PADA TANAMAN MENTIMUN (Curcumis sativus L.) DAN CABAI (Capsicum annuum L.)
HAZEN ARRAZIE KURNIAWAN
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
Dengan ini saya menyatakan tesis “ Neraca Kehidupan Kutukebul, Bemisia tabaci Gennadius (Hemiptera: Aleyrodidae) Biotipe-B dan Non-B
Pada Tanaman Mentimun (Curcumis sativus L.) dan cabai (Capsicum annuum L.)” adalah karya saya sendiri dan belum diajukan dalam
bentuk apapun ke perguruan tinggi manapun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebut dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Bogor 4 Febuari 2007
Gennadius (Hemiptera:Aleyrodidae) Biotype-B and Biotype non-B in Cucumber
(Curcumis sativus L.) and Chilli (Capsicum annum L.). Under the direction of
PURNAMA HIDAYAT and HERMANU TRIWIDODO.
Bemisia tabaci (Hemiptera: Aleyrodidae) has been widely recognized as the
important vegetable pest in Indonesia. Biological information of this pest is needed to
make control strategies of the pest. The objective of the research was to study life
table of two the biotypes of B. tabaci (biotype-B and non-B) on cucumber and chilli.
The result of the study showed that the host plant has a strong effect on the
development time of B. tabaci. Survival of both biotypes were recorded to higher in
cucumber than chilli. Developmental time of B. tabaci biotype-B from egg to adult
was about 31,27 days in cucumber, and 33,27 days in chilli. Developmental time of
B. tabaci biotype non-B from egg to adult was about 29,60 day in cucumber, and
30,86 in day chilli. Fecundity was 130,30±26,27 in cucumber and 94,48 in chilli. On
cucumber, an intrinsic rates of increase (r) was about 0,10 for biotype-B and 0,12 for
biotype non-B. On chilli, intrinsic of increase (r) was about 0,17 in biotype-B and
0,07 in biotype non-B. Doubling time of the B. tabaci biotype-B was about 3,96 in
chilli and 6,45 in cucumber. Doubling time of the B. tabaci non-B was about 5,71 in
cucumber and 9,21 in chilli.
HAZEN ARRAZIE KURNIAWAN. Neraca Kehidupan Kutukebul, Bemisia tabaci Gennadius (Hemiptera: Aleyrodidae) Biotipe-B dan non-B Pada Tanaman Mentimun (Curcumis sativus L.) dan Cabai (Capsicum Annuum L.). Dibimbing oleh PURNAMA HIDAYAT dan HERMANU TRIWIDODO.
Bemisia tabaci (Hemiptera: Aleyrodidae) merupakan salah satu hama penting yang dapat menyebabkan kerusakan langsung dengan menghisap cairan tanaman dan tidak langsung dengan menularkan penyakit pada tanaman sayuran di Indonesia. Informasi mengenai biologinya dibutuhkan untuk menyusun strategi pengendalian hama. Hasil penelitian menunjukkan tanaman inang dapat mempengaruhi waktu perkembangan B. tabaci. Daya bertahan hidup kedua biotipe tercatat lebih tinggi pada mentimun daripada cabai. Penelitian ini mempelajari neraca kehidupan dua biotipe B. tabaci (biotipe-B dan non-B) pada tanaman mentimun dan cabai. Lama hidup B. tabaci biotipe-B dari telur hingga dewasa, adalah 31,27±17,70 hari pada mentimun, dan 33,27±4,60 pada cabai. Keperidian sebesar 130±18,50 telur pada mentimun dan 82,05±22,57 telur pada cabai. Selanjutnya lama hidup B. tabaci non-B dari telur hingga dewasa adalah 29,60±3,80 hari pada mentimun dan 30,86±6,32 hari pada cabai. Keperidian sebesar 130,30±26,27 telur pada mentimun dan 94,48 telur pada cabai. Pada mentimun, nilai laju pertambahan intrinsik (r) adalah 0,10 dengan nilai waktu untuk populasi berlipat ganda (DT) 6,45 hari pada biotipe-B dan 0,12 dengan nilai berlipat populasi 5,71 hari pada biotipe non-B. Pada tanaman cabai laju pertambahan intrinsik (r) sebesar 0,17 pada B. tabaci biotipe-B, dengan nilai waktu untuk populasi berlipat ganda (DT) 3,96 hari., dan 0,07 dengan nilai berlipatan populasi 9,21 hari pada biotipe non-B.
© Hak cipta milik Institut Pertanian Bogor, tahun 2007. Hak cipta dilindungi
Dilarang mengutip dan memperbayak tampa izin tertulis dari Institut Pertanian Bogor, sebagian atau seluruhnya dalam bentuk apapun, baik cetak, fotocopy,
(HEMIPTERA: ALEYRODIDAE) BIOTIPE-B DAN NON-B PADA TANAMAN MENTIMUN (Curcumis sativus L.)
DAN CABAI (Capsicum annuum L.)
HAZEN ARRAZIE KURNIWAN
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains pada
Departemen Proteksi Tanaman
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
(Capsicum annuum L.)
Nama : Hazen Arrazie Kurniawan NIM : A451040091
Disetujui
Komisi Pembimbing
Dr. Ir. Purnama Hidayat, M.Sc. Dr. Ir. Hermanu Triwidodo M.Sc. Ketua Anggota
Diketahui
Ketua Program studi Dekan Sekolah Pascasarjana Entomologi/Fitopatologi
Dr. Ir. Sri. Hendrastuti Hidayat, M.Sc. Prof. Dr. Ir. Kahiril Anwar Notodiputro, MS.
Puji syukur penulis panjatkan kepada Allah Subhana Wata’ala atas karunia-Nya sehingga tesis dengan judul ”Neraca Kehidupan Kutukebul, B. tabaci Gennadius (Hemiptera: Aleyrodidae) Biotipe-B dan Non-B Pada Tanaman Mentimun (Curcumis sativus L.) dan Cabai (Capsicum annuum L.)” dapat diselesaikan, yang merupakan salah satu syarat untuk memperoleh gelar Magister Sains pada Fakultas Pascasarjana, Institut Pertanian Bogor.
Penulis menyampaikan penghargaan dan terima kasih kepada Dr. Ir. Purnama Hidayat, M.Sc., Dr. Ir. Hermanu Triwidodo, M.Sc., Dra. Dewi
Sartiami, M.Si., yang telah memberikan ilmu, pengarahan, dorongan dan bimbingannya selama ini. Kepada seluruh staf pengajar Departemen Perlindungan Tanaman penulis juga mengucapkan terima kasih.
Terima kasih kepada Ibunda dan Ayahanda tercinta di Medan yang telah mencurahkan tenaga, pikiran, doa, dan materi demi kelancaran pendidikan penulis di Pascasarjana Institut Pertanian Bogor. Terima kasih juga penulis sampaikan kepada Krida Laksi, SP., Raziq Rasidin Ardino, Rayhan Aghani Amiseno yang telah memberikan semangat dan dukungan dalam melewati masa studi di Bogor.
Ucapan terima kasih juga penulis sampaikan kepada rekan-rekan di Laboratorium Taksonomi (bu Aisyah, bu Eva, Echa), rekan-rekan Laboratorium Virologi (embak Tuti, pak Ray, Artha, Reno, Dedek kambing, mas Emput), rekan-rekan Laboratorium Biologi Parasitoid dan Predator (mas Jalu, mas Bandung, bu Yanti, Wina), rekan-rekan Kos (Rizal, Agung, faldi, Heru, danar) ,dan seluruh pihak yang telah memberikan saran dan masukan yang berarti.
Bogor, Febuari 2007
Armansyah dan ibunda Siti Hafsah. Penulis merupakan putra pertama dari dua bersaudara.
DAFTAR TABEL………... xii
DAFTAR GAMBAR………... xiii
DAFTAR LAMPIRAN………... xiv
PENDAHULUAN Latar Belakang……….... 1
Tujuan Penelitian……….... 3
Hipotesis………... 3
TINJAUAN PUSTAKA Taksonomi dan Morfologi………... 4
Biologi...……….... 6
Keragaman Biotipe B. tabaci... 6
Hubungan Virus dengan B. tabaci………... 8
Neraca Kehidupan………... 9
BAHAN DAN METODE Tempat dan Waktu Penelitian……….... 11
Persiapan Pengujian B. tabaci pada Tanaman Uji a. Perbanyakan Serangga... 11
b. Persiapan Tanaman Uji... 12
c. Persiapan kurungan dan Penanaman... 12
Pembuatan Preparat Mikroskop dan Identifikasi... 13
Pengujian NeracaKehidupan B. tabaci Biotipe-B dan Non-B Di Laboratorium a. Pengujian Neraca Kehidupan... 13
b. Parameter Neraca Kehidupan... 14
HASIL DAN PEMBAHASAN Identifikasi……….. 15
Siklus hidup B. tabaci Biotipe-B dan non-B pada Tanaman Mentimun dan Cabai... 15
Lama hidup dan Keperidian B. tabaci Biotipe-B dan non-B... 18
Neraca Kehidupan B. tabaci Biotipe-B dan Non-B pada Tanaman Mentimun... 23
Neraca Kehidupan B. tabaci Biotipe-B dan Non-B pada Tanaman Cabai... 23
PEMBAHASAN UMUM ... 25
KESIMPULAN DAN SARAN... 29
xi
1. Keragaman biotipe B. tabaci di berbagai negara di dunia ... 7
2. Lama stadia B. tabaci biotipe-B dan non-B pada tanaman mentimun... 16
3. Lama stadia B. tabaci biotipe-B dan non-B pada tanaman cabai... 17
4. Lama hidup dan keperidian B. tabaci biotipe-B dan non-B pada tanaman mentimun... 19
5. Lama hidup hidup dan keperidian B. tabaci biotipe-B dan non-B pada tanaman cabai... 19
6. Neraca kehidupan B. tabaci biotipe-B dan non-B pada tanaman mentimun ... 24
xii
1. Gambar imago B. tabaci…………...………. 4
2. Gambar kantung pupa B. tabaci... 16
3. Gejala penyakit daun keriting kuning pada tanaman tomat... 8
4. Gambar Kurungan pemeliharaan dan perbanyakan B. tabaci... 11
5. Gambar kurungan pemeliharaan tanaman... 12
6. Kurva lama hidup (lx) dan rataan keperidian betina per hari (mx) B. tabaci biotipe-B pada tanaman mentimun………. 21
7. Kurva lama hidup (lx) dan rataan keperidian betina per hari (mx) B. tabaci non-B pada tanaman mentimun... ... 21
8. Kurva lama hidup (lx) dan rataan keperidian betina per hari (mx) B. tabaci Biotipe-B pada tanaman cabai... 22
xiii
1. Siklus hidup B. tabaci biotipe-B di tanaman cabai………... 34
2. Siklus hidup B. tabaci non-B di tanaman mentimun………... 36
3. Siklus hidup B. tabaci non-B di tanaman cabai………... 38
4. Siklus hidup B. tabaci biotipe-B di tanaman mentimun………... 40
5. Lama hidup betina, prapeneluran, dan peneluran B. tabaci Biotipe-B pada tanaman mentimun dan cabai... 42
6. Lama hidup betina, parapeneluran, dan peneluran B. tabaci non-B pada tanaman mentimun dan cabai... 44
7. Keperidian B. tabaci biotipe-B dan non-B pada tanaman mentimun dan cabai... 46
8. Neraca kehidupan B. tabaci biotipe-B pada tanaman mentimun... 48
Latar Belakang
Sayuran merupakan tanaman hortikultura penting yang mengandung nutrisi
tinggi, terutama vitamin, mineral, dan serat yang tinggi. Beberapa jenis sayuran penting
seperti cabai, tomat, bawang merah, kentang, mentimun, dan terung mempunyai nilai
ekspor yang tinggi. Penurunan produksi sayuran, sering disebabkan oleh serangan
hama. Salah satu serangga yang dapat menyebarkan penyakit pada tanaman sayuran
adalah B. tabaci, yang diketahui dapat menimbulkan gagalnya panen di beberapa daerah
di Indonesia.
