Urban) PADA KETINGGIAN TEMPAT DAN NAUNGAN
YANG BERBEDA
BUDI MARTONO
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Dengan ini saya menyatakan bahwa disertasi Keragaman dan Tanggap Pertumbuhan serta Produksi Asiatikosida Pegagan (Centella asiatica (L.) Urban) pada Ketinggian Tempat dan Naungan yang Berbeda adalah karya saya dengan arahan komisi pembimbing dan belum diajukan dalam bentuk apapun kepada perguruan tinggi manapun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir disertasi ini.
Bogor, Juli 2011
Production of Asiatic Pennywort (Centella asiatica (L.) Urban) at Different Site Elevations and Shades. Supervised by MUNIF GHULAMAHDI, LATIFAH K. DARUSMAN, SANDRA ARIFIN AZIZ, and NURLIANI BERMAWIE
In developing products derived from asiatic pennywort there is a lack of guarantee in terms of raw material supply and quality. To support such needs, standardized plant material will ensure production level and quality. The objectives of this research were: 1) to obtain information of the genetic parameters, i.e. of asiatic pennywort morphological characters and criterion of selection to produce high content of asiaticoside and dry shoot production, obtaining asiatic pennywort accessions which have high content of asiaticoside and dry shoot production, and information about the result of asiatic pennywort selection based on independent culling level, a single selection, weighted and unweighted standardized selection indexes, 2) to analyse and evaluate genetic distances of 17 accessions asiatic pennywort based on morphological, RAPD, and the data of markers combination, and 3) to obtain response of the growth and asiaticoside production of six asiatic pennywort plant accessions in different shades on low and high elevation areas. Results of the research showed that number of leaf; length, width, and leaf area: number of vein leaf, and segment length on the longest stolon have wide genetic variability and high heritability. The length and diameter of leaf petiole: leaf thickness, number of runner, content and production of asiaticoside that showed narrow genetic variability and heritability from low to moderate. Narrow genetic variability and high heritability were shown by characters of dry shoot weight. Direct selection toward dry shoot production could be conducted through selection of leaf area. The selection indicated that 4 accessions of asiatic pennywort showed high content of asiaticoside and dry shoot weight, namely Casi 016, Casi 003, Casi 008, and Casi 002. Grouping of asiatic pennywort accessions based on morphological, RAPD, and the basis of markers combination divided the 17 accessions of asiatic pennywort into 4, 5, and 4 groups at similarity level of 0.33, 0.73, and 0.72, respectively. In high elevation, highest production of asiaticoside (2.34 g m-2) and content of asiaticoside above MMI standard (1.33%) occured in Casi 002 planted without shade.
Asiatikosida Pegagan (Centella asiatica (L.) Urban) pada Ketinggian Tempat dan Naungan yang Berbeda. Dibimbing oleh MUNIF GHULAMAHDI, LATIFAH K. DARUSMAN, SANDRA ARIFIN AZIZ, dan NURLIANI BERMAWIE.
Permasalahan dalam pengembangan produk yang berasal dari pegagan (Centella asiatica (L.) Urban) adalah tidak terjaminnya pasokan bahan baku dan mutu, karena selama ini pegagan diambil langsung dari alam tanpa usaha pembudidayaan. Untuk mengatasi masalah tersebut maka diperlukan usaha-usaha budidaya dengan menggunakan bahan tanaman unggul dan cara budidaya yang tepat untuk mendapatkan produktivitas yang optimal. Untuk melaksanakan program ini, maka diperlukan informasi tentang pendugaan parameter genetik, analisis lintas, seleksi plasma nutfah pegagan berdasarkan seleksi penyisihan bebas bertingkat, seleksi tunggal, indeks seleksi terboboti dan tidak terboboti; keragaman plasma nutfah pegagan berdasarkan penanda morfologi, molekuler, dan gabungan, serta pengujian beberapa aksesi pegagan di dataran rendah dan dataran tinggi pada naungan yang berbeda (tanpa naungan, naungan 25 dan 55%). Tujuan penelitian ini adalah: 1) menduga keragaman genetik dan heritabilitas beberapa karakter kuantitatif pegagan, mendapatkan informasi tentang karakter yang dapat digunakan sebagai kriteria seleksi kadar asiatikosida dan produksi terna kering yang tinggi, dan mendapatkan aksesi pegagan dengan kadar asiatikosida dan bobot terna kering yang tinggi berdasarkan seleksi penyisihan bebas bertingkat serta membandingkan hasil seleksi berdasarkan seleksi penyisihan bebas bertingkat, seleksi tunggal, indeks seleksi terboboti dan tidak terboboti, (2) menganalisis dan mengevaluasi jarak kedekatan genetik pada 17 aksesi pegagan berdasarkan penanda morfologi, penanda RAPD, dan data gabungan, dan (3) memperoleh informasi tentang tanggap pertumbuhan dan produksi asiatikosida 6 aksesi pegagan terhadap perlakuan naungan di dataran rendah dan tinggi.
Penelitian ini terdiri atas dua aspek, yaitu (1) keragaman plasma nutfah pegagan dan (2) tanggap perubahan karakter pegagan. Kedua aspek tersebut dikelompokkan menjadi tiga subjudul penelitian: (1) pendugaan parameter genetik, analisis lintas, dan seleksi plasma nutfah pegagan, (2) keragaman plasma nutfah pegagan berdasarkan penanda morfologi, molekuler, dan gabungan, dan (3) tanggap pertumbuhan dan produksi asiatikosida beberapa aksesi pegagan pada naungan yang berbeda di dataran rendah dan tinggi. Kajian pada subjudul 1 dan 2 dilakukan terhadap 17 aksesi pegagan dengan perlakuan naungan paranet 25%, sedangkan kajian pada subjudul 3 dilakukan terhadap 4 aksesi terpilih hasil seleksi dan 2 aksesi sebagai pembanding pada tiga tingkat naungan (tanpa naungan, naungan 25 dan 55%) dan ketinggian tempat yang berbeda (250 m dan 1500 m dpl).
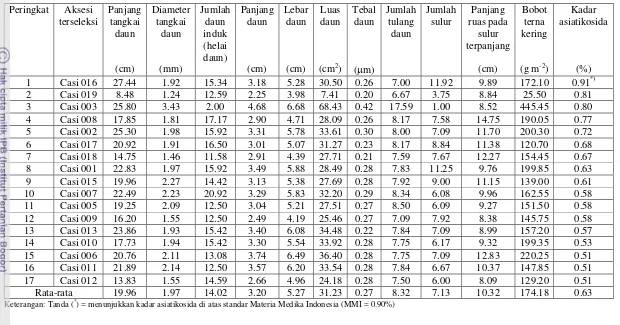
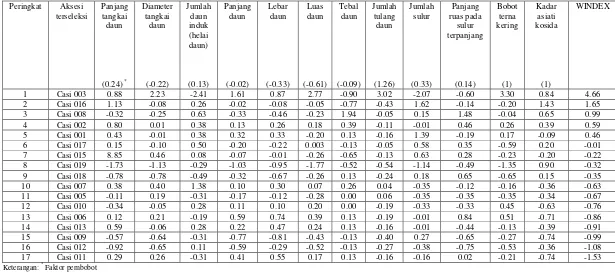
terna kering secara langsung dapat dilakukan melalui seleksi luas daun. Hasil seleksi dengan mempertimbangkan karakter kadar asiatikosida dan bobot terna kering menghasilkan 4 aksesi pegagan terpilih, yaitu Casi 016 (Boyolali), Casi 003 (Introduksi), Casi 008 (Ciwidey), dan Casi 002 (Bengkulu). Aksesi terseleksi berdasarkan seleksi penyisihan bebas bertingkat tidak selalu terseleksi pada seleksi tunggal, indeks seleksi terboboti dan tidak terboboti. Aksesi yang terseleksi sama berdasarkan seleksi penyisihan bebas bertingkat dan seleksi tunggal adalah aksesi lokal Boyolali (Casi 016). Aksesi lokal Ciwidey (Casi 008) dan lokal Bengkulu (Casi 002) merupakan aksesi yang terseleksi sama berdasarkan seleksi penyisihan bebas bertingkat dan seleksi indeks terboboti, sedangkan aksesi lokal Boyolali (Casi 016) dan lokal Bengkulu (Casi 002) merupakan aksesi yang terseleksi sama berdasarkan seleksi penyisihan bebas bertingkat dan seleksi indeks tidak terboboti. Tidak ada satupun aksesi yang terseleksi sama berdasarkan ketiga metode seleksi.
pendek; bobot terna kering dan produksi asiatikosidanya berkurang. Selain itu, kandungan klorofil a, b, dan total meningkat di dataran rendah dan berkurang di dataran tinggi. Pegagan yang tumbuh di dataran rendah pada naungan 0, 25, dan 55%, memiliki panjang tangkai daun lebih panjang (4.85–25.75 cm) daripada yang di dataran tinggi (4.85–19.25 cm). Namun demikian, daunnya lebih sempit (24.72–29.87 cm2) dibandingkan di dataran tinggi (33.23– 46.31 cm2) kecuali untuk Casi 003 lebih luas (59.46 cm2). Kandungan asiatikosida di dataran rendah (0.36–0.64%) lebih rendah dibandingkan dengan di dataran tinggi (0.80–1.38%). Produksi asiatikosida di dataran tinggi (0.42–2.34 g m-2) lebih tinggi dibandingkan dengan dataran rendah (0.17-1.62 g m-2). Produksi asiatikosida tertinggi di dataran rendah dan dataran tinggi ditemukan pada perlakuan tanpa naungan kecuali Casi 003 pada naungan 25% (1.59 g m-2 di dataran rendah dan 1.10 g m-2di dataran tinggi). Penanaman tanpa naungan di dataran tinggi dengan menggunakan Casi 002 menghasilkan kadar asiatikosida di atas standar MMI (1.33%) dan produksi asiatikosida tertinggi (2.34 g m-2). Di dataran rendah, penanaman tanpa naungan dengan menggunakan Casi 016 menghasilkan produksi asiatikosida yang tinggi (1.62 g m-2) dengan kadar asiatikosida di atas standar MMI (1.03%). Penanaman Casi 016 di dataran tinggi dapat dilakukan pada naungan 25% dan tanpa naungan dengan kadar asiatikosida di atas standar MMI (1.85 dan 1.58%) dan produksi asiatikosidanya 1.84 g m-2. Aksesi hasil eksplorasi di dataran menengah dan tinggi menghasilkan kadar asiatikosida yang lebih rendah jika ditanam di dataran rendah dibandingkan dataran tinggi.
