PEMANFAATAN TANAMAN PEMBATAS PINGGIR DAN
PREDATOR COCCINELLIDAE UNTUK PENGENDALIAN
KUTUKEBUL Bemisia tabaci (GENNADIUS) (HEMIPTERA:
ALEYRODIDAE), VEKTOR BEGOMOVIRUS PADA
PERTANAMAN CABAI MERAH (Capsicum annuum L.)
BAGUS KUKUH UDIARTO
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI DISERTASI
DAN SUMBER INFORMASI
Dengan ini saya menyatakan bahwa disertasi Pemanfaatan Tanaman Pembatas Pinggir dan Predator Coccinellidae untuk Pengendalian Kutukebul Bemisia tabaci (Gennadius) (Hemiptera: Aleyrodidae), Vektor Begomovirus pada Pertanaman Cabai Merah (Capsicum annuum L.) adalah karya saya dengan arahan komisi pembimbing dan belum diajukan dalam bentuk apapun kepada perguruan tinggi manapun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam daftar pustaka di bagian akhir disertasi ini.
Bogor, Juli 2012
ABSTRACT
BAGUS KUKUH UDIARTO, Use of Border Crops and Coccinellid Predators to Control The Whitefly Bemisia tabaci (Gennadius) (Hemiptera: Aleyrodidae), A Vector of Begomovirus on Chillipepper (Capsicum annuum L.) Under Supervision of PURNAMA HIDAYAT, AUNU RAUF, SRI HENDRASTUTI HIDAYAT and PUDJIANTO.
Whitefly, Bemisia tabaci (Hemiptera: Aleyrodidae) is an important pest on chillipepper due to its role as a vector of Begomovirus, the causal agent of yellow leaf curl disease. Many efforts has been undergone to suppress disease incidence, such as through controlling insect vector, B. tabaci. The objective of the research were to study the effect of border crops in chillipepper plantation on population of B. tabaci and incidence of yellow leaf curl disease as well as abundance and effectiveness of predator species. Field experiment was conducted using randomized completed design. Two seedling treatments (with and without cover), were combined with 4 border treatments i.e. maize, crotalaria (Fabaceae), chiffon fabric and non border. The potency of predator species was evaluated by conducting 3 consequtive assays, i.e. predation, prey preference and functional response assay. The results showed that the use of covered seedling was able to protect the seedling from B. tabaci and delay virus infection for 2 weeks. Population of B. tabaci was significantly lower in plot with combination of border crops and covered seedling. Correlation between population of B. tabaci and disease incidence was positif (r = 0.925), where as correlation between disease incidence and yield crop was negative (r = - 0.8886). Border crops especially maize could enhance the abundance of predator species with the most predators commonly found species were Menochilus sexmaculatus, Coccinella transversalis and Verania lineata. The predator species which have highest potency as natural enemy to control B. tabaci are V. lineata (Coleoptera: Coccinellidae).
RINGKASAN
BAGUS KUKUH UDIARTO. Pemanfaatan Tanaman Pembatas Pinggir dan Predator Coccinellidae untuk Pengendalian Kutukebul Bemisia tabaci (Gennadius) (Hemiptera: Aleyrodidae), Vektor Begomovirus pada Pertanaman Cabai Merah. Dibimbing oleh PURNAMA HIDAYAT, AUNU RAUF, SRI HENDRASTUTI HIDAYAT dan PUDJIANTO.
Kutukebul Bemisia tabaci (Gennadius) (Hemiptera: Aleyrodidae) merupakan salah satu hama penting pada pertanaman cabai merah karena merupakan satu-satunya vektor Begomovirus penyebab penyakit daun keriting kuning cabai. Penyakit daun keriting kuning merupakan penyakit penting pada pertanaman cabai terutama di Jawa Tengah. Kehilangan hasil akibat serangan penyakit tersebut berkisar 20 sampai 100%. Pengendalian serangga vektor merupakan strategi penting untuk menekan penyakit tersebut. Penelitian bertujuan: 1) Mengetahui pengaruh tanaman pembatas pinggir terhadap dinamika populasi B. tabaci dan insidensi penyakit daun keriting kuning pada pertanaman cabai merah; 2) Mengetahui pengaruh tanaman pembatas pinggir terhadap kelimpahan predator penting yang menyerang B. tabaci; 3) mengevaluasi efektivitas berbagai spesies predator (Coccinellidae) terhadap B. tabaci melalui uji daya pemangsaan, preferensi dan tanggap fungsional.
metode pilihan (choice), yakni dengan metode melingkar. Pengamatan dilakukan terhadap jumlah nimfa yang tersisa pada 3, 6, 12 dan 24 jam setelah pelepasan predator. Uji tanggap fungsional predator (Coccinelidae) terhadap B. tabaci. dilaksanakan dengan menggunakan Rancangan Acak Lengkap yang terdiri atas 10 perlakuan tingkat kepadatan inang, masing masing perlakuan diulang 10 kali. Kesepuluh perlakuan berbagai kepadatan nimfa B. tabaci yaitu; 1, 2, 3, 4, 6, 8, 12, 16, 22 28 nimfa. Pengamatan dilakukan terhadap jumlah nimfa yang tersisa setiap 3 jam setelah pelepasan predator selama 24 jam.
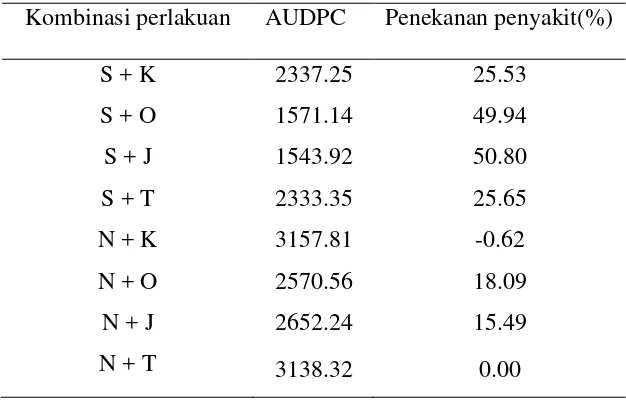
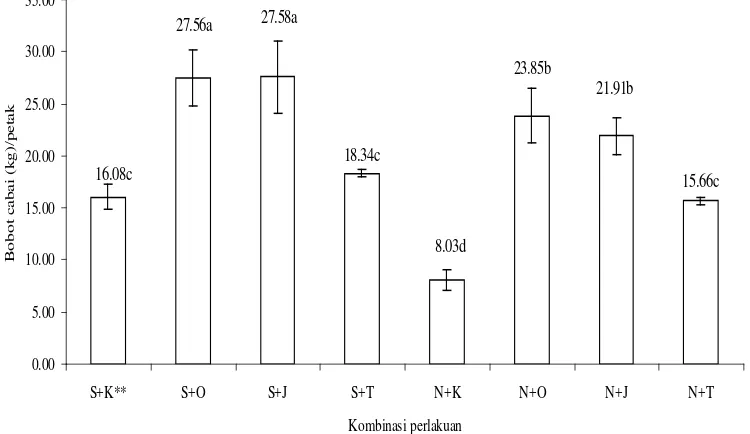
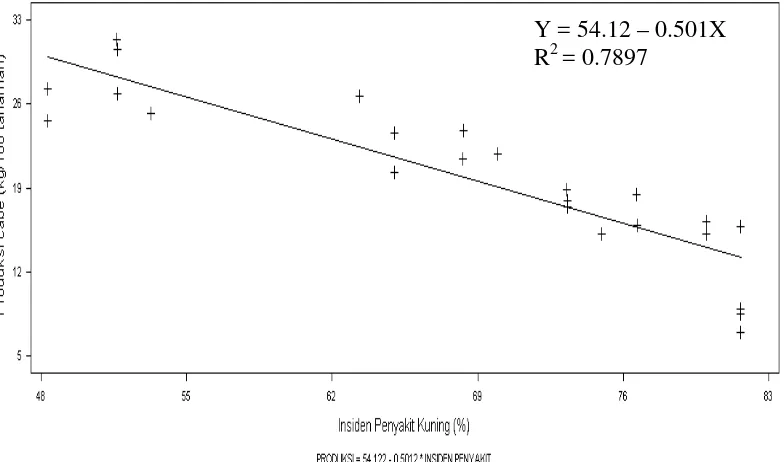
Hasil penelitian menunjukkan bahwa kehadiran B. tabaci pada pertanaman cabai tercatat sejak di pesemaian. Penggunaan sungkup dipesemaian dapat melindungi bibit cabai merah dari B. tabaci dan menunda infeksi virus selama 2 minggu. Tanaman pembatas pinggir berpengaruh nyata terhadap dinamika populasi B. tabaci dan insidensi penyakit daun keriting kuning pada pertanaman cabai merah. Kombinasi perlakuan penggunaan sungkup di pesemaian dan perlakuan pembatas pinggir tanaman jagung atau orok-orok di lahan pertanaman cabai merah dapat menekan populasi B. tabaci, insidensi penyakit daun keriting kuning cabai dan kehilangan hasil panen cabai merah akibat serangan penyakit tersebut. Insidensi penyakit keriting kuning tersebut dapat ditekan sebesar 50.80 % dan kehilangan hasil panen cabai merah dapat ditekan sebesar 80 %. Ada korelasi positif antara populasi B. tabaci dengan insidensi penyakit daun keriting kuning (r = 0.925), semakin tinggi tingkat populasi imago maka semakin tinggi tingkat insidensi penyakit daun keriting kuning cabai. Sebaliknya terdapat korelasi negatif antara insidensi penyakit daun keriting kuning dengan hasil panen cabai merah (r = - 0.8886), semakin tinggi tingkat insidensi penyakit daun keriting kuning cabai maka semakin rendah hasil panen cabai merah. Rerata populasi imago B. tabaci pada pertanaman cabai merah dengan perlakuan pembatas pinggir tanaman jagung, pembatas pinggir tanaman orok-orok, pembatas pinggir kain sifon dan tanpa pembatas pinggir berturut-turut 23.30 ekor, 24.88 ekor, 27.55 ekor dan 28.56 ekor. Rerata insidensi penyakit daun keriting kuning pada pertanaman cabai merah dengan perlakuan pembatas pinggir tanaman jagung, pembatas pinggir tanaman orok-orok, pembatas pinggir kain sifondan tanpa pembatas pinggir berturut-turut 50%, 51.56%, 75% dan 80.61%. Rerata hasil panen cabai merah dengan perlakuan pembatas pinggir tanaman jagung, pembatas pinggir tanaman orok-orok, pembatas pinggir kain sifon dan tanpa pembatas pinggir berturut-turut 27.58 kg/plot, 27.56 kg/plot, 16.08 kg/plot dan 15.66 kg/plot.
