UJI KETAHANAN TERHADAP PENYAKIT
BLAS DAUN (
Pyricularia oryzae
Cav.)
GALUR-GALUR PADI GOGO (
Oryza sativa
L.) F4:6
BERDAYA HASIL TINGGI
Oleh
Lestari Admojo
A 34401039
PROGRAM STUDI PEMULIAAN TANAMAN DAN
TEKNOLOGI BENIH
FAKULTAS PERTANIAN
INSTITUT PERTANIAN BOGOR
RINGKASAN
LESTARI ADMOJO. UJI KETAHANAN TERHADAP PENYAKIT BLAS DAUN (Pyricularia oryzae Cav.) GALUR-GALUR PADI GOGO (Oryza sativa L.) F4:6 BERDAYA HASIL TINGGI. (Dibimbing oleh DESTA WIRNAS dan TRIKOESOEMANINGTYAS).
Penelitian ini bertujuan untuk mengevaluasi galur-galur padi gogo F4:6 yang berdaya hasil tinggi untuk ketahanan terhadap blas daun dalam upaya pengembangan varietas unggul baru padi gogo. Penelitian ini dilaksanakan di Balai Besar Bioteknologi dan Sumber Daya Genetik (BB BIOGEN) Cimanggu, Bogor dari bulan Juni hingga Agustus 2005.
Bahan tanaman yang digunakan dalam penelitian ini adalah 41 galur padi gogo F4:6 yang berdaya hasil tinggi terseleksi di Cikeumeuh, Bogor, tujuh tetua yaitu Krowal, B8503E-TB-9-0-3, CT6510-24-1-3, Situgintung, Jatiluhur, IR 64 dan Gajah Mungkur, dan dua kontrol yaitu Asahan sebagai kontrol tahan dan Kencana Bali sebagai kontrol peka. Isolat yang digunakan adalah ras 033 yang mewakili ras dengan skala wilayah serangan yang luas tapi tidak begitu virulen, dan ras 173 yang mewakili ras dengan skala wilayah serangan yang sempit tapi sangat virulen.
Metode dalam penelitian ini menggunakan rancangan petak terbagi dengan dua faktor, dua ras blas yaitu ras 033 dan ras 173 digunakan sebagai petak utama dan 50 tanaman padi gogo sebagai anak petak. Data dianalisis menggunakan analisis ragam dari rancangan petak terbagi dan uji lanjut untuk data yang berbeda nyata menggunakan uji Dunnett pada taraf 5%. Data juga dihitung untuk mendapatkan nilai heritabilitas arti luas dan uji korelasi.
UJI KETAHANAN TERHADAP PENYAKIT
BLAS DAUN (
Pyricularia oryzae
Cav.)
GALUR-GALUR PADI GOGO (
Oryza sativa
L.) F4:6
BERDAYA HASIL TINGGI
Skripsi
Sebagai salah satu syarat untuk memperoleh gelar
Sarjana Pertanian pada Fakultas Pertanian
Institut Pertanian Bogor
Oleh
Lestari Admojo
A 34401039
PROGRAM STUDI PEMULIAAN TANAMAN DAN
TEKNOLOGI BENIH
FAKULTAS PERTANIAN
INSTITUT PERTANIAN BOGOR
Judul : UJI KETAHANAN TERHADAP PENYAKIT BLAS DAUN (Pyricularia oryzae Cav.) GALUR-GALUR PADI GOGO (Oryza sativa L.) F4:6 BERDAYA HASIL TINGGI
Nama : LESTARI ADMOJO
NRP : A 34401039
Menyetujui,
Pembimbing I Pembimbing II
Desta Wirnas, SP, MSi Dr. Ir. Trikoesoemaningtyas, MSc
NIP. 132 259 275 NIP. 132 169 917
Mengetahui, Dekan Fakultas Pertanian
Prof. Dr. Ir. Supiandi Sabiham, M.Agr
NIP. 130 422 698
RIWAYAT HIDUP
Penulis dilahirkan di Salatiga pada tanggal 1 Januari 1983 dari orang tua bernama Poniman Hadi Pramono (Alm) dan Siti Maryam. Penulis merupakan anak keempat dari enam bersaudara.
KATA PENGANTAR
Puji dan syukur penulis panjatkan kehadirat Allah SWT yang telah melimpahkan rahmat dan karuniaNya sehingga penulis dapat menyelesaikan skripsi ini.
Skripsi yang berjudul Uji Ketahanan Terhadap Penyakit Blas Daun (Pyricularia oryzae Cav.) Galur- galur Padi Gogo (Oryza sativa L.) F4:6 Berdaya Hasil Tinggi ini merupakan salah satu syarat untuk meraih gelar Sarjana Pertanian, Institut Pertanian Bogor.
Pada kesempatan ini penulis ingin menyampaikan terima kasih kepada :
Ø Desta Wirnas, SP, MSi dan Dr. Ir. Trikoesoemaningtyas, MSc selaku dosen pembimbing yang telah banyak meluangkan waktu, tenaga, dan pikirannya, mengarahkan, serta membimbing penulis dalam menyelesaikan skripsi ini.
Ø Willi Bayuardi, SP, MSi selaku dosen penguji, atas saran dan masukannya yang sangat berarti bagi penulis dalam perbaikan skripsi.
Ø Dr. Ir. Sriani Sujiprihati, MSc selaku konselor TPB, dan Endang Sjamsudin, Ir, MAgr. Sc selaku Pembimbing Akademik atas saran dan bimbingannya selama penulis menyelesaikan perkuliahan.
Ø Ibu Anggiani Nasution, Pak Ade, dan Pak Moko atas bantuan dan sarannya selama penelitian berlangsung.
Ø Pak Syukur, Pak Palguno, Muhtar, dan Chotim atas bantuan dan sarannya dalam pengolahan data selama penulisan skripsi.
Ø Bapak (Alm), Mamak, Mbak Atun, Mas Ndoko, Mas Tiok, Pam, Sapto, Dik Andre, Mbak Yayak, Mbak Husni, dan seluruh keluarga besar trah Karto Prawiro atas doa, perhatian, dukungan dan cintanya, sehingga perkuliahan ini bisa terselesaikan dengan baik.
Ø Keluarga besar PMTB ’38, Chotim, Aya, Noor, Pipit, Uswah, Indah, Ewi, Uci, Salha, dan rekan seperjuanganku selama penelitian, Ida atas kebersama annya selama ini.
Ø Saudari-saudariku di LISMA ’38, Ida, Aya, Uswah, Maya, Ricat, Yanti, Hikmah, Ritan, Indah, Nurlah, Iin, Rahma, Siska, Ernisa, Iha dan masih banyak lagi, atas dorongan semangat yang tiada henti.
Ø Seluruh pihak yang telah banyak membantu dalam penyelesaian skripsi ini yang tidak bisa penulis sebutkan satu persatu. Semoga Allah memberikan balasan yang terbaik. Amin.
Akhir kata semoga skripsi ini bermanfaat khususnya bagi penulis dan juga bagi semua pihak yang memerlukan.
Bogor, Januari 2006
DAFTAR ISI
Halaman
KATA PENGANTAR ... i
DAFTAR ISI ... iii
DAFTAR TABEL... v
DAFTAR GAMBAR ... vii
PENDAHULUAN... 1
Latar Belakang ... 1
Tujuan... 3
Hipotesis ... 3
TINJAUAN PUSTAKA ... 4
Klasifikasi, Morfologi, dan Daur Hidup Pyricularia oryzae Cav... 4
Cara Penyerangan dan Gejala Penyakit Blas ... 5
Keragaman Patogen dari Ras Pyricularia oryzae Cav... 6
Mekanisme Ketahanan Tanaman Padi Terhadap Blas ... 7
Metode Pengujian Terhadap Penyakit Blas ... 12
BAHAN DAN METODE ... 15
Waktu dan Tempat Penelitian ... 15
Bahan dan Alat ... 15
Metode Penelitian... 15
Pelaksanaan Penelitian ... 16
Pengamatan ... 17
Analisis Data ... 19
HASIL DAN PEMBAHASAN ... 21
Kondisi Umum ... 21
Periode Laten... 23
Jumlah Daun Terserang... 24
Jumlah Bercak Tidak Berspora ... 25
Jumlah Bercak Berspora... 27
Intensitas Serangan... 31
Heritabilitas Arti Luas... 33
KESIMPULAN DAN SARAN... 35
Kesimpulan... 35
Saran... 35
DAFTAR PUSTAKA... 36
UJI KETAHANAN TERHADAP PENYAKIT
BLAS DAUN (
Pyricularia oryzae
Cav.)
GALUR-GALUR PADI GOGO (
Oryza sativa
L.) F4:6
BERDAYA HASIL TINGGI
Oleh
Lestari Admojo
A 34401039
PROGRAM STUDI PEMULIAAN TANAMAN DAN
TEKNOLOGI BENIH
FAKULTAS PERTANIAN
INSTITUT PERTANIAN BOGOR
RINGKASAN
LESTARI ADMOJO. UJI KETAHANAN TERHADAP PENYAKIT BLAS DAUN (Pyricularia oryzae Cav.) GALUR-GALUR PADI GOGO (Oryza sativa L.) F4:6 BERDAYA HASIL TINGGI. (Dibimbing oleh DESTA WIRNAS dan TRIKOESOEMANINGTYAS).
Penelitian ini bertujuan untuk mengevaluasi galur-galur padi gogo F4:6 yang berdaya hasil tinggi untuk ketahanan terhadap blas daun dalam upaya pengembangan varietas unggul baru padi gogo. Penelitian ini dilaksanakan di Balai Besar Bioteknologi dan Sumber Daya Genetik (BB BIOGEN) Cimanggu, Bogor dari bulan Juni hingga Agustus 2005.
Bahan tanaman yang digunakan dalam penelitian ini adalah 41 galur padi gogo F4:6 yang berdaya hasil tinggi terseleksi di Cikeumeuh, Bogor, tujuh tetua yaitu Krowal, B8503E-TB-9-0-3, CT6510-24-1-3, Situgintung, Jatiluhur, IR 64 dan Gajah Mungkur, dan dua kontrol yaitu Asahan sebagai kontrol tahan dan Kencana Bali sebagai kontrol peka. Isolat yang digunakan adalah ras 033 yang mewakili ras dengan skala wilayah serangan yang luas tapi tidak begitu virulen, dan ras 173 yang mewakili ras dengan skala wilayah serangan yang sempit tapi sangat virulen.
Metode dalam penelitian ini menggunakan rancangan petak terbagi dengan dua faktor, dua ras blas yaitu ras 033 dan ras 173 digunakan sebagai petak utama dan 50 tanaman padi gogo sebagai anak petak. Data dianalisis menggunakan analisis ragam dari rancangan petak terbagi dan uji lanjut untuk data yang berbeda nyata menggunakan uji Dunnett pada taraf 5%. Data juga dihitung untuk mendapatkan nilai heritabilitas arti luas dan uji korelasi.
UJI KETAHANAN TERHADAP PENYAKIT
BLAS DAUN (
Pyricularia oryzae
Cav.)
