leydig cells of the testis produce androgens, including testosterone, in response to LH (ICSH) stimulus and in synergy with FSH and prolactin (Pineda 2003). Prolactin regulates secretion of testosterone by Leydig cells by increasing the number and affinity of the receptors for LH on the Leydig cells. Futhermore, the interaction of LH with receptors in the Leydig cells activates the adenyl cyclase system, including protein kinase activation and RNA synthesis, resulting in an increased production of pregnenolone from cholesterol by the mithocondria in the Leydig cells. Intracellular enzymatic side-chain cleavage of pregnenolone occurs in the mitochondrion and leads to the production and finally release of testosterone by the Leydic cells (Pineda 2003; King 2009).
Figure 3 Synthesis of testosterone in Leydig cells of the testis (Pineda 2003; King, 2009).
Cholesterol
17-OHPregnenolone Progesterone
Pregnenolone
Androstenediol Dehydroepiandrosterone
(DHEA)
Androstenedione 17-OH Progesterone
P450ssc
P45017 3β Δ4,5- DH -isomerase
3β - DH
Δ4,5-isomerase P45017
17,20 lyase
3β - DH
Δ4,5-isomerase 17,20 lyase
17-ketoreductase
Testosterone
3β - DH
Δ4,5
-isomerase
17-ketoreductase
Estrone
aromatase
Estradiol
17-ketoreductase aromatase
Dihydrotestosterone
Testosterone is of particular interest when studying male hormone because the hormone has various function of morphological, physiological and behavioural processes in vertebrates which can influence reproductive success in males (Baum 2002; Pineda 2003; Ostner et al. 2002). Androgens are important in the control of sexual behaviour and spermatogenesis (Nelson 1995; Pineda 2003). Moreover, testosterone is play an important part in the formation of secondary sexual characters, such as coloration of sexual skin on face, rump and genitalia in Mandrillus sphinx (Wickings & Dixson 1992; Setchell et al. 2008). Furthermore, testosterone has been associated to the expression of aggressive behaviour and social status in longtailed macaques M. fascicularis (Girard-Buttoz et al. 2009), male Maddrills Mandrillus sphinx (Setchell et al. 2008), assamese macaques M. assamensis (Ostner et al. 2011). Thus, testosterone may influence the reproductive success of individual males by modulating their behaviour and physiology (Ostner et al. 2002).
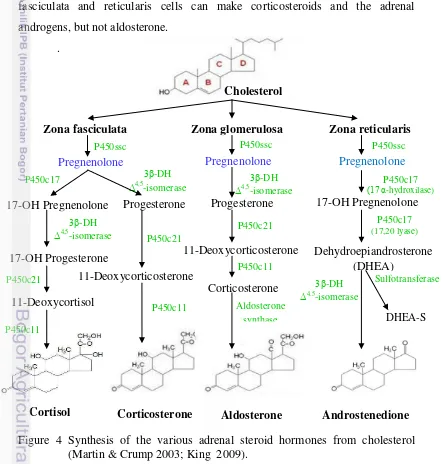
In addition to testosterone, glucocorticoids are produced by adrenal cortex which it also produces mineralcorticoids and androgens (King 2009; Martin & Crump 2003). Synthesis of glucocorticoid is shown in Figure 4 as described by Martin and Crump (2003) and King (2009) that many of the enzymes of adrenal steroid hormone synthesis are of the class called cytochrome P450 enzymes (CYP). First, cholesterol will be converted to pregnenolone in the adrenal cortex which it must be transported into the mitochondria where CYP11A1 resides. Furthermore, the conversion of pregnenolone to progesterone requires the two
enzyme activities of HSD3B2: the 3β-hydroxysteroid dehydrogenase and Δ4,5 -isomerase activities. Based on the statement by King (2009) that zona glomerulosa cells lack the P450c17 that converts pregnenolone and progesterone to their C17 hydroxylated analogs. Thus, the pathways to the glucocorticoids
glomerulosa is mainly responsible for the conversion of cholesterol to the weak mineralocorticoid, corticosterone and the principal mineralocorticoid, aldosterone. Cells of the zona fasciculata and zona reticularis lack aldosterone synthase (P450c18) that converts corticosterone to aldosterone, and thus these tissues produce only the weak mineralocorticoid corticosterone. However, both these zones do contain the P450c17 missing in zona glomerulosa and thus produce the major glucocorticoid, cortisol. Zona fasciculata and zona reticularis cells also contain P450c17, whose 17,20-lyase activity is responsible for producing the androgens, dehydroepiandosterone (DHEA) and androstenedione. Thus, fasciculata and reticularis cells can make corticosteroids and the adrenal androgens, but not aldosterone.
