i
KOMBINASI ISOLAT BAKTERI SIMBION RAYAP DENGAN
ISOLAT BAKTERI RUMEN DALAM MENINGKATKAN
NILAI GUNA PAKAN SUMBER SERAT
SKRIPSI NOVIANTO
DEPARTEMEN ILMU NUTRISI DAN TEKNOLOGI PAKAN FAKULTAS PETERNAKAN
ii RINGKASAN
NOVIANTO. D24051401. 2009. Kombinasi Isolat Bakteri Simbion Rayap Dengan Isolat Bakteri Rumen Dalam Meningkatkan Nilai Guna Pakan Sumber Serat. Skripsi. Departemen Ilmu Nutrisi dan Teknologi Pakan. Fakultas Peternakan. Institut Pertanian Bogor.
Pembimbing Utama : Ir. Anita S Tjakradidjaja, MRur. Sc. Pembimbing Anggota : Dr. Ir. Panca Dewi MHKS, MSi.
Seiring dengan peningkatan lahan pertanian terutama untuk produksi padi dan kelapa sawit maka meningkat pula hasil limbah dari sektor pertanian tersebut seperti jerami padi dan serat sawit. Kendala yang dihadapi dalam penggunaan limbah tersebut adalah kandungan nutrien yang rendah dan tingginya serat kasar. Rayap merupakan serangga sosial pendegradasi kayu yang banyak mengandung selulosa, hemiselulosa dan lignin. Dari penelitian sebelumnya telah diisolasi 13 isolat bakteri rayap dan 3 isolat bakteri rumen domba yang mampu mencerna pakan serat, dan dari ke-13 isolat bakteri rayap, isolat A {SB 53 5(3)} dan isolat D {SC 51 5(2)} adalah isolat bakteri rayap terbaik yang dapat dikombinasikan dalam mencerna pakan sumber serat, namun belum diketahui pengaruh kombinasi antara isolat bakteri rayap dengan isolat bakteri rumen dalam mencerna pakan serat. Penelitian ini bertujuan untuk mempelajari kombinasi terbaik dari isolat bakteri rayap dengan isolat bakteri rumen domba dalam mencerna pakan sumber serat yaitu serat sawit, jerami padi dan rumput gajah.
Penelitian ini menggunakan Rancangan Acak Kelompok (RAK) dengan pola faktorial 6 X 3. Faktor A adalah kombinasi isolat bakteri simbion rayap dan isolat bakteri rumen yaitu: T1= A + SE 511, T2= A + SE 512, T3= A + SE 513, T4= D + SE 511, T5= D + SE 512, dan T6= D + SE 513. Isolat A adalah SB 53 5(3)1 yang berasal dari rayap Coptotermes curvignathus Holmgren dan isolat D adalah SC 51 5(2) yang berasal dari rayap Microtermes inspiratus Kemner. Faktor B adalah bahan pakan sumber serat yaitu rumput gajah, jerami padi dan serat sawit dengan 3 ulangan sebagai kelompok. Peubah yang diamati yaitu konsentrasi amonia (NH3), produksi
asam lemak terbang (VFA) total, degradasi bahan kering (DBK), degradasi bahan organik (DBO), koefisien cerna bahan kering (KCBK) dan koefisien cerna bahan organik (KCBO). Analisis data menggunakan Sidik Ragam (ANOVA); apabila hasil uji tersebut berbeda nyata maka dilanjutkan dengan uji ortogonal kontras untuk memperoleh perlakuan terbaik.
Hasil analisis ragam memperlihatkan bahwa penggunaan pakan sumber serat berbeda sangat nyata (P<0,01) pada semua variabel kecuali VFA (P<0,05). Jerami padi mempunyai konsentrasi VFA lebih tinggi (P<0,01) dibandingkan rumput gajah dan serat sawit. Rumput gajah dan jerami padi mempunyai konsentrasi NH3 lebih
tinggi (P<0,01) dibandingkan dengan serat sawit. Persentase DBK dan DBO tertinggi (P<0,01) pada rumput gajah, sedangkan persentase KCBK atau KCBO rumput gajah dan jerami padi lebih tinggi (P<0,01) dibandingkan serat sawit. Pada kombinasi isolat bakteri, KCBK dan KCBO berbeda nyata (P<0,05), sedangkan VFA, NH3,
iii ini menunjukkan bahwa kombinasi antara isolat bakteri rayap dengan isolat bakteri rumen mempunyai hubungan yang sinergis dalam mendegradasi pakan sumber serat. Proses degradasi dan pencernaan pakan berserat terbaik diperoleh ketika isolat D (SE 51 5(2)) dikombinasikan dengan isolat SE 511, SE 512 dan SE 513. Dibandingkan rumput gajah, kokultur antara isolat bakteri rayap dan rumen domba selama 6 jam waktu inkubasi dapat memperbaiki fermentabilitas dan kecernaan jerami padi, tetapi efek yang sama belum dapat diperoleh pada serat sawit.
iv ABSTRACT
Combination Between Termite Bacterial and Rumen Bacterial Isolates In Increasing Nutrient Utilitation of Fibrous Feeds
Novianto, A. S. Tjakradidjaja and P. Dewi MHKS
The aim of this experiment was to determine the best combination between termite bacteria and rumen bacteria to degradate fibrous feeds. This experiment used randomized block design with factorial design (3x6) and 3 replications. The first factor was combination between termite bacteria and rumen bacteria {T1 (A + SE 511), T2 (A + SE 512), T3 (A + SE 513), T4 (D + SE 511), T5 (D + SE 512), and T6 (D + SE 513)}, and the second factor was fibrous feeds (napier grass, rice straw and palm press fibre). A (SB 53 5(3)1) and D (SC 51 5(2)) were isolates of termite fibrous feeds significantly affected all variables measured (P<0.01). Rice straw had higher (P<0.01) VFA concentration than napier grass and palm press fibre. Napier grass and rice straw had higher (P<0.01) NH3 concentration than palm press fibre.
Degradability of DM and OM of napier grass were higher than those of rice straw and palm press fibre (P<0.01). Differences in combination between termite bacteria and rumen bacteria significantly affected digestibilities of DM and OM. Treatments of T2, T4, T5 and T6 had higher (P<0.05) digestibility of DM than T1 and T3. Treatments of T4, T5, and T6 had higher digestibility of OM than T1, T2, and T3. It is concluded that the combination between termite bacteria and rumen bacteria have synergic relationship in degradating fibrous feeds. The best combination are T4, T5 and T6 in digesting fibrous feeds with the best isolate of termite bacteria is isolate D {SC 51 5(2)}. In comparison to napier grass, coculture between termite and sheep rumen bacterial isolates for 6 hour incubating period has improved fermentability and digestibility of rice straw, however, no such improvement was obtained in fermentability and digestibility of palm press fibre.
v
KOMBINASI ISOLAT BAKTERI SIMBION RAYAP DENGAN
ISOLAT BAKTERI RUMEN DALAM MENINGKATKAN
NILAI GUNA PAKAN SUMBER SERAT
NOVIANTO D24051401
Skripsi ini merupakan salah satu syarat untuk memperoleh gelar Sarjana Peternakan
pada Fakultas Peternakan Institut Pertanian Bogor
DEPARTEMEN ILMU NUTRISI DAN TEKNOLOGI PAKAN FAKULTAS PETERNAKAN
vi
KOMBINASI ISOLAT BAKTERI SIMBION RAYAP DENGAN
ISOLAT BAKTERI RUMEN DALAM MENINGKATKAN
NILAI GUNA PAKAN SUMBER SERAT
Oleh : NOVIANTO
D24051401
Skripsi ini telah disetujui dan disidangkan di hadapan Komisi Ujian Lisan pada tanggal 10 Agustus 2009
Pembimbing Utama Pembimbing Anggota
Ir. Anita S Tjakradidjaja, MRur. Sc. Dr. Ir. Panca Dewi MHKS, Msi.
NIP. 19610930 198603 2 003 NIP. 19611025 198703 2 002
Dekan Ketua Departemen
Fakultas Peternakan Ilmu Nutrisi dan Teknologi Pakan Institut Pertanian Bogor Fakultas Peternakan
Institut Pertanian Bogor
vii RIWAYAT HIDUP
Penulis dilahirkan di Jakarta pada tanggal 25 November 1986 dari pasangan Bapak Sukoyo dan Ibu Mujiani Susanti. Penulis merupakan anak pertama dari dua bersaudara. Penulis memulai pendidikan di taman kanak-kanak yang diselesaikan pada tahun 1993 di TK Kartika Bhakti VIII Pekayon, Jakarta dan pendidikan dasar di SDN 01 Pagi Pekayon, Jakarta yang diselesaikan pada tahun 1999. Pendidikan lanjutan diselesaikan pada tahun 2002 di SLTPN 103 Cijantung, Jakarta dan pendidikan lanjutan menengah atas diselesaikan pada tahun 2005 di SMU 39 Cijantung, Jakarta. Penulis diterima sebagai mahasiswa Tingkat Persiapan Bersama Institut Pertanian Bogor melalui jalur SPMB pada tahun 2005 dan terdaftar pada Departemen Ilmu Nutrisi dan Teknologi Pakan, Fakultas Peternakan, Institut Pertanian Bogor pada tahun 2006.
viii KATA PENGANTAR
Alhamdulillahi rabbil’alamin, puji dan syukur penulis panjatkan kehadirat Allah SWT atas rahmat, karunia dan ridho-Nya sehingga penulis dapat menyelesaikan penelitian dan penyusunan skripsi ini. Skripsi yang berjudul ” Kombinasi Isolat Bakteri Simbion Rayap Dengan Isolat Bakteri Rumen Dalam Meningkatkan Nilai Guna Pakan Sumber Serat” merupakan salah satu syarat untuk memperoleh gelar Sarjana Peternakan di Fakultas Peternakan, Institut Pertanian Bogor. Penelitian ini berlangsung dari bulan September 2008 hingga Februari 2009 di Laboratorium Biokimia, Fisiologi dan Mikrobiologi Nutrisi, dan Laboratorium Nutrisi Ternak Perah, Departemen Ilmu Nutrisi dan Teknologi Pakan, Fakultas Peternakan, Institut Pertanian Bogor.