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan yang wajar IPB
Penguji pada Ujian Terbuka: Dr. Ir. Yul H. Bahar
Urban) PADA KETINGGIAN TEMPAT
DAN NAUNGAN YANG BERBEDA
BUDI MARTONO
Disertasi
Sebagai salah satu syarat untuk memperoleh gelar Doktor pada
Program Studi Agronomi
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Nama : Budi Martono
NIM : A161060151
Disetujui Komisi Pembimbing
Dr. Ir. Munif Ghulamahdi, M.S. Prof. Dr. Ir. Latifah K. Darusman, M.S.
Ketua Anggota
Dr. Ir. Sandra Arifin Aziz, M.S. Dr. Ir. Nurliani Bermawie
Anggota Anggota
Mengetahui
Ketua Program Studi Dekan Sekolah Pascasarjana
Agronomi
Dr. Ir. Munif Ghulamahdi, M.S. Dr. Ir. Dahrul Syah, M.Sc.Agr
Puji dan syukur penulis panjatkan ke hadirat Allah SWT atas rahmat dan
karunia-Nya sehingga penulis dapat menyelesaikan karya ilmiah ini. Disertasi ini
memberikan informasi mengenai keragaman dan tanggap pertumbuhan serta
produksi asiatikosida pegagan pada ketinggian tempat dan naungan yang berbeda.
Dengan selesainya disertasi ini penulis mengucapkan banyak terima kasih
kepada Dr. Ir. Munif Ghulamahdi, M.S., selaku ketua komisi pembimbing, Prof.
Dr. Ir. Latifah K. Darusman, M.S., Dr. Ir. Sandra Arifin Aziz, M.S., dan Dr. Ir.
Nurliani Bermawie sebagai anggota komisi pembimbing yang telah banyak
memberikan bimbingan dengan cermat, terarah, dan sistematis serta motivasi yang
diberikan kepada penulis mulai dari perencanaan, pelaksanaan, sampai
penyelesaian penulisan penelitian disertasi ini.
Ucapan terima kasih juga penulis sampaikan kepada Kepala Badan
Litbang Pertanian, Ketua Komisi Pembinaan Tenaga Badan Litbang Pertanian,
Kepala Puslitbangbun, dan Kepala Balai Penelitian Tanaman Rempah dan
Tanaman Industri Lain (Balittri) yang telah memberikan kesempatan kepada
penulis untuk mengikuti pendidikan doktor di Institut Pertanian Bogor serta
fasilitas lainnya sehingga penelitian ini dapat terlaksana dengan baik. Ungkapan
terima kasih juga disampaikan kepada Dr. Ir. Made Sumertajaya, M.Si., atas
kesediannya menjadi pendamping dalam pelatihan analisis statistik. Ucapan yang
serupa disampaikan kepada semua teknisi Kebun Percobaan Cimanggu, Kebun
Percobaan Gunung Putri, Laboratorium Pascapanen Balittro, Laboratorium
Research Group on Crop Improvement (RGCI), Laboratorium Ekofisiologi Departemen AGH Fakultas pertanian, dan Laboratorium Histologi Biotrop yang
telah membantu penulis dalam pelaksanaan penelitian. Penghargaan tak terhingga
juga penulis tujukan kepada orang tua, mertua, istri (Dr. Sri Cahyorini, M.Si.),
kakak, adik, Dr. Subandi, M.Sc., Drs. Sarodjo, dan semua teman-teman yang
dengan tulus ikhlas memberikan doa dan dukungannya kepada penulis.
Akhirnya, penulis berharap semoga hasil penelitian ini bermanfaat bagi
semua pihak yang memerlukannya.
Penulis dilahirkan di Temanggung pada tanggal 7 Juni 1965, merupakan
putra keempat dari tujuh bersaudara dari ayah Soetjipto (Alm.) dan ibu Rusmi
Hadinah (Almh.). Pendidikan sarjana ditempuh di jurusan Biologi Lingkungan
Fakultas Biologi Universitas Jenderal Soedirman (UNSOED) Purwokerto, lulus
pada tahun 1989. Pada tahun 1998 penulis memperoleh beasiswa dari PAATP
untuk mengikuti program Magister Sains (S2) di Program Pascasarjana Institut
Pertanian Bogor, Program Studi Agronomi, sub program Pemuliaan Tanaman.
Kesempatan untuk melanjutkan ke program doktor pada program studi Agronomi
dan pada perguruan tinggi yang sama diperoleh pada tahun 2006. Beasiswa
pendidikan pascasarjana diperoleh dari Kementerian Pertanian Republik
Indonesia.
Sejak Desember 1990 sampai 2006, penulis bekerja sebagai staf peneliti
pemuliaan di Balai Penelitian Tanaman Rempah dan Obat (Balittro) Bogor. Pada
tahun 2006, penulis pindah tugas di Balai Penelitian Tanaman Rempah dan Aneka
Tanaman Industri (Balittri) Pakuwon, Parungkuda Sukabumi.
Selama mengikuti S3, artikel dengan judul kriteria penanda seleksi
produktivitas terna dan asiatikosida pada pegagan (Centella asiatica (L.) Urban) telah diterbitkan pada jurnal Penelitian Tanaman Industri pada tahun 2010. Artikel
lain berjudul pendugaan parameter genetik pegagan (Centella asiatica (L.) Urban), seleksi plasma nutfah pegagan (Centella asiatica(L.) Urban) berdasarkan seleksi penyisihan bebas bertingkat, seleksi tunggal, dan indeks seleksi, serta
keragaman genetik pegagan berdasarkan penanda morfologi dan Random Amplified Polymorphic DNA (RAPD) akan diterbitkan pada jurnal Pemuliaan Indonesia (Zuriat) dan jurnal Penelitian Tanaman Industri, sedangkan
pertumbuhan dan produksi asiatikosida beberapa genotipe pegagan pada berbagai
tingkat naungan di dataran tinggi dan dataran rendah akan diterbitkan pada jurnal
Agronomi Indonesia. Karya-karya ilmiah tersebut merupakan bagian dari program
DAFTAR TABEL ... xiii
Klasifikasi, Botani, dan Syarat Tumbuh Pegagan ... 7
Manfaat dan Kandungan Kimia ... 8
Parameter Genetik, Analisis Lintas, dan Seleksi ... 11
Parameter Genetik ... 11
Analisis Lintas ... 12
Seleksi ... 13
Penanda Genetik dalam Identifikasi Keanekaragaman Pegagan ... 14
Penanda morfologi ... 15
Penanda molekuler ... 15
Penanda RAPD ... 16
Pengaruh Naungan Terhadap Pertumbuhan dan Produksi Tanaman .... 17
Mekanisme Adaptasi Tanaman Terhadap Intensitas Cahaya Rendah... 19
Pengaruh Ketinggian Tempat Terhadap Pertumbuhan dan Produksi Tanaman ... 24
PENDUGAAN PARAMETER GENETIK, ANALISIS LINTAS, DAN SELEKSI PLASMA NUTFAH PEGAGAN (Centella asiatica (L.) Urban) . 26
Pendahuluan ... 27
Bahan dan metode ... 30
Hasil dan Pembahasan ... 38
Simpulan ...………...…………... 61
Pendahuluan ... 84
Bahan dan Metode ... 87
Hasil dan Pembahasan ... 91
Simpulan ….………...……….………. 122
PEMBAHASAN UMUM ... 125
SIMPULAN DAN SARAN ... 139
DAFTAR PUSTAKA ... 141
Halaman
1 Karakter morfologi dan fisiologi tanaman ternaungi ... 19
2 Analisis ragam dan harapan kuadrat tengah dari RAK untuk suatu karakter ... 35
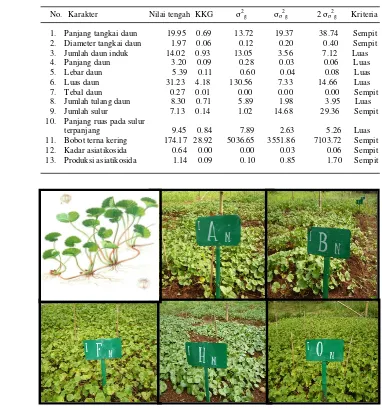
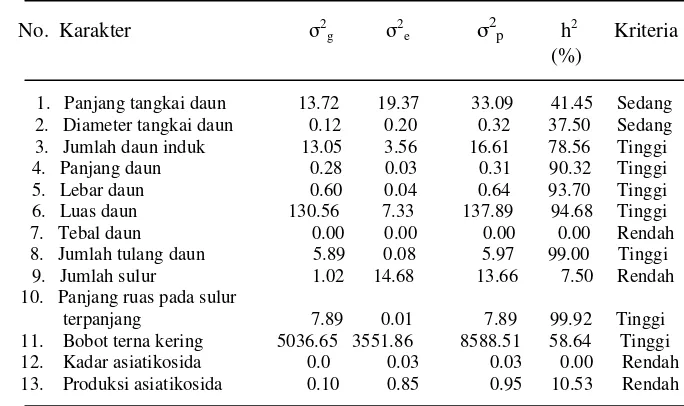
3 Nilai rata-rata, koefisien keragaman genetik (KKG), ragam genetik (σ2 g), dan standar deviasi ragam genetik (σσ2g)beberapa karakter kuantitatif plasma nutfah pegagan ... 39
4 Ragam genetik (σσ2g), ragam lingkungan (σ 2 e), ragam fenotip (σ 2 p), dan heritabilitas (h2)beberapa karakter kuantitatif plasma nutfah pegagan... 41
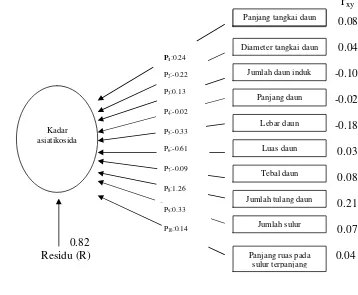
5 Nilai koefisien korelasi antara komponen pertumbuhan dan kadar asiatiko sida pada tanaman pegagan ... 42
6 Pengaruh langsung dan pengaruh tidak langsung sepuluh komponen pertumbuhan terhadap kadar asiatikosida ... 44
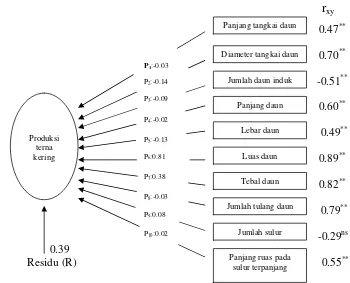
7 Nilai koefisien korelasi antara komponen pertumbuhan dan produksi terna kering pada tanaman pegagan ... 46
8 Pengaruh langsung dan pengaruh tidak langsung sepuluh komponen pertumbuhan terhadap poduksi terna kering ... 48
