KAJI AN H ABI TAT, TI N GKAH LAKU, D AN POPULASI
KALAW ET
( H y loba t e s a gilis a lbiba r bis)
D I TAM AN
N ASI ON AL SEBAN GAU KALI M AN TAN TEN GAH
YULI US D UM A
SEKOLAH PASCASARJAN A
I N STI TUT PERTAN I AN BOGOR
PERNYATAAN MENGENAI DISERTASI DAN SUMBER INFOTMASI
Dengan ini saya menyatakan bahwa segala pernyataan dalam disertasi saya berjudul :
KAJIAN HABITAT, TINGKAH LAKU, DAN POPULASI KALAWET (Hylobates agilis albibarbis) DI TAMAN NASIONAL SEBANGAU
KALIMANTAN TENGAH
adalah benar merupakan tulisan disertasi berdasarkan hasil penelitian saya sendiri dengan arahan Komisis Pembimbing dan belum diajukan dalam bentuk apapun kepada perguruan tinggi manapun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun yang tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir disertasi ini.
Bogor, Mei 2007
Yulius Duma
ABSTRAK
YULIUS DUMA. Kajian Habitat, Tingkah Laku, dan Populasi Kalawet (Hylobates agilis albibarbis) di Taman Nasional Sebangau, Kalimantan Tengah. Dibimbing oleh : SRI SUPRAPTINI MANSJOER, R.R. DYAH PERWITASARI, dan DONDIN SAJUTHI.
Penelitian dilaksanakan di Laboratorium Alam Hutan Gambut (LAHG) CIMTROP Universitas Palangka Raya, di Taman Nasional Sebangau, Kalimantan Tengah, untuk mengetahui kondisi habitat kalawet, mengamati pola aktivitas kalawet (Hylobates agilis albibarbis) memanfaatkan habitat; mendapatkan data populasi kalawet terkini; dan mengkaji vokalisasi sebagai penciri spesies Hylobates di Kalimantan. Analisis vegetasi dilakukan dengan metode garis berpetak, pengamatan aktivitas kalawet dilakukan dengan metode scan sampling, estimasi populasi dengan metode fixed point count, dan analisis vokalisasi dengan software Adobe Audition 1.5 dan Raven 1.2.1. Rata-rata tinggi pohon di tipe MSF dan TIF 26,70 m (kisaran 20-45m), sedangkan di LPF 19,46m (kisaran 15-25 m). Lebih dari 90% (kisaran 90-100%) pohon di MSF dan TIF potensial sebagai sumber pakan kalawet, sedangkan di LPF sekitar 54% (kisaran 35-73%). Kalawet KC menggunakan waktunya sekitar 41% untuk aktivitas makan, 32% istirahat, 14% berpindah, dan 13% untuk vokalisasi; dengan pola makan 73% berupa buah, 16% daun, 10% bunga, dan 1% lainnya. Aktivitas harian tersebut dilakukan dalam daerah jelajah seluas 29,5 ha. Kepadatan populasi kalawet di kawasan TN. Sebangau bervariasi menurut tipe hutan, masing-masing sebesar 2,33 kelompok/km2, dan 8,02 individu/km2 di hutan gambut campuran (MSF); 0,44 kelompok/km2, dan 1,52 individu/km2 di hutan tegakan rendah (LPF); dan 3,10 kelompok/km2 dan 10,65 individu/km2 di hutan tegakan tinggi (TIF). Jumlah kelompok dan individu kalawet yang ada di LAHG sebanyak 700 kelompok, dan 2.404 individu; dan TN. Sebangau sebanyak 7.988 kelompok, dan 27.442 individu. Analisis vokalisasi Hylobates di Kalimantan dapat dijadikan acuan dalam mengidentifikasi ‘spesies’ khususnya hibrida antara H. agilis albibarbis x H. muelleri.
ABSTRACT
YULIUS DUMA. Habitat, Behavior, and Population Analysis of Bornean Agile Gibbon (Hylobates agilis albibarbis) at Sebangau National Park, Central Kalimantan. Under Supervision of SRI SUPRAPTINI MANSJOER, R.R. DYAH PERWITASARI, and DONDIN SAJUTHI.
The research was conducted in Natural Laboratory of Peat Forest (NLPF) CIMTROP University of Palangka Raya, in Sebangau National Park, Central Kalimantan. The aim of this research is to study the habitat of Bornean agile gibbon (kalawet), its activity patterns; population density; and calling analysis to identify the species of Hylobates in Kalimantan. Kalawet habitat was analysed by block line method; the activity patterns of kalawet was conducted by scan sampling method. Population density was counted by fixed point count method, while vocalization was analysed by Adobe Audition 1.5 and Raven 1.2.1 software’s. The average tree high in MSF and TIF was 26.70 m (range: 20-45m), while in LPF it was 19.46 m (range: 15-25 m). More than 90% (range: 90-100%) trees in MSF and TIF had potential as feed source of kalawet, while in LPF it was 54% (range: 35-73%). Kalawet used the time budget was about 41% for feeding, 32% for resting, 14% for moving, and 13% for vocalization. Feeding pattern was 73% fruits, 16% leaves, 10% flowers, and 1% others. Kalawet spent time for daily activity done in home range of 29.5 ha. Population density of kalawet in Sebangau National Park varied according to forest type. In mixed swamp forest (MSF) were 2.33 groups/km2 and 8.02 individuals/km2, in low pole forest (LPF) 0.44 groups/km2, and 1.52 individuals/km2, and in tall interior forest (TIF) 3.10 groups/km2 and 10.65 individuals/km2. There were 700 groups, and 2,404 individuals in NLPF; and 7,988 groups, and 27,442 individuals in Sebangau National Park. Analysis of vocalization of Hylobates in Kalimantan can be use as reference for identifying ‘species’ especially hybrids between H. agilis albibarbis and H. muelleri.
© Hak Cipta Milik Institut Pertanian Bogor, tahun 2007 Hak Cipta Dilindungi
KAJIAN HABITAT, TINGKAH LAKU, DAN POPULASI
KALAWET (Hylobates agilis albibarbis) DI TAMAN NASIONAL
SEBANGAU KALIMANTAN TENGAH
YULIUS DUMA
Disertasi
Sebagai salah satu syarat untuk memperoleh gelar Doktor pada
Program Studi Primatologi
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Judul Disertasi : Kajian Habitat, Tingkah Laku, dan Populasi Kalawet (Hylobates agilis albibarbis) di Taman Nasional Sebangau Kalimantan Tengah
Nama : Yulius Duma
NRP : P067020021
Disetujui
Komisi Pembimbing
Dr. Ir. Sri Supraptini Mansjoer Ketua
Dr. Ir. R.R. Dyah Perwitasari, M.Sc Prof. drh. Dondin Sajuthi, MST, Ph.D Anggota Anggota
Diketahui
Ketua Program Studi Primatologi Dekan Sekolah Pascasarjana
Prof. drh. Dondin Sajuthi, MST, Ph.D Prof. Dr. Ir. Khairil A. Notodiputro, MS
PRAKATA
Segala hormat, puji dan syukur penulis panjatkan ke Hadirat Tuhan Yang
Maha Esa, karena hanya atas perkenan dan tuntunan-Nya, disertasi yang berjudul
“Kajian Habitat, Tingkah Laku, dan Populasi Kalawet (Hylobates agilis
albibarbis) di Taman Nasional Sebangau Kalimantan Tengah” dapat diselesaikan.
Penulis menyadari bahwa penelitian dan penulisan disertasi ini dapat
terselesaikan, tidak terlepas dari keterlibatan dan dukungan berbagai pihak. Untuk
itu, penulis menyampaikan terima kasih dan penghargaan kepada Komisi
Pembimbing, Dr. Ir. Sri Supraptini Mansjoer, Dr. Ir. R.R. Dyah Perwitasari, M.Sc,
dan Prof. drh. Dondin Sajuthi, MST, Ph.D, atas segala curahan waktu, tenaga,
arahan, bimbingan dan bantuan yang diberikan sejak perencanaan penelitian
sampai penulisan disertasi ini. Kepada Prof. Dr. Ir. Hadi S. Alikodra, MS selaku
Penguji Luar Komisi pada Ujian Tertutup, Prof.Ris. Dr. M. Bismark dan Dr. Ir.
Rinekso Sukmadi, M.Sc.F, selaku Penguji Luar Komisi pada Ujian Terbuka, saya
menyampikan terima kasih dan penghargaan yang besar atas arahan dan
saran-saran yang diberikan. Ucapan terima kasih yang sama, juga penulis tujukan
kepada Dekan Fakultas Pertanian dan Rektor Universitas Tadulako, yang
memberikan izin tugas belajar; Direktur Proyek DUE-like Bath III UNTAD yang
memberikan beasiswa DUE-like; Rektor IPB, Dekan Sekolah Pascasarjana IPB,
dan Ketua PS. Primatologi IPB yang memberikan kesempatan kepada penulis
sebagai mahasiswa PS. Primatologi SPs IPB; Dirjen DIKTI DEPDIKNAS yang
memberikan dana penelitian melalui penelitian Hibah Pasca; Ir. Suwido H. Limin,
MS (Direktur CIMTROP Universitas Palangka Raya) dan staf, atas izin, fasilitas,
dan bantuan yang diberikan dalam melakukan penelitian di Laboratorium Alam
Hutan Gambut (LAHG); Aurelien Brule (Direktur Kalaweit Program, Kalteng)
dan staf, atas izin dan fasilitas yang diberikan untuk melakukan perekaman
vokalisasi Hylobates di Hampapak; Dr. Susan Cheyne yang memfasilitasi
perekaman vokalisasi Hylobates hibrida di Barito Ulu; Ir. Anthon Pawarrangan
(Pimpinan Cabang PT Dwimajaya Kalteng) dan staf yang memfasilitasi survei ke
Tumbang Manggu, Katingan, Kalimantan Tengah; Erna Shinta, SHut (Unit
dalam mengidentifikasi spesies tumbuhan; Armaiki Yusmur (BTIC BIOTROP)
yang membantu dalam analisis foto lansat; PEMDA Sulawesi Tengah yang
menyediakan fasilitas asrama di Bogor; Kepala PSSP LPPM-IPB dan staf, serta
seluruh staf pengajar dan administrasi PS. Primatologi IPB, atas berbagai bentuk
bantuan dan kerjasama selama mengikuti pendidikan di IPB; rekan-rekan
mahasiswa Primatologi (HIMAPRIMA), dan rekan-rekan mahasiswa pascasarjana
Sulawesi Tengah (HIMPAST) di Bogor, atas berbagai dukungan dan kerjasama
yang baik dalam kebersamaan di Bogor; dan terima kasih kepada Ketua
PS. Primatologi yang memberikan bantuan dana penyelesaian disertasi.