Bemisia tabaci Gennadius (Hemiptera: Aleyrodidae) dengan nama umum
kutukebul tembakau mulai dikenal lebih dari 100 tahun yang lalu sebagai hama yang
merugikan pada beberbagai tanaman di daerah tropik maupun sub tropik (Oleivera et
al. 2001). Serangga ini dapat menimbulkan kerusakan langsung pada tanaman dengan
menghisap cairan makanan (Van den Ven et al. 2000), menimbulkan gangguan
fisiologis (Mecollum et al. 2000), dan memacu tumbuhnya cendawan embun jelaga
pada tanaman inangnya. B. tabaci diketahui dapat berperan sebagai vektor berbagai
virus yang dapat menyebabkan penyakit tanaman ( Legg et al. 2002).
Di Indonesia B. tabaci diketahui pertama kali pada tahun 1938 sebagai penyebab
penyakit kerupuk di tanaman tembakau di daerah Sumatra dan Jawa, yang ditularkan
dari gulma, yaitu Ageratum sp., Synedrella sp., dan Eupatorium odoratum. Namun pada
waktu itu kerusakan yang ditimbulkan oleh B. tabaci dianggap tidak berarti
(Kalshoven 1981). Hasil laporan Sulandari (2004), diketahui adanya peningkatan
kerusakan, yaitu ditemukannya 29 tanaman yang terserang penyakit keriting kuning
yang dibawa dan ditularkan oleh B. tabaci. Penyakit keriting kuning merupakan salah
satu virus yang ditularkan B. tabaci yang disebut geminivirus. Virus ini ditakuti oleh
petani, karena menyebabkan tanaman menjadi kerdil dan tidak berbuah. Beberapa
tanaman yang terserang geminivirus, seperti African Cassava Mosaic Geminivirus
(ACMV)) dapat menginfekti tanaman ubi kayu di Afrika, Catton Leaf Cruemple Virus
(CLCV) dapat menginfeksi tanaman kapas di California, dan Ageratum Yellowvein
Aidawati (2006) melaporkan minimal ada dua biotipe B. tabaci setelah
dilakukan survai ke beberapa tanaman tomat yang ada di Jawa Barat (Bogor), Jawa
Tengah (Boyolali, Mangelang, Semarang), dan Daerah Istimewa Yogykarta (Sleman,
Kuloprogo, dan Bantul), yaitu biotipe-B dan non-B. Timbulnya biotipe B. tabaci
diketahui pada tahun 1986 di Florida, pada tanaman Euphorbia pulcherrima Willdenow
menimbulkan gejala keperakan. Gejala keperakan ini terjadi di beberapa negara bagian
di Amerika. Hingga saat ini fenomena timbulnya biotipe pada B. tabaci menjadi
masalah di beberapa negara, seperti di Amerika pada tanaman kapas, di Afrika pada
tanaman ubi kayu, dan di Indonesia pada tanaman cabai dan tomat. Penentuan
biotipe-B juga didasarkan pada kemampuan berkembangbiak B. tabaci yang tinggi, daya tahan
terhadap beberapa insektisida (Harowitz et al. 1998; Belloti dan Arias 2000), dan
kemampuan penularan virus yang berbeda antar biotipe B. tabaci (Aidawati 2006).
Menurut Costa dan Brown (1990) perbedaan biologi antara B. tabaci biotipe-B dan
biotipe-A, terlihat dari tingkat keperidiannya, tanaman inang yang luas, dan ketahanan
pada beberapa insektisida yang lebih baik pada B. tabaci biotipe-B serta kemampuannya
dalam menimbulkan gejala keperakan. Hasil survai yang dilakukan oleh Lima et al.
(1991) di Brazil pada 57 lokasi dengan 27 tanaman yang berbeda di peroleh fakta bahwa
B. tabaci biotipe-B lebih dominan dijumpai dari pada biotipe-A .
Penelitian mengenai biotipe pada B. tabaci telah banyak dilakukan di beberapa
negara di Eropa. Di Indonesia informasi mengenai biologi B. tabaci biotipe-B dan
non-B masih terbatas. Oleh karena itu, penelitian mengenai neraca kehidupan B. tabaci
biotipe-B dan non-B pada tanaman mentimun dan cabai penting dilakukan. Menurut
Naranjo dan Ellworty (2005), bentuk neraca kehidupan merupakan metode untuk
menggambarkan dan mengukur kematian dalam suatu populasi. Maka diharapkan
pengujian neraca kehidupan dapat menjadi informasi dalam pengambilan keputusan
Tujuan Penelitian
Penelitian ini bertujuan untuk mengetahui neraca kehidupan B. tabaci
(Hemiptera: Aleyrodidae) biotipe-B dan non-B pada tanaman cabai
(Capsicum annuum L.) dan tanaman mentimun (Curcumis sativus L.).
Hipotesis
1. Terdapat perbedaan neraca kehidupan antara B. tabaci biotipe-B dan non-B.
2. Tanaman yang berbeda akan memberikan pengaruh pada siklus hidup dan
TINJAUAN PUSTAKA
Taksonomi dan Morfologi
Kutukebul tembakau (tobacco whitefly) atau B. tabaci (Gambar 1) digolongkan ke
dalam ordo Hemiptera, subordo Sternorrhyncha, famili Aleyrodidae termasuk ke dalam
superfamili Aleyrodoidea (Martin et al. 2000). Umumnya tersebar di daerah tropik dan
subtropik, bersifat polifag, dan diketahui sebagai vektor virus yang dapat menyebabkan
penyakit pada tanaman (Kalshoven 1981).
Menurut Kalshoven (1981) ciri morfologi B. tabaci adalah sebagai berikut: tubuh
imago berwarna kuning, sayap tertutup oleh tepung berwarna putih, panjang tubuh 1-1,5
mm. Telur yang baru diletakkan berwarna kekuning-kuningan dan biasanya tertutup
oleh lilin. Setelah 24 jam, warna telur akan berubah menjadi coklat. Nimfa instar satu
berbentuk bulat panjang, berwarna hijau cerah, dengan panjang tubuhnya 0,22 mm dan
lebar 0,13 mm. Nimfa instar dua berwarna hijau gelap dengan panjang tubuhnya 0,28
mm, lebar 0,17 mm, antena sangat pendek, dan tungkainya tereduksi. Pupanya
berbentuk bulat panjang, di bagian toraks agak melebar, cembung, dan abdomen tampak
jelas. Terdapat satu pasang seta kauda (cauda setae) pada ujung anal. Vasiform orifice
terdapat di daerah sebelum ujung posterior puparium, berbentuk segitiga, dan ukurannya
lebih panjang dari panjang alur kaudal (caudal furrow). Operkulumnya (operculum)
hampir separuh bagian menutupi bagian vasiform orifice (Gambar 2).
Lama hidup imago betina sekitar enam hari, tetapi mampu hingga 60 hari pada
kondisi tertentu. Umumnya imago lama hidup jantan lebih pendek yaitu antar 9-17 hari
(CABI 2005). Kopulasi berlangsung kurang lebih dua jam setelah serangga menjadi
dewasa, yang dapat terjadi pada saat siang dan malam hari.
1,5 mm
Gambar 2 Kantung pupa B. tabaci; keterangan : dds: dorsal disc spine, lms:
longitudinal moulting suture, ttc: thoracic tracheal, ttf: toraccic tracheal fold, r: rostrum, l : leg, tms: transverse moulting suture, m: margin, sma:
sub marginal area, vo: vasiform orifice, asms: sub marginal seta, ds:
dorsal seta, psms: posterior marginal seta, o: operculum, li: lingula, cf:
caudal furrow, cs: caudal seta (Martin 2000).
ttc ttf
r
l
tms
m
sma
vo
o
li
cf
cs asms3
asms4
psms5
psms4
psms3
psms2
Psms1 ds1
ds2
ds3
ds4
ds5 asms2
asms1
Biologi
Setelah terjadinya kopulasi antara imago jantan dan betina, selanjutnya imago
betina B. tabaci akan meletakkan telur secara satu persatu pada permukaan daun bagian
bawah, dengan cara menyisipkan telurnya ke dalam jaringan epidermis daun
menggunakan alat peletak telur. Telur-telur tersebut diletakkan secara tegak lurus pada
permukaan daun. Pada umumnya imago betina lebih memilih meletakkan telur pada
daun yang lebih muda dari pada daun yang lebih tua.
