KARAKTERISASI DAN EVALUASI AKSESI PAMELO
{C
itrus maxima
(Burm.) Merr.}
BERBIJI DAN TIDAK BERBIJI ASLI INDONESIA
ARIFAH RAHAYU
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
SUMBER INFORMASI
Dengan ini saya menyatakan bahwa disertasi Karakterisasi dan Evaluasi Aksesi Pamelo {Citrus maxima (Burm.) Merr.} Berbiji dan Tidak Berbiji Asli Indonesia adalah karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apapun kepada perguruan tinggi manapun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir disertasi ini.
Bogor, Agustus 2012
Arifah Rahayu
Indonesian Pummelo {Citrus maxima (Burm.) Merr.} Accessions. Under supervision of SLAMET SUSANTO as chairman, BAMBANG S. PURWOKO and ISWARI S. DEWI as members of the advisory committee.
Indonesia has many accessions of pummelo germplasms, but only a small number of the accessions are planted commercially. Genetic variation in the seed number per fruit in pummelo is high, ranging from seedless to very seedy. Thus pummelo is categorized as seeded, potentially seedless and seedless. The objectives of this research were: 1) to identify morphological, biochemical (isozyme) and molecular (ISSR) characters, and ploidy level of seeded and seedless pummelo accessions originated from Sumedang, Pati, Kudus and Magetan, 2) to identify fruit formation of seeded and seedless pummelo, 3) to evaluate fruit quality of seeded and seedless pummelo fruit quality during storage.
The characterization research showed that morphological characters contributed in grouping pummelo accessions were epicarp thickness, leaf lamina margin, vesicle length, epicarp color, petiole wing width and fruit shape, while based on isozyme characters were MDH (Rf 0.11 and 0.14) and ACP (Rf 0.24 and 0.33). ACP band at Rf 0.24 could be used as marker to differentiate seeded and seedless pummelo accessions. Molecular characters supported in classification of pummelo accessions were PKBT8 500 bp, PKBT8 375 bp, PKBT7 750 bp, PKBT3 750 bp, PKBT8 625 bp and PKBT12 500 bp. Separation between seeded and seedless cultivars based on morphological characters occured at similarity coefficient of 0.63, while on isoenzyme characters occured at similarity coefficient of 0.49. Both seeded and seedless pummelo accessions were diploid (2n = 2x = 18). Observation on fruit formation between selected seeded and seedless pummelo demonstrated that those pummelo accessions were able to produce parthenocarpic fruits, but bored seeded fruits when self pollination was induced, thus indicated self-compatibility. Natural self-pollination on ‘Bali Merah 1’, ‘Bali Merah 2, ‘Nambangan’ and ‘Bageng’ yielded seedless fruits, but most of ‘Jawa 3’ was seeded. Artificial self-pollination on those accessions produced seeded fruits, thus showed ovule fertility. ‘Bali Merah 2’ was shown to have high parthenocarpic degree as indicated by less seed number (<10 seed per fruit) in all pollination treatments. Therefore seedlessness in those pummelo accessions was caused by parthenocarpy, was not by pollen and ovule sterility, self-incompatibility, or poliploidy. Evaluation of pummelo fruit quality showed seedless pummelos have higher vitamin C, naringin content and acidity than seeded accessions. Vitamin C and titratable acidity (TA) content decreased, but total soluble solids (TSS) and TSS/AT ratio increased during storage period. During storage, percentage of edible portion increased, mainly due to the decrease of peel weight. Percentage of fruit axis of seedless accessions was smaller than those of seeded accessions. Sensory analysis indicated ‘Jawa 1’ seedless accessions have superior quality based on hedonic and scale quality test.
RINGKASAN
ARIFAH RAHAYU. Karakterisasi dan Evaluasi Aksesi Pamelo {Citrus maxima
(Burm.) Merr.} Berbiji dan Tidak Berbiji Asli Indonesia. Dibimbing oleh Slamet Susanto sebagai ketua, Bambang S Purwoko dan Iswari S Dewi sebagai anggota komisi pembimbing.
Indonesia memiliki banyak plasma nutfah pamelo, walaupun baru sebagian saja yang dikomersialkan. Pamelo dapat dikategorikan sebagai pamelo berbiji, potensial tidak berbiji dan tidak berbiji berdasarkan perbedaan jumlah biji per buah. Namun, untuk pasar pamelo segar, buah tidak berbiji merupakan sifat yang banyak diminati. Tanaman dianggap menghasilkan buah tidak berbiji jika mampu menghasilkan buah tanpa biji sama sekali, biji mengalami aborsi atau memiliki sejumlah biji yang tereduksi. Sementara itu, tanaman disebut potensial menghasilkan buah tidak berbiji, bila dalam satu pohon terdapat buah berbiji dan tidak berbiji.
Penelitian ini bertujuan untuk: (1) mengidentifikasi karakter morfologi, biokimia (dengan isoenzim), molekuler (dengan ISSR/Inter-Simple Sequence
Repeat), dan tingkat ploidi (set kromosom) tanaman pamelo berbiji dan tidak
berbiji; (2) mengidentifikasi pembentukan buah pamelo berbiji dan tidak berbiji melalui uji viabilitas tepung sari dan uji penyerbukan; (3) mengevaluasi karakter kualitas buah pamelo berbiji dan tidak berbiji yang berasal dari Sumedang, Kudus, Pati dan Magetan selama penyimpanan.
Hasil karakterisasi menunjukkan karakter morfologi yang berperan dalam pengelompokan pamelo adalah tebal epikarp, pinggiran helai daun, panjang kantong jus, warna kulit buah masak, lebar sayap daun dan bentuk buah, sedangkan karakter isoenzim adalah MDH (Rf 0.11 dan 0.14) dan ACP (Rf 0.24 dan 0.33). Pita ACP 0.24 dapat dijadikan penanda untuk membedakan aksesi berbiji dan tidak berbiji. Karakter molekuler yang berperan dalam pengelompokan adalah PKBT8 500 bp, PKBT8 375 bp, PKBT7 750 bp, PKBT3 750 bp, PKBT8 625 bp dan PKBT12 500 bp. Dendrogram berdasarkan karakter morfologi memisahkan kelompok aksesi berbiji dan tidak berbiji pada koefisien kemiripan 0.63, sedangkan berdasarkan isoenzim pada koefisien kemiripan 0.48. Dendrogram berdasarkan karakter morfologi dan isoenzim dapat membedakan antara aksesi pamelo berbiji, potensial tidak berbiji dan tidak berbiji. Hasil pemetaan komponen utama kongruen dengan dendrogram, yaitu dapat memisahkan aksesi berbiji maupun tidak berbiji, berdasarkan karakter morfologi, isoenzim maupun kombinasinya. Baik pamelo berbiji maupun tidak berbiji adalah diploid dengan jumlah kromosom 2n = 2x = 18.
menghasilkan buah berbiji dengan derajat yang berbeda, yang menunjukkan fertilitas ovul. ‘Bali Merah 2’ diduga memiliki derajat partenokarpi yang tinggi, dilihat dari jumlah biji yang rendah (<10 biji per buah) pada semua perlakuan penyerbukan. Dengan demikian pembentukan buah tidak berbiji pada aksesi pamelo disebabkan oleh sifat partenokarpik, karena aksesi pamelo memiliki tepung sari dan ovul yang fertil, bersifat self-compatible dan kromosomnya diploid.
Kualitas buah pamelo dipengaruhi oleh kelompok aksesi, aksesi dan masa simpan serta interaksi antara kelompok aksesi dengan masa simpan dan aksesi dengan masa simpan. Kandungan vitamin C, naringin dan pH jus buah pamelo aksesi tidak berbiji lebih tinggi dibandingkan aksesi berbiji. Penyimpanan menyebabkan kandungan vitamin C dan ATT buah pamelo menurun, sedangkan PTT dan PTT/ATT meningkat, tetapi pH jus buah menunjukkan kecenderungan berbeda antar aksesi. Selama penyimpanan persentase bobot kulit terus menurun, yang membuat persentase bobot bagian dapat dimakan meningkat. Persentase bobot aksis buah tidak berbiji lebih rendah dibandingkan buah berbiji. Hasil analisis sensori menunjukkan aksesi tidak berbiji ’Jawa 1’ memiliki banyak keunggulan, baik dari hasil uji hedonik (kesukaan) maupun uji mutu skalar, yaitu aroma jeruk yang harum, jus berwarna merah, juici, manis, agak masam, tidak getir, tekstur kantong jus halus dan sedikit graininess. Berdasarkan keungulan-keunggulan tersebut ‘Jawa 1’ dianjurkan untuk dikembangkan lebih lanjut dan dilepas sebagai kultivar pamelo baru.
© Hak Cipta milik IPB, tahun 2012 Hak Cipta dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan
atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan
pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan,
penulisan kritik ataupun tinjauan suatu masalah; dan pengutipan tersebut
tidak merugikan kepentingan yang wajar IPB
Dilarang mengumumkan dan memperbanyak sebagian atau seluruh Karya
BERBIJI DAN TIDAK BERBIJI ASLI INDONESIA
ARIFAH RAHAYU
Disertasi
Sebagai salah satu syarat memperoleh gelar Doktor
pada Program Studi Agronomi dan Hortikultura
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
BOGOR
Penguji Luar Komisi
Ujian Tertutup : Dr. Ir. Endah Retno Palupi, M.Sc.
(Staf pengajar Departemen Agronomi dan Hortikultura, Fakultas pertanian IPB)
Dr. Ir. Darda Efendi, MS
(Staf pengajar Departemen Agronomi dan Hortikultura, Fakultas pertanian IPB)
Ujian Terbuka : Dr. Ir. Agus Purwito, M.Sc.
(Ketua Departemen Agronomi dan Hortikultura, Fakultas pertanian IPB)
Dr. Ir. Winny D. Wibawa
Judul Penelitian : Karakterisasi dan Evaluasi Aksesi Pamelo {Citrus
maxima (Burm.) Merr.} Berbiji dan Tidak
Berbiji Asli Indonesia
Nama : Arifah Rahayu
NRP : A262070051
Program Studi : Agronomi dan Hortikultura
Disetujui
Komisi Pembimbing
Prof. Dr. Ir. Slamet Susanto, M.Sc. Ketua
Prof. Dr. Ir. Bambang S. Purwoko, M.Sc. Dr. Ir. Iswari S. Dewi Anggota Anggota
Diketahui
Ketua Program Studi Dekan Sekolah Pascasarjana IPB
(Dr. Ir. Munif Ghulamahdi, MS) (Dr. Ir. Dahrul Syah, M.Sc.Agr.)
PRAKATA
Puji dan syukur penulis panjatkan ke hadirat Allah SWT atas limpahan rahmat dan karunia-Nya sehingga dapat menyelesaikan disertasi ini. Rangkaian penelitian dalam disertasi berjudul ‘Karakterisasi dan Evaluasi Aksesi Pamelo
{Citrus maxima (Burm.) Merr.} Berbiji dan Tidak Berbiji Asli Indonesia’ ini
dilakukan mulai April 2009 sampai Maret 2012.
