KLONING DAN EKSPRESI GEN PENYANDI ASIL
HOMOSERIN LAKTON LAKTONASE DARI
Bacillus cereus INT1c DAN Bacillus thuringiensis SGT3g
ANJA ASMARANY R.
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA
Dengan ini saya menyatakan bahwa tesis berjudul “Kloning dan ekspresi gen penyandi asil homoserin lakton laktonase dari Bacillus cereus INT1c dan Bacillus thuringiensis SGT3g” adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apapun kepada perguruan tinggi manapun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor
Bogor, Januari 2015
RINGKASAN
ANJA ASMARANY R. Kloning dan ekspresi gen penyandi asil homoserin lakton laktonase dari Bacillus cereus INT1c dan Bacillus thuringiensis SGT3g. Dibimbing oleh IMAN RUSMANA dan ARIS TRI WAHYUDI.
Quorum sensing (QS) merupakan mekanisme komunikasi di antara sel bakteri yang diperantarai oleh molekul sinyal asil homoserin lakton (AHL) dan mekanismenya bergantung pada kepadatan jumlah sel. Pada umumnya bakteri fitopatogen menggunakan mekanisme QS untuk mengekspresikan gen virulensi pada saat menginfeksi tanaman. Penghambatan ekspresi gen virulen tersebut dapat dilakukan dengan mendegradasi senyawa AHL menggunakan AHL-laktonase.
AHL-laktonase merupakan kelompok enzim yang berfungsi untuk menghidrolisis cincin lakton pada molekul AHL. Enzim ini umumnya ditemukan pada kelompok Bacillus dan disandikan oleh gen aiiA. Isolasi bakteri dari sampel tanah dan daun asal lahan pertanian di Jawa telah berhasil mendapatkan dua isolat yang mampu menghasilkan AHL-laktonase dengan aktivitas yang tinggi. Penelitian ini bertujuan untuk mengkloning gene AHL-laktonase dari isolat Bacillus cereus INT1c and Bacillus thuringiensis SGT3g.
Amplifikasi gen AHL-laktonase dari kedua isolat dengan menggunakan primer aiiA telah berhasil mendapatkan fragmen amplikon DNA berukuran 800 bp. Kedua isolat mempunyai gen aiiA dengan sekuen lengkap pada orf 2 yang menyandikan 250 asam amino. Analisis sekuen asam amino menggunakan BLAST-X menunjukkan bahwa asam amino dari B.cereus INT1c mempunyai homologi sebesar 98% dibandingkan dengan B.cereus (nomor akses WP.000216573.1), sedangkan B.thuringiensis SGT3g mempunyai homologi sebesar 99% dibandingkan dengan B.thuringiensis serovar aizawai (nomor akses AEY70474.1).
Gen aiiA dari kedua isolat diekspresikan di dalam sel E.coli BL21(DE3). Pemotongan plasmid rekombinan dengan menggunakan BamHI dan NdeI menunjukkan adanya fragmen DNA berukuran 5708 bp (pET15b) dan 800 bp (DNA sisipan) pada gel agarosa 1.5%. Hasil ini membuktikan bahwa plasmid rekombinan telah diperbanyak di dalam sel transforman. Induksi ekspresi gen dilakukan ketika fase eksponensial menggunakan IPTG. Penambahan 1 mM IPTG ketika OD600 kultur mencapai 0.6 dapat menghasilkan protein AiiA dengan aktivitas tinggi. Sementara itu rekombinan E.coli dengan OD600 0.8 tidak menunjukkan adanya aktivitas degradasi. Hasil ini mengindikasikan bahwa protein AiiA dari E.coli rekombinan dapat mendegradasi senyawa AHL dari Chromobacterium violaceum sehingga jumlah AHL tidak mencapai quorum untuk menginduksi ekspresi gen yang bertanggung jawab untuk produksi violacein. Hasil analisis SDS-PAGE menunjukkan bahwa protein AiiA dari kedua isolat mempuyai berat 28.77 kDa.
SUMMARY
ANJA ASMARANY R. Cloning and gene expression of acyl homoserine lactone lactonase from Bacillus cereus INT1c and Bacillus thuringiensis SGT3g. Supervised by IMAN RUSMANA and ARIS TRI WAHYUDI.
Quorum sensing is a mechanism of communication between bacterial cells mediated by signal molecules acyl homoserine lactone (AHL) and the mechanism depends on density of bacterial cells. Phytopathogenic bacteria use quorum sensing mechanisms to express virulence genes when infecting the plants. Inhibition of virulence genes expression can be done by degrading the AHL succeeded to get two isolates that had ability to produce AHL-lactonase with high activity. The aim of this study was to clone AHL-lactonase genes from Bacillus cereus INT1c and Bacillus thuringiensis SGT3g.
AHL-lactonase gene amplification from the isolates using aiiA primer was succeeded to get 800 bp of DNA amplicont fragment. Both of isolate had aiiA gene with complete sequnces on orf 2 which encoding 250 amino acids. Amino acid sequences analysis using BLAST-X indicated that B.cereus INT1c amino acid had 98% of maximum identity similarity with B.cereus (access number WP.000216573.1), while B.thuringiensis SGT3g had 99% maximum identity similarity with B.thuringiensis serovar aizawai (access number AEY70474.1).
The aiiA genes from the two isolates were expressed in E.coli BL21(DE3) pLysS cells. Recombinant plasmid digestion by BamHI and NdeI showed the DNA fragment of 5708 bp (pET15b) and 800 bp (DNA inserts) size. These results are evidence that the recombinant plasmid was propagated in the transformant cells. Induction of the gene expression was done in the exponential growth phase using IPTG. 1 mM IPTG addition when the culture OD600 reached 0.6 could produce AiiA proteins with high activity. However, recombinant E.coli with 0.8 OD600 did not show AHL degradation activity. These results indicated that AiiA protein from recombinant E.coli could degrade AHL compounds produced by Chromobacterium violaceum so that the AHL amount did not reach a quorum to induce expression of the genes that responsible in violacein production. The results from SDS-PAGE analysis showed that AiiA proteins from both isolates have a molecular weight 28.77 of kDa.
©
Hak Cipta Milik IPB, Tahun 2015
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB
KLONING DAN EKSPRESI GEN PENYANDI ASIL
HOMOSERIN LAKTON LAKTONASE DARI
Bacillus cereus INT1c DAN Bacillus thuringiensis SGT3g
ANJA ASMARANY R.
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains
pada
Program Studi Mikrobiologi
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
Judul Tesis : Kloning dan Ekspresi Gen Penyandi Asil Homoserin Lakton Laktonase dari Bacillus cereus INT1c dan Bacillus thuringiensis SGT3g
Nama : Anja Asmarany R. NIM : G351120111
Disetujui oleh Komisi Pembimbing
Dr Ir. Iman Rusmana, MSi Ketua
Prof Dr Aris Tri Wahyudi, MSi Anggota
Diketahui oleh
Ketua Program Studi Mikrobiologi
Prof Dr Anja Meryandini, MS
Dekan Sekolah Pascasarjana
Dr Ir Dahrul Syah, MScAgr
PRAKATA
Puji dan syukur penulis panjatkan kepada Allah SWT atas segala karunia-Nya sehingga karya ilmiah ini berhasil diselesaikan. Tema yang dipilih dalam penelitian yang dilaksanakan sejak bulan Oktober 2013 sampai November 2014 ini ialah Kloning dan ekspresi gen penyandi asil homoserin lakton laktonase dari Bacillus cereus INT1c dan Bacillus thuringiensis SGT3g.
Terima kasih penulis ucapkan kepada Dr. Ir. Iman Rusmana, MSi sebagai ketua komisi pembimbing dan Prof. Dr. Aris Tri Wahyudi, MSi sebagai anggota komisi pembimbing, yang telah banyak memberikan nasehat, saran, motivasi, waktu konsultasi, serta solusi dari setiap permasalahan yang dihadapi penulis selama melaksanakan penelitian dan penyusunan karya ilmiah ini. Selain itu
penulis ucapkan terima kasih kepada penguji luar komisi Dr. Ir. I Made Artika, M.App.Sc dan Prof. Dr. Anja Meryandini, MS selaku Ketua Program Studi Mikrobiologi IPB, yang telah memberikan motivasi selama studi dan masukan pada saat ujian sidang tesis. Penulis mengucapkan terima kasih kepada DIKTI melalui Beasiswa Unggulan 2012/2013, terima kasih atas kepercayaannya untuk memberikan beasiswa kuliah selama menempuh pendidikan pascasarjana di IPB sehingga penelitian yang penulis lakukan dapat terlaksana dengan baik.
Penulis juga mengucapkan terima kasih kepada Ibu Heni dan Bapak Jaka selaku staf Laboratorium Mikrobiologi IPB, Dina, Vita, Asrianto, Nezharia, Wulan, Hari, Rahmi, Mahyar, Ayun, Asril, Sipriyadi, Ernin, Gege, Hamtini, Randi, Antri, Mona, Mei, Yeni, Annisa, serta seluruh teman-teman di Laboratorium Mikrobiologi IPB, atas dukungan, motivasi, dan bantuannya selama penelitian ini. Ucapan terima kasih tak terhingga juga penulis ucapkan kepada orang tua tercinta Ayah Rustamaji, Ibu Endang Sri Sulastri, dan ketiga adikku tersayang Muhammad Ibrahim Annur, Ahmad Fauzi Ghouzts, Khosyi Larasati, serta sahabat-sahabatku tersayang, atas doa, dukungan, kasih sayang, dan semangat yang diberikan. Terima kasih untuk teman-teman seperjuangan di Pascasarjana Mikrobiologi IPB angkatan 2012 serta seluruh pihak yang telah memberikan doa dan dukungannya, penulis ucapkan terima kasih.
Semoga karya ilmiah ini bermanfaat.