Telur- telur yang dihasilkan oleh imago betina B. tabaci dipengaruhi oleh faktor
tanaman inang dan kondisi suhu. Samudra dan Naito (1991) menyatakan bahwa
telur-telur yang dihasilkan oleh betina B. tabaci pada tanaman kapas adalah 81-308 telur
per betina pada suhu 25-27oC. Tsai dan Wang (1996) melaporkan bahwa, telur-telur
yang dihasilkan oleh imago betina B. tabaci dengan kondisi suhu 25oC adalah 167,6
telur per betina pada tanaman tomat, 77,5 telur per betina pada tanaman ubi kayu, dan
66 telur pada tanaman mentimun. Kisaran waktu perkembangan dari telur hingga
stadium dewasa adalah 23,6 hari pada suhu 25oC dan 17,8 hari pada suhu 27,5oC.
Powel dan Bellows (1992) melaporkan bahwa mortalitas untuk instar satu, instar
dua, instar tiga, dan instar empat pada tanaman mentimun dan tomat dengan suhu 20oC
dan 32oC, masing-masing 17-18%, 2-16%, 3-14%, dan 2-24%. Menurut Burban et al.
(1992), populasi B. tabaci yang berasal dari daerah geogerafis yang berbeda
menunjukkan perbedaan dalam kemampuan reproduksi dan makan.
Keragaman Biotipe B. tabaci
Keragaman taksonomi dimulai pada tahun 1986 dengan penemuan kutukebul
pada tanaman Euphorbia pulcherrima Willdenom, dan beberapa tanaman di daerah
Florida (tembakau, kapas, tomat, kedelai, dan terung). Gejala yang terdapat di tanaman
squash adalah daun berwarna keperakan (silver leaf) dan terjadi di beberapa negara
bagian di Amerika ( Arizona, California, Hawai, New York, Ohio, Tennessee, dan
Texas). Hasil penemuan dan pengujian genetik tersebut membuktikan bahwa, penemuan
kutukebul di tanaman E. pulcherrima Willdenom dianggap sebagai timbulnya strain
yang berbeda (Harris et al. 2001). Fakta yang mendukung timbulnya biotipe ini adalah
pengujian menggunakan elektromorphis esterase yang memperlihatkan perbedaan pita
terlihat adanya perbedaan antara B. tabaci biotipe-A dan biotipe-B, dengan tingkat
kemiripan mencapai 90%. Hasil tersebut digunakan sebagai pembanding biotipe
B. tabaci di dunia, seperti yang dilakukan oleh De barro et al. (1998) yang melakukan
survei di daerah Pasifik.
Tabel 1 Keragaman biotipe B. tabaci di berbagai negara di dunia
Biotipe Negara
A Mexico, USA
AN Australia, Niue, Papuan N.Guinea, Solomon Island, Vanuatu
B
American Samoa, Antigua, Australia, Austria, Belize, Brazil, Canada, Canary Island, Cook Island Cyprus, Denmark, Dom. Republik, Egypt, Fiji, France Grenada, Guandaloupe, Guam, Guatemala, Honduras, Iran, Israel, Italy, Japan, Jordan, Korea, Mexico, Marshall Island, N. Mariana Island, Netherland, New Celedonia, Nicaragua, Niue, Panama, Poland, Poertorico south Africa, Spain, St. Kitts, Sudan, Tobago, Trinidat, USA, Yemen
B2 Yemen
C Costa Rica
Cassava Ivory Coast
D Nicaragua
E Benin
H India
I India
J Nigeria
K Pakistan
L Sudan
M Turkey
N Puerto Rico
NA AM. Samoa, Fed. St. Micronesia, Fiji, Guam, Kiribian, Marshal Is, N. Mariana IS, Nauru, Niue, Palau, Tonga, Tuvalu, Western Samoa
Okra Ivory Coast
P Nepal
Q Spain
R Colombia
Hasil survei menyebutkan adanya tersebut terdapat perbedaan pola pita esterase
pada B. tabaci yang berasal dari daerah Sudan (biotipe-L), india (biotipe-H dan
biotipe-G), Pakistan (biotipe-K), dan New Zeland (biotipe belum diketahui). Hasil
penelitian Bark dan Markham menyatakan tentang penemuan 20 biotipe kutukebul
(Perring 2001). Keragaman biotipe B. tabaci dan lokasi di temukannya dapat dilihat
pada Tabel 1.
Hubungan Virus dengan B. tabaci
Di Amerika dan Karibian adanya populasi B. tabaci atau yang dikenal sebagai
kutukebul tembakau yang mempunyai kemampuan yang berbeda, dalam hal kisaran
inangnya, kemampuan menularkan virus, dan tingkat reproduksi yang sangat tinggi,
serta tidak dapat melakukan kopulasi dengan kutukebul tembakau yang sudah ada
sebelumnya. Biotipe baru tersebut kemudian dikenal dengan kutukebul tembakau
biotipe-B dan ada yang menyebutnya sebagai spesies B. argentifolii. Selain biotipe-B,
terdapat tujuh kelompok biotipe B. tabaci, dan biotipe-B tersebut sangat potensial dalam
menularkan Geminivirus pada berbagai tanaman budidaya (Perring 2001), CabLCV
pada tanaman kubis (Hunter et al. 1998), dan ToMV pada tanaman tomat (Gambar 3).
Secara alamiah begomovirus tidak menular melalui benih tapi hanya menular dengan
bantuan serangga B. tabaci dari tanaman satu ke tanaman lainnya.
Gambar 3 Gejala penyakit daun keriting kuning pada tanaman tomat
Peranan kutukebul tembakau sebagai vektor Geminivirus yang menyebabkan
penyakit sudah banyak dilaporkan. Pada umumnya hubungan virus dengan vektornya
(non transovarial transmission). Walaupun demikian ada pula Geminivirus yang dapat
diturunkan ke generasi berikutnya, misalnya TYLCV (Czosnek et al. 2001), dan
TYLCV-sar (Bosco et al. 2001). Geminivirus dapat diketahui melalui tehnik
PCR-RFLP yang menyerang tanaman cabai di Indonesia (Hidayat et al. 1999).
Neraca Kehidupan
Price (1997) menyatakan bahwa, untuk mengetahui pertumbuhan populasi suatu
spesies harus diketahui terlebih dahulu jumlah keturunan yang dihasilkan oleh seekor
betina pada interval umur selama hidupnya, dan jumlah individu pada setiap interval
umur tesebut. Untuk mengetahui hal tersebut, pendekatannya dapat disederhanakan
dengan hanya mengikuti betina saja dalam satu populasi.
Bila populasi betina dan perbandingan kelaminnya di ketahui maka dengan
mudah diketahui populasi jantan dan populasi total. Jika x melambangkan umur
individu pada setiap periode dan tiap bagian populasi serangga betina dalam kohort
yang dapat betahan hidup pada umur tersebut dilambangkan dengan lx, maka statistik
dari kohort serangga tersebut dapat ditabulasikan. Dari nilai lx tersebut dibuat kurva
bertahan hidup, Price (1997) berpendapat bila nilai lx di buat dalam bentuk skala
logaritma akan terlihat lebih informatif.
Kurva daya bertahan hidup secara umum dibagi dalam tiga tipe. Kurva daya
bertahan hidup tipe I dimiliki oleh populasi yang tingkat kematiannya rendah pada
umur muda dan tinggi pada umur tua, dan konstan pada semua tingkatan umur. Kurva
daya bertahan hidup tipe II dimiliki oleh populasi yang tingkat kematiannya konstan
pada semua tingkatan umur. Kurva daya bertahan hidup tipe III dimiliki oleh populasi
yang tingkat kematiannya tinggi pada umur muda dan rendah pada umur tua (Wilson
dan Bossert 1971; Cotgreave dan Forseth 2002).
Menurut Hasibuan (1988), neraca kehidupan berisi semua aspek demografi suatu
populasi, yang terdiri dari tujuh buah lajur, yaitu usia (x), daya bertahan hidup (lx), laju
kematian individu di dalam kelas usia x (dx), proporsi individu yang masuk ke dalam
kelas usia x tetapi mati di kelas usia tersebut (qx), panjang waktu hidup semua individu
yang tersisa dari semua individu yang mencapai usia x (Lx), jumlah waktu hidup yang
tersisa dari semua individu yang mencapai usia x (Tx) dan harapan hidup suatu individu
Hanya pada umur tertentu saja serangga betina mempunyai kemampuan untuk
menghasilkan keturunan sedangkan sebagian besar hidupnya adalah masa muda dan tua
yang tidak produktif. Dengan melakukan pengamatan langsung didapat nilai mx yaitu
jumlah anak betina yang di hasilkan pada umur x oleh seekor induk betina yang masih
hidup pada umur x tersebut. Total nilai mx disebut laju reproduksi kasar (gross
reproduction rate) yaitu total anak betina yang dihasilkan dari seekor induk betina
selama masa hidupnya (Price 1997).
Pertumbuhan populasi tergantung dari jumlah induk betina yang masih bertahan
hidup (lx) dan kemampuan individu dalam menghasilkan keturunan (mx) yang disebut
sebagai laju reproduksi bersih dari seekor betina di dalam populasi . Total anak betina
yang dihasilkan dari rataan induk betina di dalam populasi tersebut disebut laju
reproduksi (Ro), atau didefenisiskan sebagai jumlah anak betina yang menggantikan
secara sempurna seekor induk betina dalam satu generasi. Suatu populasi dikatakan
stabil bila Ro = 0, tetapi bila Ro > 1 populasi akan bertambah dan bila Ro < 1 populasi
akan berkurang. Bila Ro suatu spesies diketahui maka lamanya suatu generasi (T) dapat
BAHAN DAN METODE
Tempat dan Waktu
Penelitian dilakukan di laboratorium Taksonomi Departemen Proteksi Tanaman,
Fakultas Pertanian, Institut Pertanian Bogor, sedangkan perbanyakan serangga uji dan
perbanyakan tanaman uji dilakukan rumah kaca dan kasa Cikabayan. Penelitian dimulai
dari bulan April sampai Agustus 2006.