Selesainya penelitian ini tidak lepas dari bantuan berbagai pihak, untuk itu penulis mengucapkan terima kasih kepada:
1. Prof. Dr. Slamet Susanto, M.Sc. selaku ketua komisi pembimbing, Prof. Dr. Bambang S. Purwoko dan Dr. Ir. Iswari S. Dewi selaku anggota komisi pembimbing atas bantuan, arahan dan bimbingannya kepada penulis.
2. Dr. Ir. Endah Retno Palupi M.Sc dan Dr. Ir. Darda Efendi, M.S. sebagai penguji ujian tertutup, Dr. Ir. Agus Purwito, MSc. dan Dr. Ir. Winny D. Wibawa sebagai penguji ujian terbuka atas masukan dan sarannya.
3. Pimpinan, staf pengajar dan administrasi di Program Studi AGH Fakultas Pertanian Sekolah Pascasarjana IPB atas pendidikan dan layanan administrasi selama penulis studi di IPB.
4. Teknisi laboratorium di lingkungan IPB (Laboratorium RGCI, Pascapanen, Mikroteknik Departemen AGH IPB, Kebun Percobaan Cikabayan, Laboratorium Pusat Penelitian Sumberdaya Hayati IPB, Laboratorium Pusat Kajian Buah Tropika IPB), Kementerian Pertanian (Laboratorium Balai Besar Pascapanen Cimanggu Bogor), LIPI Biologi (Laboratorium Genetika Tumbuhan) dan Universitas Djuanda (Laboratorium Teknologi Pangan) atas bantuan dan dukungannya.
5. Pimpinan dan Staf Dinas Pertanian Tanaman Pangan dan Hortikultura Kabupaten Sumedang, Pati, Kudus dan Magetan
6. Direktorat Jenderal Pendidikan Tinggi, Kementerian Pendidikan dan Kebudayaan atas beasiswa yang diberikan
8. Kementerian Pertanian RI dan Kementerian RISTEK yang telah membantu dana penelitian melalui program KKP3T dan Insentif Riset Terapan.
9. Pak Sukimin dan keluarga di Magetan dan Pak Parso dan keluarga di Pati yang telah membantu dan menyediakan tanaman pamelo untuk digunakan penelitian.
10.Rekan-rekan mahasiswa S2 dan S3 AGH dan PBT angkatan 2007, terutama Ir. Kartika Ning Tyas MSi. atas bantuan, masukan dan sarannya selama penelitian dan penyelesaian studi.
11.Dr. Ir. Ika Rostika Tambunan MSi.dan Dr. Dorly atas masukan dan sarannya untuk analisis kromosom, Sulassih SP, M.Si. dan Prof. Dr. Ir. Sobir MS atas bantuannya dalam analisis ISSR dan prosedur pelepasan varietas, Prof. Dr. Ir. Sudarsono atas interpretasi data ISSR dan Setyono Ir., M.Si. atas bantuannya dalam pengolahan data.
12.Randi Ginting, SP (AGH angkatan 42), Iwan Sarjono (AGH angkatan 45), Septari (AGT Unida angkatan 2007) yang telah membantu penelitian. 13.Ayahanda Drs. H. Moh. Slamat Anwar dan Ibunda H. Sukanah (Alm.) dan
H.Yetti Mutijah, kedua mertua Drs. Sholeh Sholahuddin dan Ibu H. D. Sumiatin, serta seluruh adik-adik dan ipar atas doa dan dukungannya.
14.Suami tercinta Iwan Ridwan (Alm.) yang telah mendorong dan mendukung penulis untuk melanjutkan studi S3, anak-anak Alfi Rahmawati, Luthfan Syah Ridwan dan Fikri Ghifari Ridwan atas pengertian dan semangatnya.
15.Teman-teman sejawat di Fakultas Agribisnis dan Teknologi Pangan Universitas Djuanda, khususnya Ir. Wini Nahraeni, M.Si., Dr. Elis Dihansih, Ir. Sri Rejeki, M.Sc, dan Jaya Ismail S.Kom. yang telah memberikan dukungannya.
Besar harapan penulis, hasil penelitian ini bermanfaat bagi pengembangan ilmu pengetahuan dan pihak yang memerlukannya.
Bogor, Agustus 2012
Slamat Anwar dan Ibu Hj. Sukanah (Alm.). Penulis telah menikah dengan Ir. Iwan Ridwan MM (Alm.) pada tahun 1987 dan dikaruniai empat orang anak, Miftah Ghifari Ridwan (Alm.), Alfi Rahmawati, Luthfan Syah Ridwan dan Fikri Ghifari Ridwan.
Pendidikan sarjana di tempuh di Program Studi Agronomi Fakultas Pertanian IPB dan lulus pada tahun 1987. Pada tahun 1996 penulis melanjutkan studi di Program Studi Agronomi, Program Pascasarjana IPB dan menamatkannya pada tahun 1999. Pada tahun 2007 penulis berkesempatan meneruskan Program Doktor di Program Studi Agronomi dan Hortikultura, Sekolah Pascasarjana perguruan tinggi yang sama. Selama studi S2 dan S3 penulis mendapat beasiswa dari Kementerian Pendidikan dan Kebudayaan Indonesia.
Sejak tahun 1990 penulis bekerja sebagai staf pengajar Kopertis Wilayah IV dipekerjakan di Program Studi Agroteknologi, Fakultas Agribisnis dan Teknologi Pangan, Universitas Djuanda Bogor.
Sebagian dari disertasi ini telah disampaikan dalam Seminar Nasional Hortikultura tanggal 25-26 Nopember 2010 di Universitas Udayana Denpasar, Bali dan pada tanggal 23-24 Nopember 2011 di Balai Penelitian Tanaman Sayuran, Lembang, Bandung dan menunggu diterbitkan pada Jurnal Agronomi Indonesia edisi April 2012 dan Jurnal Hortikultura Indonesia edisi Desember 2012.
Halaman
DAFTAR TABEL ... xv
DAFTAR GAMBAR ... xvii
DAFTAR LAMPIRAN ... xx
1. PENDAHULUAN ... 1
Latar Belakang ... 1
Tujuan Penelitian ... 4
Hipotesis Penelitian ... 4
Manfaat Penelitian ... 4
2. TINJAUAN PUSTAKA ... 6
Sistematika, Asal-Usul, dan Sifat Botani Pamelo ... 6
Kultivar Pamelo ... 8
Karakterisasi Morfologi ... 8
Karakterisasi Biokimia dengan Isoenzim ... 10
Karakterisasi Molekuler dengan ISSR ... 11
Tingkat Ploidi ... 12
Pembentukan Biji dan Buah pada Pamelo ... 13
Viabilitas Tepung Sari ... 14
Peranan Zat Pengatur Tumbuh dalam Perkembangan Biji dan Buah ... 14 Evaluasi Kualitas Buah ... 15
3. KARAKTERISASI MORFOLOGI, BIOKIMIA, MOLEKULER DAN TINGKAT PLOIDI PAMELO BERBIJI DAN TIDAK BERBIJI ... 17
Karakterisasi Morfologi pada Aksesi Pamelo Berbiji dan Tidak Berbiji ... 20
Bahan dan Alat ... 20
Metode ... 21
Karakterisasi Biokimia dengan Isoenzim pada Aksesi Pamelo Berbiji dan Tidak Berbiji... 21
Bahan dan Alat ... 21
Metode ... 21
Karakterisasi Molekuler dengan ISSR pada Aksesi Pamelo Berbiji dan Tidak Berbiji... 24
Tidak Berbiji ... 27
Analisis Jumlah Kromosom ... 27
Bahan dan Alat ... 27
Karakter Morfologi Aksesi Pamelo Berbiji dan Tidak Berbiji... 28
Karakteri Isoenzim Aksesi Pamelo Berbiji dan Tidak Berbiji ... ... 44
Karakter ISSR Aksesi Pamelo Berbiji dan Tidak Berbiji 48 Karakter Gabungan Penanda Morfologi dan Isoenzim... ... 52 Karakter Gabungan Penanda Morfologi dan ISSR ... 55
Karakter Gabungan Penanda Isoenzim dan ISSR ... 57
Karakter Gabungan Penanda Morfologi, Isoenzim dan ISSR ... 60 Tingkat Ploidi Aksesi Pamelo Berbiji dan Tidak Berbiji 61 SIMPULAN ... 64
4. PEMBENTUKAN BUAH BERBIJI DAN TIDAK BERBIJI PADA PAMELO ... 65
Uji Viabilitas Tepung Sari Aksesi Pamelo Berbiji dan Tidak Berbiji ... 68
Uji Penyerbukan Aksesi Pamelo Berbiji dan Tidak Berbiji ... 69