.
Figure 4 Synthesis of the various adrenal steroid hormones from cholesterol (Martin & Crump 2003; King 2009).
Zona fasciculata Zona glomerulosa Zona reticularis
P450ssc Pregnenolone
DHEA-S
11-Deoxycortisol
17-OH Progesterone Dehydroepiandrosterone
(DHEA) Progesterone 11-Deoxycorticosterone P450ssc Pregnenolone Pregnenolone 17-OH Pregnenolone
P450c17 3β-DH
Δ4,5
-isomerase
3β-DH
Δ4,5
-isomerase P450c21
P450c21 P450c11 P450c11 Progesterone 11-Deoxycorticosterone 3β-DH Δ4,5 -isomerase Corticosterone P450c21 P450c11 Aldosterone synthase 17-OH Pregnenolone P450ssc P450c17
(17 α-hydroxilase)
P450c17 (17,20 lyase) 3β-DH Δ4,5 -isomerase Sulfotransferase Cholesterol Corticosterone
[image:34.595.95.537.272.736.2]To improve the fitness of the individual during stress, various endocrine responses are involved which the main of those hormones are glucocorticoids (GCs) and catecholamines (Möstl & Palme 2002). Palme et al. (2005) stated that the most important and biologically relevant GCs are cortisol and corticosterone. These hormones are determined as a parameter of adrenal activity (Palme et al. 2005; Martin & Crump 2003). Glucocorticoid was synthesized and released from the adrenal cortex which are controlled by the pituitary adrenocorticotrophic hormone (ACTH) and regulated by the hypothalamic corticotrophin-releasing hormone (CRH) (Palme et al. 2005; Martin & Crump 2003). However, the processes regulating the secretion of GC are complex (Palme et al. 2005). Glucocorticoids improve fitness by energy mobilisation during short-term stress, (Raynaert et al. 1976) and may change behaviour (Korte et al. 1993). However, individual fitness may decrease when severe chronic stress (long periods of high cortisol concentrations) by immunosuppression and a trophy of tissues (Munck et al. 1984). Moreover, the reproductive success of the animal is decreased (Dobson & Smith 1995).
Metabolism and Excretion of Steroid Hormones: Androgen and Glucocorticoids
Steroid hormones are metabolized by many tissues including liver, kidney, muscle and blood (Schulster et al. 1976). They are also metabolized in the intestine where the caecum is probably the major site for steroid transformation, and most of the steroidal transformations by intestinal bacteria in the gut are reductive (Honour 1984). However, the major site of steroid metabolism is the liver.
role in the conversion of the metabolites (Möstl et al. 2005). Although, steroids in the gut are reabsorbed and transported to the liver (enterohepatic circulation) and the microbial flora metabolized the steroids, but the sterane skeletal structure is not degraded (Taylor 1971; Lindner 1972; Touma & Palme 2005). Thus, in the faecal the specific steroid metabolites can be then detected (Möstl&Palme 2002; Millspaugh & Washburn 2004). In addition, a similar pattern to that found in the plasma is reflected in the faecal when a lag time between hormonal events in the plasma and in the faecal is considered (Möstl&Palme 2002; Goymann 2005). Considering these points, it is possible to use faecal hormone metabolite analyses as a non-invasive tool to assess various endocrine functions in mammals and birds (Touma & Palme 2005).
Figure 5 Scheme of the secretion, metabolism and excretion of glucocorticoids (Möstl & Palme 2002).