Bogor, Januari 2015
DAFTAR ISI
DAFTAR TABEL viii
DAFTAR GAMBAR viii
DAFTAR LAMPIRAN viii
PENDAHULUAN 1
Latar Belakang 1
Perumusan Masalah 2
Tujuan Penelitian 2
Manfaat Penelitian 2
Ruang Lingkup Penelitian 2
TINJAUAN PUSTAKA 3
Quorum Sensing 3
Asil Homoserin Lakton (AHL) 4
AHL-Laktonase 5
Ekspresi Gen Pada Escherichia coli 6
METODE 8
Kerangka Penelitian 8
Waktu dan Tempat Penelitian 8
Isolasi DNA 9
Amplifikasi Gen Penyandi AHL-Laktonase 9
Pembuatan Sel Kompeten 10
Subkloning Gen Penyandi AHL-Laktonase 10
Isolasi Plasmid Rekombinan 12
Restriksi Plasmid Rekombinan dengan EcoRI 12
Amplifikasi Plasmid Rekombinan 12
Analisis Sekuen Gen AHL-Laktonase Rekombinan 12 Ekspresi Gen AHL-Laktonase dan Bioesai 13
SDS-PAGE 13
HASIL DAN PEMBAHASAN 15
Hasil 15
SIMPULAN DAN SARAN 26 2 Sistem quorum sensing pada bakteri Gram negatif dan Gram positif 4 3 Struktur Asil Homosern Lakton (AHL) 4 4 Reaksi Biosintesis asil homoserin lakton (AHL) 5 5 Mekanisme degradasi AHL oleh AHL-laktonase 6 6 Peta dan daerah ekspresi pET15b (Novagen) 7 13 Restriksi Plasmid Rekombinan dengan EcoRI 15 14 Amplifikasi plasmid rekombinan dengan primer T7 dan SP6 16 15 Pohon filogenetik dari asam amino AHL-laktonase yang dikonstruksi menggunakan metode neighbor-joining. 17 16 Hasil analisis Open Reading Frame (ORF) 17
17 Hasil analisis sekuen gen aiiA dan asam amino dari bakteri 18
B.cereus INT1c dengan ORF finder 18 Hasil analisis sekuen gen aiiA dan asam amino dari 19
bakteri B.thuringiensis SGT3g dengan ORF finder 19 Hasil analisis domain dari isolat B.thuringiensis SGT3g 20
20 Hasil analisis domain dari isolat B.cereus INT1c 20
21 Prediksi struktur 3 dimensi protein AHL-laktonase 21 22 Prediksi titik isoelektrik (pI) dan berat molekul (Mw) AHL-laktonase dari bakteri B.thuringiensis SGT3g 21
23 Prediksi titik isoelektrik (pI) dan berat molekul (Mw) AHL-laktonase dari bakteri B.cereus INT1c 21
24 Hasil restriksi plasmid rekombinan dalam E.coli BL21(DE3) dengan enzim restriksi BamHI dan NdeI 22
25 Bioesai E.coli rekombinan dengan bakteri C.violaceum sebagai biokontrol 22
DAFTAR LAMPIRAN
1 Sekuen gen aiiA dari isolat B.cereus INT1c 33
2 Sekuen gen aiiA dari isolat B.thuringiensis SGT3g 33
3 Komposisi gel pemisah dan gel penahan 34
4 Metode pewarnaan dengan perak nitrat 34
5 Perhitungan nilai Rf marker 35
6 Kurva standar marker 35
7 Perhitungan berat molekul sampel 36
PENDAHULUAN
Latar Belakang
Quorum sensing merupakan mekanisme komunikasi diantara sel bakteri secara interseluler, bergantung pada kepadatan jumlah sel dan berperan penting dalam regulasi ekspresi gen. Bakteri memanfaatkan quorum sensing untuk regulasi pembentukan biofilm, faktor virulen, sintesis antibiotik, sporulasi, dan bioluminesen (Galloway et al. 2011). Quorum sensing diperantarai oleh suatu molekul sinyal ekstraseluler yaitu autoinduser. Molekul autoinduser akan dideteksi dan direspon oleh sel ketika konsentrasinya di lingkungan tinggi. Bakteri menghasilkan autoinduser dengan jenis yang bervariasi. Bakteri Gram negatif menghasilkan autoinduser yang disebut N-Acyl Homoserine Lactone (AHL) (Hentzer et al. 2002).
Mekanisme quorum sensing digunakan oleh bakteri fitopatogen untuk mengekspresikan gen virulen pada saat menginfeksi tanaman (Soto et al. 2006). Strategi pengendalian bakteri fitopatogen pada umumnya dilakukan dengan menggunakan pestisida maupun senyawa antimikrob. Namun penggunaan senyawa tersebut secara terus-menerus akan menimbulkan sifat resisten pada bakteri. Solusi alternatif yang dapat dilakukan untuk mengendalikan bakteri fipatogen yaitu menggunakan quorum quenching. Hentzer dan Givskov (2003) menyatakan bahwa aplikasi quorum quenching sangat menguntungkan karena dapat mengurangi munculnya sifat resisten pada bakteri. Quorum quenching tidak mempengaruhi kelangsungan hidup sel secara individual, tetapi lebih kepada sifat virulen dari populasi secara keseluruhan. Konsentrasi molekul AHL merupakan faktor utama yang mempengaruhi ekspresi gen virulen sehingga degradasi molekul AHL menjadi sasaran utama dari quorum quenching. Enzim yang mempunyai kemampuan untuk mendegradasi molekul AHL adalah AHL-laktonase (Molina et al. 2003).
Isolasi dan karakterisasi bakteri penghasil AHL-laktonase telah dilakukan menggunakan sampel tanah dan daun asal lahan pertanian Jawa. Sebanyak 12 isolat dari 261 isolat yang diperoleh positif memiliki kemampuan untuk mendegradasi AHL. Dua isolat dengan aktivitas AHL-laktonase terbaik yaitu B. cereus INT1c dan B. thuringiensis SGT3g (Afiah 2011). Kedua isolat yang diperoleh masih belum diketahui karakter gennya sehingga perlu diteliti lebih lanjut dengan melakukan kloning.
2
Perumusan Masalah
1.Bakteri patogen menggunakan mekanisme QS untuk mengeskpresikan gen virulen.
2.Penggunaan pestisida dapat menimbulkan sifat resisten pada bakteri
3.Bacillus cereus INT1c dan Bacillus thuringiensis SGT3g penghasil enzim QQ berhasil diisolasi dari tanah asal lahan pertanian Jawa.
4.Kedua isolat mempunyai aktivitas degradasi terhadap molekul sinyal AHL sehingga berpotensi untuk dijadikan sebagai agens biokontrol.
5.Penelitian mengenai isolasi dan kloning gen penyadi AHL-laktonase masih jarang dilakukan di Indonesia.
Tujuan Penelitian
Penelitian ini bertujuan untuk mengklon gen penyandi AHL-laktonase dari Bacillus cereus INT1c dan Bacillus thuringiensis SGT3g serta mengekspresikannya ke dalam sel E.coli BL21(DE3).
Manfaat Penelitian
Hasil dari penelitian ini yaitu bakteri pendegradasi asil homoserin lakton (AHL), diharapkan dapat digunakan sebagai agen biokontrol dalam pengendalian bakteri patogen pada tanaman. Aplikasi bakteri pendegradasi AHL diharapkan dapat menciptakan sistem pertanian organik dengan mengurangi penggunaan pestisida yang dapat menimbulkan sifat resisten pada bakteri.
Ruang Lingkup Penelitian
Ruang lingkup dalam penelitian ini meliputi isolasi DNA bakteri, karakterisasi serta ekspresi gen penyandi AHL-laktonase. Karakterisasi gen dilakukan melalui tahapan amplifikasi dengan primer aiiA, transformasi ke dalam sel E.coli DH5α, isolasi dan pemotongan plasmid dengan enzim EcoRI, serta amplifikasi menggunakan primer universal T7 dan SP6. Analisis ekspresi gen meliputi beberapa tahapan yaitu transformasi gen aiiA ke dalam sel E.coli BL21(DE3), restriksi dengan enzim BamHI dan NdeI, serta bioasai aktivitas protein rekombinan dengan bakteri C.violaceum sebagai biokontrol.
3
TINJAUAN PUSTAKA
Quorum Sensing
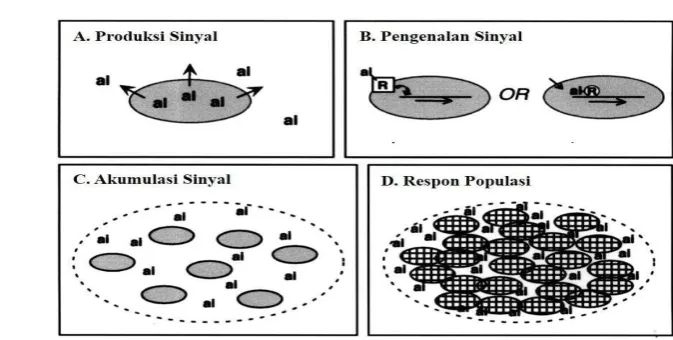
Quorum sensing (QS) merupakan mekanisme komunikasi diantara sel bakteri yang bergantung pada kepadatan populasi sel dan melibatkan sintesis serta sekresi senyawa molekul sinyal yang disebut autoinduser (Galloway et al. 2011; Waters dan Bassler 2005). Ngai (2001) menyatakan bahwa mekanisme QS pada bakteri terjadi melalui beberapa tahapan utama diantaranya sebagai berikut (Gambar 1):
a. Produksi autoinduser oleh sel bakteri, kemudian disekresikan ke lingkungan
b. Pengenalan molekul sinyal melalui interaksi dengan protein regulator c. Akumulasi sinyal seiring dengan peningkatan kepadatan populasi sel
bakteri
d. Respon populasi bakteri ketika autoinduser telah mencapai jumlah quorum
.