HASIL DAN PEMBAHASAN ... 71
Viabilitas Tepung Sari Aksesi Pamelo Berbiji dan Tidak Berbiji... ... 71
Evaluasi Kemampuan Pembentukan Buah Tidak Berbiji Melalui Uji Penyerbukan ... 72
SIMPULAN ... 80
5. EVALUASI KUALITAS BUAH PAMELO BERBIJI DAN TIDAK BERBIJI SELAMA MASA SIMPAN... 82
ABSTRAK ... 82
ABSTRACT ... 82
Metode ... 85
Peubah yang Diamati ... 86
HASIL DAN PEMBAHASAN ... 87
Karakter Fisik yang Mempengaruhi Kualitas Buah Pamelo Berbiji dan Tidak Berbiji ... 88
Karakter Kimia yang Mempengaruhi Kualitas Buah Pamelo Berbiji dan Tidak Berbiji ... 92
Analisis Sensori pada Buah Pamelo Berbiji dan Tidak Berbiji ... 99
SIMPULAN ... 104
6. PEMBAHASAN UMUM ... 105
Keragaman Pola Pengelompokan Aksesi Pamelo Berdasarkan Penanda Morfologi, Isoenzim, ISSR dan Kombinasinya ... 105
Hubungan antara Karakterisasi Morfologi, Isoenzim dan ISSR, Analisis Ploidi dan Pembentukan Buah Pamelo Berbiji dan Tidak Berbiji ... 108
Hubungan antara Karakter Morfologi dengan Karakter Kimia dan Fisik Buah Pamelo Selama Penyimpanan ... 110
Upaya Pengembangan Aksesi Pamelo yang Potensial Dijadikan Varietas Unggul Hortikultura ... 111
7. SIMPULAN DAN SARAN ... ... 114
Simpulan ... 114
Saran ... 115
DAFTAR PUSTAKA ... 116
Halaman
1. Nama dan susunan basa primer koleksi PKBT IPB ……….. 26
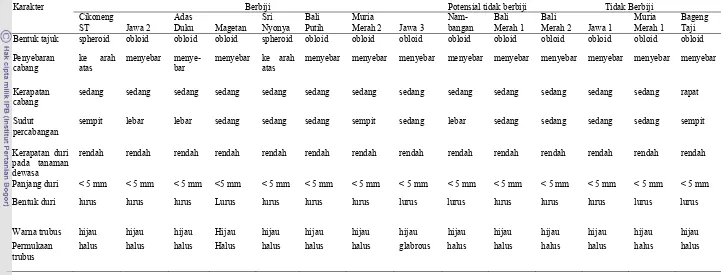
2. Deskripsi pohon 14 aksesi pamelo ………... 31
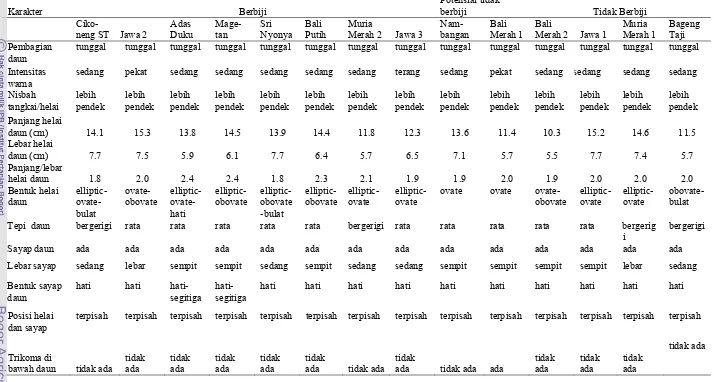
3. Deskripsi daun 14 aksesi pamelo ……….. 33
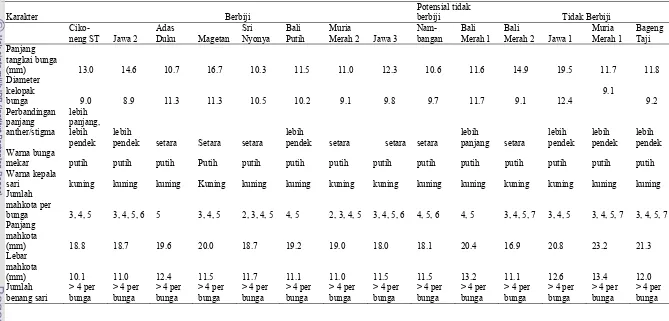
4. Deskripsi bunga 14 aksesi pamelo ………... 34
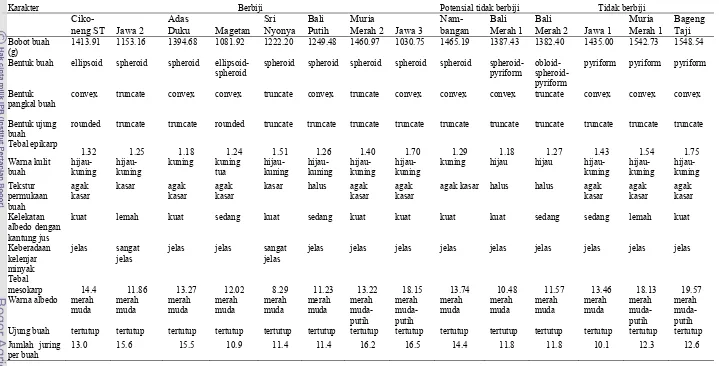
5. Deskripsi buah 14 aksesi pamelo ……….. 37
6. Nilai ciri dan dua nilai komponen utama (KU) pertama berdasarkan penanda morfologi (pohon, daun, bunga dan buah) 42 7. Tingkat kemiripan antar/di dalam kelompok aksesi pamelo berdasarkan penanda morfologi ………... 43
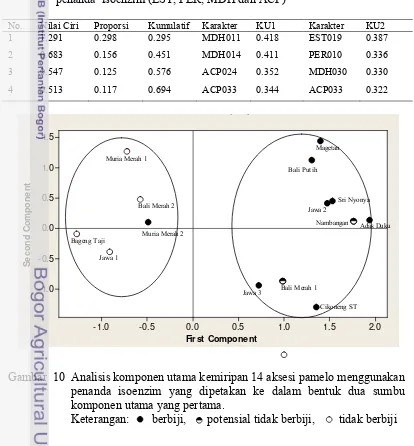
8. Nilai ciri dan dua nilai komponen utama (KU) pertama berdasarkan penanda isoenzim (EST, PER, MDH dan ACP) ... 46
9. Tingkat kemiripan antar/di dalam kelompok aksesi pamelo berdasarkan penanda isoenzim ………... 47
10. Nilai ciri dan dua nilai komponen utama (KU) pertama berdasarkan penanda ISSR ... 50
11. Tingkat kemiripan antar/di dalam kelompok aksesi pamelo berdasarkan penanda ISSR ………... 52
12. Nilai ciri dan dua nilai komponen utama (KU) pertama berdasarkan penanda morfologi dan isoenzim (EST, PER, MDH dan ACP) ………..………..………..………. 53
13. Kisaran tingkat kemiripan antar/di dalam kelompok aksesi pamelo berdasarkan penanda morfologi dan isoenzim ... 54
14. Nilai ciri dan dua nilai komponen utama (KU) pertama berdasarkan penanda morfologi dan ISSR ………... 55
15. Kisaran tingkat kemiripan antar/di dalam kelompok aksesi pamelo berdasarkan penanda morfologi dan ISSR ... 56
18. Kisaran tingkat kemiripan antar/di dalam kelompok aksesi
pamelo berdasarkan penanda morfologi, isoenzim dan ISSR ... 61
19. Hasil konfirmasi jumlah kromosom hasil analisis dengan metode Sartrosumarjo (2006) dan flow cytometry ... 63
20. Hasil uji viabilitas tepung sari pada aksesi berbiji, potensial tidak berbiji dan tidak berbiji ………..………..……… 72
21. Hasil penyerbukan pada berbagai aksesi pamelo ... 79
22 Rekapitulasi hasil penyerbukan pada berbagai aksesi pamelo … 79 23 Rekapitulasi nilai analisis ragam pengaruh kelompok aksesi, aksesi, masa simpan dan interaksi antara kelompok aksesi dan masa simpan dan aksesi dan masa simpan ... 87
24. Persentase bobot kulit buah 13 aksesi pamelo pada 0-8 MSP .... 88
25. Persentase bobot bagian buah dapat dimakan 13 aksesi pamelo pada 0-8 MSP ... 89
26. Persentase bobot sekat buah 13 aksesi pamelo pada 0-8 MSP ... 90
27. Persentase bobot aksis buah 13 aksesi pamelo pada 0-8 MSP .... 91
28. Kandungan vitamin C 13 aksesi pamelo pada 0-8 MSP ... 93
29. Kemasaman buah 13 aksesi pamelo pada 0-8 MSP ... 95
30. Kandungan PTT 13 aksesi pamelo pada 0-8 MSP ... 96
31. Kandungan ATT 13 aksesi pamelo pada 0-8 MSP ... 97
32. Nisbah PTT/ATT 13 aksesi pamelo pada 0-8 MSP ... 98
33. Kandungan naringin 13 aksesi pamelo ... 99
34. Hasil uji kesukaan (hedonik) pada 13 aksesi pamelo ... 100
35. Hasil uji mutu skalar pada aksesi pamelo ... 101
36. Nilai koefisien kemiripan tertinggi dan terendah dan nilai korelasi koefenetik MxComp (r) dengan penanda morfologi, isoenzim, ISSR dan kombinasinya ...
Halaman
1. Bagan alir penelitian Evaluasi Karakter Aksesi Pamelo {Citrus
maxima (Burm.) Merr.} Berbiji dan Tidak Berbiji Asli
Indonesia ...……... 5
2. Bentuk pohon aksesi pamelo (a) spheroid dan (b) obloid ...
29 3. Bentuk daun 14 aksesi pamelo berbiji: (a) Cikoneng ST , (b)
Jawa 2 , (c) Adas Duku , (d) Magetan , (e) Sri Nyonya , (f) Bali Putih, (g) Muria Merah 2, (h) Jawa 3, potensial tidak berbiji (i) Nambangan , (j) Bali Merah 1 dan tidak berbiji: (k) Bali Merah 2, (l) Jawa 1, (m) Muria Merah 1, (n ) Bageng
Taji ...……... 30
4. Bunga berbagai aksesi/kultivar pamelo berbiji: (a) Cikoneng ST , (b) Jawa 2 , (c) Adas Duku , (d) Magetan , (e) Bali Putih , (f) Muria Merah 2, (g) Jawa 3 , potensial tidak berbiji (h) Nambangan , (i) Bali Merah 1 dan tidak berbiji: (j) Jawa 1, (K) Muria Merah 1, (l ) Bageng Taji dan bagian-bagian
bunga pamelo ...……... 32
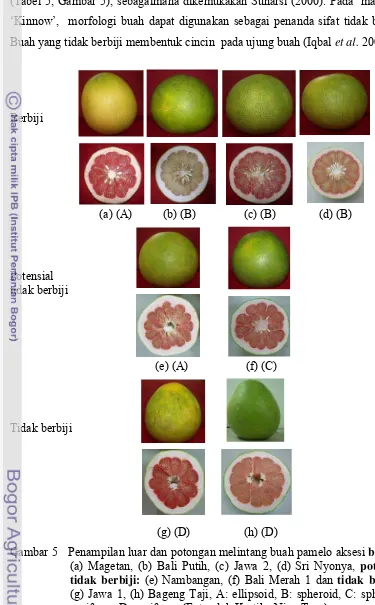
5. Penampilan luar dan potongan melintang buah pamelo aksesi
berbiji: (a) Cikoneng ST, (b) Magetan, (c) Jawa 2, (d) Sri Nyonya, potensial tidak berbiji: (e) Nambangan, (f) Bali Merah 1 dan tanpa biji, (g) Jawa 1, (h) Bageng Taji, A: ellipsoid, B: spheroid, C: spheroid-pyriform, D:
pyriform... 35
6. Perbedaan bentuk buah ‘Bali Merah 2’ dalam pohon yang
sama, kiri: obloid, kanan: spheroid-pyriform ... 40
7. Analisis komponen utama kemiripan 14 aksesi pamelo menggunakan penanda morfologi yang dipetakan ke dalam bentuk dua sumbu komponen utama yang pertama. ...
42
8. Dendogram 14 aksesi pamelo berdasarkan penanda morfologi 44
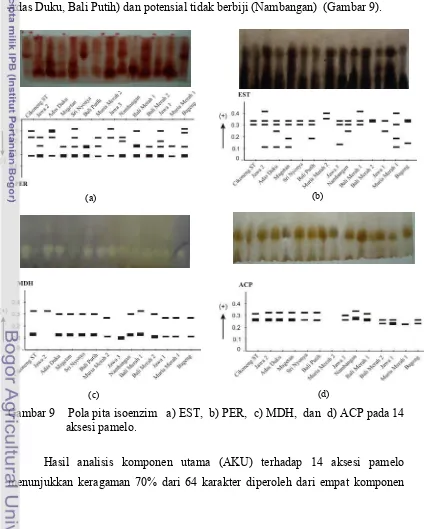
9. Pola pita isoenzim a). EST, b). PER, c). MDH, dan d).
ACP pada 14 aksesi pamelo. ... 45
10. Analisis komponen utama kemiripan 14 aksesi pamelo menggunakan penanda isoenzim yang dipetakan ke dalam
PKBT 3, (c) PKBT 4, (d) PKBT 7, (e) PKBT 8, (f) PKBT 9, (g) PKBT 11, (h) PKBT 12, (i) RED 12 dan (j) RED 17. ...