Kepada istriku terkasih Netty Tangaran, BSc dan anak-anakku tercinta Irene
Patricia Lystianti, Fanny Astria dan Olivia Christavila, penulis menyampaikan
terima kasih dan penghargaan yang besar, atas segala pengertian, pengorbanan
dan dukungan yang diberikan, serta senantiasa menyemangati penulis dalam
menyelesaikan disertasi ini. Kepada keluarga besar Tangaran-Manda, dan
Tikara-Todingpadang, serta Jemaat Gereja KIBAID Palu, penulis menyampikan banyak
terima kasih atas dukungan doa, moril, dan materil yang diberikan selama
megikuti pendidikan di IPB.
Kiranya disertasi ini bermanfaat bagi banyak pihak dan kepentingan.
Bogor, Mei 2007
RIWAYAT HIDUP
YULIUS DUMA, lahir di Rantepao, Tana Toraja, Sulawesi Selatan pada 26
Mei 1963 sebagai anak ke 3 dari 7 bersaudara, dari pasangan Ayahanda Yunus
Duma (Alm.) dan Ibunda Ludia Kombong (Almh.).
Penulis menyelesaikan pendidikan Sekolah Dasar pada tahun 1975 di SD
YPKT Mandetek, Sekolah Menengah Pertama pada tahun 1979 di SMP Katolik
Makale, dan Sekolah Menengah Atas pada tahun 1982 di SMAN 276 Makale,
Tana Toraja. Kemudian melanjutkan pendidikan di Jurusan Peternakan, Fakultas
Peternakan, Universitas Hasanuddin dan memperoleh gelar sarjana pada tahun
1987. Pada tahun 1995, penulis mendapat kesempatan mengikuti pendidikan S2 di
Program Studi Ilmu Ternak, Program Pascasarjana UGM, dan meraih gelar
Magister Pertanian pada tahun 1998. Sejak tahun 2002, penulis diterima sebagai
mahasiswa program S3 pada Program Studi Primatologi SPs IPB.
Penulis bekerja sebagai staf pengajar pada Jurusan Peternakan, Fakultas
Pertanian, Universitas Tadulako, Palu, Sulawesi Tengah, sejak tahun 1989.
Selama bekerja sebagai staf pengajar, penulis telah mengikuti beberapa kursus,
antara lain: Kursus Singkat Genetika, Kursus PEKERTI, Pemodelan Data
Bergerak, Biologi Molekuler, Bioteknologi Reproduksi, dan Field Course:
Primate Behavior and Ecology, di Pulau Tinjil (PSSP LPPM-IPB).
Penulis menikah dengan Netty Tangaran, BSc pada tahun 1987, dan telah
dikaruniai tiga orang putri, yaitu Irene Patricia Listyanti (mhs. FKG UNHAS),
DAFTAR ISI
Manfaat Penelitian ... 4
Kerangka Pemikiran ... 4
TINJAUAN PUSTAKA ... 5
Taksonomi dan Penyebaran ... 5
Morfologi ... 8
Habitat ... 10
Tingkah Laku ... 15
Populasi ... 22
Status Konservasi ... 24
Keadaan Umum Lokasi Penelitian ... 24
METODE PENELITIAN ... 30
Estimasi Populasi ... 36
Vokalisasi ... 41
Analisis Data ... 41
Analisis Vegetasi ... 41
AnalisisTingkah Laku ... 42
Estimasi Populasi ... 43
DAFTAR ISI lanjutan
Populasi... 84
Frekuensi Vokalisasi dan Jumlah Kelompok ... 84
Probabilitas Vokalisasi ... 87
Ukuran Kelompok... 89
Kepadatan Populasi... 90
Analisis Vokalisasi ... 94
Vokalisasi Betina ... 96
Vokalisasi Jantan ... 110
KONSERVASI ... 113
Habitat dan Kalawet ... 113
Perburuan ... 116
SIMPULAN DAN SARAN... 121
Simpulan... 121
Saran... 121
DAFTAR PUSTAKA ... 123
DAFTAR TABEL
Halaman
1 Estimasi densitas beberapa populasi Hylobates di Indonesia .... 23 2 Daftar peubah vokalisasi kalawet yang diamati ... 44 3 Profil vegetasi di LAHG CIMTROP di Taman Nasional
Sebangau ... 47 4 Sepuluh jenis pohon dengan indeks nilai penting (INP)
tertinggi di lokasi plot T0C di hutan gambut campuran (MSF). 53 5 Sepuluh jenis pohon dengan indeks nilai penting (INP) tertinggi di
lokasi plot T02 di hutan gambut campuran (MSF) ... 54
6 Sepuluh jenis pohon dengan indeks nilai penting (INP) tertinggi di
lokasi plot TA1 di hutan gambut campuran (MSF) ... 56
7 Sepuluh jenis pohon dengan indeks nilai penting (INP)
tertinggi di plot T05 di hutan tegakan rendah (LPF) ... 57 8 Sepuluh jenis pohon dengan indeks nilai penting (INP)
tertinggi di plot T08 tipe hutan tegakan rendah (LPF) ... 58 9 Sepuluh jenis pohon dengan indeks nilai penting (INP)
tertinggi di plot T14s di hutan tegakan tinggi (TIF) ... 60 10 Jenis tumbuhan yang dimakan oleh kalawet di LAHG ... 62 11 Profil potensi pohon sumber pakan kalawet di LAHG ... 64 12 Jenis, jumlah dan frekuensi penggunaan sebagai pohon tidur
oleh kelompok KC ... 68 13 Rata-rata panjang jelajah dan kecepatan jelajah harian kalawet
KC ... 82 14 Sebaran jumlah vokalisasi dan kelompok kalawet
teridentifikasi di setiap lokasi pengamatan ... 86 15 Probabilitas kelompok kalawet bervokalisasi selama waktu
pengamatan ... 88 16 Kepadatan kelompok kalawet pada tiap lokasi pengamatan... 91 17 Korelasi (r) antara variabel vegetasi dengan kepadatan
populasi kalawet ... 93 18 Spesies, lokasi, jumlah individu dan jumlah great call yang
analisis dalam penelitian ini ... 96 19 Nilai rata-rata peubah great call H. agilis albibarbis ... 98 20 Nilai rata-rata peubah great call Hylobates muelleri ... 102 21 Nilai rata-rata peubah great call Hibrida (Hylobates agilis
DAFTAR GAMBAR
Halaman
1 Skema kerangka pemikiran ... 4
2 Sebaran geografik spesies dari famili Hylobatidae ... 6
3 Sebaran geografik spesies dari genus Hylobates ... 7
4 Pohon pilogeni Hylobatidae berdasarkan data vokalisasi dan molekuler ... 8
5 Profil kalawet jantan (kiri) dan Betina (kanan) dewasa ... 10
6 Hubungan luas daerah jelajah dengan ukuran tubuh satwa primata ... 14
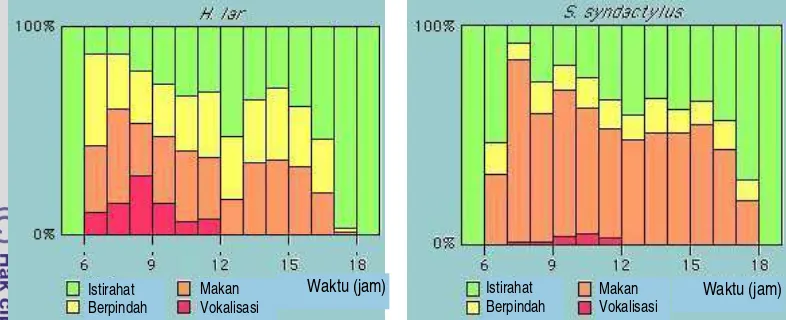
7 Perbandingan aktivitas harian H. lar dan S. syndactylus ... 18
8 Sonagram vokalisasi Hylobatidae (1) great call betina, (2) Jantan ... 20
9 Komposisi great call dari Hylobates ... 21
10 Taman Nasional Sebangau, Kalimantan Tengah ... 25
11 Profil topografi TN. Sebangau ... 26
12 Profil vegetasi di LAHG, TN. Sebangau ... 28
13 Lokasi penelitian ... 30
14 Ilustrasi tipe hutan di LAHG ... 31
15 Petak contoh bentuk garis berpetak ... 33
16 Sistem transek dan lokasi daerah jelajah KC di LAHG ... 35
17 Denah lokasi pengamatan vokalisasi dan vegetasi... 37
18 Ilustrasi peta pengamatan kepadatan kelompok ... 37
19 Sonagram great call H. a. Albibarbis ... 41
20 Contoh tampilan analisis sonagram great call menggunakan program Raven 1.2.1 ... 45
21 Jumlah jenis dan kerapatan tingkat pohon dan tiang di LAHG 48 22 Rata-rata dbh dan tinggi pohon kategori pohon dan tiang ... 51
23 Sampel pakan kalawet di LAHG ... 64
24 Sebaran lokasi pohon tidur dalam daerah jelajah kelompok KC 66 25 Profil pohon tidur kalawet (tanda panah menunjuk kalawet yang sedang vokalisasi di pohon tidur) ... 67