9 Nilai heritabilitas dalam arti luas komponen pertumbuhan pada pegagan .... 52
10 Penampilan karakter agronomi 4 aksesi pegagan hasil seleksi penyisihan bebas bertingkat (Independent culling level) ... 54
11 Penampilan karakter agronomi 17 aksesi pegagan hasil seleksi tunggal (berdasarkan kadar asiatikosida) ... 56
12 Hasil seleksi 17 aksesi pegagan berdasarkan indeks seleksi terboboti (weighted standardized selection index)... 59
13 Hasil seleksi 17 aksesi pegagan berdasarkan indeks seleksi tidak terboboti (unweighted standardized selection index) ... 60
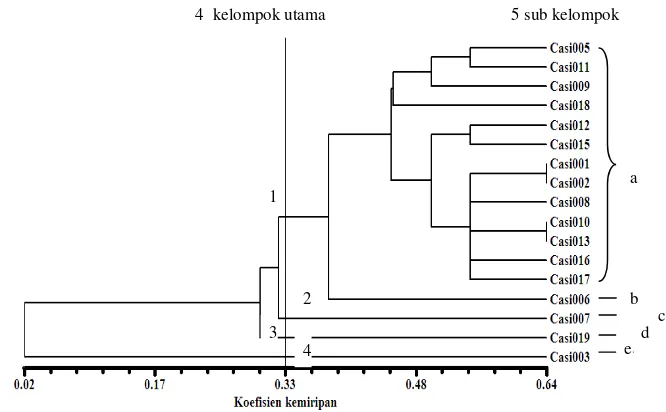
14Kelompok aksesi yang terbentuk berdasarkan dendogram morfologi …... 72
15 Nilai ciri 2 komponen utama 9 karakter morfologi ... 73
aksesi pegagan ... 75
18 Kelompok aksesi yang terbentuk berdasarkan dendogram RAPD ... 77
19 Nilai ciri 3 komponen utama pada penanda molekuler (RAPD) ... 77
20 Kelompok aksesi yang terbentuk berdasarkan data gabungan
(morfologi dan penanda molekuler)... 80
21 Nilai ciri 3 komponen utama data gabungan (morfologi dan RAPD) ... 80
22 Rangkuman hasil analisis ragam gabungan pengaruh ketinggian tempat, naungan, dan genotipe terhadap peubah yang diamati ... 92
23 Pengaruh kombinasi naungan dan genotipe terhadap luas daun ... 93
24 Pengaruh kombinasi naungan dan genotipe terhadap ketebalan daun dan panjang lapisan palisade ... 95
25 Pengaruh kombinasi naungan dengan genotipe terhadap kandungan
klorofil total (mg g-1bobot basah daun) ...……... 98 26 Pengaruh kombinasi naungan dan genotipe terhadap ratio klorofil a/b… 100 27 Pengaruh kombinasi ketinggian tempat dan naungan terhadap luas daun
pegagan (cm2) ... 101
28 Pengaruh kombinasi ketinggian tempat dan naungan terhadap ketebalan daun (m) ... 102
29 Pengaruh kombinasi ketinggian tempat dan naungan terhadap panjang lapisan palisade (m) ... 103
30 Pengaruh kombinasi ketinggian tempat dan naungan terhadap klorofil a (mg g-2bobot basah daun) ... 104
31 Pengaruh kombinasi ketinggian tempat dan naungan terhadap klorofil total (mg g-2bobot basah daun) ... 105
32 Pengaruh kombinasi ketinggian tempat dengan genotipe terhadap luas daun (cm2) ... 106
total (mg g bobot basah daun) ... 108
35 Pengaruh kombinasi ketinggian tempat dan genotipe terhadap intensitas kehijauan daun... 109
36 Pengaruh kombinasi ketinggian tempat, naungan, dan genotipe terhadap luas daun spesifik (cm2) ... 110
37 Pengaruh kombinasi ketinggian tempat, naungan, dan genotipe terhadap kerapatan stomata pada pegagan (cm2) ... 112
38 Pengaruh kombinasi ketinggian tempat, naungan, dan genotipe terhadap klorofil b (mg g-1bobot basah daun) ... 114
39 Pengaruh kombinasi ketinggian tempat, naungan, dan genotipe terhadap panjang tangkai daun pegagan (cm) ... 115
40 Pengaruh kombinasi ketinggian tempat, naungan, dan genotipe terhadap bobot terna kering pegagan (g m-2) ... 116
41 Pengaruh kombinasi ketinggian tempat, naungan, dan genotipe terhadap produksi asiatikosida (g m-2) ... 119
1 Diagram alir penelitian ... 6
2 Struktur kimia dari asiatikosida (James & Dubery 2011) ... 9
3 Skema biosintesis triterpenoid pada pegagan atauCentella asiatica(L.) Urban. (Haralampidiset al. 2002; Phillipset al. 2006; James & Dubery 2009) ... 11
4Mekanisme penghindaran terhadap kekurangan cahaya (Levitt 1980)…... 20 5 Mekanisme toleransi tanaman terhadap kekurangan cahaya
(Levitt 1980) ... 21
6 Morfologi pegagan dan keragaan beberapa aksesi pegagan: Casi 005 (A), Casi 011 (B), Casi 006 (C), Casi 002 (F), Casi 003 (H), dan Casi 016 (0) ... 39
7 Diagram analisis lintas antara komponen pertumbuhan terhadap kadar
asiatikosida... 43
8 Diagram analisis lintas antara komponen pertumbuhan terhadap produksi terna kering... 50
9 Dendogram 17 aksesi pegagan berdasarkan penanda morfologi ... 72
10 Dendogram 17 aksesi pegagan berdasarkan profil pola pita DNA dengan teknik RAPD ... 76
11 Dendogram 17 aksesi pegagan berdasarkan kombinasi penanda morfologi dan penanda RAPD ... 79
12 Penampang melintang daun pegagan terhadap naungan di dataran rendah pada perlakuan: (a) tanpa naungan, (b) naungan 25%, dan
(c) naungan 55% (perbesaran 40x) ...…... 97 13 Pola interaksi antara ketinggian tempat, naungan, dan genotipe
terhadap bobot terna kering Casi 002, Casi 003, Casi 008, Casi 012,
Casi 013, dan Casi 016 (a=dataran rendah; b=dataran tinggi)………….. 117 14 Pola interaksi antara ketinggian tempat, naungan, dan genotipe
terhadap produksi asiatikosida Casi 002, Casi 003, Casi 008, Casi 012,
Casi 013, dan Casi 016 (a=dataran rendah; b=dataran tinggi)………….. 119 15 Pola interaksi antara ketinggian tempat, naungan, dan genotipe terhadap
kadar asiatikosida Casi 002, Casi 003, Casi 008, Casi 012, Casi 013, dan
1 Data koleksi plasma nutfah pegagan hasil eksplorasi dari berbagai daerah
dan introduksi ... 157
2 Denah percobaan studi keragaman plasma nutfah pegagan (Centella asiatica(L.) Urban)... 158
3 Metode preparasi dan pengamatan struktur dan tebal daun (Sass 1951)... 159
4 Contoh perhitungan kadar asiatikosida ... 162
5 Primer yang digunakan dalamscreeningprimer RAPD... 163
6 Matriks kemiripan 17 aksesi pegagan berdasarkan penanda morfologi ... 164
7 Matriks kemiripan 17 aksesi pegagan berdasarkan penanda RAPD ... 165
8 Matriks kemiripan 17 aksesi pegagan berdasarkan kombinasi penanda morfologi dan RAPD ... 166
9 Denah percobaan tanggap pertumbuhan dan produksi asiatikosida beberapa aksesi pegagan (Centella asiatica(L.) Urban) pada naungan yang berbeda di dataran rendah dan tinggi (Lokasi: KP. Cimanggu dan KP. Gunung Putri) ... 167
10 Metode analisis kandungan klorofil (mg/g bobot basah daun)... 168
11 Metode pengamatan jumlah stomata... 169
12 Data iklim kebun percobaan Cimanggu Juli-Nopember 2008 ... 170
PENDAHULUAN
Latar Belakang
Indonesia merupakan salah satu negara mega diversity dengan keanekaragaman hayati tertinggi ke-2 setelah Brazilia dan mempunyai lebih
kurang dari 7000 spesies tumbuhan (90% dari spesies tumbuhan Asia) diketahui
berkhasiat sebagai obat (Badan POM 2001). Menurut Ditjen POM (1991) ada 283
spesies tumbuhan obat yang sudah terdaftar digunakan oleh industri obat
tradisional di Indonesia. Salah satu tumbuhan yang telah dikenal luas di dunia
sebagai obat adalah pegagan atauCentella asiatica(L.) Urban.
Terpenoid khususnya triterpenoid merupakan golongan senyawa kimia
utama pegagan, terdiri atas beberapa komponen kimia yang memiliki aktivitas
biologis. Salah satu diantaranya adalah asiatikosida, yang mempunyai khasiat
antara lain untuk antilepra dan antisifilis (Winarto & Surbakti 2003), memacu
sintesis kolagen dan mucopolisakarida untuk memperbaiki jaringan yang luka
sedangkan oksiasiatikosida dapat membunuh basilus tuberkolosis (Barnes et al. 2002; Fahmi 2002).