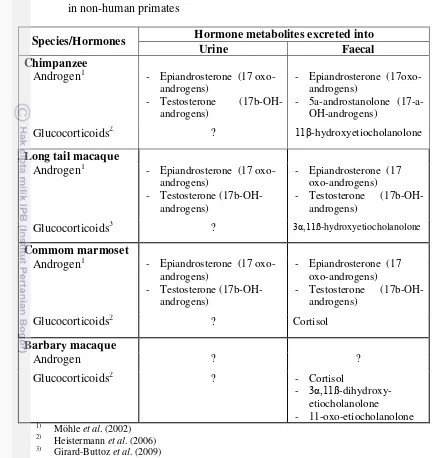
Table 1 Major urinary and faecal metabolites of androgen and glucocorticoids in non-human primates
Species/Hormones Hormone metabolites excreted into
Urine Faecal
Chimpanzee
Androgen1 - Epiandrosterone (17 oxo-androgens)
- Testosterone (17b-OH-androgens)
- Epiandrosterone (17oxo-androgens)
- 5a-androstanolone (17-a-OH-androgens)
Glucocorticoids2 ? 11β-hydroxyetiocholanolone
Long tail macaque
Androgen1 - Epiandrosterone (17 oxo-androgens)
- Testosterone (17b-OH-androgens)
- Epiandrosterone (17 oxo-androgens)
- Testosterone (17b-OH-androgens)
Glucocorticoids3 ? 3α,11ß-hydroxyetiocholanolone
Commom marmoset
Androgen1 - Epiandrosterone (17 oxo-androgens)
- Testosterone (17b-OH-androgens)
- Epiandrosterone (17 oxo-androgens)
- Testosterone (17b-OH-androgens)
Glucocorticoids2 ? Cortisol
Barbary macaque
Androgen ? ?
Glucocorticoids2 ? - Cortisol
- 3α,11ß -dihydroxy-etiocholanolone
- 11-oxo-etiocholanolone 1)
Möhle et al. (2002) 2)
Heistermann et al. (2006) 3)
Girard-Buttoz et al. (2009)
[image:37.595.95.528.100.558.2](Callithrix jacchus). They stated that T metabolites were excreted predominantly into the urine, a finding consistent with reports on the route of excretion of testosterone in other mammals (Palme et al. 1996). They mentioned that excretion of T metabolites into the faecal was much slower and more variable. On the other hand, the interval to peak excretion of T metabolites of 24–36 h obtained for the macaque and chimpanzee is similar to that reported for faecal steroid excretion in many other species (see Schwarzenberger et al. 1996, for review). The more rapid excretion of T metabolites into the faecal of the marmoset is consistent with the short gut passage time of 4–5 h (Möhle et al. 2002).
Concerning the metabolism and excretion of glucocorticoids, Lindner (1972) who was investigated the excretion of infused 14C-cortisol in the sheep had been reported that two-thirds of the radioactivity was subsequently found in the bile. The main metabolites were glucuronides of tetrahydrocortisol, tetrahydrocortisone and cortolones. In addition, C19O3 steroids (androstanes) were
also formed. Additionally, enterohepatic circulation of these metabolites occurred and cortisol metabolites were voided via the faecal. On the other hand, Möstl et al (2005) have been established a analytical validation of such non-invasive methods for assessing adrenocortical activity in droppings of birds. In general, the
metabolism of GCs include 5α or 5β reduction, hydroxylations, or reductions of
[image:38.595.107.462.520.718.2]functional groups or side-chain cleavage (C-17,20-lyase) in the case of 17α -hydroxylated metabolites (Fig. 6).
Impact of Season and Social Factors on Testosterone and Cortisol Levels in Non-Human Primates
In primates, the influence of seasonal as well as social factors on testosterone (T) and cortisol secretions have been investigated in a variety of species, although rarely in the wild. With recently non-invasive hormonal monitoring techniques (Hodges & Heistermann 2011; Whitten et al. 1998), primate field researchers are now able to examine relationships between T, aggression, mating behaviour, and social organization in naturally occurring populations of primates to examine if the Challenge Hypothesis (Wingfield et al. 1990) applies. The challenge hypothesis predicts a direct relationship between male T levels, mating systems, and reproductive strategies. In addition, they suggest that reproductive aggression related to an increase in T should be strongest during periods of social instability, including situations involving
challenges by conspecific males for access to estrous females. Thus, T levels should be positively related to the degree of inter-male aggression occurring over mating competition. The challenge hypothesis was initially applied to birds, and males in polygynous species were found to exhibit higher T concentrations for longer periods in the breeding season than did males in monogamous species
(Wingfield et al. 1990).