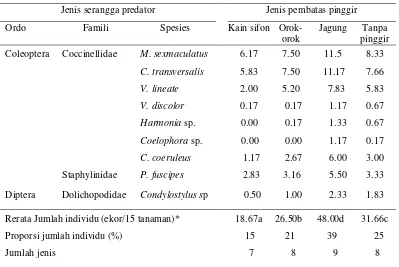
Dolichopodidae yaitu Condylostylus sp. Serangga predator tersebut yang dominan adalah M. sexmaculatus, C. transversalis dan V. lineata.
Predator M. sexmaculatus, C. transversalis dan V. lineata mempunyai daya pemangsaan terhadap B. tabaci yang sama tinggi yaitu berkisar 46 – 48 nimfa/hari atau 9 imago/hari. Predator M. sexmaculatus lebih menyukai A. gossypii dan M. persicae, predator C. transversalis lebih menyukai T. parvispinus, sedangkan V. lineata lebih menyukai B. tabaci. Hasil analisis regresi logistik (koefisien linier P1
= - 0.1006, X2 = 34.99) predator V. lineata memperlihatkan tanggap fungsional tipe II, artinya semakin meningkat kerapatan mangsa B. tabaci maka semakin meningkat mangsa yang dikonsumsi, namun proporsi mangsa yang dikonsumsi semakin menuurun. Pada kerapatan mangsa rendah (1 sampai dengan 3 nimfa) seluruh mangsa dikonsumsi (100%), artinya pada kerapatan mangsa yang rendah predator V. lineata masih mampu menemukan mangsa dan memangsanya. Hasil analisis dari persamaan cakram pada tanggap fungsional tipe II diperoleh nilai laju pencarian mangsa seketika (a) sebesar 0,3522/jam dan nilai masa penanganan mangsa (Th) sebesar 0.151jam (R2 = 0.9239). Laju pencarian mangsa seketika (a) menunjukkan proporsi dari total area yang dijelajahi predator per unit waktu jelajah. Semakin kecil nilai a, maka predator semakin agresif dan efektif dalam menemukan mangsa. Masa penanganan mangsa (Th) menunjukkan lamanya predator mengenali, mengejar, memakan, membersihkan alat mulut dan beristirahat sebelum bergerak mencari mangsa yang lain. Semakin rendah nilai Th, maka semakin tinggi daya pemangsaan maksimumnya. Predator V. lineata mempunyai daya pemangsaan maksimum 6 nimfa/jam.
Pembatas pinggir terbaik untuk pengendalian B. tabaci pada pertanaman cabai merah adalah tanaman jagung, karena mempunyai peranan ganda. Pembatas pinggir dengan tanaman jagung disamping dapat menekan populasi B. tabaci, insiden penyakit daun keriting kuning dan kehilangan hasil panen cabai, juga dapat meningkatkan kelimpahan predator penting yang menyerang B. tabaci. Serangga predator yang berpotensi untuk dikembangkan sebagai agens hayati untuk pengendalian B. tabaci adalah M. sexmaculatus, C. Transversalis dan V. lineata, namun dari ketiga predator tersebut yang paling efektif adalah V. lineata.
Disarankan varietas jagung yang digunakan sebagai tanaman pinggir sebaiknya yang mempunyai karakter tinggi tanaman lebih dari 2 m dan tajuk yang lebat serta umur yang relatif panjang. Penanaman jagung sebaiknya dilakukan 2 kali yaitu pada waktu 5 minggu sebelum dan 4 minggu setelah tanam cabai merah. Penanaman antar baris jagung sebaiknya dilakukan secara zigzag.
Penelitian lebih lanjut dapat berupa: 1) Kajian jarak dan proporsi luas tanaman pinggir dari pertanaman cabai merah serta analisis ekonominya; 2) Kajian peranan tepung sari dan mangsa alternatif pada tanaman jagung dalam mendukung konservasi predator.
© Hak cipta milik IPB, tahun 2012
Hak Cipta dilindungi undang-undang
Dilarang mengutip sebagaian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumber. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan yang wajar IPB
PEMANFAATAN TANAMAN PEMBATAS PINGGIR DAN
PREDATOR COCCINELLIDAE UNTUK PENGENDALIAN
KUTUKEBUL
Bemisia tabaci
(GENNADIUS) ( HEMIPTERA:
ALEYRODIDAE ), VEKTOR BEGOMOVIRUS PADA
PERTANAMAN CABAI MERAH (
Capsicum annuum
L.)
BAGUS KUKUH UDIARTO
Disertasi
Sebagai salah satu syarat untuk gelar Doktor
pada
Program Studi Entomologi
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Penguji pada Ujian Tertutup: Dr. Ir. I Wayan Winasa, M.Si.
Dr. Ir. Laksminiwati Prabaningrum, M.S.
Penguji pada Ujian Terbuka: Dr. Ir. Yusdar Hilman, M.Sc. Dr. Ir. Sugeng Santoso, M.Agr.
Judul Penelitian : Pemanfaatan Tanaman Pembatas Pinggir dan Predator Coccinellidae untuk Pengendalian Kutukebul Bemisia tabaci (Gennadius) (Hemiptera: Aleyrodidae), Vektor Begomovirus pada Pertanaman Cabai Merah (Capsicum annuum L.)
Nama : Bagus Kukuh Udiarto
NRP : A361070051
Program Studi : Entomologi
Disetujui Komisi Pembimbing
Dr. Ir. Purnama Hidayat, M.Sc. Prof. Dr. Ir. Aunu Rauf, M.Sc Ketua Anggota
.
Dr. Ir. Sri Hendrastuti H., M.Sc.
Anggota Anggota
Dr. Ir. Pudjianto, M.Si.
Diketahui
Ketua Program Studi Entomologi Dekan Sekolah Pascasarjana IPB
Dr. Ir. Pudjianto, M.Si. Dr. Ir. Dahrul Syah, M.Sc.Agr.
PRAKATA
Puji dan syukur penulis panjatkan ke hadirat Allah SWT, karena atas rahmat dan karunia-Nya, sehingga disertasi yang berjudul “Pemanfaatan Tanaman
Pembatas Pinggir dan Predator Coccinellidae Untuk Pengendalian Kutukebul Bemisia tabaci (Gennadius) (Hemiptera: Aleyrodidae), Vektor Begomovirus pada Pertanaman Cabai Merah (Capsicum annuum L.)” dapat penulis selesaikan.
Bagian dari disertasi ini telah diterbitkan di Jurnal Hortikultura Volume 22 No.1 Tahun 2012, dengan judul Kajian Potensi Predator Coccinellidae untuk Pengendalian B. tabaci (Gennadius) pada Cabai Merah.
Dengan selesainya penulisan disertasi ini, penulis menyampaikan rasa terimakasih dan penghargaan kepada komisi pembimbing yaitu Dr. Ir. Purnama Hidayat, M.Sc., Prof. Dr. Ir. Aunu Rauf, M.Sc., Dr. Ir. Sri Hendrastuti H., M.Sc.
dan Dr. Ir. Pudjianto, M.Si., atas bimbingan dan pengarahan kepada penulis mulai dari perencanaan penelitian hingga penyelesaian penulisan disertasi.
Ucapan terimakasih penulis sampaikan juga kepada Kepala Badan Penelitian dan Pengembangan Pertanian, Kepala Pusat Penelitian dan Pengembangan Hortikultura, Kepala Balai Penelitian Tanaman Sayuran, Dekan Sekolah Pascasarjana IPB yang telah memberikan kesempatan kepada penulis untuk melanjutkan pendidikan program doktor. Kepada Ketua Program Studi Entomologi dan seluruh dosen di Departemen Proteksi Tanaman IPB, penulis mengucapkan terimakasih atas segala arahan dan didikan.
Penulis ucapkan terimakasih kepada Badan Penelitian dan Pengembangan Pertanian melalui program Kerja Sama Kemitraan Penelitian Pertanian dengan Perguruan Tinggi (KKP3T) dan Kementerian Pendidikan dan Kebudayaan melalui program Indonesia Managing Higher Education for Relevance and Efficiency (I-MHERE B.2c.) yang telah membantu memberikan dana untuk pelaksanaan penelitian disertasi ini.
Terimakasih penulis ucapkan kepada Ir. Wiwin setiawati, M.S., Ir. Fardedi, M.Si., Rahmini, S.P., M.P. dan Rahmawati S.P., M.Si., atas dukungan dan kerjasama yang baik, serta kepada ibu Yuke Wulandari, S.P. dan Bapak Sagimen yang telah membantu pelaksanaan penelitian di lapangan.
Kepada orangtua, ayahanda Soehiro (alm) dan ibunda Yetty Soebaningsih, serta mertua M.E. Kurniadi beserta istri disampaikan terimakasih atas pendidikan, dukungan, nasihat dan doa sehingga penulis dapat menyelesaikan pendidikan ini. Kepada istri Asih Setiasih, S.H. dan ketiga putra tercinta Destylana Agusti Putri, Hadian Dwi Nugroho dan Rizqi Wisudanto serta seluruh keluarga yang selalu mendoakan dan memberikan dukungan dalam menyelesaikan pendidikan ini, penulis mengucapkan terimakasih.
Semoga karya ilmiah ini bermanfaat.
Bogor, Juli 2012
RIWAYAT HIDUP
Penulis dilahirkan di Surabaya pada tanggal 31 Januari 1962 dari ayah Soehiro (alm) dan ibu Yetty Subaningsih. Penulis merupakan putra keempat dari lima bersaudara.
Pada tahun 1982 penulis lulus dari SMA Negeri 10 Bandung, Jawa Barat dan pada tahun 1983 lulus seleksi masuk Universitas Padjadjaran Bandung, Jurusan Hama dan Penyakit Tumbuhan melalui jalur ujian masuk perguruan tinggi negeri (UMPTN). Tahun 1988 penulis mendapat gelar Sarjana Pertanian. Penulis bekerja sebagai staf peneliti pada Kelompok Peneliti Hama dan Penyakit Tanaman, Balai Penelitian Tanaman Sayuran di Lembang, Bandung, Jawa Barat sejak tahun 1990. Penulis mendapat kesempatan untuk melanjutkan studi Program Magister Pertanian di Universitas Gajah Mada pada tahun 1999 dengan beasiswa dari Badan Penelitian dan Pengembangan Pertanian melalui proyek ARMP-II. Penulis lulus dan mendapat gelar Magister Pertanian (M.P.) pada tahun 2002. Penulis diterima sebagai mahasiswa Program Doktor di Sekolah Pascasarjana Institut Pertanian Bogor pada tahun 2007 dengan beasiswa dari Badan Penelitian dan Pengembangan Pertanian, Kementrian Pertanian.