Persiapan Pengujian B. tabaci pada Tanaman Uji a. Perbanyakan Serangga
Serangga yang digunakan adalah B. tabaci biotipe-B yang berasal dari tanaman
brokoli (Brassica oleraceae var. Sakata), sedangkan B. tabaci non-B berasal dari
tanaman terung (Solanum melongena L.). Kedua biotipe tersebut merupakan koleksi
dari laboratorium Virologi Departemen Proteksi Tanaman, Fakultas Pertanian, Institut
Pertanian Bogor, yang sengaja dikembangbiakkan dan diperbanyak untuk kepentingan
penelitian. Perbanyakan kedua biotipe tersebut dilakukan di rumah kaca dan kasa
Departemen Proteksi Tanaman di Cikabayan. B. tabaci biotipe-B dipelihara pada
tanaman brokoli, sedangkan B. tabaci non-B dipelihara pada tanaman tembakau
(Nicotiana tabacum L.) di dalam sebuah kurungan kasa yang berbeda (Gambar 4).
b. Persiapan Tanaman Uji
Tanaman yang diuji adalah cabai (Capsicum annuum L.) var TM-888
(Solanaceae), dan mentimun (Curcumis sativus L.) var Venus (Cucurbitaceae). Selain
pertimbangan famili, pemilihan tanaman uji juga didasarkan pada jenis tanaman yang
umumnya ditanam petani di Jawa Barat (Bogor) secara tumpang sari, benih cabai dan
mentimun yang akan digunakan diperoleh dari kios pertanian.
c. Persiapan Kurungan dan Penanaman
Benih tanaman mentimun dan cabai di semai menggunakan nampan (bak
penyemaian) dengan media semai adalah tanah kompos yang diperoleh dari kios
pertanian. Pemindahan tanaman mentimun ke dalam kantung plastik (tinggi 20 cm,
diameter 20 cm) dilakukan pada saat umur tanaman berumur 20 hari, dan cabai berumur
30 hari. Pemberian ajir bambu setinggi kurungan pada tanaman mentimun, digunakan
sebagai penunjang.
Setelah benih dipindahkan ke kantung plastik, kemudian di masukkan ke dalam kurungan kasa berbentuk kubus (panjang 1,5 m, tinggi 1,5 m, dan lebar 1,5 m). Bahan
penutup bagian depan kurungan menggunakan plastik kaca tipis, sedangkan di bagian
sisi lainya menggunakan kain kasa (Gambar 5). Tujuan pemakaian kurungan kasa
adalah untuk sterilisasi tanaman uji dari organisme pengganggu tanaman sebelum
digunakan.
Pembuatan Preparat Mikroskop dan Identifikasi
Puparium yang dikoleksi dipindahkan ke dalam alkohol 95% yang ditempatkan
pada gelas arloji, lalu dipindahkan secara hati-hati ke dalam tabung reaksi yang berisi
larutan KOH 10% kemudian dipanaskan selama 5-10 menit (hingga transparan), setelah
itu di cuci dengan akuades sebanyak dua kali, kemudian masukkan ke dalam alkohol
50% selama 10 menit. Selanjutnya ditambahkan tiga tetes fuchsin dan diamkan selama
20 menit, lalu tambahkan lagi dengan satu tetes asam asetat glasial, diamkan selama
± 10 menit. Setelah itu puparium dimasukkan ke dalam alkohol 80% selama 5-10 menit
lalu diganti dengan alkohol absolut selama 10 menit. Kemudian masukkan ke dalam
asam asetat glasial selama 10 menit, lalu diganti lagi dengan alkohol absolut selama
5-10 menit. Selanjutnya dimasukkan ke dalam minyak cengkeh selama 10 menit.
Setelah itu puparium ditempatkan pada gelas objek dengan menggunakan
canada balsam. Preparat mikroskop yang telah jadi, dikeringkan di atas pemanas dan
diidentifikasi dengan kunci identifikasi Martin (1987).
Pengujian Neraca Kehidupan B. tabaci Biotipe-B dan Non-B di Laboratorium a. Pengujian Neraca Kehidupan
Pengujian neraca kehidupan B. tabaci biotipe-B dan non-B dilakukan di
laboratorium Departemen Proteksi Tanaman, Fakultas Pertanian, Institut Pertanian
Bogor. Untuk memperoleh telur B. tabaci biotipe-B dan non-B, pada tanaman cabai
digunakan plastik mika berbentuk tabung selinder (tinggi 25 cm, diameter 8 cm),
bagian atasnya dilapisi oleh kain kasa. Pada bagian tengah dari plastik mika terdapat
lubang yang berfungsi untuk memasukkan imago B. tabaci. Ukuran tabung pada
tanaman mentimun disesuaikan dengan tinggi tanaman. Setelah itu ke dalam tabung
plastik dimasukkan lima pasang B. tabaci. Tahap awal pengujian dan pengamatan
dimulai dari fase telur , nimfa, pupa, dan imago. Telur yang digunakan untuk pengujian
adalah sebanyak 100 telur dan diamati setiap hari dengan bantuan mikroskop binokuler,
untuk mengetahui usia, kematian, lama waktu hidup, jumlah generasi yang dihasilkan
satu betina, dan ukuran setiap stadianya. Pengujian di laboratorium dilakukan dengan
menggunakan B. tabaci biotipe-B dan non-B generasi ke dua dari masing-masing
b. Parameter Neraca Kehidupan
Parameter neraca kehidupan digunakan untuk melihat hubungan preferensi
kutukebul terhadap tanaman yang diujikan. Untuk menentukan kesesuaian inang
biasanya digunakan parameter pertumbuhan seperti lama stadia pradewasa, lamanya
stadia imago, dan keperidian.
Cara demikian sering menimbulkan kesulitan dalam pengambilan keputusan jika
parameter tersebut saling bertentangan. Penggunaan statistik demografi mampu
mengatasi kesulitan tersebut. Statistik demografi serangga oleh Zeng et al. (1983)
diartikan sebagai analisis secara kuantitatif populasi serangga hama dalam hubungannya
dengan kelangsungan hidup, keperidian, dan pola pertumbuhan populasi. Adapun data
mengenai daya kemampuan hidup dan keperidian disusun dalam bentuk neraca
kehidupan (life table). Parameter demografi yang dihitung (Birch 1948) meliputi :
1. Laju Reproduksi Bersih (Ro), dihitung dengan rumus:
Ro = ∑lxmx
2. Laju Reproduksi Kotor (GRR), dihitung dengan rumus :
GRR = ∑mx
3. Laju pertambahan Intrinsik (r) dihitung secara eterasi dihitung dengan rumus:
∑lxmx e- rx = 1
4. Rataan masa generasi (T) dihitung dengan rumus:
T = ∑xlxmx/∑lxmx
5. Populasi berlipat ganda dihitung dengan rumus :
DT = ln (2)/r
Keterangan :
x = Kelas umur kohor (hari).
lx = Proporsi individu yang hidup pada umur x.
mx = Keperidian spesifik individu-individu pada kelas umur x atau jumlah.
anak betina perkapita yang lahir pada kelas x.
Peubah biologi yang diamati meliputi: 1) lama waktu perkembangan yang di
butuhkan sejak telur di letakkan oleh imago hingga menetas menjadi nimfa instar satu;
3) lama waktu perkembangan yang dibutuhkan nimfa instar empat hingga menjadi pupa
; 4) lama waktu perkembangan yang dibutuhkan pupa hingga menjadi imago ; 5) lama
hidup imago sejak keluar dari pupa sampai mati ; 6) masa sebelum peletakkan telur
HASIL DAN PEMBAHASAN
Identifikasi
Hasil identifikasi dengan menggunakan preparat mikroskop pada kantung pupa
kutukebul berdasarkan kunci identifikasi Martin (1987), ditemukan ciri morfologi
B. tabaci yang spesifik berupa seta kauda yang kokoh, umumnya sama panjang dengan
vasiform orifice, dan tidak banyak variasi diantara individu. Vasiform orifice terdapat di
daerah sebelum ujung posterior, berbentuk segitiga, ukurannya lebih panjang dari alur
kauda (caudal furrow) dan bagian samping orifice hampir lurus. Lingula berbentuk
seperti lidah, serta subdorsum tampa pori. Kutikula kantung pupa berwarna pucat,
dengan bentuk bervariasi tergantung bentuk permukaan daun.
Kantung pupa kutukebul antara biotipe-B dan non-B memiliki beberapa
perbedaan morfologi. Ciri morfologi spesifik bagi biotipe-B adalah tidak adanya
submarginal seta (ASMS4) di bagian anterior adanya lapisan lilin di anterior kiri dan
kanan tubuh yang berukuran pendek, serta lapisan lilin yang sedikit di bagian posterior.
Ciri morfologi spesifik biotipe non-B adalah terdapat submarginal seta (ASMS4) di
bagian anterior. Pada bagian anterior kiri dan kanan terdapat lapisan lilin yang panjang,
sedangkan bagian posterior terdapat lapisan lilin yang lebih melebar.
Siklus Hidup B. tabaci Biotipe-B dan Non-B pada Tanaman Mentimun dan Cabai
Siklus hidup adalah selang waktu sejak telur diletakkan hingga saat imago betina
meletakkan telur untuk pertama kalinya. Hasil pengamatan harian menunjukkan, siklus
hidup B. tabaci biotipe-B dan non-B pada mentimun (Tabel 2), berturut-turut adalah
31,27 hari dan 29,60 hari. Pada tanaman cabai siklus hidup B. tabaci biotipe-B dan
non-B (Tabel 3) yaitu berturut-turut 33,27 pada hari dan 30,86 hari. Pada tanaman
mentimun imago betina B. tabaci biotipe-B mulai meletakkan telur pada hari ke-25,
dengan masa peneluran 14,86 hari dan jumlah telur yang diletakkan 130 butir per
betina. Sementara itu di tanaman cabai, telur mulai diletakkan pada hari ke-27, dengan
masa peneluran 15,58 hari, dan jumlah telur yang diletakkan 82,05 butir per betina.
