INJEKSI
BOVINE SOMATOTROPIN
(bST) DAN
PENAMBAHAN KONSENTRAT PADA SAPI
HOLSTEIN LAKTASI DI DATARAN TINGGI CIKOLE,
LEMBANG
ELVIA HERNAWAN
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
ABSTRAK
ELVIA HERNAWAN. Injeksi Bovine Somatotropin (bST) dan Penambahan Konsentrat pada Sapi Holstein Laktasi di Dataran Tinggi Cikole, Lembang. Di bawah bimbingan WASMEN MANALU, SYAHRUN HAMDANI NASUTION, dan NASTITI KUSUMORINI
Sebanyak 24 ekor sapi Holstein laktasi yang dipelihara di dataran tinggi telah digunakan untuk mempelajari respons fisiologis terhadap injeksi somatotropin. Penelitian dirancang menggunakan rancangan acak kelompok dengan pola faktorial 3x2. Faktor pertama adalah injeksi somatotropin (bST) dengan 3 level, yang terdiri atas injeksi nonbST atau kontrol (plasebo yang diberi injeksi 1 mL sesame oil/ekor/hari), injeksi bST harian (36 mg/ekor/hari), dan injeksi bST selang 14 hari. (500 mg/ekor/14hari). Faktor ke dua adalah penambahan konsentrat dengan 2 level, yang terdiri atas pakan standar dan pakan standar ditambah 25% konsentrat. Parameter yang diukur meliputi denyut jantung, frekuensi pernapasan, suhu tubuh, nilai hematokrit, kadar hemoglobin, glukosa, trigliserida, nitrogen urea, kortisol, T4, T3 dalam darah, produksi susu, produksi 4% FCM (fat corrected milk), komposisi susu, bobot tubuh, pertambahan bobot tubuh, konsumsi bahan kering, efisiensi produksi susu dan nilai kondisi ternak, Injeksi somatotropin nyata meningkatkan denyut jantung dan frekuensi pernapasan demikian pula pemberian pakan standar ditambah 25% konsentrat nyata meningkatkan konsumsi bahan kering pakan. Terdapat interaksi antara injeksi bST dan penambahan 25% konsentrat pada produksi susu dan pertambahan bobot tubuh. Injeksi somatotropin pada sapi yang diberi pakan standar (sesuai dengan kebutuhan) dapat meningkatkan produksi susu sebesar 17-26%, sedangkan injeksi bST dan pakan yang ditambah 25% konsentrat cenderung menurunkan produksi susu, namun meningkatkan bobot tubuh, khususnya pada sapi yang mendapat injeksi bST selang 14 hari.
3
ABSTRACT
ELVIA HERNAWAN. Bovine Somatotropin Injection and Concentrate Supplementations in Lactating Holstein Cows in the Highland Cikole, Lembang. Under the supervision of WASMEN MANALU, SYAHRUN HAMDANI NASUTION, and NASTITI KUSUMORINI
Twenty four lactating Holstein cows were used to study the effect of bovine somatotropin injection and concentrate supplementations in lactating cows in the highland Cikole Lembang. The experimental cows were assigned into a Randomized Block Design with a 3 x 2 factorial arrangement. The first factor was somatotropin injection with three levels (nonbST injection, daily injection, and biweekly injection). The second factor was concentrate supplementation with two levels (fed standard ration and standard ration plus 25% concentrate.) Parameters measured were heart rate, respiration frequency, rectal temperature, hematocrit, hemoglobin, glucose, triglyceride, blood urea nitrogen, cortisol, thyroxine, triiodothyronine concentrations in the blood, weight gain, dry matter intake, milk production efficiency, and body condition score, milk production, 4% FCM (fat corrected milk), milk composition, weight gain, dry matter intake, milk production efficiency, and body condition score. Bovine Somatotropin injection significantly increased heart rate and respiration rate. Concentrate supplementation increased dry matters intake. There were an interaction between bST and concentrate supplementation on milk production and weight gain. Bovine somatotropin injection in cows fed standart ration increased milk production by 17–26%, but injection in cows fed ration with 25% concentrate decreased milk production but showed increase weight gain. Daily and biweekly somatotropin supplementation did not significantly affect milk production.
PERNYATAAN MENGENAI DISERTASI
Saya menyatakan dengan sebenar-benarnya bahwa disertasi saya dengan
judul Injeksi Bovine Somatotropin (bST) dan Penambahan Konsentrat pada Sapi
Holstein Laktasi di Dataran Tinggi Cikole, Lembang adalah benar-benar asli
karya saya sendiri dengan arahan komisi pembimbing, dan bukan hasil jiplakan
atau tiruan dari tulisan siapa pun serta belum diajukan dalam bentuk apa pun
kepada perguruan tinggi mana pun.
Bogor, Mei 2007
Elvia Hernawan
5
INJEKSI
BOVINE SOMATOTROPIN
(bST) DAN
PENAMBAHAN KONSENTRAT PADA SAPI
HOLSTEIN LAKTASI DI DATARAN TINGGI
CIKOLE, LEMBANG
ELVIA HERNAWAN
Disertasi
Sebagai salah satu syarat untuk memperoleh gelar Doktor pada
Program Studi Biologi
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
7
PENGUJI LUAR KOMISI
I. Ujian Tertutup tanggal 24 April 2007
Dr. Ir. Bagus Pryo Purwanto
Staf Pengajar Departemen Ilmu Produksi Ternak, Fakultas Peternakan, Institut
Pertanian Bogor.
II. Ujian Terbuka tanggal 28 Mei 2007
1. Dr. Ir. Chalid Talib MS
Peneliti dari Pusat Penelitian dan Pengembangan Peternakan , Departemen
Pertanian.
2. Dr. Ir. Suryahadi DEA
Staf Pengajar Departemen Nutrisi dan Makanan Ternak, Institut Pertanian
RIWAYAT HIDUP
Penulis dilahirkan di Bandung pada tanggal 25 Desember 1951 sebagai
anak ke empat dari 12 bersaudara pasangan R. H. Ma’mun Hernawan dan R. Hj.
Siti Halimah Zyin.
Pendidikan SD diselesaikan pada tahun 1963 di SD Palasari Bandung,
SMP diselesaikan pada tahun 1966 di SMPN V Bandung, dan SMA di selesaikan
pada tahun 1969 di SMAN II Bandung .
Pada tahun 1970 penulis tercatat sebagai mahasiswa Fakultas Peternakan
Universitas Padjadjaran Bandung dan menyandang gelar Sarjana Peternakan pada
tahun 1977. Pada tahun 1985 penulis melanjutkan pendidikan S2 pada Program
Studi Ilmu Ternak, Fakultas Pascasarjana Institut Pertanian Bogor, melalui
beasiswa TMPD dari Ditjen DIKTI, DEPDIKBUD dan memperoleh gelar
Magister Sain (MS) pada tahun 1989. Pada bulan September 2002 penulis
tercatat sebagai mahasiwa Program Doktor pada Program Studi Biologi, Fakultas
Pascasarjana Institut Pertanian Bogor dengan status swadana. Pada bulan
September 2003 mendapat beasiswa dari BPPS Ditjen DIKTI, DEPDIKNAS.
Penulis menikah dengan Ir. Burhanudin Soelaeman pada bulan Desember
1979 dan dikaruniai tiga anak, yaitu Gia Ginanjar S. Burhanudin, Gerry Nugraha
S. Burhanudin, dan Gistya Gemma Rahayu S. Burhanudin. Penulis tercatat
menjadi staf pengajar di Laboratorium Fisiologi dan Biokimia, Jurusan Nutrisi
9
PRAKATA
Alhamdulillah wa syukurillah atas berkah dan rahmat Allah SWT, penulis
dapat berhasil menyelesaikan karya ilmiah ini. Karya ilmiah yang berjudul Injeksi
Bovine Somatotropin (bST) dan Penambahan Konsentrat pada Sapi Holstein
Laktasi di Dataran Tinggi Cikole Lembang yang telah dilaksanakan sejak bulan
Juni 2005 sampai Maret 2006.
Pada kesempatan ini tak lupa penulis menyampaikan ucapan terima kasih
dan penghargaan setinggi-tingginya kepada
1. Prof. Ir. Wasmen Manalu Ph.D Ketua komisi pembimbing yang telah
memberikan arahan dan bimbingan selama penulis mengikuti program S3.
2. Drh. Syahrun Hamdani Nasution Ph.D anggota komisi pembimbing yang telah
memberikan saran, dorongan, dan koreksi dalam penyempurnaan penelitian
disertasi.
3. Dra. Nastiti Kusumorini Ph.D anggota komisi pembimbing yang telah
memberikan arahan dan koreksi dalam penyempurnaan penelitian disertasi.
4. Dr. Ir. Bagus Pryo Purwanto penguji luar komisi pada ujian tertutup yang
telah memberikan saran dan koreksi dalam penyempurnaan disertasi.
5. Dr.Ir. Chalid Talib MS penguji luar komisis pada ujian terbuka yang telah
memberikan saran dan koreksi dalam penyempurnaan disertasi.
6. Dr. Ir. Suryahadi DEA penguji luar komisi pada ujian terbuka yang telah
memberikan masukan dalam penyempurnaan disertasi.
7. Rektor Institut Pertanian Bogor, Dekan Sekolah Pascasarjana Insitut Pertanian
Bogor yang telah memberikan fasilitas yang terbaik selama penulis
menimba ilmu di program S3.
8. Departemen Pendidikan Nasional dalam hal ini Direktorat Jenderal
Pendidikan Tinggi yang telah mendanai penulis melalui BPPS (Beasiswa
Pendidikan program Pascasarjana) saat mengikuti pendidikan Program
Doktor di Insitut Pertanian Bogor .
9. Rektor Universitas Padjadjaran, Dekan Fakultas Peternakan, Universitas
Padjadjaran, dan Kepala Laboratorium Fisiologi dan Biokimia, Fakultas
kesempatan untuk mengikuti program S3 dan dorongan yang tak
henti-hentinya.
10. Kepala Dinas Peternakan Provinsi Jawa Barat dan Kepala Balai Peningkatan
Produksi Ternak Perah, Balai Pelatihan, Balai Pengujian Sarana Produksi
Peternakan Pakan, dan Balai Penyidikan Penyakit dan Kesmavet di Cikole
Lembang, yang telah memberikan izin dan fasilitas penelitian pada saat
penulis melaksanakan pengumpulan data
11. Bunda, Ibu Hj. Siti Halimah Zyin Hernawan, Keluarga besar Bpk R.H.
Mamun Hernawan dan Keluarga besar Bpk H.Soelaeman yang telah
memberikan doa, dorongan bantuan berupa materi dan spriritual yang tidak
ternilai kepada penulis.