49
13. Analisis komponen utama kemiripan 14 aksesi pamelo menggunakan penanda ISSR yang dipetakan ke dalam bentuk
dua sumbu komponen utama yang pertama. ... 51
14. Dendogram 14 aksesi pamelo berdasarkan penanda ISSR ...
52 15. Analisis komponen utama kemiripan 14 aksesi pamelo
menggunakan penanda morfologi dan isoenzim yang dipetakan ke dalam bentuk dua sumbu komponen utama yang
pertama. ... 53
16. Dendogram 14 aksesi pamelo berdasarkan penanda morfologi
dan isoenzim. ... 55
17. Analisis komponen utama kemiripan 14 aksesi pamelo menggunakan penanda morfologi dan ISSR yang dipetakan ke dalam bentuk dua sumbu komponen utama yang pertama. ...
56
18. Dendogram 14 aksesi pamelo berdasarkan penanda morfologi
dan ISSR ... 57
19. Analisis komponen utama kemiripan 14 aksesi pamelo menggunakan penanda isoenzim dan ISSR yang dipetakan ke dalam bentuk dua sumbu komponen utama yang pertama. ...
59
20. Dendogram 14 aksesi pamelo berdasarkan penanda isoenzim
dan ISSR ... 60
21. Dendogram 14 aksesi pamelo berdasarkan penanda morfologi,
isoenzim dan ISSR ... 61
22. Kromosom (a) Bageng, (b) Muria Merah 1, (c) Muria Merah
2, (d) Sri Nyonya, (e) Adas Duku, (f) Nambangan ... 62
23. Hasil konfirmasi ploidi antar (a) ‘Nambangan-‘Sri Nyonya’, (b) ‘Nambangan’- ‘Magetan’, dan (c) ‘Sri Nyonya’-‘Muria
Merah 2. ... 63
24. Perbedaan warna tepung sari yang (a) viable (merah) dan
nonviable (kuning), dan (b) tepung sari yang berkecambah ... 72
26 Potongan melintang buah pamelohasil penyerbukan sendiri secara alami (a) ‘Jawa 3’, (b) ‘Nambangan’, (c) ‘Bali Merah 1’,(d) ‘Bali Merah 2’, (e) ‘Bageng Taji’, hanya ‘Jawa 3’ yang
buahnya sebagian berbiji ... 75
27 Potongan melintang buah pamelo hasil penyerbukan sendiri secara buatan (a) Bali Merah 2, (b) Bageng, (b) Nambangan,
semuanya mampu menghasilkan biji ... 76
28 Potongan melintang buah pamelo hasil penyerbukan terbuka, (a) Bali Merah 2 x Nambangan, (b) Bageng x Cikuning, (c) Bali Merah 1 x Jawa 3, semuanya berbiji banyak kecuali Bali
Merah 2 ... 76
29. Potongan melintang buah pamelo hasil penyerbukan silang alami (a) Bali Merah 2, berbiji sedikit, (b) Bageng tidak
berbiji, (c) Nambangan berbiji ... 76
30. Diagram laba-laba uji mutu skalar aksesi pamelo
berbiji... 102
31. Diagram laba-laba uji mutu skalar aksesi pamelo potensial tidak berbiji... ...
103
Halaman
1. Descriptor list pamelo ... 132
2. Komposisi larutan penyangga gel pH 6.0 (Horry 1989) ... 142
3. Komposisi larutan pengekstrak untuk volume 40 ml (Horry 1989) ... 142
4. Komposisi larutan penyangga elektroda pH 6.0 (Horry 1989) .... 142
5. Komposisi larutan penyangga fosfat pH 7.0 ... 142
6. Komposisi larutan AAT ... 142
7. Komposisi larutan CTAB 10%... 142
8. Komposisi larutan kloroform:isoamilalkohol (24:1)... 142
9. Komposisi buffer Tris-HCl 1 M pH 8.0 ... 143
10. Komposisi buffer TAE 50 x... ... 143
11. Komposisi loading dye... 143
12. Komposisi larutan etanol 70% ... 143
13. Komposisi larutan NaCl ... 143
14. Komposisi buffer ekstraksi untuk Isolasi DNA ... 143
15. Komposisi buffer TE (Tris-HCl EDTA) ... 143
16. Komposisi larutan EDTA 0.5 M pH 8.0 ... 144
17. Komposisi larutan natrium asetat 3 M pH 5.2 ... 144
18. Kuisioner uji hedonik rasa, aroma dan penerimaan pamelo umum buah ... 144
Latar Belakang
Pamelo {Citrus maxima (Burm.) Merr.} merupakan salah satu di antara
tiga spesies Citrus yang asli, disamping sitrun (C. medica L.) dan mandarin (C.
reticulata Blanco). Spesies Citrus lainnya merupakan hasil persilangan ketiga
kelompok tersebut (Campos et al. 2005). Pamelo potensial dikembangkan di
Indonesia, karena karakteristiknya yang khas, yaitu berukuran besar, memiliki
rasa segar, dan daya simpan yang lama sampai 4 bulan (Susanto 2004). Di
samping itu kriteria buah pamelo yang potensial dikembangkan adalah memiliki
warna jus merah, rasa getir rendah, mudah dikupas dan tidak berbiji. Bila
dibungkus kertas dalam kotak yang berventilasi baik, buah pamelo dapat tetap
baik kondisinya selama 6-8 bulan (Orwa et al. 2009).
Di antara faktor yang turut menentukan keberhasilan pengembangan
pamelo adalah terdapatnya kultivar unggul yang dirakit dan/atau diseleksi dari
berbagai aksesi plasma nutfah yang tersedia. Plasma nutfah pamelo banyak
ditemukan di berbagai daerah di Indonesia, dengan nama daerah yang berbeda,
yang dikenal sebagai kultivar lokal. Meskipun demikian baru sebagian kecil
kultivar yang dimanfaatkan secara komersial, antara lain ‘Magetan’,
‘Nambangan’, ‘Raja’, ‘Ratu’ dan ‘Sri Nyonya’ (Pusat Penelitian dan
Pengembangan Hortikultura 2006).
Berbagai plasma nutfah tersebut merupakan sumber keragaman genetik
dalam spesies pamelo yang penting sebagai bahan pemuliaan. Keragaman genetik
ini dapat diukur pada berbagai tingkatan fungsional, mulai dari fenotipe hingga
cetak biru (blueprint), yaitu dari keragaman morfologi, keragaman kromosom
(inversi, translokasi dan poliploidi), lokus enzim dan penanda DNA (Mallet
1996). Plasma nutfah pamelo dapat diperoleh antara lain melalui eksplorasi ke
sentra produksi, dan selanjutnya dilakukan karakterisasi dan evaluasi.
Karakterisasi merupakan kegiatan dalam rangka mengidentifikasi
sifat-sifat penting yang bernilai ekonomi, atau yang merupakan penciri suatu
aksesi/kultivar/varietas yang bersangkutan. Evaluasi adalah kegiatan yang
reaksi varietas tanaman terhadap cekaman faktor biotik dan abiotik (Somantri et
al. 2008). Kegiatan karakterisasi dan evaluasi memiliki arti dan peran penting
yang akan menentukan nilai guna varietas tanaman yang bersangkutan.
Hingga kini karakterisasi dan evaluasi pamelo di Indonesia belum tuntas
dilakukan, meskipun pada beberapa aksesi pamelo asal Magetan telah dilakukan
karakterisasi secara morfologi (Purwanto et al. 2002a), dengan isoenzim
(Purwanto et al. 2002b; Purwanto et al. 2002c), secara morfologi dan isoenzim
(Rakhman 2008). Analisis biokimia dengan isoenzim pada tanaman jeruk, antara
lain digunakan untuk membedakan kultivar jeruk asam (Rahman et al. 2001),
mendapatkan informasi genetik pada jeruk liar (Hirai et al. 1990), dan
mengidentifikasi bibit zigotik dan nuselar pada jeruk hasil persilangan antar ploidi
yang berbeda (Tusa et al. 2002).
Karakterisasi morfologi dan biokimia akan lebih akurat bila disertai
dengan analisis molekuler. Pada aksesi pamelo dari berbagai daerah di Indonesia
telah dilakukan analisis RAPD (Agisimanto dan Supriyanto 2007), sedangkan di
Turki pembedaan antara aksesi grapefruit dan pamelo digunakan ISSR (Uzun et
al. 2010). Baik teknik RAPD (Randomly Amplified Polymorphic DNA) maupun
ISSR (Inter-Simple Sequence Repeat) sama-sama sederhana dan tidak
memerlukan pengetahuan tentang sekuens genom yang akan diuji (Pharmawati et
al. 2004). Kelebihan ISSR adalah mempunyai daya ulang (reproduceability) yang
lebih tinggi, karena menggunakan primer yang lebih panjang (16-25 mer)
dibandingkan dengan primer RAPD (10 mer) (Reddy et al. 2002).
Evaluasi kualitas selama penyimpanan yang meliputi kandungan padatan
terlarut total (PTT), asam tertitrasi total (ATT), vitamin C, dan rasa pahit
(bitterness) aksesi-aksesi pamelo belum banyak dilakukan. Rasa pahit pada
pamelo terutama disebabkan oleh flavonoid (naringin) dan limonoid (limonin)
(Pichaiyongvongdee dan Haurenkit 2009a). Evaluasi mengenai kandungan
senyawa ini menarik untuk dilakukan, karena Tisserat dan Berhow (1998)
menyatakan bahwa terdapat hubungan antara tingkat ploidi dengan kandungan
flavonoid pada grapefruit (Citrus paradisi Macf.). Kandungan naringin pada
grapefruit tetraploid lebih tinggi dibanding pada grapefruit diploid. Hal demikian
Berdasarkan jumlah biji per buah dapat diamati adanya aksesi pamelo
yang buahnya berbiji, tidak berbiji, atau potensial tidak berbiji. Buah tidak berbiji
merupakan sifat yang banyak diminati untuk pasar pamelo segar. Hal ini
berkaitan dengan kemudahan penggunaannya. Selain mudah dikonsumsi,
proporsi bagian dapat dimakan dari buah jeruk tidak berbiji lebih besar dibanding
buah berbiji (Yamashita 1976).