GALUR-GALUR PADI GOGO (
Oryza sativa
L.) F4:6
BERDAYA HASIL TINGGI
Skripsi
Sebagai salah satu syarat untuk memperoleh gelar
Sarjana Pertanian pada Fakultas Pertanian
Institut Pertanian Bogor
Oleh
Lestari Admojo
A 34401039
PROGRAM STUDI PEMULIAAN TANAMAN DAN
TEKNOLOGI BENIH
FAKULTAS PERTANIAN
INSTITUT PERTANIAN BOGOR
Judul : UJI KETAHANAN TERHADAP PENYAKIT BLAS DAUN (Pyricularia oryzae Cav.) GALUR-GALUR PADI GOGO (Oryza sativa L.) F4:6 BERDAYA HASIL TINGGI
Nama : LESTARI ADMOJO
NRP : A 34401039
Menyetujui,
Pembimbing I Pembimbing II
Desta Wirnas, SP, MSi Dr. Ir. Trikoesoemaningtyas, MSc
NIP. 132 259 275 NIP. 132 169 917
Mengetahui, Dekan Fakultas Pertanian
Prof. Dr. Ir. Supiandi Sabiham, M.Agr
NIP. 130 422 698
RIWAYAT HIDUP
Penulis dilahirkan di Salatiga pada tanggal 1 Januari 1983 dari orang tua bernama Poniman Hadi Pramono (Alm) dan Siti Maryam. Penulis merupakan anak keempat dari enam bersaudara.
KATA PENGANTAR
Puji dan syukur penulis panjatkan kehadirat Allah SWT yang telah melimpahkan rahmat dan karuniaNya sehingga penulis dapat menyelesaikan skripsi ini.
Skripsi yang berjudul Uji Ketahanan Terhadap Penyakit Blas Daun (Pyricularia oryzae Cav.) Galur- galur Padi Gogo (Oryza sativa L.) F4:6 Berdaya Hasil Tinggi ini merupakan salah satu syarat untuk meraih gelar Sarjana Pertanian, Institut Pertanian Bogor.
Pada kesempatan ini penulis ingin menyampaikan terima kasih kepada :
Ø Desta Wirnas, SP, MSi dan Dr. Ir. Trikoesoemaningtyas, MSc selaku dosen pembimbing yang telah banyak meluangkan waktu, tenaga, dan pikirannya, mengarahkan, serta membimbing penulis dalam menyelesaikan skripsi ini.
Ø Willi Bayuardi, SP, MSi selaku dosen penguji, atas saran dan masukannya yang sangat berarti bagi penulis dalam perbaikan skripsi.
Ø Dr. Ir. Sriani Sujiprihati, MSc selaku konselor TPB, dan Endang Sjamsudin, Ir, MAgr. Sc selaku Pembimbing Akademik atas saran dan bimbingannya selama penulis menyelesaikan perkuliahan.
Ø Ibu Anggiani Nasution, Pak Ade, dan Pak Moko atas bantuan dan sarannya selama penelitian berlangsung.
Ø Pak Syukur, Pak Palguno, Muhtar, dan Chotim atas bantuan dan sarannya dalam pengolahan data selama penulisan skripsi.
Ø Bapak (Alm), Mamak, Mbak Atun, Mas Ndoko, Mas Tiok, Pam, Sapto, Dik Andre, Mbak Yayak, Mbak Husni, dan seluruh keluarga besar trah Karto Prawiro atas doa, perhatian, dukungan dan cintanya, sehingga perkuliahan ini bisa terselesaikan dengan baik.
Ø Keluarga besar PMTB ’38, Chotim, Aya, Noor, Pipit, Uswah, Indah, Ewi, Uci, Salha, dan rekan seperjuanganku selama penelitian, Ida atas kebersama annya selama ini.
Ø Saudari-saudariku di LISMA ’38, Ida, Aya, Uswah, Maya, Ricat, Yanti, Hikmah, Ritan, Indah, Nurlah, Iin, Rahma, Siska, Ernisa, Iha dan masih banyak lagi, atas dorongan semangat yang tiada henti.
Ø Seluruh pihak yang telah banyak membantu dalam penyelesaian skripsi ini yang tidak bisa penulis sebutkan satu persatu. Semoga Allah memberikan balasan yang terbaik. Amin.
Akhir kata semoga skripsi ini bermanfaat khususnya bagi penulis dan juga bagi semua pihak yang memerlukan.
Bogor, Januari 2006
DAFTAR ISI
Halaman
KATA PENGANTAR ... i
DAFTAR ISI ... iii
DAFTAR TABEL... v
DAFTAR GAMBAR ... vii
PENDAHULUAN... 1
Latar Belakang ... 1
Tujuan... 3
Hipotesis ... 3
TINJAUAN PUSTAKA ... 4
Klasifikasi, Morfologi, dan Daur Hidup Pyricularia oryzae Cav... 4
Cara Penyerangan dan Gejala Penyakit Blas ... 5
Keragaman Patogen dari Ras Pyricularia oryzae Cav... 6
Mekanisme Ketahanan Tanaman Padi Terhadap Blas ... 7
Metode Pengujian Terhadap Penyakit Blas ... 12
BAHAN DAN METODE ... 15
Waktu dan Tempat Penelitian ... 15
Bahan dan Alat ... 15
Metode Penelitian... 15
Pelaksanaan Penelitian ... 16
Pengamatan ... 17
Analisis Data ... 19
HASIL DAN PEMBAHASAN ... 21
Kondisi Umum ... 21
Periode Laten... 23
Jumlah Daun Terserang... 24
Jumlah Bercak Tidak Berspora ... 25
Jumlah Bercak Berspora... 27
Intensitas Serangan... 31
Heritabilitas Arti Luas... 33
KESIMPULAN DAN SARAN... 35
Kesimpulan... 35
Saran... 35
DAFTAR PUSTAKA... 36
DAFTAR TABEL
Nomor Teks Halaman
1. Penentuan Ras P. oryzae dan Distribusi Patogenisitas Berdasarkan
Reaksinya Terhadap Tujuh Varietas Diferensial di Indonesia... 7
2. Klasifikasi serangan P. oryzae berdasarkan IRRI (1996... 18
3. Sidik Ragam dari Rancangan Petak Terbagi ... 19
4. Rekapitulasi Sidik Ragam Peubah yang diamati pada 9 HSI... 24
5. Rata-rata Hasil Peubah Galur-galur Padi Gogo F4:6 yang diamati pada 9 HSI untuk ras 033 ... 28
6. Rata-rata Hasil Peubah Galur-galur Padi Gogo F4:6 yang diamati pada 9 HSI untuk ras 173 ... 29
7. Tingkat Ketahanan Galur-galur Padi Gogo F4:6 terhadap Penyakit Blas Daun pada 9 HSI ... 30
8. Hasil Uji Korelasi Terhadap Intensitas Serangan pada 9 HSI ... 33
9. Nilai Haritabilitas Arti Luas Peubah yang diamati pada 9 HSI ... 34
Lampiran 1. Rata-rata Jumlah Daun Terserang Ras 033 ... 40
2. Rata-rata Jumlah Daun Terserang Ras 173 ... 41
3. Rata-rata Jumlah Bercak Tidak Berspora... 42
4. Rata-rata Jumlah Bercak Berspora ... 43
5. Rata-rata Intensitas Serangan Ras 033 ... 44
6. Rata-rata Intensitas Serangan Ras 173 ... 45
7. Tingkat Ketahanan Galur-galur Padi Gogo F4:6 Terhadap Ras 033 .... 46
8. Tingkat Ketahanan Galur-galur Padi Gogo F4:6 Terhadap Ras 173 .... 47
9. Nilai Heritabilitas Arti Luas Seluruh Peubah Padi Gogo F4:6 yang diamati ... 48
10. Sidik Ragam Jumlah Daun Terserang pada 3 HSI ... 49
11. Sidik Ragam Jumlah Daun Terserang pada 6 HSI ... 49
12. Sidik Ragam Jumlah Daun Terserang pada 9 HSI ... 49
DAFTAR GAMBAR
Nomor Halaman
PENDAHULUAN
Latar Belakang
Padi merupakan tanaman pangan utama yang kebutuhannya selalu meningkat di Indonesia seiring pertambahan penduduk. Biro Pusat Statistik memperkirakan akan terjadi penurunan produksi beras nasional karena meningkatnya jumlah penduduk yang tidak diimbangi perluasan areal persawahan dan peningkatan produktivitas. Berdasarkan data BPS (2000) dalam Adiratna (2004), peningkatan penduduk mencapai 1.495 % dan terjadi penyusutan luas areal pertanian. Penyusutan tersebut terutama terjadi karena konversi lahan pertanian menjadi non pertanian. Menurut Irawan (2003), pada tahun 1983-1993 konversi lahan mencapai 1.28 juta ha dan sebesar 78 % terjadi di Pulau Jawa, dimana sebagian besar adalah lahan persawahan. Konversi lahan sawah pada tahun 1978-1998 menyebabkan hilangnya peluang produksi sebesar 8.83 juta ton, atau sekitar 31 % yang seharusnya bisa dicapai pada tahun 1998. Pasandaran (2003) juga menyatakan bahwa sejak awal 1980-an, Pulau Jawa sangat sulit untuk memperluas areal persawahan. Kecenderungan yang sama terjadi di luar Pulau Jawa pada tahun 1990-an. Menurut data BPS tahun 2001, luas panen padi mencapai 11.79 juta ha pada tahun 2000 dan menurun menjadi 11.50 juta ha pada 2001, atau sekitar 2,49%. Pada tahun yang sama terjadi penurunan produktivitas sebesar 0,30%, yaitu dari 44,01 ku/ha menjadi 43,88 ku/ha. Salah satu cara bagi pemerintah untuk memenuhi kebutuhan nasional adalah dengan terus berusaha meningkatkan produksinya, baik dengan intensifikasi maupun ekstensifikasi.
Datta (1975) dalam Puslitbangtan (2003), menyatakan bahwa pengembangan tanaman padi kemungkinan akan dilakukan di lahan kering. Datta menyebutkan, Indonesia memiliki lahan kering yang berpeluang untuk dimanfaatkan sebesar 9 juta ha, antara lain 480 ribu ha di Sumatera, 254 ribu ha di Kalimantan, 91 ribu ha di Sulawesi, 5 ribu ha di Maluku, dan sekitar 1.3 juta ha tersebar di wilayah lain. Hal ini pula yang melatarbelakangi adanya program peningkatan produksi lahan marjinal dengan pendekatan Pengelolaan Tanaman Terpadu (PTT) yang meliputi lahan kering, sawah tadah hujan, sawah pasang surut, dan lebak yang menjadi alternatif pengadaan pangan masa kini dan masa depan (Puslitbangtan, 2004).