DAFTAR ISI
Halaman
DAFTAR TABEL... xiii
DAFTAR GAMBAR... xiv
DAFTAR LAMPIRAN... xv
I. PENDAHULUAN... 1
Latar Belakang... 1
Alur Penelitian... 4
Tujuan Penelitian... 7
Daftar Pustaka... 7
II. TINJAUAN PUSTAKA... 10
Biologi dan Kisaran Inang B. tabaci... 10
Peranan B. tabaci dalam Penularan Penyakit Daun Keriting Kuning Cabai... 13
Peranan Teknik Budidaya Khususnya Pemanfaatan Tanaman Pinggir untuk Pengendalian Hama dan Konservasi Predator... 15 Peranan Predator dalam Pengendalian B. tabaci... 17
Preferensi Predator terhadap Spesies Mangsa... 19
Tanggap Fungsional Predator terhadap Mangsa... 19
Daftar Pustaka... 21
III. PENGARUH PEMANFAATAN TANAMAN PEMBATAS PINGGIR DI PERTANAMAN CABAI MERAH TERHADAP DINAMIKA POPULASI KUTUKEBUL B. tabaci DAN INSIDENSI PENYAKIT DAUN KERITING KUNING... 27
Abstrak... 27
Abstract……… 28
Pendahuluan………. 28
Bahan dan Metode……… 30
Hasil dan Pembahasan... 33
Simpulan... 42
Daftar Pustaka... 43
IV. PENGARUH PEMANFAATAN TANAMAN PEMBATAS PINGGIR TERHADAP KELIMPAHAN PREDATOR PENTING B. tabaci PADA PERTANAMAN CABAI MERAH.. 46
Abstrak... 46
Abstract……… 46
Pendahuluan………. 47
Bahan dan Metode……… 48
Hasil dan Pembahasan... 50
Simpulan... 55
V. DAYA PEMANGSAAN, PREFERENSI DAN TANGGAP FUNGSIONAL PREDATOR TERHADAP B. tabaci PADA
TANAMAN CABAI MERAH... 58 Abstrak... 58 Abstract... 59 Pendahuluan... 59 Bahan dan Metode... 61 Hasil dan Pembahasan... 65 Simpulan... 71 Daftar Pustaka... 72
VI. PEMBAHASAN UMUM... 74
VII. SIMPULAN DAN SARAN... 81 DAFTAR PUSTAKA... 84
LAMPIRAN... 91
DAFTAR TABEL
Halaman
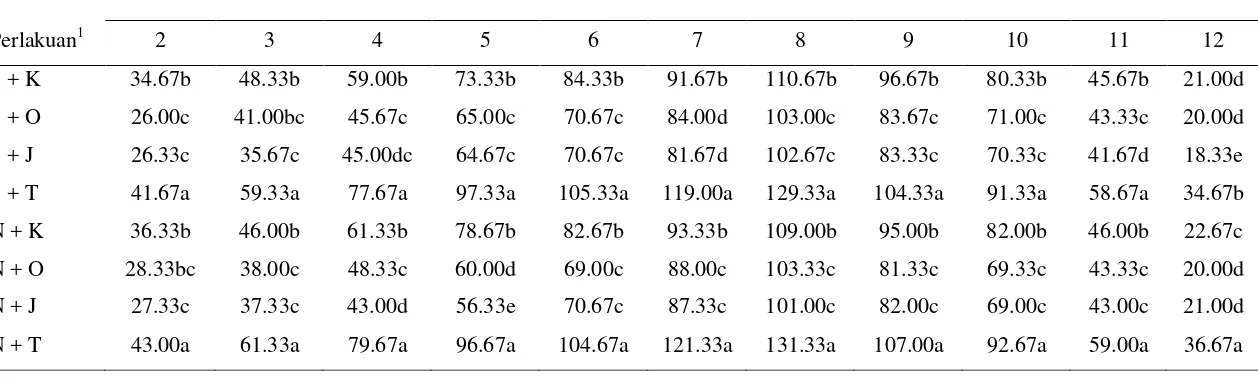
3.1 Rerata jumlah imago B. tabaci/perangkap di pertanaman cabai merah dengan kombinasi perlakuan pesemaian dan spesies
tanaman pembatas pinggir... 35
3.2 Rerata jumlah nimfa B. tabaci/daun di pertanaman cabai merah dengan kombinasi perlakuan pesemaian dan spesies tanaman
pembatas pinggir... 37
3.3 Pengaruh perlakuan terhadap nilai AUDPC dan penekanan penyakit daun keriting kuning (%) di pertanaman cabai pada
pengamatan 2 - 12 minggu setelah tanam... 40
4.1 Spesies dan jumlah serangga predator B. tabaci/15 tanaman/6 pengamatan di pertanaman cabai merah dengan perlakuan
berbagai spesies tanaman pembatas pinggir... 52
5.1 Daya pemangsaan beberapa predator terhadap nimfa dan imago
B. tabaci di laboratorium... 66 5.2 Rerata jumlah B. tabaci, Trips dan Kutudaun yang dimangsa oleh
berbagai spesies predator Coccinellidae dan index preferensi berbagai spesies predator terhadap berbagai spesies mangsa pada
Cabai merah... 68 5.3 Rerata mangsa yang dikonsumsi (x ± SD) predator V. Lineata
DAFTAR GAMBAR
Halaman
1.1 Alur penelitian potensi pemanfaatan tanaman pembatas pinggir dan predator unntuk pengendalian Bemisia tabaci (Gennadius)
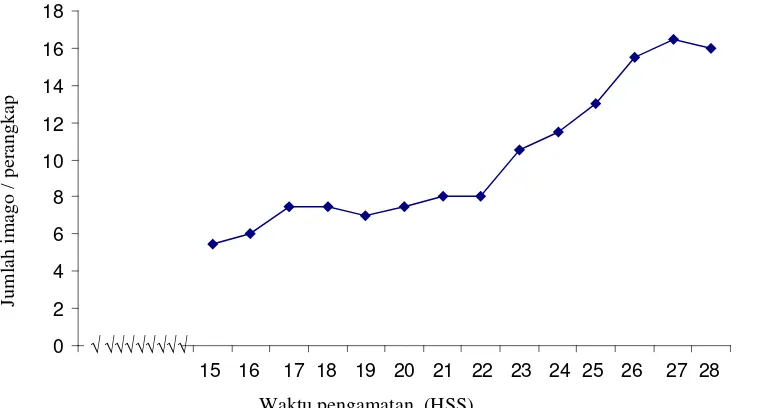
pada tanaman cabai merah,... 6 3.1 Rerata jumlah imago B. tabaci pada perangkap kuning di
pesemaian cabai merah... 33
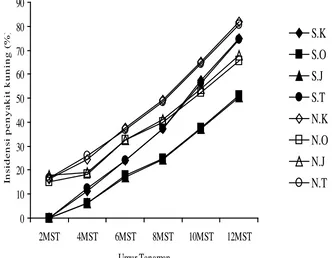
3.2 Rerata insidensi penyakit daun keriting kuning cabai (%) pada pertanaman cabai merah dengan kombinasi perlakuan pesemaian
dan jenis tanaman pembatas pinggir... 39
3.3 Regresi populasi B. tabaci dengan insidensi penyakit daun
keriting kuning cabai... 40
3.4 Rerata bobot buah cabai per petak (kg) pada pertanaman cabai merah dengan kombinasi perlakuan pesemaian dan spesies
tanaman pembatas pinggir... 41
3.5 Regresi antara insidensi penyakit daun keriting kuning dengan
bobot buah cabai merah... 42
4.1 Spesies serangga predator B. tabaci yang ditemukan di
pertanaman cabai merah dengan tanaman pembatas pinggir... 51
4.2 Kelimpahan predator/15tanaman dan jumlah nimfa B. tabaci/daun di pertanaman cabai merah dengan berbagai pembatas pinggir:
(A) predator dan (B) nimfa B. tabaci... 54
4.3 Regresi populasi B. tabaci dengan kelimpahan predatornya di
pertanaman cabai merah... 55 5.1 Rerata nilai pengamatan proporsi mangsa yang dimakan (titik)
dan penduga (garis) berdasarkan hasil analisis regresi logistik... 70
5.2 Kurva tanggap fungsional predator V. lineata terhadap
peningkatan kerapatan mangsa B. tabaci... 70
6.1 Model strategi pengendalian B. tabaci di pertanaman cabai merah dengan cara perpaduan antara penggunaan tanaman jagung
DAFTAR LAMPIRAN
Halaman
1 Perlakuan penggunaan sungkup di pesemaian dan spesies tanaman pembatas pinggir di lahan pertanaman cabai merah: (a) Penggunaan sungkup di pesemaian; (b) Pesemaian tanpa sungkup; (c) Pembatas pinggir kain sifon; (d) Jagung sebagai tanaman pinggir; (e) Orok-orok sebagai tanaman pinggir; (f) Tanpa
pembatas pinggir... 92
2 Tata letak petak percobaan pengaruh pemanfaatan tanaman pembatas pinggir terhadap populasi B. tabaci dan kelimpahan
predator... 93
3 Cara pengamatan terhadap imago dan nimfa B. tabaci serta predator pada pertanaman cabai merah: (a) Pengamatan imago pada perangkap kuning; (b) Pengamatan nimfa pada daun cabai; (c) Pengambilan predator dengan menggunakan penyedot
serangga... 94 4 Imago dan nimfa B. tabaci serta gejala penyakit daun keriting
kuning cabai: (a) Imago B. tabaci pada perangkap kuning; (b)
Nimfa B. tabaci; (c) Gejala penyakit daun keriting kuning cabai... 95 5 Pelaksanaan penelitian kajian potensi predator Coccinellidae
terhadap B. tabaci: (a) Pembiakan massal B. tabaci dan kutudaun pada pertanaman cabai merah; (b) Percobaan uji daya pemangsaan;
(c) Percobaan uji tanggap fungsional; (d) Percobaan uji preferensi.. 96 6 Predator Verania lineada: (a) Imago betina; (b) Imago jantan; (c)
I. PENDAHULUAN
Latar Belakang
Cabai merah (Capsicum annuum L.) merupakan komoditas sayuran yang
mempunyai nilai ekonomi tinggi dan dapat memacu peningkatan sumber
pendapatan petani, substitusi impor dan penghasil devisa. Areal pertanaman cabai
merah dari tahun 2005 – 2010 selalu menduduki areal terluas di antara tanaman
sayuran yang diusahakan di Indonesia, dan pada tahun 2010 luas panen komoditas
cabai merah mencapai 194.64 ha (Direktorat Jendral Tanaman Pangan dan
Hortikultura 2010). Produktivitas cabai merah selama 5 tahun terakhir
menunjukkan peningkatan yang berarti, dan pada tahun 2010 mencapai
6.94ton/ha. Namun demikian, tingkat produktivitas cabai merah tersebut masih
lebih rendah dibandingkan dengan potensi produksi cabai merah yang dapat
mencapai 10 ton/ha (Suwandi et al. 1989). Hal ini disebabkan antara lain adanya
serangan organisme pengganggu tumbuhan (OPT). Duriat et al. (2005)
melaporkan bahwa terdapat 15 jenis OPT yang menyerang tanaman cabai merah.