26 Daerah jelajah KC ... 71
27 Pola aktivitas harian kalawet KC ... 73
28 Sebaran pola aktivitas harian kalawet KC pada waktu (jam) yang berbeda sepanjang hari ... 74
29 Komposisi pakan kalawet KC ... 77
30 Sebaran frekuensi makan per komponen pakan pada waktu (jam) yang berbeda sepanjang hari ... 78
31 Arah dan jalur jelajah harian kalawet KC selama pengamatan.. 81
32 Variasi besar kelompok kalawet di LAHG ... 89
33 Peta tutupan lahan Taman Nasional Sebangau ... 92
34 Sonagram great callHylobates agilis albibarbis... 99
35 Sonagram great callH. muelleri ... 101
DAFTAR GAMBAR Lanjutan
36 Sonagram great call Hibrida (Hylobates agilis albibarbis x H. muelleri) ...
104
37 Gelombang suara great call Hibrida (Hylobates agilis albibarbis x H. muelleri) ...
105
DAFTAR LAMPIRAN
Halaman
1 Jenis tumbuhan dan nilai variabel vegetasi tingkat pohon, tiang, pancang dan semai, di plot T0C di LAHG CIMTROP
TN Sebangau ... 131 2 Jenis tumbuhan dan nilai variabel vegetasi tingkat pohon,
tiang, pancang dan semai, di plot T02 di LAHG CIMTROP
TN Sebangau ... 135 3 Jenis tumbuhan dan nilai variabel vegetasi tingkat pohon,
tiang, pancang dan semai, di plot TA1 di LAHG CIMTROP
TN Sebangau ... 140 4 Jenis tumbuhan dan nilai variabel vegetasi tingkat pohon,
tiang, pancang dan semai, di plot T05 di LAHG CIMTROP
TN Sebangau ... 144 5 Jenis tumbuhan dan nilai variabel vegetasi tingkat pohon,
tiang, pancang dan semai, di plot T08 di LAHG CIMTROP
TN Sebangau ... 148 6 Jenis tumbuhan dan nilai variabel vegetasi tingkat pohon,
tiang, pancang dan semai, di plot T14 di LAHG CIMTROP
TN Sebangau ... 151 7 Jumlah dan jenis tumbuhan tingkat pohon yang yang terdapat
di setiap lokasi ... 154 8 Jumlah dan jenis tumbuhan tingkat tiang yang yang terdapat di
setiap lokasi ... 156 9 Kepadatan populasi kalawet dan kondisi vegetasi tingkat
pohon di LAHG ... 158 10 Peta sebaran vokalisasi kelompok kalawet di setiap lokasi
Pengamatan ... 159 11 Rata-rata durasi kalawet KC berada di pohon tidur (PT),
aktivitas harian total, dan durasi aktivitas di luar pohon tidur,
selama 12 hari pengamatan full-day follow ... 162 12 Frekuensi aktivitas harian kalawet KC di LAHG TN Sebangau 163 13 Sebaran frekuensi makan per komponen pakan kalawet KC
pada waktu yang berbeda ... 164 14 Hasil analisis ragam peubah vokalisasi great call Hylobates di
Hylobates merupakan jenis kera kecil yang hidup di hutan tropis Asia
Selatan dan Tenggara, di antaranya Thailand, Malaysia dan Indonesia (Fleagle
1988). Indonesia sendiri mempunyai beberapa spesies Hylobates, yaitu H. lar dan
H. agilis di Sumatera, H. klossii di Pulau Mentawai, H. moloch di Jawa, H. agilis
dan H. muelleri di Kalimantan. H. agilis terdiri dari tiga subspesies, yaitu H. agilis
agilis yang menyebar di dataran tinggi Sumatera bagian barat, H. agilis ungko di
dataran rendah Sumatera bagian timur, dan H. agilis albibarbis di Kalimantan
Barat dan Tengah (Marshall & Sugardjito 1986; Supriatna & Wahyono 2000).
Selain itu, di daerah Barito Ulu, sebagai perbatasan daerah sebaran antara H.
agilis albibarbis dan H. muelleri, telah terjadi persilangan di antara kedua spesies
tersebut (McConkey 2003), yang kemungkinan bakal menjadi spesies tersendiri.
Sebagian besar genus Hylobates khususnya yang ada di Indonesia kini
diperkirakan dalam keadaan terancam yang disebabkan terutama oleh rusak dan
hilangnya hutan sebagai habitatnya, serta perburuan dan perdagangan ilegal.
Populasi Hylobates, khususnya H. agilis albibarbis dan H. muelleri yang ada di
Kalimantan, diperkirakan menurun dari tahun ke tahun seiring dengan penyusutan
hutan habitatnya serta perburuan dan perdagangan satwa ini secara ilegal.
Berdasarkan data Departemen Kehutanan (1997), telah terjadi deforestasi di
kawasan hutan Kalimantan sebesar 25,9% selama tahun 1985–1997. Aktivitas
illegal logging yang terus terjadi, mengindikasikan deforestasi terus berlangsung
dari tahun ke tahun sampai saat ini. Terjadinya deforestasi berarti hilangnya
habitat bagi banyak satwa liar, termasuk satwa primata di Kalimantan. Kerusakan
dan hilangnya habitat akan berdampak negatif terhadap populasi satwa primata.
Buckley (2004) melaporkan kepadatan populasi kalawet (H. agilis albibarbis) di
hutan gambut Sebangau (bekas konsesi HPH) Kalimantan Tengah, sebesar 7,4
ekor/km2, jauh lebih rendah dibanding hasil penelitian Mitani (1990) di Taman Nasional Gunung Palung, Kalimantan Barat, sebesar 13,5–15,6 ekor/km2.
Suatu keuntungan bagi konservasi kalawet (H. agilis albibarbis) bahwa di
daerah sebarannya, kini terdapat empat taman nasional, yaitu TN. Gunung Palung,
PHKA 2006b). Akan tetapi, perambahan dan penebangan liar tetap terjadi dalam
kawasan taman nasional tersebut (Alikodra & Syaukani 2004).
Kehilangan dan rusaknya hutan khususnya di Kalimantan merupakan
pembunuhan tidak langsung terhadap satwa yang hidup di dalamnya termasuk
H. agilis albibarbis dan H. muelleri, serta silangannya di daerah Barito Ulu.
Eksploitasi perusahaan HPH, penebangan liar, konversi hutan dan musibah
kebakaran hutan yang terjadi di Kalimantan selama ini semakin mempersempit
habitat dan fragmentasi habitat Hylobates. Habitat yang semakin sempit dibarengi
dengan fragmentasi habitat yang tinggi semakin memperkecil ruang ekologis bagi
Hylobates dalam mendapatkan teritori tempat tinggalnya. Itu berarti terjadi
persaingan yang tinggi baik intra maupun inter spesies untuk mendapatkan teritori
dan pakan.
Kondisi demikian diperkirakan berdampak pada aspek biologis dan
populasi kalawet. Sifat Hylobates yang arboreal pada kanopi tengah dan atas, dan
lokomosi secara brakhiasi membuatnya sulit untuk mencari dan mengembangkan
wilayah jelajahnya pada kondisi kualitas habitat yang rendah dan terfragmentasi.
Daerah jelajah tiap kelompok menjadi sempit, dan ketersediaan pakan menjadi
terbatas sehingga akan berpengaruh terhadap populasi dan biologis kalawet.
Fragmentasi habitat diperkirakan berpengaruh pula terhadap luas jangkauan
kalawet muda meninggalkan kelompok keluarga asalnya untuk mencari teritori
dan pasangan kawinnya membentuk kelompok baru. Hal ini berarti, peluang
terjadinya inbreeding semakin besar, selanjutnya akan mempengaruhi tatanan
genetik populasi.
Informasi ekologi kalawet (H. agilis albibarbis) yang ada kini masih
sangat terbatas, khususnya tentang kondisi habitat, kepadatan populasi, dan
aktivitas kalawet dalam memanfaatkan habitatnya. Informasi-informasi tersebut
sangat dibutuhkan sebagai dasar pertimbangan dalam upaya-upaya konservasi
kalawet ke depan, dalam rangka mengantisipasi ancaman kepunahan. Satwa
primata tersebut dikategorikan oleh IUCN (International Union for Conservation
of Nature and Natural Resources) sebagai spesies yang masih berisiko rendah
Selain kerusakan dan hilangnya sebagian habitat, kegiatan perburuan juga
merupakan ancaman bagi populasi kalawet (H. agilis albibarbis) dan H. muelleri
serta silangannya di Kalimantan. Satwa primata ini tergolong spesies yang
dilindungi, dan terdaftar dalam Appendix I oleh CITES (Convention on
International Trade in Endangered Species of Wild Fauna and Flora) sebagai
satwa yang tidak boleh diperdagangkan (Soehartono & Mardiastuti 2002); namun
tetap diburu oleh masyarakat untuk diperdagangkan dan dipelihara sebagai pet
animal. Sebagai satwa yang dilindungi, terdapat upaya-upaya merehabilitasi
kalawet dan Hylobates lainnya yang dipelihara oleh masyarakat untuk
dikembalikan ke alam hidupan liarnya.
Fenotipe ketiga jenis Hylobates yang ada di Kalimantan (H. agilis
albibarbis, H. muelleri, dan hibrida antara H. agilis albibarbis x H. muelleri)
hampir sama, sehingga cukup sulit untuk mengidentifikasi dan memisahkannya
dalam program rehabilitasi dan reintroduksi. Hal ini dapat diatasi melalui analisis
vokalisasi, sebagaimana dinyatakan Mitani (1987) bahwa variasi vokalisasi
Hylobates mengindikasikan perbedaan genetik minimal pada level spesies
sehingga dapat digunakan untuk membedakan spesies.
Penelitian ini dirancang untuk mendapatkan informasi mutahir tentang
aspek ekologi (habitat, tingkah laku, dan populasi) kalawet, di TN. Sebangau,
Kalimantan Tengah, dan vokalisasi sebagai penciri spesies Hylobates di
Kalimantan.
Tujuan
1. Mengkaji kondisi habitat kalawet di TN Sebangau.
2. Mengkaji pola tingkah laku kalawet, meliputi aktivitas harian, jelajah harian,
vokalisasi, dan komposisi jenis pakan.
3. Mendapatkan informasi populasi kalawet terkini di kawasan hutan rawa
gambut TN Sebangau, melalui observasi vokalisasi.
4. Mengkaji karakteristik suara Hylobates untuk dijadikan salah satu penciri
Manfaat Penelitian
1. Membantu mengidentifikasi spesies Hylobates melalui analisis suara yang
sangat berguna untuk program rehabilitasi dan reintroduksi Hylobates di
Kalimantan.
2. Hasil penelitian merupakan informasi mutahir yang dapat dijadikan dasar
pertimbangan dalam merumuskan kebijakan konservasi populasi kalawet
beserta habitat.
Kerangka Pemikiran
Gambar 1 Skema kerangka pemikiran
TINJAUAN PUSTAKA
Taksonomi dan Penyebaran Hylobates
Hylobates merupakan salah satu genus dari famili Hylobatidae, ordo
Primates. Genus Hylobates terdiri dari enam spesies, dan beberapa subspesies.
Salah satu subspesies dari spesies H. agilis adalah H. agilis albibarbis yang
terdapat di Kalimantan dengan klasifikasi selengkapnya sebagai berikut:
Kingdom : Animalia
Filum : Chordata
Klas : Mammalia
Ordo : Primates
Famili : Hylobatidae
Genus : Hylobates
Spesies : Hylobates agilis
Subspesies : H. agilis albibarbis
Nama lokal : Kalawet, Owa-owa
(Napier & Napier 1985; Geissmann 2003; Brandon-Jones et al. 2004).