Penanaman padi gogo terus dilakukan pada lahan kering dan marjinal sebagai salah satu upaya ekstensifikasi pertanian. Padi gogo adalah padi yang ditanam di lahan kering dengan sumber air terutama dari air hujan. Padi gogo dapat tumbuh pada ketinggian mulai diatas permukaan laut hingga 2500 mdpl (Datta, 1975 dalam Puslitbangtan, 2003). Lahan marjinal memiliki kendala antara lain tingkat kemasaman tanah yang tinggi, tercekam Al, dan lingkungan kering yang memudahkan Pyricularia oryzae menyerang pertanaman padi gogo. Tingkat produksi yang rendah pada padi gogo antara lain disebabkan daerah pertanaman yang kurang berkembang, tidak adanya jaringan irigasi yang memadai, gulma yang mengganas di lahan kering, serta serangan hama dan penyakit terutama penyakit blas yang disebabkan oleh cendawan Pyricularia oryzae (Partohardjono, 1992). Menurut Puslitbangtan (2003), produksi padi gogo petani menurun karena teknologi budidaya tradisional, penanaman varietas lokal, pengendalian gulma yang kurang intensif, dan munculnya serangan penyakit blas.
cara yang paling murah dan paling aman, apalagi jika didukung sistem budidaya yang tepat, sehingga bisa menghambat terbentuknya ras P. oryzae yang baru. Optimalisasi lahan marjinal tersebut dapat diupayakan dengan menciptakan varietas unggul baru padi gogo yang tahan terhadap penyakit blas, dan berdaya hasil tinggi.
Researh Genetics and Crops Improvement (RGCI), Departemen Agronomi dan Hortikultura, Institut Pertanian Bogor, memiliki sejumlah galur padi gogo F4:6 yang berdaya hasil tinggi. Galur-galur tersebut yang akan diuji dalam penelitian ini untuk evaluasi ketahanan terhadap penyakit blas daun dengan menyertakan varietas pembanding. Berdasarkan penelitian ini diharapkan akan mendapatkan galur- galur yang teruji ketahanannya terhadap penyakit blas daun, untuk selanjutnya dikembangkan melalui serangkaian uji daya hasil lanjutan menjadi varietas unggul baru padi gogo.
Tujuan
Mengevaluasi galur-galur padi gogo F4:6 yang berdaya hasil tinggi untuk ketahanan terhadap blas daun dalam upaya pengembangan varietas unggul baru padi gogo.
Hipotesis
1. Terdapat perbedaan ketahanan dibandingkan kontrol peka terhadap penyakit blas daun diantara galur- galur F4:6 padi gogo.
2. Terdapat paling tidak satu galur F4:6 padi gogo yang tahan dibandingkan kontrol peka terhadap serangan penyakit blas daun.
TINJAUAN PUSTAKA
Klasifikasi, Morfologi, dan Daur Hidup Pyricularia oryzae Cav.
Blas merupakan penyakit padi tertua yang penyebarannya meliputi semua negara penanam padi dan merupakan penyakit utama pada padi gogo. Penyakit ini telah menyerang lebih dari 70 negara penghasil padi di dunia (Thurston, 1984). Berdasarkan laporan pertama tahun 1637 dalam buku Agronomic Practices,
diketahui bahwa sejak zaman dinasti Ming di Cina sudah ditemukan penyakit demam padi oleh Soong Ying Shin yang kemudian disebut blas. Nama penyebab penyakit ini ditemukan oleh Cavara pada tahun 1891 dan sebutan blas pertama kali diberikan oleh Mitkalf pada tahun 1907 (Ou, 1972).
Pyricularia oryzae Cav. penyebab penyakit blas merupakan golongan cendawan yang secara morfologi tidak berbeda dengan Pyricularia grisea yang diketahui banyak menyerang jenis rumput-rumputan dan gulma (Rush, 1992). Menurut Alexopoulus dan Mims (1979), klasifikasi Pyricularia oryzae adalah sebagai berikut : termasuk Kingdom Myceteae, dengan Divisi Amastigomycota, Sub Divisi Deuteromycotina, Kelas Deuteromycetes, Ordo Moniliales, dan Famili Moniliaceae.
Konidiofor dari P. oryzae bersekat, antara 2-4 sekat, berkelompok dari setiap stroma, jarang bercabang, kelabu, licin dan membentuk konidia pada ujungnya (Semangun, 1971). Miselium septat, multinukleat, hifa bercabang. Konidiofor dan konidia dapat dengan mudah dihasilkan dari luka keabu-abuan. Konidiofor simpel, jarang bercabang, keabuan, septat dan ramping. Konidia oval, 2-septat ketika dewasa, dan meruncing hingga tumpul pucuknya, tergantung ras funginya. Sel-sel konidium multinukleat. Setiap inti memiliki dua kromosom besar dan kecil ( Chou dan Li, 1965 dalam Singh, 1978).
angin. Cendawan yang tumbuh kemudian menghasilkan lebih banyak spora. Konidia atau spora yang terbang terbawa angin kemudian jatuh pada daun padi. Pembentukan spora mencapai puncak pada 3-8 hari setelah timbul gejala awal. Spora dihasilkan oleh bercak sekitar 6 Hari Setelah Inokulasi (HSI).
Satu bercak bisa mencapai 2000-6000 spora per hari dalam dua minggu. Spora umumnya dilepas dini hari, sekitar pukul 02.00-06.00. Daerah tropis juga bisa terjadi pelepasan spora pada siang hari. Peranan air hujan sangat penting untuk pelepasan spora. Banyaknya spora yang ditangkap oleh daun tergantung pada kecepatan angin dan posisi atau sudut kemiringan daun (Semangun, 1991). Spora kemudian berkecambah dan cendawan menembus permukaan daun atau masuk ke dalam daun melalui stomata. Cendawan lalu tumbuh dan setelah 4-5 hari menimbulkan bercak pada daun. Bercak juga bisa terjadi pada buku dan bagian malai. Bercak memanjang runcing pada kedua ujungnya (belah ketupat) dengan ukuran dan bentuk berbeda menurut varietas padinya (Balitbangtan, 1991).
Cara Penyerangan dan Gejala Penyakit Blas
Konidiospor berkecambah pada permukaan daun dan menghasilkan apresoria coklat pada pucuk pembuluh, dimana infeksi akan berkembang dan melakukan penetrasi pada sel-sel epidermal melalui kutikula. Setelah invasi, fungi berhasil berkembang melalui percabangannya dan memasuki epidermal tetangga atau sel-sel tetangga dengan penetrasi langsung (Yoshi, 1937 dalam Matsuo et. al, 1995).
daripada tanaman yang masih muda. Infeksi pada bagian lain dari tanaman adalah pada pangkal malai, pelepah daun dan ruas batang (Singh, 1978).
Keragaman Patogen dari Ras Pyricularia oryzae Cav.
Tahun 1922, Sasaki di Jepang mencatat keberadaan ras-ras patogen P. oryzae. Sejak 1950-an, beberapa negara terutama Jepang, Amerika, dan Taiwan kemudian mempelajari ras-ras tersebut secara lebih intensif. Masing- masing negara menggunakan set yang berbeda dari varietas yang beragam. Banyak ras yang diidentifikasi di setiap negara tersebut. Kemudian pada 1960-an ada juga laporan mengenai ras-ras berikutnya dari Philipina, Indonesia, Korea, dan Kolombia (Ou, 1975 dalam Bonman, 1992).
P. oryzae diketahui mempunyai banyak ras fisiologi yang berbeda-beda sifat dan virulensinya. Pada tahun 1975, IRRI sudah melaporkan adanya 260 ras fisiologi (Semangun, 1991). Konidium-konidium yang dihasilkan oleh suatu biakan murni yang berasal dari konidium tunggal dapat berkembang menjadi banyak ras (Thurston, 1984). Patogenisitas dipengaruhi oleh perbedaan mekanisme metabolisme dan senyawa-senyawa kimia yang terdapat pada cendawan P. oryzae (Otsuka et al., 1963). Para peneliti Jepang dan India membuktikan bahwa P. oryzae mampu membentuk beberapa macam toksin, antara lain ß-picolinic acid, pyricularin, pyriculol, dan tenuazonic acid (Ou, 1985). Keragaman patogen dari P. oryzae sangat kompleks. Perubahan patogenisitas berlanjut seiring pertumbuhan fungi. Konidia dihasilkan pada luka tunggal, kultur konidia tunggal, atau dari pertumbuhan hifa ujung. Pada satu dari tiga sel konidia mungkin bisa berbeda rasnya. Satu tipe luka blas memproduksi 2000-6000 konidia. Jika lebih dari satu isolat monokonidial diuji, lebih dari 150 ras diketahui di Philipina yang diidentifikasi dari luka tunggal atau kultur tunggal.
Edwina dan Amir (1987) menemukan 27 ras blas berdasarkan perbedaan reaksinya dengan tujuh varietas diferensial di Indonesia, yaitu Asahan, Cisokan, IR 64, Krueng Aceh, Cisadane, Cisanggarung, dan Kencana Bali. Mogi et al.
(1993) dalam Makmur (2003), menemukan 15 ras baru dari 20 ras yang diidentifikasi di Taman Bogo, Lampung dan Kerang Agung, Sumatera Utara.
Tabel 1. Penentuan Ras P. oryzae dan Distribusi Patogenisitas Berdasarkan Reaksinya Terhadap Tujuh Varietas Diferensial di Indonesia.
Varietas
Ras dominan Nilai
skor 001 003 023 041 133 173 201
Asahan Cisokan IR 64 K. Aceh Cisadane Cisanggarung Kencana Bali
T T T T T T R
T T T T T R R
T T T R T R R
T T R T T T R
T R T R R R R
T R R R R R R
R R T T T T R
200 100 40 20 10 2 1 Keterangan : Ras-ras semakin virulen searah tanda panah
T : Tahan R : Rentan
Sumber : Mogi et al., (1991)
Mekanisme perubahan keragaman yang sangat cepat belum banyak diketahui. Studi saat ini menunjukkan bahwa banyak sel-sel dalam miselium dan konidia adalah uninukleat. Anastomosis dari hifa telah sering diamati. Beberapa peneliti awal percaya bahwa sel-sel multinukleat dan heterokariotik (Ou, 1975
dalam Bonman, 1992).
Mekanisme Ketahanan Tanaman Padi Terhadap Blas
sel-sel tersilifikasi per unit area meningkat secara cepat pada daun yang paling atas. Polimerisasi asam silikat memenuhi sela pada misel selulosa yang membentuk membran selulosa silikat. Tanaman padi yang tumbuh pada kondisi yang bebas silikon menunjukkan tingkat respirasi yang abnormal. Berdasarkan hasil- hasil penelitian tersebut dapat disimpulkan bahwa transportasi dan deposisi silikon pada tanaman padi berhubungan erat dengan transpirasi, karena silikon berada pada sepanjang jalur transpirasi. Lapisan silika gel pada epidermis juga berfungsi untuk mengontrol transpirasi serta menghalangi invasi fungi dan serangga. Aplikasi ammonium sulfat mampu mengurangi kehilangan ketahanan dari epidermis daun sehingga mampu menahan serangan blas.
Menurut Amir (1985), sistem budidaya juga berpengaruh terhadap sifat kerentanan padi gogo terhadap blas. Beberapa kebiasaan petani seperti penggunaan pupuk yang melebihi dosis anjuran terutama urea, membiarkan jerami padi, serta penggunaan pestisida yang tidak tepat bisa memicu meluasnya serangan blas, bahkan bisa menyebabkan terbentuknya ras baru yang semula tidak ditemukan.