Kutukebul Bemisia tabaci (Gennadius) (Hemiptera: Aleyrodidae)
merupakan salah satu hama penting pada tanaman sayuran terutama pada famili
Solanaceae termasuk cabai merah. Hama ini pertama kali ditemukan di Indonesia
pada tahun 1938 pada tanaman tembakau di Deli, Sumatera Utara (Kalshoven
1981). Kutukebul menjadi hama yang sangat penting terutama pada pertanaman
cabai merah di Indonesia sejak tahun 2001 (Sulandari et al. 2001). Kutukebul
dapat menimbulkan kerusakan secara langsung yaitu berupa bercak nekrotik,
klorosis dan daun gugur, dan secara tidak langsung yaitu sebagai vektor virus
penyebab penyakit daun keriting kuning cabai (Pepper Yellow Leaf Curl
Begomovirus/PepYLCV) (Byrne dan Bellows 1990; Oliveira et al. 2001; Jones
2003; Hidayat et al. 2006). Penyakit daun keriting kuning bersifat epidemik pada
pertanaman cabai di berbagai daerah seperti di Jawa Barat, Jawa Tengah, DI
Yogyakarta, Sumatera Selatan dan Lampung (Sulandari et al. 2004; Hidayat et al.
2006). Perkembangan luas serangan penyakit daun keriting kuning di beberapa
daerah di Indonesia terutama di pulau Jawa sangat cepat. Pada tahun 2003 luas
serangan penyakit berkisar antara 6.2 ha sampai 60 ha dan pada tahun 2008
Program Tanaman Pangan dan Hortikultura 2010). Kerusakan akibat serangan
penyakit daun keriting kuning pada pertanaman cabai dapat sangat berat sehingga
kerugian ekonomi dapat mencapai 20% sampai 100% (Brown 1994; Sulandari et
al. 2006). Penularan Begomovirus hanya terjadi melalui imago B. tabaci dan tidak
dapat melalui kontak atau biji (Aidawati et al. 2002; Jones 2003; Sulandari 2004;
Hidayat dan Rahmayani 2007). Oleh karena itu pengendalian terhadap B. tabaci
merupakan salah satu strategi untuk menekan kejadian penyakit daun keriting
kuning cabai.
Upaya pengendalian B. tabaci yang umum dilakukan petani selama ini
adalah dengan penggunaan insektisida secara intensif. Penggunaan insektisida
yang berlebihan, disamping merupakan pemborosan juga dapat membahayakan
manusia dan menimbulkan berbagai kerugian terhadap lingkungan, antara lain
terbunuhnya organisme bukan sasaran seperti predator dan parasitoid, dan
terjadinya resistensi hama terhadap insektisida. Sugiyama (2005) dan Setiawati et
al. (2007) melaporkan bahwa B. tabaci sudah mulai menunjukkan gejala resisten
terhadap beberapa jenis insektisida seperti golongan organofosfat, karbamat dan
piretroid sintetik. Untuk itu diperlukan upaya pengendalian yang lebih
berlandaskan pendekatan ekologi dan ekonomi, yaitu tidak mencemari
lingkungan, aman bagi pemakai dan konsumen cabai merah, relatif murah, tetapi
juga efektif terhadap hama B. tabaci.
Salah satu konsep pengendalian yang lebih berlandaskan pada pendekatan
ekonomi dan ekologi adalah pengendalian hama terpadu (PHT). Perkembangan
konsep PHT saat ini dan dimasa mendatang mengarah pada rakitan teknologi yang
bersifat bio – intensif, yang berupaya memanfaatkan sumberdaya hayati yang ada
di alam, seperti musuh alami, varietas tahan, pestisida nabati, dan tanaman
penolak, tanaman penarik, atau tanaman pinggir (Frisbie dan Smith 1991; Hoddle
et al. 1998).
Pemanfaatan tanaman pembatas pinggir (border crops) merupakan salah
satu alternatif pengendalian yang dapat menekan populasi B. tabaci di pertanaman
cabai merah dan aman terhadap lingkungan. Menurut Difanzo et al. (1996) dan
Fereres (2000) pemanfaatan tanaman pembatas pinggir dapat menekan kejadian
3
Perfecto dan Sediles (1992) penggunaan tanaman bukan inang dalam sistem pola
tanam tumpangsari, selain berperan sebagai samaran yang membuat tanaman
inang sulit ditemukan, juga berperan sebagai penghalang fisik bagi hama untuk
menemukan tanaman yang diusahakan. Pemanfaatan tanaman jagung (Zea mays)
sebagai pembatas pinggir di pertanaman kentang dapat menekan kejadian
penyakit virus Y kentang (PVY) yang ditularkan oleh kutudaun. Selanjutnya
dilaporkan oleh Muthomi et al. (2010) tanaman jagung yang ditanam dengan
jarak 0,5 dan 1 m dari pertanaman kentang dapat menekan populasi kutudaun dan
kejadian penyakit virus kentang sampai 48 %. Hasil penelitian lain menunjukkan
bahwa pemanfaatan orok-orok (Crotalaria juncea) sebagai tanaman pembatas
pinggir di pertanaman “zucchini” dapat menekan populasi kutudaun dan kejadian
penyakit virus bercak cincin papaya (PRSV) strain semangka (Roshan dan Cerruti
2011).
Pemanfaatan tanaman pembatas pinggir merupakan salah satu alternatif
pengendalian yang kompatibel jika dipadukan dengan musuh alami dalam hal ini
predator. Jenis tanaman pembatas pinggir yang dipilih harus mempunyai fungsi
ganda yaitu, sebagai penghalang masuknya imago B. tabaci ke pertanaman cabai
merah, dan dapat mendorong konservasi musuh alami seperti predator, yaitu
sebagai tanaman refugia yang berfungsi untuk berlindung sementara dan penyedia
polen untuk makanan alternatif jika mangsa utama populasinya rendah atau tidak
ada (Untung 2006). Teknik konservasi bertujuan menghindarkan
tindakan-tindakan yang dapat menurunkan populasi musuh alami termasuk predator.
Teknik augmentasi yang memanipulasi atau modifikasi ekosistem sehingga lebih
mendorong peningkatan populasi dan efektifitas serta efisiensi musuh alami,
dimasukkan ke dalam teknik konservasi (Stehr 1982). Menurut Pedigo (1991)
populasi predator dan parasitoid terutama yang generalis pada vegetasi yang
beragam relatif stabil, dan bertahan lama, sebab makanan (tepung sari dan nektar)
tersedia lebih berkesinambungan, serta adanya tempat berlindung dan
mikrohabitat yang sesuai. Untung (2006) mengungkapkan dalam penerapan PHT
konservasi musuh alami terutama predator dan parasitoid merupakan teknik
Studi musuh alami Bemisia spp. di Brasil menemukan sekitar 14 spesies
predator, diantaranya dari jenis predator kumbang kubah ( lady beetle ),
Nephaspis hydra Gordon dan Delphastus davidsoni Gordon diketahui pertama
kali sebagai pemangsa Bemisia spp. (Gerling 1990; Olsen 2001). Setiawati
(2005), mengungkapkan beberapa spesies predator yang diketahui efektif terhadap
B. tabaci antara lain Coccinella transversalis, Menochilus sexmaculatus,
Coenosia attenuate, Delphastus pusillus, Deracocoris pallens, Euscius hibisci,
Orius albidipennis, Scymus syriacus. dan Chrysoperla carnea. Sudrajat (2009)
melaporkan bahwa dari hasil explorasi musuh alami di Kabupaten Bandung dan
Karawang pada tahun 2005 ditemukan beberapa jenis predator yang mempunyai
potensi untuk mengendalikan hama kutukebul (B. tabaci) pada pertanaman
sayuran. Jenis predator kutukebul yang ditemukan dan mempunyai potensi adalah
beberapa spesies dari famili Coccinellidae (Menochilus sexmaculatus, Coccinella
transfersalis, Harmonia sp., Curinus sp. dan Delphastus sp.) dan famili
Stapilinidae (Paederus fusipes).
Pemanfaatan tanaman jagung atau orok-orok sebagai pembatas pinggir pada
pertanaman cabai merah untuk pengendalian B. tabaci telah dilakukan oleh para
petani terutama di daerah Jawa Tengah. Meskipun demikian dalam cara
pelaksanaannya seperti jarak dan waktu tanam antara tanaman pembatas pinggir
dan tanaman cabai merah kurang tepat, sehingga hasilnya kurang maksimal. Oleh
karena itu perlu dilakukan kajian secara ilmiah mengenai pengaruh tanaman
pembatas pinggir terhadap populasi B. tabaci dan insidensi penyakit daun keriting
kuning cabai serta kelimpahan predator penting.
Alur Penelitian
Pengendalian hama terpadu (PHT) terdiri atas 5 komponen pengendalian
yaitu penggunaan insektisida, penanaman varietas tahan, pengendalian secara fisik
dan mekanik, teknik bercocok tanam dan pengendalian hayati (musuh alami).
Untuk pengendalian hama B. tabaci sebagai vektor Begomovirus pada
pertanaman cabai merah, penggunaan insektisida tidak dianjurkan karena
disamping membahayakan lingkungan juga telah banyak dilaporkan bahwa B.
5
mekanik seperti pengunaan perangkap warna dengan memperhitungkan sifat
biologi dan ekologi hama dapat menekan populasi hama, tetapi pengendalian ini
masih mengundang kontroversi. Sebagian petani berpendapat kedua pengendalian
tersebut kurang praktis dan justru mengundang hama masuk ke lahan pertanaman
yang diusahakan. Sementara penanaman varietas tahan hama atau penyakit untuk
pengendalian B. tabaci dan Begomovirus yang ditularkannya belum dapat
dikembangkan karena sampai saat ini belum diperoleh varietas tanaman cabai
merah yang benar-benar tahan terhadap Begomovirus.