Hylobatidae terdiri dari empat genus dan 12 spesies yang hidup menyebar di
kawasan hutan tropik Asia Tenggara, yaitu (1) Genus Hylobates Illiger, 1811
terdiri dari enam spesies: (a) H. klossii di Kepulauan Mentawai; (b) H. lar
menyebar di Sumatera bagian utara, Semenanjung Malaysia, Thailand, Myanmar
dan Yunnan (Smith 1999); (c) H. moloch di pulau Jawa; (d) H. agilis di Sumatera,
Kalimantan, dan Malaysia; (e) H. muelleri di Kalimantan; dan (f) H. pileatus di
Thailand dan Kamboja; (2) Genus Nomascus Miller, 1933: spesies N. concolor,
N. gabriellae, N. leucogenys, dan N. nasutus, menyebar di China, Hainan, Laos,
Vietnam dan Kamboja; (3) Genus Symphalangus Glonger, 1841: spesies
S. syndactylus di Sumatera dan Malaysia; (4) Genus Bunopithecus Matthew dan
Granger, 1923: spesies B. hoolock di Assam, Bangladesh, dan Myanmar
(Geissmann 2003; Brandon-Jones et al. 2004). Untuk jelasnya, sebaran geografik
spesies dari famili Hylobatidae, dan genus Hylobates tersebut dapat dilihat pada
Gambar 2 Sebaran geografik spesies dari famili Hylobatidae
Kini, spesies B. hoolock dianggap tidak tepat menggunakan nama
Bunopithecus, karena ternyata karakter dental dari spesimen fragmen rahang
Bunopithecus sangat berbeda dengan karakter dental Hylobates yang ada saat ini.
Bunopithecus dianggap sebagai genus dari Hylobatidae yang sudah punah. Oleh
karena itu, nama genus dari spesies B. hoolock diganti dengan Hoolock, sehingga
nama spesiesnya menjadi Hoolock hoolock (Mootnick & Groves 2005;
Gambar 3 Sebaran geografik spesies dari genus Hylobates
Berdasarkan data vokalisasi dan molekuler, Geissmann (2006a) menyusun
pohon filogenetik Hylobatidae, dilengkapi dengan vernacular names
masing-masing spesies, seperti pada Gambar 4. Jumlah kromosom diploid antar genus
bervariasi, masing-masing genus Hylobates 44, Hoolock 38, Nomascus 52, dan
genus Symphalangus 50 (Geissmann 2006a).
H. agilis terdiri dari tiga subspesies, yaitu (a) H. agilis agilis yang menyebar
di dataran tinggi Sumatera bagian barat dan semenanjung Malaysia, (b) H. agilis
Gambar 4 Pohon filogeni Hylobatidae berdasarkan data vokalisasi dan molekuler (Geissmann 2006a)
di Kalimantan Barat dan Tengah (Marshall & Sugardjito 1986, Supriatna &
Wahyono 2000, GCC 2004, Geissmann 2006a). Penyebaran H. agilis albibarbis
di Kalimantan Tengah ke arah timur dibatasi oleh Sungai Barito dan ke utara
dibatasi Sungai Kapuas (Supriatna & Wahyono 2000).
H. muelleri menyebar hampir di seluruh pulau Kalimantan, kecuali di
daerah barat Sungai Barito dan selatan Sungai Kapuas, sebagai daerah penyebaran
H. agilis albibarbis seperti pada Gambar 3 (Geissmann 1995). H. muelleri juga
terdiri dari tiga subspesies, yaitu (a) H. muelleri muelleri di Kalimantan Timur
dan Selatan, (b) H. muelleri abbotti di Kalimantan Barat dan sebelah utara Sungai
Kapuas, (c) H. muelleri funereus di daerah Serawak (Geissmann 2006a).
Di daerah hulu Sungai Barito (Barito Ulu), sebagai batas daerah sebaran
H. agilis albibarbis dengan H. muelleri dilaporkan telah terjadi hibrida di antara
keduanya yang diketahui sejak tahun 1979 oleh Dr. Marshall (Marshall &
Sugardjito 1986, McConkey et al. 2003). Selain di Barito Ulu, hibrida juga
terjadi di semenanjung Malaysia antara H. agilis dengan H. lar, dan di Thailand
antara H. lar dengan H. pileatus (Brockelman & Gittins 1984).
Morfologi
Hylobates sering disebut sebagai kera kecil (small apes) karena postur
tubuhnya yang kecil dengan bobot badan sekitar 4-8 kg. H. agilis memiliki bobot
badan rata-rata berkisar 4-7 kg dengan panjang badan antara 45-55 cm,
H. muelleri 5-6.4 kg dan panjang badan 42-47 cm, H. moloch 4-8 kg dan panjang
badan 75-80 cm (Supriatna & Wahyono 2000), H. lar dan H. pileatus 5-7 kg
dengan panjang badan 45-65 cm (Smith 1999, Sullivan 2004). Pada umumnya
jantan sedikit lebih besar dari pada betina, namun Fleagle (1988) tidak
mengkategorikan perbedaan ukuran tubuh tersebut sebagai dimorfisme seksual.
Bobot badan yang ringan dibutuhkan untuk menunjang sifatnya yang arboreal.
Warna rambut H. agilis mulai dari coklat muda kemerahan sampai hitam,
terutama sekitar pergelangan tangan dan kaki sampai jari-jari berwarna hitam dan
umumnya lebih gelap dari warna bagian tubuh lainnya, sehingga sering juga
disebut black/dark-handed gibbon. Rambut H. agilis Sumatera pada umumnya
berwarna hitam, sedangkan H. agilis Kalimantan bervariasi dari coklat muda
sampai coklat hitam dan bagian ventral lebih gelap daripada dorsal (Groves 2001,
GCC 2004). Pada umumnya agilis memiliki garis lengkung putih pada alis (white
brow) sampai pipi (cheek patches). Garis lengkung putih ini lebih kontras di
sekeliling wajah pada jantan daripada betina, bahkan garis putih di pipi pada
betina berangsur hilang menjelang dewasa pada umur sekitar enam tahun.
Kalawet betina dewasa yang dipelihara pada kondisi cahaya yang kurang atau
defisiensi nutrisi dapat kehilangan garis putih pada alis (GCC 2004). Profil
kalawet jantan dan betina, serta H. muelleri dapat dilihat pada Gambar 5.
H. agilis mempunyai lengan yang lebih panjang dari kaki (indeks
intermembralnya >100) dilengkapi dengan empat jari yang panjang dan ibu jari
yang lebih kecil. Kondisi ini memungkinkan H. agilis dapat bergerak secara
brakhiasi/berayun dengan lincah dari pohon ke pohon menggunakan tangannya
seperti menggantung pada dahan tanpa menggenggam. Kaki sebagai salah satu
alat gerak kurang difungsikan pada Hylobates sehingga menjadi lebih pendek,
tidak berkembang sebaik lengan yang berfungsi sebagai alat gerak utama dalam
brakhiasi (Fleagle 1988, Kuester 2000). Metode lokomosi demikian
memungkinkan H. agilis dapat berpindah sejauh tiga meter sekali berayun. Selain
itu, H. agilis juga dapat berpindah dengan cara memanjat, melompat, dan berjalan
bipedal sambil mengangkat kedua lengannya (Napier & Napier 1967, Fleagle
Kalawet jantan dewasa Kalawet betina dewasa
Kalawet betina dewasa Kalawet betina muda H. muelleri betina dewasa
H. muelleri remaja
Gambar 5 Profil kalawet (H. agilis albibarbis) dan H. muelleri (a,bLAHG Sebangau, c,eHampapak, dPalangka Raya, fNijman 2005b)
Habitat
Vegetasi
Suatu populasi menempati suatu wilayah tertentu yang dapat memenuhi
segala kebutuhannya, khususnya kebutuhan dasar populasi yaitu untuk
berlindung, berkembangbiak, makanan dan air serta untuk pergerakan. Suatu
kawasan yang dapat memenuhi kebutuhan dasar populasi tersebut dikenal sebagai
habitat (Alikodra 2002).
H. agilis yang ada di Indonesia menempati habitat hutan dataran tinggi di
Sumatera (H. agilis agilis) dan hutan dataran rendah (H. agilis ungko) di Sumatera
dan (H. agilis albibarbis) di Kalimantan Tengah dan Barat (Supriatna & Wahyono
a b
2000). Dataran rendah di Kalimantan berupa hutan rawa dan hutan dataran rendah
kering terbentang sangat luas mencapai ratusan kilometer ke arah hulu atau
pedalaman. Hutan rawa dibedakan dalam tiga kelompok utama, yaitu hutan bakau
yang dipengaruhi air laut, hutan rawa air tawar yang mendapat air dari sungai, dan
hutan rawa gambut tadah hujan, sedangkan hutan dataran rendah kering
merupakan hutan Dipterocarpaceae (MacKinnon et al. 2000).
Hutan dataran rendah Kalimantan, kecuali hutan bakau, antara Sungai
Kapuas dan Sungai Barito merupakan habitat kalawet (H. agilis albibarbis),
sedangkan H. muelleri yang endemik Kalimantan tersebar di bagian lainnya dari
pulau Kalimantan (MacKinnon et al. 2000, Supriatna & Wahyono 2000). Selain
Hylobates, Pulau Kalimantan juga dihuni oleh beberapa jenis satwa primata
lainnya, seperti orangutan (Pongo pygmaeus), bekantan (Nasalis larvatus),
beberapa spesies Presbytis, di antaranya P. baricunda, P. cristata, P. frontata, P.
hosei, dan P. melalophos); Macaca nemestrina dan M. fascicularis.
Dalam kawasan habitat penyebaran kalawet (H. agilis albibarbis), sampai
saat ini terdapat empat taman nasional (TN), yaitu TN Gunung Palung seluas
90.000 ha di Kalimantan Barat, TN Bukit Baka-Bukit Raya (181.090 ha) di
Kalimantan Barat dan Tengah, TN Tanjung Puting (415.040 ha), dan TN
Sebangau (568.700 ha) di Kalimantan Tengah. Selain itu, juga terdapat beberapa
cagar alam (CA), antara lain: CA Bukit Tangkiling (2.061 ha), CA Pararawen I/II
(5.855 ha), dan CA Bukit Sapat Hawung (239.000 ha) (Ditjen PHKA 2006b).