12. Suami tercinta Ir. Burhanudin Soelaeman MM, buah hati tersayang Gia
Ginanjar BS, Gerry Nugraha BS, dan Gistya Gemma Rahayu BS, yang telah
memberikan pengertian dan dorongan dengan penuh keikhlasan, serta
kesempatan yang luas, sehingga banyak waktu-waktu keluarga yang tersita
selama penulis mengikuti program S3.
13. Bapak K.H Aan Mustafa Kamil beserta keluarga besar Tariqoh Assadzaliah
yang telah memberi semangat dan dorongan spiritual, siraman rohani yang
menyejukkan selama mengikuti program S3.
14. Dr. Ir. Aslamiyah MS dan Ir. Arief Nasution MS (dari Universitas
Hasanudin), teman seperjuangan menimba ilmu di program S3, yang telah
banyak membantu dan memberi semangat selama mengikuti program S3.
15. Grup Somatotropin (Gatot Muslim, SPt, M.Si, La Eddy SPd M.Si dari Ternate, Dr. Ir. Hudiana Hernawan MS, Ni Wayan M.Si dari Udayana) yang telah bahu membahu dalam persiapan dan pelaksanaan penelitian, dengan suasana penuh canda tawa dan kekeluargaan.
16. Kepala Dinas Peternakan dan Perikanan Kabupaten Bogor beserta jajarannya, yang telah banyak memberikan fasilitas dan bantuan kepada penulis selama mengikuti program S3
17. Poppy, Yudi, Titta, Ida, Nining, Caca, Cecen, Dedih, Mang Ade, Ayun dan
Poniman, group Cikole adalah kawan- kawan setia yang telah menemani dan
11
18. Ibu Asmarida, Ibu Sri dan Pak Wawan dari Laboratorium Fisiologi,
Departemen AFF, FKH IPB, yang telah banyak membantu penulis dalam
penyelesaian penelitian di laboratorium.
19. Dinda Dr. Dra. Iis Arifiantini MSi beserta keluarga yang telah banyak tersita
waktunya untuk membantu saat akhir penyelesaian disertasi ini.
20. Rekan-rekan dan semua pihak baik secara langsung maupun tidak langsung
yang terlibat dalam pelaksanaan dan penulisan disertasi ini.
Akhirulkalam semoga amal dan budi baik yang telah diberikan kepada
penulis menjadikan pahala dan mendapat balasan yang berlipat ganda dari
Allah SWT, Sang Maha Pemberi dan Pengabul. Kunfayakun! Semoga Karya
Ilmiah ini dapat bermanfaat bagi perkembangan khasanah biologi khususnya
peternakan di Indonesia.
`
Bogor, Mei 2007
DAFTAR ISI
Halaman
DAFTAR ISI ... i
DAFTAR TABEL ... ii
DAFTAR GAMBAR ... iii
DAFTAR LAMPIRAN ... v
PENDAHULUAN ... 1
Latar Belakang ... 1
Tujuan dan Manfaat Penelitian ... 5
Hipotesis ... 6
TINJAUAN PUSTAKA ... 7
Sapi Perah di Indonesia ... 7
Masa Laktasi ... 9
Homeostasis ... 10
Somatotropin dan Mekanisme Kerja ... 11
Somatotropin, IGF-I, dan Produksi Susu ... 16
Somatotropin, Metabolisme, dan Produksi Susu ... 18
Somatotropin, Kesehatan Ternak dan Keamanan Pangan ... 22
MATERI DAN METODE PENELITIAN ... 25
Materi Penelitian ... 25
Metode Penelitian ... 25
Alir Penelitian ... 27
Peubah yang Diamati dan Cara Pengukuran ... 29
Analisis Statistik ... 34
HASIL ... 36
Status Faali dan Hematologi ... 36
Metabolit Darah dan Hormon Metabolisme ... 40
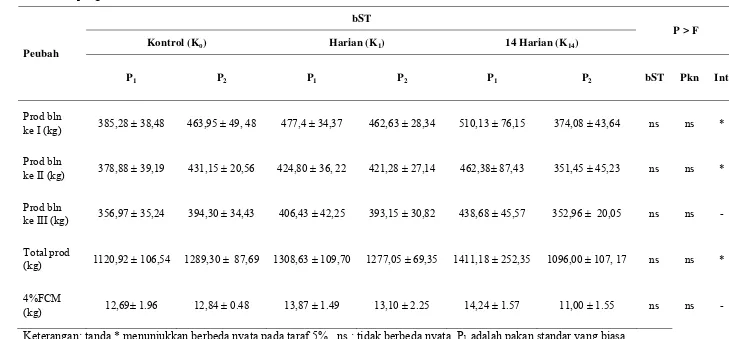
Produksi Susu... 46
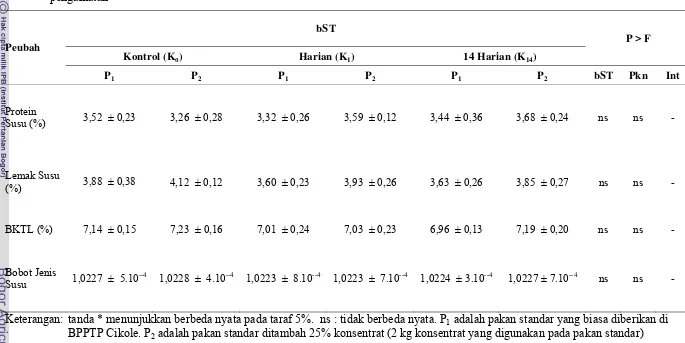
Komposisi Susu dan Berat Jenis Susu ... 53
Tubuh, Konsumsi Bahan Kering, Efisiensi Produksi Susu, dan Nilai Kondisi Ternak ... 57
PEMBAHASAN ... 65
SIMPULAN ... 89
DAFTAR PUSTAKA ... 90
13
DAFTAR TABEL
Halaman
Tabel 1 Komposisi asam-amino somatotropin manusia, sapi, domba, dan babi ... 13
Tabel 2 Komposisi pakan sapi perah laktasi di BPPTP Cikole ... 28
Tabel 3 Kandungan nutrisi pakan penelitian, imbangan hijauan dan konsentrat pakan, dan angka kebutuhan nutrien sapi penelitian .. 28
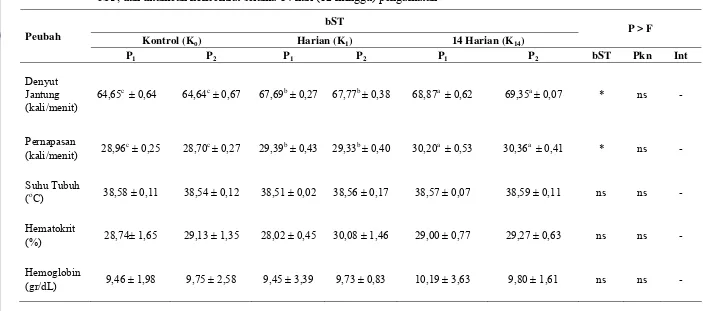
Tabel 4 Rataan denyut jantung, frekuensi pernapasan, suhu tubuh, nilai hematoktit dan kadar hemoglobin sapi yang diinjeksi bST dan
ditambah konsentrat selama 84 hari (12 minggu) pengamatan .... 37
Tabel 5 Rataan kadar metabolit dan hormon metabolisme pada sapi yang diinjeksi bST dan ditambah konsentrat selama 84 hari (12 minggu) pengamatan ... 41
Tabel 6 Rataan produksi susu dan 4% FCM pada sapi yang
disuplementasi bST dan pakan selama 84 hari (12 minggu) pengamatan ... 47
Tabel 7 Hubungan produksi susu dan waktu perlakuan dari masing-masing kombinasi perlakuan selama 84 hari (12 minggu) pengamatan ... 52
Tabel 8 Rataan komposisi dan bobot jenis susu pada sapi yang diinjeksi bST dan ditambah konsentrat selama 84 hari (12 minggu) pengamatan ... 54
DAFTAR GAMBAR
Halaman
Gambar 1 Kurva Produksi Susu selama Laktasi (Alim et al. 2001) ... 9
Gambar 2 Aktivitas langsung dan tidak langsung ST pada pertumbuhan
dan metabolisme (Kamil et al. 2001) ... 15
Gambar 3 Koordinasi antarjaringan dalam adaptasi metabolik pada sapi
laktasi yang diinjeksi bST (Aker 2002) ... 21
Gambar 4 Alir Penelitian ... 27
Gambar 5 Rataan denyut jantung efek utama pakan (a), bST (b), dan kombinasi perlakuan (c), selama 84 hari (12 minggu) pengamatan ... 36
Gambar 6 Rataan frekuensi pernapasan efek utama pakan (a), bST (b), dan kombinasi perlakuan (c), selama 84 hari (12 minggu) pengamatan ... 38
Gambar 7 Rataan suhu tubuh efek utama pakan (a), bST (b), dan kombinasi perlakuan (c), selama 84 hari (12 minggu) pengamatan ... 39
Gambar 8 Rataan nilai hematokrit efek utama pakan (a), bST (b), dan kombinasi perlakuan (c), selama 84 hari (12 minggu) pengamatan ... 39
Gambar 9 Rataan kadar hemoglobin efek utama pakan (a), bST (b), dan kombinasi perlakuan (c), selama 84 hari (12 minggu) pengamatan ... 40
Gambar 10 Rataan kadar glukosa serum efek utama pakan (a), bST (b), dan kombinasi perlakuan (c), selama 84 hari (12 minggu) pengamatan ... 42
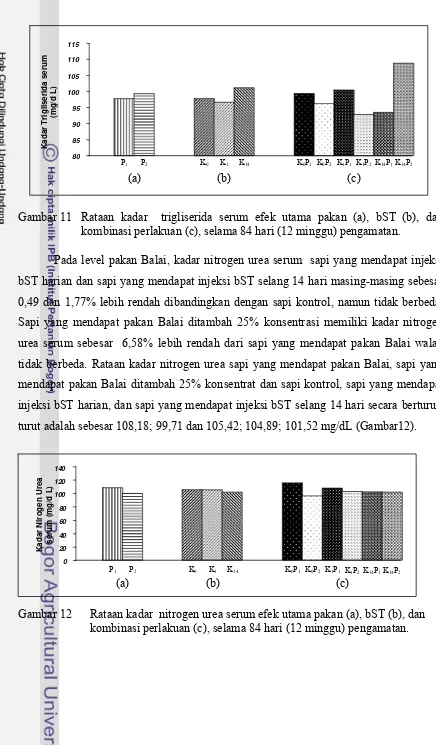
Gambar 11 Rataan kadar trigliserida serum efek utama pakan (a), bST (b), dan kombinasi perlakuan (c), selama 84 hari (12 minggu) pengamatan ... 43
Gambar 12 Rataan kadar nitrogen urea serum efek utama pakan (a), bST (b), dan kombinasi perlakuan (c), selama 84 hari (12 minggu)
pengamatan ... 43
Gambar13 Rataan kadar kortisol serum efek utama pakan (a), bST (b), dan kombinasi perlakuan (c), selama 84 hari (12 minggu) pengamatan ... 44
Gambar 14 Rataan kadar T4 serum efek utama pakan (a), bST (b), dan kombinasi perlakuan (c), selama 84 hari (12 minggu) pengamatan ... 45
15
Gambar 16 Rataan produksi susu (kg/minggu) masing-masing kombinasi
perlakuan selama penelitian ... 46
Gambar 17 Interaksi injeksi bST harian, injeksi bST selang 14 hari, dan pakan satandar (P1), pakan standar ditambah 25% konsentrat
(P2) ... 49
Gambar 18 Hubungan produksi susu dan waktu perlakuan dari masing-yang lebih rendah masing kombinasi perlakuan selama 84 hari
(12 minggu) ... 51
Gambar19 Rataan produksi susu 4% FCM efek utama pakan (a), bST (b), dan kombinasi perlakuan (c), selama 84 hari (12 minggu) pengamatan ... 53
Gambar 20 Rataan protein susu (%) efek utama pakan (a), bST (b), dan kombinasi perlakuan (c), selama 84 hari (12 minggu) pengamatan ... 55
Gambar21 Rataan lemak susu (%) efek utama pakan (a), bST (b), dan kombinasi perlakuan (c), selama 84 hari (12 minggu) pengamatan ... 55
Gambar 22 Rataan BKTL susu (%) efek utama pakan (a), bST (b), dan kombinasi perlakuan (c), selama 84 hari (12 minggu) pengamatan ... 56