Gambar 1 Mekanisme quorum sensing pada bakteri
Konsentrasi autoinduser di lingkungan berkaitan dengan kepadatan populasi sel bakteri (Federle dan Blasser 2003). Senyawa autoinduser ini disintesis secara intraseluler oleh sel bakteri dan disekresikan ke lingkungan selama masa pertumbuhannya. Hal ini menyebakan konsentrasi molekul sinyal di dalam sel dan di lingkungan meningkat hingga jumlahnya mencapai ambang batas konsentrasi (quorum). Pada saat senyawa autoinduser mencapai jumlah quorum, autoinduser akan mengaktifkan ekspresi gen-gen tertentu dalam sel bakteri (Ryan dan Dow 2008; Weber dan Buceta 2013; Lade et al. 2014).
4
positif molekul sinyal berupa oligonukleotida yang merupakan modifikasi dari asam amino tertentu. Autoinduser ini sering disebut sebagai autoinducing peptides (AIP). Deteksi senyawa oligopeptida terjadi melalui dua komponen sinyal tranduksi. AIP yang telah berikatan dengan sensor kinase akan menyebabkan terjadinya autofosforilasi. Fosfat yang dihasilkan selanjutnya ditransfer pada protein respon regulator dan mengaktifkan transkripsi gen target (xyz) (Federle dan Bassler 2003).
Gambar 2 Sistem quorum sensing pada bakteri Gram negatif (a) dan positif (b)
Asil Homoserin Lakton (AHL)
Sebanyak lebih dari 70 genus bakteri Gram negatif dari kelompok proteobakteria dilaporkan menghasilkan senyawa AHL sebagai sinyal untuk berkomunikasi. Struktur AHL tersusun atas cincin homoserin lakton dan rantai samping asil dengan panjang yang bervariasi yaitu antara 4 sampai 18 rantai karbon (Gambar 3). AHL dengan rantai terpendek dan terpanjang yang dapat ditemukan di alam yaitu C4-HSL dan C18-HSL. Karakteristik molekul AHL dapat dipengaruhi oleh beberapa faktor diantaranya jumlah rantai asil dan jenis gugus substitusi pada molekulnya. Rantai asil dari AHL dapat bersifat jenuh maupun tak jenuh serta dapat mempunyai substitusi berupa gugus fungsional hidroksil atau oxo pada atom C nomor 3 (Decho et al. 2011; Tang dan Zhang 2014). Beberapa penelitian juga telah melaporkan penemuan senyawa AHL dengan struktur spesifik diantaranya isovaleryl-HSL, aryl-HSL (p-coumaroyl-HSL dan cinnamoyl-HSL) dan N-carboxyl-acyl-HSL (Schaefer et al. 2008; Ahlgren et al. 2011;Lindemann et al. 2011; Zhang et al. 2012).
5 Kelarutan, kemampuan berdifusi, serta stabilitas dari senyawa AHL bergantung pada panjang pendek dari rantai asil (Tang dan Zhang 2014). Molekul AHL dengan rantai asil pendek dapat berdifusi secara bebas melalui membran sel, sedangkan molekul AHL dengan rantai asil panjang berdifusi dengan menggunakan mekanisme transpot aktif (Parsek dan Greenberg 2000; Khmel dan Metlitskaya 2006). Senyawa 3OC6-HSL merupakan contoh senyawa AHL dengan rantai asil pendek yang dihasilkan oleh bakteri Vibrio fischeri. Senyawa ini mempunyai kelarutan dan kemampuan berdifusi yang tinggi sehingga dapat berdifusi keluar dan keluar sel secara bebas. Sementara itu senyawa 3OC12-HSL yang dihasilkan oleh Pseudomonas aeruginosa masih memerlukan transport aktif untuk berdifusi (Ng dan Bassler 2009). Modifikasi gugus substitusi pada atom C3 juga dapat meningkatkan kelarutan AHL. Selain itu karakteristik molekul AHL juga dapat dipengaruhi oleh kondisi pH dan suhu di lingkungan. Perubahan pH dan suhu di lingkungan akan mempengaruhi hidrolisis abiotik dari molekul AHL (Yates et al. 2002).
Senyawa AHL dihasilkan oleh AHL sintase dengan S-adenosyl-L-methionine (SAM) dan acyl-ACP sebagai substratnya (Watson et al. 2002). Substrat SAM tersebut dibutuhkan untuk pembentukan cincin homoserin lakton, sedangkan acyl-ACP berperan sebagai pembawa gugus asil (Khmel dan Metlitskaya 2006). Kedua substrat tersebut akan berikatan dengan enzim AHL sintase dan menyebabkan terjadinya reaksi asilasi dan laktonisasi yang menghasilkan produk berupa AHL, holo-ACP serta 5- methylthioadenosine (Gambar 4) (Watson et al. 2002).
Gambar 4. Reaksi Biosintesis asil homoserin lakton (AHL) (Watson et al. 2002).
AHL-Laktonase
6
AHL-laktonase juga dilaporkan dihasilkan oleh bakteri dari kelompok Agrobacterium tumefaciens, Arthrobacter sp., Rhodococcus erythropolis, R. Eutropha (Dong et al. 2002; Zhang et al. 2002; Park et al. 2006; Uroz et al. 2008).
Gambar 5 Mekanisme degradasi AHL oleh AHL-laktonase (Czajkowski dan Jafra 2009)
Ekspresi Gen Pada Escherichia coli
Escherichia coli merupakan bakteri yang sering digunakan sebagai inang untuk produksi protein rekombinan karena kemampuan tumbuhnya yang cepat, mudah dimanipulasi secara genetik, serta banyaknya jumlah vektor kloning yang tersedia (Marisch et al. 2013). Permasalahan yang sering dihadapi ketika menggunakan E.coli sebagai inang ekspresi yaitu rendahnya ekspresi gen asing yang diintroduksikan ke dalam sel dan kelarutan dari protein rekombinan yang dihasilkan. Beberapa strategi yang dapat dilakukan untuk meningkatkan ekspresi dan kelarutan protein yaitu mengganti vektor dan galur inang yang digunakan, penambahan senyawa kimia selama proses induksi serta co-expression (Gopal dan Kumar 2013).
Saat ini telah tersedia berbagai jenis inang E.coli yang telah dimodifikasi secara genetik agar sesuai digunakan untuk ekspresi gen asing. Namun yang paling sering digunakan sebagai inang adalah kelompok E.coli galur BL21, salah satunya E.coli BL21(DE3). Bakteri E.coli BL21(DE3) mempunyai gen penyandi T7 polimerase yang diintroduksikan ke dalam genomnya. Selain itu bakteri tersebut telah dihilangkan kedua gen penyandi proteasenya yaitu lon dan Ompt sehingga tidak dapat mendegradasi protein asing (Ratelade et al. 2009; Gopal dan Kumar 2013).
Upaya penggantian vektor juga menjadi solusi alternatif dalam produksi protein rekombinan. Penggantian vektor bertujuan untuk mengubah promoter dari
7
8
METODE
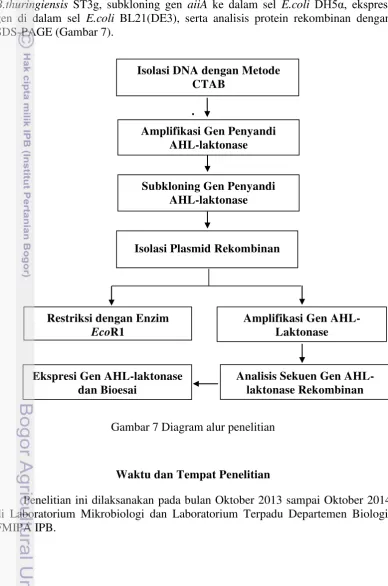
Kerangka Penelitian
Kerangka penelitian ini meliputi isolasi DNA dari B.cereus INT1c dan B.thuringiensis ST3g, subkloning gen aiiA ke dalam sel E.coli DH5α, ekspresi gen di dalam sel E.coli BL21(DE3), serta analisis protein rekombinan dengan SDS-PAGE (Gambar 7).
.
Gambar 7 Diagram alur penelitian
Waktu dan Tempat Penelitian
Penelitian ini dilaksanakan pada bulan Oktober 2013 sampai Oktober 2014 di Laboratorium Mikrobiologi dan Laboratorium Terpadu Departemen Biologi, FMIPA IPB.
Isolasi DNA dengan Metode CTAB
Amplifikasi Gen Penyandi AHL-laktonase
Subkloning Gen Penyandi AHL-laktonase
Analisis Sekuen Gen AHL-laktonase Rekombinan Ekspresi Gen AHL-laktonase
dan Bioesai Restriksi dengan Enzim
EcoR1
9
Isolasi DNA
Isolasi DNA bakteri dilakukan mengikuti metode standar (Sambrook dan Russel 2001). Isolasi DNA dilakukan melalui beberapa tahapan yaitu tahap pencucian sel, lisis, purifikasi, dan pengendapan DNA. Isolat bakteri ditumbuhkan pada 25 mL LB dan diinkubasi selama 18 jam pada suhu 37 °C. Sebanyak 1.5 mL kultur bakteri disentrifugasi pada 12.000 rpm selama 10 menit. Selanjutnya pelet dicuci dua kali dengan 760 uL bufer TE (10 mM Tris HCl; 1 mM EDTA, pH 8.0) dan disentrifugasi pada 12.000 rpm selama 10 menit. Tahap lisis sel diawali dengan penambahan 200 µL bufer TE dan 45 µL lisozim pada pelet, kemudian diinkubasi selama 1 jam pada suhu 55 °C. Selanjutnya sampel ditambah dengan 100 µL larutan sodium dodecyl sulfate (SDS) 10% dan 50 µL proteinase K, diinkubasi pada suhu 55 °C selama 1 jam. Sampel ditambahkan 100 µL cetyl trimetyl amonium bromide (CATB) dan 100 µL larutan 5 M NaCl, kemudian diinkubasi pada suhu 65 °C selama 30 menit. Setelah sel mengalami lisis maka dilakukan tahapan purifikasi dan pengendapan DNA.