Jumlah telur yang diletakkan imago betina B. tabaci biotipe non-B pada tanaman
mentimun dan cabai masing masing 130,30 butir per betina, dengan masa peneluran
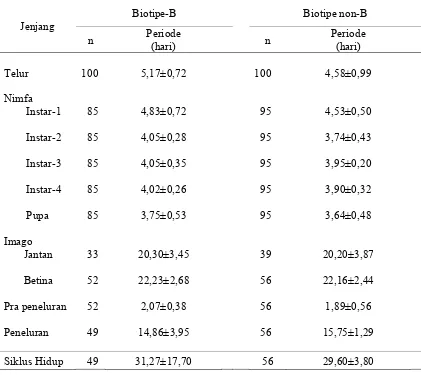
Tabel 2 Lama stadia B. tabaci biotipe-B dan non-B pada tanaman mentimun
Jenjang
Biotipe-B Biotipe non-B
n Periode
(hari) n
Periode (hari)
Telur 100 5,17±0,72 100 4,58±0,99
Nimfa
Instar-1 85 4,83±0,72 95 4,53±0,50
Instar-2 85 4,05±0,28 95 3,74±0,43
Instar-3 85 4,05±0,35 95 3,95±0,20
Instar-4 85 4,02±0,26 95 3,90±0,32
Pupa 85 3,75±0,53 95 3,64±0,48
Imago
Jantan 33 20,30±3,45 39 20,20±3,87
Betina 52 22,23±2,68 56 22,16±2,44
Pra peneluran 52 2,07±0,38 56 1,89±0,56
Peneluran 49 14,86±3,95 56 15,75±1,29
Siklus Hidup 49 31,27±17,70 56 29,60±3,80
Powell dan Bellows (1992) menyatakan bahwa siklus hidup B. tabaci pada
tanaman mentimun berkisar 17,4 hari dengan suhu 25oC, dan tingkat keberhasilan telur
menetas adalah 86,80%. Sementara itu Tsai dan Wang (1996) melaporkan siklus hidup
B. tabaci pada tanaman mentimun berkisar 19,3 hari pada suhu 25,5oC, dengan tingkat
keberhasilan telur menetas mencapai 95%.
Perbandingan sex rasio antara B. tabaci biotipe-B dan non-B pada tanaman cabai
dan tanaman mentimun adalah satu jantan banding dua betina, artinya ke dua biotipe
B. tabaci lebih banyak menghasilkan keturunan betina daripada jantan. Imago betina
hidup lebih lama daripada imago jantan yang dapat dilihat pada Tabel 2 dan Tabel 3.
Mortalitas pradewasa antara B. tabaci non-B dengan biotipe-B mempunyai kesamaan,
Mortalitas tertinggi B. tabaci non-B di tanaman cabai terjadi pada fase telur adalah
25%, dan nimfa instar satu sebesar 16%. Tingkat mortalitas pada B. tabaci biotipe-B
[image:31.612.104.524.176.552.2]ditanaman cabai terjadi pada fase telur sebesar 30%, dan nimfa instar satu 15,71%.
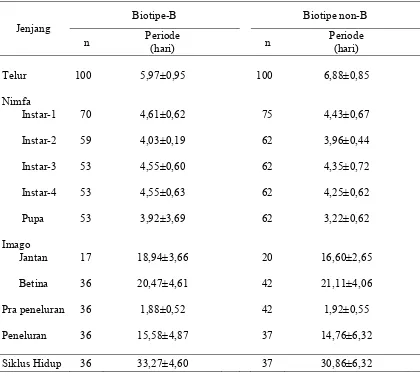
Tabel 3 Lama stadia B. tabaci biotipe-B dan non-B pada tanaman cabai
Jenjang
Biotipe-B Biotipe non-B
n Periode
(hari) n
Periode (hari)
Telur 100 5,97±0,95 100 6,88±0,85
Nimfa
Instar-1 70 4,61±0,62 75 4,43±0,67
Instar-2 59 4,03±0,19 62 3,96±0,44
Instar-3 53 4,55±0,60 62 4,35±0,72
Instar-4 53 4,55±0,63 62 4,25±0,62
Pupa 53 3,92±3,69 62 3,22±0,62
Imago
Jantan 17 18,94±3,66 20 16,60±2,65
Betina 36 20,47±4,61 42 21,11±4,06
Pra peneluran 36 1,88±0,52 42 1,92±0,55
Peneluran 36 15,58±4,87 37 14,76±6,32
Siklus Hidup 36 33,27±4,60 37 30,86±6,32
Hasil uraian di atas terlihat bahwa perbedaan tanaman berpengaruh nyata
terhadap jenjang perkembangan B. tabaci biotipe-B dan non-B. Nilai keperidian
B. tabaci biotipe-B dan non-B dapat dilihat pada Tabel Lampiran 7.
Lama Hidup dan Keperidian B. tabaci Biotipe-B dan Non-B
Setiap organisme mempunyai variasi jangka hidup yang terbatas, yang
menentukan karakteristik kelangsungan hidupnya di dalam suatu populasi. Lama hidup
sama. Kemampuan bertahan hidup B. tabaci biotipe-B dan non-B pada tanaman
mentimun yaitu berturut-turut adalah 53 hari dan 51 hari. Laju kematian biotipe-B dan
non-B terjadi pada fase telur yang ditunjukkan dengan penurunan jumlah
masing-masing pada hari ke-8 dan ke-9. Hal ini mengisyaratkan bahwa fase telur sangat
rentan terhadap gangguan luar, termasuk gangguan mekanis pada saat gugurnya daun.
Pada tanaman cabai kemampuan hidup B. tabaci biotipe-B dan non-B masing-masing
adalah 54 hari dan 53 hari. Laju kematian tertinggi pada kedua biotipe terjadi pada fase
telur, setelah itu nimfa instar I dan nimfa instar II. Nutrisi merupakan salah satu faktor
yang penting yang mempengaruhi kehidupan serangga. Pengaruhnya antara lain pada
proses pertumbuhan, perkembangbiakan, reproduksi, dan kebugaran imago. Faktor yang
lainnya adalah faktor fisik dan kimia tanaman yang umumnya tidak berkerja secara
tunggal tetapi bersama-sama membentuk suatu sistem pertahanan terhadap serangga
(Wiseman 1985).
Tipe bertahan hidup B. tabaci biotipe-B dan non-B memperlihatkan pola yang
dikenal dengan tipe II. Menurut Price (1978), ada tiga jenis kurva bertahan hidup,
yaitu: tipe I, tipe II, dan tipe III. Kurva tipe I adalah kematian organisme dalam jumlah
yang sedikit ketika umur muda dan kematian dalam jumlah besar sewaktu organisme
lebih tua, tipe II menunjukkan laju kematian yang konstan, sedangkan tipe III
memperlihatkan kematian yang besar yang terjadi di waktu muda.
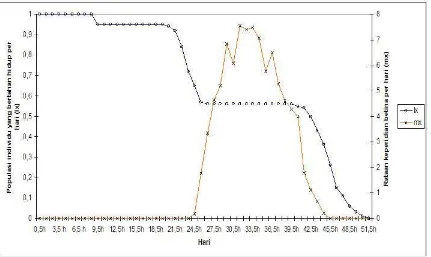
Banyaknya telur yang diletakkan setiap harinya disajikan dalam bentuk mx
(laju reproduksi kotor). Adapun nilai mx menunjukkan banyaknya telur betina yang
dihasilkan induk yang berumur hari ke-x, dan dihitung setelah mempertimbangkan
nisbah kelamin. Nilai mx B. tabaci biotipe-B dan non-B pada tanaman mentimun dapat
dilihat pada Tabel 4, sedangkan pada tanaman cabai disajikan pada Tabel 5. Puncak
kurva nilai mx pada B. tabaci non-B untuk tanaman cabai dicapai pada hari ke-43
dengan awal meletakkan telur pada hari ke-27 dengan nilai mx adalah 38,67 individu per
betina, sedangkan untuk tanaman mentimun dicapai pada hari ke-33 dengan awal
peletakan telur hari ke-25 dan nilai mx adalah 91,71 individu per betina. Sementara itu
puncak kurva dan nilai mx pada B. tabaci biotipe-B untuk tanaman cabai dan mentimun,
berturut-turut hari ke-35 dengan nilai mx adalah 75,79 individu dan hari ke-31 dengan
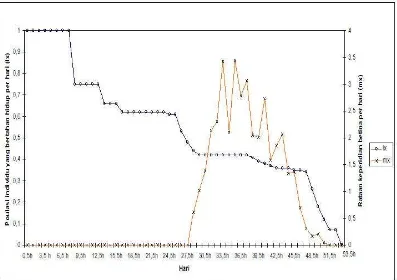
nilai mx adalah 49,36 individu. Kurva lama hidup dan keperidian B. tabaci pada
pada Gambar 7. Kurva bertahan hidup dan keperidian B. tabaci pada tanaman cabai
[image:33.612.107.513.177.374.2]biotipe-B dapat dilihat pada Gambar 8, dan untuk B. tabaci non-B pada Gambar 9.
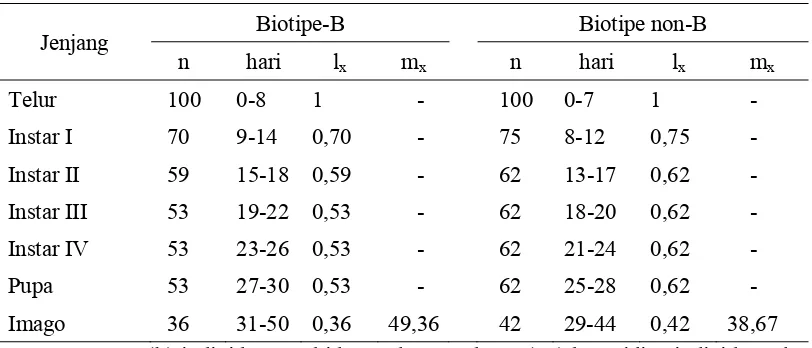
Tabel 4 Lama hidup dan keperidian B. tabaci biotipe-B dan non-B pada tanaman mentimun
Jenjang Biotipe-B Biotipe non-B
n hari lx mx n hari lx mx
Telur 100 0-7 1 - 100 0-8 1 -
Instar I 85 8-12 0,85 - 95 9-13 0,95 -
Instar II 85 13-16 0,85 - 95 14-17 0,95 -
Instar III 85 17-20 0,85 - 95 18-20 0,95 -
Instar IV 85 21-24 0,85 - 95 21-23 0,95 -
Pupa 85 25-28 0,85 - 95 24-26 0,95 -
Imago 52 29-48 0,52 75,79 56 27-44 0,56 91,71
Keterangan : (lx) individu yang hidup pada umur ke-x, (mx) keperidian individu
pada umur ke-x.