Patterns of testosterone excretion in adult males are highly variable in seasonally breeding polygynous primate groups in the wild. For examples, in male Japanese macaques, and red-fronted brown lemurs, faecal testosterone (fT) levels are higher either at the onset, or during mating periods than at other times, and in tufted capucihins male fT levels rise during the peak of female estrus synchrony
after the birth season (Barrett et al. 2002; Lynch et al. 2002; Ostner et al. 2002).
In Verreaux‟s sifaka, fT peaks 1 month before the breeding season and remains relatively constant for a 3-month period encompassing mating and male transfer (Brockman et al. 1998). Conversely, in male muriquis, only a slight increase in fT occurs at the onset of mating season, and no overall difference is found between mating and non-mating periods (Strier et al. 1999).
chimpanzees, and Verreaux‟s sifaka, dominant males exhibit higher T levels than
do lower-ranking individuals (Kraus et al. 1999; Sannen et al. 2003; Muehlenbein et al. 2004; Muller & Wrangham 2004a; Marshall & Hohmann 2005). In
Verreaux‟s sifaka, high-ranking males have been found to exhibit higher T levels during socially stable periods (Brockman et al. 1998). In contrast, in Japanese macaques, tufted capuchins, and red-fronted brown lemurs, no relationship has been found between male rank and fT levels (Barrett et al. 2002; Lynch et al. 2002; Ostner et al. 2002).
Social relationships may also factor into the regulation of the „„stress‟‟ hormone cortisol. Studies in several primates species have shown that the relationship between social and an individual's glucocorticoid output is not consistent. There was no difference in cortisol between dominant and subordinate
males in Wied‟s marmosets (Schaffner & French, 2004), golden lion tamarin (Bales et al. 2006), rhesus macaques (Bercovitch & Clarke 1995), cotton-top tamarins (Ginther et al. 2002), or tufted capuchins (Lynch et al. 2002), although male capuchins did display increases in cortisol during the time of highest female sexual activity. In contrast, a recent study of males sifakas (Propithecus verreauxi), dominant males had higher cortisol (Fichtel et al. 2007) suggests that elevated glucocorticoid concentrations in high-ranking males may reflect increased metabolic costs associated with dominant male reproductive strategy. Other studies in wild groups of chimpanzees (Muller &Wrangham 2004b), and female ring-tailed lemurs (Cavigelli, 1999) have found high-ranking animals to have higher glucocorticoid levels than low-ranking animals, suggesting that the benefits of dominance may be offset by certain costs.
Although many studies on the impact of season on endocrine patterns have been already conducted in seasonally breeding primates, study in non-seasonally breeding primates is extremely limited. A studies in captivity of stumptailed macaques M. arctoides (Nieuwenhuijsen et al. 1987), Hanuman langur Semnopithecus entellus (Lohiya et al. 1998) showed no seasonal variation in male testosterone levels or testicular parameters such as testis size, seminiferous tubule diameter, nuclear diameter of Sertoli cells and Leydig cells, and the cholesterol,
long tailed macaques (M. fascicularis) showed that male long tailed macaques exhibit a distinct annual variation in both androgen and glucocorticoid levels (Girard-Buttoz et al. 2009). Futhermore they stated that androgen level were significantly elevated during the conception period in association with elevated rates of male male aggression and copulatory activity, both strongly related to the number of cycling females in the group. In addition, neither glucocorticoid nor androgen levels were related to male dominance rank or to the environmental parameters investigated. Interestingly, levels of both hormones started to increase in the late birth season and thus 1-2 months prior to the mating season, suggesting that male long-tailed macaques go through pre breeding hormonal changes in preparation for prospective challenges.