Tahap purifikasi dilakukan dengan penambahan 600 µL larutan fenol:kloroform:isoamil alkohol (24:24:1). Selanjutnya sampel disentrifugasi pada 12000 rpm selama 10 menit sehingga terbentuk 2 lapisan. Lapisan atas dipindahkan ke tabung baru dan ditambahkan 600 µL kloroform:isoamil alkohol (24:1). Sampel disentrifugasi kembali pada 12000 rpm selama 10 menit. Lapisan atas dipindahkan ke tabung mikro baru, kemudian dilakukan proses pengendapan DNA. Tahap pengendapan DNA dilakukan dengan penambahan 0.6 kali volume etanol absolut. Sampel diinkubasi selama semalam pada suhu 37 °C, kemudian disentrifugasi pada 12000 rpm selama 10 menit. Pelet DNA dicuci dengan 1 mL etanol 70% dan disentrifugasi kembali pada 12000 rpm selama 10 menit. Pelet dikeringanginkan dan ditambah 50 µL ddH2O. Kontaminasi RNA dibersihkan dengan penambahan 0.1 kali volume RNase dan diinkubasi selama 1 jam pada suhu 37 C.
Amplifikasi Gen Penyandi AHL-Laktonase
Amplifikasi gen aiiA dilakukan dengan menggunakan primer aiiAF (5’ -GGG GGA CAT ATG ACA GTA AAG AAG CTT TAT TTC G-3’) dan aiiAR
10
Pembuatan Sel Kompeten
Sebanyak 2 mL biakan E.coli DH5α umur 24 jam disubkultur ke dalam 50 mL media LB. Kultur diinkubasi selama 2-3 jam pada suhu 37 C sampai nilai Optical Density (OD600) mencapai 0.45-0.5. Selanjutnya kultur diambil sebanyak 1,5 mL dan disentrifugasi pada kecepatan 5000 rpm selama 15 menit. Pelet diresuspensi dengan 1 mL bufer transformasi. Sampel diinkubasi di dalam es selama 15 sampai 30 menit, kemudian disentrifugasi pada 5000 rpm selama 5 menit dengan suhu 4 C. Pelet diresuspensi kembali dengan 250 µL bufer transformasi (Sambrook dan Russel 2001).
Subkloning Gen AHL-laktonase
Subkloning dilakukan dengan menggunakan vektor pGEMT-Easy (Gambar 8). Subkloning dimulai dengan meligasikan DNA hasil amplifikasi pada plasmid pGEMT-Easy di daerah MCS (Gambar 9). Reaksi ligasi terdiri dari 2.5 µL fragmen DNA (15 ng/ µL), 1 µL vektor pGEMT-Easy (50 ng), 5 µL bufer ligasi, 1 µL T4 DNA ligase dan 0.5 µL ddH2O. Proses ligasi dilakukan dengan inkubasi sampel pada suhu 10 C selama 16 jam. DNA rekombinan hasil ligasi ditransformasikan ke dalam suspensi sel kompeten (E.coli DH5α) (Gambar 10). Sampel diinkubasi dalam es selama 45 menit dan setiap 15 menit dijentik secara perlahan. Selanjutnya sampel diberi perlakuan renjatan panas pada suhu 42 C selama 45 detik, kemudian diinkubasi dalam es selama 15 menit.
Sampel diresuspensi dengan 250 µL media LB dan diinkubasi selama 1 jam pada suhu 37 C. Sampel disentrifugasi pada kecepatan 3000 rpm selama 5 menit, kemudian diresuspensi dengan 100 µL media LB dan ditumbuhkan pada media LA. Media LA yang digunakan mengandung 50 mg/mL ampisilin, 40 µg/mL IPTG, dan 40 µg/mL X-Gal. Biakan diinkubasi pada suhu 37 C selama 24 jam (Sambrook dan Russel 2001).
11
Gambar 9 Sekuen multiple cloning site (MCS) dari plasmid pGEMT-Easy
12
Isolasi Plasmid Rekombinan
Isolasi plasmid rekombinan dilakukan mengikuti prosedur High Speed Plasmid Mini Kit (Geneaid). Koloni E.coli DH5α rekombinan ditumbuhkan dalam 2 mL media LB yang mengandung 50 mg/mL ampisilin dan diinkubasi pada suhu 37 C selama semalam. Sebanyak 1.5 mL kultur disentrifugasi dengan kecepatan 13.000 selama 1 menit. Pelet ditambahkan 200 µL bufer PD1, kemudian diresuspensi dengan 200 µL bufer PD2. Selanjutnya sampel dibolak balik 10 kali dan diinkubasi selama 2 menit pada suhu ruang. Sampel ditambah 300 µL bufer PD3 dan disentrifugasi pada kecepatan 13.000 rpm selama 3 menit. Cairan jernih dibagian atas dipindahkan ke dalam tabung kolom dan disentrifugasi pada 13.000 rpm selama 30 detik. Setelah itu ditambahkan 400 µL bufer W1 dan disentrifugasi pada 13.000 rpm selama 30 detik. Larutan bagian bawah dibuang dan kolom dicuci dengan 600 µL bufer pencuci. Sampel disentrifugasi pada 13.000 rpm selama 30 detik, kemudian larutan bagian bawah dibuang. Kolom dikeringkan dengan disentrifugasi pada 13.000 rpm selama 3 menit. Selanjutnya DNA dielusi dengan 50 µL bufer elusi dan diinkubasi selama 3 menit pada suhu ruang, kemudian disentrifugasi pada 13.000 rpm selama 2 menit.
Pemotongan Plasmid Rekombinan dengan EcoRI
Konfirmasi DNA sisipan dalam plasmid pGEMT-Easy dilakukan menggunakan enzim EcoRI. Plasmid dipotong dengan mereaksikan 5 µL DNA plasmid, 1 µL 10x bufer enzim restriksi, 1 µL enzim restriksi dan 3 µL ddH2O. Sampel diinkubasi pada suhu 37 C selama semalam. Fragmen DNA yang telah dipotong dengan enzim dianalisis melalui elektroforesis pada gel agarosa 1.5 %.
Amplifikasi Plasmid Rekombinan
Konfirmasi hasil transformasi juga dilakukan dengan amplifikasi plasmid rekombinan menggunakan primer T7 dan SP6. Reaksi PCR terdiri dari 2 µL template DNA, 1.25 µL primer T7, 1.25 µL primer SP6, 12.5 µL Go Taq Green Ready Mix (Promega), dan 8 µL ddH2O. Tahapan PCR yang digunakan yaitu pre-denaturasi 94 C selama 2 menit, pre-denaturasi 94 C selama 30 detik, annealing 50 C selama 30 detik, elongasi 72 C selama 1 menit, dan elongasi akhir 72 C selama 10 menit. Siklus PCR berlangsung sebanyak 30 siklus.
Analisis Sekuen Gen AHL-laktonase Rekombinan
13 tiga dimensi di situs prosite (http://protein model portal.org). Analisis nilai titik isoelektrik dan berat molekul juga dilakukan pada sekuen asam amino (http://web.expasy.org/compute_pi/).
Ekspresi Gen AHL-laktonase dan Bioesai
Plasmid rekombinan yang telah subkloning dipotong menggunakan enzim restriksi BamHI dan NdeI, kemudian diligasikan dalam plasmid pET15b. Plasmid rekombinan ditransformasikan ke dalam bakteri E.coli BL21(DE3) (Gambar 11). Sel transforman ditumbuhkan di media LB yang mengandung 50 mg/mL ampisilin. Kultur diinkubasi selama 2-3 jam pada suhu 37C. Ekpresi gen diinduksi dengan penambahan 0.1 mM IPTG ketika OD600 kultur mencapai 0.6, kemudian diinkubasi selama 2-3 jam pada suhu 37C. Kultur E.coli BL21(DE3) dan E.coli BL21(DE3) rekombinan tanpa penambahan IPTG digunakan sebagai kontrol.
Bioesai aktivitas AHL-laktonase dilakukan secara kualitatif dengan menggunakan bakteri C.violaceum sebagai biokontrol. Sebanyak 120 μL kultur E.coli rekombinan diteteskan pada kertas cakram, kemudian diletakkan di atas media luria agar semi padat yang telah mengandung 1% C.violaceum. Biakan diinkubasi selama 24 sampai 48 jam pada suhu ruang. Ekspresi gen penyandi AHL-laktonase ditunjukkan dengan terbentuknya zona hambat disekitar kertas cakram.
SDS-PAGE
Sebanyak 500 µL kultur E.coli BL21(DE3) dilisis menggunakan sonikator. Sonikasi dilakukan pada frekuensi 13.000 Hz selama 1 menit dengan jeda 10 detik. Sel yang telah lisis diambil 100 µL dan dicampur dengan 30 µL 5x larutan bufer sampel. Selanjutnya sampel divortek dan direbus pada suhu 100 C selama 5 menit. Sampel yang telah terdenaturasi dielektroforesis pada 12% gel poliakrilamida selama 4 jam dengan tegangan 60 V dan arus 22 mA (Lampiran 3). Gel hasil elektroforesis diwarnai menggunakan Silver Staining (Lampiran 4). Berat molekul AHL-laktonase dihitung dengan menggunakan persamaan linear berdasarkan nilai Rf (mobilitas relatif) marker dan log BM (berat molekul) (Lampiran 5). Nilai Rf dihitung dengan menggunakan rumus:
Rf =
Jarak migrasi pita protein
14
15
HASIL DAN PEMBAHASAN
Hasil
Amplifikasi Gen aiiA dan Konfirmasi E.coliDH5α Rekombinan
Isolasi DNA dengan metode CTAB berhasil mendapatkan DNA dengan konsentrasi dan nilai kemurnian yang baik. Konsentrasi DNA dari B.thuringiensis SGT3g yaitu 361.9 ng/µL dengan nilai kemurnian pada A260/280 sebesar 2.02. Sementarai itu B.cereus INT1c mempunyai konsentrasi DNA sebesar 280.9 ng/µL dengan nilai kemurnian 1.99. Deteksi gen penyandi AHL-laktonase dari kedua isolat dengan primer aiiA berhasil mengamplifikasi fragmen DNA berukuran 800 bp (Gambar 12). Jumlah amplikon dari isolat B.cereus INT1c sebanyak 918 bp (Lampiran 1) dan B.thuringiensis SGT3g sebanyak 903 bp (Lampiran 2).