Skripsi ini disusun dengan harapan isoat bakteri rayap dapat dijadikan sumber probiotik untuk ternak ruminansia sehingga dapat mencerna pakan berserat lebih baik lagi. Penulis menyadari bahwa penulisan skripsi ini masih banyak kekurangan, oleh sebab itu penulis mengharapkan adanya sumbangan pemikiran, baik berupa kritik maupun saran sehingga skripsi ini menjadi lebih baik. Penulis mengucapkan banyak terima kasih kepada seluruh pihak yang telah ikut berperan sehingga penulisan skripsi ini dapat terselesaikan. Semoga skripsi ini bermanfaat terutama di dunia peternakan Amin.
Bogor, Agustus 2009
x
Degradasi Bahan Kering dan Degradasi Bahan Organik ... 27
Pencernaan Hidrolisis Aerob ... 28
Koefisien Cerna Bahan Kering (KCBK) ... 28
Koefisien Cerna Bahan Organik (KCBO) ... 29
HASIL DAN PEMBAHASAN ... 30
Konsentrasi VFA Total ... 30
Konsentrasi NH3 ... 32
Degradasi Bahan Kering ... 34
Degradasi Bahan Organik ... 37
Koefisien Cerna Bahan Kering ... 39
Koefisien Cerna Bahan Organik ... 41
KESIMPULAN DAN SARAN ... 44
Kesimpulan... 44
Saran ... 44
UCAPAN TERIMA KASIH ... 45
DAFTAR PUSTAKA ... 46
i
KOMBINASI ISOLAT BAKTERI SIMBION RAYAP DENGAN
ISOLAT BAKTERI RUMEN DALAM MENINGKATKAN
NILAI GUNA PAKAN SUMBER SERAT
SKRIPSI NOVIANTO
DEPARTEMEN ILMU NUTRISI DAN TEKNOLOGI PAKAN FAKULTAS PETERNAKAN
ii RINGKASAN
NOVIANTO. D24051401. 2009. Kombinasi Isolat Bakteri Simbion Rayap Dengan Isolat Bakteri Rumen Dalam Meningkatkan Nilai Guna Pakan Sumber Serat. Skripsi. Departemen Ilmu Nutrisi dan Teknologi Pakan. Fakultas Peternakan. Institut Pertanian Bogor.
Pembimbing Utama : Ir. Anita S Tjakradidjaja, MRur. Sc. Pembimbing Anggota : Dr. Ir. Panca Dewi MHKS, MSi.
Seiring dengan peningkatan lahan pertanian terutama untuk produksi padi dan kelapa sawit maka meningkat pula hasil limbah dari sektor pertanian tersebut seperti jerami padi dan serat sawit. Kendala yang dihadapi dalam penggunaan limbah tersebut adalah kandungan nutrien yang rendah dan tingginya serat kasar. Rayap merupakan serangga sosial pendegradasi kayu yang banyak mengandung selulosa, hemiselulosa dan lignin. Dari penelitian sebelumnya telah diisolasi 13 isolat bakteri rayap dan 3 isolat bakteri rumen domba yang mampu mencerna pakan serat, dan dari ke-13 isolat bakteri rayap, isolat A {SB 53 5(3)} dan isolat D {SC 51 5(2)} adalah isolat bakteri rayap terbaik yang dapat dikombinasikan dalam mencerna pakan sumber serat, namun belum diketahui pengaruh kombinasi antara isolat bakteri rayap dengan isolat bakteri rumen dalam mencerna pakan serat. Penelitian ini bertujuan untuk mempelajari kombinasi terbaik dari isolat bakteri rayap dengan isolat bakteri rumen domba dalam mencerna pakan sumber serat yaitu serat sawit, jerami padi dan rumput gajah.
Penelitian ini menggunakan Rancangan Acak Kelompok (RAK) dengan pola faktorial 6 X 3. Faktor A adalah kombinasi isolat bakteri simbion rayap dan isolat bakteri rumen yaitu: T1= A + SE 511, T2= A + SE 512, T3= A + SE 513, T4= D + SE 511, T5= D + SE 512, dan T6= D + SE 513. Isolat A adalah SB 53 5(3)1 yang berasal dari rayap Coptotermes curvignathus Holmgren dan isolat D adalah SC 51 5(2) yang berasal dari rayap Microtermes inspiratus Kemner. Faktor B adalah bahan pakan sumber serat yaitu rumput gajah, jerami padi dan serat sawit dengan 3 ulangan sebagai kelompok. Peubah yang diamati yaitu konsentrasi amonia (NH3), produksi
asam lemak terbang (VFA) total, degradasi bahan kering (DBK), degradasi bahan organik (DBO), koefisien cerna bahan kering (KCBK) dan koefisien cerna bahan organik (KCBO). Analisis data menggunakan Sidik Ragam (ANOVA); apabila hasil uji tersebut berbeda nyata maka dilanjutkan dengan uji ortogonal kontras untuk memperoleh perlakuan terbaik.
Hasil analisis ragam memperlihatkan bahwa penggunaan pakan sumber serat berbeda sangat nyata (P<0,01) pada semua variabel kecuali VFA (P<0,05). Jerami padi mempunyai konsentrasi VFA lebih tinggi (P<0,01) dibandingkan rumput gajah dan serat sawit. Rumput gajah dan jerami padi mempunyai konsentrasi NH3 lebih
tinggi (P<0,01) dibandingkan dengan serat sawit. Persentase DBK dan DBO tertinggi (P<0,01) pada rumput gajah, sedangkan persentase KCBK atau KCBO rumput gajah dan jerami padi lebih tinggi (P<0,01) dibandingkan serat sawit. Pada kombinasi isolat bakteri, KCBK dan KCBO berbeda nyata (P<0,05), sedangkan VFA, NH3,
iii ini menunjukkan bahwa kombinasi antara isolat bakteri rayap dengan isolat bakteri rumen mempunyai hubungan yang sinergis dalam mendegradasi pakan sumber serat. Proses degradasi dan pencernaan pakan berserat terbaik diperoleh ketika isolat D (SE 51 5(2)) dikombinasikan dengan isolat SE 511, SE 512 dan SE 513. Dibandingkan rumput gajah, kokultur antara isolat bakteri rayap dan rumen domba selama 6 jam waktu inkubasi dapat memperbaiki fermentabilitas dan kecernaan jerami padi, tetapi efek yang sama belum dapat diperoleh pada serat sawit.
iv ABSTRACT
Combination Between Termite Bacterial and Rumen Bacterial Isolates In Increasing Nutrient Utilitation of Fibrous Feeds
Novianto, A. S. Tjakradidjaja and P. Dewi MHKS
The aim of this experiment was to determine the best combination between termite bacteria and rumen bacteria to degradate fibrous feeds. This experiment used randomized block design with factorial design (3x6) and 3 replications. The first factor was combination between termite bacteria and rumen bacteria {T1 (A + SE 511), T2 (A + SE 512), T3 (A + SE 513), T4 (D + SE 511), T5 (D + SE 512), and T6 (D + SE 513)}, and the second factor was fibrous feeds (napier grass, rice straw and palm press fibre). A (SB 53 5(3)1) and D (SC 51 5(2)) were isolates of termite fibrous feeds significantly affected all variables measured (P<0.01). Rice straw had higher (P<0.01) VFA concentration than napier grass and palm press fibre. Napier grass and rice straw had higher (P<0.01) NH3 concentration than palm press fibre.
Degradability of DM and OM of napier grass were higher than those of rice straw and palm press fibre (P<0.01). Differences in combination between termite bacteria and rumen bacteria significantly affected digestibilities of DM and OM. Treatments of T2, T4, T5 and T6 had higher (P<0.05) digestibility of DM than T1 and T3. Treatments of T4, T5, and T6 had higher digestibility of OM than T1, T2, and T3. It is concluded that the combination between termite bacteria and rumen bacteria have synergic relationship in degradating fibrous feeds. The best combination are T4, T5 and T6 in digesting fibrous feeds with the best isolate of termite bacteria is isolate D {SC 51 5(2)}. In comparison to napier grass, coculture between termite and sheep rumen bacterial isolates for 6 hour incubating period has improved fermentability and digestibility of rice straw, however, no such improvement was obtained in fermentability and digestibility of palm press fibre.
v
KOMBINASI ISOLAT BAKTERI SIMBION RAYAP DENGAN
ISOLAT BAKTERI RUMEN DALAM MENINGKATKAN
NILAI GUNA PAKAN SUMBER SERAT
NOVIANTO D24051401
Skripsi ini merupakan salah satu syarat untuk memperoleh gelar Sarjana Peternakan
pada Fakultas Peternakan Institut Pertanian Bogor
DEPARTEMEN ILMU NUTRISI DAN TEKNOLOGI PAKAN FAKULTAS PETERNAKAN
vi
KOMBINASI ISOLAT BAKTERI SIMBION RAYAP DENGAN
ISOLAT BAKTERI RUMEN DALAM MENINGKATKAN
NILAI GUNA PAKAN SUMBER SERAT
Oleh : NOVIANTO
D24051401
Skripsi ini telah disetujui dan disidangkan di hadapan Komisi Ujian Lisan pada tanggal 10 Agustus 2009
Pembimbing Utama Pembimbing Anggota
Ir. Anita S Tjakradidjaja, MRur. Sc. Dr. Ir. Panca Dewi MHKS, Msi.