Keberadaan taman nasional dan cagar alam tersebut sebagai kawasan konservasi
dalam habitat penyebaran kalawet, merupakan hal yang menguntungkan dalam
upaya konservasi kalawet. Namun demikian, kawasan konservasi tersebut, tidak
terlepas dari perambahan dan penebangan liar (Alikodra & Syaukani 2004;
Simbolon & Mirmanto 2000), terlebih lagi kawasan hutan yang tidak termasuk
kawasan konservasi.
Alikodra & Syaukani (2004) menyatakan bahwa deforestasi (penghancuran
hutan) di Indonesia saat ini sudah berada pada titik yang sangat membahayakan.
Tingkat deforestasi pada zaman Orde Baru berkisar 0,8-1,0 juta hektar per tahun,
dan kini di era reformasi, meningkat menjadi 1,6-2,5 juta hektar per tahun.
kawasan hutan menjadi perkebunan terutama kelapa sawit, dan kebakaran (Rieley
& Ahmad-shah 1996, Boehm et al. 2003, Drasospolino 2004). Data Departemen
Kehutanan (1997), menunjukkan telah terjadi deforestasi di kawasan hutan
Kalimantan sebesar 25,9% selama tahun 1985–1997. Deforestasi ini terus
berlangsung dari tahun ke tahun sampai saat ini.
Hampir seluruh kawasan hutan di Kalimantan Tengah, sebelumnya telah
dimanfaatkan sebagai penghasil kayu dengan sistem tebang pilih. Akibatnya,
hutan rawa gambut yang tersisa saat ini, termasuk kawasan yang dilindungi,
sebelumnya pernah ditebangi minimal sekali (Simbolon & Mirmanto 2000).
Selanjutnya, Simbolon & Mirmanto (2000) menyatakan bahwa hutan rawa
gambut dalam kawasan TN Tanjung Puting, masih terjadi penebangan liar secara
intensif, membuat hutan tersebut semakin rusak dan mendorong terjadinya
kebakaran. Hal yang sama juga terjadi di TN Gunung Palung. Alikodra &
Syaukani (2004) menyatakan bahwa sekitar 80% kawasan TN Gunung Palung
rusak parah akibat penebangan liar yang didorong oleh dibukanya ekspor kayu
bulat di era reformasi.
Deforestasi yang cukup tinggi juga terjadi di kawasan hutan rawa gambut
Sebangau, antara Sungai Sebangau dan Sungai Katingan, dan kawasan proyek
lahan gambut sejuta hektar (MRP mega rice project). Berdasarkan analisis foto
lansat, Boehm et al. (2003) mendapatkan data deforestasi di kawasan tersebut
sebesar 33% dalam jangka waktu 10 tahun, antara tahun 1991 – 2001. Itu berarti,
deforestasi di kawasan tersebut rata-rata 3,3%/tahun. Deforestasi antara tahun
1991-1997 berkisar 1,9%/tahun, meningkat menjadi 6,5%/tahun antara tahun
1997-2000. Peningkatan luas kawasan hutan rawa gambut yang mengalami
kerusakan tersebut, disebabkan oleh kebakaran hutan pada tahun 1997, aktivitas
penebangan liar, dan aktivitas mega rice project.
Pada kondisi normal, vegetasi hutan rawa air tawar lebih kaya akan jenis
flora dan tinggi tajuk dari pada hutan rawa gambut. Hal ini memberi gambaran
bahwa hutan rawa air tawar lebih kaya unsur hara karena disuplai oleh aliran
sungai, dibandingkan dengan rawa gambut yang hanya tadah hujan (Mackinnon
et al. 2000). Hal ini berarti, semakin jauh dari sungai, semakin miskin hara, dan
Analisis vegetasi pada hutan rawa gambut di Sebangau sebagai kawasan
hutan bekas konsesi perusahaan HPH telah dilakukan oleh Shepherd et al. (1997).
Kerapatan pohon (dbh diameter at breast high ≥ 7 cm) di daerah marginal ±2 km
dari sungai, 1.693 pohon/ha, daerah peralihan 6,5 km dari sungai, 2.500 pohon/ha,
dan daerah interior 18 km dari sungai, 1.347 pohon/ha. Tinggi pohon dan
diameter batang (dbh) yang lebih besar, lebih banyak pada daerah interior dari
pada daerah peralihan dan marginal.
Struktur fisik hutan terbentuk oleh adanya perbedaan tinggi pohon menurut
jenis, umur dan sifat tumbuhnya. Kondisi ini membentuk stratifikasi menjadi
relung ekologi tertentu bagi suatu jenis satwa, seperti adanya perbedaan
ketinggian tempat makan dalam suatu habitat bagi beberapa primata simpatrik.
Untuk analisis struktur fisik vegetasi hutan, Soerianegara & Indrawan (1998)
membedakan stadium tumbuh vegetasi, sebagai berikut:
a. semai (seedling) mulai dari kecambah sampai setinggi 1,5 m,
b. pancang (sapling) tumbuhan berkayu yang tingginya lebih dari 1,5 m dan
diameter kurang dari 10 cm,
c. tiang (pole) tumbuhan berkayu dengan diameter 10 – <20 cm, dan
d. pohon dewasa yang berdiameter ≥20 cm.
Daerah Jelajah
Pada umumnya suatu kelompok keluarga Hylobates menempati suatu
wilayah teritori tertentu dalam habitatnya yang cenderung dipertahankan secara
ketat dari gangguan individu atau kelompok lainnya (Leighton 1987, Mitani
1987, Nowak 1997, Geissmann 2003). Wilayah teritorinya ditandai dengan
vokalisasi, khususnya pada pagi hari melalui duet call sebagai tanda
keberadaannya kepada kelompok-kelompok tetangganya (Leighton 1987, Smith
1999).
Selain teritori, satwa primata dalam memanfaatkan habitat, juga dikenal
adanya daerah jelajah (home range) sebagai area habitat yang digunakan untuk
seluruh aktivitas hidup suatu kelompok satwa primata; area inti (core area)
merupakan area yang paling banyak/sering di tempati dalam melakukan sebagian
kelompok melakukan aktivitasnya dalam satu hari (NRC. 1981, Fleagle 1988,
Collinge 1993, Rowe 1996).
Perilaku menjelajah satwa primata sangat terkait dengan kebutuhan pakan
(Oates 1986). Spesies yang folivorous cenderung mempunyai daerah jelajah yang
lebih sempit karena ketersediaan dedaunan lebih bersifat umum dan merata,
dibandingkan dengan spesies yang frugivorous, dimana ketersediaan buah lebih
terbatas; dan spesies dengan ukuran tubuh yang besar cenderung membutuhkan
daerah jelajah yang lebih luas untuk mendukung kebutuhan hidupnya,
dibandingkan dengan ukuran tubuh yang lebih kecil, seperti diilustrasikan pada
Gambar 6 (Fleagle 1988).
Gambar 6 Hubungan luas daerah jelajah dengan ukuran tubuh satwa primata (simbol tertutup = folivorous, simbol terbuka = frugivorous, segi tiga = nokturnal, segi empat = diurnal terestrial, dan bulat = diurnal arboreal) (Fleagle 1988).
Itu sebabnya, luas daerah jelajah di antara spesies dan kelompok Hylobates
cukup bervariasi. Chivers (2001) merangkum luas daerah jelajah beberapa spesies
Hylobates sebagai berikut: H. hoolock 38 ha, H. lar 41 ha, H. concolor 46 ha,
H. klossii 32 ha, H. pileatus 36 ha, H. muelleri 44 ha, H. agilis 29 ha, hibrida
H. agilis x H. muelleri 18 dan 34 ha, H. moloch 17 ha, dan siamang 31 ha. Ahsan
(2001) mendapatkan variasi luas daerah jelajah di antara tiga kelompok
H. hoolock yang diamati, masing-masing 40,7 ha, 86 ha, dan 25,7 ha. Selain itu,
Ahsan (2001) juga mencatat bahwa H. hoolock lebih aktif berpindah dengan
panjang jelajah yang lebih tinggi pada pagi hari.
Rowe (1996) menyatakan bahwa daerah jelajah satwa primata dapat berubah
dari tahun ke tahun, tergantung perubahan iklim, ketersediaan sumber pakan dan
air, persaingan antar kelompok dalam spesies yang sama, perburuan dan degradasi
habitat. Hal ini didukung oleh Collinge (1993) bahwa luas daerah jelajah dapat
berubah tergantung pada ketersediaan sumber pakan dan air, dan tempat
berlindung. Hasil penelitian Iskandar (2007) pada owa Jawa (H. moloch)
membuktikan bahwa daerah jelajah dipengaruhi oleh perubahan iklim dan tipe
hutan. Luas daerah jelajah H. moloch di Taman Nasional Gunung Halimun-Salak
pada musim hujan di hutan primer sebesar 16,58 ha, sedikit lebih sempit,
dibandingkan pada hutan sekunder sebesar 17,61 ha. Demikian halnya pada
musim kemarau, luas daerah jelajah di hutan primer 18,91 ha, lebih sempit
dibandingkan pada hutan sekunder sebesar 21,13 ha. Selanjutnya Iskandar (2007)
menjelaskan bahwa daerah jelajah owa Jawa yang lebih sempit pada musim hujan
dan di hutan primer dibandingkan pada musim kemarau dan di hutan sekunder,
disebabkan oleh ketersediaan sumber pakan berupa buah yang lebih banyak pada
musim hujan dan pada hutan primer. Hal ini berarti, semakin melimpah
ketersediaan pakan, semakin sempit daerah jelajah, dan sebaliknya.
Chivers (2001) merangkum beberapa hasil penelitian dan menyimpulkan
bahwa Hylobates mempertahankan sekitar 80-90% dari daerah jelajahnya sebagai
teritori. Hylobates menempati dan menetap pada daerah teritori dengan luas antara
20-40 ha (Geissmann 2003), daerah jelajah siamang 47 ha dan H. Lar 57 ha
(Raemaekers 1979), H. muelleri 33-43 ha (Leighton 1987), hibrida H. Muelleri
dengan H. agilis albibarbis 43-46 ha (McConkey 1999 dalam McConkey et al.
2003), H. klossii 33 ha (Whitten 1980 dalam Bismark 2006). Teritori tersebut
dipertahankan secara ketat dan tidak akan pindah ke wilayah lain ketika wilayah
tersebut mengalami gangguan (Shneider 1995 dalam Geissmann 2003). Perilaku
demikian menyebabkan Hylobates mudah terancam ketika habitatnya terganggu.