Gambar 23 Rataan bobot jenis susu efek utama pakan (a), bST (b), dan kombinasi perlakuan (c), selama 84 hari (12 minggu) pengamatan ... 56
Gambar 24 Rataan bobot tubuh efek utama pakan (a), bST (b), dan kombinasi perlakuan (c), selama 84 hari (12 minggu) pengamatan ... 57
Gambar 25 Interaksi injeksi bST harian, injeksi bST selang 14 hari, dan pakan standar (P1), pakan standar ditambah 25% konsentrat
(P2) pada penambahan bobot tubuh ... 59
Gambar 26 Rataan konsumsi bahan kering efek utama pakan (a), bST (b), dan kombinasi perlakuan (c), selama 84 hari (12 minggu) pengamatan ... 59
Gambar 27 Rataan efisiensi produksi susu (EPS) efek utama pakan (a), bST (b), dan kombinasi perlakuan (c), selama 84 hari (12 minggu) pengamatan ... 60
Gambar 28 Rataan nilai kondisi ternak (NKT) efek utama pakan (a), bST (b), dan kombinasi perlakuan (c), selama 84 hari (12 minggu)
DAFTAR LAMPIRAN
Halaman
Lampiran 1 Analisis ragam rataan denyut jantung selama 84 hari pengamatan ... 91
Lampiran 2 Analisis ragam rataan frekuensi pernapasan selama 84 hari
pengamatan ... 91
Lampiran 3 Analisis ragam rataan suhu tubuh selama 84 hari pengamatan ... 91
Lampiran 4 Analisis ragam rataan nilai hematokrit selama 84 hari pengamatan ... 92
Lampiran 5 Analisis ragam rataan kadar hemoglobin selama 84 hari pengamatan ... 92
Lampiran 6 Analisis ragam rataan glukosa serum selama 84 hari pengamatan ... 92
Lampiran 7 Analisis ragam rataan nitrogen urea serum selama 84 hari
pengamatan ... 92
Lampiran 8 Analisis ragam rataan trigliserida serum selama 84 hari pengamatan ... 93
Lampiran 9 Analisis ragam rataan kortisol serum selama 84 hari pengamatan ... 93
Lampiran 10 Analisis ragam rataan T4 serum selama 84 hari pengamatan ... 93
Lampiran 11 Analisis ragam rataan T3 serum selama 84 hari pengamatan ... 93
Lampiran 12 Analisis ragam total produksi susu bulan ke-1 perlakuan ... 89
Lampiran 13 Analisis ragam total produksi susu bulan ke-2 perlakuan ... 89
Lampiran 14 Analisis ragam total produksi susu bulan ke-3 perlakuan ... 89
Lampiran 15 Analisis ragam total produksi susu selama 84 hari pengamatan ... 89
Lampiran 16 Analisis ragam produksi susu 4% FCM selama 84 hari pengamatan ... 90
Lampiran 17 Analisis ragam rataan kadar protein susu selama 84 hari
pengamatan ... 90
Lampiran 18 Analisis ragam rataan kadar lemak susu selama 84 hari pengamatan ... 90
Lampiran 19 Analisis ragam rataan kadar berat kering tanpa lemak selama 84 hari pengamatan ... 90
17
Lampiran 21 Analisis ragam bobot tubuh awal sapi uji ... 94
Lampiran 22 Analisis ragam bobot tubuh akhir ... 94
Lampiran 23 Analisis ragam pertambahan bobot tubuh selama penelitin
... 94
Lampiran 24 Analisis ragam rataan bobot tubuh selama 84 hari (12 minggu) pengamatan ... 94
Lampiran 25 Analisis ragam rataan konsumsi bahan kering selama 84
hari pengamatan ... 94
Lampiran 26 Analisis ragam efisiensi produksi susu selama 84 hari (12
minggu) pengamatan ... 95
Lampiran 27 Analisis ragam rataan nilai kondisi ternak 1-4 minggu pengamatan ... 95
Lampiran 28 Analisis ragam rataan nilai kondisi ternak 5-8 minggu pengamatan ... 95
Lampiran 29 Analisis ragam rataan nilai kondisi ternak 8-12 minggu pengamatan ... 95
Lampiran 30 Analisis homogenitas koefisien regresi hubungan antara produksi susu dan waktu pengamatan antarkombinasi perlakuan ... 108
Lampiran 31 Analisis homogenitas koefisien regresi hubungan antara produksi susu dan waktu pengamatan antarkombinasi perlakuan ... 108
Lampiran 32 Analisis homogenitas koefisien regresi hubungan antara produksi susu dan waktu pengamatan antarkombinasi perlakuan ... 109
Lampiran 33 Analisis homogenitas koefisien regresi hubungan antara produksi susu dan waktu pengamatan antarkombinasi perlakuan ... 109
Lampiran 34 Analisis homogenitas koefisien regresi hubungan antara produksi susu dan waktu pengamatan antarkombinasi perlakuan ... 109
PENDAHULUAN
Latar Belakang
Angka konsumsi susu di Indonesia saat ini tercatat paling rendah di
Asean, yaitu 7 kg/kapita/tahun atau setara dengan 19 cc/kapita/hari, sedangkan
Malaysia, Thailand dan Bangladesh mempunyai rataan konsumsi susu di atas 20
kg/kapita/tahun. Berdasarkan data produksi susu segar yang dilaporkan Direktorat
Jenderal Peternakan (2005) menunjukkan adanya peningkatan produksi susu dari
tahun 2002, 2003, 2004, dan 2005 yang secara berturut-turut sebesar 493 375;
553 442; 550 000; dan 550 000 ton. Namun demikian, produksi susu dalam
negeri baru dapat mensuplai sekitar 30% saja sehingga kekurangannya yaitu
70% atau sebanyak 1 425 200 ton diimpor dalam bentuk susu olahan dari
Selandia Baru, Australia, Irlandia, Belanda, Filipina, dan Thailand. Kegiatan
impor susu ditengarai sebagai akibat rendahnya produksi susu dalam negeri, dan
di samping itu juga akibat kesenjangan yang kian melebar antara angka
kebutuhan konsumsi dan produksi susu dalam negeri.
Pada tahun 2005 populasi sapi perah di Indonesia adalah sekitar 374 000
ekor dan hampir 91,1% dikelola dalam usaha yang berbasis peternakan rakyat
dengan produksi susu yang masih rendah, yaitu berkisar antara 8 dan 10
L/ekor/hari atau antara 2 500 dan 3 000 L/laktasi. Sejumlah 8,9% lainnya
dikelola dalam perusahaan komersial dengan produksi susu antara 3 000 dan
5 000 L/laktasi. Berdasarkan asal usulnya, sapi perah yang banyak dipelihara di
Indonesia adalah bangsa FH yang berasal dari negara beriklim
subtropis/temperate, yang telah mengalami proses adaptasi lingkungan. Namun,
hasil produksinya masih jauh di bawah rataan sapi-sapi perah di negara asalnya,
yakni 7 342 kg/laktasi (Talib et al. 2003).
Indonesia termasuk daerah tropis lembab yang dicirikan oleh suhu
lingkungan, kelembaban, dan curah hujan yang tinggi. Sapi perah termasuk hewan
homoioterm yang akan selalu menjaga keseimbangan antara panas tubuhnya
dengan lingkungan disekeliling. Perubahan lingkungan luar akan segera diikuti
dengan perubahan lingkungan dalam tubuh, dan dikembalikan ke kondisi semula
dengan proses homeostasis. Konsep homeostasis melibatkan keseimbangan panas,
kegiatan organ sirkulasi, kardiovaskular, dan lain-lain.
Perubahan lingkungan dalam tubuh akan menimbulkan serangkaian
perubahan reaksi biokimia dalam tubuh, yang berdampak pada aktivitas sel dan
kerja organ yang pada gilirannya menyebabkan banyak energi yang digunakan
hanya untuk mempertahankan kondisi tubuh atau untuk hidup pokok. Proses yang
jika berlangsung cukup lama dan berkesinambungan ini akan mempengaruhi
penggunaan energi, sedangkan produksi yang dihasilkan ternak merupakan
kelebihan energi setelah digunakan untuk mengelola atau mempertahankan
kebutuhan hidup pokok. Ternak dalam kondisi tersebut di atas cenderung
menurunkan level konsumsi pakan bila berada di daerah panas. Proses tersebut
diyakini sebagai penyebab rendahnya produktivitas ternak di daerah panas karena
serangkaian proses metabolisme tubuh yang harus dilakukan untuk
mempertahankan sistem kerja organ-organ tubuh.