Suatu tanaman dianggap menghasilkan buah tidak berbiji jika mampu
menghasilkan buah tidak berbiji sama sekali, biji mengalami aborsi atau memiliki
sejumlah biji yang tereduksi (Vardi et al. 2008). Pada jeruk mandarin, buah
disebut tidak berbiji jika jumlah biji per buah kurang dari lima (Varoquaux et al.
2000), sedangkan pada grapefruit 0-6 biji (Chacoff dan Aizen 2007).
Buah tidak berbiji dapat dihasilkan dengan cara pemuliaan, teknik
budidaya dan secara alami. Dengan cara pemuliaan, buah tidak berbiji didapat
dengan mengembangkan aksesi tidak berbiji melalui produksi bibit hibrida
triploid (Yamashita 1976), kultur endosperma (Raza et al. 2003) dan melalui
iradiasi untuk mendapatkan mutan (Sutarto et al. 2009). Secara kultur teknis,
produksi buah tidak berbiji dapat dilakukan dengan menggunakan zat pengatur
tumbuh, antara lain asam giberelin (GA) (Ben-Cheikh et al. 1997). Pengembangan
aksesi tidak berbiji dapat pula dilakukan dengan memanfaatkan aksesi tidak
berbiji sebagai tetua. Menurut Yamamoto et al. (1995), terdapat hubungan yang
erat antara kemampuan berbiji tetua dengan turunan hibridanya. Tetua yang tidak
berbiji atau berbiji sedikit umumnya menghasilkan keturunan tidak berbiji,
sementara tetua yang berbiji atau berbiji banyak hanya menghasilkan sedikit
keturunan tidak berbiji.
Kajian mekanisme pembentukan buah tidak berbiji telah dilakukan oleh
Yamamoto et al. (1995), yang mempelajari hubungan antara sterilitas jantan dan
betina, dan ketidakserasian-sendiri (self-incompatibility) terhadap jumlah biji pada
berbagai aksesi jeruk, termasuk pamelo. Sementara itu Yamamoto dan Tominaga
(2002) mempelajari hal ini pada Citrus keraji, dan mendapatkan bahwa buah tidak
berbiji diakibatkan oleh sterilitas betina, self-incompatibility dan partenokarpi.
Pada aksesi pamelo tidak berbiji di Indonesia mekanisme ini belum diketahui.
kromosomnya. Tanaman yang kromosomnya triploid (3n) akan menghasilkan
buah tidak berbiji, sedangkan yang diploid (2n) berbiji. Serangkaian penelitian
dilakukan dengan skema seperti tercantum pada Gambar 1.
Tujuan
Secara umum penelitian ini bertujuan untuk mengkarakterisasi dan
mengevaluasi aksesi pamelo yang dapat digunakan sebagai karakter dalam
pengembangan pamelo tidak berbiji. Secara khusus penelitian ini bertujuan
untuk:
1. Mengidentifikasi karakter morfologi, biokimia, molekuler dan tingkat
ploidi tanaman pamelo berbiji dan tidak berbiji.
2. Mengidentifikasi pembentukan buah pamelo berbiji dan tidak berbiji.
3. Mengevaluasi karakter kualitas buah pamelo berbiji dan tidak berbiji
selama masa simpan.
Hipotesis
Hipotesis yang diajukan dalam penelitian ini adalah:
1. Terdapat perbedaan karakter morfologi, biokimia, molekuler dan tingkat
ploidi antar aksesi pamelo berbiji dan tidak berbiji.
2. Terdapat perbedaan karakter yang berhubungan dengan pembentukan buah
pamelo berbiji dan tidak berbiji
3. Terdapat perbedaan karakter kualitas buah pamelo berbiji dan tidak berbiji
selama masa simpan.
Manfaat Penelitian
Hasil penelitian ini diharapkan dapat mengungkapkan berbagai informasi
penting yang bermanfaat dalam pengembangan kultivar pamelo tidak berbiji,
melalui hasil karakterisasi morfologi, biokimia, molekuler dan tingkat ploidi,
pembentukan buah pamelo berbiji dan tidak berbiji serta evaluasi kualitas buah
Gambar 1 Bagan alir penelitian Karakterisasi dan Evaluasi Aksesi Pamelo {Citrus maxima (Burm.) Merr.} Berbiji dan Tidak Berbiji Asli Indonesia.
Kajian Pembentukan Buah Berbiji dan Tidak Berbiji pada Pamelo
Evaluasi Kualitas Buah: - Nutrisi selama masa
simpan
- Analisis Sensori Karakterisasi:
- Morfologi - Isoenzim - Molekuler - Tingkat ploidi
Kelompok Aksesi: - Berbiji
- Potensial tidak berbiji - Tidak berbiji
Karakter-karakter tanaman yang dapat membedakan aksesi pamelo berbiji dan tidak berbiji, yang dapat digunakan sebagai karakter
dalam pengembangan pamelo tidak berbiji Eksplorasi Berbagai
2. TINJAUAN PUSTAKA
Sistematika, Asal-Usul, dan Sifat Botani Pamelo
Pamelo (Citrus maxima (Burm.) Merr.) memiliki sinonim C. grandis (L.)
Osbeck, C. decumana L., C. aurantium var. grandis L. dan C. aurantium var.
decumana L. (Manner et al. 2006). Tanaman ini termasuk genus Citrus, sub suku
(sub-tribe) Citrinae, tribe Citriae, sub famili Aurantioideae, famili Rutaceae
(Direktorat Tanaman Buah 2003). Citrinae dicirikan dengan buah hesperidium
dengan adanya kulit keras dan kaku yang mengelilingi bagian dalam buah dan
kantong jus (Hamilton et al. 2008). Adanya kulit yang keras dan kaku ini
melindungi buah dari kerusakan selama penanganan pascapanen dan pengeringan
selama penyimpanan (Albrigo dan Carter 1977).
Pamelo berasal dari Malesia, kemudian menyebar ke Indo-Cina, Cina
Selatan, Jepang Selatan, India Barat, Mediterania dan Amerika Tropik
(Niyomdham 1992). Walaupun tempat asal pamelo yang tepat tidak diketahui,
tetapi kemungkinan besar dari Malaysia, Thailand dan Indonesia, karena di daerah
ini banyak dijumpai kerabat liarnya (Thulaja 2003). Kini pamelo telah diproduksi
secara komersial di 74 negara baik untuk memenuhi kebutuhan lokal maupun
ekspor (Talon dan Gmitter Jr. 2008).
Pusat produksi pamelo dunia terdapat di Cina bagian Selatan, Thailand,
Vietnam, Malaysia, Indonesia, Taiwan dan Jepang (Hodgson 1967). Di
Indonesia, sentra produksi pamelo utama terdapat di Kabupaten Magetan,
sedangkan sentra produksi potensial antara lain di Kabupaten Sumedang, Pati,
Kudus, Pangkajene dan Kepulauan (Pangkep) dan Bireun (Aceh).
Tinggi tanaman pamelo sekitar 5-15 m dengan batang yang besar (10-30
cm), percabangan rendah dan tidak beraturan (Morton 1987). Cabang muda
bersudut, seringkali berambut halus, pendek dan biasanya berduri (Orwa et al.
2009). Tajuk pohon berbentuk bulat (Christman 2008).
Daun pamelo berbentuk bulat telur sampai jorong dengan ukuran 5-20 cm
x 2-12 cm. Pangkal daun membundar sampai menjantung, tepi rata sampai
beringgit dangkal, dan ujung daun lancip sampai tumpul. Pada permukaan daun
mencapai 7 mm, berwarna hijau kuning, helai daun bagian bawah berbulu,
berwarna agak suram (Niyomdham 1992).
Bunga pamelo berbau harum, tunggal atau terdiri atas 2-10 kuntum bunga
di ketiak daun atau kadang-kadang berjumlah 10-15 kuntum di ujung cabang.
Mahkota bunga pamelo berjumlah 4-5 helai, berwarna putih-kekuningan, panjang
1.5-3.5 cm, berbulu halus pada bagian luar. Benang sari berwarna putih,
sedangkan serbuk sari jingga (Morton 1987). Kotak sari bunga pamelo terletak
berhadapan dengan permukaan kepala putik dan dapat melepaskan serbuk sarinya
sebelum kuncup bunga mekar. Meskipun demikian, sebagian besar aksesi pamelo
bersifat self-incompatible (Niyomdham 1992). Sifat self-incompatible pada
pamelo memberi peluang untuk dihasilkannya buah tidak berbiji (Yamamoto dan
Tominaga 2002). Bunga jeruk mekar pada pagi hingga sore dan mencapai
puncaknya pada tengah hari. Kepala putik sudah reseptif sebelum bunga mekar
(Ashari 2004).
Bunga jeruk bersifat pentamerous (berbilangan lima) dan hermaprodit.
Tangkai bunga memiliki dua daerah absisi, yaitu pada axil dan yang lain dekat
dengan kelopak bunga (Ortiz 2002). Bunga jeruk dibedakan atas bunga tanpa
daun (leafless bloom) dan bunga berdaun (leafy bloom). Bunga berdaun cenderung
mampu membentuk buah (fruit set) lebih tinggi dibanding bunga tanpa daun.
Pamelo biasanya menghasilkan banyak bunga pada musim utama, yang
jumlahnya bergantung pada aksesi, umur pohon dan kondisi lingkungan.
Walaupun demikian persentase fruit set relatif rendah, berkisar 0.8-1.1% pada
bunga tidak berdaun dan 4.8 – 6.0% pada bunga berdaun (Nakajima et al. 1993).
Buah pamelo berukuran besar, dengan diameter rata-rata 15-22 cm,
bahkan ada yang lebih dari 30 cm, dengan warna kulit kuning. Daging buah
berwarna putih, kekuningan atau merah muda. Bobot buah rata-rata sekitar 1-2
kg, kadang-kadang dapat mencapai 9 kg (Christman 2008). Biji pamelo tidak
banyak, berukuran besar dengan permukaan keriput, warnanya kekuningan dan
Kultivar Pamelo
Di Indonesia terdapat banyak kultivar pamelo, antara lain Nambangan,
Besar Nambangan, Magetan, Srinyonya, Cikoneng ST, Pangkajene Merah, Buton,
Bulat Hijau, Lonjong Hijau, Duku, Papermus Obesi I, Papermus Obesi II, Besar
Merah, Besar Putih, Putih tanpa biji (Agismanto dan Supriyanto 2007). Selain
itu dikenal pula pamelo Raja, Ratu dan Pangkep (Direktorat Tanaman Buah
2007), dan berbagai aksesi yang belum dilepas sebagai kultivar.
Di negara lain dikenal kultivar Banpeiyu (asal Malaya, diintroduksi ke
Taiwan dan Jepang), Chandler (berkembang di India dan California), Hirado
(Jepang), Dang Ai Chaa, Hoem Bai Toey, Kao Lang Sat, Kao Pan, Kao Phuang,
Kao Ruan Tia, Kao Yai, Khun Nok, Thong Dee (Thailand), Siamese Sweet
(diintroduksi oleh USDA dan ditanam di California), Tahitian (diduga berasal dari
Kalimantan dibawa ke Tahiti kemudian Hawaii) dan Tresca (dari Bahama,
ditanam secara komersial di California) (Morton 1987).