Konfirmasi plasmid rekombinan dalam sel E.coli DH5α melalui pemotongan dengan EcoRI menghasilkan 2 fragmen DNA berukuran 3015 bp dan 800 bp (Gambar 13). Enzim EcoRI tersebut memisahkan DNA sisipan berukuran 800 bp dari plasmid pGEMT-Easy.
Gambar 12 Produk PCR gen aiiA pada gel agarosa 1.5%. Sumur dari kiri: (M) marker 1 kb, (1) B.cereus INT1c, (2) B.thuringiensis SGT3g.
16
Konfirmasi dengan primer T7 dan SP6 juga berhasil mengamplifikasi DNA sisipan dalam plasmid pGEMT-Easy berukuran 1000 bp. Fragmen tersebut merupakan hasil amplifikasi DNA sisipan dan sekuen situs restriksi di MCS (Gambar 14).
Gambar 14 Hasil amplifikasi plasmid rekombinan. Sumur dari kiri: (M) marker 1kb, (1) B.cereus INT1c, (2) B.thuringiensis SGT3g.
Sekuen Asam Amino dan Filogenetik AHL-Laktonase B.cereus INT1c dan
B.thuringiensis SGT3g
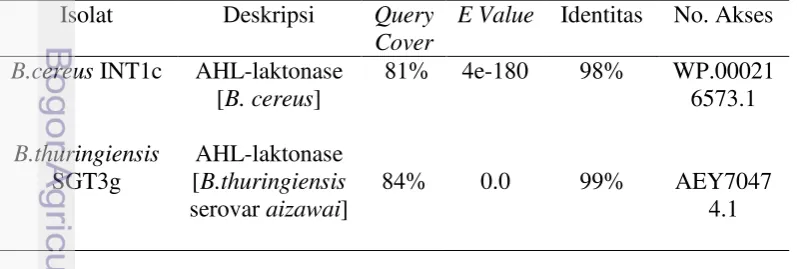
Bakteri B.cereus INT1c dan B.thuringiensis SGT3g mempunyai sekuen penyandi AHL-laktonase yang mirip setelah disejajarkan dengan MEGA5 software. Analisis sekuen dengan program BLAST-X menunjukkan bahwa asam amino dari B.cereus INT1c memiliki homologi 98% dengan B.cereus nomor akses WP.000216573.1, sedangkan B.thuringiensis SGT3g memiliki homologi 99% dengan B.thuringiensis serovar aizawai nomor akses AEY70474.1 (Tabel 1). Tabel 1 Hasil BLAST-X dari sekuen gen aiiA
Analisis jarak evolusioner kedua isolat dengan pohon filogenetik menghasilkan distribusi homologi gen aiiA yang beragam dengan kelompok Bacillus yang lain (Gambar 15). Kedua isolat memiliki kekerabatan dekat dengan B.amyloliquefaciens, B.weihenstephanensis, dan B.thuringiensis serovar kim.
17 Bakteri Pseudomonas aeruginosa (nomor akses WP023464371.1) dipilih sebagai outer group yang merupakan bakteri Gram negatif penghasil AHL-asilase.
Gambar 15 Pohon filogenetik dari asam amino AHL-laktonase yang dikonstruksi menggunakan metode neighbor-joining.
Open Reading Frame (ORF) Gen AHL-Laktonase B.cereus INT1c dan
B.thuringiensis SGT3g
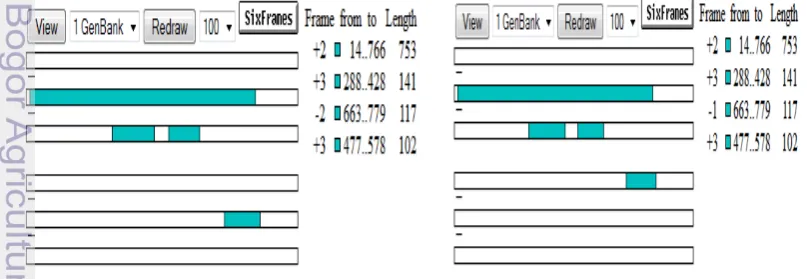
Hasil analisis dengan ORF Finder menunjukkan bahwa kedua isolat bakteri mempunyai 1 ORF yang diduga dapat mengekspresikan AHL-laktonase yaitu pada orf2 (Gambar 16). Kedua isolat mempunyai orf2 yang sama yang dimulai dari basa ke-14 dan diakhiri pada basa ke 766. Orf2 terdiri atas 752 bp yang menyadikan 250 asam amino. Sekuen gen aiiA dari kedua isolat telah lengkap karena mempunyai start kodon (ATG) dan stop kodon (TAG) (Gambar 17 dan 18).
18
14 atgacagtaaagaagctttatttcgtcccagcaggtcgttgtatg M T V K K L Y F V P A G R C M 59 ttagatcattcttctgttaatagtacactcgcgccggggaattta L D H S S V N S T L A P G N L 104 ttgaacttacctgtatggtgttatcttttggagacagaagagggg L N L P V W C Y L L E T E E G 149 cctattttagtagatacaggtatgccagaaagtgcagttaataat P I L V D T G M P E S A V N N 194 gaagggatttttaacggtacatttgttgaaggacagattttaccg E G I F N G T F V E G Q I L P 239 aaaatgactgaagaagatagaatcgtgaatatattaaagcgtgta K M T E E D R I V N I L K R V 284 gggtatgagccggacgaccttttatatattattagttctcactta G Y E P D D L L Y I I S S H L 329 cattttgatcatgcaggaggaaacggtgcttttacaaatacaccg H F D H A G G N G A F T N T P 374 attattgtgcagcgagcggaatatgaggcagcacttcatagagaa I I V Q R A E Y E A A L H R E 419 gaatatatgaaagaatgtatattaccgcatttgaactacaaaatt E Y M K E C I L P H L N Y K I 464 attgaaggggattatgaagtggtaccaggtgttcaattattgtat I E G D Y E V V P G V Q L L Y 509 acgccaggtcattctccaggccatcagtcgttattcattgagacg T P G H S P G H Q S L F I E T 554 gagcaatccggttcagttttattaacaattgatgcatcgtacacg E Q S G S V L L T I D A S Y T 599 aaagagaattttgaagatgaagtgccgttcgcaggatttgatcca K E N F E D E V P F A G F D P 644 gaattagctttatcttcaatcaaacgcttaaaagaagttgtgaca E L A L S S I K R L K E V V T 689 aaagagaaatcgattgttttctttggtcatgatatagagcaggaa K E K S I V F F G H D I E Q E 734 aagggttgtagagtgttcccggagtatatatag 766 K G C R V F P E Y I *
19 14 atgacagtaaagaagctttatttcgtcccagcaggtcgttgtatg
M T V K K L Y F V P A G R C M 59 ttagatcattcttctgttaatggtacactcgcgccggggaattta L D H S S V N G T L A P G N L 104 ttgaacttacctgtatggtgttatcttttggagacagaagagggg L N L P V W C Y L L E T E E G 149 cctattttagtagatacaggtatgccagaaagtgcagttaataat P I L V D T G M P E S A V N N 194 gaagggctttttaacggtacatttgttgaaggacagattttaccg E G L F N G T F V E G Q I L P 239 aaaatgactgaagaagatagaatcgtgaatatattaaagcgtgta K M T E E D R I V N I L K R V 284 gggtatgagccggacgaccttttatatattattagttctcactta G Y E P D D L L Y I I S S H L 329 cattttgatcatgcaggaggaaacggtgcttttacaaatacaccg H F D H A G G N G A F T N T P 374 attattgtgcagcgaacggaatatgaggcagcacttcatagagaa I I V Q R T E Y E A A L H R E 419 gaatatatgaaagaatgtatattaccgcatttgaactacaaaatt E Y M K E C I L P H L N Y K I 464 attgaaggggattatgaagtggtaccaggtgttcaattattgtat I E G D Y E V V P G V Q L L Y 509 acgccaggtcattctccaggccatcagtcgctattcattgagacg T P G H S P G H Q S L F I E T 554 gagcaatccggctcagttttattaacaattgatgcatcgtacacg E Q S G S V L L T I D A S Y T 599 aaagagaattttgaagatgaagtgccgttcgcaggatttgatcca K E N F E D E V P F A G F D P 644 gaattagctttatcttcaattaaacgtttaaaaggagttgtggcg E L A L S S I K R L K G V V A 689 aaagagaaaccaattgttttctttggtcatgatatagagcaggaa K E K P I V F F G H D I E Q E 734 aagggttgtagagtgttccctgagtatatatag 766 K G C R V F P E Y I *
20
Domain dan Struktur Model Protein AHL-Laktonase B.cereus INT1c dan
B.thuringiensis SGT3g
AHL-laktonase dari kedua isolat termasuk dalam domain metallo-β -lactamase superfamily. Perbedaan domain kedua isolat terletak pada situs fosforilasi kasein kinase II. Bakteri B.cereus INT1c tidak mempunyai residu treonin nomor 126. Situs aktif pada domain terletak pada urutan asam amino nomor 149-153. Daerah tersebut merupakan situs forsforilasi tirosin kinase (Gambar 19 dan 20).
MTVKKLYFVPAGRCMLDHSSVNGTLAPGNLLNLPVWCYLLETEEGPILVDTGMPE SAVNNEGLFNGTFVEGQILPKMTEEDRIVNILKRVGYEPDDLLYIISSHLHFDHA GGNGAFTNTPIIVQRTEYEAALHREEYMKECILPHLNYKIIEGDYEVVPGVQLLY TPGHSPGHQSLFIETEQSGSVLLTIDASYTKENFEDEVPFAGFDPELALSSIKRL KGVVAKEKPIVFFGHDIEQEKGCRVFPEYI
Gambar 19 Hasil analisis domain dari isolat B.thuringiensis SGT3g. Daerah berwarna biru: situs fosforilasi kasein kinase II, dan daerah berwarna kuning: merupakan situs aktif yaitu situs forsforilasi tirosin kinase.