Penelitian ini dirancang untuk mengefaluasi penggabungan pemanfaatan
tanaman pembatas pinggir dengan potensi predator untuk menekan populasi B.
tabaci dan insidensi penyakit daun keriting kuning pada pertanaman cabai merah
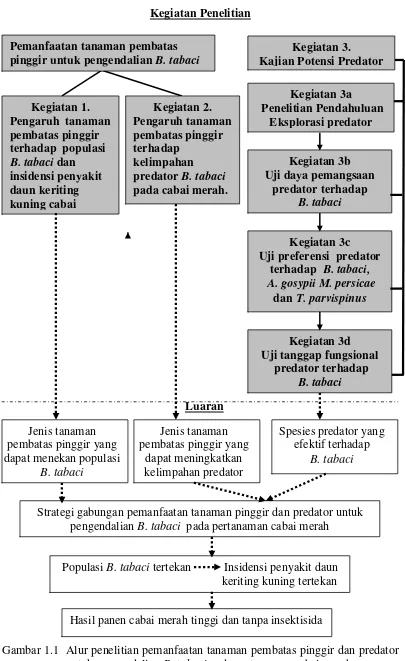
(Gambar 1.1).
Penelitian dibagi menjadi 2 kelompok. Kelompok pertama adalah penelitian
pemanfaatan tanaman pembatas pinggir untuk pengendalian B. tabaci dan
kejadian penyakit daun keriting kuning pada pertanaman cabai merah, serta
peranannya dalam konservasi predator. Luaran yang diharapkan dalam penelitian
ini ialah diperoleh jenis tanaman pembatas pinggir yang efektif dalam
mengendalikan B. tabaci dan menekan insidensi penyakit daun keriting kuning
pada pertanaman cabai, tetapi tidak membahayakan bahkan mendorong dalam
konservasi predator B. tabaci.
Kelompok kedua adalah penelitian potensi predator untuk pengendalian B.
tabaci. Penelitian ini terdiri atas empat tahapan percobaan, yaitu: 1) Eksplorasi
predator B. tabaci di daerah sentra produksi cabai merah, 2) Uji daya pemangsaan
berbagai predator (yang dominan ditemukan dalam eksplorasi) terhadap B. tabaci;
3) Uji preferensi berbagai predator (yang mempunyai daya pemangsaan tinggi
dari percobaan 2) terhadap B. tabaci, Thrips dan kutudaun; 4) Uji tanggap
fungsional predator (yang mempunyai preferensi tertinggi terhadap B. tabaci)
terhadap B. tabaci. Secara keseluruhan luaran dari penelitian ini adalah dapat
diperolehnya jenis predator yang benar-benar efektif untuk pengendalian B.
Kegiatan Penelitian
Luaran
[image:34.595.83.489.79.740.2]
Gambar 1.1 Alur penelitian pemanfaatan tanaman pembatas pinggir dan predator untuk pengendalian B. tabaci pada pertanaman cabai merah
Kegiatan 1. Pengaruh tanaman pembatas pinggir terhadap populasi
B. tabaci dan insidensi penyakit daun keriting kuning cabai
Kegiatan 3b Uji daya pemangsaan
predator terhadap
B. tabaci
Strategi gabungan pemanfaatan tanaman pinggir dan predator untuk pengendalian B. tabaci pada pertanaman cabai merah
Hasil panen cabai merah tinggi dan tanpa insektisida Populasi B. tabaci tertekan Insidensi penyakit daun
keriting kuning tertekan Kegiatan 3a Penelitian Pendahuluan Eksplorasi predator Kegiatan 2. Pengaruh tanaman pembatas pinggir terhadap kelimpahan predator B. tabaci
pada cabai merah. Pemanfaatan tanaman pembatas
pinggir untuk pengendalian B. tabaci
Kegiatan 3. Kajian Potensi Predator
Kegiatan 3c Uji preferensi predator
terhadap B. tabaci, A. gosypiiM. persicae
dan T. parvispinus
Kegiatan 3d Uji tanggap fungsional
predator terhadap
B. tabaci
Jenis tanaman pembatas pinggir yang dapat menekan populasi
B. tabaci
Jenis tanaman pembatas pinggir yang
dapat meningkatkan kelimpahan predator
Spesies predator yang efektif terhadap
7
Tujuan Penelitian
Tujuan penelitian untuk memperoleh strategi teknik pemanfaatan tanaman
pembatas pinggir yang digabungkan dengan pemanfaatan predator untuk
mengendalikan B. tabaci dan insidensi penyakit daun keriting kuning pada
pertanaman cabai merah. Secara khusus penelitian bertujuan: 1) Mengetahui
pengaruh tanaman pinggir terhadap dinamika populasi B. tabaci dan insidensi
penyakit daun keriting kuning cabai; 2) Mengetahui pengaruh tanaman pinggir
terhadap kelimpahan predator penting yang menyerang B. tabaci; 3)
Mengidentifikasi dan mengetahui dominansi spesies predator yang menyerang B.
tabaci di pertanaman cabai merah; 4) Mengevaluasi efektivitas berbagai spesies
predator (dominan dari eksplorasi) terhadap B. tabaci melalui uji daya
pemangsaan, preferensi dan tanggap fungsional.
Daftar Pustaka
Aidawati N, Hidayat SH, Suseno R, Sosromarsono S. 2002. Transmission of an Indonesian isolate of tobacco leaf curl virus (Begomovirus) by Bemisia tabaci Genn. (Hemiptera: Aleyrodidae) Plant Pathology 18: 231 – 236.
Brown JK. 1994. Current status of Bemisia tabaci as a plant pest and virus vector in agroecosystems word wide. Plant Protection Bulletin. 42: 3 – 32.
Byrne DN, Bellows TS. 1990. Whitefly biology. Annnual Review of Entomology
36 : 431 – 457.
Difanzo CD, Rogsdale DW, Radcliffe NC, Sencor GA. 1996. Crop borders reduce potato virus Y incidence in seed potato. Annals of Applied Biology
129: 289–302.
Direktorat Jendral Tanaman Pangan dan Hortikultura. 2010. Hasil pembangunan Pertanian Tanaman Pangan : Aspek Areal Produksi dan Faktor-faktor
Produksi. Jakarta (ID): Direktorat Jendral Tanaman Pangan dan
Hortikultura.
Duriat AS, Ratnawati ML, Kirana R, Widjaja ES, Sulastrin I, Gunaen N, Gunawan OS, Gaswanto R, Wulandari AW, Murtiningsih RR, van der Wolf JM, van der Zouwen PS. 2005. The most important pests and seed-borne diseases of vegetables in Indonesia. A Progress report in 2003-2005. Hortin Project. Indonesian - Netherlands research colaboration. 26 pages.
Frisbie RE, Smith Jr JW. 1991. Biologically intensive integrated pest management : the future. Di dalam: Menn JJ dan Steinhaner AL. International Progress and Perspective for 21th
Gerling D. 1990. Natural enemies of whitelies; predator and parasitoids. Di dalam: Garling D. Whiteflies: Their Bionomics, Pest Status and
Management. Andover (US): Intercept Ltd. hlm 147–185.
Century. Maryland (US): Entomol. Soc. Amer. Maryland.
Hidayat SH. 2003. Rangkuman Hasil Penelitian Begomovirus di Indonesia : Sebagai Bahan Diskusi Untuk Menghadapi Peningkatan Infeksi Begomovirus Pada Cabai. Makalah pada Seminar Sehari Pengenalan dan Pengendalian Penyakit Virus Pada Cabai. Dir. Perlindungan Hortikultura, Dir. Jen. Bina Produksi Hortikultura. Jakarta. 4 hal.
Hidayat SH, Chatchawankanpanich O, Rusli E, Aidawati N. 2006. Begomovirus associated with pepper yellow leaf curl diseases in West Java, Indonesia.
Journal Indonesian Microbiology 11(2): 87 – 89.
Hidayat SH, Rahmayani E. 2007. Transmission of tomato leaf curl begomovirus by two different species of whitefly (Hemiptera: Aleyrodidae). J. Plant Pathol. 23 (2): 57– 61.
Hoddle MS, van Driesche RG, Sanderson JP. 1998. Biology and use of the whitefly parasitoid Encarsia formosa. Annual Review of Entomology 43 : 645 – 669.
Jones D. 2003. Plant viruses transmitted by whiteflies. European Journal Plant Pathology 10(9): 197- 221.
Kalshoven LGE. 1981. The Pests of Crops in Indonesia. Laan PA van der, penerjemah. Jakarta (ID): Ichtiar Baru- van Hoeve. Terjemahan dari: De Plagen vande Cultuurgewassen in Indonesie.
Oliveira MRV, Henneberry TJ, Anderson P. 2001. History, current status and collaborative research projects for Bemisia tabaci. Crop Protection 20:709– 723.
Olsen LV. 2001. The behavior of the Ladybird and its ability as a predator.
Available at http:
Pedigo LP. 1991. Entomologi and pest management. MacMillan Publishing company New York. Collier MacMillan Publishers. London. 646p.
Perfecto I, Sediles A. 1992. Vegetational diversity, ants (Hymenoptera: Formicidae), and herbivorous pest in a neotropical agroecosystem.
Environment Entomology 21(1):61– 67.
9
Pertemuan Apresiasi Penerapan Penganggulangan Virus Cabai, Yogyakarta, 14–15 April 2005.
Setiawati W, Udiarto BK, Soetiarso TA. 2007. Selektivitas Beberapa Insektisida terhadap Hama Kutu Kebul (Bemisia tabaci Genn.) dan Predator Menochilus sexmaculatus Fabr. Jurnal Hortikultura Vol.17, No. 2, Tahun 2007.
Sudrajat. 2009. Eksplorasi Musuh Alami Kutukebul (Bemisia tabaci) di Jawa Barat (Pangalengan, Ciwidae, Lembang dan Krawang) pada Tanaman Sayuran. Laporan Sementara Hasil Penelitian untuk Disertasi S-3. Universitas Padjadjaran.
Sugiyama K. 2005. Management of whitefly for commercial tomato production in greenhouses in Shizuoka, Japan. In. Proc. of the International Seminar on Whitefly Management and Control Strategy. Taichung, Taiwan. Oct 3 – 8, 2005. pp.81 – 91.
Sulandari S, Hidayat SH, Suseno R, Jumanto H, Sosromarsono S. 2001. Keberadaan virusgemini pada cabai di DIY. Konggres Nasional dan
Seminar Ilmiah PFI ke XVI. Bogor, Agustus 2001.
Sulandari S, Suseno R, Hidayat SH, Harjosudarno J, Sosromarsono S. 2006. Deteksi dan kajian kisaran inang virus penyebab penyakit daun keriting kuning cabai. Journal Hayati 13(4): 1 – 6.
Sulandari S. 2004. Karakterisasi biologi, serologi, dan analisa sidik jari DNA virus penyebab penyakit daun kuning keriting cabai [disertasi]. Bogor (ID): Sekolah Pascasarjana, Institut Pertanian Bogor.
Suwandi, Nurteka N, Sahat S. 1989. Bercocok tanam sayuran dataran rendah. Lembang (ID): Laporan Balai Penelitian Hortikultura Lembang dan Proyek ATA 395. 31 – 36.