Pegagan banyak terdapat di Indonesia dan telah banyak dimanfaatkan
dalam ramuan tanaman obat atau jamu. Kebutuhan industri untuk bahan baku
pegagan mencapai 100 ton/th, namun pada tahun 2005 baru dapat dipasok 4 ton
(IPB 2005). Permasalahan dalam pengembangan produk yang berasal dari
pegagan adalah tidak terjaminnya pasokan bahan baku dan mutu. Untuk memasok
kebutuhan industri, selama ini pegagan diambil langsung dari alam tanpa usaha
pembudidayaan sehingga jaminan pasokan bahan baku dan mutunya tidak
terjamin. Dengan meningkatnya minat masyarakat terhadap obat bahan alam,
diperlukan pasokan bahan baku yang konsisten dengan mutu yang sesuai
kebutuhan industri melalui usaha budidaya.
Salah satu upaya untuk mendapatkan produk pegagan yang bermutu yaitu
diperlukannya bahan tanaman yang terjamin tingkat produksi dan mutunya.
Sampai saat ini baru ada 2 varietas unggul pegagan yang dilepas (Castina 1 dan
Castina 2), sehingga keadaan ini mendorong para pemulia untuk terus
Aromatik (BALITTRO) telah memiliki 17 nomor aksesi pegagan yang berasal
dari introduksi dan eksplorasi dari berbagai daerah dengan kondisi agroekologi
berbeda. Dari 17 aksesi tersebut belum diketahui karakter morfologi dan agronomi
secara menyeluruh sehingga perlu dilakukan identifikasi untuk melengkapi
informasi data deskripsi masing-masing aksesi. Selain karakterisasi dan
identifikasi, untuk program perbaikan genetik pegagan diperlukan informasi
mengenai parameter genetik.
Sebagai bahan genetik untuk pembentukan varietas unggul terutama
melalui persilangan, selain deskripsi koleksi plasma nutfah, perlu diketahui juga
hubungan kekerabatan antara aksesi pegagan. Hubungan kekerabatan atau jarak
genetik tersebut akan menentukan keberhasilan persilangan. Namun sampai saat
ini jarak genetik antar aksesi ini belum banyak diketahui sehingga perlu dilakukan
analisis keanekaragaman genetik.
Untuk mempelajari keanekaragaman genetik dapat menggunakan penanda
morfologi. Penanda morfologi telah lama digunakan sebagai penanda genetik
untuk mengatasi masalah duplikasi plasma nutfah di lapang (Simmond & Sheperd
1955). Namun karakterisasi yang didasarkan pada penanda morfologi biasanya
dipengaruhi lingkungan makro dan mikro, serta umur tanaman. Agar dapat
memperkuat informasi data penanda morfologi maka diperlukan dukungan
penanda molekuler karena memperjelas perbedaan dan hubungan kekerabatan
antar aksesi berdasarkan karakteristik molekuler (Jarret & Gawel 1995).
Salah satu penanda molekuler berbasis DNA yang telah banyak
diaplikasikan sebagai penanda genetik tanaman adalah RAPD (Random Amplified Polymorphic DNA). Penanda ini menjadi salah satu alternatif penanda genetik berbasis DNA untuk tanaman karena tekniknya lebih cepat, lebih mudah dan lebih
murah dibandingkan dengan RFLP, SSR, dan AFLP (Darmono 1996). Dengan
dasar ini maka dilakukan analisis keanekaragaman genetik berdasarkan penanda
morfologi bersama dengan penanda RAPD. Diharapkan dengan menggunakan dua
penanda tersebut akan memberikan informasi yang saling mendukung sehingga
diperoleh informasi yang lebih akurat.
Pegagan dapat ditemukan di dataran rendah sampai dengan ketinggian
pada tempat yang terbuka dan ternaungi, namun sampai sejauh mana
kemampuannya tumbuh dalam kondisi ternaungi perlu dipelajari. Rachmawaty
(2005) melaporkan bahwa tanaman ini dapat tumbuh dengan baik pada tingkat
naungan 25%, bahkan pada naungan 75% masih menunjukkan pertumbuhan yang
baik meskipun terjadi penurunan produksi pegagan. Oleh karena itu, pegagan
dapat dikembangkan sebagai tanaman sela musiman maupun tahunan, misalnya di
antara tanaman jagung, kelapa, kelapa sawit, dan buah-buahan yang tidak terlalu
rindang. Sejauh ini, informasi tentang respon pertumbuhan dan produksi
asiatikosida terhadap perlakuan naungan di dataran rendah dan tinggi pada
pegagan masih belum banyak dilaporkan. Pengaruh naungan tersebut sangat
penting untuk dipelajari karena kondisi yang ideal untuk tanaman obat adalah
kombinasi biomassa dan bahan aktif yang tinggi.
Berdasarkan hal tersebut di atas, pada penelitian ini akan dilakukan studi
keragaman dan tanggap pertumbuhan serta produksi asiatikosida beberapa aksesi
pegagan pada naungan dan ketinggian tempat yang berbeda. Informasi tersebut
sangat penting bagi pengembangan bahan tanaman pegagan yang memiliki kadar
dan produksi asiatikosida yang tinggi.
Tujuan Penelitian
Secara umum, penelitian ini bertujuan untuk mendapatkan informasi
mengenai keragaman plasma nutfah pegagan dan mendapatkan aksesi serta
naungan terbaik di dataran rendah dan tinggi dalam rangka menghasilkan
produksi asiatikosida yang tinggi. Penelitian ini terdiri dari beberapa tahapan
percobaan yang bertujuan untuk: (1) mendapatkan informasi parameter genetik
plasma nutfah pegagan, mendapatkan informasi tentang karakter yang dapat
digunakan sebagai kriteria seleksi kadar asiatikosida dan produksi terna kering
yang tinggi, mendapatkan aksesi pegagan dengan kadar asiatikosida dan produksi
terna kering yang tinggi, serta membandingkan hasil seleksi plasma nutfah
pegagan berdasarkan seleksi penyisihan bebas bertingkat, seleksi tunggal, indeks
seleksi terboboti dan tidak terboboti, (2) menganalisis dan mengevaluasi jarak
kedekatan genetik pada 17 aksesi pegagan berdasarkan penanda morfologi,
pertumbuhan dan produksi asiatikosida 6 aksesi pegagan terhadap perlakuan
naungan di dataran rendah dan tinggi.
Manfaat Penelitian
Manfaat dari hasil penelitian ini adalah diperoleh: (1) informasi tentang
keragaman genotipik, diketahui karakter yang dapat digunakan sebagai kriteria
seleksi untuk perbaikan kadar asiatikosida dan produksi terna kering serta
didapatkan aksesi pegagan dengan kadar asiatikosida dan produksi terna kering
yang tinggi dan informasi tentang hasil seleksi plasma nutfah pegagan
berdasarkan seleksi penyisihan bebas bertingkat, seleksi tunggal, indeks seleksi
terboboti dan tidak terboboti, (2) informasi jarak kedekatan genetik 17 aksesi
pegagan berdasarkan penanda morfologi, penanda RAPD, dan data gabungan,
dan (3) ilmu pengetahuan untuk meningkatkan produksi asiatikosida yang tinggi
melalui perlakuan naungan dan ketinggian tempat.
Hipotesis Penelitian
Terdapat keragaman dan perbedaan tanggap pertumbuhan serta produksi
asiatikosida pegagan pada naungan dan ketinggian tempat yang berbeda. Setiap
tahapan percobaan dapat ditarik beberapa hipotesis sebagai berikut:
1. Terdapat keragaman genotipik dari plasma nutfah pegagan, diperoleh karakter
agronomi yang dapat dijadikan sebagai kriteria seleksi untuk perbaikan
kadar asiatikosida dan produksi terna kering serta didapatkan aksesi pegagan
dengan kadar asiatikosida dan produksi terna kering yang tinggi, dan diketahui
urutan peringkat aksesi berdasarkan seleksi penyisihan bebas bertingkat,
seleksi tunggal, indeks seleksi terboboti dan tidak terboboti.
2. Diperoleh jarak kedekatan genetik 17 aksesi pegagan berdasarkan penanda
morfologi, penanda RAPD, dan data gabungan.
3. Terdapat perbedaan tanggap pertumbuhan dan produksi asiatikosida 6 aksesi
Ruang Lingkup Penelitian
Untuk mencapai tujuan penelitian di atas, maka strategi penelitian yang
dilakukan harus mempunyai keterkaitan antara penelitian yang satu dengan
penelitian lainnya. Penelitian ini terdiri atas dua aspek: (1) keragaman plasma
nutfah pegagan dan (2) tanggap perubahan karakter pegagan. Kedua aspek
tersebut dikelompokkan menjadi tiga subjudul penelitian: (1) pendugaan
parameter genetik, analisis lintas, dan seleksi plasma nutfah pegagan (Centella asiatica(L.) Urban), (2) keragaman plasma nutfah pegagan (Centella asiatica(L.) Urban) berdasarkan penanda morfologi, molekuler, dan gabungan, dan (3)
tanggap pertumbuhan dan produksi asiatikosida beberapa aksesi pegagan
(Centella asiatica (L.) Urban) pada naungan yang berbeda di dataran rendah dan tinggi.
Keragaman genetik dan heritabilitas beberapa karakter kuantitatif pegagan
dan karakter yang dapat dijadikan sebagai kriteria seleksi untuk meningkatkan
kadar asiatikosida dan produksi terna kering berdasarkan nilai korelasi, pengaruh
langsung dan tidak langsung serta nilai heritabilitas dipelajari dalam subjudul
penelitian pertama. Selain itu, pada penelitian pertama juga dilakukan seleksi
plasma nutfah pegagan untuk mendapatkan aksesi pegagan yang mempunyai
kadar asiatikosida dan produksi terna kering tinggi berdasarkan seleksi penyisihan
bebas bertingkat dan membandingkannya dengan seleksi berdasarkan kadar
asiatikosida, indeks seleksi terboboti dan tidak terboboti. Analisis kedekatan
genetik untuk mengetahui tingkat kedekatan atau keragaman genetik dari koleksi
plasma nutfah pegagan berdasarkan penanda morfologi, molekuler dan gabungan
dilakukan pada subjudul penelitian kedua. Aksesi terpilih dari hasil seleksi
berdasarkan kadar asiatikosida dan produksi terna kering yang dilakukan pada
subjudul pertama dan telah diketahui tingkat kedekatannya dari penelitian pada
subjudul penelitian kedua dipelajari pertumbuhan dan produksi asiatikosidanya
pada taraf naungan yang berbeda di dataran rendah dan tinggi dalam penelitian
ketiga.