Efek dari bentuk N yang diserap mungkin berdampak secara langsung terhadap kerentanan tanaman. Osuna-Canizalez et al., (1991) dalam Bonman (1992) membuktikan bahwa tanaman yang diberi NO3-N pada nutrisi kulturnya lebih rentan daripada yang diberikan NH4+-N. Mereka menyimpulkan bahwa bentuk N mempengaruhi mekanisme pertumbuhan yang menyebabkan kerentanan terhadap blas.
Lama pengembunan juga berpengaruh secara langsung pada patogen. Pada temperatur optimum (sekitar 25oC), patogen dapat menginfeksi tanaman setelah 6-8 jam pengembunan. Pada suhu sekitar 16oC infeksi terjadi kemudian setelah 16-20 jam. Temperatur di atas 28oC mendukung pertumbuhan P. oryzae tapi juga merangsang ketahanan inang. Tanaman lebih rentan pada temperatur rendah, sekitar 20oC, akan mendukung kemudahan infeksi walaupun pertumbuhan patogen saat itu rendah.
tahan pada ras patogen tertentu. Ketahanan parsial jika interaksi antara varietas tanaman dan berbagai ras patogen sama, sehingga tanaman bisa menahan berbagai serangan ras patogen. Ketahanan lengkap dapat diintroduksikan ke tanaman relatif mudah dan dampaknya akan terlihat dengan cepat yaitu adanya reaksi hipersensitifitas tanaman. Ketahanan lengkap berhubungan dengan plastisitas patogen. Jika tanaman memiliki ketahanan lengkap dan mendapat serangan ras patogen, maka patogen bisa menjadi plastis dan dapat memproduksi gen mematikan yang menyerang dan membentuk gen resistansi sehingga ras patogen lebih kuat menyerang atau membentuk ras yang lebih baru. Ketahanan parsial sebaliknya tidak berhubungan dengan plastisitas patogen. Patogen tidak mendapat keuntungan dengan membentuk ras baru karena tanaman mampu menahan serangan seluruh ras patogen. Kultivar dengan ketahanan parsial tetap menjadi kultivar yang tahan walaupun ras patogen menjadi beragam (Roumen, 1993).
Van der Plank (1975) mendefinisikannya sebagai ketahanan vertikal untuk ketahanan lengkap dan katahanan horisontal untuk ketahanan parsial. Ketahanan vertikal ditentukan oleh karakter kualitatif yang dikontrol oleh beberapa gen mayor atau monogenik sedangkan ketahanan horizontal dikontrol oleh gen-gen minor atau poligenik. Kriteria dari ketahanan vertikal adalah tipe luka, bukan jumlah luka. Ketahanan horizontal berdampak pada keduanya yaitu jumlah dan tipe luka dimana tipe luka memperlihatkan banyaknya sporulasi. Jumlah luka pada tanaman yang memiliki ketahanan horizontal lebih sedikit daripada varietas yang rentan pada kondisi yang sama dan diinokulasi dengan jumlah spora yang sama. Waktu yang diperlukan untuk luka baru hingga membentuk spora (periode antara inokulasi dan sporulasi selanjutnya) pada tanaman yang memiliki ketahanan horizontal lebih lama dibanding pada tanaman dengan ketahanan vertikal. Banyaknya sporulasi pada luka lebih sedikit pada ketahanan horizontal daripada vertikal (Van der Plank, 1975).
Kemampuan parasit adalah kemampuan dalam memanfaatkan tanaman sebagai sumber nutrisi, termasuk menemukan lokasi inang (Littlefield dan Heath, 1979
dalam Parlevliet, 1993). Penelitian kemudian diarahkan untuk mendapat gen-gen ketahanan yang mampu menahan serangan beberapa ras sekaligus melalui pendekatan bioteknologi. Laporan pertama pada studi genetik ketahanan terhadap blas disampaikan oleh Sasaki pada tahun 1922. Setelah 66 tahun kemudian penelitian terbagi menjadi dua fase. Fase pertama pada awal kemajuan penelitian, percobaan dilakukan untuk memperkirakan jumlah gen- gen yang mengontrol ketahanan. Pada fase kedua dimana studi intensif telah dilakukan untuk mengidentifikasi gen-gen ketahanan, strain-strain fungi yang telah diketahui patogenisitasnya digunakan untuk inokulasi. Penelitian yang dilakukan Sasaki membuka jalan untuk studi genetik hubungan inang-patogen yang berkembang pada 1960-an.
Distribusi geografis gen-gen ketahanan yang dihasilkan antara lain Pi-a
diidentifikasi pada varietas dari Jepang, Korea, Cina, India dan Pakistan. Pi-I
pada varietas dari Jepang, Korea dan USA, Pi-k dan Pi-km dari varietas Jepang dan Cina. Pi-kp dari varietas Jepang dan Pakistan, Pi-kh dari varietas Jepang, Vietnam, India dan USA, Pi-ks dari varietas Jepang, China dan USA. Pi-ta, Pi-ta2
dari varietas Jepang, China dan Philipina. Pi-z dari varietas Jepang, USA dan Perancis, Pi-z’ dari varietas India, Jepang, Thailand dan Malaysia. Pi-b, Pi-t dari varietas Jepang, Malaysia dan Indonesia dan Pi-sh dari varietas Jepang (Matsuo et al, 1995).
Studi pewarisan menunjukkan bahwa ketahanan dikendalikan oleh gen utama dominan atau gen utama dominan dan resesif yang dilatarbelakangi gen-gen minor (Makmur, 2003). Makmur juga menyatakan metode silang balik untuk mendapatkan ketahanan gen- gen utama sukar dilaksanakan, oleh karena itu metode untuk menghimpun gen- gen ketahanan lebih memungkinkan.
bahwa yang mengendalikan ketahanan untuk kedua ras tersebut berbeda dan bersifat independent satu dengan yang lainnya.
Utami (2000) dalam Rusdiansyah (2002) menyimpulkan bahwa dari hasil pengujian pada progeni BC2F3 hasil keturunan silang balik IR 64/Oryza
rufipogon/IR 64 terhadap tiga ras 173, 033, dan 001 menunjukkan bahwa sifat ketahanan pada O. rufipogon dikendalikan oleh banyak gen. Penelitian terbaru yang dilakukan Utami (2005) menunjukkan bahwa karakter ketahanan terhadap patogen blas bersifat kuantitatif dengan aksi gen epistasis duplikat untuk ras 001 dan 033, dan epistasis komplementer untuk ras 173.
Para tim ahli di International Rice Blast Genome Consortium and the Whitehead Institute for Genome Research di Cambridge, Massachusetts bekerjasama dengan U. S. Department of Agriculture-National Science Foundation Microbial Genome Sequencing Program, telah memetakan dan mempublikasikan genom P. oryzae yang bisa diakses via internet untuk memudahkan para peneliti dari negara lain memanfaatkannya. Para ahli mengidentifikasi ada sekitar 40 juta pasang basa pada genom P. oryzae yang tersimpan dalam fragmen DNA yang disebut BACs (Bacterial Artificial Chromosomes) (Dalke, 2002).
Metode Pengujian Terhadap Penyakit Blas
Upaya pengembangan metode pengujian atau teknik inokulasi sangat penting untuk menganalisis gen-gen ketahanan (Kiyosawa dan Ando, 1970 dalam
Pada metode inokulasi pelepah daun biasanya dilakukan pada tanaman tua. Tanaman dipotong dan dengan menurunkan suspensi spora yang ditempatkan di dalam pelepahnya. Pelepah yang diinokulasi dinkubasi sekitar 40 jam di petridish pada suhu 28o C. epidemis pada permukaan dalam kemudian dikerik keluar dan diuji dibawah mikroskop. Jumlah perkembangan miselia dari konidia tunggal pada sel-sel epidermal mengindikasikan tingkat kerentanan. Suspensi spora juga dapat disuntikkan pada pelepah daun paling atas yang mendekati malai ketika setengah malai telah muncul. Metode ini memberi persentase infeksi yang tinggi hingga 100%.
Menggunakan metode inokulasi daun, Takahashi (1963) menganalisa segregasi pada generasi F2 hasil persilangan antara varietas yang berbeda keahanannya. Hasilnya mengindikasikan varietas tahan memiliki satu,dua dan tiga gen-gen dominan. Abumiya (1959) dalam Takahashi (1963) kemudian menyatakan adanya resiko yang didapat dengan menggunakan metode inokulasi daun untuk pengujian rasio segregasi. Asai et. al. (1975) dalam Matsuo et. al,
(1995), juga melaporkan varietas yang diuji memperlihatkan perbedaan reaksi pada inokulasi semprot dan daun.
Efisiensi Infeksi Relatif (RIE) adalah komponen penting dari ketahanan parsial sesuai dengan hasil studi komponen ketahanan parsial, yaitu jumlah luka bersporulasi yang berkembang, ukuran luka dan periode laten. Genotipe-genotipe dengan tingkat ketahanan parsial yang tinggi memperlihatkan luka yang lebih sedikit dengan area sporulasi lebih kecil dibandingkan genotipe yang rentan. Analisis genetik menunjukkan bahwa RIE dikontrol oleh beberapa gen minor. Ketahanan parsial secara cepat meningkat sesuai umur daun. Semakin tinggi tingkat ketahanan parsial pada genotipe, semakin cepat jaringan daun menjadi tahan. Tingginya tingkat ketahanan terutama diekspresikan pada penurunan secara cepat terhadap luka bersporulasi (Roumen, 1993).
luka yang biasanya memiliki bercak berspora lebih kecil diandingkan yang rentan (Roumen,1993).
BAHAN DAN METODE
Waktu dan Tempat Penelitian
Penelitian dilaksanakan di rumah kasa Balai Besar Bioteknologi dan Sumberdaya Genetik Pertanian (BB BIOGEN), Cimanggu, Bogor. Penelitian ini dilaksanakan dari bulan Juni hingga Agustus 2005.
Bahan dan Alat
Bahan tanaman yang digunakan adalah 41 galur padi gogo generasi F6 yang berdaya hasil tinggi hasil seleksi di Cikeumeuh pada generasi F4 (galur-galur F4 : 6), tujuh tetua, dan dua varietas pembanding yaitu Kencana Bali dan Asahan sebagai pembanding peka dan tahan. Galur-galur tersebut berasal dari 21 populasi bersegregasi hasil persilangan half dialel dari tujuh tetua terpilih, yaitu Krowal, B8503E-TB-9-0-3, CT6510-24-1-3, Situgintung, Jatiluhur, IR 64 dan Gajah Mungkur. Bahan lain yang digunakan adalah isolat blas ras 033 yang mewakili ras dengan skala wilayah serangan yang luas tapi tidak begitu virulen, dan ras 173, yang mewakili ras dengan skala wilayah serangan yang sempit tapi sangat virulen. Media tanam menggunakan tanah dan pupuk N, P, dan K dengan dosis 5 gram Urea, 1.5 gram TSP, dan 1.2 gram KCl per 10 kg tanah kering.
Alat yang digunakan antara lain bak tanam, erlenmeyer, cawan petri, inkubator, kamar lembab, karung goni, kain blacu, inokulum aplikator, sprinkler
embun, mikroskop, haemacytometer, rumah kaca, dan rumah kasa.