Stehr DW. 1982. Parasitoid and Predator in Pest Management. Dalam Metcalf and Luckmann WH (ed.) Introduction to Insect Pest Management. John Wiley and Sons. New York. 135 – 173p.
Biologi dan Kisaran Inang Bemisia tabaci (Gennadius)
Bemisia tabaci (Gennadius) digolongkan ke dalam ordo Hemiptera, subordo
Sternorrhyncha, superfamili Aleyrodoidea, famili Aleyrodidae (Martin et al.
2000). B. tabaci pertama kali ditemukan di Indonesia pada tahun 1938 pada
tanaman tembakau (Kalshoven 1981). Pada tahun 2001 B. tabaci telah menjadi
hama utama terutama pada berbagai jenis tanaman sayuran dan tersebar ke seluruh
Pulau Jawa, Sumatera, Kalimantan dan Sulawesi, dan sejak awal tahun 2004 telah
pula menjadi hama penting di Bali (Sulandari 2001; Setiawati et al. 2005).
Perkembangan B. tabaci terdiri atas empat stadia, yaitu dimulai dari telur,
nimfa, pupa dan imago. Telur B. tabaci . bentuknya lonjong (oval), warnanya
putih bening ketika baru diletakkan, kemudian kecokelatan menjelang menetas.
Telur berdiameter 0,25 mm, dan biasanya diletakkan pada permukaan bawah daun.
Jumlah telur yang dihasilkan seekor betina mencapai 28 sampai 300 butir
tergantung pada tanaman inang dan suhu lingkungan (Hirano et al., 2002). Pada
kapas, B. tabaci. rata-rata bertelur 81 butir pada suhu 26,7ºC atau 72 butir pada
suhu 32,2ºC dengan masa inkubasi telur 5 hari (Butler et al., 1983). Stadia telur
pada tanaman tomat adalah 6,8–8,7 hari pada suhu 25 0C dan RH 65% (Salas dan
Mendoza 1995). Rata-rata stadia telur pada Hibiscus rosa-sinensis kultivar Pink
Versicolor adalah 6,3 hari dan Brilliant Red adalah 6,7 hari pada suhu 26,7 0
Nimfa B. tabaci terdiri dari tiga instar dan instar ke-4 dianggap sebagai
transisi dan dinamakan instar ke-4 atau pupa karena peralihan antara dua stadia
yang singkat dan sulit untuk dipisahkan (Salas dan Mendoza 1995). Byrne dan
Bellows (1991) menyatakan bahwa nimfa instar ke-4 biasanya dikenal sebagai
pupa. Nimfa instar satu (panjang + 0,223 mm, lebar + 0,131 mm) berbentuk bulat
panjang, berwarna hijau cerah, pada pinggir tubuhnya terdapat bulu bulu halus
dan lapisan lilin yang tipis. Nimfa instar 1 yang baru keluar dari telur aktif
bergerak dan mengisap cairan makanan pada permukaan bawah daun selama 1-2
hari, dan setelah mendapatkan tempat yang sesuai akan menetap dan tidak
bergerak lagi (Badri 1983). Nimfa instar 2 (panjang + 0,283 mm, lebar + 0,178 C dan
11
mm), nimfa instar 3 (panjang + 0,470 mm, lebar + 0,312 mm) dan nimfa instar 4
(umum disebut pupa) tidak bergerak, berwarna hijau gelap, tungkai tereduksi,
pada bagian dorsal terdapat tiga pasang duri (Badri. 1983). Stadia nimfa instar
pertama pada tanaman tomat adalah 4,0 hari, nimfa instar kedua dan ketiga pada
tanaman tomat adalah 2,7 dan 2,5 hari (Salas dan Mendoza 1995). Rata-rata stadia
nimfa instar pertama pada Hibiscus rosa-sinensis kultivar Pink Versicolor adalah
4,2 hari dan kultivar Brilliant Red adalah 4,3 hari pada suhu 26,7 0C dan RH 55%
(Liu dan Stansly 1998). Menurut Gameel (1977) perkembangan nimfa secara
keseluruhan berlangsung selama 12-15 hari pada suhu 28o-32oC, dan 28 – 32 hari
pada suhu 20o-24oC. Pada suhu lebih tinggi yaitu 30o-34oC periode
perkembangan lebih cepat, dan sebaliknya menjadi lebih lama apabila suhu
mencapai 18o - 22o
Imago B. tabaci berwarna kekuningan dan tubuhnya tertutup oleh sekresi
seperti tepung lilin yang berasal dari kelenjar lilin yang terletak pada ruas
abdomen pertama dan kedua pada imago jantan, sedangkan pada imago betina
terletak pada ruas abdomen ke tiga dan ke empat. Sayap depan berwarna putih dan
mempunyai pembuluh radial sektor yang bercabang satu dan pembuluh kubitus
lurus. Antena tujuh ruas dengan ruas terakhir meruncing dan ditutupi oleh rambut
rambut yang halus. Mata majemuk berkembang sempurna (Kalshoven 1981).
Imago B. tabaci berukuran 1,0-1,5 mm. Panjang sayap depan + 0,673 mm dan
lebar + 0,246 mm, sedangkan panjang sayang belakang + 0,572 dan lebar + 0,209
mm (Badri 1983). Lama hidup imago dipengaruhi oleh lingkungan dan faktor
faktor lainnya. Lama hidup imago jantan pada umumnya lebih pendek dibanding C .
Nimfa instar 4 umumnya disebut pupa. Pupa B. tabaci berbentuk bulat
panjang berwarna kuning, bagian toraks agak melebar dan cembung, ruas
abdomen tampak jelas. Pinggir puparium tidak rata dan pada bagian dorsal
terdapat tujuh pasang seta (duri) dan satu pasang pada ujung anal. Vasiform orifice
berbentuk segi tiga dan memanjang, operkulum menutupi hampir separuh bagian
dari vasiform orifice (Kalshoven 1981). Panjang pupa + 0,613 mm dan lebar +
0,421 mm. Lamanya stadium pupa rata rata 2,51 + 0,16 hari (Badri 1983). Stadia
betina. Lama hidup imago jantan berkisar antara 9,54-17,20 hari, sedangkan
betina mencapai 37,75- 74,20 hari (Gameel 1977).
Tanaman inang adalah tanaman yang dapat memenuhi kebutuhan hidup
serangga baik yang berhubungan dengan makanan (kebutuhan gizi) maupun
dalam hubungannya dengan perilaku. Serangga yang hidup pada tanaman inang
yang sesuai berkembang biak lebih cepat dari pada yang hidup pada tanaman
inang yang kurang sesuai (Beck 1965; House 1965).
B. tabaci bersifat polifag (Costa dan Brown 1990), sejumlah besar spesies
tanaman tahunan dan setahun yang telah dibudidayakan maupun yang belum
dibudidayakan sesuai untuk makan dan/atau reproduksi (Bedford et al. 1992;
Brown et al. 1992). B. tabaci mempunyai kisaran inang lebih dari 600 spesies
tanaman (Greathead 1986) yang berasal dari 63 famili tanaman, dan sebanyak
50% spesies tanaman yang merupakan inang kutukebul berasal dari famili
Fabaceae, Asteraceae, Malvaceae, Solanaceae dan Euphorbiaceae (Mound dan
Halsey 1978). Di antara famili tersebut 99% spesies tanaman yang merupakan
inang kutukebul adalah Fabaceae (Basu 1995).
Di Brazil, kutukebul juga telah menginfestasi gulma sebagai inang. Gulma
yang menjadi inang kutukebul tersebut adalah: Cleome espinosa (Cleomaceae),
Senna obtusifolia (Fabaceae), Herisanthia hemoralis (Malvaceae), Richardia
grandiflora, Borreriaverticilliata (Rubiaceae), Waltheria indica, W. Rotundifolia
(Sterculicaceae), dan Stachytarpheta sanguinea (Verbenaceae) (Lima et al. 2000).
Selanjutnya Simmon et al. (2000) melaporkan bahwa inang baru kutukebul di
Amerika Serikat adalah: Hyperium perfolatum (Hypericaceae), Valeriana
officinalis (Valerianaceae), Tanacetaum parthenium, Echinaceae pallida, E.
purpurea (Asteraceae).
Henridval et al. (2011) melaporkan kisaran inang B. tabaci yang tumbuh di
sekitar pertanaman cabai merah di Indonesia tepatnya di Kecamatan Pakem,
Kabupaten Sleman, DI Yogyakarta terdiri atas 22 spesies gulma dan lima spesies
tanaman budidaya lainnya. Inang tersebut meliputi 13 famili yaitu Amaranthaceae,
Araceae, Asteraceae, Brassicaceae, Capparidaceae, Convolvulaceae,
Euphorbiaceae, Lamiaceae, Oxalidaceae, Rubiaceae, Papilionaceae, Solanaceae,
13
populasi B. tabaci paling banyak dibandingkan dengan famili lainnya. Gulma A.
conyzoides berperan sebagai inang B. tabaci, inang alternatif Geminivirus, dan
reservoir parasitoid Eretmocerus sp. Gulma A. boehmerioides hanya berperan
sebagai inang B. tabaci dan inang alternatif Geminivirus.
Hama B. tabaci dilaporkan telah ada sejak tahun 1800 di Amerika Serikat.
Hama ini ditemukan di bagian selatan Amerika dan bermigrasi ke California
Utara, dan akhirnya ditemukan pula di bagian barat Amerika dan Canada.
Selanjutnya hama ini menyebar ke pulau Caribia, Amerika Serikat dan Amerika
Selatan serta Mexico, kemudian menyebar ke Eropa, Afrika, India, Australia (Mc
Auslane 2005).
Sebelum tahun 1986, hama B. tabaci hanya bersifat sebagai hama sekunder.
Pada tahun 1986 di Florida, serangga ini menjadi serangga yang merugikan secara
ekonomi. Hama ini berkembang dengan cepat di Amerika Serikat (Texas, Arizona
dan California). Pada tahun 1994, ditemukan spesies baru, yaitu B. argentifolii
(Mc Auslane 2005).
Peranan B. tabaci dalam Penularan Begomovirus
Peranan B. tabaci sebagai vektor Begomovirus yang menyebabkan penyakit
pada tanaman sayuran termasuk cabai merah sudah banyak dilaporkan. Menurut
Cohen dan Berlinger (1986) populasi kutukebul yang sangat rendah sudah dapat
menyebabkan kerusakan tanaman, karena merupakan vektor virus tanaman. Pada
umumnya hubungan virus dengan vektornya bersifat persisten akan tetapi pada
umumnya tidak diturunkan ke generasi berikutnya melalui telur (non transovarial
transmission), walaupun diketahui terdapat begomovirus yang dapat diturunkan
ke generasi berikutnya, misalnya TYLCV (Czosnek et al. 1988), dan TYLCV-sar
(Bosco et al. 2001). Ghanim et al. (1998) dan Ghanim dan Czosnek (2000)
melaporkan bahwa TYLCV-Israel mampu ditularkan secara transovarial oleh B.
tabaci selama dua generasi dan melalui kopulasi antar individu.