Karakterisasi Morfologi
Menurut Idris dan Saad (2001), karakterisasi plasma nutfah memiliki
beberapa manfaat. Data karakterisasi memiliki nilai diagnostik, yaitu sebagai alat
untuk mengidentifikasi bahan yang dikoleksi atau menguji keaslian plasma
nutfah, untuk membedakan homonim atau nama yang hampir sama dan
mengidentifikasi atau menyeleksi spesies, klon, kultivar atau varietas. Di samping
itu hasil karakterisasi juga bermanfaat untuk mengklasifikasikan spesies, klon,
kultivar atau varietas dan mendeteksi karakteristik yang berkaitan yang
kemungkinan dapat memiliki nilai praktis.
Tahap awal identifikasi tanaman, biasanya dilakukan secara morfologi.
Sifat morfologi ini dapat berupa sifat kualitatif maupun kuantitatif, yang memiliki
tipe dan aksi gen yang berbeda (Fitmawati 2008). Sifat kualitatif antara lain
bentuk tajuk, daun, dan buah; warna daun, bunga, kulit buah, dan daging buah,
sedangkan sifat kuantitatif, diantaranya panjang dan lebar daun, panjang mahkota
dan kelopak bunga serta bobot bagian-bagian buah. Penanda morfologi masih
dan buah aksesi pamelo terbaik (Rahman et al. 2003), dan mendeteksi poliploidi
pada jeruk (Bilquess 2004).
Malik et al. (2006) mendapatkan adanya keragaman ukuran daun, buah
dan biji pada koleksi aksesi C. indica Tanaka dan C. macroptera Montr. yang
dikaraktersisasi secara morfologi. Hasil penelitian Hardiyanto et al. (2007)
menunjukkan karakter morfologi dapat membedakan jeruk varietas lokal (keprok
Cinakonde, manis Punten dan besar Nambangan) dari kelompok lain dalam
populasi spesies yang sama, sehingga varietas lokal ini dapat dipisahkan pada
tingkatan takson di bawah spesies. Lebih jauh Hamilton et al. (2008) menyatakan
bahwa morfologi biji, khususnya topografi permukaan biji bermanfaat sebagai alat
identifikasi taksonomi pada jeruk liar Australia.
Meskipun karakterisasi secara morfologi kurang akurat, karena
dipengaruhi oleh stadia pertumbuhan tanaman dan faktor lingkungan, tetapi tetap
diperlukan sebagai tahap awal untuk mengetahui keragaman genetik tanaman dan
untuk melengkapi data hasil analisis biokimia dan molekuler. Karakterisasi
morfologi juga penting dilakukan, karena keragaman morfologi pada jeruk,
terutama mandarin tidak bergantung pada keragaman genetiknya, yang tampak
dari korelasi kofenetik yang rendah antara dendrogram morfologi dan molekuler
(Koehler-Santos et al. 2003; Campos et al. 2005). Beberapa karakter hortikultura
penting juga dikendalikan oleh banyak gen (Campos et al. 2005), dan
heritabilitasnya rendah. Dalam melakukan karakterisasi perlu diperhatikan
karakter-karakter yang dapat diturunkan, dapat mudah diamati dengan mata
telanjang, dan diekspresikan pada semua kondisi atau lingkungan (Perry dan
Battencourt 1997). Biasanya karakter ini bersifat kualitatif dan stabil pada
berbagai kondisi lingkungan, contohnya warna bunga dan bentuk buah.
Menurut Suharsi (2000) perbedaan antar aksesi pamelo dapat diidentifikasi
berdasarkan ciri-ciri buahnya, antara lain ukuran dan bentuk buah, bentuk ujung
dan pangkal buah, warna dan tekstur flavedo (epicarp), ketebalan dan warna
albedo (mesocarp), warna endokarpium, warna dan rasa vesicula atau daging
buah, aroma minyak atsiri, jumlah buah pada setiap pohon dan jumlah biji pada
tidak berbiji (hasil penyerbukan sendiri) mempunyai bobot buah lebih ringan
dibandingkan buah berbiji (hasil penyerbukan terbuka) (Yahata et al. 2005).
Kelemahan penanda morfologi adalah dipengaruhi oleh tahap
perkembangan tanaman dan lingkungan. Kadang-kadang sulit membedakan
genotipe yang diamati, karena secara morfologi tampak sama, walaupun
sebenarnya genotipe tersebut berbeda. Hal ini terjadi akibat sifat resesif tertutup
oleh sifat dominan (Bakhtiar 2002).
Karakterisasi Biokimia dengan Isoenzim
Isoenzim merupakan enzim yang terdiri atas molekul-molekul yang
mempunyai struktur kimia yang berbeda akan tetapi mengkatalisis reaksi kimia
yang sama. Sejumlah isoenzim diketahui berasosiasi atau terkait dengan karakter
agronomi, namun jumlah isoenzim yang terbatas membuat penggunaan isoenzim
menjadi terbatas pula.
Dibandingkan dengan ciri-ciri morfologi, isoenzim dan sifat-sifat biokimia
lain dapat lebih menunjukkan genotipe suatu organisme. Pita-pita isoenzim dapat
dievaluasi dari segi genetik (yaitu frekuensi alela pada setiap lokus) dan dari segi
fenotipe (yaitu dengan menganggap setiap pita tunggal sebagai suatu karakter
kodominan, yang keberadaannya dinyatakan dengan skor) (Gonzalez-Andrez et
al. 1996).
Fungsi utama isoenzim adalah mengendalikan aktivitas metabolik suatu
organisme. Isoenzim-isoenzim dikode oleh gen-gen yang berbeda, dan setiap gen
dapat memiliki alela berbeda pada lokus yang sama. Perbedaan ukuran,
konfigurasi dan muatan ion di antara isoenzim membuatnya dapat dideteksi dan
ditampilkan dengan berbagai cara pemisahan melalui elektroforesis.
Kebanyakan isoenzim menunjukkan lokus kodominan yang terdistribusi
mengikuti hukum Mendel dan banyak lokus diekspresikan pada seluruh stadia
siklus hidup tanaman (Hamrick 1989). Walaupun demikian, isoenzim juga dapat
menghasilkan pola pita yang kompleks terutama ketika enzim multimerik terlibat,
yang membuat interpretasi menjadi sulit. Sistem isoenzim juga bergantung pada
pewarnaan histokimia, sehingga memerlukan jumlah enzim yang optimal dari
keuntungan antara lain peralatan dan bahan yang digunakan relatif murah, dapat
menganalisis jumlah sampel yang banyak dalam waktu singkat, dan dapat
dilakukan pada fase bibit, sehingga menghemat waktu, tempat dan biaya (Hadiati
dan Sukmadjaja 2002).
Karakterisasi Molekuler dengan ISSR
Penanda molekuler merupakan alat yang bermanfaat untuk menunjukkan
keragaman genetik, menentukan tetua dan mengetahui hubungan filogenetik di
antara berbagai spesies jeruk (Uzun et al. 2010). Salah satu metode analisis
molekuler yang dapat mengatasi kekurangan pada metode lainnya, adalah
Inter-Simple Sequence Repeat (ISSR). Teknik ISSR merupakan metode berbasis PCR
yang melibatkan amplifikasi segmen DNA yang ada pada jarak yang dapat
diamplifikasi di antara dua wilayah mikrosatelit berulang yang menuju arah
berlawanan (Reddy et al. 2002).
Metode ISSR dapat mengatasi daya ulang (reproduceability) yang rendah
pada RAPD, biaya tinggi pada AFLP (Amplified Fragment Length Polymorphism)
dan perlunya pengetahuan tentang sekuens DNA tanaman yang akan dianalisis
pada SSR (Reddy et al. 2002). Pada analisis keragaman jeruk siam, primer ISSR
mendeteksi lebih banyak pita DNA dibandingkan dengan primer RAPD, dan
mampu membangun dendrogram yang memperlihatkan keragaman besar
(Agisimanto et al. 2007). Teknik ISSR juga cepat, dapat dipercaya, hanya
memerlukan sejumlah kecil DNA, dan tidak bersifat radioaktif (Jabbarzadeh et al.
2010), karena selain menggunakan poliakrilamida yang dikombinasikan dengan
radioaktif, hasil PCR-ISSR juga dapat dideteksi menggunakan agarose-etidium
bromida (Reddy et al. 2002). Walaupun demikian pada tanaman tertentu, seperti
tanaman obat Scutellaria baicalensis, tetap diperlukan pemilihan primer, karena
primer yang sama dapat menunjukkan hasil amplifikasi berbeda pada spesies yang
berbeda (Guo et al. 2009).
Lebih lanjut Behera et al. (2008) melaporkan bahwa pada berbagai aksesi
paria (Momordica charantia L.) terdapat korelasi tinggi antara hasil analisis
dengan penanda ISSR dan RAPD. Kemungkinan karena sistem penanda ini
fragmen-fragmen yang dideteksi hampir semuanya berukuran sama. Menurut Kumar et al.
(2009) untuk identifikasi dan sertifikasi plasma nutfah jeruk penanda ISSR lebih
baik dibandingkan RAPD.
Shahnavar et al. (2007) berhasil menggunakan penanda ISSR untuk
membedakan aksesi-aksesi jeruk berkerabat dekat dan menjelaskan hubungan
kekerabatan antara genotipe jeruk yang belum teridentifikasi dengan kultivar yang
telah diketahui. Penanda ISSR juga dapat membedakan bibit jeruk nuselar dan
zigotik (Krueger 2003). Sementara Emel (2010) memanfaatkan ISSR untuk
mengidentifikasi DNA unik yang dapat menjadi penanda untuk menentukan
kultivar gandum.
Tingkat Ploidi
Jumlah kromosom setiap sel pada semua individu dari setiap spesies
adalah konstan. Spesies-spesies yang mempunyai hubungan kekerabatan dekat
lebih kurang mempunyai jumlah kromosom sama, sedangkan spesies yang tidak
mempunyai hubungan, memiliki jumlah kromosom berbeda (Sudarnadi 1989).
Hasil penelitian Frost (1925a) menunjukkan kultivar jeruk berbiji bersifat
diploid, dengan jumlah kromosom 2n = 18. Selain itu terdapat pula kultivar jeruk
yang tetraploid (Frost 1925b) dan triploid. Tanaman triploid dapat diperoleh dari
hasil persilangan antara tanaman diploid dengan tetraploid (Fatima et al. 2002),
hibridisasi somatik antara kultivar diploid dan haploid (Kobayashi et al. 1997),
kultur endosperma (Raza et al. 2003), iradiasi (Zhang et al. 1988) atau
terbentuk secara spontan (Jaskani et al. 2007). Pada jeruk, triploid spontan juga
terdapat pada bibit zigotik seksual (Raza et al. 2003).