Kajian pada subjudul penelitian 1 dan 2 dilakukan terhadap 17 aksesi
pegagan dengan perlakuan naungan paranet 25%. Subjudul penelitian 3, kajian
sebagai pembanding) yang dilakukan pada tiga tingkat naungan (tanpa naungan,
naungan paranet 25 dan 55%) dan ketinggian tempat yang berbeda (dataran
rendah dan tinggi). Garis besar seluruh kegiatan penelitian disajikan dalam
TINJAUAN PUSTAKA
Klasifikasi, Botani, dan Syarat Tumbuh Pegagan
Pegagan (Centella asiatica (L.) Urban) sin. Hydrocotyle asiatica (L.) termasuk ke dalam divisi Spermatophyta, kelas monocotiledoneae, ordo
Umbillales, dan famili Umbelliferae atau Apiaceae. Tanaman ini berasal dari
daerah Asia Tropik, tersebar luas di Asia Tenggara (Indonesia), India, Tiongkok,
Jepang, dan Australia. Nama daerah atau lokalnya adalah pegagan, daun kaki
kuda, daun penggaga, rumput kaki kuda, pegagan, kaki kuda, pegago, pugago
(Sumatera); cowet gompeng, antanan, antanan bener, antanan gede (Sunda);
gagan-gagan, ganggagan; kerok batok, panegowang, panigowang, rendeng,
calingan rambat, pacul gowang, gan gagan (Jawa); bebele, paiduh, penggaga,
kelai lere (Nusa Tenggara); sarowati, kolotidi manora (Maluku); pagaga,
wisu-wisu, cipubalawo, hisu-hisu, (Sulawesi); dogauke, gogauke, sandanan (Papua)
(Departemen Kesehatan RI 1977; Winarto & Surbakti 2003).
Tanaman pegagan merupakan herba menahun yang tidak berbatang
dengan akar rimpang pendek dan akar merayap atau menjalar, dengan panjang
stolon yang bisa mencapai 2,5 m (De Paduaet al. 1999). Akar terdapat pada buku yang menyentuh tanah, akarnya tunggang bercabang-cabang, sedangkan akar
serabut tumbuh dari buku-buku stolon (geragih). Daun tunggal, letak basalis atau
rosette dengan 2-10 daun. Bentuk daun seperti ginjal (reniformis), ukuran 2-5 cm
x 3-7 cm, tangkai daun tegak dan sangat panjang ukurannya 9-17 cm, bagian
dalam tangkai daun berlubang. Tepi daun bergerigi dengan penampang 1-7 cm
dan kadang berambut (Wijayakusuma et al. 1994). Pangkal dari tangkai daun melekuk ke dalam dan melebar seperti pelepah. Tulang daun menjari (palmitus).
Helaian daun biasanya berwarna hijau dan hijau muda. Bunga putih atau merah
muda berbentuk payung, tunggal atau 3-5 bunga secara bersama keluar dari ketiak
daun (Santa & Prayogo 1992; Wijayakusuma et al. 1994), dengan tangkai bunga (pedunculus) lebih pendek daripada tangkai daun. Buahnya kecil bergantung
jumlah kromosom pegagan adalah 2n=18, 22, 22+1B, 22+2B, dan 33 (Mitsukuri
& Kurahori 1959; Bell 1960; Das & Mallick 1991).
Pegagan dapat diperbanyak secara vegetatif dengan tunas akar serta dapat
pula diperbanyak dengan biji atau secara generatif. Hingga saat ini perbanyakan
menggunakan stek tunas akar lebih banyak dilakukan dibandingkan perbanyakan
dengan biji. Perbanyakan dengan biji atau benih jarang dan bahkan belum pernah
dilakukan, karena selain ukuran bijinya yang terlalu kecil juga sangat sulit untuk
mendapatkan biji tersebut (Januwati & Muhammad 1992).
Pegagan merupakan tumbuhan kosmopolit atau memiliki daerah
penyebaran sangat luas, terutama di daerah tropis dan subtropis. Tanaman ini
tumbuh dengan baik di tempat-tempat terbuka atau cukup sinar matahari atau agak
terlindung yang tanahnya subur dan agak lembab. Pegagan menyebar liar dan
dapat tumbuh dari dataran rendah sampai dataran tinggi sampai dengan ketinggian
1-2500 m dpl, namun tanaman ini tumbuh baik di dataran menengah pada
ketinggian sekitar 700 m dpl. Kelembaban udara yang diinginkan antara 70-90%
dengan rata-rata temperatur udara antara 20-250C dan tingkat kemasaman tanah
netral (pH) antara 6-7 (LBN 1980; Dalimartha 2000; Winarto & Surbakti 2002).
Manfaat dan Kandungan Kimia
Di Jawa Barat, kadang-kadang pegagan dipakai sebagai tanaman penutup
tanah perkebunan teh. Di Indonesia, sejak dulu rebusan daunnya digunakan untuk
bermacam-macam penyakit antara lain untuk mengobati keracunan jengkol,
peluruh air seni, dan diaforetika, penyakit saluran empedu, wasir, batuk kering
pada anak-anak, pendarahan hidung, tukak lambung, sakit ginjal dan sebagai obat
kumur pada sariawan (LBN 1980). Selain itu, pegagan digunakan untuk obat
diare, radang usus, bronchitis dan keputihan. Penggunaan lokal, yaitu untuk
mengobati pembengkakan buah zakar, kaki gajah, luka baru atau borok (Heyne
1987). Di India pegagan digunakan untuk mengobati sipilis dan lepra (Martindale
1967).
Kandungan utama pegagan adalah triterpen asam asiatat dan asam
madekasat, serta turunan dari triterpen ester glikosida (tidak kurang dari 2%),
oksi-asiatikosida, asam madesiatat, asam brahmat, asam sentelat, asam sentoat,
asam indosentoat, asam thankunat, asam isobrahmat, asam sentat, madekassol,
sentellosa, dan alkaloid (Anonim 1993, 1999).
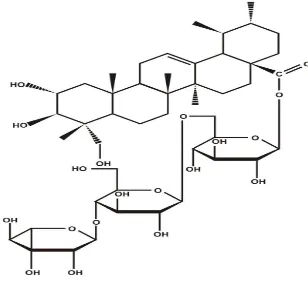
Asiatikosida (C48H78O19) termasuk ke dalam golongan glikosida
triterpenoid turunan dari-amyrin dengan molekul gula, terdiri atas 2 glukosa dan
1 rhamnosa. Aglikon triterpennya disebut asam asiatikat yang mempunyai gugus
alkohol primer, glikol, dan sebuah karboksilat teresterifikasi dengan gugus gula
(Vickery & Vickery 1981; Talalaj & Czechowics 1989; Maeda et al. 1994). Stuktur kimia asiatikosida dapat dilihat pada Gambar 2.
Gambar 2 Struktur kimia dari asiatikosida (James & Dubery, 2011)
Triterpenoid merupakan senyawa terpenoid yang kerangka karbonnya
berasal dari enam satuan isoprena dan secara biosintesis diturunkan dari
hidrokarbon C30 asiklik. Seluruh senyawa terpenoid yang ada di alam dibangun
dari kondensasi unit isoprena aktif yang disebut isopentenil pirofosfat (IPP) dan
dimetilalil pirofosfat (DMAPP). Dua lintasan biosintesis yaitu lintasan mevalonat
dan non-mevalonat (deoksisilulosa difosfat: DXP) adalah lintasan yang penting
dalam biosintesis isoprena. Tumbuhan ini memiliki kedua jalur biosintesis
isoprena tersebut secara bersamaan, perbedaannya hanya pada organ sel tempat
berlangsungnya proses reaksi biosintesis. Beberapa data baru memperlihatkan
bahwa proses biosintesis isoprena melalui mevalonat lebih aktif terjadi pada
diterpena. Jalur biosintesis isoprena melalui non mevalonat terjadi di plastida
(Agusta 2006).
Goldstein dan Brown (1990), Dewick (1997), serta Burke et al. (1999) menuliskan bahwa pada lintasan mevalonat, proses pertama meliputi reaksi
kondensasi dua molekul asetil-coenzim A (asetil-CoA) menjadi asetoasetil-CoA
yang dikatalisasi oleh enzim asetil-CoA asetiltransferase. Selanjutnya
asetoasetil-CoA berkondensasi lagi dengan satu unit asetil-asetoasetil-CoA untuk membentuk molekul
-hidroksi--metilgutaril-CoA (CoA) yang dikatalisasi oleh enzim
HMG-CoA sintase. Menurut Choi et al. (1992), Newmann dan Chappel (1999), serta Schnee et al. (2002) bahwa enzim HMG-CoA tersebut pada tumbuhan terdapat pada retikulum endoplasma yang regulasinya dapat dipicu oleh adanya luka pada
organ tumbuhan atau terjadinya infeksi oleh patogen. Proses kedua adalah reduksi
HMG-CoA oleh NADPH dengan katalisasi oleh enzim HMG-CoA reduktase
menjadi asam mevalonat (MVA). Proses berikutnya, dengan bantuan enzim
mevalonat kinase dan enzim fosfomevalonat kinase, asam mevalonat dikonversi
menjadi 5-fosfomevalonat dan 5-difosfomevalonat. Selanjutnya enzim
difosfomevalonat dekarboksilase akan mengubah 5-difosfomevalonat menjadi
isopentenil pirofosfat (IPP). Proses selanjutnya IPP dengan bantuan enzim IPP
isomerase akan membentuk reaksi kesetimbangan menjadi dimetilalil pirofosfat
(DMAPP). Kondensasi IPP dan DMAPP akan membentuk geranil pirofosfat
(GPP, C-10) yang merupakan senyawa antara untuk semua monoterpen.
Penggabungan satu unit IPP dengan GPP menghasilkan Farnesil pirofosfat (FPP,
C-15) yang merupakan senyawa antara bagi sesquiterpenoid, masing-masing
dikatalisasi oleh geranil pirofosfat sintase dan farnesil pirofosfat sintase. Senyawa
triterpenoid terbentuk jika dua molekul sesquiterpenoid bergabung.