Perring (2001) menyatakan bahwa terdapat 7 kelompok biotipe B. tabaci,
dan biotipe B yang sangat potensial dalam menularkan begomovirus pada
berbagai tanaman budidaya, misal CabLCV pada tanaman kubis dan ToMV pada
melalui benih tapi hanya menular dengan bantuan serangga B. tabaci dari tanaman
satu ke tanaman lainnya.
Sulandari (2004) melaporkan bahwa B. tabaci adalah suatu vektor yang
sangat efektif, karena pada percobaan di rumah kaca hanya dengan menggunakan
satu imago B. tabaci setelah mengisap sumber inokulum selama 48 jam dan
melalui periode inokulasi 24 jam sudah dapat menularkan begomovirus penyebab
penyakit daun keriting kuning cabai (PepYLCV) sebesar 40%. dan efektifitas
penularan meningkat 2 kali apabila digunakan 3 imago. Semakin banyak imago B.
tabaci yang digunakan, efektivitas penularan makin meningkat dan masa
inkubasinya lebih singkat. Brown (1994) menunjukkan bahwa satu ekor B. tabaci
yang telah diberi perlakuan periode makan akuisisi (PMA) selama 48 jam dan
periode makan inokulasi (PMI) selama 3 hari, mampu menularkan chino del
tomato virus (CdTV) dengan jumlah tanaman terinfeksi 15%. Mehta et al. (1994)
melaporkan bahwa satu ekor B. tabaci biotipe B mampu menularkan tomato
yellow leaf curl virus-Mesir (TYLCV-Mesir), dan efisiensi penularannya
meningkat 4 kali jika jumlah serangga ditingkatkan hingga 5 ekor pertanaman.
Uzcategui dan Lastra (1978) melaporkan bahwa efisiensi penularan virus oleh B.
tabaci yang dipelihara pada suhu 30 – 34oC adalah 93%, sedangkan yang
dipelihara pada suhu 20-30o
Imago B. tabaci betina lebih efektif dalam menularkan PepYLCV
dibandingkan yang jantan. Semua tanaman (100%) tertular oleh PepYLCV yang
dibawa imago betina dengan masa inkubasi yang lebih pendek (6 sampai 8 hari)
sementara yang jantan hanya mampu menularkan sebanyak 60% dengan masa C hanya 75%.
Hubungan PepYLCV dengan B. tabaci bersifat persisten akan tetapi pada
umumnya tidak diturunkan ke generasi berikutnya melalui telur (non transovarial
transmission). Periode akuisisi dan inokulasi yang optimal dari B. tabaci untuk
menularkan Pep.YLCV adalah 3 sampai 6 jam, memerlukan periode laten di
dalam tubuh vektor selama minimal 9 jam dan periode retensinya sampai serangga
mati (Sulandari 2004). Untuk PepYLCV di Thailan, periode makan akuisisi imago
B. tabaci selama 1 jam merupakan waktu yang optimal untuk penularannya, dan
15
inkubasi yang lebih panjang (10 sampai 18 hari), dan gejala yang ditimbulkannya
juga lebih ringan, tanaman cabai tidak menjadi kerdil (Sulandari 2004).
Peranan Teknik Budidaya Khususnya Pemanfaatan Tanaman Pembatas Pinggir Untuk Pengendalian Hama dan Konservasi Predator
Pengendalian hama secara budidaya atau bercocok tanam bertujuan
mengelola lingkungan tanaman sedemikian rupa sehingga menjadi kurang cocok
bagi kehidupan dan perkembangan hama serta dapat mengurangi laju peningkatan
populasi hama dan kerusakan tanaman. Disamping itu, diusahakan juga agar
lingkungan dapat mendorong berfungsinya musuh alami sebagai pengendali hama
yang efektif (Untung 2006).
Mula mula teknik pengendalian secara bercocok tanam yang diterapkan
merupakan teknik yang kurang dilihat perpaduannya dengan teknik-teknik
pengendalian hama yang lain seperti pemanfaatan musuh alami. Saat ini dalam
rangka sistem PHT teknik pengendalian hama secara bercocok tanam mencakup
pengertian yang lebih luas yaitu pengelolaan ekologi lingkungan pertanaman
(Pedigo 1991). Untuk meningkatkan keefektifan dan efisiensi hasil pengendalian
teknik pengendalian secara bercocok tanam perlu dipadukan dengan teknik-teknik
pengendalian hama lainnya sesuai dengan prinsip-prinsip PHT (Untung 2006).
Menurut Pedigo (1991) sebagian besar teknik pengendalian secara bercocok
tanam berdasarkan sasaran yang akan dicapai dapat dikelompokkan menjadi 4 dan
salah satunya adalah mengalihkan populasi hama menjauhi tanaman. Dengan
mempelajari sifat preferensi dan kemampuan serangga hama bergerak menjauhi
tanaman yang tidak disukai dan menuju ke jenis tanaman yang disenangi dapat
diusahakan mengalihkan perhatian hama untuk menjauhi tanaman yang sedang
diusahakan.
Penanaman tanaman border di sekitar tanaman yang diusahakan, berfungsi
menolak atau menghalangi agar hama menjauhi atau tidak sampai ke tanaman
yang diusahakan. Penanaman tanaman border dilakukan dengan menanam jenis
tanaman bukan inang atau yang tidak disukai oleh hama sasaran di sekeliling
tanaman yang diusahakan. Tanaman bukan inang selain berperan sebagai samaran
yang membuat tanaman inang sulit ditemukan, juga berperan sebagai penghalang
jagung di sekeliling tanaman cabai dapat menghalangi pemencaran trips (Pedigo
1991). Agar diperoleh hasil dengan baik, waktu penanaman tanaman border harus
disesuaikan dengan fenologi hama terutama waktu pemunculan fase hidup hama
yang merusak tanaman. Disamping itu perlu dipertimbangkan juga postur tinggi
tanaman border harus lebih tinggi dari tanaman utama dan kemampuan ketinggian
terbang hama sasaran (Untung 2006).
Menurut Settle et al. (1996) dan Untung (2006), pemanfaatan tanaman
border dapat mendorong stabilitas ekosistem sehingga populasi hama dapat
ditekan berada dalam kesetimbangannya. Fenomena ini terjadi karena suatu lahan
pertanian yang ditanamai berbagai jenis tanaman dapat meningkatkan keragaman
vegetasi yang terdapat di dalam ekosistem, sehingga membuat ekosistem tersebut
kurang sesuai bagi serangga hama, dan sebaliknya sesuai bagi musuh alami.
Beberapa hasil penelitian menunjukkan bahwa populasi serangga hama ditemukan
lebih tinggi pada pertanaman monokultur dari pada pertanaman polikultur.
Tingginya populasi serangga hama pada pertanaman dengan vegetasi yang lebih
seragam disebabkan oleh laju kolonisasi serangga hama lebih tinggi dan hambatan
fisik (barier) serta kimia yang lebih rendah dalam menemukan tanaman inang
(Altieri 1991).
Di sisi lain, teknik bercocok tanam seperti penanaman tanaman border dapat
mendorong konservasi musuh alami seperti predator. Hal ini terjadi karena
pemanfaatan tanaman border merupakan teknik pengendalian hama secara
budidaya yang dapat meningkatkan keragaman vegetasi (Untung 2006). Vegetasi
yang beragam sangat sesuai bagi kehidupan musuh alami seperti predator yang
pada umumnya bersifat generalis. Predator generalis dapat memangsa
bermacam-macam mangsa yang tersedia dalam waktu yang lama. Selain itu predator juga
dapat memelihara reproduksinya dan meningkatkan lama hidup, karena
tersedianya nektar dan tepung sari serta mikrohabitat yang sesuai (Andow 1991;
Perfecto dan Sediles 1992; Untung 2006). Dengan demikian, pada vegetasi yang
beragam populasi predator generalis itu dapat bertahan dalam tingkat yang lebih
tinggi dan dalam waktu yang relatif lama, sehingga predator generalis lebih kuat
menekan populasi hama (Risch et al. 1983). Norris dan Kogan (2005)
17
dan parasitoid terutama yang generalis pada vegetasi yang beragam relatif stabil,
dan bertahan lama, sebab makanan (tepung sari dan nektar) tersedia lebih
berkesinambungan serta adanya tempat berlindung dan mikrohabitat yang sesuai.
Peranan Predator dalam Pengendalian B. tabaci
Menurut Pedigo (1991) predator adalah organisasi yang hidup bebas dan
memakan binatang lain, memangsa kadang-kadang secara utuh dan cepat. Istilah
“predatisme” adalah suatu bentuk simbiosis dari dua individu yang salah satu di
antara individu tersebut menyerang atau memakan individu lainnya satu atau lebih
spesies, untuk kepentingan hidupnya yang dapat dilakukan dengan
berulang-ulang. Individu yang diserang disebut mangsa. Menurut Smith (1978) dan
Huffaker et al. (1976) dalam pemanfaatan predator, bisa dilakukan dengan
mengintroduksi maupun yang sudah ada di suatu daerah kemudian dikelola untuk
mengendalikan serangga hama.
Menurut De Bach (1979) dan Untung (2006), populasi hama yang
meningkat sehingga menimbulkan kerugian secara ekonomik disebabkan oleh
keadaan lingkungan yang kurang memberi kesempatan bagi predator untuk
menjalankan fungsi alaminya. Apabila kepada musuh alami dalam hal ini predator
kita berikan kesempatan berfungsi, antara lain dengan jalan rekayasa lingkungan
seperti introduksi predator, memperbanyak dan melepaskannya, serta mengurangi
berbagai dampak negatif terhadap predator, maka predator akan dapat
melaksanakan fungsinya dengan baik
Serangga predator pada umumnya bersifat polifag atau oligofag. Coppel and
Mertins (1977) menyatakan bahwa spesies predator yang polifag kurang
tergantung pada kerapatan populasi serangga hama. Walaupun demikian predator
yang polifag memiliki kemampuan yang tinggi dalam beradaptasi terhadap
lingkungan, dan memiliki kemampuan memencar yang cepat serta mampu beralih
mangsa apabila mangsa utama berkurang. Wiedenmann and Smith (1997)
menambahkan bahwa predator yang potensial harus mempunyai kemampuan
berkompetisi dan menyabar yang tinggi. Pada tanaman semusim, kolonisasi
predator dapat terjadi secara periodik. Pada saat di lapangan tidak ada tanaman
saat di lapang ada tanaman maka predator harus mampu dengan cepat dapat
mengolonisasi habitat itu.