Sapi perah asal subtropis akan berkembang biak secara baik dengan
produksi susu yang cukup tinggi pada daerah yang mempunyai ketinggian dari
750 sampai 1 250 m di atas permukaan laut, yang memiliki kisaran suhu udara
antara 17 dan 22°C dan kelembaban di atas 55% (Atmadilaga 1979). Hasil
penelitian di Thailand, yang juga negara tropis, menunjukkan bahwa sapi-sapi
perah subtropis dapat beraklimatisasi dengan baik pada suhu di bawah 18°C dan
kelembaban di atas 55% (Siregar et al. 2003). Dengan alasan ini, populasi sapi
perah di Indonesia tersebar di daerah-daerah tertentu saja, khususnya yang
mempunyai kesamaan iklim dengan daerah asalnya.
Dalam menanggulangi permasalahan tentang kesenjangan antara produksi
susu dan kebutuhan konsumsi susu nasional, pemerintah dalam kebijakan
peningkatan produksi dan kualitas susu tahun 2010 mencanangkan peningkatan
suplai susu dalam negeri dari 30 menjadi 40% melalui peningkatan produksi susu
dari 8-10 L menjadi 15L/ekor/hari, kepemilikan ternak dari 4-6 ekor menjadi 10
ekor/kepala keluarga (Departemen Pertanian 2006). Sebagai tindak lanjut maka
berbagai upaya peningkatan produktivitas ternak telah banyak dilakukan, di
antaranya dengan memanipulasi lingkungan, nutrisi, bahkan reproduksi melalui
inseminasi buatan dan trasfer embrio. Namun, hasilnya dipandang belum
3
dilaporkan, sementara di luar negeri penggunaan produk bioteknologi seperti
hormon pertumbuhan rekombinan sudah luas di kalangan peternak.
Somatotropin sapi rekombinan atau bovine somatotropin (bST)
rekombinan telah dikenal sejak tahun 80-an, yang merupakan hasil bioteknologi
khususnya dalam manipulasi genetik, reproduksi dan fisiologi, dan aplikasinya
sangat mewarnai perubahan dalam industri peternakan. Mekanisme kerja
somatotropin dalam peningkatan produksi susu pada sapi laktasi secara langsung
terlibat dalam penggalangan nutrisi. Sementara aktivitas sintesis susu dalam
kelenjar susu tidak secara langsung oleh kerja ST karena di dalam sel-sel kelenjar
susu hampir tidak ditemukan reseptor ST, namun reseptor IGF-1 banyak djumpai
sehingga para peneliti meyakini bahwa perangsangan produksi susu melalui
injeksi bST pada sapi laktasi dimediasi oleh insulin like growth factor-1 (IGF-1).
Hal ini terlihat jelas adanya peningkatan konsentrasi IGF-1 mencapai 3 sampai 4
kali lipat setelah diberi perlakuan ST, dan peningkatan akan terus berlanjut selama
perlakuan.
Somatotropin pada sapi laktasi berperan dalam proses-proses yang
berkaitan erat dengan penggalangan nutrien untuk kebutuhan sintesis susu
sehingga perangsangan produksi susu melalui injeksi ST berdampak pada
metabolisme. Peningkatan sintesis susu memerlukan sejumlah tambahan nutrien
baik substrat atau prekursor sehingga sebagai konsekuensinya aliran darah menuju
kelenjar susu perlu ditingkatkan. Dalam penggalangan nutrien, ST berperan
sebagai agen homeorhesis dengan melakukan serangkaian adaptasi metabolisme
dalam tubuh.
Semakin tersedianya ST hasil isolasi pituitari atau rekayasa genetika,
penggunaan ST dalam industri peternakan semakin berkembang pesat. Hasil
ulasan Peel dan Bauman (1987), Chalupa dan Galligan (1989) menyimpulkan
bahwa penggunaan ST tidak memberikan dampak yang merugikan pada ternak,
bahkan akan tampak seperti produksi susu dari jenis genetik unggul jika diberikan
pada sapi yang berproduksi tinggi. Injeksi ST pada sapi-sapi yang berproduksi
rendah dengan manajemen yang baik akan memberikan respons yang lebih baik
dibandingkan dengan sapi yang berproduksi tinggi.
Bovine somatotropin (bST) semakin populer di kalangan peternak, dan
waktu tertentu (14 atau 28 hari). Cara injeksi tersebut di atas akan berdampak
pada konsentrasi ST dalam darah yang pada gilirannya akan berdampak pada
kerja ST dalam penggalangan nutrisi. Injeksi bST selang 14 hari dilaporkan
menghasilkan produksi susu yang lebih rendah, yaitu hanya 70-79%
dibandingkan dengan cara injeksi bST harian, namun dipandang lebih efektif
untuk aplikasi di lapangan (Chilliard 1989, Jenny et al.1992, Etherton&Bauman
1998). Penambahan hormon eksogen berpengaruh pada konsentrasi hormon–
hormon lain yang saling berkaitan secara metabolis. Respons sapi laktasi terhadap
injeksi bST akan meningkatkan sekresi susu harian atau mingguan, dan
peningkatan produksi susu akan dipertahankan selama pemberian yang
berkesinambungan (Akers 2002), tetapi akan cepat kembali ke level basal ketika
injeksi bST dihentikan ( Manalu 2001; Akers 2002). Injeksi ST dalam industri
peternakan saat ini dianggap merupakan suatu terobosan dari hasil bioteknologi
yang cukup ekonomis dan efisien, sekalipun dibandingkan dengan upaya
pemberantasan penyakit, vaksinasi atau penggunaan bioteknologi reproduksi
(Hardjopranjoto 2001).
Somatotropin sangat menjanjikan peningkatan produktivitas serta efisiensi
penggunaan pakan pada sapi perah. Penggunaan bST pada tingkat peternak dapat
meningkatkan rataan produksi susu hingga 5 kg/hari atau berkisar antara 15 dan
20% tanpa menimbulkan penyakit metabolis atau perubahan kualitas susu yang
berarti (Manalu 1994), bahkan laporan terakhir peningkatan produksi susu dapat
mencapai antara 30 dan 40% bergantung pada dosis pemakaian (Akers 2002).
Namun, demikian pasokan pakan yang diberikan perlu mendapat perhatian,
khususnya dalam hal keseimbangan nutrisi pakan agar bisa mendukung
penggalangan pasokan bahan sintesis susu.
Pakan sapi perah terdiri atas penyusun pakan terbesar berupa hijauan dan
konsentrat sebagai tambahan. Keseimbangan antara hijauan dan konsentrat akan
berpengaruh pada produksi dan komposisi susu. Pakan melalui proses fermentasi
rumen akan menghasilkan sejumlah asam lemak terbang (asam asetat, asam
propionat, asam butirat) yang akan digunakan sebagai sumber energi utama
(Akers 2002). Hijauan secara kuantitas cenderung berpengaruh pada produksi
asam asetat dan berkaitan erat dengan kadar lemak susu serta penyediaan rangka
5
propionat yang melalui proses glukoneogenesis dalam hati akan diubah menjadi
glukosa. Glukosa merupakan substrat dan prekursor yang sangat dibutuhkan
dalam proses sintesis susu khususnya dalam sintesis laktosa. Untuk dapat
meningkatkan produksi susu perlu dilakukan penambahan sumber glukosa yang
tiada lain dari konsentrat.
Peningkatan produksi susu melalui injeksi bST pada usaha sapi perah
kiranya dapat dijadikan salah satu jalan pintas bagi penyediaan kebutuhan
konsumsi susu dalam negeri. Namun, perlu penjajagan ke arah tersebut. Untuk
mendapatkan data konkrit khususnya bagi peternakan di Indonesia yang berlatar
belakang iklim tropis, dan dicirikan rendahnya produktivitas yang diakibatkan
oleh suhu lingkungan dan kelembaban tinggi, maka perlu dilakukan penelitian
tentang injeksi somatotropin (bST) pada sapi laktasi di dataran tinggi.
Tujuan dan Manfaat Penelitian
Penelitian ini bertujuan mengkaji potensi sapi perah yang sudah
beradaptasi dengan iklim Indonesia dalam merespons injeksi bST dan
penambahan konsentrat dalam upaya peningkatan produktivitas ternak. Tujuan
penelitian secara rinci :
1. Membandingkan efektivitas selang waktu injeksi bST dan penambahan 25%
konsentart pada profil metabolit dan hormon metabolisme darah yang erat
kaitannya dengan sintesis susu.
2. Membandingkan pengaruh selang waktu injeksi bST dan penambahan 25%
konsentrat pada produksi susu dan komposisi susu.
3. Membandingkan efisiensi produksi susu masing-masing selang waktu injeksi
bST dan penambahan 25% konsentrat.
Diharapkan hasil penelitian ini dapat menambah khasanah biologi dalam
penerapan produk bioteknologi, yaitu somatotropin rekombinan dan selanjutnya
dapat dijadikan informasi dasar bagi para peternak atau penentu kebijakan dalam
Hipotesis
Injeksi bST selang 14 hari dan penambahan 25% konsentrat pada sapi
Holstein laktasi akan menghasilkan produksi susu dan komposisi yang sama
dengan hasil injeksi bST harian dan pakan standar, yang dinilai melalui
perubahan status faali, hematologi, metabolit, hormon metabolisme, bobot tubuh,
7
TINJAUAN PUSTAKA
Sapi Perah di Indonesia
Rumpun sapi perah Fries Holstein (FH) banyak dipelihara di Indonesia.
Rumpun sapi perah ini berasal dari daerah subtropis atau lebih jelasnya dari
provinsi Holland Utara dan Friesland Barat Belanda. Berdasarkan asal usulnya,
rumpun sapi ini dikembangkan dari nenek moyang sapi liar Bos (Taurus) Typicus
primigenius dan telah diternakkan di negeri Belanda sekurang-kurangnya 2000
tahun.
Sapi FH mempunyai ciri-ciri kepala panjangnya sedang, mulut lebar
dengan hidung terbuka lebar, rahang kuat, dahi lebar, leher panjang dan pipih.
Selain itu, sapi FH mempunyai punggung kuat dan rata dengan ruas-ruas tulang
belakang yang rapih hubungannya, dan pinggang lebar dan kuat. Kedudukan
keempat kaki sapi FH membentuk empat persegi panjang, dan bentuk kaki halus
tetapi tampak kuat. Warna tubuh pada umumnya belang hitam putih, tetapi di
negeri Belanda bagian timur dan di Jerman terdapat kelompok sapi yang berwarna
belang merah putih (Sosroamidjojo & Soeradji 1978).
Pada umumnya sapi-sapi Eropa mempunyai kisaran suhulingkungan
nyaman yang rendah sehingga lebih toleran terhadap suhu lingkungan dingin
dibandingkan suhu lingkungan panas. Agar sapi perah FH dapat memberikan
produksi maksimal sesuai dengan kemampuan genetiknya, perlu lokasi yang
mempunyai lingkungan mikro yang hampir menyamai tempat asalnya. Wilayah
yang sesuai untuk pengembangan sapi perah adalah daerah yang memiliki suhu
lingkungan antara 0 dan 20ºC, sedangkan suhu kritis untuk sapi Holstein, Brown
Swiss, dan Brahman secara berturut-turut adalah 21; 24-27; 32ºC (Hafez 1968).