MTVKKLYFVPAGRCMLDHSSVNSTLAPGNLLNLPVWCYLLETEEGPILVDTGMPE SAVNNEGIFNGTFVEGQILPKMTEEDRIVNILKRVGYEPDDLLYIISSHLHFDHA GGNGAFTNTPIIVQRAEYEAALHREEYMKECILPHLNYKIIEGDYEVVPGVQLLY TPGHSPGHQSLFIETEQSGSVLLTIDASYTKENFEDEVPFAGFDPELALSSIKRL KEVVTKEKSIVFFGHDIEQEKGCRVFPEYI
Gambar 20 Hasil analisis domain dari isolat B.cereus INT1c. Daerah berwarna biru: situs fosforilasi kasein kinase II, dan daerah berwarna kuning: merupakan situs aktif yaitu situs forsforilasi tirosin kinase.
21
Gambar 21 Prediksi struktur 3 dimensi protein AHL-laktonase, kiri: lipatan αβ/βα dan kanan: motif HXHXDH pada AHL-laktonase.
Titik isoelektrik merupakan pH ketika muatan suatu molekul protein bernilai nol. Bakteri B.thuringiensis SGT3g diprediksi mempunyai titik isoelektrik sebesar 4.81, sedangkan B.cereus INT1c mempunyai titik isoelektrik 4.7. Kedua isolat juga diprediksi mempunyai berat molekul yang sama (Gambar 22 dan 23).
10 20 30 40 50 60
IGSMTVKKLY FVPAGRCMLD HSSVNGTLAP GNLLNLPVWC YLLETEEGPI LVDTGMPESA 70 80 90 100 110 120 VNNEGLFNGT FVEGQILPKM TEEDRIVNIL KRVGYEPDDL LYIISSHLHF DHAGGNGAFT 130 140 150 160 170 180 NTPIIVQRTE YEAALHREEY MKECILPHLN YKIIEGDYEV VPGVQLLYTP GHSPGHQSLF 190 200 210 220 230 240 IETEQSGSVL LTIDASYTKE NFEDEVPFAG FDPELALSSI KRLKGVVAKE KPIVFFGHDI 250
EQEKGCRVFP EYI
pI/Mw: 4.81 / 28.24617
Gambar 22 Prediksi titik isoelektrik (pI) dan berat molekul (Mw) AHL-laktonase dari bakteri B.thuringiensis SGT3g.
10 20 30 40 50 60 MTVKKLYFVP AGRCMLDHSS VNSTLAPGNL LNLPVWCYLL ETEEGPILVD TGMPESAVNN 70 80 90 100 110 120 EGIFNGTFVE GQILPKMTEE DRIVNILKRV GYEPDDLLYI ISSHLHFDHA GGNGAFTNTP 130 140 150 160 170 180 IIVQRAEYEA ALHREEYMKE CILPHLNYKI IEGDYEVVPG VQLLYTPGHS PGHQSLFIET 190 200 210 220 230 240 EQSGSVLLTI DASYTKENFE DEVPFAGFDP ELALSSIKRL KEVVTKEKSI VFFGHDIEQE 250
KGCRVFPEYI
pI/Mw: 4.77 / 28.08093
Gambar 23 Prediksi titik isoelektrik (pI) dan berat molekul (Mw) AHL-laktonase dari bakteri B.cereus INT1c.
Ekspresi Gen aiiA danAkvifitas AHL-laktonase Hasil Kloning
22
BL21(DE3) telah terligasi dengan fragmen gen aiiA dari B.cereus INT1c dan B.thuringiensis SGT3g. Penambahan 1 mM IPTG dapat menginduksi ekspresi gen aiiA dibawah kontrol T7 lac promoter. Penambahan 1 mM IPTG ketika OD600 kultur mencapai 0.6 menghasilkan protein AiiA dengan aktivitas lebih tinggi dibandingkan saat OD600 0.8. Bakteri E.coli rekombinan dengan nilai OD600 0.6 mempunyai aktivitas penghambatan terhadap bakteri C.violaceum. Hal ini ditunjukkan dengan terbentuknya zona berwarna opaque disekitar kertas cakram. Sementara itu bakteri E.coli rekombinan dengan nilai OD600 0.8 tidak menunjukkan aktivitas degradasi terhadap senyawa AHL. Sementara itu pada E.coli BL21(DE3) non rekombinan dan E.coli BL21(DE3) tanpa perlakuan IPTG tidak menunjukkan adanya aktivitas AHL-laktonase (Gambar 25).
Gambar 24 Hasil restriksi plasmid rekombinan dalam E.coli BL21(DE3) dengan enzim restriksi BamHI dan NdeI.
23 Analisis SDS-PAGE menunjukkan bahwa overexpression gen aiiA dari kedua isolat berhasil dilakukan dalam sel E.coli BL21(DE3) rekombinan dengan induksi IPTG. Hasil ini ditandai dengan terbentuknya pita yang lebih tebal pada sampel protein kedua isolat dibandingkan dengan pita kontrol dan pita E.coli BL21(DE3) rekombinan tanpa perlakuan IPTG. Protein AiiA dari kedua isolat mempunyai berat molekul yang sama dan diprediksi berukuran sekitar 28.77 kDa (Gambar 26).
Gambar 26 Hasil analisis SDS-PAGE protein AiiA rekombinan. Sumur dari kiri ke kanan: (M) Marker, (1) E.coli BL21(DE3) non rekombinan, (2 dan 4) E.coli BL21(DE3) rekombinan yang tidak diinduksi dengan IPTG, (3 dan 5) E.coli BL21(DE3) rekombinan yang diinduksi dengan IPTG, (2 dan 3) E.coli rekombinan dengan gen aiiA dari B.cereus INT1c, (4 dan 5) E.coli rekombinan dengan gen aiiA dari B.thuringiensis SGT3g.
Pembahasan
Enzim pendegradasi AHL dilaporkan banyak terdapat pada kelompok proteobakteria dan disandikan oleh berbagai macam gen seperti aiiA, aiiB, attM, ah1D, dan aiiD (Carlier et al. 2003; Dong et al. 2000; Lin et al. 2003; Park et al.
2003). Isolat B.cereus INT1c dan B.thuringiensis SGT3g terdeteksi mempunyai aktivitas degradasi AHL yang tinggi dari hasil uji kualitatif dengan C.violaceum sebagai biokontrol. Beberapa hasil penelitian juga melaporkan bahwa AHL-laktonase dengan spektrum degradasi substrat yang luas telah banyak ditemukan pada bakteri B.cereus dan B.thuringiensis (Dong et al. 2002; Lee et al. 2002; Flores et al. 2014). Aktivitas degradasi substrat dengan kisaran luas berkaitan dengan mekanisme hidrolisis AHL-laktonase yang tidak dipengaruhi oleh panjang rantai asil dari AHL (Cao et al. 2012; Chen et al. 2013; Lade et al. 2014).
24
2007; Anzhou et al. 2013; Dong et al. 2002; Lee et al. 2002). Fragmen berukuran 800 bp tersebut setelah dianalisis dengan ORF Finder hanya 752 bp yang diprediksi dapat mengekspresikan protein AiiA. Penelitian sebelumnya juga melaporkan bahwa sekuen gen aiiA dari genus Bacillus berukuran sekitar 750 bp yaitu Bacillus sp. 240B1 (750 bp), B.subtilis BS-1 dan Bacillus sp. B546 (753 bp) serta B. thuringiensis 147-11516 (754 bp) (Dong et al. 2002; Pan et al. 2008; Chen et al. 2010; Flores et al. 2014). Eksplorasi gen aiiA dari bakteri kelompok Bacillus akan sangat membantu untuk pengembangan agen biokontrol terhadap bakteri fipatogen penghasil senyawa AHL. Sementara itu beberapa mikroba dilaporkan tidak hanya menghasilkan enzim pendegradasi AHL sebagai mekanisme pertahanan melainkan juga menggunakan molekul hasil degradasinya digunakan untuk pertumbuhan sel (Tinh et al. 2007).
Homologi AiiA telah banyak dikarakterisasi pada bakteri Gram positif dan Gram negatif. Sekuen penyandi aiiA yang beragam membuat gen ini sulit diamplifikasi menggunakan primer spesifik (Anzhou et al. 2013). Primer aiiA yang digunakan untuk deteksi gen penyandi AHL-laktonase dari kedua isolat merupakan primer yang spesifik mengamplifikasi daerah kromosom. Dong et al. (2000) melaporkan bahwa lokasi gen aiiA bukan terletak pada plasmid melainkan pada DNA kromosom. Hal ini dibuktikan dengan kemampuan B.thuringiensis subsp. kurstaki B2 dan B.thuringiensis subsp. israelensis strain B23 untuk menghasilkan AHL-laktonase meskipun telah kehilangan plasmidnya.
Konfirmasi DNA sisipan di dalam plasmid rekombinan dengan primer T7 dan SP6 menghasilkan fragmen DNA berukuran 1000 bp. Perubahan ukuran fragmen DNA terjadi karena adanya penambahan nukleotida. Amplifikasi plasmid rekombinan dengan primer SP6 dan T7 meningkatkan ukuran amplikon sebesar 182 bp. Hal ini sesuai dengan jumlah nukleotida tambahan di daerah MCS yang terletak antara ujung 5' primer T7 dan SP6 (Mukhopadhyaya et al. 2006).
Bakteri B.cereus INT1c dan B.thuringiensis SGT3g mempunyai kesamaan sekuen gen penyandi AHL-laktonase setelah disejajarkan dengan MEGA 5.0 software serta mempunyai karakteristik morfologi yang mirip. Analisis dengan BLAST-X dan pohon filogenetik menunjukkan bahwa keduanya memiliki hubungan kekerabatan yang dekat. Taksonomi dari bakteri B.cereus dan B.thuringiensis sampai saat ini masih menjadi perdebatan. Studi berdasarkan analisis kromosom belum bisa membedakan genetik dari kedua bakteri. Kedua bakteri belum bisa dibedakan termasuk kedalam spesies yang berbeda atau varietas dari spesies yang sama (Ivanova et al. 2003). Hal ini disebabkan B.cereus and B.thuringiensis mempunyai kesamaan pada fenotip dan genotipnya (Oh et al. 2011). Bakteri B.thuringiensis dibedakan dari B.cereus karena mempunyai gen cry di dalam plasmidnya (Helgason et al. 2000).