NIP. 19610930 198603 2 003 NIP. 19611025 198703 2 002
Dekan Ketua Departemen
Fakultas Peternakan Ilmu Nutrisi dan Teknologi Pakan Institut Pertanian Bogor Fakultas Peternakan
Institut Pertanian Bogor
vii RIWAYAT HIDUP
Penulis dilahirkan di Jakarta pada tanggal 25 November 1986 dari pasangan Bapak Sukoyo dan Ibu Mujiani Susanti. Penulis merupakan anak pertama dari dua bersaudara. Penulis memulai pendidikan di taman kanak-kanak yang diselesaikan pada tahun 1993 di TK Kartika Bhakti VIII Pekayon, Jakarta dan pendidikan dasar di SDN 01 Pagi Pekayon, Jakarta yang diselesaikan pada tahun 1999. Pendidikan lanjutan diselesaikan pada tahun 2002 di SLTPN 103 Cijantung, Jakarta dan pendidikan lanjutan menengah atas diselesaikan pada tahun 2005 di SMU 39 Cijantung, Jakarta. Penulis diterima sebagai mahasiswa Tingkat Persiapan Bersama Institut Pertanian Bogor melalui jalur SPMB pada tahun 2005 dan terdaftar pada Departemen Ilmu Nutrisi dan Teknologi Pakan, Fakultas Peternakan, Institut Pertanian Bogor pada tahun 2006.
viii KATA PENGANTAR
Alhamdulillahi rabbil’alamin, puji dan syukur penulis panjatkan kehadirat Allah SWT atas rahmat, karunia dan ridho-Nya sehingga penulis dapat menyelesaikan penelitian dan penyusunan skripsi ini. Skripsi yang berjudul ” Kombinasi Isolat Bakteri Simbion Rayap Dengan Isolat Bakteri Rumen Dalam Meningkatkan Nilai Guna Pakan Sumber Serat” merupakan salah satu syarat untuk memperoleh gelar Sarjana Peternakan di Fakultas Peternakan, Institut Pertanian Bogor. Penelitian ini berlangsung dari bulan September 2008 hingga Februari 2009 di Laboratorium Biokimia, Fisiologi dan Mikrobiologi Nutrisi, dan Laboratorium Nutrisi Ternak Perah, Departemen Ilmu Nutrisi dan Teknologi Pakan, Fakultas Peternakan, Institut Pertanian Bogor.
Skripsi ini disusun dengan harapan isoat bakteri rayap dapat dijadikan sumber probiotik untuk ternak ruminansia sehingga dapat mencerna pakan berserat lebih baik lagi. Penulis menyadari bahwa penulisan skripsi ini masih banyak kekurangan, oleh sebab itu penulis mengharapkan adanya sumbangan pemikiran, baik berupa kritik maupun saran sehingga skripsi ini menjadi lebih baik. Penulis mengucapkan banyak terima kasih kepada seluruh pihak yang telah ikut berperan sehingga penulisan skripsi ini dapat terselesaikan. Semoga skripsi ini bermanfaat terutama di dunia peternakan Amin.
Bogor, Agustus 2009
x
Degradasi Bahan Kering dan Degradasi Bahan Organik ... 27
Pencernaan Hidrolisis Aerob ... 28
Koefisien Cerna Bahan Kering (KCBK) ... 28
Koefisien Cerna Bahan Organik (KCBO) ... 29
HASIL DAN PEMBAHASAN ... 30
Konsentrasi VFA Total ... 30
Konsentrasi NH3 ... 32
Degradasi Bahan Kering ... 34
Degradasi Bahan Organik ... 37
Koefisien Cerna Bahan Kering ... 39
Koefisien Cerna Bahan Organik ... 41
KESIMPULAN DAN SARAN ... 44
Kesimpulan... 44
Saran ... 44
UCAPAN TERIMA KASIH ... 45
DAFTAR PUSTAKA ... 46
xi DAFTAR TABEL
Nomor Halaman
1. Kandungan Nutrien Jenis Substrat ... 13
2. Konsentrasi VFA Total oleh Isolat Bakteri Rayap dan Rumen ... 30
3. Konsentrasi NH3 oleh Isolat Bakteri Rayap dan Rumen ... 33
4. Degradasi Bahan Kering oleh Isolat Bakteri Rayap dan Rumen... 35
5. Degradasi Bahan Organik oleh Isolat Bakteri Rayap dan Rumen ... 38
6. Koefisien Cerna Bahan Kering oleh Isolat Bakteri Rayap dan Rumen . 39
xii DAFTAR GAMBAR
Nomor Halaman
1. Analisis Serat... 5 2. Struktur Selulosa ... 6 3. Struktur Hemiselulosa... 8 4. Struktur Lignin ... 9 5. Ringkasan Pencernaan Karbohidrat pada Ternak Ruminansia... 17 6. Proses Metabolisme Karbohidrat di dalam Rumen Ternak
xiii DAFTAR LAMPIRAN
Nomor Halaman
1. ANOVA Pengaruh Perlakuan terhadap Produksi VFA ... 51 2. ANOVA Pengaruh Perlakuan terhadap Produksi NH3 ... 51
1 PENDAHULUAN
Latar Belakang
Indonesia adalah suatu negara yang dikenal sebagai negara agraris, sehingga pertanian harus menjadi landasan utama dari setiap pembangunan yang ada di Indonesia, baik pembangunan dalam bidang ekonomi, budaya, politik bahkan bidang peternakan. Seiring dengan peningkatan lahan pertanian maka meningkat pula hasil sampingan atau limbah dari sektor pertanian. Limbah-limbah dari sektor pertanian ini masih belum banyak dimanfaatkan oleh masyarakat. Padahal, limbah-limbah tersebut berpotensi besar untuk dijadikan pakan ternak.
Beberapa limbah pertanian yang dapat dimanfaatkan sebagai pakan ternak adalah jerami padi dan serat sawit. Jerami padi sangat berpotensi sebagai pakan ternak sebab jumlah setiap tahunnya meningkat begitu pula dengan serat sawit. Peningkatan limbah serat sawit disebabkan meningkatnya perkebunan kelapa sawit yang ada di Indonesia. Kendala yang dihadapi dari limbah-limbah tersebut adalah kandungan serat kasar yang sangat tinggi dan tingkat kecernaannya yang rendah. Serat kasar yang terkandung dalam serat sawit dan jerami padi tersebut sangat sulit dicerna oleh ruminansia dibandingkan dengan serat kasar yang ada di dalam rumput. Sulitnya mikroorganisme rumen mencerna serat tersebut menyebabkan rendahnya nilai kecernaan dari limbah tersebut.
2 merupakan serangga sosial pendegradasi kayu yang mengandung banyak selulosa, hemiselulosa dan lignin.
Penelitian Setianegoro (2004) menunjukkan bahwa mikroba simbion rayap dapat mencerna pakan sumber serat walaupun hasilnya masih lebih rendah daripada yang diperoleh dari mikroba rumen. Rayap yang digunakan dalam penelitian tersebut adalah Macrotermes gilvus Hagen, Coptotermes curvignathus Holmgren dan Microtermes inspiratus Kemner. Penelitian selanjutnya mendapatkan 13 isolat murni bakteri rayap terbaik dalam mencerna selulosa (Widyastuti, 2005). Berdasarkan uji kemampuan mendegradasi pakan sumber serat (jerami padi, serat sawit, dan rumput gajah) telah diperoleh lima isolat terbaik dalam mendegradasi pakan sumber serat tersebut (Sulistiani, 2005). Pradana (2006) dan Solihat (2006) menguji kemampuan hidup kelima isolat tersebut dalam kondisi rumen. Dari hasil penelitian tersebut didapatkan tiga isolat bakteri simbion rayap terbaik yang berasal dari Coptotermes curvignathus Holmgren yaitu isolat A (SB53 5(3)1) dan isolat C (SB 53 1(3)), dan yang berasal dari Microtermes inspiratus Kemner yaitu isolat D (SC 51 5(2)). Penelitian Sopandi (2007) yang mempelajari kombinasi dari isolat tersebut, memperoleh hasil yaitu isolat A dan isolat D merupakan kombinasi terbaik dalam mencerna pakan sumber serat. Hasil tersebut masih perlu diuji kemampuan dari isolat A dan isolat D yang dikombinasikan dengan isolat yang berasal dari rumen domba.
Perumusan Masalah
3 Serat sawit dan jerami padi adalah salah satu hasil samping yang dapat dimanfaatkan untuk pakan ternak. Kendala dari hasil samping tersebut adalah tingginya serat kasar yang terkandung di dalamnya, kandungan nutrien yang rendah dan nilai kecernaan kedua pakan tersebut juga rendah. Oleh sebab itu perlu adanya penelitian-penelitian untuk meningkatkan nilai guna dari limbah tersebut.
Dari penelitian-penelitian sebelumnya telah ditemukan bakteri simbion rayap yang dapat meningkatkan pemanfaatan pakan yang berasal dari hasil samping tanaman padi dan sawit melalui perbaikan proses pencernaan pakan sumber serat tersebut. Bakteri ini telah diisolasi dari rayap dan dipilih yang terbaik sehingga dapat hidup dalam kondisi rumen. Namun demikian isolat yang diperoleh masih perlu dikaji kembali kemampuannya dalam mencerna serat apabila isolat bakteri simbion rayap tersebut dikombinasikan dengan isolat bakteri rumen.
Tujuan
4 TINJAUAN PUSTAKA
Serat Kasar
Serat kasar adalah semua zat-zat organik yang tidak dapat larut dalam larutan H2SO4 0,3 N dan dalam larutan NaOH 1,5 N yang berturut-turut dimasak selama 30
menit. Serat kasar mengandung selulosa, lignin, dan sebagian dari pentosan-pentosan (Anggorodi, 1979). Menurut Sofyan et al. (2000), serat kasar adalah fraksi dari karbohidrat yang tidak larut dalam larutan basa maupun larutan asam encer setelah pendidihan masing-masing selama 30 menit. Sedangkan Tillman et al. (1989) menyatakan bahwa bagian-bagian dari karbohidrat adalah serat kasar dan bahan ekstrak tanpa nitrogen (BETN) yang telah dipisahkan dengan cara analisis kimia sederhana. Serat kasar terdiri atas selulosa, hemiselulosa dan lignin (Gambar 1). Selulosa dan hemiselulosa adalah komponen dalam dinding sel tanaman dan tidak dapat dicerna oleh hewan-hewan monogastrik (Tillman et al., 1989). Hal ini disebabkan monogastrik tidak menghasilkan enzim untuk mencerna bahan-bahan tersebut.