Sapi perah akan berproduksi maksimal pada suhu lingkungan antara 4,4 dan
21,1ºC dengan tingkat kelembaban berkisar antara 60 dan 80%. Kenaikan suhu
lingkungan akan menurunkan konsumsi pakan dan produksi susu sementara
konsumsi air meningkat (Schmidt 1971).
Sekitar abad ke-19 sapi FH pertama kali didatangkan ke Indonesia oleh
pemerintah Belanda untuk memenuhi kebutuhan susu bagi bangsa Belanda yang
ada di Indonesia. Indonesia terletak di daerah tropis yang sangat sedikit daerah
sehingga kombinasi tersebut menyebabkan fluktuasi suhu harian dan kelembaban
yang sangat luas. Secara geografis, Indonesia terletak antara 5 dan 7º Lintang
Utara dan Lintang Selatan, yang mempunyai suhu yang relatif konstan (± 27ºC),
curah hujan yang tinggi berkisar dari 2 032 sampai 3 048 mm (Williamson &
Payne 1993). Dalam kurun waktu yang cukup panjang sapi-sapi asal Belanda
tersebut mengalami aklimatisasi dengan lingkungan tropis Indonesia, bahkan
pada saat ini telah berkembang pesat peranakan FH yang telah lebih beradaptasi
dengan lingkungan.
Suhu optimal untuk produksi susu sapi yang berasal dari daerah subtropis
adalah 10°C (Williamson & Payne 1993). Penampilan produksi masih cukup baik
walau suhu lingkungan meningkat sampai 21,1°C (Sutardi, 1981). Suhu kritis
yang berdampak pada penurunan produksi susu yang tajam adalah antara 21
sampai 27°C (Williamson & Payne 1993). Tidak mengherankan apabila
pemeliharaan sapi FH sebagian besar terkonsentrasi pada daerah-daerah dataran
tinggi (±1000 m dpl) seperti Garut, Lembang, Pangalengan, Pujon, Nongkojajar,
yang lingkungannya hampir menyamai daerah asalnya (Siregar et al. 2003).
Populasi sapi perah di Indonesia menunjukkan perkembangan. Selama
kurun waktu 1970 hingga 2001 terjadi peningkatan populasi dari 52 000 ekor
menjadi 347 000 ekor, sedangkan sejak tahun 1994 produksi susu tercatat
426 727 ton dan meningkat menjadi 550 000 ton pada tahun 2004 (Direktorat
Jenderal Peternakan 2005).
Sapi Fries Holstein mempunyai produksi susu yang tinggi (4 000 sampai
5 000 L/tahun), dan di daerah tropis dalam kisaran dari 2 500 sampai 5 000 kg/
laktasi (Pane 1986). Tampaknya sapi Fries Holstein mengalami penurunan
produktivitas khususnya pada peternakan rakyat: produksi susu mencapai 8 L/hari
dan pada peternakan sapi perah komersial berkisar dari 3 000 sampai 5 000 L/305
hari, sedangkan di negara asalnya rata-rata produksi mencapai 7 342 kg/laktasi
9
Masa Laktasi
Masa laktasi seekor sapi perah menunjukkan lamanya waktu sapi perah
tersebut menghasilkan susu dalam satu periode (Bath et al.1978). Periode laktasi
normal berlangsung kira-kira 44 minggu atau 305 hari. Produksi susu sapi perah
pada awal laktasi umumnya meningkat secara dramatis dan mencapai puncaknya
pada minggu keempat hingga kedelapan setelah partus dan menurun secara
perlahan sampai ahir masa laktasi (NRC 1988). Produksi susu selama 2 bulan
pertama laktasi mencapai 145% dari rataan produksi satu periode laktasi,
sedangkan pada bulan ketiga dan keempat menunjukkan penurunan menjadi
120%, bulan laktasi kelima dan keenam produksi susu sama dengan rataan
produksi dalam satu periode laktasi dan setelah itu produksi menjadi 78% pada
bulan laktasi ketujuh dan kedelapan, akan menurun hingga 70% ketika menjelang
beranak (Tanuwiria 2004). Sebagai gambaran kurva produksi susu selama laktasi
disajikan pada Gambar 1.
Produksi
Susu
(
Kg)
BULAN
0 1 2 3 4 5 6 7 8 9 10
Gambar 1 Kurva Produksi Susu selama Laktasi (Alim et al. 2001)
Produksi susu setiap periode laktasi akan meningkat sejalan dengan
bertambahnya umur, dan puncak prestasi produksi dicapai pada umur antara 6 dan
7 tahun. Laktasi pertama sapi betina yang sehat akan terjadi pada umur dua tahun
dengan produksi susu 75% dari produksi umur dewasa, laktasi kedua terjadi pada
umur tiga tahun dengan produksi 85% dari produksi umur dewasa, laktasi ketiga
92% dan 98% dari produksi umur dewasa. Umur dewasa dicapai pada 6 tahun,
keadaan ini sedikit bervariasi di antara sapi perah. Jika sapi betina berumur dari 8
sampai 9 tahun atau lebih maka produksi susu akan menunjukkan penurunan
setiap laktasi (Schmidt et al. 1988).
Jumlah produksi susu per laktasi dari seekor sapi perah sangat dipengaruhi
oleh bangsa sapi, umur, musim kelahiran, geografis, masa laktasi, manajemen,
nutrien, dan frekuensi pemerahan (Homan & Wattiaux 1996). Selain itu, jumlah
produksi susu per laktasi dipengaruhi juga oleh manajemen sehingga lambatnya
pengaturan perkawinan dalam periode laktasi akan memungkinkan periode laktasi
lebih panjang.
Homeostasis
Semua ternak merupakan hewan homeotermis yang mempunyai suhu
tubuh yang relatif tetap atau mempunyai kisaran suhu tubuh yang sempit. Untuk
dapat mempertahankan suhu tubuh yang tetap, ternak harus mampu mengatur
keseimbangan antara panas yang diproduksi dari hasil metabolisme seluler dan
yang diperoleh dari lingkungan dengan pengeluaran panas dari tubuh (Hafez
1968).
Produksi panas tubuh berlangsung secara berkesinambungan di dalam
protoplasma sebagai hasil oksidasi zat-zat makanan terutama yang terjadi di
dalam tenunan-tenunan otot dan kelenjar. Ternak tetap memerlukan energi untuk
menyelenggarakan proses-proses untuk mendukung hidup pokok, yaitu sirkulasi
darah, respirasi, gerak otot, ekskresi, dan lain-lainnya (Soeharsono 1984). Selain
itu, panas diperoleh secara langsung dari luar tubuh berupa penyerapan melalui
permukaan tubuh dari panas matahari (Cole 1962; Hafez 1968). Proses fisiologis
hanya akan berlangsung baik apabila kondisi lingkungan luar tubuh (millieu
exterieur) dan di dalam tubuh (millieu interieur) berada dalam keseimbangan.
Perubahan kondisi lingkungan luar tubuh akan mengubah kondisi lingkungan di
dalam tubuh.
Pada ternak homoiotermis, perubahan dari luar akan berdampak pada
serangkaian proses dalam tubuh untuk mengembalikan ke dalam keadaan yang
relatif tetap. Proses tersebut dikenal dengan homeostasis. Setiap individu akan
11
environment) dan lingkungan dalam tubuh (internal enviroment). Perubahan
lingkungan eksternal akan menimbulkan banyak perubahan reaksi biokimia yang
berdampak pada gangguan kerja sel dan kerja organ yang mengakibatkan banyak
energi yang digunakan hanya untuk mempertahankan kondisi tubuh atau hidup
pokok (Soeharsono 1984).
Konsep homeostasis menyangkut keseimbangan panas, pengaturan suhu
tubuh, keseimbangan kimiawi air, persenyawaan karbon, elektrolit, kardiovaskular
dan lain-lain. Kondisi luar yang menimbulkan perubahan di dalam tubuh terjadi
terus menerus sehingga lambat laun ternak akan terbiasa dengan perubahan di luar
tersebut dan kegiatan proses homeostasis semakin ringan menandakan ternak telah
mengalami proses penyesuaian fisiologis. Dalam jangka panjang akan terjadi
proses adaptasi, namun tidak memperlihatkan produktivitas yang tinggi
(Soeharsono 1984).
Somatotropin dan mekanisme kerja
Somatotropin (ST) adalah nama ilmiah hormon pertumbuhan (growth
hormone atau GH) yang merupakan hormon protein atau hormon polipeptida
dengan rangkaian 190-191 residu asam amino yang membentuk satu molekul
polipeptida. Somatotropin disintesis dan disekresikan oleh sel-sel somatotrof yang
terletak dalam lobus anterior kelenjar pitiutari (Djojosoebagio 1990; Manalu
1994; Soeharsono 2001), dan sekresinya sangat dipengaruhi oleh faktor neural,
metabolik, dan hormonal (Djojosoebagio 1990).
Fungsi fisiologis hormon ini adalah mempengaruhi proses metabolisme
yang menyangkut pertumbuhan melalui stimulasi sintesis protein, meningkatkan
transportasi asam amino ke dalam sel, mempengaruhi metabolisme karbohidrat,
glukoneogenesis dalam hati, memacu mobilisasi lemak tubuh, dan mempengaruhi
metabolisme mineral (Hardjopranjoto 2001; Koentjoko 2001; Soeharsono 2001).