Tabel 5 Lama hidup dan keperidian B. tabaci biotipe-B dan non-B pada tanaman cabai
Jenjang Biotipe-B Biotipe non-B
n hari lx mx n hari lx mx
Telur 100 0-8 1 - 100 0-7 1 -
Instar I 70 9-14 0,70 - 75 8-12 0,75 -
Instar II 59 15-18 0,59 - 62 13-17 0,62 -
Instar III 53 19-22 0,53 - 62 18-20 0,62 -
Instar IV 53 23-26 0,53 - 62 21-24 0,62 -
Pupa 53 27-30 0,53 - 62 25-28 0,62 -
Imago 36 31-50 0,36 49,36 42 29-44 0,42 38,67
Keterangan : (lx) individu yang hidup pada umur ke-x, (mx) keperidian individu pada
[image:33.612.107.513.438.612.2]Gambar 6 Kurva lama hidup (lx) dan rataan keperidian betina per hari (mx) B. tabaci
biotipe-B pada tanaman mentimun.
Gambar 7 Kurva lama hidup (lx) dan rataan keperidian betina per hari (mx) B. tabaci
[image:34.612.110.541.367.624.2]Gambar 8 Kurva lama hidup (lx) dan rataan keperidian betina per hari (mx) B. tabaci
biotipe-B pada tanaman cabai.
Gambar 9 Kurva lama hidup (lx) dan retaan keperidian betina per hari (mx) B. tabaci
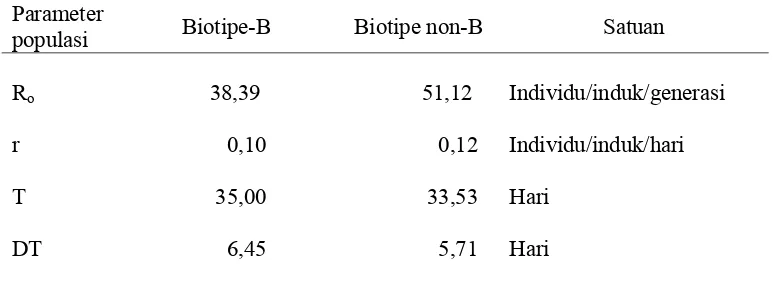
[image:35.612.109.506.389.669.2]Neraca Kehidupan B. tabaci Biotipe-B dan Non-B pada Tanaman Mentimun
Pada tanaman mentimun (Tabel 6), nilai Ro untuk B. tabaci non-B per individu
per generasinya adalah 51,12 per individu, sedangkan biotipe-B adalah 38,39
per individu. Laju pertambahan intrinsik (r) merupakan pertambahan populasi pada
lingkungan konstan dan sumberdaya yang tidak terbatas (Birch 1948). Keturunan yang
dihasilkan B. tabaci non-B dan biotipe-B berturut-turut adalah 0,12 dan 0,10 betina per
induk per hari. Tingginya nilai r disebabkan oleh tingginya keperidian, rendahnya
mortalitas pradewasa, dan masa dewasa. Birch (1948) menyatakan bahwa semangkin
tinggi persentase telur diletakkan pada kelompok umur muda, maka akan semangkin
besar nilai laju pertambahan intrinsik organisme tersebut.
Rataan masa generasi (T) adalah rataan waktu yang dibutuhkan sejak telur
diletakkan hingga saat imago betina menghasilkan separuh keturunannya. Berdasarkan
dua jenis tanaman (mentimun dan cabai) yang diujikan, terlihat bahwa waktu yang
dibutuhkan B. tabaci non-B untuk berkembangbiak pada tanaman mentimun lebih
singkat 1,2 kali daripada biotipe-B. Menurut Oka (1998), arti penting nilai Ro dan T
adalah untuk membandingkan nilai r. Misalkan dua populasi spesies yang berbeda
dengan nilai Ro yang sama tinggi, tetapi salah satu spesies nilai T-nya pendek. Maka
laju pertambahan intrinsik kedua populasi tersebut dalam satuan waktu tertentu akan
berbeda. Waktu yang dibutuhkan populasi untuk berlipat ganda (DT) adalah 6,45 hari
pada biotipe-B dan 5,71 hari pada non-B.
Neraca Kehidupan B. tabaci Biotipe-B dan Non-B Pada Tanaman Cabai
Pada tanaman cabai (Tabel 7) diperoleh nilai laju reproduksi kotor (Ro) B. tabaci
biotipe-B dan non-B berturut-turut adalah 15,30 dan 15,41 per individu. Hasil tersebut
diartikan. Nilai r yang diperoleh pada B. tabaci biotipe-B dan non-B masing-masing
0,17 dan 0,07 per induk per hari. Rataan masa generasi (T) yang diperoleh merupakan
rataan waktu yang dibutuhkan sejak telur diletakkan hingga saat imago menghasilkan
separuh keturunan masing- masing 37,69 hari pada biotipe-B dan 37,14 hari pada
non-B. Waktu yang dibutuhkan untuk populasi berlipat ganda (DT) adalah 3,96 hari pada
Tabel 6. Neraca kehidupan B. tabaci biotipe-B dan non-B pada tanaman mentimun
Parameter
populasi Biotipe-B Biotipe non-B Satuan
Ro 38,39 51,12 Individu/induk/generasi
r 0,10 0,12 Individu/induk/hari
T 35,00 33,53 Hari
DT 6,45 5,71 Hari
Keterangan : (Ro) Laju reproduksi bersih, (r) laju pertambahan intrinsik , (T) rataan
masa generasi, (DT) waktu untuk populasi berlipat ganda
Tabel 7. Neraca kehidupan B. tabaci biotipe-B dan non-B pada tanaman cabai
Parameter
populasi Biotipe-B Biotipe non-B Satuan
Ro 15,30 15,41 Individu/induk/generasi
r 0,17 0,07 Individu/induk/hari
T 37,69 37,14 Hari
DT 3,96 9,21 Hari
Keterangan : (Ro) Laju reproduksi bersih, (r) laju pertambahan intrinsik , (T) rataan
[image:37.612.102.486.306.458.2]PEMBAHASAN UMUM
Hasil identifikasi menggunakan preparat mikroskop kantung pupa B. tabaci
berdasarkan kunci identifikasi Martin (1987) dan Martin (2000), diperoleh adanya
perbedaan secara morfologi. Perbedaan tersebut terlihat pada adanya submarginal seta
(ASMP4) pada B. tabaci biotipe-B, sedangkan pada non-B tidak ditemukan. Kemudian
perbedaan yang lainnya adalah lapisan lilin pada bagian posterior B. tabaci biotipe-B
lebih sedikit dan lebih pendek pada bagian anterior tubuhnya, berbeda dengan
biotipe-non B lapisan lilin bagian posterior tubuh lebih banyak dan lebih melebar pada
bagian anteriornya. Calvert et al. (2001), menemukan adanya perbedaan morfologi
lainnya yaitu pada bagian mata dan antena. Adapun metode identifikasi lainnya adalah
dengan menggunakan tehnik polymesterase chain reaction (PCR). Aidawati (2006)
melaporkan hasil identifikasi, dengan menggunakan metode tehnik PCR-RAPD
menemukannya bahwa B. tabaci biotipe-B diperoleh pada tanaman brokoli yang berasal
dari daerah Bogor, sedangkan B. tabaci non-B diperoleh dari tanaman cabai, terung,
mentimun, kedelai, dan edamame. Pengujian lainnya adalah menggunakan tanaman
indikator (tanaman squash), gejala yang timbul pada tanaman ini adalah dengan
perubahan warna daun menjadi keperak-perakan.
Siklus hidup B. tabaci biotipe-B dan non-B baik ditanaman mentimun maupun
di tanaman cabai tidak berbeda. Kesimpulan tersebut bedasarkan kisaran waktu yang
dibutuhkan dari telur hingga dewasa pada tanaman mentimun antara biotipe-B dan
non-B , masing-masing 31,27±17,70 hari dan 29,60±3,80 hari. Pada tanaman mentimun
kisaran waktu yang dibutuhkan dari telur hingga dewasapada B. tabaci biotipe-B dan
non-B, berturut-turut 33,27±4,60 hari dan 30,86±6,32 hari. Hasil tersebut berbeda
dengan yang dilaporkan oleh Powell dan Bellows (1996) yang menyatakan bahwa,
kisaran waktu yang dibutuhkan dari telur hingga dewasa oleh B. tabaci pada tanaman
mentimun masing-masing 38,2 hari dengan suhu 20oC; 20,2 hari dengan suhu 25oC, dan
17,4 hari dengan suhu 32oC. Setelah terjadinya kopulasi antara jantan dan betina,
selanjutnya imago betina meletakkan telur di permukaan daun. Jumlah telur yang
diletakkan oleh imago betina B. tabaci biotipe-B dan non-B pada tanaman mentimun
masing-masing sebanyak 130 telur dan 130,30 telur. Sedangkan pada tanaman cabai
butir untuk non-B. Adapun perbedaan jumlah telur yang dihasilkan oleh betina B. tabaci
diduga karena adanya faktor fisik dan kimi tanaman. Menurut Harris (2001),
kemampuan imago betina B. tabaci dalam menghasilkan telur dan siklus hidupnya
dipengaruhi oleh jenis tanaman inang dan kondisi suhu.
Perbandingan sex rasio antara B. tabaci biotipe-B dan non-B dengan perlakuan
mentimun dan cabai adalah 1:2 (satu jantan banding dua betina). Menurut Samudra dan
Naito (1991) B. tabaci bersifat arrhenotokous artinya lebih banyak menghasilkan betina
daripada jantan. Dengan demikian jumlah individu betina dapat mempengaruhi nilai
keperidian, yang terlihat pada B. tabaci biotipe non-B dengan perlakuan mentimun
(Tabel 2). Jumlah imago betina yang lebih banyak daripada imago jantan dapat
menimbulkan masalah jika serangga tersebut menimbulkan kerugian. Hal ini
disebabkan perkembangan populasi di suatu habitat ditentukan oleh banyaknya imago
betina, artinya semangkin banyak imago betina maka populasi akan semangkin
meningkat.