Tillman et al. (1989) menyatakan bahwa hewan tidak menghasilkan enzim untuk mencerna selulosa dan hemiselulosa, tetapi mikroorganisme dalam suatu saluran pencernaan menghasilkan selulase dengan hemiselulase yang dapat mencerna selulosa dan hemiselulosa, juga dapat mencerna pati dan karbohidrat yang larut dalam air menjadi asam-asam asetat, propionat dan butirat. Namun lignin tidak dapat dicerna baik oleh ruminansia maupun mikroorganisme. Lignin inilah yang menyebabkan turunnya produksi pada ternak. Apalagi bila lignin ini berikatan dengan selulosa dan hemiselulosa.
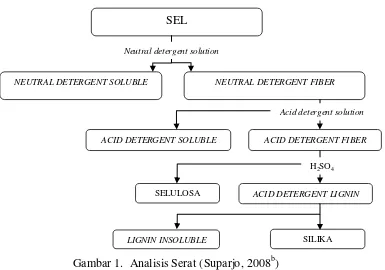
5 Gambar 1. Analisis Serat (Suparjo, 2008b)
Menurut Tillman et al. (1989), hasil akhir dari proses pencernaan golongan hemiselulosa dan selulosa berupa asam asetat, asam propionat dan asam butirat. Walaupun fungsi hemiselulosa dan selulosa dalam saluran pencernaan tidak spesifik, tetapi penting dalam meningkatkan gerak peristaltik pada pencernaan hewan golongan non ruminansia, juga merupakan sumber energi bagi mikroorganisme dalam lambung dan sebagai bahan pengisi lambung. Golongan lignin tidak memiliki hasil akhir dari proses pencernaan dan keberadaannya dapat menghambat proses pencernaan pada ternak.
Selulosa
Anggorodi (1979) mengatakan bahwa selulosa adalah suatu polisakarida yang mempunyai formula umum seperti pati (C6H10O5)n. Selulosa sebagian besar terdapat
dalam dinding sel dan bagian-bagian berkayu dari tumbuh-tumbuhan. Kapas hampir merupakan selulosa murni. Menurut Lehninger (1982), selulosa adalah senyawa seperti serabut, tidak larut dalam air dan ditemukan dalam dinding sel pelindung tumbuhan, terutama pada tangkai, batang, dahan dan semua bagian berkayu dari jaringan tumbuhan. Selulosa adalah bagian terbesar dari komponen lignoselulosa tanaman dan sebagai komponen utama penyusun dinding sel tanaman selain hemiselulosa dan lignin.
NEUTRAL DETERGENT SOLUBLE NEUTRAL DETERGENT FIBER
ACID DETERGENT SOLUBLE ACID DETERGENT FIBER
SELULOSA ACID DETERGENT LIGNIN
LIGNIN INSOLUBLE SILIKA
SEL
Acid detergent solution
H2SO4
6 Menurut Irawadi (1990), struktur selulosa sebagai polimer karbohidrat atau polisakarida tersusun dari anhidroglukopiranosa yang memiliki rumus C6H10O5.
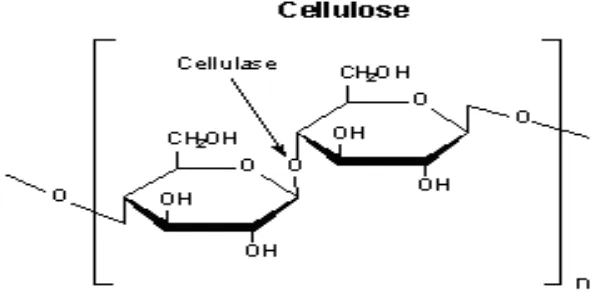
Selulosa membentuk dinding sel tanaman (Gambar 2). Pada tanaman, ikatan selulosa dibentuk dengan cara yang tersusun untuk memproduksi kumpulan padat (mikrofibril) yang disatukan bersamaan baik oleh ikatan molekul hidrogen inter maupun intra (McDonald et al., 2002).
Gambar 2. Struktur Selulosa (Carpita dan McCann, 2000)
Menurut Suparjo (2008a), ikatan β-1,4 glukosida pada serat selulosa dapat dipecah menjadi monomer glukosa dengan cara hidrolisis asam atau enzimatis. Kesempurnaan pemecahan selulosa pada saluran pencernaan ternak tergantung pada ketersediaan enzim pemecah selulosa yaitu selulase. Saluran pencernaan manusia dan ternak non ruminansia tidak mempunyai enzim yang mampu memecah ikatan β -1,4 glukosida sehingga tidak dapat memanfaatkan selulosa. Ternak ruminansia dengan bantuan enzim yang dihasilkan mikroba rumen dapat memanfaatkan selulosa sebagai sumber energi. Pencernaan selulosa dalam sel merupakan proses yang kompleks yang meliputi penempelan sel mikroba pada selulosa, hidrolisis selulosa dan fermentasi yang menghasilkan asam lemak terbang.
7 selulosa maupun hemiselulosa. Hubungan lignin-karbohidrat lebih berperan dalam mencegah hidrolisis polimer selulosa (Sa’id, 1994). Selulosa lebih tahan terhadap reagensia kimia daripada pati. Asam lemah dan alkali lemah mempunyai pengaruh kecil terhadap selulosa, tetapi zat tersebut dapat dihidrolisa oleh asam kuat menjadi glukosa (Anggorodi, 1979).
Hemiselulosa
Istilah hemiselulosa menunjukkan segolongan zat-zat termasuk didalamnya pentosa dan berbagai hexosan, yang kurang peka terhadap zat-zat kimia daripada selulosa. Golongan zat tersebut biasanya didefinisikan sebagai zat kabohidrat yang tidak larut dalam air mendidih, tetapi larut dalam alkali encer dan hancur dalam asam encer (Anggorodi, 1979). Sedangkan menurut McDonald et al. (1988), hemiselulosa adalah polisakarida pada dinding sel tanaman yang larut dalam alkali dan menyatu dengan selulosa. Hemiselulosa terdiri atas unit D-glukosa, D-galaktosa, D-manosa, D-xylosa dan L-arabinosa yang terbentuk bersamaan dalam kombinasi dan ikatan glikosidik yang bermacam-macam.
8 Gambar 3. Struktur Hemiselulosa (Carpita dan McCann, 2000)
Hidrolisis hemiselulosa akan menghasilkan tiga jenis monosakarida yaitu xylan, arabinosa dalam jumlah yang lebih banyak dan glukosa dalam jumlah yang lebih sedikit. Hidrolisa hemiselulosa dapat dilakukan dengan fermentasi oleh beberapa macam mikroorganisme yang mampu menggunakan gula pentosa sebagai subtratnya. Produk biokonversi selulosa dan hemiselulosa antara lain metana, asam organik, alkohol dan lain-lain (Gong, 1981 dalam Sa’id, 1994).
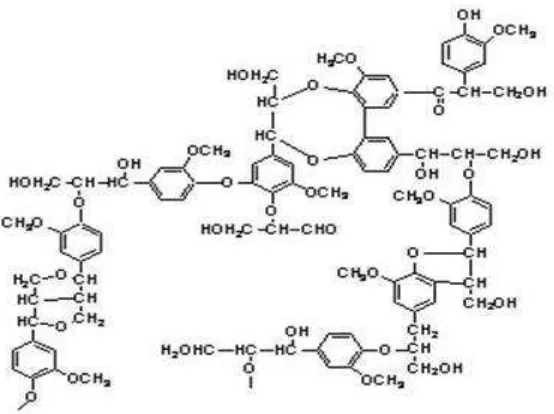
Lignin
9 tidak larut tersebut. Lignin bukan karbohidrat, tetapi sangat berhubungan erat dengan senyawa-senyawa karbohidrat (Departemen Fisika IPB, 2009).
Gambar 4. Struktur Lignin (Lora, 2006)
10 Pakan Sumber Serat
Jerami Padi
Menurut Wardhani et al. (1983), jerami padi merupakan hijauan yang berasal dari limbah pertanian yaitu bagian vegetatif tanaman padi yang telah diambil bulir padinya. Jerami padi dapat dimanfaatkan sebagai sumber energi bagi ternak ruminansia, namun menyebabkan penampilan produksinya kurang memuaskan akibat adanya lingnoselulosa yang tinggi (Laconi, 1992). Hal ini sejalan dengan yang dikatakan oleh Nitis (1979) yaitu penggunaan jerami padi sebagai pakan mempunyai keterbatasan dari nilai protein, daya cernanya rendah dan juga kurang palatabel. Dinding sel jerami padi sebagian besar tersusun dari lignin, selulosa, dan hemiselulosa. Faktor-faktor yang menghambat penggunaan jerami padi sebagai pakan ternak ruminansia yang terutama adalah rendahnya kandungan zat makanan, nilai kecernaanya maupun palatabilitasnya. Menurut Pramudyati (1983), jerami padi mempunyai daya cerna sekitar 35-40%. Hal ini disebabkan tanaman padi yang dipanen pada umur tua mempunyai kandungan dinding sel yang tinggi dan tingkat lignifikasi yang sempurna sehingga sulit dirombak oleh mikroba rumen (Wardhani et al., 1983).
Sutardi (1980) mengatakan, rendahnya kecernaan jerami padi disebabkan oleh tanaman padi yang dipanen pada umur tua mempunyai kandungan lignin yang tinggi sehingga sulit dirombak oleh mikroba rumen. Selain itu menurut Sutardi (1980), jerami padi sebagai pakan ternak masih terbatas sekali pemanfaatannya, karena hanya berperan sebagai bulk dan menggantikan tidak lebih dari 25% kebutuhan ternak akan rumput.
11 Jerami padi sangat mudah diperoleh dalam jumlah yang besar dan pemberiannya ke dalam ransum dalam jumlah tertentu akan mengurangi ongkos makanan serta dapat mempertahankan kondisi optimal bagi sapi-sapi pada periode kering dimana kebutuhan makanan relatif rendah (NRC, 1976). Doyle et al. (1986) mengatakan, perbandingan antara produksi padi dan jerami padi diperkirakan 1:1. Serat Sawit (Elaeis guineensis Jacq.)