Senyawa-senyawa triterpenoid berasal dari MVA melalui skualene dan biasanya melalui 2,3
epoksiskualen, kemudian terbentuklah senyawa dammarenediol. Lupeol dan
-amyrin dibiosintesis dari dammarene, dari -amyrin selanjutnya akan diperoleh
derifat dari-amyrin, yaitu asiatikosida (Manitto 1981; Vickery & Vickery 1981;
IPP FPS SQS SQE OSC
+ FPP Squalen 2,3-Oxidosqualen [Dammarenyl kation] DMAP
Betulinic acid lupeol [lupenyl kation]
Asiatic acid -Amyrin
/-AS [oleanyl kation]
Centellasapogenol A -Amyrin
Keterangan:
FPS : Farnesyl diphosphate synthase SQS : Squalen synthase
SQE : Squalen epoksidase OSC : Oxidosqualen cyclase
/-AS:/-amyrin synthase
Gambar 3 Skema biosintesis triterpenoid pada pegagan atauCentella asiatica(L.) Urban (Haralampidiset al. 2002; Phillipset al. 2006; James & Dubery 2009)
Parameter Genetik, Analisis Lintas, dan Seleksi
Parameter genetik
Pendugaan parameter genetik penting dalam proses pemuliaan tanaman
karena terkait dengan proses seleksi dalam tahapan selanjutnya. Beberapa
parameter genetik yang umum diduga dalam penelitian pemuliaan adalah
keragaman dan heritabilitas.
Seleksi merupakan dasar dari seluruh perbaikan tanaman untuk
mendapatkan varietas unggul baru. Dalam perakitan varietas unggul, keragaman
genetik memegang peranan yang sangat penting karena semakin tinggi keragaman
genetik semakin tinggi pula peluang untuk mendapatkan sumber gen bagi karakter
yang akan diperbaiki. Disamping itu, keragaman genetik yang tinggi juga dapat
meningkatkan respons seleksi, karena respons seleksi berbanding lurus dengan
keragaman genetik (Kuswantoro et al. 2006). Ragam yang diukur dari suatu populasi untuk karakter tertentu merupakan ragam fenotipe. Ragam fenotipe
terdiri dari ragam genetik, ragam lingkungan, serta ragam interaksi antara genetik
(σ2A), ragam genetik dominan (σ2D), dan ragam genetik epistasis (σ2E) atau σ2G=
σ2A+σ2D+σ2E.
Heritabilitas adalah hubungan antara ragam genetik dengan ragam
fenotipenya, hubungan ini menggambarkan seberapa jauh fenotipe yang tampak
merupakan refleksi dari genotipe. Berdasarkan komponen ragamnya, heritabilitas
dibedakan menjadi heritabilitas dalam arti luas (broad sense heritability) dan heritabilitas dalam arti sempit (narrow sense heritability). Heritabilitas dalam arti luas merupakan perbandingan antara ragam genetik total dan ragam fenotipe
(h2(BS)=σ2G / σ2P), sedangkan heritabilitas arti sempit merupakan perbandingan
antara ragam aditif dan ragam fenotipe (h2(NS)=σ2A/ σ2P)(Mangoendidjojo 2003).
Secara teori nilai heritabilitas berkisar antara nol sampai dengan satu. Kategori
mengenai besar kecilnya nilai heritabilitas, dikemukakan oleh Whirter (1979)
yaitu: 0.50 h21.00 (tinggi); 0.20 h2 0.50 (sedang) dan 0.00 h2 0.20 (rendah), heritabilitas biasanya dinyatakan dalam persen (%). Heritabilitas dapat
menduga peningkatan kemajuan genetik yang mungkin diperoleh bila dilakukan
seleksi terhadap sifat tertentu. Jika heritabilitas suatu sifat memiliki nilai tinggi
berarti penampilan indivitu lebih banyak dipengaruhi oleh faktor genetik
dibanding dengan faktor lingkungan dan seleksi berdasarkan individu efektif.
Heritabilitas tinggi juga menandakan aksi gen aditif penting untuk sifat tersebut
dan sebaliknya jika heritabilitas rendah, maka mungkin aksi gen seperti lewat
dominan (over dominance), dominan, dan epistasis lebih penting (Lasley, 1978).
Analisis Lintas
Analisis lintas merupakan pengembangan metode analisis korelasi.
Analisis lintas dapat menjelaskan keeratan hubungan antar karakter dengan cara
menguraikan koefisien korelasi menjadi pengaruh langsung dan tidak langsung.
Jika dibandingkan dengan analisis korelasi, maka analisis lintas tidak hanya
memberikan informasi tentang keeratan hubungan antar karakter, tetapi juga
menjelaskan mekanisme hubungan kausal antar karakter. Mekanisme hubungan
kausal tersebut diperoleh dari penguraian koefisien korelasi menjadi pengaruh
langsung dan tidak langsung masing-masing karakter melalui karakter lain
dalam menarik kesimpulan pada analisis lintas, yaitu: 1) Jika koefisien korelasi
(rxij) hampir sama besar dengan pengaruh langsungnya (C) maka koefisien
korelasi tersebut seutuhnya mengukur derajat keeratan hubungan Xi dan Y,
artinya seleksi berdasarkan variabel Xi sangat efektif, 2) Jika koefisien korelasi
(rxij) bernilai positif tapi pengaruh langsungnya negatif atau dapat diabaikan, maka
pengaruh tidak langsungnya (Cirij) menjadi penyebab korelasi. Semua variabel
bebas X harus diperhatikan dan diperhitungkan secara serempak, dan 3) Jika
koefisien korelasi (rxij) bernilai negatif, tapi pengaruh langsungnya (C) bernilai
positif dan besar, maka pengaruh tidak langsung yang tidak dikehendaki dibatasi,
sehingga dalam penafsirannya pengaruh langsung benar-benar dapat
dimanfaatkan.
Analisis lintas sangat bermanfaat dalam menentukan strategi pemuliaan
tanaman yang efektif dan efisien. Penggunaan analisis lintas untuk
mengembangkan kriteria seleksi telah banyak dilakukan pada berbagai jenis
tanaman, seperti pada kedelai, gandum, sorgum, cabe, peartmillet (Asadi et al. 2004; Budiartiet al. 2004; Bizetiet al. 2004; Ezeaku 2006; Ganefiantiet al. 2006; Wirnas et al. 2006; Vetriventhan & Nirmalakumari 2007). Bizeti et al. (2004) telah berhasil mengembangkan kriteria seleksi yang efektif untuk meningkatkan
daya hasil pada kedelai. Kriteria seleksi yang digunakan adalah jumlah buku per
tanaman karena pada setiap buku akan terdapat polong.
Seleksi
Seleksi merupakan satu tahapan untuk mendapatkan genotipe yang sesuai
dengan target lingkungan produksi. Pada dasarnya seleksi terbagi atas seleksi
langsung dan seleksi tidak langsung. Seleksi langsung diartikan sebagai pemilihan
genotipe-genotipe terbaik berdasarkan karakter-karakter yang memenuhi kriteria
seleksi, sedangkan seleksi tidak langsung diartikan sebagai pemilihan secara tidak
langsung genotipe-genotipe terbaik berdasarkan karakter-karakter yang memiliki
hubungan dengan tujuan akhir dari program pemuliaan. Seleksi langsung
dikategorikan ke dalam seleksi langsung berdasarkan satu sifat dan seleksi
langsung terhadap beberapa sifat. Seleksi beberapa sifat secara simultan yang
seleksi simultan (independent culling level), dan seleksi indeks (Bariet al. 1981). Seleksi indeks adalah yang terbaik karena dalam penyusunan indeks
mempertimbangkan koefisien korelasi fenotipik, koefisien korelasi genotipik, dan
nilai heritabilitas. Selain itu, diperhitungkan pula ragam fenotipik dan peragam
genotipik serta nilai ekonomi setiap sifat. Bobot ekonomis dalam seleksi indeks
diharapkan dapat memberikan keseimbangan pada perolehan nilai genetik yang
proporsional (Smith 1936, diacu dalam Sutresna 2008). Namun demikian terdapat
beberapa kelemahan dalam seleksi indeks yaitu permasalahan dalam perolehan
ragam dan peragam, kemungkinan perubahan parameter dengan seleksi dan
kesulitan penetapan kepentingan relatif suatu sifat (Lin, 1978). Sutresna (2008)
menyatakan bahwa kriteria seleksi dengan menggunakan indeks seleksi
menunjukkan efisiensi yang lebih tinggi dibandingkan dengan seleksi yang
dilakukan terhadap satu sifat. Keunggulan indeks seleksi akan meningkat dengan
peningkatan jumlah sifat yang diseleksi diantara sifat tersebut, dan menurun
dengan peningkatan perbedaan-perbedaan kepentingan masing-masing sifat secara
nisbi. Menurut Moeljoprawiro (2002), bahwa indeks seleksi padi yang dihasilkan
terhadap beberapa sifat secara simultan lebih efisien dibandingkan dengan seleksi
yang didasarkan atas satu atau kombinasi dari dua sifat. Indeks seleksi yang
melibatkan sifat panjang malai, panjang gabah, dan lebar gabah lebih berpeluang
untuk perbaikan hasil tanaman.
Penanda Genetik dalam Identifikasi Keanekaragaman Pegagan
Pengungkapan informasi sifat genetik tanaman dapat dilakukan dengan
melakukan identifikasi dari setiap aksesi plasma nutfah. Identifikasi untuk
pembedaan identitas antar varietas dapat dideteksi melalui beberapa penanda
genetik (genetic marker). Penanda genetik merupakan karakter yang dapat diturunkan yang berhubungan dengan genotipe tertentu dan dapat digunakan
untuk mengkarakterisasi genotipe tersebut (Asieduet al. 1989)
Penanda genetik dapat digolongkan atas penanda morfologi (Livneh &
Vardi 1998), penanda agronomi (Melchinger 1990), penanda isoenzim dan
penanda agronomi dimasukkan pada kategori penanda fenotipik, sedangkan
penanda isoenzim dimasukkan sebagai penanda protein atau biokimia.
Penanda morfologi
Penanda morfologi digunakan dalam deskripsi taksonomi karena lebih
mudah, lebih cepat, sederhana dan lebih murah (Cross 1990). Disamping itu,
dalam proses koleksinya tidak membutuhkan teknologi yang mahal (Maxtedet al. 1997). Sifat-sifat morfologi yang diamati haruslah sifat-sifat yang memiliki nilai
heritabilitas yang tinggi dan stabil pada beberapa lokasi percobaan karena
umumnya penampakan sifat pada morfologi tanaman sangat dipengaruhi
lingkungan. Penanda morfologi yang selama ini hanya dipergunakan sebagai cara
cepat untuk mengidentifikasi varietas diharapkan dapat digunakan untuk menilai
kekerabatan antar aksesi sehingga lebih bermanfaat dalam program pemuliaan
tanaman, sedangkan penanda agronomi lebih dipertimbangkan dalam usaha
pemilihan tetua persilangan berdasarkan sifat hasil dan komponen hasil.