25 Asp191 dan ion hidroksida (OH-) (Liu et al. 2005; Momb et al. 2008). Analisis struktur kristal dari B.cereus menunjukkan bahwa residu histidin pada motif HXHXDH terlibat dalam pengikatan ion Zn2+, sedangkan residu aspartat terlibat dalam reaksi katalisis (Carfi et al. 1995). Dong et al. (2000) juga melaporkan bahwa penggantian residu histidin pada motif HXHXDH dengan residu serin dan leusin menyebabkan akifitas AHL-laktonase menurun. AHL-laktonase merupakan enzim yang sangat spesifik dengan motif katalitik 106HXDH109~H169. Aktivitas enzim tidak bergantung pada ion Zn atau ion logam meskipun AHL-laktonase mempunyai ion Zn2+ pada pusat katalitiknya,. Hal ini yang menyebabkan AHL-laktonase dibedakan dengan kelompok metallo-β-lactamase superfamily yang lain walaupun mempunyai struktur dan motif yang identik (Wang et al. 2004; Chen et al. 2013).
Hasil prediksi titik isoelektrik mengindikasikan bahwa AHL-laktonase yang dihasilkan oleh B.thuringiensis SGT3g dan B.cereus INT1c merupakan protein yang bersifat asam dengan titik isoelektrik sebesar 4.7 dan 4.8. Wang et al. (2004) juga melaporkan bahwa protein AiiA merupakan protein yang bersifat asam dengan titik isoelektrik 4.17. Pada titik isoelektriknya struktur protein menjadi lebih hidrofobik dan lebih kompak, namun menjadi kurang stabil karena tidak adanya gaya tolak antar partikel (Salgin et al. 2006; Patil et al. 2007). Hal ini membuat protein lebih mudah membentuk agregat dan mengendap pada titik isoelektriknya (Salgin et al. 2012). Interaksi elektrostatik diantara muatan residu asam amino berperan penting dalam mempertahankan konformasi protein AHL-laktonase. Struktur konformasi asimetris AHL-laktonase tidak berubah pada pH 7-9, sedangkan perubahan pH menjadi 5 membuat AHL-laktonase menjadi kehilangan struktur asimetrisnya (Wang et al. 2004).
Bioesai aktivitas AHL-laktonase menunjukkan bahwa E.coli BL21(DE3) rekombinan dapat menghambat produksi pigmen violacein pada bakteri C.violaceum. Bakteri C.violaceum merupakan bakteri Gram negatif yang menggunakan mekanisme quorum sensing untuk membentuk pigmen violacein. Bakteri tersebut menghasilkan senyawa AHL sebagai molekul sinyal yang disintesis oleh CviI (McClean et al. 1997; Duran dan Menck 2001; Stauff dan Blasser 2011). Gen penyandi AHL-laktonase dari kedua isolat diekspresikan setelah kultur diinduksi dengan IPTG pada fase eksponensial. Induksi dengan IPTG ketika OD600 kultur mencapai 0.6 dapat menghasilkan protein AiiA rekombinan dengan aktivitas yang tinggi, sedangkan induksi IPTG ketika OD600 kultur mencapai 0.8 tidak dapak menginduksi ekspresi gen aiiA. Induksi E.coli rekombinan dengan IPTG pada awal fase eksponensial dapat meningkatkan aktivitas protein rekombinan yang dihasilkan, sedangkan induksi pada akhir fase lag dapat menghambat pertumbuhan sel (Kwon et al. 2002; Olaofea et al. 2012).
26
sisanya terdapat dalam bentuk inclusion bodies. Kondisi lingkungan akan sangat mempengaruhi aktivitas protein serta jumlah protein terlarut dan tidak larut yang dihasilkan oleh E.coli rekombinan. Ekspresi protein rekombinan dilaporkan dipengaruhi oleh berbagai faktor seperti waktu induksi, lama inkubasi setelah induksi, suhu inkubasi, jenis vektor serta inang yang digunakan (Picaud et al. 2007; Dang et al. 2012; Fang et al. 2014; Piubelli et al. 2013). Molina et al. (2003) melaporkan bahwa mekanisme degradasi AHL oleh protein AiiA yang tidak disekresikan keluar sel dapat terjadi di dalam sitoplasma. AHL merupakan molekul sinyal yang dapat keluar masuk melalui membran sel secara difusi maupun melalui transport aktif. AHL yang telah terakumulasi di lingkungan akan berdifusi kembali ke dalam sel dan didegradasi oleh protein AiiA di dalam sitoplasma. Dengan demikian jumlah molekul AHL dilingkungan dapat berkurang tanpa perlu mensekresikan protein AiiA keluar sel.
Ekspresi gen aiiA dari kedua isolat dalam sel E.coli BL21(DE3) menghasilkan protein AiiA dengan berat molekul berukuran 28.77 kDa. Berat molekul tersebut hampir sama dengan berat molekul hasil prediksi di situs EXPASY berdasarkan sekuen asam amino dari kedua isolat (http://web.expasy.org/compute_pi/). Lee et al. (2002) melaporkan bahwa gen aiiA dari isolat B.thuringiensis subsp. morrisoni dan B.thuringiensis subsp. kyushuensis berhasil dieskspresikan dalam sel E.coli BL21(DE3) dan menghasilkan protein AiiA dengan berat molekul sekitar 28 kDa. Bakteri B.thuringiensis 47-11516 dan Bacillus sp. 240B1 juga dilaporkan menghasilkan protein AiiA dengan berat molekul sekitar 28 kDa (Flores et al. 2012; Dong et al. 2000).
SIMPULAN DAN SARAN
Simpulan
Gen aiiA dari B.cereus INT1c dan B.thuringiensis SGT3g dapat diekspresikan di dalam sel E.coli BL21(DE3) rekombinan. Protein AiiA yang dihasilkan oleh E.coli rekombinan dapat menghambat produksi violacein pada C.violaceum melalui degradasi senyawa AHL. Ekspresi gen AHL-laktonase rekombinan dan aktivitas proteinnya dipengaruhi oleh waktu induksi IPTG. Protein AiiA rekombinan dari kedua isolat mempunyai berat molekul 28.77 kDa.
Saran
27
DAFTAR PUSTAKA
Afiah TSN. 2011. Isolasi dan karakterisasi bakteri penghasil acyl-homoserin lactonase (AHL-laktonase) asal lahan pertanian di Jawa [skripsi]. Bogor (ID): Institut Pertanian Bogor.
Ahlgren NA, Harwood CS, Schaefer AL, Giraud E, Greenberg EP. 2011. Aryl-homoserine lactone quorum sensing in stem-nodulating photosynthetic bradyrhizobia. Proc Natl Acad Sci USA. 108:7183-7188.doi:10.1073/pnas. 1103821108.
Ang G, Gui-ying M, Shun L, Ping W, Qun T, Yan-ping L,Hui W, Xiao-min A, Li-qun Z, Xiao-xue Y, Dong-cai L. 2013. High-resolution structures of AidH complexes provide insights into a novel catalytic mechanism for N-acyl homoserine lactonase. Acta Cryst. 69:82-91.
Anzhou M, Di L, Xuliang Z, Guoqiang Z. 2013. Quorum quenching in culturable phyllosphere bacteria from tobacco. Int J Mol Sci. 14:14607-14619.doi:10. 3390/ijms140714607.
Cao Y, He S, Zhou Z, Zhang M, Mao W, Zhang H, Yao B. 2012. Orally administered thermostable N-acyl homserine lactonase from Bacillus sp. strain AI96 attenuates Aeromonas hydrophila infection in zebrafish. Appl Environ Microbiol. 78: 1899-1908.doi:10.1128/AEM.06139-11.
Carfi A, Pares S, Duee E, Galleni M, Duez C, Frere JM, Dideberg O. 1995. The 3-D structure of a zin metallo-β-lactamase from Bacillus cereus reveals a new type of protein fold. EMBO J. 14:4914-4921.
Carlier A, Uroz S, Smadja B, Fray R, Latour X, Dessaux Y, Faure D. 2003. The Ti plasmid of Agrobacterium tumefaciens harbors an attM-paralogous gene, aiiB, also encoding N-acyl homoserine lactonase activity. Appl Environ Microbiol. 69:4989-4993.doi:10.1128/AEM.69.8.
Chan KG, Tiew SZ, Ng CC. 2007. Rapid isolation method of soil Bacilli and screening oftheir quorum quenching activity. Asia Pac J Mol Biol. 15:153-156.
Chen F, Gao Y, Chen X, Yu Z, Li X. 2013. Quorum quenching enzymes and their application in degrading signal molecules to block quorum sensing dependent infection. Int J Mol Sci. 14:17477-17500.
Chen X, Huang Z, Zhou B, Wang H, Jia G, Qiao J. 2014. Expression and purification of porcine Akirin2 in Escherichia coli. Turk J Biol. 38:339-345.
Chen R, Zhou Z, Cao Y, Bai Y, Yao B. 2010. High yield expression of an AHL-lactonase from Bacillus sp. B546 in Pichia pastoris and its application to reduce Aeromonas hydrophila mortality in aquaculture. Microb Cell Fact. 1-10.doi:10.1186/1475-2859-9-39.
Czajkowski R, Jafra S. 2009. Quenching of acyl-homoserine lactone dependent quorum sensing by enzymatic disruption of signal molecules. Acta Biochim Pol. 5:1-16.
28
Decho AW, Frey RL, Ferry JL. 2011. Chemical challenges to bacterial AHL signaling in the environment. Chem Rev. 111:86-99.doi:10.1021/cr100 311q.
Dong YH, Gusti AR, Zhang Q, Ling Xu J, Hui Zhang L. 2002. Identification of quorum quenching N-acyl homoserine lactonases from Bacillus species. Appl Environ Microbiol. 68:1754-1759.doi:10.1128/AEM.68.4.