Menurut Tomlinson (1961), taksonomi dari kelapa sawit digolongkan ke dalam filum : Angiospermae, sub filum : Monocotyledonae, kelas : Corolliferae, ordo : Palmae, famili : Arecaceae, sub famili : Cocoineae, genus : Elaeis dan spesies : Elaeis guineensis Jacq. Mansjur (1980) mengatakan bahwa Indonesia merupakan negara produsen kelapa sawit yang besar. Tanaman kelapa sawit (Elaeis guineensis Jacq.) bukan merupakan tanaman asli Indonesia, tanaman ini berasal dari daerah yang terletak antara Guinea dan Angola di Afrika Barat.
Sofyan et al. (2000) mengatakan bahwa terdapat dua tahap pengolahan kelapa sawit. Tahap pertama pengolahan kelapa sawit dari buah kelapa sawit yang menghasilkan minyak kelapa sawit dan lumpur kelapa sawit. Tahap kedua adalah pengolahan inti kelapa sawit dan bungkil kelapa sawit. Irawadi (1990) mengatakan bahwa hasil ikutan dan limbah pengolahan kelapa sawit yang dapat dijadikan sumber pakan ternak adalah bungkil inti sawit (palm kernel cake), serabut kelapa sawit (palm press fibre) dan lumpur minyak sawit (palm oil sludge).
12 Menurut Tomlinson (1961), serat kelapa sawit merupakan limbah pengolahan kelapa sawit yang dipisahkan dari buah setelah pengambilan minyak dan biji sawit pada proses pemerasan. Sedangkan menurut Aritonang (1986), serat sawit adalah hasil ikutan pengolahan kelapa sawit yang dipisahkan dari buah setelah pengambilan minyak dan biji dalam proses pemerasan.
Aritonang (1986) mengatakan bahwa serat sawit mengandung serat kasar yang tinggi. Komponen serat kasar yang tinggi (40,5-41,5 %) terdiri dari bagian-bagian berupa lignin, hemiselulosa dan abu. Serat sawit merupakan limbah yang mengandung ikatan lignoselulolitik, dimana selulosa tidak terdapat dalam bentuk bebas melainkan berikatan dengan lignin. Selain itu, Agustin (1991) menyatakan bahwa serat sawit kurang palatabel dan hanya mampu menggantikan rumput 23% dalam ransum ternak ruminansia.
Oleh karena itu, untuk mengatasi kelemahan-kelemahan penggunaan limbah kelapa sawit ini diperlukan pengolahan yang lebih efektif sehingga mampu memutuskan ikatan lignoselulosa dan secara tidak langsung membantu meningkatkan nutrien limbah tersebut (Irawadi, 1990). Menurut Akhirani (1998), usaha untuk meningkatkan kecernaan atau fermentabilitas pakan serat ini dapat dilakukan dengan memberi perlakuan pada pakan berkualitas rendah sebelum diberikan pada ternak, baik secara fisik (pemotongan, penggilingan, perendaman dan pemeletan), secara kimia (penambahan larutan basa atau amoniasi) dan juga secara biologi berupa fermentasi, penambahan enzim, menumbuhkan jamur dan bakteri.
Rumput Gajah (Pennisetum purpureum)
Menurut Reksohadiprojo (1985), taksonomi dari rumput gajah adalah divisio : Spermatophyta, sub divisio : Angiospermae, kelas : Monocotyledoneae, ordo : Glumifora, famili : Gramineae, sub famili : Panicodea, genus : Pennisetum dan spesies : Pennisetum purpureum Schumach. Secara alami rumput gajah hidup di daerah-daerah dengan curah hujan yang tinggi sampai 2500 mm tiap tahun atau tidak kurang dari 40 inchi setahun, kecuali pada pinggir sungai.
13 Selain itu juga Sofyan et al. (2000) menambahkan bahwa batang rumput gajah menjadi bertambah tebal dan keras apabila sudah menua, ditutupi perisai yang agak berbulu, rangkum bunga bertipe tandan dengan warna keemasan. Reksohadiprojo (1985) mengatakan bahwa rumput gajah memiliki ciri yang cukup spesifik. Rumput ini tumbuh sangat tegak dan berakar sangat dalam. Tinggi rumput gajah dapat mencapai lebih dari 4,5 meter. Hal yang serupa juga dikatakan oleh McIlroy (1976) yaitu rumput gajah merupakan tanaman tahunan, tingginya dapat mencapai 4,5 meter dan berumpun-rumpun.
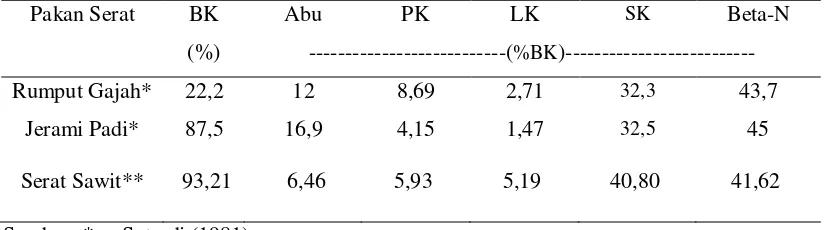
Menurut McIlroy (1976), kandungan nilai gizi jenis pakan hijauan dipengaruhi oleh perbandingan daun/batang, fase pertumbuhan pada waktu dipotong atau digembalai, kesuburan tanah dan pemupukan serta keadaan iklim. Sedangkan daya cerna hijauan makanan ternak pada ruminansia dan nilai gizi yang tinggi mungkin tergantung pada tercapainya imbangan yang tepat antara kandungan karbohidrat yang dapat larut dengan kandungan nitrogen. Menurut Sofyan et al. (2000), rumput gajah umumnya mengandung bahan kering (BK) yang rendah yaitu 12-18 %. Serat kasar berkisar dari 26-40,5 %, BETN sekitar 30,4-49,8 % dengan kandungan lemak kasar 1,0-3,6 %. Kandungan TDN berkisar antara 40-67 % dengan kecernaan BK sekitar 48-71 %. Tabel 1 adalah kandungan nutrien dari rumput gajah, jerami padi dan serat sawit (Sutardi, 1981; Agustin, 1991).
Tabel 1. Kandungan Nutrien Jenis Substrat
Pakan Serat BK Abu PK LK SK Beta-N
(%) ---(%BK)---
Rumput Gajah* 22,2 12 8,69 2,71 32,3 43,7
Jerami Padi* 87,5 16,9 4,15 1,47 32,5 45
Serat Sawit** 93,21 6,46 5,93 5,19 40,80 41,62
14 Rayap
Rayap merupakan serangga sosial dan terdapat pembagian pekerjaan diantara kastanya (Sigit et al., 2006). Hasan (1986) mengatakan bahwa rayap termasuk filum anthropoda, kelas insekta, dari ordo isopteran yang dalam perkembangan hidupnya mengalami metamorfosa bertahap.
Sigit et al. (2006) mengatakan rayap dalam hidupnya mengalami perkembangan metamorfose secara bertahap dari mulai telur yang dihasilkan oleh kasta reproduktif primer maupun sekunder. Nimfa yang berhasil menetas dari telur mengalami beberapa kali perubahan bentuk sampai menjadi salah satu kasta. Hasan (1986) mengatakan kelompok binatang ini pertumbuhannya melalui tiga tahap, yaitu tahap telur, tahap nimfa dan tahap dewasa. Setelah menetas dari telur, nimfa akan menjadi dewasa dengan melalui beberapa instar yaitu bentuk diantara dua masa perubahannya. Perubahan bentuk ini berlangsung secara bertahap, sehingga baik bentuk badan pada umumnya, cara hidup maupun makanan pokok antara nimfa dewasa adalah serupa. Di seluruh dunia ini terdapat lebih dari 2000 spesies rayap telah dikenal dan 120 diantaranya merupakan hama. Di Indonesia sendiri terdapat 200 spesies yang telah dikenal dan 20 diantaranya adalah hama perusak kayu dan hama hutan atau pertanian (Tarumingkeng, 2001).
Di alam, rayap sangat berguna mengubah kayu mati dan bahan organik lainnya yang mengandung selulosa untuk dijadikan humus (Sigit et al., 2006). Makanan utama rayap adalah kayu atau bahan yang terdiri atas selulosa. Berdasarkan bahan yang digunakan, maka dapat dikatakan rayap termasuk golongan perombak bahan mati yang sebenarnya sangat bermanfaat bagi kelangsungan kehidupan dalam ekosistem manusia (Tarumingkeng, 2001). Rayap tanah masih lebih mengutamakan kayu untuk dimakan sebagai sumber makanan (Sigit et al., 2006). Rayap mampu mencerna selulosa dan melumatkan serta menyerapnya sehingga sebagian besar ekskremen hanya tinggal lignin saja. Kerja protozoa (flagelata) dalam usus bagian belakang dari berbagai jenis rayap, terutama rayap tingkat rendah Rhinotermitidae, yang berperan sebagai simbion untuk melumatkan selulosa sehingga rayap mampu mencerna dan menyerap selulosa (Tarumingkeng, 2001).
15 banyak mengandung selulosa (kurang lebih 40-45% bahan kering). Berdasarkan tempat dan bahan yang didegradasi, maka mikroba selulolitik yang ada di dalam saluran pencernaan rayap, gajah, kerbau, dan sapi mempunyai karakteristik yang berbeda-beda, misalnya ada yang mempunyai aktivitas CMC-ase tinggi, tetapi aktivitas eksoglukanase maupun β-glukosidasenya rendah, demikian pula sebaliknya (Prabowo et al., 2007).
Rayap-rayap bersarang dalam tanah terutama dekat pada bahan organik yang mengandung selulosa seperti kayu, serasah dan humus. Jenis rayap tanah tersebut adalah rayap Termitidae yang paling umum menyerang bangunan dan objek-objek berjarak sampai 200 m dari sarangnya yaitu dengan bantuan enzim yang dikeluarkan dari mulutnya (Tarumingkeng, 2001).
Pencernaan Rayap
Ada beberapa perbedaan dan persamaan antara sistem pencernaan dari rayap dan ruminansia. Menurut Oldeson dan Breznak (1983), rayap memiliki kesamaan aktivitas dengan ruminansia dalam proses makanannya, diantaranya sama-sama dapat memanfaatkan sumber pakan berasal dari serat kasar berupa selulosa, terdapat mikroorganisme pendegradasi serat kasar dalam saluran pencernaannya, produk fermentasi yang dihasilkan dari proses pencernaan pakan dan kondisi dalam alat pencernaannya yaitu rumen dalam ruminansia dan usus belakang dalam rayap sama-sama dalam kondisi anaerob. Selain itu juga terdapat perbedaan aktivitas pencernaan makanan antara ruminansia dan rayap, yaitu dalam rumen lebih didominasi oleh bakteri, sedangkan dalam rayap lebih didominasi oleh protozoa. Pada rayap, produk fermentasinya tidak selengkap pada ruminansia.