Setiap spesies tanaman mempunyai deskripsi morfologi yang spesifik.
Deskripsi khusus tanaman tersebut telah diterbitkan oleh International Plant Genetic Resources Institute (IPGRI) untuk memudahkan dalam identifikasi karakter morfologi dan agronomi tanaman.
Penanda morfologi ini telah lama dan banyak digunakan terutama untuk
mengatasi masalah duplikasi plasma nutfah di lapang (Simmond & Sheperd
1955). Disamping itu juga digunakan untuk identifikasi kekerabatan dan
keragaman genetik antar klon/kultivar dan masih terus digunakan sampai saat ini.
Penanda molekuler
Keunggulan penanda molekuler adalah kemampuan membedakan setiap
spesies tanaman atau genotipe tanaman tanpa dipengaruhi lingkungan. Umumnya
sifat kuantitatif pada tanaman dikendalikan oleh banyak gen (poligen) dan
dipengaruhi oleh faktor lingkungan sehingga perbedaan antara klon atau spesies
berkerabat dekat sulit untuk diamati. Dengan kemajuan dalam bidang biologi
molekuler permasalahan di atas telah terbukti mampu diatasi. Penggunaan
penanda molekuler sangat bermanfaat untuk membandingkan berbagai klasifikasi
dengan analisis berdasarkan penanda lainnya, seperti SSR (Simple Sequence Repeat), RFLP (Restriction Fragment Length Polymorphism) dan AFLP (Amplified Fragment Length Polymorphism) sehingga hasil klasifikasi lebih akurat (Crouchet al. 1998).
Pemanfaatan penanda molekuler DNA akhir-akhir ini lebih banyak
digunakan karena memiliki beberapa kelebihan dibandingkan penanda morfologi
dan isoenzim (Wattimena 1999), yaitu: (1) sangat akurat dan tidak dipengaruhi
oleh lingkungan yang mempengaruhi ekspresi dari gen tersebut, (2) dapat diuji
pada semua tingkat perkembangan tanaman, (3) pada pengujian ketahanan hama
dan penyakit tidak bergantung pada organisme penganggu, dan (4) seleksi pada
tingkat genotipe dapat mempercepat proses seleksi dan hemat pada pengujian
selanjutnya di lapang.
Penanda RAPD
RAPD merupakan salah satu metode yang sering digunakan untuk analisis
profil DNA genom. Teknik RAPD ini merupakan suatu metode analisis DNA
genom dengan cara melihat pola pita DNA yang dihasilkan setelah DNA genom
diamplifikasi menggunakan primer acak. Metode ini didasarkan pada teknik
polimerisasi berantai (PCR).
Secara umum RAPD lebih mudah, lebih murah dan tekniknya lebih cepat
dibandingkan RFLP, SSR dan AFLP (Darmono 1996). Hal ini karena adanya
beberapa alasan: (1) tidak memerlukan pengetahuan latar belakang genom yang
akan diteliti, (2) primer secara universal dapat digunakan untuk organisme
prokariot maupun eukariot, (3) mampu menghasilkan karakter yang relatif tidak
terbatas jumlahnya, (4) bahan-bahan yang digunakan relatif murah, (5) mudah
dalam hal preparasi, dan (6) memberikan hasil lebih cepat dibandingkan dengan
analisis keragaman molekuler lainnya.
Disamping kelebihan, teknik RAPD juga memiliki kelemahan, yaitu: (1)
pemunculan pita DNA kadang-kadang tidak konsisten, hal ini lebih sering terjadi
jika suhu annealing (penempelan primer) yang digunakan terlalu tinggi. Dalam analisis kekerabatan hal ini dapat diatasi dengan menggunakan primer yang lebih
diperlukan dalam analisis sangat bergantung pada tujuan atau jenis informasi yang
diinginkan. Jika tujuannya untuk mengungkapkan hubungan kekerabatan, maka
analisisnya tidak dapat bertumpu pada satu atau beberapa karakter saja. Halldenet al. (1994) menyatakan bahwa semakin banyak jumlah primer yang digunakan semakin rendah nilai koefisien keragaman hasil analisis yang diperoleh. Sepuluh
sampai 20 primer dianggap sudah mencukupi untuk keperluan analisis
kekerabatan karena dengan 10 primer pengaruh kesalahan percobaan telah dapat
diperkecil hingga mendekati nol.
Pengaruh Naungan Terhadap Pertumbuhan dan Produksi Tanaman
Salisbury dan Ross (1992) menyatakan bahwa cahaya matahari
mempunyai peranan dalam proses fisiologi tanaman seperti fotosintesis, respirasi,
pertumbuhan dan perkembangan, menutup dan membukanya stomata, dan
perkecambahan tanaman. Cahaya matahari berperan penting dalam metabolisme
tanaman hijau, sehingga ketersediaan cahaya matahari menentukan tingkat
produksi tanaman. Tanaman hijau memanfaatkan cahaya matahari melalui proses
fotosintesis. Naungan merupakan salah satu faktor yang membatasi proses
fotosintesis.
Penurunan intensitas cahaya akan mempengaruhi pertumbuhan dan hasil
tanaman serta terganggunya berbagai proses metabolisme tanaman. Beberapa
hasil penelitian tentang pengaruh naungan telah banyak dilaporkan. Rachmawaty
(2005) melaporkan bahwa tanaman pegagan dapat tumbuh baik pada tingkat
naungan 25%, sedangkan pada naungan 75% terjadi penurunan produksi pegagan.
Penelitian Tarore (1992) pada kencur menunjukkan bahwa naungan berpengaruh
nyata terhadap pertumbuhan dan hasil tanaman kencur. Naungan 40%
menghasilkan luas daun, bobot kering daun, kadar klorofil total yang lebih tinggi
dibanding dengan perlakuan naungan 60% dan tanpa naungan. Tingkat naungan
60% memberikan hasil tanaman terendah.
Asadiet al. (1997) melaporkan bahwa penurunan hasil biji kedelai dari 28 galur yang diuji di bawah naungan 33% berkisar 2-45% dibandingkan dengan
tanpa naungan, sedangkan Sunarlim (1997) melaporkan bahwa dengan pemberian
sebelumnya juga telah diketahui bahwa, pemberian naungan dapat mempengaruhi
kandungan bioaktif tanaman.
Pada tanaman daun jinten, kadar kumarat dan fanilat tertinggi terdapat
pada naungan 75% (Urnemi et al. 2002). Pigmentasi antosianin pada kedelai meningkat pada persentase naungan yang semakin tinggi (Muhuria et al. 2006). Pada penelitian periode pencahayaan, Nirwan (2007) melaporkan bahwa naungan
50% selama 1 dan 3 bulan pada cahaya 100% dapat menghasilkan kandungan
total flavonoid dan antosianin daun dewa tertinggi. Periode pencahayaan yang
menghasilkan produksi total flavonoid dan kuersetin per tanaman tertinggi adalah
naungan 50% selama 3 dan 1 bulan cahaya 100%, sedangkan produksi antosianin
tertinggi diperoleh pada naungan 25% selama 2 bulan dan 2 bulan cahaya 100%.
Bermawieet al. (2006b) melaporkan bahwa pada tingkat naungan 45% dan tanpa naungan, kadar asiatikosida dari tiga aksesi pegagan yang dipanen seluruh bagian
tanaman bervariasi antara 1.01-1.89%, sedangkan pegagan yang dipanen daun dan
tangkainya bervariasi antara 0.16-1.16%. Pada tanaman pule pandak (Raufolia serpentina), Sulandjari et al. (2005) melaporkan bahwa pada naungan 50% sampai dengan 80%, kadar reserpina lebih tinggi daripada naungan 20%, tetapi
bobot akar pertanaman tertinggi diperoleh pada tingkat naungan 20%.
Tanaman akan memberikan tanggap terhadap naungan, tanggap
pertumbuhan tanaman yang ternaungi dapat dilihat pada Tabel 1 (Anderson &
Tabel 1 Karakter morfologi dan fisiologi tanaman ternaungi
No. Karakter morfologi No. Karakter fisiologi
1. Batang lebih kecil karena xilem kurang 1. Kandungan klorofil lebih berkembang tinggi
2. Luas daun per tanaman lebih besar 2. Laju fotosintesis lebih rendah 3. Jarak antara buku menjadi lebih panjang 3. Laju respirasi lebih rendah 4. Jumlah cabang lebih sedikit 4. Kandungan air lebih tinggi 5. Sel-sel pada daun berukuran lebih besar 5. Transpirasi lebih lambat
sehingga helai daun menjadi lebih besar dan tipis
6. Endodermis lebih berkembang 6. C/N rendah
7. Kutikula dan dinding lebih berkembang 7. Kemampuan berbunga dan 8. Kloroplas lebih banyak dan berukuran lebih berbuah kurang bagus
besar
9. Jaringan palisade kurang berkembang 8. Bunga muncul lebih lambat
10. Jaringan mesofil lebih berkembang 9. Kurang tahan terhadap
11. Jarak antar sel lebih besar stress suhu, kekeringan,
12. Akar lebih pendek dan ratio akar/tajuk dan penyakit lebih rendah
13. Pada tanaman legume, bintil akar lebih sedikit dan lebih kecil
Sumber: Anderson & Osmond (1987)
Mekanisme Adaptasi Tanaman Terhadap Intensitas Cahaya Rendah
Setiap tanaman mempunyai cara tersendiri dalam menghadapi kondisi
kekurangan cahaya. Tanaman dalam menghadapi cekaman terhadap kekurangan
cahaya memiliki dua cara penyesuaian, yaitu melalui mekanisme penghindaran
(avoidance) dan mekanisme toleransi (Levitt 1980). Mekanisme penghindaran berkaitan dengan perubahan anatomi dan morfologi daun agar lebih efisien dalam
melakukan fotosintesis. Pada umumnya kemampuan tanaman dalam mengatasi
cekaman naungan tergantung pada kemampuannya melakukan fotosintesis secara
normal pada kondisi kekurangan cahaya. Dengan penurunan titik kompensasi
cahaya yang lebih rendah, tanaman dapat mengakumulasi produk fotosintesis
pada level cahaya yang lebih rendah dibandingkan dengan yang dicapai oleh
Ketiadaan kutikula, lilin Ketiadaan pigmen
dan bulu pada permukaan daun non kloroplas
Peningkatan Peningkatan kandungan
kandungan kloroplas pigmen per kloroplas
Peningkatan kandungan Kloroplas dalam sel
kloroplas per sel mesofil epidermis
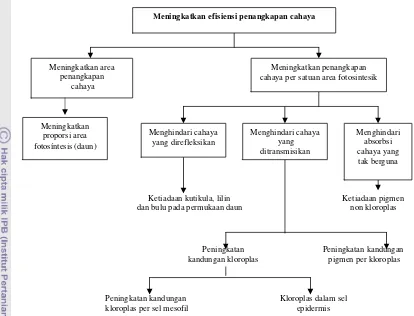
Gambar 4 Mekanisme penghindaran terhadap kekurangan cahaya (Levitt 1980)
Penghindaran kekurangan radiasi dapat dicapai dengan meningkatkan
efisiensi dari radiasi yang ditangkap tanaman, yaitu dengan cara: 1) meningkatkan
intersepsi cahaya total melalui luas penangkapan cahaya (luas daun), dengan
demikian maka daun-daun yang ternaungi menjadi tipis dengan permukaan
fotosintetik maksimum, dan 2) meningkatkan persentase cahaya yang diintersepsi
yang digunakan untuk fotosintesis, sehingga perlu mengurangi proporsi cahaya
yang direfleksi dan ditransmisikan (Hale & Orcutt 1987). Beberapa faktor yang
berperanan pada penghindaran kekurangan cahaya disajikan pada Gambar 4.