Bovine somatotropin (bST) mampu mempercepat pengangkutan asam amino
melalui dinding sel ke dalam sitoplasma sehingga dapat menambah konsentrasi
asam amino dalam sel. Selain itu, bST juga mampu meningkatkan pembentukan
asam ribonukleat (RNA) dalam inti sel sehingga dapat mendorong proses
transkripsi dan translasi dalam ribosom di dalam sitoplasma sel (Hardjopranjoto
Secara spesifik terdapat 3 macam bentuk somatotropin. Bentuk pertama
adalah ST dengan 191 asam amino, dengan berat molekul 22 kDa, yang
mengandung dua jembatan disulfida yang menghubungkan asam amino 53 dan
165 membentuk suatu loop besar dan asam amino 182 dan 189 dekat terminus
karboksil dari peptida membentuk loop yang kecil. Bentuk ST ini banyak terdapat
dalam kelenjar pituitari. Bentuk kedua adalah ST yang mempunyai urutan asam
amino yang sama dengan bentuk pertama, tapi kehilangan 15 asam amino nomor
32-46 dari terminal Amino, dengan berat molekul 20 kDa, dan terdapat sekitar 10
sampai 15% dari hormon pituitari. Bentuk ketiga merupakan dimerisasi 2 bentuk
22 kDa peptida yang dihubungkan dengan ikatan disulfida antar-rantai dengan
berat molekul 45 kDa dan hanya ditemukan sekitar 1% dari jumlah hormon
pituitari. Secara keseluruhan perbedaan bentuk tersebut menyebabkan adanya
perbedaan fungsi biologis somatotropin. Bentuk 20 kDa mempunyai ikatan yang
kurang efektif terhadap reseptor hati dan kelenjar susu dibandingkan dengan
bentuk 22 kDa, walaupun kedua hormon tersebut sama-sama merangsang
pertumbuhan (Kamil et al. 2001). Komposisi asam amino penyusun ST berbeda
antarspesies (Tabel 1) sehingga aktivitas biologis ST dari satu spesies tidak akan
mempunyai pengaruh apabila disuntikkan ke spesies lain, atau dengan kata lain
setiap somatotropin dari satu spesies mempunyai kekhususan sendiri (species
specifity) (Turner & Bagnara l988; Djojosoebagio 1990; Soeharsono 2001).
Sekresi ST oleh kelenjar hipofisa dilakukan sesuai kebutuhan fisiologis
melalui dua faktor, yakni growth hormone releasing factor (GHRF) dan growth
hormone inhibiting factor (GHIF) atau somatostatin (Gambar 2). Somatotropin
(ST) setelah dikeluarkan oleh pituitari akan diangkut melalui sistem aliran darah,
namun dengan sifat molekulnya yang besar hormon ini tidak dapat menerobos
membran sel sehingga hormon protein memerlukan kehadiran reseptor spesifik di
13
Tabel 1. Komposisi asam amino somatotropin manusia, sapi, domba, dan babi
Asam Amino Manusia Sapi Domba Babi
Lisina 9 11 13 11
Histidina 3 3 3 3
Arginina 11 13 13 12 Aspartat 20 16 16 15 Treonina 10 12 12 7 Serina 18 13 12 14 Glutamat 26 24 25 24 Prolina 8 6 8 7 Glisina 8 10 10 8 Alanina 7 15 14 16 Sistina 4 4 4 4 Valina 7 6 7 8
Metionina 3 4 4 3
Isoleusina 8 7 7 6
Leusina 26 27 22 24 Tirosina 8 6 6 7
Fenilalanina 13 13 13 12
Triptofan 1 1 1 1
Sumber : Hariadi et al. (2001)
Reseptor ST adalah suatu glikoprotein berantai tunggal dari 620 asam
amino dengan suatu domain ekstraseluler yang luas (246 residu asam amino),
domain transmembran tunggal (24 residu), dan domain sitoplasmik yang panjang
(350 residu). Reseptor ST terdapat dalam dua bentuk, yaitu reseptor yang
berafinitas tinggi dan rendah. Reseptor berafinitas tinggi sangat spesifik dikaitkan
dengan pertumbuhan. Bagian ekstraseluler meliputi hormone binding site dan
suatu daerah yang penting untuk dimerisasi reseptor. Reseptor ST banyak
didapatkan pada permukaan sel seperti hati, otot, tulang, dan jaringan adiposa
(Granner 1997; Kamil et al, 2001).
Somatotropin atau growth hormone yang beredar dalam aliran darah
diikat oleh suatu growth hormone binding proteins (GHBP). Fungsi GHBP ini
masih belum jelas, tetapi diyakini bahwa kehadiran GHBP dapat meningkatkan
waktu paruh ST, memodifikasi distribusinya, mengurangi efek biologis ST pada
preadiposit, dan mengurangi efek biologis sekresi ST pulsatif dengan mengurangi
(GHBP) berada dalam sirkulasi darah beberapa spesies termasuk manusia, kera,
kelinci, babi, kuda, sapi, domba, anjing, tikus, ayam, dan kalkun (Kamil et al.
2001). Dalam keadaan normal, kadar hormon di dalam darah dipengaruhi oleh
kecepatan sintesis dan degradasi hormon serta kecepatan sekresi, sedangkan kerja
hormon dipengaruhi oleh keadaan reseptor pada sel target dan protein pengangkut
di dalam darah.
Ikatan hormon dengan reseptor (HR) pada permukaan membran sel akan
meningkatkan permiabilitas membran plasma sehingga meningkatkan
permiabilitas ion-ion dan zat-zat lain yang mempunyai konfigurasi sedemikian
rupa. Selain itu, HR dapat menyebabkan hiperpolarisasi membran plasma
sehingga memungkinkan pemasukan ion-ion dan zat-zat anorganik ke dalam sel.
Selanjutnya, perubahan ini akan diikuti dengan proses-proses dalam sitoplasma
dan juga modulasi enzim adenilat siklase yang akan merangsang sintesis cyclic
adenosine 3, 5–monophosphate (cAMP), yaitu caraka kedua (second messenger),
yang bekerja mempengaruhi fungsi sel (Djojosoebagio 1990; Manalu 2001;
Shahib 2001).
Meskipun ST telah diketahui sejak tahun 30-an, pada permulaan tahun
1937 para peneliti mendapatkan bahwa sapi-sapi yang disuntik dengan ekstrak ST
dari hipofisis sapi yang diperoleh dari rumah potong hewan (RPH)
memperlihatkan peningkatan produksi susu. Sampai dengan tahun 70-an,
kemajuan yang dicapai hanya sebatas aplikasi ST pada ternak untuk tujuan
komersial. Hal tersebut disebabkan oleh keterbatasan produksi ST. Untuk
memperoleh ST yang akan diberikan kepada seekor sapi dibutuhkan ekstrak
hipofisis yang berasal dari 200 ekor sapi (Hardjopranjoto 2001). Setelah 50 tahun,
pemakaian ST berkembang pesat khususnya ketika ditemukannya sistem
rekombinan, ST banyak digunakan untuk meningkatkan produksi ternak daging
dan susu (Kamil et al. 2001).
Dewasa ini berkembang hormon rekombinan antara lain recombinant
growth hormone (rbGH) atau recombinant soamtotropin (rbST). Salah satu
produk tersebut dibuat oleh Monsanto Corporation melalui rekayasa genetika,
yakni rbGH dengan nama dagang Posilac yang juga dikenal sebagai bST atau
15
dalam penggunaannya dewasa ini, yaitu bovine Somatotropin (bST), ovine
Somatotropin (oST), dan porcine Somatotropin (pST) (Hardjopranjoto 2001).
HIPOTALAMUS
GHRH SOMATOSTATIN
SOMATOTROF
AKSI LANGSUNG
EFEK ANTI INSULIN
METABOLISME KARBOHIDRAT LIPOLISIS
FFA DAN
GLISEROL GLUKOSA
AKTIVITAS DIABETOGENIK AKSI TAK LANGSUNG
HATI
PERTUMBUHAN TULANG SOMATOMEDIN
KHONDROSIT
LIPOGENESIS SINTESIS PROTEIN OTOT SEL LEMAK
SEL LEMAK PROLAKTIN
KHORIONIK SOMATOTROPIN INSULIN
MEKANISME UMPAN BALIK
(–) (+)
KELENJAR PITUITARI
SOMATOTROPIN
PEMBENTUKAN KARTILAGO
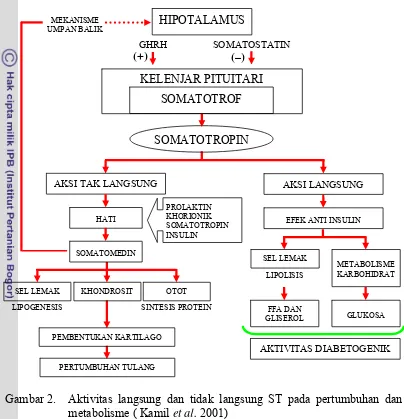
Gambar 2. Aktivitas langsung dan tidak langsung ST pada pertumbuhan dan metabolisme ( Kamil et al. 2001)
Somatotropin berpengaruh pada metabolisme tubuh yang pada gilirannya
akan merangsang proses produksi dan proses adaptasi homeostatis yang akan
menyediakan nutrien yang digunakan dalam proses tersebut (Vernon 1989).
Somatotropin mempunyai dua pengaruh utama. Pengaruh pertama berkaitan
dengan proses-proses produksi yang mungkin sebagian diperantarai oleh IGF-I.
Pengaruh yang kedua adalah penyediaan zat-zat makanan yang diperantarai oleh
ST sendiri, akan tetapi pengaruh ini bergantung pada keadaan fisiologis hewan
[image:33.595.108.512.155.574.2]mengakibatkan peningkatan akresi protein otot sementara selama laktasi
perlakuan ST dapat mengakibatkan kehilangan protein tubuh (Peel et al. 1981;
Akers 2002).
Somatotropin, IGF-I, dan Produksi susu
Telah disepakati bahwa somatotropin tidak saja berperan dalam
pengontrolan pertumbuhan, namun juga berperan dalam produksi susu (Manalu
1994). Somatotropin sering juga digunakan sebagai agen galaktopoietis yang
dapat meningkatkan produksi susu tanpa memberikan implikasi pada kesehatan
konsumen atau ternak itu sendiri (Tucker 2000).
Somatomedins atau IGF-I (Insulin-like growth factor I) mempunyai
rangkaian 70 asam amino dengan berat molekul 7 649 Dalton sedangkan IGF-II
mengandung 67 asam amino dengan berat molekul 7 471 Dalton. Urutan asam
amino IGF-I dan IGF-II memiliki kemiripan sekitar 70% dan juga mempunyai
struktur yang sama dengan proinsulin. Aktivitas biologis IGF bergantung pada
reseptor IGF yang spesifik yang ada pada sel target. Reseptor IGF ada dua
macam, tipe I (reseptor IGF-I) dan tipe II (reseptor IGF-II). Reseptor IGF-I
merupakan glikoprotein yang mempunyai dua subunit ekstra selluler dan dua
subunit B transmembran. Walaupun mempunyai persamaan affinitas ikatan,
reseptor IGF-I lebih menyukai ikatan dengan IGF-I diikuti dengan IGF-II dan
insulin. Tipe II atau reseptor IGF-II merupakan polipeptida rantai tunggal yang
disebut sebagai reseptor mannosa-6-fosfat. Secara struktural reseptor ini tidak
berkaitan dengan reseptor IGF-I atau insulin. Reseptor tipe II mengikat IGF-II
dengan afinitas yang lebih tinggi daripada IGF-I dan tidak mengenal insulin (
Kamil et al.2001).