Dong YH, Xu JL, Li XZ, Zhang LH. 2000. AiiA, an enzyme that inactivates the acyl homoserine lactone quorum-sensing signal and attenuates the virulence of Erwinia carotovora. Proc Natl Acad Sci USA. 97:3526-3531. doi:10.1073/pnas.97.7.3526.
Dubendorff JW, Studier FW. 1991. Controlling basal expression in an inducible T7 expression system by blocking the target T7 promoter with lac repressor. J Mol Biol. 219:45-59.doi:10.1016/0022-2836(91)90856-2. Durán N, Menck CFM. 2001. Chromobacterium violaceum: a review of
pharmacological and industrial perspectives. Crit Rev Microbiol. 27:201-222.doi:10.1080/20014091096747.
Fang XJ, Tanga ZX, Lia ZH, Zhanga ZL, Shia LE. 2014. Production of a new non-specific nuclease from Yersinia enterocolitica subsp. palearctica: optimization of induction conditions using response surface methodology. Biotechnol Biotec Eq. 28:559-566. doi: 10.1080/13102818.2014.915612. Federle MJ, Bassler BL. 2003. Interspecies communication in bacteria. J Clin
Invest. 112:1291-1299. doi:10.1172/JCI200320195.
Flores AM, González A, Pedroza CJ, Correa E, Rueda NJ, Orduz S. 2014. Identification,cloning and lactonase activity of recombinant protein of N-acyl homoserine lactonase (AiiA) from Bacillus thuringiensis 147-11516 strain. Rev Colomb Biotecnol. 16:153-162.
Galloway WRJD, Hodgkinson JT, Bowden SD, Welch M, Spring DR. 2011. Quorum sensing in gram-negative bacteria: small-molecule modulation of ahl and AI-2 quorum sensing pathways. Chem Rev. 111:28-67.doi:10. 1021/cr100109t.
Gopal GJ, Kumar A. 2013. Strategies for the Production of recombinant protein in Escherichia coli. Protein J. 32: 419-425. doi:10.1007/s10930-013-9502-5. Helgason E, Okstad OA, Caugant DA, Johansen HA, Fouet A, Mock M, Ian H,
Kolstø AB. 2000. Bacillus anthracis, Bacillus cereus, and Bacillus thuringiensis : One species on the basis of genetic evidence. Appl Environ Microbiol. 66:2627-2630.doi:10.1128/AEM.66.6.
Hentzer M, Givskov M, Parsek MR. 2002. Targeting quorum sensing for treatment of chronic bacterial biofilm infections. Lab Medicine. 33:295-303. Hentzer M, Givskov M. 2003. Pharmacological inhibition of quorum sensing for
the treatment of chronic bacterial infections. J Clin Invest. 112:1300-1307. doi:10.1172/JCI200320074.
Ivanova N, Sorokin A, Anderson I, Galleron N, Candelon B, Kapatral V, Bhattacharyya A, Reznik G, Mikhailova N, Lapidus A et al. 2003. Genome sequence of Bacillus cereus and comparative analysis with Bacillus anthracis. Nature. 423:87-91.
29 Kwon, Mi-Jung, Park SL, Kim SK, Nam SW. 2002. Overproduction of Bacillus macerans cyclodextrin glucanotransferase in E.coli by coexpression of GroEL/ES chaperone. J Microbiol Biotechnol. 12:1002-1005.
Lade H, Paul D, Kweon JH. 2014. N-Acyl homoserine lactone-mediated quorum sensing with special reference to use of quorum quenching bacteria in membrane biofouling control. Biomed Res Int. 2014:1-25.doi:org/10.1155/ 2014/162584.
Lee SJ, Park SY, Lee JJ, Yum DY, Koo BT, Lee JK. 2002 Genes encoding the N-acyl homoserine lactone degrading enzyme are widespread in many subspecies of Bacillus thuringiensis. Appl Environ Microbiol. 68:3919-3924.doi:10.1128/AEM.68.8.
Lin YH, Xu JL, Hu J, Wang LH, Ong SL, Leadbetter JR, Zhang LH. 2003. Acyl-homoserine lactone acylase from Ralstonia strain XJ12B represents a novel and potent class of quorum-quenching enzymes. Mol Microbiol. 47: 849-860.
Lindemann A, Pessi, G, Schaefer AL, Mattmann ME, Christensen QH, Kessler A, Hennecke H, Blackwell HE, Greenberg EP, Harwood CS. 2011. Isovaleryl-homoserine lactone, an unusual branched-chain quorum-sensing signal from the soybean symbiont Bradyrhizobium japonicum. Proc Natl Acad Sci USA. 108:16765-16770. doi:10.1073/pnas.1114125108.
Liu D, Lepore BW, Petsko GA, Thomas PW, Stone EM, Fast W, Ringe D. 2005. Three-dimensional structure of the quorum-quenching N-acyl homoserine lactone hydrolase from Bacillus thuringiensis. Proc Natl Acad Sci USA. 102:1882-11887.doi:10.1073/pnas.0505255102.
Marisch K, Bayer K, Puschmann MC, Luchner M, Striedner. 2014. Evaluation of three industrial Escherichia coli strains in fed-batch cultivations during high-level SOD protein production. Microb Cell Fact. 12:1-11.doi:10. 1186/1475-2859-12-58.
McClean KH, Winson MK, Fish L, TaylorA, Chhabra SR, Camara M, DaykinM, Lamb JH, Swift S, Bycroft BW, Stewart GSAB, Williams P. 1997. Quorum sensing and Chromobacterium violaceum: exploitation of violacein production and inhibition for the detection of N-acylhomoserine lactones. Microbiology. 143:3703-3711.
MukhopadhyayaPN, Jha M, MuraleedharanP, GuptaRR, RathodRN, MehtaHH, Khod VK. 2006. Simulation of normal, carrier and affected controls for large scale genotyping of cattle for factor XI deficiency. Genet Mol Res. 5:323-332.
Molina L, Constantinescu F, Michel L, Reimmann C, Duffy B, Defago, G. 2003. Degradation of pathogen quorum-sensing molecules by soil bacteriaa preventive and curative biological control mechanism. FEMS Microbiol Ecol. 45:71-81.doi:10.1016/S0168-6496(03)00125-9.
Momb J, Wang C, Liu D, Thomas PW, Petsko GA, Guo H, Ringe D, Fast W. 2008. Mechanism of the quorum-quenching lactonase (AiiA) from Bacillus thuringiensis. 2. Substrate modeling and active site mutations. Biochemistry. 47:7715-725.
30
Ngai MJM. 2001. Identifying ‘prime suspects’: symbioses and the evolution of multicellularity. Comp Biochem Physiol B. 129:711-723.
Oh MH, Ham JS, Cox JM. 2011. Diversity and toxigenicity among members of the Bacillus cereus group isolated from rice products. Int J Food Microbiol. 152:1-8.doi:10.1021/bi8003704.
Olaofea OA, Burtona SG, Cowanb DA, Harrisona STL. 2010. Improving the production of a thermostable amidase through optimising IPTG induction in a highly dense culture of recombinant Escherichia coli. Ferment Bioeng. 52:19-24.doi:10.1007/s00253-013-4933-7.
Pan J, Huang T, Yao F, Huang Z, Powell C, Qiua S, Guana X. 2007. Expression and characterization of aiiA gene from Bacillus subtilis BS-1. Microbiol Res. 163:711-716.doi:10.1016/j.micres.2007.12.002.
Parsek MR, Greenberg EP. 2000. Acyl-homoserine lactone quorum sensing in Gram-negative bacteria: A signaling mechanism involved in associations with higher organisms. Proc Natl Acad Sci USA. 97:8789-8793.
Park SY, Hwang BJ, Shin MH, Kim JA, Kim HK, Lee JK. 2006. N-acylhomoserine lactonase-producing Rhodococcus spp. with different AHL- degrading activities. FEMS Microbiol Lett. 261:102-108.
Park SY, Lee SJ, Oh TK, Oh JW, Koo BT, Yum DY, Lee JK. 2003. AhlD, an N-acylhomoserine lactonase in Arthrobacter sp. and predicted homologues in other bacteria. Microbiology. 149:541-1550.doi:10.1099/mic.0.26269-0. Patil S, Sandberg A, Heckert E, Self W, Seal S. 2007. Protein adsorption and
cellular uptake of cerium oxide nanoparticles as a function of zeta potential.
Biomaterials. 28:4600-4607. doi:10.1016/j.biomaterials.2007.07. 029. Picaud S, Olsson ME, Brodelius PE. 2007. Improved conditions for production of
recombinant plant sesquiterpene synthases in Escherichia coli. Protein Expr Purif. 51:71-79.doi:10.1016/j.pep.2006.06.025.
Piubelli L, Campa M, Temporini C, Binda E, Mangione F, Amicosante M, Terreni, Marinelli F, Pollegioni L. 2013. Optimizing Escherichia coli as a protein expression platform to produce Mycobacterium tuberculosis immunogenic proteins. Microb Cell Fact. 12:1-14.doi:10.1186/1475-2859-12-115.
Ratelade J, Miot MC, Johnson E, Betton JM, Mazodier P, Benaroudj N. 2009. Production of recombinant proteins in the ion-deficient BL21(DE3) strain of Escherichia coli in the absence of the DnaK chaperone. Appl Environ Microbiol. 75:3803-3807.doi:10.1128/AEM.00255-09.
Ryan RP, Dow JM. 2008. Diffusible signals and interspecies communication in bacteria. Microbiology. 154: 1845-1858.doi:10.1099/mic.0.2008/017871-0.
Salgın S, Salgın U, Bahadır S. 2012. Zeta Potentials and isoelectric points of biomolecules: The effects of ion types and ionic strengths. Int J Electroche. Sci.7:12404-12414.
Salgın S, Takaç S, Özdamar TH. 2006. Adsorption of bovine serum albumin on polyether sulfone ultrafiltration membranes: Determination of interfacial interaction energy and effective diffusion coefficient. J Membrane Sci. 278: 251-260.doi:10.1016/j.memsci.2005.11.008.