Metode Penelitian
Penelitian dilakukan di rumah kasa menggunakan rancangan petak terbagi dengan dua faktor. Faktor isolat bla s ras 033 dan 173 sebagai petak utama, dan 50 tanaman padi gogo sebagai anak petak dengan tiga ulangan, sehingga terdapat 300 satuan percobaan. Benih padi gogo ditanam pada bak percobaan yang berukuran 40 x 28 x 7 cm3. Setiap bak terdiri dari 10 baris tana man untuk delapan galur dan dua pembanding. Masing- masing galur ditanam sebanyak 10 tanaman dengan satu benih per lubang tanam.
Yijk = µ +α i +βj +δi j + γk + (βγ)j k + εijk
i = Ulangan (i = 1,2,3)
j = Isolat Pyricularia oryzae (j =1,2) k = Galur padi gogo (k = 1,2,3,...,50)
Yijk = Nilai pengamatan pada isolat ke-j dari nomor galur ke-k dan ulangan ke- i
µ = Nilai rataan umum
αi = Pengaruh aditif ulangan ke- i
βj = Pengaruh aditif perlakuan ke-j faktor isolat
δij = Pengaruh galat yang muncul pada taraf ke-j faktor isolat dan
ulangan ke- i
γk = Pengaruh aditif dari taraf ke-k faktor galur padi gogo
(βγ)jk = Pengaruh interaksi taraf ke-j faktor isolat dan taraf ke-k faktor
galur
εijk = Pengaruh galat dari ula ngan ke- i yang memperoleh taraf ke-j
faktor isolat dan taraf ke- k faktor galur padi gogo
Pelaksanaan Penelitian
Penelitian diawali dengan persiapan bibit. Benih yang terpilih dipisahkan dan dioven dengan suhu 30°C selama lima hari. Selanjutnya benih ditanam dalam bak plastik berukuran 40 x 28 x 7 cm3 dengan 10 baris untuk delapan galur dan dua pembanding. Setiap galur ditanam 10 tanaman dengan satu benih per lubang tanam. Media tanam menggunakan tanah dan pupuk N, P, K. Pemupukan dilakukan sehari sebelum tanam dengan dosis pupuk 5 g Urea, 1.5 g TSP, dan 1.2 g KCl tiap 10 kg tanah kering.
Metode inokulasi yang digunakan adalah metode semprot. Tanaman yang sudah berumur 18 hari disemprot dengan inokulum blas sebanyak 50 ml/bak. Cara inokulasi adalah dengan menyemprotkan inokulum menggunakan labu erlenmeyer yang diberi glass automizer dan disambungkan dengan kompressor. Kerapatan spora yang digunakan ± 3 x 106 spora/ liter.
dinding ruangan. Selanjutnya tanaman dipindahkan ke dalam rumah kasa yang dindingnya dilapisi kain blacu dan bawahnya dialasi karung goni. Suhu rumah kasa antara 25-28oC dan kelembaban dipertahankan diatas 90% dengan cara penyiraman menggunakan sprinkler embun.
Pengamatan
Pengamatan dilakukan pada : 1. Periode laten
Periode laten adalah rentang waktu antara inokulasi hingga munculnya bercak blas. Periode laten dihitung sejak tanaman dikeluarkan dari kamar lembab. Daun telah bergejala bila telah muncul bercak walaupun sebesar ujung jarum. Bila salah satu tanaman telah bergejala maka itu disebut periode laten.
2. Jumlah daun terserang
Daun dianggap diserang apabila telah muncul bercak walaupun sebesar ujung jarum. Jumlah daun terserang dihitung mulai hari ketiga setelah inokulasi. Pengamatan selanjutnya dilakukan pada hari keenam, kesembilan dan 15 hari setelah inokulasi.
3. Jumlah bercak berspora
Bercak berspora adalah bercak yang berdiameter paling kecil 3 mm dan dikelilingi warna abu-abu dibagian luar. Jumlah bercak berspora dihitung pada tiga daun teratas untuk setiap rumpun padi. Tanaman diamati pada hari kesembilan dan kelimabelas setelah inokulasi.
4. Jumlah bercak tidak berspora
Bercak tidak berspora ditandai dengan bercak berwarna coklat, berdiameter kurang dari 3 mm dan tidak terdapat warna abu-abu dibagian luar. Jumlah bercak tidak berspora dihitung pada hari kesembilan dan kelimabelas setelah inokulasi.
5. Tingkat serangan penyakit
inokulasi. Pengamatan selanjutnya dilakukan pada hari keenam, kesembilan dan kelimabelas setelah inokulasi. Skor ketahanan untuk penyakit blas daun terdapat pada Tabel 2.
Gambar 1. Penampakan gejala serangan blas daun
Tabel 2. Klasifikasi serangan P. oryzae berdasarkan IRRI (1996).
Skor Kerusakan Daun Klasifikasi
0 1 2 3
4
5 6 7 8
9
Tidak ada bercak
Bercak sebesar ujung jarum
Bercak lebih besar dari ujung jarum
Bercak nekrotik, abu-abu, bundar, sedikit me manjang 1-2 mm, tepi berwarna cokelat
Bercak khas blas (belah ketupat), panjang 1-2 mm dan luas daun terserang kurang dari 2 %
Bercak khas blas, luas daun terserang 2-10 % Bercak khas blas, luas daun terserang 11-25 % Bercak khas blas, luas daun terserrang 26- 50 %
Bercak khas blas, luas daun terserang 51-75 % dan beberapa daun mulai mati
Semua daun mati
Tahan Tahan Tahan Tahan
Moderat
Moderat Moderat
Peka Peka
Peka
6. Intensitas serangan
Berdasarkan nilai skor ketahanan dapat dihitung nilai intensitas serangan (IS), yaitu kemampuan patogen menyerang dalam suatu populasi tanaman. IS dapat dihitung berdasarkan rumus dari IRRI (1996).
IS = S nxv X 100 % NxV
Keterangan : IS = intensitas serangan
N = jumlah rumpun yang diamati V = skor serangan yang tertinggi
Tanaman tergolong tahan jika persentase serangan 0-25%, 25-75% tergolong moderat, dan rentan bila diatas 75%.
Analisis Data
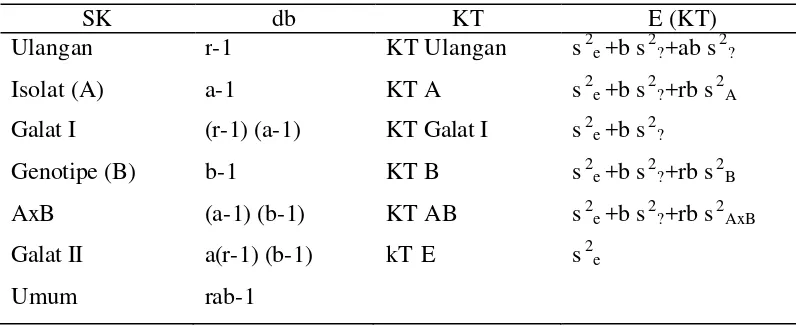
Data yang diperoleh dianalisis dengan analisis ragam menggunakan sidik ragam dan pendugaan ragam dari rancangan petak terbagi. Berdasarkan sidik ragam tersebut dapat dihitung nilai heritabilitas arti luas. Dirumuskan secara kuantitatif sebagai berikut :
Vg h2bs = Vg + Ve
Keterangan :
[image:42.596.113.511.514.677.2]h2bs = heritabilitas arti luas Vg = ragam genotipe Ve = ragam lingkungan
Tabel 3. Sidik Ragam dari Rancangan Petak Terbagi
SK db KT E (KT)
Ulangan Isolat (A) Galat I Genotipe (B) AxB
Galat II Umum
r-1 a-1 (r-1) (a-1) b-1
(a-1) (b-1) a(r-1) (b-1) rab-1
KT Ulangan KT A KT Galat I KT B KT AB kT E
s2e +b s2?+ab s2? s2e +b s2?+rb s2A s2e +b s2?
s2e +b s2?+rb s2B s2e +b s2?+rb s2AxB s2e
KT B-KT E (b-1) Vg =
r a
Ve = KT E
KT B = Kuadrat Tengah genotipe KT E = Kuadrat Tengah galat II r = Jumlah ulangan
a = Jumlah isolat b = Jumlah galur
HASIL DAN PEMBAHASAN
Kondisi Umum
Penelitian dilakukan di rumah kasa dengan suhu berkisar 25-28oC dan kelembaban diatas 90% yang diupayakan dengan cara penyiraman terus- menerus menggunakan springkler embun. Tanaman diinokulasi pada saat berumur 18 hari kemudian dipindahkan ke kamar lembab selama 48 jam untuk mempercepat penetrasi patogen ke tanaman inang. Pemeliharaan dilakukan terhadap tanaman sebelum inokulasi, yaitu minimal dua kali penyiraman setiap hari, karena kondisi rumah kaca saat tanam yang kering dan panas sehingga transpirasi sangat tinggi.
Untuk mengatasi keterbatasan tempat dan fasilitas, ulangan dilakukan dalam waktu yang berbeda dengan selang satu minggu antar ulangan. Kondisi tanaman berbeda tiap ulangan. Banyak galur dalam satu baris tanaman pada ulangan satu yang tidak tumbuh serempak. Ketidakserempakan tumbuh diduga karena perbedaan kedalaman benih saat tanam. Ulangan kedua dan ketiga dilakukan upaya antisipasi dengan menanam benih per baris lebih banyak dari ulangan pertama.
Kondisi pada ulangan kedua jauh lebih baik dibanding pada ulangan pertama. Tanaman tumbuh serempak walaupun ada satu atau dua tanaman yang tumbuh terlambat, namun tidak menjadi masalah karena dalam satu baris ditanam lebih dari 10 tanaman sehingga mampu mengatasi keterlambatan tumbuh benih. Pada ulangan ketiga ada beberapa galur yang pucuk daunnya memutih. Hal ini diduga terserang Aphelenchoides besseyi karena menunjukkan gejala yang sama, namun tidak menimbulkan kerusakan yang berarti.
Kerapatan spora yang digunakan adalah 4.8x105, 4.3x105, dan 5.3x105 untuk ras 033. Kerapatan spora untuk ras 173 adalah 2.3x105, 3.6x105, dan 3.5x105. Metode inokulasi yang digunakan adalah metode semprot dengan inokulum blas sebanyak 50 ml/bak. Pada pengamatan perkecambahan spora, terlihat bahwa spora sudah mulai berkecambah sehari setelah inokulasi.
kematian karena daun mengering atau parahnya serangan. Tanaman pada 15 hari setelah inokulasi sebagian besar mengalami recovery (kemunculan daun baru) dan beberapa tanaman mengalami kematian terutama karena tingkat serangan yang tinggi.
[image:45.596.177.448.374.530.2]Gambar 2. Tanaman yang siap diinokulasi dengan ras blas
Gambar 3. Tanaman yang ditempatkan di dalam kamar lembab
[image:45.596.176.448.575.737.2]Periode Laten
Periode laten adalah rentang waktu antara inokulasi hingga munculnya gejala serangan. Periode laten dihitung sejak tanaman dikeluarkan dari kamar lembab atau tiga Hari Setelah Inokulasi (HSI). Perhitungan berdasarkan waktu tercepat yang muncul diantara ketiga ulangan. Periode laten yang lama diharapkan menunjukkan tingkat ketahanan yang lebih baik, karena tanaman yang tahan memiliki mekanisme tertentu baik dari morfologi maupun fisiologinya yang mampu menghambat patogen untuk menginfeksi jaringan tanaman, sehingga walaupun terserang, gejala akan muncul lebih lama.