Hipotesis klasik yang dikemukakan oleh peneliti terdahulu bahwa aksi ST
menstimulasi hati untuk memproduksi somatomedin atau insulin-like growth
faktor I (IGF-I) yang selanjutnya akan diangkut melalui darah untuk bereaksi
pada organ sasaran yang spesifik (Sharma et al. 1994). Reseptor ST banyak
dijumpai pada hati sehingga diyakini sebagian besar peneliti bahwa ST
memodulasi hati untuk mensintesis IGF-I. Insulin-like growth factor I (IGF-I)
mempunyai peranan penting dalam mengatur metabolisme sel mammalia,
pertumbuhan, dan diferensiasi. Insulin-like growth factor I (IGF-I) hampir
17
jaringan dapat mensintesisnya sehingga IGF-I bukan saja bereaksi secara endokrin
tetapi juga sebagai autokrin atau parakrin (Prosser & Mepham 1989; Sharma et al.
1994; Tucker 2000; Kamil et al. 2001) sehingga level plasma IGF-I merupakan
produksi kumulatif dari beberapa jaringan (Tucker 2000). Produksi IGF-I akut
sensitif terhadap status nutrien (Prosser & Mepham 1989).
Hampir sebagian besar hormon dalam aksi biologisnya mempunyai organ
target. Namun, tidak demikian halnya dengan ST. Hormon ini tidak memiliki
organ target yang spesifik, tetapi sebagian besar sel tubuh dapat merespons ST
(Djojosoebagio 1990). Efek ST pada peningkatan produksi susu sangat
kontradiktif karena pada jaringan kelenjar susu hampir tidak ditemukan reseptor
untuk ST (Prosser & Mepham 1989; Akers 2002), sedangkan untuk melakukan
aktivitasnya kehadiran reseptor spesifik sangat mutlak (Shahib 2001) sehingga
diyakini bahwa ST tidak mempunyai efek langsung pada fungsi sel sekretori
kelenjar susu (Prosser & Mepham 1989). Pernyataan tersebut didasarkan pada
hasil pengamatan in vitro bahwa pemberian infus ST langsung melalui pembuluh
darah arteri kelenjar susu mengakibatkan terjadi pengurangan efek ST pada
jaringan kelenjar susu dan stimulasi sekresi susu.
Hipotesis lain menyatakan bahwa aksi ST dimediasi oleh somatomedin,
khususnya IGF-I. Pernyataan ini didasari pengamatan secara in vitro bahwa efek
langsung IGF-I pada pertumbuhan kelenjar susu dan perkembangan sistem
transport glukosa pada sel-sel epitel kelenjar susu diyakini sebagai aksi ST dalam
kelenjar susu yang dimediasi oleh IGF-I. Bukti lain adalah bahwa konsentrasi
IGF-I plasma meningkat 3 sampai 4 kali pada sapi laktasi yang diberi injeksi ST
dalam jangka waktu singkat (short term). Peningkatan IGF-I jika dihitung
berdasarkan peningkatan aliran darah ternyata dapat mencapai 5 kali dari jumlah
IGF-I yang beredar dalam kelenjar susu (Prosser & Mepham 1989).
Hasil pengamatan in vivo tentang aksi IGF-I untuk meningkatkan sekresi
susu pada ternak kambing, dengan cara penyuntikan IGF-I pada arteri tertutup ke
dalam satu kelenjar dengan laju 1,1 ηmol/menit selama 6 jam, ternyata dapat
meningkatkan laju sekresi sebesar 30 ± 5% dibandingkan dengan yang tidak
mendapat infus hanya sebesar 15 ± 4% (Prosser & Mepham 1989). Konsentrasi
IGF-I dalam susu telah meningkat sebesar 32 ± 22% dibandingkan dengan yang
kelenjar susu dan peningkatan sekresi susu juga diamati secara in vitro ketika
ternak ruminansia kecil diinfus IGF-I melalui pembuluh darah yang langsung
menuju kelenjar susu (Sharma et al. 1994). Efek galaktopoietik ST mengantar
peningkatan sekresi IGF-I ke dalam susu, walaupun pada puncak produksi masih
terdapat dalam rentang fisiologis normal untuk ruminansia penghasil susu.
Somatotropin, Metabolisme, dan Produksi Susu
Penggunaan somatoropin pada sapi laktasi dapat memodifikasi hampir
seluruh aspek metabolisme baik melalui pengaruh langsung atau tidak langsung
(Vernon 1989). Pengaruh langsung khususnya dalam rangka penggalangan zat-zat
makanan, sedangkan pengaruh tidak langsung bukan berkaitan dengan
proses-proses produksi yang mungkin sebagian diperantarai oleh IGF-1. Kerja
somatoropin bergantung pada keadaan fisiologis hewan percobaan. Perlakuan ST
pada hewan yang sedang tumbuh akan mengakibatkan peningkatan akresi protein
otot, sedangkan pada hewan laktasi menyebabkan kehilangan protein tubuh (Peel
et al.1981).
Glukosa merupakan prekursor laktosa utama dan hampir 80% glukosa
darah pada sapi laktasi digunakan untuk sintesis laktosa. Pada ternak ruminansia,
konsentrasi glukosa dalam sistem sirkulasi berasal dari ekstraksi propionat dalam
hati (Baldwin & Smith 1983; Annison et al. 1984; Collier 1985). Laktosa atau
gula susu adalah karbohidrat susu berupa disakarida yang tersusun dari satu
molekul glukosa dan satu molekul galaktosa yang bersatu dalam ikatan karbon 1-4
sebagai β-galaktosida (Larson 1985). Kadar laktosa dalam susu relatif konstan dan
dipertahankan pada kisaran yang tetap. Laktosa merupakan osmoregulator dan
determinan untuk volume susu sehingga produksi susu akan sejalan dengan
produksi laktosa.
Jalur sintesis laktosa adalah sebagai berikut :
Glukosa + ATP --- (1) Æ Glukosa -6-fosfat + ADP
Glukosa-6-fosfat --- (2) Æ Glukosa-1-fosfat
Uridine trifosfat + glukosa-1-P -- (3) Æ Uridine difosfat-glukosa + Pirofosfat
Uridine difosfat glukosa --- (4) Æ Uridine Difosfat-galaktosa
19
(1) Heksokinase
(2) Fosfoglukomutase
(3) Uridine difosforil glukosa pirofosforilase
(4) Uridine difosforil galaktosa-4-epimerase
(5) Laktosa sintetase ( terdiri atas galaktosil transferase dan α-laktalbumin)
Protein susu disintesis dari asam amino yang beredar dalam darah hasil
penyerapan dari saluran pencernaan maupun hasil perombakan protein tubuh.
Glukosa dan beberapa sumber nitrogen diperlukan untuk sintesis asam amino di
kelenjar susu ruminansia (Annison et al. 1984; Collier 1985).
Sintesis lemak susu terjadi dalam sitoplasma sel-sel kelenjar susu. Pada
ternak ruminansia, asetat dan β-OH butirat (hasil metabolisme asam butirat),
selain untuk sumber energi juga digunakan untuk sintesis lemak di dalam kelenjar
susu oleh enzim asetil KoA karboksilase (Annison et al. 1984; Collier 1985).
Oksidasi glukosa melalui siklus pentosa oleh enzim glukosa 6-p dehidrogenase
menghasilkan NADPH yang diperlukan untuk sintesis asam lemak di kelenjar
susu.
Laju sintesis komponen air susu terutama laktosa, lemak, dan protein akan
semakin berkurang jika ketersediaan substrat di kelenjar susu semakin sedikit
sehingga dalam keadaan seperti ini sejumlah besar cadangan zat-zat makanan
akan dimobilisasi untuk mempertahankan produksi susu (Bines & Hart 1982).
Neraca nutrien negatif akan terjadi bila tingkat konsumsi tidak memenuhi
kebutuhan untuk sintesis susu, dan keadaan ini sering dijumpai pada awal laktasi
karena hampir 65% sampai 83% dari metabolisme energi akan diubah untuk
sintesis susu selama laktasi (Gardner & Hogue 1985).
Hasil mobilisasi asam lemak menyebabkan penumpukan asetil KoA yang
tak dapat memasuki siklus asam sitrat sehingga akan diubah menjadi benda-benda
keton seperti aseton, β-OH butirat, maupun asetoasetat sebagai hasil kondensasi
2 molekul asetil KoA (Foot et al. 1984). Pada ternak yang berada dalam neraca
nitrogen negatif akan terjadi katabolisme protein yang ditandai dengan
peningkatan konsentrasi nitrogen urea darah (Peel et al. 1981).
Produksi susu merupakan fungsi jumlah sel-sel sekretoris yang aktif dan
hanya akan bergantung pada jumlah sel-sel sekretoris yang aktif dan tersedianya
subtrat (zat-zat makanan) untuk sintesis komponen air susu (Davis & Collier
1983). Lebih lanjut dikemukakan Sumaryadi & Manalu (1996) bahwa dengan
meningkatkan aliran substrat, sel-sel sekretoris akan lebih aktif sehingga keadaan
tersebut diduga akan dapat memperlambat laju involusi sel-sel sekretoris.
Proses modulasi aliran substrat ke kelenjar susu sangat ditentukan oleh
konsentrasi substrat dan laju aliran darah ke kelenjar susu. Substrat atau zat-zat
makanan yang berada dalam sistem sirkulasi berasal dari penyerapan dari sistem
saluran pencernaan (Davis & Collier 1983) dan mobilisasi cadangan energi tubuh
(Collier 1985; Vernon 1989), yang selanjutnya masuk ke dalam sel-sel sekretori
dengan sistem transportasi melalui pengaturan hormonal.
Hormon-hormon metabolisme, melalui pengaktifan beberapa enzim pada
saat laktasi, akan meningkatkan aktivitas sintesis komponen susu dalam sel-sel
sekretoris yang telah siap bersama semua perangkat enzimnya untuk merangkai
substrat berupa glukosa, asam amino, asam lemak, dan gliserol secara berturut
menjadi laktosa, protein (kasein), dan lemak untuk menghasilkan susu (Anderson
1985).
Penggunaan bST dalam memacu produksi akan menyebabkan terjadinya
adaptasi metabolik yang sangat dinamis dan sangat variatif sesuai dengan status
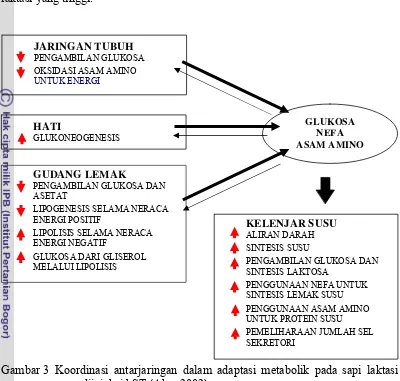
hewan yang bersangkutan. Koordinasi antarjaringan dalam regulasi homeorhetik
stimulasi bST pada sintesis susu dapat dilihat pada Gambar 3. Kehadiran reseptor
ST pada hati (Pocious & Herbein 1986) menyebabkan pemberian ST akan
berpengaruh langsung pada metabolisme glukosa, dan ST pada situasi ini mampu
bekerja mirip insulin, khususnya dalam meningkatkan glukosa darah, sementara
dalam keadaan lain ST akan bersifat antiinsulin dengan cara mengurangi glukosa
dengan mengorbankan cadangan lemak (Manalu 1994).