Studi musuh alami Bemisia spp. di Brasil menemukan sekitar 14 spesies
predator (Jervis dan Kidd 1996). Dari jenis predator kumbang kubah (lady beetle),
Nephaspis hydra Gordon dan Delphastus davidsoni Gordon diketahui pertama
kali sebagai pemangsa Bemisia spp. Gerling (1990) menambahkan bahwa ada 19
spesies serangga tercatat sebagai predator yang memangsa B. tabaci yang
termasuk pada empat famili yaitu Chrysopidae, Miridae, Anthocoridae dan
Stigmaeidae. Setiawati (2005), mengungkapkan beberapa spesies predator yang
diketahui efektif terhadap B. tabaci antara lain Coccinella transversalis,
Menochilus sexmaculatus, Coenosia attenuate, Delphastus pusillus, Deracocoris
pallens, Euscius hibisci, Orius albidipennis, Scymus syriacus. dan Chrysoperla
carnea.
Dari hasil penelitian eksplorasi musuh alami di Kabupaten Bandung dan
Karawang pada tahun 2005 ditemukan beberapa jenis predator dan parasitoid
yang mempunyai potensi untuk mengendalikan hama kutukebul (B. tabaci) pada
pertanaman sayuran. Jenis predator kutukebul yang ditemukan adalah beberapa
spesies dari famili Coccinellidae (Menochilus sexmaculatus, Coccinella
transversalis, Harmonia sp., Curinus coerulus dan Delphastus sp.), Paederus
fusipes (Coleoptera: Stapilinidae) dan Condilastylus sp. (Diptera). Jenis parasitoid
kutukebul yang ditemukan adalah beberapa spesies dari famili Aphelenidae
(seperti Eritmoserus sp., Neopompalae sp. dan Encarsia sp.). (Sudrajat 2009).
Berdasarkan hasil pengujian skala laboratorium jenis predator yang potensial
untuk dikembangkan adalah M. sexmaculatus, C. transfersalis, Harmonia sp., C.
coerulus dan Paederus fuscipes, dengan daya pemangsaan rata-rata berkisar 65%.
(Muharam dan Setiawati 2007).
Keefektifan predator dalam mengendalikan populasi hama dapat diukur dari
daya pemangsaannya (Kharboutli dan Mack 1993). Berdasarkan daya pemangsaan
tersebut dapat dinilai kemampuan predator dalam mengatur keseimbangan
populasi mangsa. Menurut Tarumingkeng (1994), terdapat beberapa faktor yang
menentukan tingkat laju pemangsaan (konsumsi) oleh suatu predator terhadap
19
kerapatan mangsa hubungannya dengan kemampuan mencari yang tinggi
terutama pada saat populasi mangsa rendah (tanggap fungsional).
Preferensi Predator terhadap Jenis Mangsa
Berbagai spesies predator mempunyai preferensi terhadap mangsa tertentu.
Preferensi diartikan sebagai derajat kesukaan predator terhadap mangsa tertentu,
yang ditunjukkan oleh jumlah individu mangsa yang dimakan dan terbunuh oleh
suatu individu predator dalam satu satuan waktu. Menurut Cineros dan
Rosenheim (1998) preferensi dapat merupakan seleksi terhadap satu tipe mangsa
terhadap proporsi mangsa yang tersedia dalam satu lingkungan tertentu. Untuk
menilai preferensi tersebut dapat digunakan nilai indeks linier pemilihan mangsa
yang dapat diduga dengan persamaan yang dikembangkan oleh Houck (1986)
sebagai berikut : Li = ri - pi. Li adalah indeks linier pemilihan mangsa, ri adalah
proporsi mangsa yang dimangsa oleh predator, dan pi adalah proporsi mangsa
yang tersedia (Houck 1986). Secara simetris nilai indeks linier pemilihan mangsa
bervariasi antara -1 s/d + 1. Nilai positif menunjukan adanya preferensi
(kesukaan) terhadap mangsa tertentu dan nilai negatif menunjukan tidak ada
kesukaan (penolakan) terhadap mangsa tertentu atau mangsa relatif dihindari.
Beberapa komponen yang berpengaruh pada preferensi terhadap mangsa
adalah ketertarikan dan kesesuaian terhadap mangsa, pengenalan terhadap
mangsa, keputusan menyerang atau tidak, menangkap serta mengkonsumsi
mangsa. Walaupun tingkat penerimaan terhadap dua jenis mangsa sama, tetapi
karena kemampuan menghindar dari kedua mangsa tersebut berbeda maka
frekuensi pertemuan predator dan mangsa dapat berbeda, akibatnya preferensi
terhadap kedua mangsa tersebut berbeda (Cineros dan Rosenheim 1998).
Tanggap Fungsional Predator terhadap Mangsa
Hubungan antara predator dan mangsa sangat erat dalam kaitannya dengan
perubahan kerapatan mangsa. Perubahan kerapatan populasi mangsa dapat
mengubah perilaku predator dalam hal pemangsaan dan keefektifan predator
dicerminkan oleh intensitas tanggapnya terhadap kerapatan populasi mangsa
Elseth dan Baumgardner (1981) memperkenalkan dua jenis tanggap
predator terhadap perubahan kerapatan populasi mangsa yaitu tanggap fungsional
dan tanggap numerik. Tanggap fungsional menunjukkan perubahan jumlah
mangsa yang diserang atau dimakan (laju pemangsaan) per individu predator
akibat perubahan kerapatan mangsa per satuan waktu (Hassel 1966). Tanggap
numerik menunjukkan perubahan kerapatan predator sebagai akibat dari
perubahan kerapatan mangsa. Peningkatan kerapatan populasi predator sebagai
tanggap terhadap peningkatan kerapatan populasi mangsa. Peningkatan kerapatan
mangsa dan predator disebabkan oleh imigrasi atau reproduksi yang meningkat
atau keduanya (Brewer 1979).
Tanggap fungsional kemudian menjadi salah satu ukuran untuk menentukan
keefektivan suatu predator dalam mengendalikan hama atau mengatur
keseimbangan populasi hama (Hassell et al. 1977). Tanggap fungsional
merupakan aspek penting untuk dipelajari, sehingga diperoleh gambaran tentang
kemampuan predator dalam menangani mangsanya (Pervez dan Omkar 2005).
Holling (1959) mengidentifikasi tiga macam tipe tanggap fungsional yang
menggambarkan hubungan predator-mangsa yaitu; (1) Tanggap fungsional tipe I.
(linier) yaitu proporsi mangsa yang dimakan bersifat konstan, sehingga hubungan
antara banyaknya mangsa yang dimakan predator dan kerapatan mangsa bersifat
linier. Jumlah mangsa yang dimakan meningkat secara proporsional mengikuti
peningkatan kerapatan mangsa sampai batas maksimum kemudian mendatar
sejalan dengan peningkatan kerapatan mangsa; (2) Tanggap fungsional Tipe II.
(hiperbolik) yaitu proporsi mangsa yang dimakan/terbunuh menurun tajam dengan
meningkatnya kerapatan populasi mangsa. Jumlah mangsa yang dimakan makin
banyak dengan makin meningkatnya populasi mangsa, kemudian pertambahan
jumlah mangsa yang dimakan tersebut mulai menurun dan penurunan itu makin
cepat sejalan dengan makin meningkatnya populasi mangsa; (3) Tanggap
fungsional Tipe III. (sigmoid) yaitu proporsi mangsa yang dimakan/terbunuh
awalnya meningkat, tetapi kemudian secara berangsur menurun dengan
meningkatnya kerapatan populasi mangsa. Pertambahan jumlah mangsa yang
21
mangsa, kemudian makin cepat dan lambat kembali dengan meningkatnya
populasi mangsa.
Untuk memperoleh parameter tanggap fungsional laju pencarian mangsa
seketika (a) dan waktu penanganan mangsa (Th) digunakan model persamaan
cakram dari Holling (1959) atau persamaan acak dari Rogers (1972). Untuk
tanggap fungsioan tipe I adalah: Persamaan cakram Ne = aN0 + b; dan
Persaman acak Ne = aT.No. Untuk tanggap fungsional tipe II adalah:
Persamaan cakram Ne = a.T.N0/(1+a.Th.N0); Persamaan acak Ne = N0
{1–exp[a(Th.Ne–T)]}. Untuk tanggap fungsional tipe III adalah: Persamaan
cakram Ne = a.T.N02/(1+cN0+b.Th.N02); Persamaan acak Ne = N0 {1–exp
[(d+bN0)(Th.Ne–T)/(1+cN0)]}. Ne adalah banyaknya mangsa yang dimakan oleh
per individu predator, N0 banyaknya mangsa yang tersedia, T lama waktu mangsa
terpapar pada predator waktu pencarian mangsa, Th waktu penanganan mangsa, a
laju pencarian seketika(penyerangan) dan b, c dan d adalah konstan yang
diturunkan dari parameter a (Hassell et al. 1977). Persamaan cakram Holling
digunakan bila selama percobaan berlangsung, mangsa yang dikonsumsi oleh
predator diganti sehingga kepadatan mangsa konstan. Jika mangsa yang
dikonsumsi tidak diganti, maka persamaan acak lebih cocok untuk digunakan
(Rogers 1972).
Tanggap fungsional dapat dipengaruhi oleh beberapa faktor seperti stadia
mangsa, fase pertumbuhan tanaman, cuaca, kehadiran mangsa alternatif dan
kompetisi antara predator, disamping itu keragaman fisik dari suatu habitat juga
mempengaruhi tanggap fungsional predator terhadap mangsa. Hal ini ada kaitanya
dengan habitat sebagai tempat berlindung dan sumber makanan alternatif bagi
serangga predator (Horn 1981).
Daftar Pustaka
Altieri MA. 1991. Ecology of tropical herbivores in polycultural agroecosystem. P.607 – 617. In Price PC, T.M. Lewinsohn, G.W. Fernandes, W. Benson (ed.) Plant-Animals interaction: evolutionary ecology in tropical and temperate regions. John Wiley dan Sons, Inc New York 545p.
Andow D.A. 1991. Vegetational diversity and arthropod population response.
Badri I. 1983. Identification of the Aleyrodid on soybean from two location in west Jafa and some bionomics of Bemisia tabaci Genn. (Homoptera : Aleyrodidae) on three soybean varieties. Biotrop, Seameo-Regional Center for tropical Biology. Bogor, Indonesia. 62 pp.
Basu AN. 1995. Bemisia tabaci (Gennadius): Crop Pest and Principal Whitefly Vector 0f Plant Viruses. New Delhi: Westview Press
Beck, S.D. 1965. Resist