Menurut Breznak (1982), masing-masing mikroorganisme mempunyai peran yang berbeda dalam mencerna selulosa tergantung kepada kelas rayap dimana mikroorganisme tersebut berdiam. Pada rayap kelas rendah, protozoa mempunyai peran lebih besar daripada bakteri dalam mencerna sumber serat. Namun hal sebaliknya terjadi pada rayap kelas tinggi dimana bakteri menjadi mikroba dominan dalam mencerna pakan.
16 lain pihak mikroorganisme menyumbang enzim selulase untuk membantu proses pencernaan serat kasar bagi rayap. Proses fermentasi selulosa di dalam saluran pencernaan rayap-rektum dilakukan oleh mikroba simbion yang terdiri atas protozoa atau flagelata, bakteri dan spirochaeta (Nakamura, 1989 dalam Sopandi, 2007).
17 Pencernaan Ruminansia
Pada umumnya kesanggupan hewan untuk mencerna selulosa atau serat kasar tergantung dari macamnya alat pencernaan yang dimiliki hewan tersebut dan tergantung pula dari mikroorganisme yang terdapat di dalam alat pencernaan. Dalam hal ini ruminansia mempunyai alat pencernaan yang paling sempurna untuk bekerjanya mikroorganisme terhadap serat kasar dan selulosa (Anggorodi, 1979).
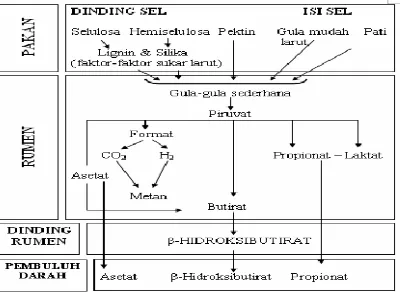
Untuk proses pencernaannya sendiri, pada ternak ruminansia terjadi secara mekanis (di dalam mulut), secara fermentatif (oleh enzim-enzim yang berasal dari mikroba rumen) dan secara hidrolisis (oleh enzim-enzim pencernaan hewan induk semang). Lokasi (posisi) proses pencernaan fermentatif bervariasi antar jenis ternak. Posisi tersebut akan menentukan karakteristik pakan yang sesuai untuk jenis ternak bersangkutan (Sutardi, 1980). Ruminansia mempunyai mikroorganisme di dalam retikulorumen yang mensekresikan enzim-enzim sehingga dapat mencerna makanan yang masuk. Bagian terbesar karbohidrat terdiri dari: yang mudah larut (gula dan pati) dan yang sukar larut (selulosa dan hemiselulosa, misal hijauan dan limbah serat). Keduanya ini difermentasikan oleh mikroba rumen membentuk VFA di dalam
18 rumen dan retikulum. Pemecahan karbohidrat menjadi VFA terjadi di rumen yang terdiri dari 2 tahap: 1). Hidrolisis ekstraseluler dari karbohidrat kompleks (selulosa, hemiselulosa, pektin) menjadi oligosakarida rantai pendek terutama disakarida (selobiosa, maltosa, pentosa) dan gula-gula sederhana. 2). Pemecahan oligosakarida dan gula-gula sederhana menjadi VFA oleh aktifitas enzim intraseluler (Departemen Fisika IPB, 2009).
Pada sistem pencernaan ruminansia juga dikenal suatu proses yang disebut memamahbiak (ruminasi). Pakan berserat (hijauan) yang dimakan ditahan untuk sementara di dalam rumen. Pada masa hewan tersebut beristirahat, pakan dalam rumen lalu dikembalikan ke mulut (proses regurgitasi) untuk dikunyah kembali (proses remastikasi), kemudian pakan ditelan kembali (proses redeglutasi). Selanjutnya pakan tersebut dicerna lagi oleh enzim-enzim mikroba rumen (microbial attack). Kontraksi retikulum rumen yang terkoordinasi dalam rangkaian proses tersebut bermanfaat pula untuk memadukan digesta, inokulasi digesta dan penyerapan nutrien. Selain itu kontraksi retikulo rumen juga bermanfaat untuk pergerakan digesta meninggalkan retikulo rumen melalui retikulo-omasal orifice (Erwanto, 1995). Gambar 5 adalah proses sederhana dalam pencernaan kabohidrat dalam ternak ruminansia.
Mikroorganisme dalam rumen merombak selulosa untuk membentuk asam-asam lemak terbang. Mikroorganisme tersebut mencerna pula pati, gula, lemak, protein dan nitrogen bukan protein untuk membentuk protein mikrobial dan vitamin B (Anggorodi, 1979). Menurut Arora (1989), di dalam rumen spesies-spesies bakteri dan protozoa yang berbeda saling berinterakasi melalui hubungan simbiosa dan menghasilkan produk-produk yang khas seperti selulosa, hemiselulosa dan pati melalui pencernaan polimer tumbuhan. Bakteri-bakteri tertentu yang bertanggung jawab dalam proses fermentasi pregastrik membentuk asetat, propionat, butirat, CO2
dan H2. Spesies bakteri metanogenik menggunakan CO2, H2 dan format untuk
membentuk metan.
19 mikroorganisme terhadap protein atau ikatan-ikatan lainnya yang mengandung nitrogen. Asam-asam tersebut masuk ke dalam abomasum untuk mengalami pencernaan dan akhirnya masuk ke dalam usus untuk kemudian diserap masuk peredaran darah.
Arora (1989) menyatakan bahwa proses fermentasi pakan di dalam rumen menghasilkan VFA dan NH3, serta gas-gas (CO2, H2 dan CH4) yang dikeluarkan dari
rumen melalui proses eruktasi. Menurut Sutardi (1988), produk fermentasi di rumen akan diserap melalui dinding rumen, sedangkan produk pencernaan hidrolitik diserap melalui dinding usus halus. Zat yang diserap selanjutnya akan masuk ke dalam sistem peredaran darah.
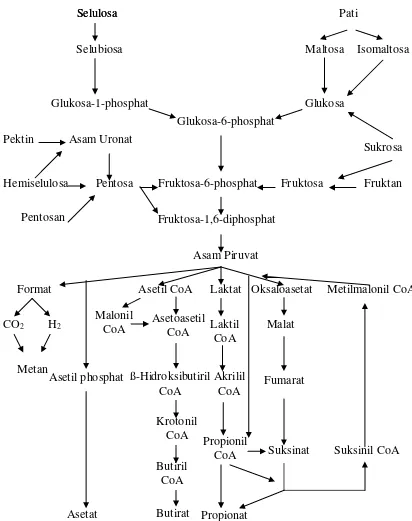
Volatile Fatty Acid (VFA)
Menurut Parakkasi (1999), proses fermentasi karbohidrat dalam rumino-retikulum akan menghasilkan asam lemak atsiri (asam lemak terbang atau asam lemak berantai pendek = Volatile Fatty Acid atau VFA) terutama asetat, propionat, n-butirat, laktat (dengan ransum yang kaya akan biji-bijian), dan format (dengan hay yang tua). McDonald et al. (2002) menyatakan bahwa proses fermentasi karbohidrat dalam rumen terjadi melalui dua tahap, yaitu pemecahan karbohidrat kompleks menjadi gula sederhana dan fermentasi gula sederhana menjadi asam asetat, asam propionat, asam butirat, CO2 dan CH4. Gambar 6 merupakan alur pencernaan
karbohidrat menjadi VFA pada ternak ruminansia.
Asam-asam asetat, propionat dan butirat, CO2 dan gas metan adalah hasil
20 Gambar 6. Proses Metabolisme Karbohidrat di dalam Rumen Ternak Ruminansia
(McDonald et al., 2002)
Sebagian besar VFA diserap langsung melalui dinding rumen; hanya sedikit asetat, beberapa propionat dan sebagian besar butirat termetabolisme dalam dinding rumen. VFA yang terbentuk merupakan sumber energi utama yang merupakan salah satu ciri khas dari ruminan (Parakkasi, 1999). VFA kemudian diserap melalui
Asetil CoA Laktat Oksaloasetat Metilmalonil CoA
21 dinding rumen melalui penonjolan-penonjolan yang menyerupai jari yang disebut vili. Lebih lanjut dikemukakan bahwa sekitar 75 % dari total VFA yang diproduksi akan diserap langsung di retikulo-rumen masuk ke darah, sekitar 20 % diserap di abomasum dan omasum, dan sisanya sekitar 5 % diserap di usus halus (McDonald et al., 2002). VFA yang terserap selain dipakai sebagai sumber energi, juga dipakai
sebagai bahan pembentuk glikogen di hati, lemak, karbohidrat dan hasil-hasil yang dibutuhkan ternak (Anggorodi, 1979).
Amonia (NH3)
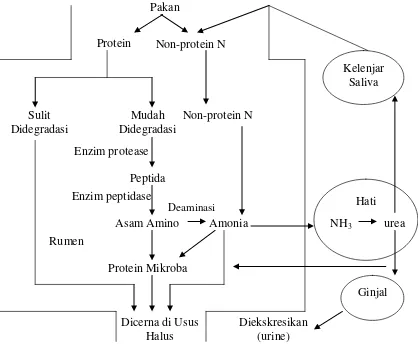
Seluruh protein yang berasal dari makanan pertama kali dihidrolisa oleh mikroba rumen. Tingkat hidrolisa protein tergantung dari daya larutnya yang berkaitan dengan kenaikan amonia (Arora, 1989). Sebagaimana yang tercantum pada Gambar 7, McDonald et al. (2002) mengatakan bahwa protein bahan makanan yang masuk ke dalam rumen pada awalnya akan mengalami proteolisis oleh enzim-enzim protease menjadi peptida, lalu dihidrolisa menjadi asam amino yang kemudian secara cepat dideaminasi menjadi amonia. Keduanya akan digunakan oleh mikroba rumen dalam pembentukan protein mikroba. Menurut Arora (1989), hidrolisa protein menjadi asam amino diikuti oleh proses deaminasi untuk membebaskan amonia. Kecepatan deaminasi biasanya lebih lambat daripada proteolisis.