Non-Invasive Methods for Assessing Hormone
Non-invasive methods, based on the measurement of hormone metabolites in urine and faecal are preferable, particularly as they have been shown to be as accurate and reliable as blood hormone analysis for monitoring the reproductive status in many animal species (Heistermann et al. 1995; Hodges & Heistermann 2002). Therefore, to date urinary and/or faecal hormone analysis is widely used in many species of both domestic and wildlife, particularly when working with
animals which are difficult to handle, potentially dangerous, and sensitive to
physical and social disruption (Heistermann 2010; Hodges & Heistermann 2011). These methods are now imperially used to investigate hormone-behaviour relationship, as well as questions in the fields of reproduction, animal welfare, ecology, conservation biology, and biomedicine (Schwarzenberger et al. 1996; Möstl & Palme 2002; Millspaugh & Washburn 2004; Palme et al. 2005; Touma & Palme 2005).
also provides the only feasible option for longitudinal studies in the wild and it is predominantly for this reason that interest in faecal assay methodologies has increased so markedly over the last 5-10 years (Hodges & Heistermann 2002).
On the basis of the diverse array of metabolites present in the faecal, it is advisable to apply group-specific immunoassays for their measurement (Schwarzenbergeret al. 1996, Möstl et al. 2005, Palme et al. 2005). Radioimmunoassays or enzyme immunoassays are most commonly used (Möstl et al. 2005). Because of several advantages, the latter are becoming more popular. However, Möstl et al. (2005) have been declared that all immunoassays must be validated analytically with regard to sensitivity, accuracy, precision, and specificity (cross-reactions) with the 5α5β-reduced metabolites present in the faecal. After the analytical validation of an immunoassay, a physiological validation must be conducted to demonstrate that an assay technique is capable of detecting changes in the levels of faecal steroid metabolites compared to respective changes of steroid concentrations in the blood (Palme 2005). In the case of GCs, a widely used method is an adrenocorticotrophic hormone (ACTH) challenge test (Goymann 2005, Touma & Palme 2005).
REFERENCES
Agil M. 2007. Reproductive biology of the Sumatran Rhinoceros Dicerorhinus sumatrensis (FISCHER 1814) [Dissertation]. Bogor: Study Program of Reproduction Biology, Postgraduate School, Bogor Agricultural University.
Albrecht GH. 1978. The Craniofacial Morphology of the Sulawesi Macaques: Multivariate Approach to Biological Problems. S Karger, Basel.
Bales KL, French JA, McWilliams J, Lake RA, Dietz JM. 2006. Effect of social status, age, and season on androgen and cortisol levels in wild male golden lion tamarins (Leontopithecus rosalia). Horm Behav 49:88-95.
Barret GM, Shimizu K, Bardi M, Asaba S, Mori A. 2002. Endocrine correlates of rank, reproduction, and female directed aggression in male Japanese macaques (Macaca fuscata). Horm Behav 42: 85-96.
Baum MJ. 2002. Neuroendocrinology of sexual behavior in the male. In: Becker JB, Breedlove SM, Crews D, McCarthy MM, editors. Behavior Endocrinology. Cambridge: The MIT Press. pp: 153-198.
Bercovitch FB, Clarke AS. 1995. Dominance rank, cortisol concentrations, and reproductive maturation in male rhesus macaques. Physiol Behav 58:215– 221.
Brockman DK, Whitten PL, Richard AF, Schneider A. 1998. Reproduction in free-ranging male Propithecus verreauxi: the hormonal correlates of mating and aggression. Am J Phys Anthropol 105: 137–151.
Cavigelli S. 1999. Behavioural patterns associated with faecal cortisol levels in free-ranging female ring-tailed lemurs, Lemur catta. Anim Behav 57:935– 944.
Dixson AF. 1977. Observation on the displays, menstrual cycles and sexual
behaviour of the “black ape” of Celebes (Macaca nigra). J Zool 182: 63-84.
Dobson H, Smith RF. 1995. Stress and reproduction in farm animals. J Reprod Fertil Suppl 49:451–61.