Tingkah Laku
Struktur sosial
Hylobates merupakan jenis kera kecil yang hidup di hutan hujan tropis Asia
1998) secara arboreal pada kanopi tengah dan atas. Mereka hidup secara
monogami dalam kelompok kecil sebagai kelompok keluarga (family group) yang
biasanya terdiri dari dua sampai enam individu, sepasang jantan dan betina
dewasa, dan beberapa anak pada umur yang berbeda (Leighton 1987, Fleagle
1988, Reichard 1998, Geissmann 2003, Sullivan 2004, Nijman 2004).
Kelompok umur Hylobates dalam satu kelompok keluarga dapat dibedakan
atas: (a) bayi atau infant dari lahir sampai 2 tahun, (b) anak-anak atau juvenile,
umur 2-4 tahun, (c) remaja atau adolescent, umur 4-6 tahun, (d) muda atau
sub-adult, umur enam tahun atau lebih tetapi belum kawin, dan (e) dewasa umur di
atas enam tahun saat mencapai dewasa kelamin (Leighton 1987, Sullivan 2004).
Jantan dan betina muda menjelang dewasa atau mencapai dewasa kelamin akan
meninggalkan kelompoknya dan hidup mandiri dengan pasangannya sebagai
kelompok keluarga yang baru (Kirkwood & Stathatos 1992).
Brockleman et al. (1998) dalam penelitiannya pada H. lar, mendapati
anggota kelompok subadult jantan (F1) mulai melakukan solo songs di sekitar
batas teritori kelompoknya (F) sebagai perilaku khas Hylobates. Dua tahun
kemudian, jantan muda (F1) tersebut meninggalkan kelompoknya dan menjadi
pasangan kawin dengan induk di kelompok A menggantikan jantan A yang
kemungkinan dikalahkan oleh F1.
Walaupun Hylobates pada umumnya hidup monogami, Fuentes (2000)
mendapatkan adanya individu yang kawin dengan betina bukan pasangannya di
alam. Hal ini merupakan upaya untuk menjamin keberhasilan reproduksi dalam
rangka mempertahankan eksistensi genetiknya (genetic fitness) di bawah tekanan
stres lingkungan seperti terbatasnya ketersediaan pakan.
Aktivitas Harian
Waktu aktivitas Hylobates sedikit berbeda dengan jenis primata diurnal
lainnya. Hylobates mulai beraktivitas sebelum matahari terbit, tetapi lebih cepat
istirahat di pohon tidur pada sore hari (Chivers, 2001). Aktivitas harian utama
satwa primata meliputi aktivitas makan, melakukan perjalanan/berpindah,
istirahat, bersuara (Chivers 1973, MacKinnon & MacKinnon 1980). Selain itu,
waktu istirahat. Chivers (2001) menyatakan bahwa aktivitas harian Hylobates
berlangsung selama kurang lebih 9,5 jam dalam sehari, dari pukul 06:19 pagi
sampai pukul 15:43 sore hari.
Chivers (2001) merangkum aktivitas harian beberapa spesies Hylobates,
dengan pola sebagai berikut: 38% alokasi waktu untuk makan, berpindah 12%,
istirahat 42% (termasuk aktivitas menelisik 3%), dan 8% untuk vokalisasi.
Aktivitas harian H. agilis rata-rata selama sembilan jam sehari yang dimulai pada
pukul 06.15-07.30 pada pagi hari sampai pukul 13.10-17.40, dengan aktivitas
utama adalah makan, berpindah, bersuara, dan istirahat (Gittins & Raemaekers
1980). H. lar aktif rata-rata selama 8-9 jam dan berada di pohon tidur rata-rata
selama 15 jam per hari (Reichard 1998).
Aktivitas bersuara pada pagi hari merupakan awal aktivitas harian kelompok
Hylobates yang berfungsi untuk menunjukkan teritorialnya sekaligus sebagai
pengaturan ruang antar kelompok (Bates 1970). Aktivitas bersuara pada ungko
sebagian besar dilakukan pada pagi hari yang dapat terdengar sampai 1 km
(Gittins & Raemaekers 1980), bahkan terdengar sampai 2 km (O’Brien et al.
2004).
Setelah turun dari pohon tidur, kelompok Hylobates memulai aktivitas
makannya pada pohon yang sedang berbuah. Lamanya kegiatan makan pada suatu
pohon bervariasi bergantung pada jenis dan kelimpahan pakan. Aktivitas makan
ungko cenderung lebih banyak pada ketinggian tajuk menengah, 15-25 m, karena
pada kanopi tengah ini merupakan jenis pohon yang menyediakan lebih banyak
buah. Sebaliknya, tajuk yang tinggi atau kanopi atas pada umumnya merupakan
jenis-jenis Dipterocarpaceae yang menghasilkan lebih banyak daun (Gittins &
Raemaekers 1980).
Pola aktivitas harian Hylobatidae dalam memanfaatkan daerah jelajahnya
bervariasi di antara spesies, dan terkait pula dengan sebaran dan jumlah sumber
pakan, dan sebaran pohon tidur. Hal ini dicontohkan oleh MacKinnon &
MacKinnon (1980) bahwa H. lar menggunakan waktu untuk makan lebih sedikit
Gambar 7 Perbandingan aktivitas harian H. lar dan S. syndactylus
Palombit (1997) dalam penelitiannya di TN Gunung Leuser, Sumatera
Utara, mendapatkan aktivitas harian yang sedikit berbeda antara H. lar dengan
S. syndactylus. H. lar menggunakan waktu untuk aktivitas makan sebesar 34%,
berpindah 16%, istirahat 45%, vokalisasi 3% dan interaksi antar kelompok 2%;
sedangkan S. syndactylus menggunakan waktu untuk aktivitas makan sebesar
40%, berpindah 12%, istirahat 44%, vokalisasi 1%, dan interaksi antar kelompok
3%.
Selain pola aktivitas harian, pola makan di antara spesies Hylobatidae juga
cukup bervariasi, seperti dirangkum oleh Conklin-Brittain et al. (2001) sebagai
berikut: H. hoolock, H. agilis, H. klossii, H. lar dan H. pileatus, rata-rata 72%
buah, 15% daun, 6% bunga, dan 7% insekta; H. moloch, dan H. muelleri, rata-rata
60% buah, 37% daun, 1% bunga, dan 2% insekta; dan S. syndactylus (siamang)
40% buah, 49% daun, 6% bunga, dan 5% insekta, serta N. concolor 21% buah,
71% daun, dan 7% bunga. S. syndactylus dan N. concolor cenderung
mengkonsumsi lebih banyak daun dibandingkan dengan spesies lainnya. Kedua
spesies Hylobatidae ini lebih banyak hidup pada kawasan hutan dengan
ketinggian sedang sampai pegunungan, dimana ketersediaan pohon buah semakin
terbatas, sedangkan spesies Hylobatidae lainnya lebih banyak di kawasan hutan
dataran rendah yang lebih kaya dengan keragaman pohon dan pohon buah
(Chivers 2001, Conklin-Brittain et al. 2001).
Aktivitas harian satwa primata pada umumnya dipelajari melalui pendekatan
alokasi waktu (time budget) setiap jenis aktivitas. Penelitian pola makan satwa
Istirahat Berpindah
Makan Vokalisasi
Istirahat Berpindah
Makan Vokalisasi
primata di habitatnya umumnya didasarkan pada persentase alokasi waktu untuk
aktivitas makan (Conklin-Brittain et al. 2001, Ahsan 2001). Beberapa metode
yang dapat digunakan dalam mempelajari berbagai aktivitas satwa disajikan oleh
Martin & Bateson (1993), di antaranya ad libitum sampling, focal sampling,
instantaneous sampling dan scan sampling.
Vokalisasi
Semua spesies dari Hylobatidae menghasilkan suara atau vokalisasi
menyerupai nyanyian dengan pola yang spesifik untuk spesies dan jenis kelamin,
biasanya dilakukan pada pagi hari (Geissmann 1995, Geissmann & Nijman 2006).
Aktivitas bersuara di pagi hari merupakan awal aktivitas harian kelompok
Hylobates. Vokalisasi berfungsi antara lain: untuk menunjukkan teritorinya,
sekaligus sebagai pengaturan ruang antar kelompok, atraksi kawin, dan untuk
mempererat hubungan sebagai pasangan kawin (Bates 1970, Leighton 1987,
Cowlishaw 1992). Vokalisasi dari Hylobatidae cukup nyaring melengking
sehingga dapat terdengar sampai 1 km (Gittins & Raemaekers 1980), bahkan
terdengar sampai 2 km (O’Brien et al. 2004).
Pola vokalisasi Hylobates dapat divisualisasikan dalam bentuk sonagram,
dan menunjukkan variasi di antara individu dan spesies, antara lain dalam hal
durasi, frase suara, jumlah not, dan frekuensi, seperti pada Gambar 8 (Geissmann
1995, Geissmann 2006a). Variasi vokalisasi tersebut, mengindikasikan perbedaan
genetik minimal pada level spesies (Brockelman & Schilling 1984, Geissmann
1984), sehingga dapat digunakan sebagai penciri spesies (Mitani 1987). Pada
dewasa ini, karakteristik vokalisasi masing-masing spesies dapat digunakan untuk
memperkirakan hubungan sistematik spesies dari Hylobatidae dan menyusun
pohon filogeninya (Geissmann 2002a).
Hylobatidae betina menghasilkan vokalisasi yang sangat menonjol, nyaring
melengking dan terbagi dalam beberapa pase, disebut great call; biasanya terdiri
dari 6-100 not, tergantung spesies, dengan durasi 6-30 detik. Bentuk not dan
interval antar not dari great call merupakan pola spesifik spesies (Geissmann
Gambar 8 Sonagram vokalisasi Hylobatidae (1) great call betina, (2) Jantan, a. H. agilis, b. H. lar, c. H. moloch, d. H. muelleri, e. H. pileatus, f. H. klossii, g. H. hoolock, h. N. Concolor, i. N. Leucogenys, j. N. Gabriellae, dan k. S. syndactylus (Geissmann 1995, Geissmann 2006a)
menghasilkan nyanyian yang lasim disebut duet call, kecuali pada H. moloch dan
H. klossii (Dallmann & Geissmann 2001, Geissmann 2006a). Vokalisasi great
call, biasanya didahului oleh serangkaian not-not singkat sebagai pengantar,
kemudian disambung dengan not-not great call. Interval antar great call sekitar 2
menit, diisi dengan not-not singkat sebagai selingan (Geissmann 2006a).