Menurut Arora (1989), amonia yang dibebaskan dalam rumen sebagian dimanfaatkan oleh mikroba untuk mensintesis protein mikroba. Bahkan amonia yang dibebaskan dari urea atau garam-garam amonium lain dapat dipergunakan untuk sintesa protein mikroba.
22
Gambar 7. Proses Metabolisme Protein di dalam Rumen Ternak Ruminansia (McDonald et al., 2002)
Pada umumnya sebagian protein ransum tidak akan terdegradasi dalam rumen dan akan digunakan di dalam segmen saluran pencernaan pascarumen. Bila sebagian protein ransum tersebut disubtitusi dengan NPN, maka protein bebas degradasi rumen tersebut tentu akan berkurang pula dengan segala pengaruhnya terhadap penggunaan NPN itu sendiri (Parakkasi, 1999).
23 METODE
Waktu dan Tempat
Penelitian ini telah dilaksanakan dari bulan September 2008 sampai Februari 2009, bertempat di Laboratorium Biokimia, Fisiologi dan Mikrobiologi Nutrisi, dan Laboratorium Nutrisi Ternak Perah, Departemen Ilmu Nutrisi dan Teknologi Pakan, Fakultas Peternakan, Institut Pertanian Bogor.
Materi Bahan
Penelitian ini menggunakan bahan yaitu pakan sumber serat, bahan kimia dan sumber inokulum yang berasal dari bakteri simbion rayap dan bakteri rumen serta cairan rumen sapi dari Rumah Potong Hewan (RPH) sebanyak 3 ekor yang telah diautoclave. Bahan pakan yang digunakan antara lain rumput gajah, jerami padi dan serat sawit yang telah digiling halus. Sumber inokulum bakteri simbion rayap yaitu dua isolat bakteri simbion rayap terbaik yang berasal dari rayap yang berbeda yaitu isolat A {SB 53 5(3)1}, berasal dari rayap Coptotermes curvignathus Holmgren dan isolat D {SC 51 5(2)} yang merupakan bakteri simbion rayap terbaik yang berasal dari rayap Microtermes inspiratus Kemner (Sopandi, 2007). Sumber inokulum bakteri rumen adalah isolat mikroba yang berasal dari cairan rumen domba yaitu SE 511, SE 512 dan SE 513.
Bahan kimia yang digunakan adalah larutan McDougall, gas CO2, larutan
pepsin/HCl 0,2 %, larutan asam borat, larutan HgCl2, larutan Na2CO3 jenuh, larutan
H2SO4 0,005 N, vaselin, larutan H2SO4 15 %, larutan NaOH 0,5 N, larutan HCl 0,5
N dan aquades.
Alat
24 Metode
Perlakuan
Perlakuan dalam penelitian ini yaitu kombinasi berbagai isolat bakteri simbion rayap dan isolat bakteri rumen domba, yang terdiri dari 6 perlakuan dan 3 ulangan, yaitu:
1. A+ SE 511 2. A + SE 512 3. A + SE 513 4. D + SE 511 5. D + SE 512 6. D + SE 513
Populasi bakteri yang diinokulumkan dari setiap isolat yaitu isolat A : 0,68 x 1012 CFU/ml, isolat D : 0,11 x 1012 CFU/ml, SE 511 : 6,53 x 1012 CFU/ml, SE 512 : 4,34 x 1012 CFU/ml, dan SE513 : 1,49 x 1012 CFU/ml. Kombinasi isolat tersebut kemudian diperlakukan pada tiga (3) jenis bahan pakan sumber serat yaitu jerami padi, serat sawit dan rumput gajah.
Peubah yang Diamati
Peubah yang diamati dalam penelitian ini yaitu konsentrasi NH3, produksi
VFA total, degradasi bahan kering (DBK), degradasi bahan organik (DBO), koefisien cerna bahan kering (KCBK) dan koefisien cerna bahan organik (KCBO).
Rancangan Percobaan
Rancangan percobaan yang digunakan dalam penelitian ini adalah Rancangan Acak Kelompok (RAK) dengan pola faktorial 6 x 3. Faktor A adalah kombinasi isolat bakteri simbion rayap dan isolat bakteri rumen yaitu : A + SE 511, A + SE 512, A + SE 513, D + SE 511, D + SE 512, dan D + SE 513. Isolat A adalah SB 53 5(3)1 dan isolat D adalah SC 51 5(2). Faktor B adalah bahan pakan sumber serat yaitu rumput gajah, jerami padi dan serat sawit.
Adapun model matematikanya adalah :
25 Keterangan:
Yijk = efek blok ke-k, kombinasi isolat bakteri rayap dan bakteri rumen ke-i
dan pakan sumber serat ke-j µ = rataan umum
σk = efek utama kelompok
αi = efek utama kombinasi dari kedua isolat bakteri ke-i
βj = efek utama bahan pakan sumber serat ke-j
αiβj = efek interaksi kombinasi dari kedua isolat bakteri ke-i dan bahan pakan
sumber serat ke-j
εijk = error blok ke-k, kombinasi isolat bakteri rayap dan bakteri rumen ke-i
dengan pakan sumber serat ke-j i = 1, ..., 6
j = 1, 2, 3
Analisis data menggunakan Sidik Ragam (ANOVA); apabila hasil uji tersebut berbeda nyata maka dilanjutkan dengan uji ortogonal kontras untuk memperoleh perlakuan terbaik (Steel dan Torrie, 1993).
Prosedur Pelaksanaan Peremajaan Bakteri
Media basal (BHI) sebanyak 10 ml dimasukkan ke dalam tabung reaksi dengan dialiri gas, kemudian tabung ditutup dengan prop karet dan diselotip agar keadaan media tumbuh bakteri tetap dalam keadaan anaerob. Selanjutnya sumber inokulum bakteri dimasukkan ke dalam tabung tersebut. Tabung lalu dimasukkan ke dalam inkubator (suhu 39 0C) selama 24 jam. Mikroba ini selanjutnya digunakan sebagai inokulum pada uji fermentasi dan kecernaan in vitro.
Analisis NH3 dan VFA Pencernaan Fermentatif
26 McDougall diturunkan pHnya hingga 6,5-6,8, kemudian sebanyak 12 ml dimasukkan ke dalam tabung fermentor tersebut disertai dengan penambahan gas CO2. Cairan
rumen sapi yang telah diautoclave dimasukkan ke dalam tabung tersebut sebanyak 6 ml dan diikuti dengan memasukkan sumber inokulum sebanyak 2 ml. Larutan dialiri gas CO2 selama 30 detik, kemudian ditutup dengan prop karet yang berventilasi.
Setelah itu, tabung tersebut dimasukkan ke dalam penangas bergoyang (shaker water bath) dengan suhu 39 0C dan difermentasikan selama 6 jam. Setelah 6 jam, tutup karet dibuka kemudian ditambahkan 0,2 ml larutan HgCl2 jenuh untuk mematikan
mikroba sehingga tidak ada aktifitas mikroba lagi dalam tabung tersebut. Cairan dalam tabung fermentor diambil dengan menggunakan pipet dan dipindahkan ke dalam tabung sentrifuse. Tabung sentrifuse yang berisi cairan tersebut disentrifuse dengan kecepatan 3000 rpm selama 15 menit dan supernatannya ditampung dalam tabung film untuk dianalisis konsentrasi VFA dan NH3.
Pengukuran Konsentrasi NH3
Supernatan dari pencernaan fermentatif diambil sebanyak 1 ml dan ditempatkan pada salah satu ujung alur cawan, sedangkan Na2CO3 jenuh ditempatkan
pada ujung alur cawan sebelahnya (kedua bahan tersebut tidak boleh bercampur sebelum tutup cawan ditutup rapat). Larutan asam borat berindikator sebanyak 1 ml ditempatkan dalam cawan kecil yang terletak di tengah cawan Conway. Selanjutnya cawan Conway yang telah diolesi vaselin pada bibir dan tutupnya ditutup rapat agar tidak ada NH3 yang keluar. Larutan Na2CO3 jenuh dicampurkan dengan supernatan
sampel hingga merata dengan cara menggoyang-goyangkan dan memiringkannya. Sebelum dititrasi cawan Conway dibiarkan selama 24 jam pada suhu kamar. Setelah 24 jam tutup cawan dibuka dan asam borat dititrasi dengan H2SO4 0,005 N sampai
warnanya berubah dari biru menjadi kemerah-merahan. Saat titrasi, asam borat tidak boleh terkontaminasi oleh supernatan karena warna asam borat tidak akan berubah menjadi merah. Berikut ini adalah rumus pengukuran konsentrasi NH3 :
27 Pengukuran Konsentrasi VFA (Steam Destilation)
Supernatan yang sama dengan analisis NH3 diambil 5 ml dan dimasukkan ke
dalam tabung destilasi. Sebanyak 1 ml H2SO4 15 % ditambahkan ke dalam
supernatant lalu segera ditutup dengan tutup karet yang mempunyai lubang dan dapat dihubungkan dengan labu pendingin. Setelah itu, tabung destilasi dimasukkan ke dalam labu penyulingan yang berisi air mendidih (dipanaskan terus selama destilasi). Uap air panas akan mendorong asam lemak terbang dan terkondensasi dalam proses pendinginan. Selanjutnya cairan yang terbentuk ditampung dalam labu Erlenmeyer yang berisi 5 ml NaOH 0,5 N, sampai mencapai 300 ml. Indikator phenolphtalein ditambahkan sebanyak 2-3 tetes dan dititrasi dengan HCl 0,5 N, sampai warna titrat berubah dari merah jambu menjadi tidak berwarna. Produksi VFA total dapat dihitung dengan rumus sebagai berikut :
VFA(mM) = (a – b) ml x N-HCl x (1000/5) bobot sampel x BK sampel
Keterangan:
a = volume titran blanko b = volume titran sampel
Degradasi Bahan Kering (DBK) dan Degradasi Bahan Organik (DBO)
Residu yang diperoleh dari proses fermentasi selama 6 jam dikeringkan di dalam oven 1050C selama 24 jam, kemudian ditimbang setelah didinginkan di dalam eksikator. Penimbangan dilakukan untuk mengetahui bobot kering residu yang digunakan untuk menentukan degradasi bahan kering. Setelah penimbangan, sampel residu dimasukan ke dalam tanur 6000C selama 6 jam. Sampel didinginkan di dalam eksikator. Setelah dingin, sampel residu ditimbang untuk mengetahui bobot abu residu, sedangkan bobot bahan organik dihitung dengan mengurangi bobot kering residu dengan bobot abu residu.