Hasil pengujian menggunakan ras 033 menunjukkan periode laten yang hampir sama diantara galur yang diuji. Sebanyak 36 galur memperlihatkan periode laten yang sama, yaitu 3 HSI. Hasil tersebut tidak berbeda dengan Kencana Bali sebagai kontrol peka. Empat galur memperlihatkan periode laten pada 4 HSI, yaitu PMT 06-C-C-09-9, PMT 10-C-C-08-5, PMT 12-C-C-42-7, dan PMT 05-C-C-03-10. Galur PMT 11-C-C-47-7 adalah satu-satunya galur dengan periode laten terlama yaitu 5 HSI, namun masih lebih cepat jika dibandingkan dengan Asahan sebagai kontrol tahan yaitu 6 HSI. Hasil pengamatan dengan menggunakan ras 173 menunjukkan rata-rata periode laten hampir semuanya sama diantara galur yang diuji. Sebanyak 40 galur dan Kencana Bali memperlihatkan periode laten yang sama, yaitu 3 HSI. Hanya satu galur yang memperlihatkan periode laten lebih panjang, yaitu galur PMT 21-C-C-09-10 pada 4 HSI. Galur PMT 11-C-C-47-7 juga menunjukkan tingkat ketahanan yang lebih baik, begitu pula galur PMT 21-C-C-09-10. Hal ini terlihat pada pengamatan intensitas serangan untuk kedua ras, dimana hasil pengamatan pada 3, 6, 9, dan 15 HSI menunjukkan reaksi tahan (Tabel 5. dan Tabel 6. Lampiran 1).
ditentukan oleh lamanya daun padi basah karena embun. Tanaman yang memiliki lapisan kutikula yang tebal menyebabkan lebih sukar ditembus cendawan P. oryzae. Menurut Thurston (1984), penetrasi kebanyakan terjadi secara langsung dengan menembus kutikula, walaupun jamur juga dapat melakukan penetrasi melalui mulut kulit.
[image:47.596.113.514.262.372.2]Jumlah Daun Terserang
Tabel 4. Rekapitulasi Sidik Ragam Peubah yang diamati pada 9 HSI
Peubah Ras Galur Interaksi
Jumlah daun terserang Jumlah bercak tak berspora Jumlah bercak berspora Intensitas serangan
sn sn tn sn
sn sn sn sn
sn tn tn n
Keterangan : Data ditransformasi dengan √(x+0,5)
sn : sangat nyata n : nyata
tn : tidak nyata
Hasil sidik ragam pada 9 HSI (Tabel 4) menunjukkan pengaruh yang sangat nyata diantara ras yang digunakan dan galur-galur yang diuji, artinya terdapat perbedaan tingkat virulensi ras blas yang digunakan dan terdapat perbedaan respon diantara galur- galur yang diuji terhadap jumlah daun yang terserang. Terdapat interaksi yang sangat nyata antara ras dan galur yang diuji artinya setiap ras memberikan pengaruh yang berbeda terhadap galur yang diuji pada peubah jumlah daun terserang.
yaitu 38 galur tahan dan tiga galur peka untuk ras 033 (Tabel 5), dan 39 galur tahan dan 2 galur peka untuk ras 173 (Tabel 6). Pada 15 HSI sebanyak 40 galur tahan dan satu galur peka untuk ras 033, sedangkan pada ras 173 sebanyak 37 galur tahan dan 4 galur peka (Tabel Lampiran 1 dan Tabel Lampiran 2).
Rata-rata jumlah daun terserang menurun pada pengamatan 15 HSI dibandingkan pada pengamatan sebelumnya. Menurut Roumen (1993), semakin tinggi tingkat ketahanan pada genotipe, semakin cepat jaringan daun menjadi tahan. Roumen juga menyatakan bahwa daun yang lebih tua memiliki tingkat ketahanan yang lebih tinggi. Pernyataan tersebut juga senada dengan Notteghem (1993) dimana ketahanan pada blas daun meningkat seiring meningkatnya umur daun. Menurut Kuilman (1940) dalam Semangun (1991), pada masa batang padi tumbuh memanjang (hingga berumur sekitar 55 hari), tanaman menjadi sangat rentan terhadap infeksi daun oleh cendawan P. oryzae.
Ada beberapa galur yang tahan berdasarkan intensitas serangan, namun tidak tahan untuk peubah jumlah daun terserang. Hal ini dimungkinkan karena walaupun lebih banyak jumlah daun yang terserang, namun lebih sedikit jumlah lukanya maka skor yang di dapatkanpun akan kecil, sehingga termasuk tahan berdasarkan intensitas serangan.
Jumlah Bercak Tidak Berspora
Tanaman tahan jika lebih sedikit jumlah bercak tak berspora karena semakin sedikit peluang untuk menjadi bercak berspora. Berdasarkan uji Dunnett pada taraf 5% tanaman tahan jika berbeda nyata lebih kecil dari kontrol peka.
Hasil sidik ragam pada 9 HSI (Tabel 5) menunjukkan pengaruh yang sangat nyata diantara ras dan galur-galur yang diuji. Hal ini berarti terdapat perbedaan virulensi ras yang digunakan, dan terdapat perbedaan respon diantara galur-galur yang diuji. Tidak terdapat interaksi yang nyata antara ras dan galur yang diuji terhadap peubah jumlah bercak tidak berspora. Hal ini berarti setiap ras tidak memberikan pengaruh yang berbeda pada galur yang diuji terhadap peubah jumlah bercak tidak berspora.
HSI sebanyak 34 galur tahan dan tujuh galur peka (Tabel 6). Pada 15 HSI terdapat 39 galur tahan dan dua galur peka untuk pengujian dengan ras 033, sedangkan pada ras 173 sebanyak 24 galur tahan dan 17 galur peka (Tabel Lampiran 3)
Tanaman yang tahan akan merespon secara cepat invasi fungi yang masuk, sehingga fungi tidak akan berkembang bahkan mati sebelum sempat bereproduksi. Itulah mengapa bercak tidak berkembang lagi pada tahap selanjutnya. Ketahanan tersebut diperlihatkan melalui reaksi hipersensitif inang. Reaksi dimana setelah fase invasi fungi ke tanaman inang, inang secara cepat merespon sehingga antara ras dan kultivar tidak kompatibel. Patogen akan mati diikuti kematian jaringan daun yang terserang. Pada studi yang dilakukan dimana ras dan kultivar tidak kompatibel, Koga dan Kobayashi (1982) dalam Matsuo et al. (1995) menyatakan bahwa hipersensitivitas dicirikan lepasnya secara cepat membran sitoplasma dari sel inang ketika fungi melakukan invasi. Hal ini antara lain disebabkan banyaknya
toxic active oxygens yang dilepas sel inang dalam merespon invasi fungi, yang menyebabkan hancurnya membran sitoplasma. Akumulasi dari toxic super oksida
yang tinggi tersebut menyebabkan kematian pada sel inang diikuti oleh kematian parasit. Hal ini yang menyebabkan bercak tidak berspora atau tidak mampu berkembang lagi.
Berdasarkan hasil pengamatan ada beberapa galur yang tahan berdasarkan intensitas serangan, namun tidak tahan untuk peubah jumlah bercak tidak berspora. Bercak tidak berspora walaupun berperan dalam penentuan skor ketahanan akan tetapi tidak menjamin tingginya skor, karena dipengaruhi juga oleh banyaknya bercak berspora. Berdasarkan hal itu maka galur yang tahan untuk intensitas serangan belum tentu tahan berdasarkan bercak tak berspora.
berspora tidak diikuti tingginya bercak berspora pada saat yang sama, sehingga skor yang didapatkanpun kecil, dan berdasarkan intensitas serangan ermasuk tahan.
Jumlah Bercak Berspora
Hasil sidik ragam pada 9 HSI (Tabel 4) menunjukkan pengaruh yang sangat nyata diantara galur-galur yang diuji, dan tidak nyata untuk ras dan interaksi untuk peubah jumlah bercak berspora. Hal ini berarti, terdapat perbedaan respon dari galur-galur yang diuji, tidak terdapat perbedaan virulensi dari ras yang digunakan, dan tidak ada perbedaan pengaruh dari ras yang digunakan terhadap galur-galur yang diuji untuk peubah jumlah bercak berspora.
Bercak berspora merupakan salah satu komponen terpenting dalam menentukan ketahanan tanaman terhadap blas daun. Roumen (1993) menyatakan bahwa berdasarkan studi mendetail dari komponen ketahanan mengindikasikan bahwa jumlah luka adalah komponen yang paling penting.
Tanaman tahan jika lebih sedikit jumlah bercak berspora, karena bercak berspora mempengaruhi nilai intensitas serangan. Semakin banyak jumlah bercak berspora, akan semakin tinggi pula skor yang didapatkan sehingga intensitas serangan akan semakin tinggi. Berdasarkan uji dunnett pada taraf 5%, tanaman tahan jika berbeda nyata lebih kecil dari kontrol peka.