Vokalisasi dari genus Bunopithecus (Hoolock), Nomascus dan
Symphalangus, hanya berupa duet call dari pasangan kawin, sedangkan pada
genus Hylobates, kecuali H. moloch dan H. klossii, dapat berupa duet call, solo
betina dan solo jantan. Vokalisasi H. moloch dan H. klossii, hanya berupa solo
betina termasuk great call, dan solo jantan; tidak menghasilkan duet call
(Geissmann & Nijman 2006).
Vokalisasi great call dibedakan atas tiga fase, yaitu fase pre-trill, fase trill
dan fase post-trill, pada spesies H. moloch (Dallmann & Geissmann 2001, 2001a),
H. klossii (Haimoff & Tilson 1985), dan H. agilis (Haimoff & Gittins 1985),
seperti pada Gambar 9 (Dallmann & Geissmann 2001).
Gambar 9 Komposisi great call dari Hylobates
Hasil penelitian Dallmann dan Geissmann (2001) pada H. moloch
mendapatkan adanya perbedaan yang signifikan great call betina antar individu
dan antar populasi. Keragaman vokalisasi great call antar individu pada spesies
H. moloch, lebih tinggi dibandingkan dengan H. klossii, tetapi lebih rendah dari
Populasi
Populasi didefinisikan sebagai kelompok organisme yang terdiri dari
individu-individu satu spesies yang saling berinteraksi dan melakukan
perkembangbiakan pada suatu tempat dan waktu tertentu (Anderson 1985).
Batasan populasi tersebut, disesuaikan oleh Alikodra (2002) untuk digunakan
dalam pengelolaan satwa liar, menjadi “kelompok organisme yang terdiri dari
individu-individu satu spesies yang mampu menghasilkan keturunan yang sama
dengan tetuanya”. Tarumingkeng (1994) mendefinisikan populasi adalah
sehimpunan individu atau kelompok individu suatu jenis makhluk hidup yang
tergolong dalam satu spesies (atau kelompok lain yang dapat melangsungkan
interaksi genetik dengan jenis yang bersangkutan), dan pada suatu waktu tertentu
menghuni suatu wilayah atau tata ruang tertentu.
Pada umumnya populasi primata di hutan hujan tropis menghadapi ancaman
karena destruksi dan fragmentasi habitat, dan kegiatan perburuan untuk konsumsi
dan perdagangan hewan piaraan (Meijaard et al. 1999, Robinson & Bennett 2000,
Cowlishaw & Dunbar 2000). Tidak terkecuali, populasi satwa primata di
Indonesia dihadapkan pada ancaman oleh aktivitas legal/illegal logging,
perambahan dan konversi hutan, kebakaran hutan dan perburuan satwa (Meijaard
et al. 1999, Apriadi 2001, Andayani et al. 2001, O’Brien et al. 2004). Alikodra
(2002) menyatakan bahwa populasi satwa liar, termasuk primata, semakin
terdesak oleh aktivitas kehidupan manusia. Satwa liar banyak yang diburu untuk
berbagai keperluan antara lain: diperdagangkan, konsumsi daging, keperluan
pertunjukan, dan dipelihara sebagai hewan kesayangan (pet).
Suatu populasi memiliki sifat-sifat khas yaitu kepadatan (densitas), laju
kelahiran (natalitas), laju kematian (mortalitas), sebaran (distribusi), pemencaran
(dispersi), struktur umur, potensi biotik, sifat genetik, nisbah kelamin dan perilaku
(Tarumingkeng 1994, Alikodra 2002). Kepadatan populasi itu sendiri dipengaruhi
oleh beberapa parameter demografi antara lain natalitas, mortalitas, struktur
populasi, nisbah kelamin, dan migrasi (Alikodra 2002).
Studi populasi Hylobates, khususnya ukuran populasi dan karakteristik
demografik di habitatnya, masih relatif jarang dilakukan. Hal ini tidak terlepas
populasi yang tidak terhabituasi cukup sulit untuk dihitung dan diteliti
(Brockelman & Srikosamatara 1993, O’Brien et al. 2004, Nijman 2004).
Akibatnya, informasi kepadatan populasi dan karakteristik demografik Hylobates
khususnya di Indonesia masih relatif sedikit (Tabel 1).
Perilaku vokalisasi Hylobates di pagi hari, dapat dimanfaatkan dalam
melakukan survei populasi satwa primata tersebut (Brockelman dan
Srikosamatara 1993, Nijman 2004). Estimasi kepadatan kelompok dan populasi
dapat dilakukan dengan metode fixed point count berdasarkan vokalisasinya
(O’Brien et al. 2004, Buckley 2004). Selain itu, metode yang umum digunakan
dalam estimasi besar populasi Hylobates adalah line transect (Kool 1992).
Tabel 1 Estimasi densitas beberapa populasi Hylobates di Indonesia
Status Konservasi
Semua famili Hylobatidae di Indonesia merupakan satwa yang dilindungi
sesuai Surat Keputusan Menteri Kehutanan No. 301/Kpts-II/1991 dan Peraturan
Pemerintah No. 7 tahun 1999. Selain itu, H. agilis dikategorikan oleh IUCN
(International Union for Conservation of Nature and Natural Resources) sebagai
spesies yang masih berisiko rendah namun hampir terancam (low risk : near
threatened) (Eudey 2000); sedangkan menurut CITES (Convention on
International Trade in Endangered Species of Wild Fauna and Flora) termasuk
dalam Appendix I sebagai satwa yang tidak boleh diperdagangkan (Soehartono &
Mardiastuti 2002).
Keadaan Umum Lokasi Penelitian
Geografi
Taman Nasional Sebangau Kalimantan Tengah sebelumnya merupakan
kawasan hutan produksi yang kemudian dirubah fungsinya menjadi taman
nasional berdasarkan Surat Keputusan Menteri Kehutanan No.
423/Menhut-II/2004 Tanggal 19 Oktober 2004, seluas 568.700 ha. Kawasan Taman Nasional
Sebangau berada pada tiga wilayah daerah tingkat II, yaitu Kabupaten Katingan,
Kabupaten Pulang Pisau, dan Kota Palangka Raya, dengan koordinat 113°
18'-114° 03' BT dan 01° 55'-03° 07' LS. Kawasan ini terletak di antara Sungai
Katingan dan Sungai Sebangau, dan dialiri oleh beberapa sungai kecil yang
bermuara di kedua sungai tersebut (Gambar 10) (Drasospolino, 2004).
Sebelum ditetapkan sebagai taman nasional, sebagian kawasan hutan
Sebangau dikelola oleh CIMTROP (Centre for International Cooperation in
Management of Tropical Peatland) Universitas Palangka Raya sebagai
Laboratorium Alam Hutan Gambut (LAHG) seluas ±50.000 ha. LAHG
CIMTROP Universitas Palangka Raya berada pada daerah aliran sungai Sebangau
yang membatasinya dengan Desa Kereng Bangkirai, kecamatan Pahandut, Kota
LAHG CIMTROP
Univ. Palangka Raya
Luas : ±50.000 ha
Gambar 10 Taman Nasional Sebangau, Kalimantan Tengah
Sebagai taman nasional yang baru, data informasi ekologi TN Sebangau
masih sangat terbatas, kecuali di kawasan Laboratorium Alam Hutan Gambut
(LAHG) CIMTROP Universitas Palangka Raya yang kini merupakan bagian dari
TN Sebangau, sudah dilakukan beberapa penelitian, seperti vegetasi (Rieley et al.
1996, Shepherd et al. 1997), dan biodiversitas fauna (Rieley et al. 1996, Page
et al. 1997).
Topografi
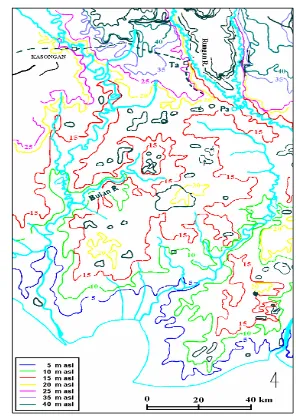
Secara umum topografi TN Sebangau relatif datar dengan ketinggian antara
5-20 m dari permukaan laut (dpl) seperti terlihat pada Gambar 11, dan ketebalan
gambut antara 0-12 m (CIMTROP 2002). Keadaan ini membentuk beberapa tipe
hutan dengan vegetasi yang berbeda. Tipe hutan beserta vegetasinya yang ada di
LAHG berbeda menurut jaraknya dari Sungai Sebangau.
Gambar 11 Profil topografi TN Sebangau (CIMTROP 2002)
Rieley et al. (1996), Rieley & Ahmad-Shah (1996), Shepherd et al. (1997),
dan Page at al. (1999), membedakan beberapa tipe hutan di LAHG, yaitu (1)
hutan rivarian (RF) dengan jarak sampai 1 km dari sungai, (2) hutan rawa gambut
campuran (MSF) 1-5 km, (3) hutan tegakan rendah (LPF) 5-13 km, dan (4) hutan
tegakan tinggi (TIF) >13 km dari pnggir sungai.
Kedalaman muka air tanah (water table) di bawah permukaan gambut di
LAHG yang diukur pada akhir musim kemarau di beberapa lokasi, bervariasi
menurut tipe hutan. Pengukuran kedalaman muka air tanah di MSF (2 lokasi)
masing-masing 39,0 cm; di LPF (3 lokasi) masing-masing 34,3 cm, 23,7 cm dan
Iklim
Iklim di Pulau Kalimantan adalah iklim hujan tropis. Iklim ini memiliki
delapan bulan basah dan tidak memiliki bulan kering yang nyata. Jumlah hujan
pada bulan paling kering lebih dari 60 mm (Ditjen PHKA 2006a). Musim hujan di
Palangka Raya dan sekitarnya termasuk LAHG mulai pada bulan Oktober sampai
bulan Juni, sedangkan musim kemarau dari bulan Juli sampai bulan September.
Curah hujan tahunan maksimum mencapai 2.600 mm, dengan curah hujan
bulanan bervariasi dari 22-525 mm (CIMTROP 2002). Temperatur udara di
LAHG rata-rata 27,20C dengan kisaran 23,10C-34,70C di areal terbuka (sekitar camp LAHG), sedangkan temperatur udara di dalam hutan rata-rata 25,60C dengan kisaran 23,00C-29,60C (Realey et al. 1996).