Siklus hidup B. tabaci non-B pada tanaman mentimun lebih pendek 1,04 kali
daripada tanaman cabai, dengan keperidian lebih banyak 1,37 kali. Sementara itu siklus
hidup pada B. tabaci biotipe-B pada tanaman mentimun lebih cepat 1,06 kali daripada
tanaman cabai, dengan keperidian lebih banyak 1,58 kali pada tanaman mentimun
daripada tanaman cabai. Keadaan ini disebabkan oleh kebutuhan makan, perilaku seekor
serangga, dan jenis dimakannya, yang selanjutnya akan menentukan keperidian dan
strategi reroduksinya (Caroline et al. 2002). Menurut Naranjo dan Ellsworth (2005),
tanaman inang merupakan salah satu faktor biotik yang dapat mempengaruhi aspek
biologi dan kelangsungan hidup suatu organisme.
Isaacs et al. (1998) menyatakan bahwa pemberian variasi terhadap kualitas
tanaman melon dapat memberikan pengaruh terhadap mekanisme fisiologi dan perilaku
B. tabaci. Mekanisme fisiologi diperlihatkan dengan penurunan berat tubuh yang
sejalan dengan penurunan konsentrasi karbohidrat. Blackmer et al. (2002) melaporkan
adanya pengaruh terhadap lama hari dan tingginya intensitas penyinaran terhadap
peletakkan telur dan daya bertahan hidup. Menurut Kogan (1982), faktor fisik dan kimia
tanaman memegang peranan penting dalam pemilihan dan penentuan inang tersebut.
Faktor ini umumnya tidak berkerja secara tunggal, tetapi bersama-sama membentuk
Waktu yang dibutuhkan untuk populasi berlipat ganda (DT) pada B. tabaci
biotipe-B adalah 3,96 hari, sedangkan yang terendah pada B. tabaci non-B dengan
perlakuan tanaman cabai. Walaupun nilai laju reproduksi bersih B. tabaci biotipe-B
dengan perlakuan cabai rendah yaitu 15,30 per generasi, tetapi dengan nilai DT yang
tinggi dapat menyebabkan meningkatnya laju reproduksi kotor (GRR) dan nilai laju
reproduksi bersih (Ro) dalam satuan waktu tertentu. Menurut Birch (1948), nilai
berlipat populasi ganda yang tinggi pada suatu individu dapat menyebabkan penurunan
sumber-sumber daya lingkungan dan mempengaruhi pada nilai laju pertambahan
intrinsik (r).
Nilai rataan masa generasi (T) diartikan sebagai rataan waktu yang dibutuhkan
sejak telur diletakkan hingga saat imago menghasilkan keturunan. Pada B. tabaci non-B
dengan perlakuan tanaman mentimun diperoleh nilai laju reproduksi bersihnya (Ro) dan
nilai laju reproduksi kotornya (GRR) masing-masing 51,12 per induk per generasi dan
91,71 per betina, dengan nilai rataan masa generasinya 33 hari. Nilai tersebut diartikan
sebagai bentuk respon dari nilai rataan masa generasi yang pendek, yang menyebabkan
imago betina meningkatkan nilai Ro dan GRR. Laba (2005) menyatakan bahwa nilai T
yang rendah dapat meyebabkan meningkatnya nilai Ro pada kepik renda lada
(Dicoronis hewetti) pada varietas chunuk. Nilai Ro dan GRR yang tinggi
memperlihatkan tingkat kesesuian hidup pada suatu tanaman.
Pada perlakuan tanaman cabai antara B. tabaci biotipe-B dan non-B terlihat nilai
Ro dan T yang tidak berbeda (Tabel 7). Menurut Oka (1998), jika kedua populasi dari
spesies yang berbeda mempunyai nilai Ro yang sama tetapi salah satu spesies
mempunyai nilai T yang rendah sedangkan yang kedua mempunyai nilai T yang tinggi,
maka nilai laju pertambahan intrinsik dalam satuan waktu tertentu akan berbeda.
Spesies populasi yang mempunyai nilai T yang rendah akan tumbuh lebih cepat di
bandingkan spesies yang mempunyai nilai T yang tinggi.
Laju pertambahan intrinsik merupakan kapasitas suatu populasi untuk
peningkatan, nilai yang diperoleh ditentukan oleh berbagai aspek yang berhubungan
dengan sejarah kehidupan organisme, yaitu kematian, kelahiran, dan waktu
perkembangan. Nilai laju pertambahan instrinsik (r) yang tertinggi terlihat pada
B. tabaci biotipe-B dengan perlakuan tanaman cabai sebesar 0,17 per induk per hari.
dewasa dengan reproduksi yang awal. Hal ini terlihat dari B. tabaci non-B dengan
perlakuan mentimun (Tabel 4). Menurut Birch (1948), jika nilai r lebih kecil dari nilai r
yang minimum maka spesies tersebut gagal dalam bertahan hidup. Tetapi nilai laju
pertambahan intrinsik (r) yang tinggi pada suatu spesies tidak selalu diartikan sebagai
tingkat keberhasilan dalam suatu habitat. Hal tersebut berdasarkan adanya proses seleksi
dari spesies tersebut agar nilai r-nya menjadi relatif tinggi untuk mampu berkompetisi
dengan spesies yang lain. Hasil laporan Ginting (1996), menunjukkan bahwa perbedaan
nilai r antara kohor bunga dan kohor polong pada tanaman kacang panjang disebabkan
adanya berbedaan nilai mx, sedangkan pada nilai lx dan T tidak banyak berbeda. Brewer
(1979) membagi dalam empat faktor yang menyebabkan tinggi dan rendahnya nilai r,
yaitu jumlah keturunan per periode perkembangan, jumlah yang bertahan hidup dan
selama masa reproduktif, usia saat reproduktif dimulai, dan lama usia reproduktif. Dari
nilai neraca kehidupan yang diperoleh baik B. tabaci biotipe-B maupun non-B dengan
perlakuan tanaman mentimun dan cabai memperlihatkan adanya hubungan pengaruh
tanaman inang dengan strategi untuk bertahan hidup pada suatu habitat, oleh karena itu
dapat disimpulkan bahwa tanaman inang merupakan salah satu komponen faktor yang
KESIMPULAN
B. tabaci biotipe-B dan non-B memiliki waktu generasi lebih cepat pada
tanaman mentimun daripada tanaman cabai. Begitu juga kedua biotipe memiliki nilai
laju reproduksi yang lebih besar pada tanaman mentimun dibandingkan dengan tanaman
cabai.
B. tabaci biotipe-B pada tanaman cabai mempunyai laju pertumbuhan intrinsik
tertinggi yaitu 0,17 individu per induk per hari dan waktu untuk populasi berlipat ganda
tercepat yaitu 3,96 hari. B. tabaci non-B pada tanaman cabai mempunyai laju
pertumbuhan intrinsik paling rendah yaitu 0,07 individu per induk per hari dan memiliki
waktu untuk populasi berlipat ganda 9,12 hari.
SARAN
Untuk mengetahui neraca kehidupan B. tabaci pada berbagai keadaan,
disarankan untuk melakukan penelitian neraca kehidupan B. tabaci pada berbagai inang
DAFTAR PUSTAKA
Aidawati N 2006. Pengkajian Keanekaragaman Geminivirus Pada Tomat Dan Serangga Vektornya, Bemisia tabaci Gennadius (Hemiptera:Aleyrodidae), Serta Pengujian Ketahanan Genotipe Tomat Terhadap Strain Geminivirus. [Disertasi]. Program Pascasarjana, Institut Pertanian Bogor.
Belloti AC, Arias B. 2000. Host plant resistence to whitefly white emphasis on cassava as a case study. Cassava entomology. hlm : 1-23
Birch LC. 1948. The intristic rate of natural increase of an insect population. Journal Animal Ecology. hlm: 15-26
Blackmer JL, Lee LL, Henneberry J. 2002. Factor affecting egg hatch, development, and survival of B.argentifolii (Homoptera:Aleyrodidae) reared on artificial feeding. Journal Environment Entomology. 31(2): 306-312
Bosco D, Mason G, Accotto GP. 2001. Investigation on transovarial transmission of TYLCV-sar by Bemisia tabaci (Hemiptera: Aleyrodidae) European Whitefly Symp
Ragusa (Sicilia, Italy), 27 th Feb-3 rd March 2001
Burban C, Fishpool LDC, Fauquet C, Fargette D, Thouvenel JC. 1992. Host Assocoated biotypes within West African Population of the Whitefly Bemisia tabaci (Genn) (Homoptera: Aleyrodidae). J Appl. Entomol. 113: 416-432
Brewer R. 1979. Principles of ecology. W.B Sounders Co. Phidelphia. hlm : 299
Brown JK. 1990. An update on thw whitefly-transmitted geminiviruses in Americas and Carabian Basin. FAO Plant Prot. Bull. hlm : 33-58
[CABI] Center of Agricultural and Biologia Institute 2005. Crop Protection Comedium 2005 edition. London : CABI
Calvert LA, Cuervo M, Arroyave JA, Constantino AL, Belloti A, Frohclich D. 2001. Morphological and mitocondrial DNA marker analyses of whitefly (Homoptera: Aleyrodidae) colonizing cassava and beans in colombia. Journal of Annual Entomology society. 94(4):512-519
Carrey JR. 1993. Applied demograhy for biologist with special emphasis on insect. Oxford University Press. New York. hlm: 11-41
Caroline S A, Simon RL. 2002. Host plant quality and fecundity in herbivorous insect. [Review]. Annual review entomol. 47:817-835
Costa HS, Brown JK. 1990. Valibility in biology characteristics, is isozyme patterns and virus transmission among population of Bemisia tabaci in Arizona.
Journal Phytopathology. 61: 211-219
Czosnek H, Fridma V, Levy A, Morin S, Rubinstein G, Sobol I, Zeidan M. 2001. Interaction of whiteflies with geminiviruses from the old world. European Whitefly Symp. Ragusa (Sicilia, Italy), 27 th Feb-3rd March 2001.