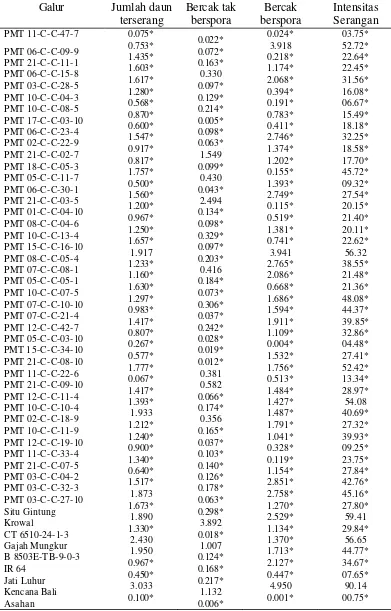
Tabel 5. Rata-rata Hasil Peubah Galur- galur Padi Gogo F4:6 yang diamati pada 9 HSI untuk ras 033
Galur Jumlah daun terserang Bercak tak berspora Bercak berspora Intensitas Serangan PMT 11-C-C-47-7 PMT 06-C-C-09-9 PMT 21-C-C-11-1 PMT 06-C-C-15-8 PMT 03-C-C-28-5 PMT 10-C-C-04-3 PMT 10-C-C-08-5 PMT 17-C-C-03-10 PMT 06-C-C-23-4 PMT 02-C-C-22-9 PMT 21-C-C-02-7 PMT 18-C-C-05-3 PMT 05-C-C-11-7 PMT 06-C-C-30-1 PMT 21-C-C-03-5 PMT 01-C-C-04-10 PMT 08-C-C-04-6 PMT 10-C-C-13-4 PMT 15-C-C-16-10 PMT 08-C-C-05-4 PMT 07-C-C-08-1 PMT 05-C-C-05-1 PMT 10-C-C-07-5 PMT 07-C-C-10-10 PMT 07-C-C-21-4 PMT 12-C-C-42-7 PMT 05-C-C-03-10 PMT 15-C-C-34-10 PMT 21-C-C-08-10 PMT 11-C-C-22-6 PMT 21-C-C-09-10 PMT 12-C-C-11-4 PMT 10-C-C-10-4 PMT 02-C-C-18-9 PMT 10-C-C-11-9 PMT 12-C-C-19-10 PMT 11-C-C-33-4 PMT 21-C-C-07-5 PMT 03-C-C-04-2 PMT 03-C-C-32-3 PMT 03-C-C-27-10 Situ Gintung Krowal CT 6510-24-1-3 Gajah Mungkur B 8503E-T B-9-0-3 IR 64 Jati Luhur Kencana Bali Asahan 0.075* 0.753* 1.435* 1.603* 1.617* 1.280* 0.568* 0.870* 0.600* 1.547* 0.917* 0.817* 1.757* 0.500* 1.560* 1.200* 0.967* 1.250* 1.657* 1.917 1.233* 1.160* 1.630* 1.297* 0.983* 1.417* 0.807* 0.267* 0.577* 1.777* 0.067* 1.417* 1.393* 1.933 1.212* 1.240* 0.900* 1.340* 0.640* 1.517* 1.873 1.673* 1.890 1.330* 2.430 1.950 0.967* 0.450* 3.033 0.100* 0.022* 0.072* 0.163* 0.330 0.097* 0.129* 0.214* 0.005* 0.098* 0.063* 1.549 0.099* 0.430 0.043* 2.494 0.134* 0.098* 0.329* 0.097* 0.203* 0.416 0.184* 0.073* 0.306* 0.037* 0.242* 0.028* 0.019* 0.012* 0.381 0.582 0.066* 0.174* 0.356 0.165* 0.037* 0.103* 0.140* 0.126* 0.178* 0.063* 0.298* 3.892 0.018* 1.007 0.124* 0.168* 0.217* 1.132 0.006* 0.024* 3.918 0.218* 1.174* 2.068* 0.394* 0.191* 0.783* 0.411* 2.746* 1.374* 1.202* 0.155* 1.393* 2.749* 0.115* 0.519* 1.381* 0.741* 3.941 2.765* 2.086* 0.668* 1.686* 1.594* 1.911* 1.109* 0.004* 1.532* 1.756* 0.513* 1.484* 1.427* 1.487* 1.791* 1.041* 0.328* 0.119* 1.154* 2.851* 2.758* 1.270* 2.529* 1.134* 1.370* 1.713* 2.127* 0.447* 4.950 0.001* 03.75* 52.72* 22.64* 22.45* 31.56* 16.08* 06.67* 15.49* 18.18* 32.25* 18.58* 17.70* 45.72* 09.32* 27.54* 20.15* 21.40* 20.11* 22.62* 56.32 38.55* 21.48* 21.36* 48.08* 44.37* 39.85* 32.86* 04.48* 27.41* 52.42* 13.34* 28.97* 54.08 40.69* 27.32* 39.93* 09.25* 23.75* 27.84* 42.76* 45.16* 27.80* 59.41 29.84* 56.65 44.77* 34.67* 07.65* 90.14 00.75*
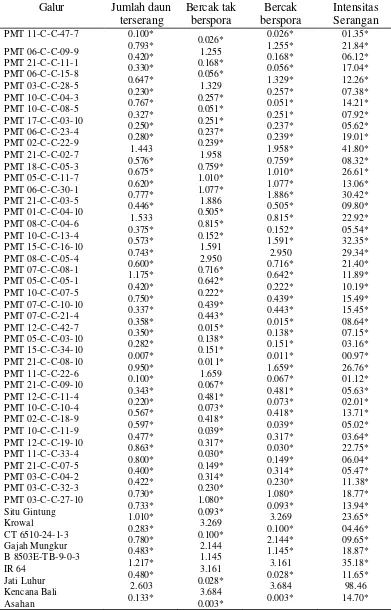
Tabel 6. Rata-rata Hasil Peubah Galur- galur Padi Gogo F4:6 yang diamati pada 9 HSI untuk ras 173
Galur Jumlah daun terserang Bercak tak berspora Bercak berspora Intensitas Serangan PMT 11-C-C-47-7 PMT 06-C-C-09-9 PMT 21-C-C-11-1 PMT 06-C-C-15-8 PMT 03-C-C-28-5 PMT 10-C-C-04-3 PMT 10-C-C-08-5 PMT 17-C-C-03-10 PMT 06-C-C-23-4 PMT 02-C-C-22-9 PMT 21-C-C-02-7 PMT 18-C-C-05-3 PMT 05-C-C-11-7 PMT 06-C-C-30-1 PMT 21-C-C-03-5 PMT 01-C-C-04-10 PMT 08-C-C-04-6 PMT 10-C-C-13-4 PMT 15-C-C-16-10 PMT 08-C-C-05-4 PMT 07-C-C-08-1 PMT 05-C-C-05-1 PMT 10-C-C-07-5 PMT 07-C-C-10-10 PMT 07-C-C-21-4 PMT 12-C-C-42-7 PMT 05-C-C-03-10 PMT 15-C-C-34-10 PMT 21-C-C-08-10 PMT 11-C-C-22-6 PMT 21-C-C-09-10 PMT 12-C-C-11-4 PMT 10-C-C-10-4 PMT 02-C-C-18-9 PMT 10-C-C-11-9 PMT 12-C-C-19-10 PMT 11-C-C-33-4 PMT 21-C-C-07-5 PMT 03-C-C-04-2 PMT 03-C-C-32-3 PMT 03-C-C-27-10 Situ Gintung Krowal CT 6510-24-1-3 Gajah Mungkur B 8503E-T B-9-0-3 IR 64 Jati Luhur Kencana Bali Asahan 0.100* 0.793* 0.420* 0.330* 0.647* 0.230* 0.767* 0.327* 0.250* 0.280* 1.443 0.576* 0.675* 0.620* 0.777* 0.446* 1.533 0.375* 0.573* 0.743* 0.600* 1.175* 0.420* 0.750* 0.337* 0.358* 0.350* 0.282* 0.007* 0.950* 0.100* 0.343* 0.220* 0.567* 0.597* 0.477* 0.863* 0.800* 0.400* 0.422* 0.730* 0.733* 1.010* 0.283* 0.780* 0.483* 1.217* 0.480* 2.603 0.133* 0.026* 1.255 0.168* 0.056* 1.329 0.257* 0.051* 0.251* 0.237* 0.239* 1.958 0.759* 1.010* 1.077* 1.886 0.505* 0.815* 0.152* 1.591 2.950 0.716* 0.642* 0.222* 0.439* 0.443* 0.015* 0.138* 0.151* 0.011* 1.659 0.067* 0.481* 0.073* 0.418* 0.039* 0.317* 0.030* 0.149* 0.314* 0.230* 1.080* 0.093* 3.269 0.100* 2.144 1.145 3.161 0.028* 3.684 0.003* 0.026* 1.255* 0.168* 0.056* 1.329* 0.257* 0.051* 0.251* 0.237* 0.239* 1.958* 0.759* 1.010* 1.077* 1.886* 0.505* 0.815* 0.152* 1.591* 2.950 0.716* 0.642* 0.222* 0.439* 0.443* 0.015* 0.138* 0.151* 0.011* 1.659* 0.067* 0.481* 0.073* 0.418* 0.039* 0.317* 0.030* 0.149* 0.314* 0.230* 1.080* 0.093* 3.269 0.100* 2.144* 1.145* 3.161 0.028* 3.684 0.003* 01.35* 21.84* 06.12* 17.04* 12.26* 07.38* 14.21* 07.92* 05.62* 19.01* 41.80* 08.32* 26.61* 13.06* 30.42* 09.80* 22.92* 05.54* 32.35* 29.34* 21.40* 11.89* 10.19* 15.49* 15.45* 08.64* 07.15* 03.16* 00.97* 26.76* 01.12* 05.63* 02.01* 13.71* 05.02* 03.64* 22.75* 06.04* 05.47* 11.38* 18.77* 13.94* 23.65* 04.46* 09.65* 18.87* 35.18* 11.65* 98.46 14.70*
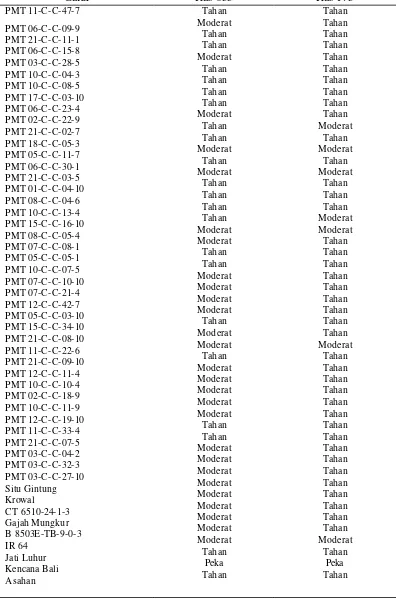
Tabel 7. Tingkat Ketahanan Galur-galur Padi Gogo F4:6 terhadap Penyakit Blas Daun pada 9 HSI
Galur Ras 033 Ras 173
Genotipe-genotipe dengan tingkat ketahanan yang tinggi menunjukkan sedikit luka yang biasanya memiliki bercak berspora yang lebih sedikit dibandingkan yang rentan. Ou (1975) menyatakan bahwa pada varietas dengan tingkat ketahanan yang tinggi, patogen tidak membentuk bercak atau bercak hanya berupa bintik cokelat sebesar kepala jarum. Menurut Takahashi (1951)
dalam Matsuo et al. (1995), tanaman tahan dicirikan dengan adanya reaksi hipersensitif inang, yaitu hanya terjadi bintik-bintik kuning muda dan hifa yang menginfeksi hanya bisa melakukan penetrasi. Tanaman moderat dicirikan terjadinya perubahan warna cokelat gelap pada sel yang terinfeksi dan sel tetangganya, dan tanaman mampu menekan petumbuhan fungi yang menginfeksi. Tanaman peka dicirikan adanya respon nekrotik secara lambat pada sel, diikuti perubahan warna cokelat tua, dan selanjutnya fungi tumbuh menginfeksi sel tetangga.
Intensitas Serangan
Intensitas serangan menunjukkan tingkat serangan ras blas daun yang diinokulasikan menginfeksi galur-galur padi gogo yang diuji. Intensitas serangan merupakan salah satu indikator ketahanan tanaman terhadap serangan penyakit. Perhitungan berdasarkan rumus dari IRRI dimana nilai intensitas serangan antara 0-25% tergolong tahan, 26-75% tergolong moderat, dan diatas 75% tergolong rentan.
Hasil sidik ragam pada 9 HSI (Tabel 4) menunjukkan pengaruh yang sangat nyata diantara ras dan galur-galur yang diuji, dan terdapat interaksi yang nyata antara ras dan galur yang diuji untuk peubah intensitas serangan. Hal ini berarti bahwa terdapat perbedaan virulens i dari ras yang digunakan, terdapat perbedaan respon dari galur- galur yang diuji, dan terdapat perbedaan pengaruh dari ras yang digunakan terhadap galur-galur yang diuji untuk peubah intensitas serangan.