Jeruk tidak berbiji di Indonesia kemungkinan terbentuk secara spontan,
sebagai hasil persilangan alami antara kultivar diploid dan tetraploid atau mutasi
alami, karena mutasi alami dan sport sering terjadi pada jeruk (Raza et al. 2003).
Secara morfologi, terdapat perbedaan antara tanaman jeruk yang tetraploid,
triploid dan diploid. Tanaman jeruk tetraploid tumbuh lebih cepat, memiliki daun
lebih lebar, lebih tebal, dan berwarna lebih gelap dibanding tanaman triploid dan
diidentifikasi karena ukuran bijinya yang 1/3 sampai 1/6 kali lebih kecil dari biji
diploid (Esen dan Soost 1971).
Viabilitas Tepung Sari
Penentuan viabilitas tepung sari berperan besar dalam proses reproduksi
tanaman, karena dapat menunjukkan kemampuan butir tepung sari untuk
menghantarkan sel sperma ke kantong embrio pada proses penyerbukan.
Pengujian viabilitas ini dapat dilakukan dengan berbagai cara, antara lain dengan
metode pewarnaan tepung sari dan perkecambahan tabung sari secara in vitro.
Selain itu viabilitas tepung sari juga dapat diduga dari ukuran diameternya. Pada
Collinsia verna (Scrophulariaceae; 2n = 14), tepung sari yang hidup (viable)
memiliki diameter lebih besar dibanding yang non-viable (Kelly et al. 2002).
Metode pewarnaan banyak digunakan untuk menduga viabilitas tepung
sari, karena mudah dilakukan, walaupun cara ini memiliki derajat ketepatan yang
lebih rendah dibandingkan metode perkecambahan. Di antara berbagai metode
pewarnaan (acetic-orcein, acetic-carmin, IKI, acridine orange, tetrazolium
klorida), penggunaan tetrazolium klorida pada tepung sari Helleborus niger
berkorelasi lebih baik dengan kemampuan tepung sari berkecambah
(Heslop-Harrison et al. 1984).
Pembentukan Biji dan Buah pada Pamelo
Peran penyerbukan dalam proses pembentukan buah jeruk bervariasi.
Aksesi dengan derajat partenokarpi tinggi (jeruk nipis ‘Tahiti’, jeruk keprok
‘Satsuma’) tidak memerlukan penyerbukan untuk membentuk buah. Sementara
itu, pada aksesi dengan derajat partenokarpi rendah (grapefruit ‘Star-ruby’)
penyerbukan diperlukan untuk membentuk buah (fruit set) (Varoquaux et al.
2000). Aksesi dengan derajat partenokarpi tinggi biasanya menghasilkan buah
tidak berbiji (Iglesias et al. 2007), karena bakal buah mampu berkembang tanpa
pembuahan pada bakal biji (Varoquaux et al. 2000).
Pada jeruk yang berbiji penyerbukan amat mempengaruhi keberhasilan
fruit set dan perkembangan buah selanjutnya. Pada aksesi berbiji, bila sel telur
maka tidak akan terjadi perkembangan biji, dan kantong induk megaspora akan
gugur ketika bunga mengalami senesen. Hal ini menunjukkan peranan biji yang
amat penting untuk pertumbuhan dan perkembangan buah, karena biji merupakan
sumber fitohormon (Ben-Cheikh et al. 1997).
Sifat tidak berbiji pada jeruk disebabkan oleh sterilitas jantan (Yamamoto
et al. 1995) dan self-incompatibility (Yamamoto et al. 2006). Self-incompatibility
pada jeruk dapat disebabkan oleh pertumbuhan tabung tepung sari yang lambat,
diduga akibat adanya inhibitor pada tangkai putik. Hal ini menyebabkan tangkai
putik gugur sebelum tabung tepung sari dapat mencapai kantung embrio dan
melepas inti sperma ke kantung induk megaspora (Krezdorn tanpa tahun). Aksesi
yang self-incompatible menunjukkan derajat partenokarpi rendah, sehingga
dianggap memiliki ’facultative parthenorcapy’. Dalam hal ini buah tidak berbiji
hanya terbentuk ketika tidak terjadi pembuahan (Iglesias et al. 2007). Sementara
itu jantan steril dapat disebabkan oleh perkembangan benang sari (stamen) yang
tidak sempurna atau perkembangan tepung sari yang terganggu. Sterilitas tepung
sari ditemukan pada berbagai tingkat pada banyak aksesi jeruk (Jackson dan
Gmitter tanpa tahun). Pada beberapa spesies, buah tidak berbiji terbentuk sebagai
hasil partenokarpi atau stenospermokarpi, yaitu pembuahan yang diikuti dengan
aborsi pasca-zigotik (Gomez-Alverado et al. 2004).
Peranan Zat Pengatur Tumbuh dalam Perkembangan Biji dan Buah
Perkembangan biji dan buah merupakan proses yang saling berhubungan
erat dan dipengaruhi oleh fitohormon (Gillaspy et al. 1993). Fitohormon yang
terlibat antara lain auksin, sitokinin dan giberelin (GA), yang berperan pada tahap
awal perkembangan buah, terutama pada fase perkembangan bakal buah dan
periode pembelahan sel secara cepat (Srivastava 2002). Hal ini sejalan dengan
ditemukannya peningkatan kandungan auksin, giberelin dan sitokinin pada
organ-organ bunga setelah bakal buah mengalami fertilisasi, sehingga ketiga fitohormon
ini efektif untuk menginduksi perkembangan buah tanpa fertilisasi
(partenokarpik), seperti pada tomat dan terong (Gillaspy 1993). Walaupun
demikian banyak hasil penelitian menunjukkan giberelin yang paling berpengaruh
Menurut Talon et al. (1992) GA endogen pada bakal buah yang sedang
berkembang berperan mengendalikan perkembangan buah partenokarpik (jeruk
mandarin ’Satsuma’ dan ’Clementine’). Disamping itu konsentrasi GA pada buah
partenokarpik (mandarin ’Satsuma’) lebih tinggi dibandingkan buah
non-partenokarpik, seperti ’Hyuganatsu’ (Citrus unshiu) (Kojima 1997). Lebih lanjut,
Altaf dan Khan (2007) menunjukkan bahwa jeruk mandarin ’Kinnow’ berbiji
sedikit (0-10 biji/buah) memiliki derajat partenokarpi rendah, karena kandungan
GA endogennya rendah. Pada aksesi demikian pembentukan buah tidak berbiji
dapat dilakukan dengan penyemprotan GA untuk menginduksi partenokarpi.
Evaluasi Kualitas Buah
Evaluasi agronomi aksesi ditujukan untuk mempermudah pemanfaatan
plasma nutfah berdasarkan sifat agronomi, antara lain berupa kualitas buah.
Menurut IPGRI (1999), kualitas buah yang diamati dapat berupa kandungan
minyak esensial pada kulit buah, kandungan asam tertitrasi total (ATT), gula, pH,
nisbah padatan terlarut total (PTT)/ATT dan kandungan asam askorbat buah.
Disamping itu evaluasi kegetiran (bitterness) merupakan hal penting pada pamelo,
karena rasa getir mempengaruhi kesukaan konsumen terhadap pamelo dan
prospek pemanfaatannya dalam industri jus.
Hasil penelitian Mahardika dan Susanto (2003) pada pamelo ‘Sri Nyonya’,
‘Nambangan’ dan ‘Bali Merah’ menunjukkan ‘Sri Nyonya’ memiliki kandungan
PTT relatif lebih tinggi dibanding ‘Nambangan’ dan ‘Bali Merah’. Nisbah
PTT/ATT ‘Nambangan’ dan ‘Bali Merah’ lebih tinggi dibanding ‘Sri Nyonya’.
Disamping itu, Ketsa (1989) menunjukkan ketebalan kulit buah pada tangerine
(Citrus reticulata Blanco) tidak berpengaruh terhadap kandungan PTT dan asam
askorbat, tetapi tangerine berkulit tipis memiliki ATT lebih rendah dan nisbah
PTT/ATT lebih tinggi dibanding yang berkulit tebal. Hal ini membuat rasa
tangerine berkulit tipis lebih enak dibanding tangerine yang berkulit tebal. Selain
dipengaruhi oleh faktor genetik, kandungan PTT pada buah juga dipengaruhi oleh
nisbah jumlah daun/buah. Pada tomat peningkatan rasio daun/buah akan
Kualitas buah juga berhubungan dengan warna jus dan rasa getir. Pamelo
dengan warna jus merah memiliki kandungan fenolik total dan karotenoid lebih
tinggi dibandingkan yang warna jusnya putih, sehingga merupakan sumber
antioksidan yang baik dan lebih efisien dalam menangkap berbagai bentuk radikal
bebas (Tsai et al. 2007). Rasa getirpada buah dan jus jeruk terutama disebabkan
oleh senyawa dari kelompok flavonoid dan limonoid. Flavonoid pada buah jeruk
terdiri atas flavanon (naringin), flavon (nobiletin) dan flavonol (quercetin).
Naringin merupakan flavanon paling tinggi kandungannya pada jus pamelo
(Pichaiyongvongdee dan Haruenkit 2009b). Pada grapefruit naringin disintesis di
daun muda yang sedang tumbuh, dan ditranslokasikan ke bagian tanaman lain
(Moriguchi et al 2003).
Limonoid pada jeruk berada dalam bentuk aglikon limonoid dan glukosida
limonoid. Dari 16 kultivar pamelo matang rata-rata mengandung 18 ppm limonin
dan 29 ppm limonoid glukosida total. Dibandingkan dengan jus lain, pamelo
mengandung limonin dengan konsentrasi amat tinggi dan konsentrasi limonoid
glukosida amat rendah (Ohta dan Hasegawa 2006).
Konsentrasi senyawa pembuat rasa getir tertinggi umumnya ditemukan
pada buah mentah. Konsentrasi senyawa penyebab rasa getir semakin menurun
dengan makin masaknya buah (Hasegawa et al. 1996). Hasil penelitian
Pichaiyongvongdee dan Haruenkit (2009a) menunjukkan bahwa naringin dan
limonin pada pamelo tersebar pada flavedo, albedo, selaput pembungkus, jus dan
3. KARAKTERISASI MORFOLOGI, BIOKIMIA, MOLEKULER DAN TINGKAT PLOIDI AKSESI PAMELO BERBIJI DAN TIDAK BERBIJI
(Morphological, Biochemical, Molecular Marker and Ploidy Level of Seeded and Seedless Pummelo Accessions)
ABSTRAK
Penelitian ini bertujuan untuk mengevaluasi karakter morfologi, biokimia (isoenzim), molekuler (ISSR) dan tingkat ploidi aksesi pamelo berbiji dan tidak berbiji asal Sumedang, Kudus, Pati dan Magetan. Karakter morfologi yang berperan dalam pengelompokan aksesi pamelo adalah tebal epikarp, pinggiran helai daun, panjang kantong jus, warna kulit buah masak, lebar sayap daun dan bentuk buah, sedangkan karakter isoenzim adalah MDH (Rf 0.11 dan 0.14) dan ACP (Rf 0.24 dan 0.33). Pita ACP Rf 0.24 dapat dijadikan penanda untuk membedakan aksesi berbiji dan tidak berbiji. Karakter molekuler yang berperan dalam pengelompokan adalah PKBT8 500 bp, PKBT8 375 bp, PKBT7 750 bp, PKBT3 750 bp, PKBT8 625 bp dan PKBT12 500 bp. Dendrogram berdasarkan karakter morfologi memisahkan kelompok aksesi berbiji dan tidak berbiji pada koefisien kemiripan 0.63, dengan isoenzim pada koefisien kemiripan 0.48, sedangkan berdasarkan molekuler belum dapat memisahkan antara aksesi berbiji dan tidak berbiji. Dendrogram berdasarkan karakter morfologi dan isoenzim dapat membedakan antara aksesi berbiji, potensial tidak berbiji dan tidak berbiji. Hasil pemetaan komponen utama kongruen dengan dendrogram, yaitu dapat memisahkan aksesi berbiji maupun tidak berbiji, berdasarkan karakter morfologi, isoenzim maupun kombinasinya. Baik pamelo berbiji maupun tidak berbiji memiliki kromosom diploid (2n = 2x = 18).