De Barro PJ, Liebregts W, Carver M. 1998. Distribution and identiy of biotype of
Bemisia tabaci (Gennadius) using ribosomal ITS1. Mol. Phylogeny . Evol. 37:214-218
Drost YC, van Lenteren JC, van Roermund HJW. 1998. Life-history parameters of different biotypes of Bemisia tabaci (Hemiptera:Aleyrodidae) in relaction to temperature and host plant: a selective review. [Review]. Bulliten of entomological research. 88:219-229
Ginting BB. 1996. Biologi dan statistik demografi Maruca testulalis Geyer (Lepidoptera: Pyralidae) pada bunga dan polong kacang panjang. [Tesis]. Program Pascasarjana, Institut Pertanian Bogor.
Harris KF. Smith OP, Duffus JE. 2001. Virus-Insect-Plant-Interaction. Academic Press. hlm: 250-254
Hasibuan KM. 1988. Permodelan Matematika di dalam biologi populasi: Dinamika populasi. Pusat antar Universitas IPB. Bogor. hlm : 170
Hidayat SH, Rusli ES, Nooraidawati. 1999. Penggunaan primer universal dalam Polymerase chain reaction untuk mendeteksi virusgemini pada cabe. Kongres Nasional XV dan Seminar Ilmiah PFI, Purwakerto, 16-18 September 1999
Hunter WB, Heibert E, Webb SE, Tsai JH, Polston JE. 1998. Location of Geminivirus
in the whitefly Bemisia tabaci (Homoptera;Aleyrididae). Plant Disiase. 82:1147-1151
Horowitz AR. 1986. Population dynamics of Bemisia tabaci (Gennadius): With special emphasis on cotton field. Journal of Agricultural Ecosistem and Environment. 17: 37-47
Horowitz AR, Weintrayb PG, Ishaaya I. 1998. Status of pesticide resistence in arthropod pest in Israel. [mini review]. Dalam artikel Phytoparasitica:// www. phytoparasitica.org. (13 Des 2005)
Kalshoven LGE. 1981. The Pest of Crop in Indonesia. PA van der Laan. Penerjemah. Jakarta: Ichtiar Baru-van Houve. Terjemahan dari : De Plagen van de Cultuurgewassen in Indonesia
Kogan. M. 1982. Plant resistence in pest management. Introduction to insect management. New York. John wiley and Son
Laba IW. 2005, Kepik renda lada, Diconocoris hewetti Dist (Hemiptera: Tingiatae). Biologi, kelimpahan populasi, dan pengaruhnya terhadap kehilangan hasil. [Disertasi]. Program Pascasarjana, Institut Pertanian Bogor.
Legg JP. French R, Rogan D, Akao-Akuja, Browns K. 2002. A distinct Bemisia tabaci
(Gennadius) (Hemiptera:Sternorrhycha:Aleyrodidae) genotype cluster is associated with epidemic of severe cassava mosaic virus disease in Uganda.
Molecular ecology. 11:1219-1229
LimaLHC, Návia D, Inglis PW , de Oliveira MRV. 2000. Survey of Bemisia tabaci
(Gennadius) (Hemiptera: Aleyrodidae) biotypes in Brazil using RAPD markers.
Journal of Genetics and Molecular Biology. 23:781-785
Martin JH. 1987. An identification guide to common whitefly pest species of the word (Homoptera: Aleyrodidae). Tropical Pest Management. 33:298-322
Martin JH, Mifsud D, Rapisarda C. 2000. The whiteflies (Hemiptera: Aleyrodidae) of
eourope and mediterranean basin. Buletin of Entomological Reseach. 86:407-448
McCollum TG, Stoffella PJ, Powell CA, Cantliffe DJ, Hanif-khan S. 2000. Effect of silverleaf whitefly feeding on tomato fruit ripening. Postharvest Biology and Technology. 31: 183-190
Naranjo SE, Ellsworty PC. 2005. Mortality dinamics and population regulation in
Bemisia tabaci. Entomologia exsperimentalalis et aplicata. hlm: (116) 93-108. [NaranjoEllswortEEA.2005pdf].Dalamartikel http//.www.
ars.usda.gov/SP2UserFiles/person/4056/NaranjoEllsworthEEA .2005pdf [20.April. 2006]
Oka IN. 1998. Pengendalian hama terpadu dan implementasinya di Indonesia. Yogyakarta. Gagjah Mada University Press.
Oleivera MRV, Henneberry TJ, Anderson P. 2001. History, current status, and collaborative projects for Bemisia tabaci. Species issue: Challenges and opportunities for pest management of Bemisia tabaci in the new century.
Journal of Crop Protection. 20(9):709-723
Perring MT. 2001. The Bemisia tabaci species complex. Special issue: Challenges and opportunities foe pest management of Bemisia tabaci in the new century.
Perring TM, Farrar CA, Bellows TS, Cooper AD, Rodriguez RJ. 1993. Evidence for a new species of whitefly: UCR findings and aplications. California. 48:7-8
Price PW. 1997. Insect ecology (3rd edn). John Wiley. New York. hlm : 474-511
Powell DA, Bellows TS, Jr. 1992. Preimaginal development and survival of
Bemisia tabaci on cotton and cucumber. Journal Environment Entomology. 21:359-363
Sulandari S. 2004. Karakteristik, biologi, serologi, dan analisis sidik jari DNA Virus Penyebab Penyakit Daun Keriting Kuning Cabai. [Disertasi]. Program Pascasarjana, Institut Pertanian Bogor.
Samudra IM, Naito A. 1991. Variental resistence of soybean to whitefly Bemisia tabaci Genn. Dalam. Proceeding of final seminar on the strenghening of pioneering research for palawija crop production (ATA-378). Central Researcj Institute for food crop. Bogor. Indonesia. hlm : 51-55
Tsai JH, Wang K. 1996. Development and reproduction of Bemisia argentifolii
(Homoptera:Aleyrodidae) on five host plant. Journal Environtment Entomology. 25: 810-816
Van den Ven WTG, Levesque CS, Perring TM, Walling LL. 2000. Local and systemic chages in squash gene exspession in response to silverleaf whitefly feeding. The plant cell. 12:1409-1423
Wilson EO, Bossert WH. 1971. Primer population biology. Sinauer Associstes Inc. Sunderlan. hlm : 111-114
Wiseman BR. 1984. Type and mechanisms of host plant resistance to insect attack. Insect science and its application. 6(3): 239-248
Tabel lampiran 1 Siklus hidup B. tabaci biotipe-B pada tanaman cabai
Individu Hari
telur Instar I Instar II Instar III Instar IV Pupa Imago
1 5 4 4 6 5 4 4
2 5 5 4 6 5 4 5
3 5 5 4 4 5 5 5
4 5 5 4 4 5 5 4
5 5 5 4 4 5 4 6
6 5 5 4 5 5 3 5
7 5 4 4 4 5 4 7
8 5 4 4 4 3 4 6
9 5 4 4 4 3 4 6
10 5 5 4 4 6 4 7
11 5 4 4 4 4 4 5
12 6 4 4 5 5 4 6
13 6 4 4 5 4 4 7
14 6 4 5 4 5 4 7
15 7 5 4 4 5 3 5
16 7 6 4 5 4 3 6
17 7 5 4 5 4 3 6
18 7 5 4 5 4 4 6
19 5 5 4 5 5 5 6
20 5 5 4 5 5 5 6
21 7 4 4 4 5 3 6
22 7 4 4 4 5 5 5
23 7 6 4 4 4 4 5
24 7 6 4 4 4 4 5
25 6 4 4 4 4 4 7
26 5 5 4 5 5 5 6
27 5 5 4 5 5 4 5
Lanjutan………..
Individu Hari Telur Instar I Instar II Instar III Instar IV Pupa Imago
29 7 5 4 5 5 3 5
30 5 5 4 5 5 4 5
31 5 5 4 4 4 4 7
32 5 4 4 5 4 3 4
33 7 5 4 4 4 3 7
34 7 5 4 4 4 3 7
35 5 5 4 5 4 5 6
36 5 5 4 4 5 5 5
Total 208 170 145 163 164 142 206
Tabel Lampiran 2 Siklus hidup B. tabaci non-B pada tanaman mentimun
Individu Hari telur instar I Instar II Instar III Instar IV Pupa Imago
1 4 5 4 4 4 3 4
2 4 5 4 4 4 3 4
3 4 5 5 4 4 4 5
4 4 6 4 4 3 3 5
5 4 5 4 4 4 4 5
6 4 4 5 4 4 4 5
7 4 5 4 4 4 4 5
8 5 5 4 4 4 3 5
9 5 5 4 4 4 4 5
10 5 6 4 4 4 4 4
11 5 4 4 4 4 4 5
12 5 5 4 4 4 5 5
13 5 4 4 4 4 4 3
14 5 4 4 4 4 4 5
15 5 4 4 4 4 4 5
16 5 4 4 4 4 4 5
17 4 5 4 4 4 4 5
18 5 5 4 4 4 4 4
19 5 5 4 4 4 4 5
20 5 4 4 4 4 4 6
21 5 6 4 4 4 4 6
22 5 3 4 5 4 4 10
23 5 5 4 4 5 3 5
24 5 4 4 4 4 3 5
25 5 5 4 4 4 4 5
26 5 5 4 4 4 4 5
27 5 4 4 4 4 3 4
28 6 4 4 4 4 3 5
29 6 5 4 4 4 4 5
30 5 6 4 4 4 4 8
31 6 6 4 4 4 3 4
32 6 6 4 4 4 3 5
33 6 5 5 5 4 4 5
34 6 5 4 4 4 4 5
35 6 4 4 4 4 4 5
36 6 5 4 4 4 4 5
37 6 5 4 4 4 4 5
38 6 5 4 3 5 4 5
39 6 5 4 5 4 3 5
40 6 5 4 4 4 4 5
41 6 5 4 4 4 4 5
42 6 5 5 4 4 4 5
Lanjutan………..
Individu Hari Telur Instar I Instar II Instar III Instar IV Pupa Imago
44 6 5 4 4 4 4 6
45 6 6 4 4 4 3 6
46 6 6 4 4 4 3 6
47 6 5 4 5 4 5 6
48 6 5 4 6 4 4 5
Total 252 236 196 197 193 181 246
Tabel Lampiran 3 Siklus hidup B.tabaci non-B p