Perlakuan bST akan menyebabkan perubahan dalam metabolisme
karbohidrat karena terjadinya peningkatan kebutuhan glukosa yang tinggi untuk
sintesis susu. Dengan demikian, terjadi peningkatan produksi glukosa hati dan
terjadi penurunan oksidasi oleh jaringan tubuh dalam upaya meningkatkan
glukosa untuk sintesis susu. Pada hewan ruminan, produk fermentasi rumen
adalah volatile fatty acid (VFA) dan hanya sedikit persentase (15%) pada glukosa
21
glukoneogenesis hati yang mana produksinya dapat bertambah 3 kg/hari pada sapi
laktasi yang tinggi.
KELENJAR SUSU
ALIRAN DARAH SINTESIS SUSU
PENGAMBILAN GLUKOSA DAN SINTESIS LAKTOSA
PENGGUNAAN NEFA UNTUK SINTESIS LEMAK SUSU PENGGUNAAN ASAM AMINO UNTUK PROTEIN SUSU PEMELIHARAAN JUMLAH SEL SEKRETORI
HATI
GLUKONEOGENESIS
JARINGAN TUBUH
PENGAMBILAN GLUKOSA OKSIDASI ASAM AMINO
UNTUK ENERGI
GUDANG LEMAK
PENGAMBILAN GLUKOSA DAN ASETAT
LIPOGENESIS SELAMA NERACA ENERGI POSITIF
LIPOLISIS SELAMA NERACA ENERGI NEGATIF
GLUKOSA DARI GLISEROL MELALUI LIPOLISIS
GLUKOSA NEFA ASAM AMINO
Gambar 3 Koordinasi antarjaringan dalam adaptasi metabolik pada sapi laktasi yang diinjeksi bST (Aker 2002)
Pada sapi yang mendapat injeksi bST akan terjadi peningkatan
glukoneogenesis hati, yang paling tidak menjelaskan kemampuan antiinsulin untuk menghambat glukoneogenesis. Glukosa akan digunakan sebagai substrat
untuk sintesis laktosa di dalam kelenjar susu, dan pada sapi laktasi dengan
produksi yang tinggi hampir mendekati 85% total glukosa digunakan untuk
sintesis susu. Injeksi bST dapat meningkatkan pemanfaatan netto glukosa pada
jaringan kelenjar susu dan menurunkan penggunaan glukosa pada jaringan
nonkelenjar susu. Adaptasi metabolik ini cukup untuk memenuhi kebutuhan
glukosa untuk sintesis susu (Akers 2002).
Menurut Cohick et al. (1989) bahwa pada ruminansia, peningkatan
kehilangan karbon secara irrevesibel mempunyai implikasi peningkatan glukosa,
[image:39.595.112.512.115.496.2]peningkatan ini bisa disebabkan oleh peningkatan glikogenolisis, akan tetapi
dalam tempo beberapa hari keadaan itu pasti disebabkan oleh glukoneogenesis.
Beberapa hormon yang terlibat dalam metabolisme dalam tubuh adalah
tiroksin, insulin, prolaktin, dan somatotropin (McDonald & Pineda 1980; Collier
1984; Bauman et al. 1985) dan beberapa hormon yang turut andil dalam
metabolisme mineral dalam penyediaan aliran mineral ke kelenjar susu di
antaranya, paratiroid, kalsitonin, dan vitamin D (McDonald & Pineda 1980;
Collier et al. 1984; Tucker 1985).
Tiroksin (triiodotironin dan tetraiodotironin) berperan sangat vital dalam
penyediaan energi ATP (McDonald & Pineda 1980; Ultiger 1987) bagi perakitan
glukosa, asam amino, asam lemak, dan gliserol menjadi glikogen, protein, dan
lemak. Hormon tiroksin telah diketahui dapat mempengaruhi aktivitas
metabolisme pada kelenjar susu (Sumaryadi & Manalu 1995b).
Kortisol berfungsi dalam memobilisasi glukosa, asam amino, dan asam
lemak untuk tujuan sintesis pada jaringan dan menghambat penggunaan zat-zat
makanan untuk tujuan oksidasi dalam sel (McDonald & Pineda 1980; Manalu &
Sumaryadi 1995b) dengan merombak cadangan energi tubuh yang akan
dibebaskan ke dalam sistem sirkulasi untuk mendukung proses sintesis di kelenjar
susu (Annison et al. 1984).
Insulin mempunyai peran dalam mengangkut glukosa, asam amino, dan
asam lemak ke dalam sel (Bines & Hart 1982). Namun, insulin tidak memiliki
reseptor pada sel-sel kelenjar susu sehingga tidak menunjukkan pengaruh pada
pengangkutan substrat ke dalam sel-sel sekretoris kelenjar susu (Manalu 1994).
Prolaktin sangat spesifik dan selalu dikaitkan dengan produksi susu,
namun berperan hanya sebatas mengaktifkan (menginisiasi) bukan memelihara
atau mempertahankan sintesis air susu (Forsyth 1986).
Somatotropin, Kesehatan Ternak, dan Keamanan Pangan
Sebelumnya telah diramalkan bahwa penggunaan bST akan menimbulkan
pengaruh yang membahayakan bagi kesehatan sapi yang mengakibatkan
permasalahan ketosis, milk fever, dan “burn-out” (Manalu 2001).
Injeksi bST pada sapi perah menunjukkan terjadinya perubahan
23
seiring dengan peningkatan produksi (Akers 2002). Peningkatan pengaliran
subtrat ke kelenjar susu membawa konsekuensi peningkatan aliran darah, namun
selama penggunaan bST ini tidak nyata berpengaruh pada pengukuran klinis
seperti suhu tubuh ataupun pernapasan (Eppard et al. 1987; Manalu et al. 1991),
tapi denyut nadi tampak sedikit meningkat khususnya pada pemakaian dosis
tinggi (Soderholm et al. 1988). Perubahan-perubahan kimia darah yang terjadi
selama perlakuan bST umumnya berkaitan dengan peningkatan produksi dan tetap
berada dalam kisaran normal yang dapat diterima secara klinis. Angka
hemoglobin dan nilai hematokrit termasuk dalam kategori ini, dan kelihatannya
penurunan tersebut terlalu kecil untuk menggambarkan anemia fungsional
(Manalu 1994).
Dari hasil beberapa penelitian yang dilakukan di Amerika, pemakaian
hingga dosis 40,5 mg/hari tidak menunjukkan kejadian mastitis (Manalu 1994).
Injeksi bST secara berlebihan dapat mendorong terjadinya pemakaian lemak yang
berlebihan sehingga menghasilkan asam asetoasetat dalam hati yang akan
dikeluarkan ke dalam cairan tubuh dan menyebabkan terjadinya penyakit
metabolik seperti ketosis (Harjopranjoto 2001).
Injeksi bST tidak menghasilkan pengaruh yang konsisten pada kejadian
mastitis klinis atau hitungan sel somatis (Phipps 1989). Pada suatu percobaan
selama dua tahun di U.K, hitungan sel somatis untuk kontrol dan sapi perlakuan
lebih rendah dibandingkan rata-rata kejadian secara nasional (Phipps 1989).
Bahkan, bovine somatotropin (bST) secara positif bisa mempengaruhi pemulihan
fungsi kelenjar susu setelah kejadian mastitis (Phipps 1989).
Hasil penelitian dari Cornell University menunjukkan bahwa sapi-sapi
yang diinjeksi bST setiap hari, mulai hari ke-84 sampai hari ke-272 postpartum,
ternyata tidak menunjukkan adanya gangguan pada kesehatan maupun status
reproduksi (Hariadi et al. 2001). Lebih lanjut diungkapkan bahwa hasil tersebut di
atas diperoleh dari sapi-sapi yang dikelola dengan baik, namun sampai sejauh
mana injeksi bST bila diberikan pada sapi-sapi yang dikelola secara sederhana
masih belum diketahui (Hariadi et al. 2001).
Hasil diskusi The Standing Senate Committee on Agriculture and Forestry,
Canada pada tahun 1999 yang dikutip Soeharsono (2001) menyatakan bahwa
menyebabkan adanya metabolit IGF-I yang disebut miotropin pada susu yang
dapat menyebabkan mastitis pada sapi perah. Lebih lanjut dikemukakan bahwa
penyuntikan bST/rbGH di California (sebelum tahun 1995) dapat menurunkan
harapan hidup sapi perah, meningkatkan risiko penyakit, penurunan bobot hidup,
kadang-kadang menyebabkan sapi jadi infertile dan lebih peka terhadap mastitis.
Dampak injeksi bST pada keamanan pangan sudah banyak dipublikasikan,
namun masih menyisakan kontroversi di antara peneliti. Injeksi bST pada sapi
perah dapat meningkatkan produksi susu secara tajam, namun bagaimana
pengaruhnya pada orang yang mengkonsumsi produk susu tersebut masih perlu
kajian lebih lanjut. Kandungan hormon, faktor pertumbuhan, dan peptida lainnya
telah banyak diteliti. Konsentrasi ST pada susu sapi hampir sama dengan susu
manusia masing-masing kurang dari 1 ηg/mL. Konsentrasi IGF-I, suatu mediator
untuk aksi ST berkisar antara 1,5 sampai 8 ηg/mL pada susu sapi dan
1 sampai 3 ηg/mL pada susu manusia, konsentrasi tersebut akan meningkat pada
penggunaan bST antara 2 sampai 5 ηg/mL, sedangkan konsentrasi IGF-II tidak
meningkat (Budinuryanto 2001). Lebih lanjut diungkapkan adanya kontroversi
antarpeneliti, khususnya yang berkaitan dengan jumlah IGF-I dan kanker.
Insulin-like growth factor I dan II diidentifikasi merupakan regulator pertumbuhan
autokrin dan endokrin yang ikut memacu berbagai karsinoma. Insulin- like growth
factor I (IGF-I) memainkan peranan sangat penting pada proliferasi sel kanker
pankreas, serta berperan dalam metabolisme glukosa dalam tumor, bahkan
pasteurisasi yang menginaktifkan dan merusak bST tidak mempengaruhi IGF