Fichtel C, Kraus C, Ganswindt A, Heistermann M. 2007. Influence of reproductive season and rank on fecal glucocorticoid levels in free-ranging male Verreaux's sifakas (Propithecus verreauxi). Horm Behav 51:640–8.
Fooden J. 1969. Taxonomy and Evaluation of the Monkeys of Celebes (Primates:Cercopithecidae). S Karger, Basel.
Girard-Buttoz C, Heistermann M, Krummel S, Engelhardt A. 2009. Seasonal and social influences on fecal androgen and glucocorticoid excretion in wild male long tailed macaques (Macaca fascicularis). Physiol Behav 30:8-21. Goymann W. 2005. Noninvasive monitoring of hormones in bird droppings:
physiological validation, sampling, extraction, sex differences, and the influence of diet on hormone metabolite levels. Ann NY Acad Sci 1046: 35–53.
Groves C. 2001. Primate Taxonomy. Washington: Smithsonian Institution Press. Hadidian J. 1980. Yawning in an old world monkey, Macaca nigra
(Primates:Cercopithecidae). Behav 75: 134-147.
Heistermann M. 2010. Non-invasive monitoring of endocrine status in laboratory primates: methods, guidelines and applications. Adv Sci Res 1:1-9. Heistermann M, Mostl E, Hodges JK. 1995. Non-invasive endocrine monitoring
of female reproductive status: methods and applications to captive breeding and conservation of exotic species. In: Ganslo er U, Hodges JK, Kaumanns W, Editors. Research and Captive Propagation. Filander Verlag: Erlangen. pp. 36-48.
Heistermann M, Palme R, Ganswindt A. 2006. Comparison of different enzymeimmunoassays for assessment of andrenocortical activity in primates based on fecal analysis. Am J Primatol 68:257-273.
Hodges JK, Heistermann M. 2002. Noninvasive assesment of reproductive fuction in primates. Evol Anthropol.
Hodges JK, Heistermann M. 2011. Field endocrinology: monitoring hormonal changes in free-ranging primates. In: Setchell JM, Curtis DJ (eds) Field and Laboratory Methods in Primatology: A Practical Guide. Cambridge: Cambridge University Press 353–370.
Honour JW. 1984. Billiary excretion and enterohepatic circulation. In: Makin HIJ, editor. Biochemistry of Steroid Hormones. 2nd Ed. London: Blackwell Scientific Publication. Oxford. pp. 382-393.
International Union for Conservation of Nature (IUCN). 2009. Crested Macaques (Macaca nigra). http://www.iucnredlist.org [2 January 2010].
King MW. 2009. Steroid hormones. The Medical Biochemistry. http://www.med.unibs.it/-marchesi/sterhorm.html [19 December 2009].
Kinnaird MF, O‟brien TG. 1999. A contextual analysisof the loud call of the
Sulawesi crested black macaque, Macaca nigra. Tropic Biodiver 6: 37-42. Korte SM, Bouws GAH, Bohus B. 1993. Central actions of
Kraus C, Heistermann M, Kappeler PM. 1999. Physiological suppression of sexual function of subordinate males: a subtle form of intrasexual competition among male sifakas (Propithecus verreauxi)?. Physiol Behav 66: 855–861.
Lang C. 2006 . Crested black macaque (Macaca nigra) Taxonomy, Morphology, & Ecology . http://pin.primate.wisc.edu [2009 20 December 2009].
Lindner HR. 1972. Enterohepatic circulation and patterns of urinary excretion of cortisol metabolites in the ewe. J Endocrinol 52: 19-20.
Lohiya NK, Sharma RS, Manivannan B, Kumar MA. 1998. Reproductive exocrine and endocrine profiles and their seasonality in male langur monkeys (Presbitis entellus entellus). J Med Primatol 27:15-20.
Lynch JW, Ziegler TE, Strier KB. 2002. Individual and seasonal variation in fecal testosterone and cortisol levels of wild male tufted capuchin monkeys (Cebus paella nigritus). Horm Behav 41:275-287.
MacKinnon J, MacKinnon K. 1980. Cagar Alam Gunung Tangkoko Duasudara, Sulawesi Utara Management Plan 1981–1986. Bogor, Indonesia.