ABSTRACT
PENDAHULUAN
Pamelo (Citrus maxima (Burm.) Merr.) merupakan spesies jeruk
berukuran paling besar, dan populer di Cina bagian Selatan, Thailand dan
negara-negara Asia Tenggara (Christmann 2008), termasuk Indonesia (Morton 1987). Hal
inilah yang membuat aksesi pamelo dibedakan atas kelompok Thailand, Cina dan
Indonesia (Davies dan Albrigo 1994). Selain ukuran buahnya yang khas, aksesi
pamelo juga memiliki jumlah biji beragam, mulai dari tidak berbiji hingga berbiji
banyak (Ladaniya 2008). Buah tidak berbiji lebih banyak diminati oleh
konsumen, karena biji menyebabkan rasa pahit dan merepotkan saat
mengkonsumsi buah (Altaf dan Khan 2007), sehingga pengembangan jeruk
diarahkan pada aksesi tidak berbiji.
Upaya pengembangan aksesi tidak berbiji melalui program pemuliaan dan
pemanfaatan plasma nutfah, memerlukan informasi keanekaragaman genetik dan
hubungan kekerabatan antar kultivar pamelo. Informasi ini dapat diperoleh
melalui karakterisasi menggunakan berbagai metode analisis. Kombinasi
informasi dari tipe analisis berbeda diharapkan dapat meminimalkan risiko
kesalahan dalam menginterpretasikan data (Petrokas dan Stanys 2008).
Identifikasi tanaman secara morfologi dilakukan karena adanya
kemudahan dalam mengamati perbedaan-perbedaan antara tanaman secara visual.
Selain itu kebanyakan karakter hotikultura penting dikendalikan oleh banyak gen
(Campos et al. 2005), dan memiliki derajat heritabilitas rendah, sehingga sebagian
karakter hortikultura tidak dapat dievaluasi melalui penanda molekuler (Dorji dan
Yapwattanaphun 2011). Hal ini membuat karakterisasi morfologi tetap
diperlukan.
Karakter morfologi merupakan ekspresi fenotipe dari individu dan
populasi, diregulasi dan ditentukan oleh gen dan interaksinya dengan lingkungan.
Beberapa peneliti telah menggunakan karakterisasi morfologi pada jeruk, antara
lain untuk meningkatkan jumlah genotipe yang potensial dalam program
pemuliaan atau untuk dilepas sebagai kultivar baru jeruk keprok (Citrus spp.)
genetik spesies jeruk (Hardiyanto et al. 2007), dan untuk mendapatkan kultivar
unggul pamelo (Ara et al. 2008).
Di lain pihak penggunaan karakter morfologi relatif sulit membedakan
sifat antar aksesi jeruk, karena beberapa aksesi hanya dapat dibedakan melalui
sifat buahnya (Quang et al. 2011). Sementara panen raya pamelo hanya terjadi
sekali dalam setahun dan masa vegetatif jeruk relatif panjang. Oleh karena itu
data karakter morfologi, perlu didukung dengan analisis lain, diantaranya
isoenzim, molekuler dan set kromosom.
Isoenzim merupakan enzim yang terdiri atas molekul-molekul yang
mempunyai struktur kimia yang berbeda akan tetapi mengkatalisis reaksi kimia
yang sama. Sebagai protein, isoenzim secara langsung dapat menunjukkan
perubahan dalam sekuen DNA melalui perbedaan komposisi asam amino.
Seringkali perbedaan komposisi asam amino akan mengubah muatan enzim, yang
akan menyebabkan perubahan dalam mobilitas elektroforesis (Weeden dan
Wendel 1989). Hal ini merupakan indikator yang baik untuk keragaman genetik,
sehingga isoenzim dapat digunakan untuk mengidentifikasi aksesi pada berbagai
tanaman, seperti pamelo (Phan et al. 2006), Citrus junos dan kerabat jeruk asam
(Rahman et al. 2001), nenas (Hadiati dan Sukmadjaja 2002), dan padi (Abdullah,
2001). Analisis isoenzim juga digunakan untuk membedakan bibit jeruk triploid
dan diploid (King et al. 1996) dan batang bawah jeruk nuselar dan zigotik (Stykes
2011). Di lain pihak belum diketahui kemampuan isoenzim dalam membedakan
antara aksesi berbiji dan tidak berbiji pada pamelo.
Teknologi penanda molekuler berperan penting dalam membedakan
keragaman genetik, mengidentifikasi hubungan kekerabatan dan memetakan sidik
jari plasma nutfah. Salah satu metode analisis molekuler yang telah digunakan
secara luas adalah Inter-Simple Sequence Repeat (ISSR). Menurut Kumar et al.
(2009) marka ISSR dapat membedakan individu aksesi dalam setiap spesies jeruk
(C. indica, C. medica, C. latipes dan C. sp. ‘Memang athur’) dengan polimorfisme
sedang (38-56 persen). ISSR mampu mendeteksi keragaman genetik yang lebih
besar pada Vigna umbellata dibandingkan RAPD (Muthusamy et al. 2008), dan
ISSR juga lebih berguna untuk identifikasi plasma nutfah jeruk dibandingkan
Analisis set kromosom diperlukan untuk mengetahui kemungkinan adanya
aksesi tidak berbiji yang triploid, karena jumlah kromosom pada jeruk
mempengaruhi pembentukan dan perkembangan biji. Tanaman jeruk triploid (3n)
biasanya menghasilkan buah tidak berbiji (Toolapong et al. 1995). Kondisi
triploid ini menyebabkan meiosis yang abnormal dan aborsi embrio (Zhu et al.
2009).
Penelitian ini bertujuan untuk mengidentifikasi karakter morfologi,
isoenzim, molekuler dan tingkat ploidi pamelo berbiji, potensial tidak berbiji dan
tidak berbiji dan mendapatkan informasi keanekaragaman genetik dan hubungan
kekerabatan antar aksesi pamelo.
BAHAN DAN METODE Waktu dan Tempat
Karakterisasi morfologi dilakukan pada bulan April 2009 sampai April
2010 di sentra produksi pamelo Sumedang, Gunung Muria (Kudus dan Pati) dan
Magetan. Analisis isoenzim dikerjakan pada bulan Maret 2011 di Laboratorium
Pusat Penelitian Sumberdaya Hayati dan Bioteknologi IPB. Analisis molekuler
dengan ISSR dilakukan pada bulan Maret-Juni 2011 di Laboratorium Pusat Kajian
Buah Tropika IPB. Konfirmasi tingkat ploidi dengan analisis jumlah kromosom
dilakukan di Laboratorium Mikroteknik, Departemen Agronomi dan Hortikultura
IPB pada bulan September 2010 sampai Pebruari 2011, dan analisis flow
cytometry dilakukan di Laboratorium Genetika Tumbuhan, LIPI Biologi,
Cibinong pada bulan Pebruari 2012.
Karakterisasi Morfologi pada Aksesi Pamelo Berbiji dan Tidak Berbiji Bahan dan Alat
Bahan yang digunakan ialah bagian-bagian vegetatif dan reproduktif
tanaman 14 aksesi tanaman pamelo asal Sumedang: Cikoneng ST, Magetan: Jawa
1, Jawa 2, Jawa 3, Magetan, Sri Nyonya, Adas Duku, Bali Putih, Nambangan,
Bali Merah 1, Bali Merah 2, Kudus: Muria Merah 1, Muria Merah 2 dan Pati:
Bageng Taji yang tumbuh di kebun petani. Alat yang digunakan ialah pH meter
Metode
Identifikasi pamelo secara morfologi dilakukan berdasarkan descriptor list
IPGRI (1999) yang telah dimodifikasi (Lampiran 1). Tanaman pamelo yang
diamati telah dewasa (telah berbuah), berumur lebih dari 6 tahun, dengan asumsi
cir-ciri morfologi dan fisiologi tanaman telah stabil (secara visual sifat genetik
telah tampak dan stabil) dan pertumbuhan tanaman tampak normal, tidak ada
indikasi atau gejala kekurangan hara. Metode pemilihan contoh dilakukan secara
terarah, yaitu dari setiap lokasi yang dipilih dalam satu daerah diamati sebanyak
tiga pohon contoh yang dapat mewakili populasi tanaman.
Pengamatan karakter kuantitatif dan kualitatif dievaluasi dari 10 daun, 10
bunga dan 10 buah dari setiap tanaman. Dalam penelitian ini aksesi pamelo
dikelompokkan tidak berbiji, bila jumlah biji per buah kurang dari 10, dimasukkan
kelompok potensial tidak berbiji, jika dalam satu aksesi terdapat buah berbiji dan
tidak berbiji, dan kelompok buah berbiji jika jumlah biji per buah ≥10.
Karakterisasi Biokimia dengan Isoenzim pada Aksesi Pamelo Berbiji dan Tidak Berbiji
Bahan dan Alat
Bahan tanaman yang digunakan untuk analisis ialah daun muda pamelo
aksesi berbiji (Cikoneng ST, Jawa 2, Jawa 3, Magetan, Sri Nyonya, Adas Duku,
Bali Putih dan Muria Merah 2), tidak berbiji (Muria Merah 1, Bali Merah 2,
Bageng Taji, Jawa 1) dan potensial tidak berbiji (Nambangan dan Bali Merah 1),
yang diperoleh dari hasil eksplorasi di tiga sentra produksi pamelo di Sumedang,
Gunung Muria (Pati dan Kudus) dan Magetan.
Metode
Teknik analisis isoenzim esterase (EST), malat dehidrogenase (MDH),
peroksidase (PER), dan asam fosfatase (ACP) mengikuti metode Horry (1989),
sedangkan aspartat amino transferase (AAT) atau glutamat oksaloasetat