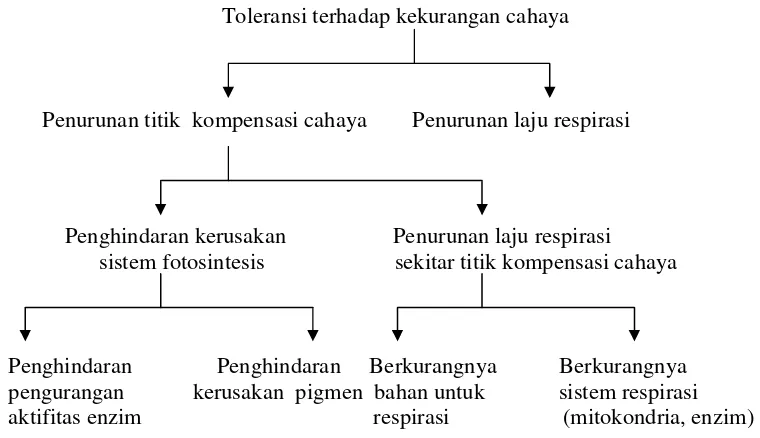
Toleransi terhadap kekurangan cahaya pada tanaman yang ternaungi dapat
dicapai dengan cara menurunkan titik kompensasi cahaya dan menurunkan laju
respirasi. Mekanisme toleransi kekurangan cahaya pada tanaman naungan
Toleransi terhadap kekurangan cahaya
Penurunan titik kompensasi cahaya Penurunan laju respirasi
Penghindaran kerusakan Penurunan laju respirasi
sistem fotosintesis sekitar titik kompensasi cahaya
Penghindaran Penghindaran Berkurangnya Berkurangnya pengurangan kerusakan pigmen bahan untuk sistem respirasi
aktifitas enzim respirasi (mitokondria, enzim)
Gambar 5 Mekanisme toleransi tanaman terhadap kekurangan cahaya (Levitt 1980)
Perubahan Anatomi dan Morfologi.Cahaya dapat mempengaruhi anatomi dan morfologi tanaman. Perubahan rasio luas daun terhadap bobot daun menunjukkan
adanya perubahan anatomi di dalam lapisan mesofil dan palisade. Pada kondisi
kurang cahaya atau ternaungi rasio luas daun terhadap bobot daun menjadi tinggi,
atau lapisan daun menjadi tipis. Pada umumnya lapisan palisade akan berkurang
dari 2-3 sel menjadi satu sel pada daun-daun yang ternaungi atau daun yang
resisten terhadap naungan (Fitter & Hay 1998). Taiz & Zeiger (1991) menyatakan
bahwa lapisan palisade dapat berubah sesuai kondisi cahaya agar tanaman
menjadi efisien dalam menyimpan energi cahaya yang dibutuhkan untuk
perkembangannya. Peran yang kontras antara sel palisade dan sel bunga karang,
yaitu sel palisade dapat menyebabkan cahaya lewat dan sel bunga karang
menangkap cahaya sebanyak mungkin, menyebabkan absorbsi cahaya yang lebih
seragam di dalam daun.
Perubahan Kandungan Klorofil. Kandungan kloroplas tanaman dipengaruhi oleh intensitas cahaya matahari, jumlahnya akan berbeda antara tanaman yang
mendapat sinar matahari penuh dengan tanaman yang dinaungi. Tanaman
ternaungi mempunyai grana yang lebih besar, sekitar 100 tilakoid per granum
rasio membrane tilakoid/stroma lebih besar sehingga kandungan klorofil per luas
daun lebih tinggi dibandingkan dengan tanaman yang mendapat sinar matahari
penuh.
Dalam keadaan normal, peralatan fotosintetik seperti klorofil akan
mengalami proses kerusakan, degradasi dan perbaikan. Kemampuan melawan
degradasi klorofil sangat penting dan akan menentukan daya adaptasi terhadap
naungan. Tanaman yang mampu beradaptasi pada kondisi kurang cahaya atau
ternaungi memiliki kemampuan meningkatkan jumlah kloroplas per satuan luas
daun (Hale & Orchut 1987) dan juga meningkatkan jumlah klorofil dalam
kloroplas (Okadaet al. 1992). Hal ini ditunjukkan oleh genotipe padi gogo toleran yakni memiliki kandungan klorofil a dan b lebih tinggi dibanding yang peka
(Chowdury et al. 1994; Sulistyono 1998). Sejalan dengan itu, Hidema et al. (1992) melaporkan bahwa terjadinya penurunan rasio klorofil a/b karena
meningkatnya klorofil b pada tanaman yang dinaungi, yang berkaitan dengan
peningkatan klorofil a/b pada LHC II (Light-Harvesting ComplexII). Penyesuaian tanaman terhadap radiasi yang rendah juga dicirikan dengan membesarnya antena
untuk fotosistem II (LHCIIb). Membesarnya antenna untuk fotosistem II akan
meningkatkan efisiensi pemanenan cahaya. LHCIIb merupakan kompleks trimerik
yang mengikat sekitar 60% klorofil PSII (PhotosystemII). Sebagian besar klorofil b merupakan komponen pemanenan cahaya dari PSII, maka perubahan pada
jumlah kompleks pemanen cahaya (LHC) pada PSII dan PSI. Khumaida (2002)
menyatakan bahwa genotipe kedelai toleran naungan memiliki kapasitas
penangkapan cahaya yang lebih besar dari pada genotipe peka karena memiliki
kemampuan yang lebih tinggi dalam mengkonversi klorofil a menjadi klorofil b.
Park et al. (1996) menyatakan bahwa PSII merupakan kompleks apparatus fotosintetik yang paling peka (vulnerable) terhadap stress cahaya. Perilaku PSII sangat ditentukan oleh dosis foton atau intensitas cahaya yang diterima. Taiz dan
Zeiger (1991) menyatakan bahwa klorofil b berfungsi sebagai antena yang
mengumpulkan cahaya untuk kemudian ditransfer ke pusat reaksi. Pusat reaksi
tersusun dari klorofil a. Energi cahaya akan diubah menjadi energi kimia di pusat
Penambahan klorofil b akan sangat bermanfaat bagi tanaman yang ternaungi
untuk memperoleh energi cahaya yang lebih banyak.
Perubahan Fisiologi dan Biokimia.Pada umumnya, kemampuan tanaman dalam mengatasi intensitas cahaya rendah tergantung pada kemampuan tanaman dalam
melakukan fotosintesis dalam kondisi defisit cahaya.
Faktor lain yang membatasi fotosintesis adalah resistensi stomata terhadap
CO2dan rendahnya aktivitas enzim rubisco pada kondisi ternaungi. Pertumbuhan
dan hasil tanaman pada kondisi intensitas cahaya rendah juga dipengaruhi oleh
kemampuan tanaman untuk memfiksasi CO2 yang ada di atmosfir. Jika
konsentrasi CO2 yang ada di udara rendah maka fotosintesis bersih (net
photosynthesis) antara yang ternaungi dengan yang kontrol hampir sama. Pada konsentrasi CO2 di udara yang sama maka tanaman yang dinaungi akan
memfiksasi CO2 jauh lebih sedikit dibandingkan dengan yang tidak ternaungi
karena resistensi stomata yang tinggi terhadap CO2 sehingga fotosintesis
bersihnya jauh lebih rendah dibandingkan dengan tanaman yang mendapat
intensitas cahaya penuh (Salisbury & Ross 1985; Mohr & Schoper 1995).
Intensitas cahaya rendah mengakibatkan perubahan fisiologi dan biokimia pada
tanaman. Rubisco merupakan enzim yang berperan dalam fiksasi CO2. Intensitas
cahaya rendah mengakibatkan penurunan kandungan dan aktivitas enzim rubisco
dan berbagai enzim lainnya (Bruggeman & Danborn 1993). Genotipe kedelai
yang toleran terhadap intensitas cahaya rendah memiliki aktivitas rubisco yang
lebih tinggi dibandingkan dengan yang peka pada kondisi ternaungi.
Lebih jauh intensitas cahaya rendah mengakibatkan penurunan karbohidrat
dalam tanaman, terutama fruktosa dan sukrosa dan selanjutnya akan terjadi
berbagai perubahan dalam proses metabolisme pada tanaman (Kephart et al. 1992). Hal ini disebabkan oleh tingginya resistensi stomata dan sel-sel mesofil
terhadap pertukaran gas CO2 serta penurunan kandungan dan aktivitas