Marshall AJ, Hohmann G. 2005. Urinary testosterone levels of wild male bonobos (Panpaniscus) in the Lomako Forest, Democratic Republic of Congo. Am J Primatol 65:87–92.
Martin PA, Crump MH. 2003. The adrenal gland. In: Pineda MH, Dooley MP, editors. Veterinary Endocrinology and Reproduction. 5th Ed. USA: Lowa State Press. pp 165-200.
Millspaugh JJ, Washburn BE. 2004. Use of fecal glucocorticoid metabolite measures in conservation biology research: considerations for application and interpretation. Gen Comp Endocrinol 138: 189–199.
Möhle U, Heistermann M, Palme R, Hodges JK. 2002. Characterization of urinary and fecal metabolites of testosterone and theirmeasurement for assessing gonadal endocrine function in male nonhuman primates. Gen Comp Endocr 129:135–145.
Möstl E, Palme R. 2002. Hormones as indicators of stress. Dom Anim Endocrinol 23: 67–74.
Möstl E, Rettenbacher ES, Palme R. 2005. Measurement of corticosterone
metabolites in birds‟ droppings: an analytical approach. Ann NY Acad Sci 1046: 17–34.
Muehlenbein MP, Watts DP, Whitten PL. 2004. Dominance rank and fecal testosterone levels in adult male chimpanzees (Pan troglodytes schweinfurthii) at Ngogo, Kibale National Park, Uganda. Am J Primatol 64:71–82
Muller MN, Wrangham RW. 2004a. Dominance, aggression and testosterone in
Muller MN, Wrangham RW. 2004b. Dominance, cortisol and stress in wild chimpanzees (Pan troglodytes schweinfurthii). Behav Ecol Sociobiol 55:332– 340.
Munck A, Guyre PM, Holbrook NI. 1984. Physiological functions of glucocorticoids in stress and their relationship to pharmacological actions. Endocr Rev 5:25–44.
Nieuwenhuijsen K, Deneef KJ, Tenboschjjv, Slob AK. 1987. Testosterone, testis size, seasonality, and behavior in group living stumptail macaques (Macaca arctoides). Horm Behav 21:153–169.
Nelson RJ. 1995. An Introduction to Behavioral Endocrinology. Sinauer Associates Inc., Sunderland, MA.
O'Brien TG, Kinnaird MF. 1997. Behavior, diet, and movements of the Sulawesi crested black macaque (Macaca nigra). Int J Primatol 18:321 351.
Ostner J, Heistermann M, Schülke O. 2011. Male competition and its hormonal correlates in Assamese macaques (Macaca assamensis). Horm Behav 59:105-113.
Ostner J, Kappeler PM, Heistermann M. 2002. Seasonal variation and social correlates of androgen excretion in male redfronted lemurs (Eulemur fulvus rufus). Behav Ecol Sociobiol 52:485–495.
Palme R. 2005. Measuring fecal steroids guidelines for practical application. Ann NY Acad Sci 1046: 75-80.
Palme R, Fischer P, Schildorfer H, Ismail MN. 1996. Excretion of infused, 14C-steroid hormones via faeces and urine in domestic livestock. Anim Reprod Sci 43: 43–63.
Palme R, Rettenbacher S, Touma C, El-Bahr SM, Möstl E. 2005. Stress hormones in mammals and birds: comparative aspects regarding metabolism, excretion, and noninvasive measurement in fecal samples. Ann NY Acad Sci 1040: 162–171.
Pineda MH. 2003. Male reproductive system. In: Pineda MH, Dooley MP, editors. Veterinary Endocrinology and Reproduction. 5th Ed. USA: Lowa State Press. pp 239-282.
Raynaert R, De Paepe M, Peeters G. 1976. Influence of stress, age and sex on serum growth hormone and free fatty acids in cattle. Horm Metab Res 8:109–14.
Rosenbaum B, O‟brien TG, Kinnaird M, Supriatna J. 1998. Population densities
of Sulawesi crested black macaques (Macaca nigra) on Bacan and Sulawesi: Effects of habitat disturbance and hunting. Am J Primatol 44:89-106.