Flora
Terdapat beberapa jenis flora di kawasan ini, antara lain ramin (Gonystilus
bancanus), jelutung (Dyera costulata), dan belangeran (Shorea belangeran),
bintangur (Calophyllum sclerophyllum), jinjit (Calophyllum canum), meranti
(Shorea sp.), nyatoh (Palaquium sp.), keruing (Dipterocarpus sp.), agathis
(Agathis sp.), menjalin (Xanthophyllum sp), bengaris (Kompassia malaccensis),
hangkang (Palaquium leiocarpum), tumih (Combretocarpus rotundatus),
jambu-jambu (Eugenia sp.), galam tikus (E. spicata), manggis-manggis (Gracinia sp.),
malam-malam (Diospyros pseudomalabarica dan D. siamang), medang (Ixora
sp.), kenari (Blumeodendron takbrai), mahang kerume (Ternstroemia magnifica),
Lithocarpus dasystachys, milas merah (Xanthophyllum amoenum) , mendarah
merah (Knema intermedia), kopi-kopi (Randia sp.), Aglaia rubiginosa,
Parastemon sp., Polyalthia sp., Neoscortechinia kingii, Litsea sp. Xylopia fusca,
Aromadendron nutans, Horsfieldia crassifolia, Cotylelobium lanceolatum,
Licania splendens, Campnosperma coriaceum, Tetractomia tetrandra, Syzygium
clavatum, Castanopsis foxworthyii. Gymnostoma sumatrana, Ilex hypoglauca,
Palaquium pseudorostratum, Tetramerista glabra, dan Syzygium remotifolium
(Shepherd et al. 1997, Ditjen PHKA 2006a). Profil vegetasi hutan di kawasan
Gambar 12 Vegetasi di LAHG, TN Sebangau
Fauna
Jenis Mamalia yang terdapat di kawasan hutan Sebangau, antara lain
orangutan (Pongo pygmaeus), bekantan (Nasalis larvatus), kalawet (H. agilis),
monyet ekor panjang (M. fascicularis) beruk (M. nemestrina), kalasi (Presbytis
baricunda), beruang madu (Helarctos malayanus), babi hutan (Sus barbatus), rusa
Sambar (Cervus unicolor), kijang mas (Muntiacus atheroides), kancil (Tragulus
javanicus), macan dahan (Neofelis nebulosa), tupai (Tupaia sp.), loris (Nycticebus
coucang), dan tarsius (Tarsius bancanus). Jenis burung antara lain pecuk ular
(Anhinga melanogaster), cangak laut (Ardea sumatrana), cangak merah
(A. purpurea), elang hitam (Ictinaetus malayensis), pergam (Ducula bicolor),
enggang berjambul putih (Aceros comatus), enggang gunung (A. undulatus),
enggang gading (Buceros vigil), enggang badak (B. rhinoceros), dan bangau
tong-tong (Leptoptilus javanicus), layang-layang api (Hirundo rustica), dan
layang-layang bulu (H. tahitica). Jenis reptil antara lain sanca (Python
rufus), cobra (Naja sumatrana), ular hijau (Ahaetulla prasina), ular coklat malaya
(Xenelaphis hexagonatus), cicak terbang (Draco sp.), biawak (Varanus
borneensis), kura-kura kotak (Cuora amboinensis), dan kura-kura berduri
METODE PENELITIAN
Tempat Penelitian
Penelitian ini dilaksanakan di kawasan Laboratorium Alam Hutan Gambut
(LAHG) seluas ±50.000 ha, yang dikelola oleh CIMTROP (Centre for
International Cooperation in Management of Tropical Peatland) Universitas
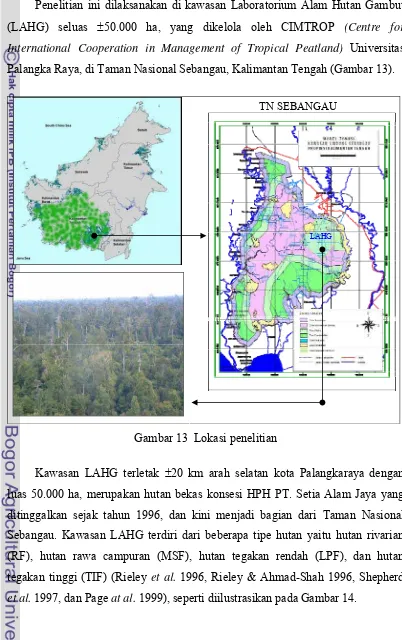
Palangka Raya, di Taman Nasional Sebangau, Kalimantan Tengah (Gambar 13).
Gambar 13 Lokasi penelitian
Kawasan LAHG terletak ±20 km arah selatan kota Palangkaraya dengan
luas 50.000 ha, merupakan hutan bekas konsesi HPH PT. Setia Alam Jaya yang
ditinggalkan sejak tahun 1996, dan kini menjadi bagian dari Taman Nasional
Sebangau. Kawasan LAHG terdiri dari beberapa tipe hutan yaitu hutan rivarian
(RF), hutan rawa campuran (MSF), hutan tegakan rendah (LPF), dan hutan
tegakan tinggi (TIF) (Rieley et al. 1996, Rieley & Ahmad-Shah 1996, Shepherd
et al. 1997, dan Page at al. 1999), seperti diilustrasikan pada Gambar 14.
TN SEBANGAU
1 km 5 km 13 km
Gambar 14 Ilustrasi tipe hutan di LAHG
Hutan rivarian merupakan daerah genangan air ketika sungai meluap
(musim hujan) dengan jarak sampai 1 km dari pinggir sungai, didominasi oleh
tumbuhan herba dan semak. Hutan rawa gambut campuran (MSF) terletak antara
1-5 km dari pinggir sungai dengan kedalaman gambut 2-6 m. Lantai hutan selalu
tergenang oleh luapan sungai di musim hujan rata-rata 0,3 m di atas permukaan
gambut, dan pada musim kering muka air tanah 0,4 m di bawah permukaan
gambut. MSF mempunyai vegetasi yang relatif tinggi (sampai 35 m) dengan
lapisan kanopi yang bertingkat. Hutan tegakan rendah (LPF) terletak antara 5-13
km dari pinggir sungai dengan kedalaman gambut 6-10 m. Pada musim hujan,
lantai hutan LPF juga tergenang air lebih dari 0,3 m di atas permukaan gambut,
dan pada musim kering muka air tanah hanya 0,24 m di bawah permukaan
gambut. Karakteristik vegetasi di LPF, antara lain: tinggi kanopi jauh lebih rendah
dibanding MSF dan TIF, kanopi lapisan atas maksimum 20 m, sangat terbuka dan
didominasi oleh tumih (Combretocarpus rotundatus); tinggi kanopi lapisan bawah
rata-rata di bawah 15 m yang sebagian besar merupakan vegetasi tingkat tiang;
dan lantai hutan rata-rata ditumbuhi oleh pandan yang cukup padat. Hutan tegakan
tinggi (TIF) berjarak lebih dari 13 km dari pinggir sungai dengan kedalaman
gambut 8-12 m. Berbeda kontras dengan MSF dan LPF, TIF tidak tergenang di
musim hujan, hanya muka air tanah yang naik menjadi 0,2-0,3 m di bawah
permukaan gambut, sedangkan di musim kering muka air tanah sekitar 1,5 m di
bawah permukaan gambut. Karakteristik vegetasi di TIF, antara lain: terdiri atas
pohon besar dengan kanopi yang tinggi (sampai 45 m), dan lapisan kanopi yang
bertingkat dan relatif lebih tertutup. Lantai hutan relatif bersih dari pandan dan
semak (Realey et al. 1996, Rieley & Ahmad-Shah 1996, Shepherd et al. 1997).
MSF sungai
Waktu Penelitian
Peninjauan lapangan dan survei awal dilakukan pada bulan Oktober
sampai dengan November 2004. Pengumpulan data dilakukan pada bulan Juli
sampai dengan November 2005.
Materi Penelitian
Materi yang digunakan sebagai objek penelitian dalam analisis vegetasi
adalah vegetasi yang terdapat di habitat kalawet di enam lokasi pengamatan di
LAHG, sedangkan materi pengamatan populasi adalah 47 kelompok kalawet yang
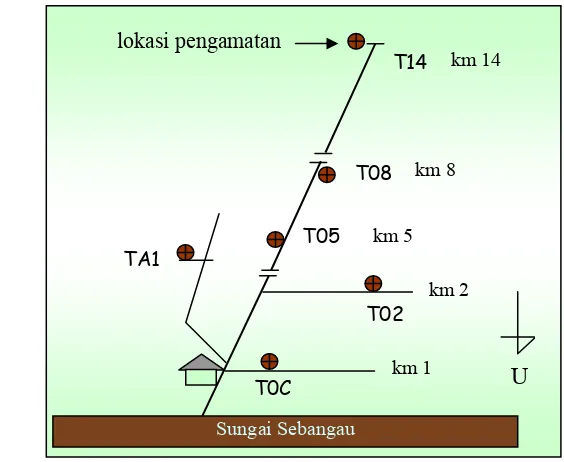
teridentifikasi di enam lokasi pengamatan di LAHG TN. Sebangau (Gambar 17).
Untuk pengamatan tingkah laku harian, hanya satu keluarga yaitu kelompok KC
(4 ekor) yang terdiri dari sepasang jantan-betina dewasa, seekor remaja dan seekor
bayi. Untuk analisis vokalisasi, dilakukan perekaman suara dari 13 kelompok
H. agilis albibarbis di LAHG, dua kelompok hibrida H. agilis albibarbis x
H. muelleri di Barito Ulu, dua pasang dan satu betina dewasa H. muelleri di
kandang rehabilitasi ‘Kalaweit Program’ Hampapak, Kalimantan Tengah.
Alat yang digunakan berupa 2 unit binokuler, 1 unit GPS (Global
Positioning System) Etrex Garmin 12, 2 unit kompas, 1 unit rekorder kaset, 1 unit
rekorder digital, dan 1 unit kamera digital, serta foto landsat dan sejumlah
software pendukung.
Metode Penelitian
Habitat
1. Analisis vegetasi
Analisis vegetasi dilakukan pada daerah jelajah kalawet yang akan diamati.
Pengamatan dilakukan dengan menggunakan metode garis berpetak (Gambar
15). Petak contoh berbentuk empat persegi berukuran 20x20 m sebagai petak
utama (untuk tingkat pohon), 10x10 m (tingkat tiang), dan 5x5 m (tingkat
pancang), dan 2x2 m (tingkat semai) (Soegianto 1994; Soerianegara & Indrawan