HSI sebanyak 21 galur menunjukkan reaksi tahan dan 20 galur menunjukkan reaksi moderat untuk ras 033 (Tabel 5), sedangkan untuk ras 173 sebanyak 35 galur menunjukkan reaksi tahan dan enam galur menunjukkan reaksi moderat (Tabel 6). Pada pengamatan 15 HSI sebanyak 36 galur menunjukkan reaksi tahan dan lima galur peka untuk ras 033, dan untuk pengujian menggunakan ras 173 sebanyak 37 galur menunjukkan reaksi tahan dan sebanyak empat galur menunjukkan reaksi moderat (Tabel Lampiran 7 dan Tabel Lampiran 8).
Pada pengamatan 3 HSI dan 6 HSI belum bisa dijadikan standar penentuan ketahanan karena penetrasi patogen belum menyeluruh ke jaringan tanaman. Singh (1978) menyebutkan, bahwa spora dari bercak akan berkembang hingga sekitar 6 HSI. Pada 15 HSI sebagian besar tanaman mengalami recovery atau kemunculan daun-daun baru yang tidak terinfeksi lagi oleh patogen, sehingga tidak bisa dijadikan standar penentuan tingkat ketahanan. Berdasarkan hal tersebut maka 9 HSI dijadikan waktu yang tepat untuk penentuan tingkat ketaha nan, dimana penetrasi patogen sudah dianggap sempurna dan tanaman sudah cukup kuat untuk menangkal serangan patogen.
Menurut hasil tersebut, maka pada 9 HSI dari 41 galur yang diuji diperoleh 17 galur yang tergolong tahan untuk kedua ras, empat galur tergolong moderat untuk kedua ras, dua galur hanya tahan untuk ras 033 dan 18 galur hanya tahan terhadap ras 173. Hasil tersebut memperlihatkan lebih banyak galur yang tahan terhadap ras 173 walaupun tingkat virulensi ras 173 lebih tinggi jika dibandingkan ras 033 (Tabel 7) . Hal tersebut diduga karena kerapatan spora yang digunakan, dimana kerapatan spora ras 033 lebih tinggi dibandingkan ras 173. Konsentrasi yang tinggi tersebut diduga meningkatkan serangan patogen walaupun virulensinya lebih rendah (Nasution, 2005).
berkorespondensi dengan gen avirulen dari patogen yang menyerang, begitupun sebaliknya. Tingkat ketahanan yang tinggi secara umum diatur oleh satu atau beberapa gen mayor yang hanya efisien dengan isolat P. oryzae yang avirulen (Kiyo sawa, 1986 dalam Dioh et al., 2000). Gen- gen avirulen tersebut mengkode protein-protein melalui sinyal peptida untuk sekresi yang diekspresikan hanya selama infeksi (Dioh et al., 2000)
Uji Korelasi
[image:56.596.110.508.402.487.2]Korelasi memperlihatkan hubungan antara satu karakter dengan karakter lainnya. Hasil uji korelasi dari jumlah daun terserang, jumlah bercak tak berspora, dan jumlah bercak berspora terhadap intensitas serangan pada 9 HSI menunjukkan nilai yang positif sangat nyata. Hal ini berarti apabila karakter tersebut meningkat, maka akan meningkatkan intensitas serangan.
Tabel 8. Hasil Uji Korelasi Terhadap Intensitas Serangan pada 9 HSI
Peubah Korelasi
Jumlah Daun Terserang pada 9 HSI Jumlah Bercak Tak Berspora pada 9 HSI Jumlah Bercak Berspora pada 9 HSI
0.842** 0.200** 0.423** Keterangan : (**) : sangat nyata
(*) : nyata
Berdasarkan uji korelasi tersebut, semakin banyak jumlah daun yang terserang, jumlah bercak tak berspora, dan jumlah bercak berspora, maka intensitas serangan akan semakin tinggi. Hal ini berarti kita dapat melakukan seleksi pada peubah jumlah daun terserang, karena lebih mudah dalam pengamatan dibandingkan peubah lainnya.
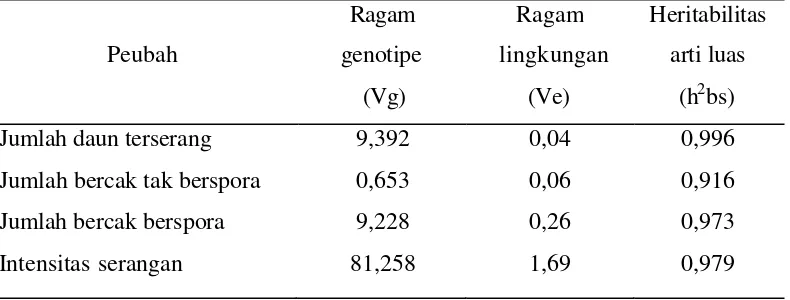
Heritabilitas Arti Luas
menunjukkan kekuatan dari hubungan banyak gen terhadap lingkungan. Nilai dihitung berdasarkan sidik ragam dari rancangan petak terbagi.
Tabel 9. Nilai Heritabilitas Arti Luas Peubah yang diamati pada 9 HSI
Peubah
Ragam genotipe
(Vg)
Ragam lingkungan
(Ve)
Heritabilitas arti luas
(h2bs) Jumlah daun terserang
Jumlah bercak tak berspora Jumlah bercak berspora Intensitas serangan
9,392 0,653 9,228 81,258
0,04 0,06 0,26 1,69
0,996 0,916 0,973 0,979
Menurut Mc Whirter (1979) dalam Rachmadi (1999), penduga heritabilitas suatu karakter termasuk tinggi apabila nilainya diatas 50%, termasuk sedang apabila nilainya antara 20-50%, dan termasuk rendah apabila nilainya dibawah 20%. Nilai heritabilitas yang tinggi mengindikasikan bahwa variasi yang muncul antara individu tanaman lebih disebabkan oleh faktor genetik, sehingga besar harapan akan diperolehnya kemajuan secara genetik.
KESIMPULAN DAN SARAN
Kesimpulan
Terdapat perbedaan ketahanan dari galur- galur padi gogo yang diuji dibandingkan dengan kontrol peka terhadap serangan ras blas daun ras 033 dan 173. Pengamatan intensitas serangan pada 9 HSI untuk ras 033 terdapat 20 galur yang menunjukkan reaksi tahan. Pada pengujian dengan ras 173 terdapat 35 galur yang menunjukkan reaksi tahan. Berdasarkan hasil tersebut, sebanyak 17 galur menunjukkan reaksi tahan untuk kedua ras dan dua galur menunjukkan reaksi moderat untuk kedua ras.
Saran
DAFTAR PUSTAKA
Adiratna, E. R. 2004. Seri Agriwawasan Stop Tanam Padi. Penebar Swadaya, Jakarta. 116 p.
Alexopoulos, C. J. and C. W. Mims. 1979. Introductory Mycology. John Wiley and Sons, New York. 631 p.
Allard, R. W. 1960. Principles of Plant Breeding. New John Wiley and Sons. New York.
Amir, M. 1985. Blast Disease ( Pyricularia oryzae Cav.) Control on Dry Land (Seminar of May, 1985). Directory of Food Crops Protection. Pasar Minggu. Jakarta North. 24 p.
Balitbangtan. 1981. Padi Buku 1. Balitbangtan. Bogor.
Balitbangtan. 1991. Padi Buku 3. Balitbangtan. Bogor.
Bonman, J. M. 1992. Durable resistance to Rice Blast disease-environment influences p. 115-124 In R. Johnson and G. J. Jellis (eds.) Proceed. of a Breeding for Disease Resistance. Kluwer Academic Publishers. Nethe rland. 205 p.
BPS. 1999. Survey Pertanian Luas Lahan menurut Penggunaannya di Indonesia. BPS. Jakarta. 195 p.
BPS. 2001. Produksi Tanaman Padi dan Palawija di Indonesia. BPS. Jakarta. 108p.
Dalke, K. 2002. Fighting Blast Disease: Rice Pathogen Sequenced. Annual Review of Microbiology. Vol. 50. USA..
Dioh, W., D. Tharreau, J. L. Notteghem, M. Orbach and M. Lebrun. 2000. Molecular Plant Microbe Interaction. The American Phithopathology Society. Vol 13(2) : 217-227. Saint Paul, USA.
Edwina, R. dan M. Amir. 1987. Identifikasi ras jamur Pyricularia oryzae Cav. Dari beberapa daerah di Indonesia. Kongres Nasional IX Perhimpunan Fitopatologi Indonesia p. 209-212.
Irawan, B. 2003. Konversi Lahan Sawah di Indonesia dan Dampaknya Terhadap Produksi Padi Sawah. Prosiding Seminar Nasional Ekonomi Padi dan Beras Indonesia. Jakarta, November 2003.
IRRI. 1996. Standard Evaluation System for Rice. 4th ed. IRRI. Los Banos, Philiphines. 52 p.
Kiyosawa, S. 1972. Genetics of Blast Resistance p. 203-236. In IRRI (ed). Rice Breeding. Los Banos, Philiphines.
Kuckuck, H. G. Kobabe, G. Wenzel. 1991. Fundamentals of Plant Breeding. Springer-Verlag, Berlin Heidelberg, Germany. 236 p.
Makmur, A. 2003. Pemuliaan Tanaman bagi Lingkungan Spesifik. IPB Press, Bogor. 53 p.
Matsuo, T., K. Kumazawa, and R. Ishii. 1995. Science of the Rice Plant Vol. II Physiology. Food and Agriculture Research Center, Tokyo. 1239 p.
Mogi, Shizuo, S. Zaenuddin, B. S. Wibowo, E. Ros dan C. Irawan. 1991. Establishment of the Differential Variety Series for Pathogenic Race Identification of Rice Blast Fungus and Distribution of Race Based on the New Differentials in Indonesia. Rice Disease Study Group. Karawang, Indonesia.
Nasution, A. 1995. Pencarian Resistensi Terhadap Blas Daun dan Leher Malai. Risalah Kongres Nasional XII dan Seminar Ilmiah Perhimpunan Fitopatologi Indonesia. Yogyakarta, 6-8 September 1993. p. 140-145.
Nasution, 2005. Komunikasi Pribadi.
Nelson, R R. 1975. Horizontal resistance in plant : Concepts, Controversies and Applications. Proceed. Of the Seminar on Horizontal Resistance to Blast Disease of Rice. CIAT.
Notteghem, J. L.1993. Durable Resistance to Rice Blast Disease p. 125-131 In R. S. Zeigler, S. A. Leong, P. S. Teng (eds.) Proceed. of a Rice Blas Disease. International Rice Research Institute. Los Banos, Philiphines. 626 p.
Otsuka, H., Kinjiro T., Nagahiro O. 1963. Variability of Pyricularia oryzae in Culture in the Rice Blast Disease. Proceed. Of a symposium at the International Rice Research Institute. The Johns Hopkins Press. Baltimore, Maryland. In Rice Blast Disease. Proceed. Of a symposium at the International Rice Research Institute. John Hopkins Press. USA.
Ou, S. H. 1972. Rice Disease. Commonwealth Mycologycal Institute. England.
Ou, S. H. 1975. A Hand Book of Rice Diseases in the Tropics. IRRI. 58 p.
Parlevliet, J. E., R. E. Nicks, and P. R. Ellis. 1993. Resistance to Parasites. p.422-445 In Hayward, M. D., N. O. Bosemark and I. R