28 % DBK = BoK sampel –(BoK residu-BoK Blanko) x 100%
BoK sampel
% DBO = BoO sampel –(BoO residu-BoO Blanko) x 100% BoO sampel
Keterangan: BoK: Bobot kering BoO: Bobot Organik
Pencernaan Hidrolisis Aerob
Produk fermentasi yang telah dipisahkan supernatannya pada percobaan fermentabilitas (24 jam), ditambah larutan enzim pepsin 0,2 % (20 ml), kemudian dilakukan proses inkubasi lanjutan secara aerob sampai 24 jam dalam penangas bergoyang (shaker water bath). Setelah 24 jam campuran tersebut disaring dengan kertas saring Whatman No. 41 menggunakan pompa vakum untuk mendapatkan residunya.
Koefisien Cerna Bahan Kering (KCBK)
Residu yang telah dipisahkan dari supernatannya, kemudian dimasukkan ke dalam cawan porselin untuk diuapkan airnya di dalam oven 105 0C selama 24 jam. Dari penguapan dengan oven 105 0C tersebut didapatkan bahan kering. Bobot kering sampel dalam cawan porselin kemudian ditimbang. Selanjutnya untuk menentukan besaran koefisien kecernaan bahan kering dapat dihitung menggunakan cara berikut ini.
% KCBK = BoK asal - (BoK residu – BoK blanko) x 100% BoK asal
Keterangan: BoK: Bobot Kering BoO: Bobot Organik
29 Koefisien Cerna Bahan Organik (KCBO)
Besaran koefisien cerna bahan organik pakan diperoleh dari bobot abu residu. Untuk mengetahui bobot abu residu, sampel dalam cawan porselin yang telah dikeringkan pada analisa KCBK, kemudian dimasukkan ke dalam tanur suhu 600 0C selama 6 jam. Setelah 6 jam, cawan porselin tersebut diangkat dan ditimbang bobot abunya setelah terlebih dahulu didinginkan di dalam eksikator. Bobot bahan organik diperoleh dengan mengurangi bobot kering dengan bobot abu residu. Besaran koefisien cerna bahan organik dapat dihitung dengan cara berikut ini:
% KCBO = BoO asal - (BoO residu – BoO blanko) x 100% BoO asal
Keterangan: BoK: Bobot Kering BoO: Bobot Organik
30 HASIL DAN PEMBAHASAN
Konsentrasi VFA Total
Pada ruminansia, sumber energi utamanya berasal dari fermentasi serat kasar yaitu berupa asam lemak terbang (VFA). Menurut Arora (1989), VFA dalam rumen diproduksi dari hasil perombakan karbohidrat dan digunakan oleh mikroba rumen sebagai sumber energi bagi inang maupun kerangka karbon untuk perkembangbiakan mikroba. Proses fermentasi karbohidrat dalam rumen terjadi melalui dua tahap, yaitu pemecahan karbohidrat kompleks menjadi gula sederhana dan fermentasi gula sederhana menjadi asam asetat, asam propionat, asam butirat, CO2 dan CH4
(McDonald et al., 2002).
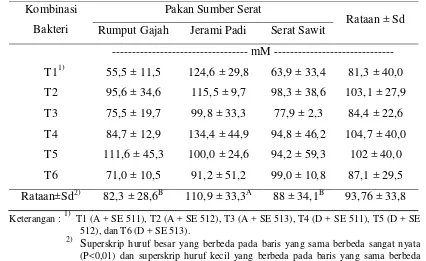
Tabel 2. Konsentrasi VFA Total oleh Isolat Bakteri Rayap dan Rumen
Keterangan : 1) T1 (A + SE 511), T2 (A + SE 512), T3 (A + SE 513), T4 (D + SE 511), T5 (D + SE pakan sumber serat (P<0,05), sedangkan untuk kombinasi isolat bakteri dan interaksi antara pakan sumber serat dengan kombinasi isolat bakteri tidak menyebabkan efek yang nyata terhadap konsentrasi VFA total. Hasil uji ortogonal kontras memperlihatkan bahwa penggunaan jerami padi (110,9±33,3 mM) mempunyai konsentrasi VFA total tertinggi (P<0,01), sedangkan penggunaan serat sawit
Kombinasi Bakteri
Pakan Sumber Serat
Rataan ± Sd Rumput Gajah Jerami Padi Serat Sawit
31 (88±34,1 mM) dan rumput gajah (82,3±28,6 mM) menunjukkan hasil yang tidak berbeda nyata pada waktu inkubasi 6 jam.
Hasil penelitian ini menunjukkan produksi VFA dari jerami padi paling tinggi dibandingkan dengan rumput gajah dan serat sawit. Hasil ini sejalan dengan penelitian yang dilakukan oleh Sulistiani (2005) yang menggunakan isolat murni dari bakteri rayap yang memperlihatkan produksi VFA total untuk jerami padi 134,39±8,04 mM, serat sawit 114,89±6,29 mM dan rumput gajah 105,66±3,21 mM. Penelitian Sopandi (2007) yang mengkombinasikan isolat bakteri rayap menghasilkan produksi VFA yang lebih tinggi dibandingkan pada penelitian ini yaitu rumput gajah 127,9±35,44 mM, jerami padi 121,07±40,83 mM dan serat sawit 108,09±33,99 mM. Hal ini menunjukkan bahwa konsentrasi produksi VFA jerami padi pada 6 jam (waktu inkubasi) dari kultur bakteri tunggal lebih tinggi daripada rumput gajah dan serat sawit, tetapi tidak terjadi perbedaan yang nyata dari ketiga pakan berserat tersebut pada saat isolat bakteri rayap dikulturkan secara bersama dan isolat bakteri rayap lebih baik aktifitasnya bila dikulturkan dengan isolat bakteri rayap.
32 Waktu inkubasi selama 6 jam memberikan kesempatan kepada bakteri untuk memaksimalkan mencerna serat kasar terutama pada jerami padi. Produksi VFA yang rendah pada serat sawit menunjukkan bahwa serat sawit membutuhkan waktu inkubasi yang lebih lama. Hal ini disebabkan kandungan lignin serat sawit lebih tinggi daripada jerami padi. Menurut Sopandi (2007), kandungan lignin dari jerami padi sebesar 13,16% dan serat sawit sebesar 25,90%. Secara keseluruhan bahan pakan limbah membutuhkan waktu yang lebih lama untuk mencapai produksi gas yang mendekati rumput gajah (Toha, 2006).
Kombinasi isolat bakteri yang hasilnya tidak berbeda nyata terhadap konsentrasi VFA total menunjukkan bahwa kombinasi dari isolat bakteri rayap dan isolat bakteri rumen sama-sama dapat mencerna serat kasar dari ketiga pakan sumber serat dengan baik. Hal ini dapat dilihat dari konsentrasi VFA total yang dihasilkan berkisar antara 55,5-134,4 mM. Menurut Sutardi (1980), kadar VFA yang dibutuhkan untuk menunjang pertumbuhan optimal rumen adalah 80-160 mM.
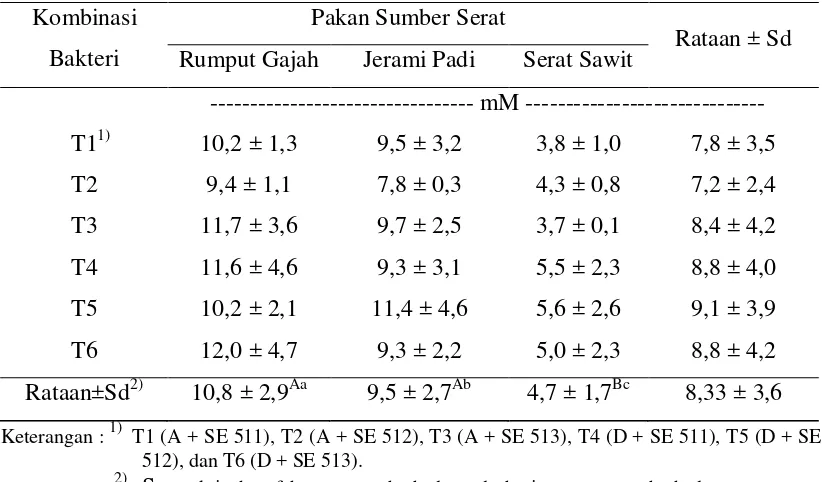
Konsentrasi NH3
Amonia dalam rumen diproduksi dari hasil perombakan asam amino yang merupakan hasil perombakan protein secara fermentatif (McDonald et al., 1988). Protein bahan makanan yang masuk ke dalam rumen pada awalnya akan mengalami proteolisis oleh enzim-enzim protease menjadi peptida, lalu dihidrolisa menjadi asam amino yang kemudian secara cepat dideaminasi menjadi amonia. Keduanya akan digunakan oleh mikroba rumen dalam pembentukan protein mikroba (McDonald et al., 2002). Secara kuantitatif kadar amonia dalam cairan rumen penting karena pemakaian amonia oleh mikroba terus meningkat (Arora, 1989).
Pada penelitian ini konsentrasi NH3 (Tabel 3) tidak dipengaruhi oleh perlakuan
dari kombinasi isolat bakteri dan interaksi antara isolat bakteri dan bahan pakan sumber serat, sedangkan untuk bahan pakan sumber serat berpengaruh sangat nyata lebih tinggi (P<0,01) terhadap konsentrasi NH3. Hasil uji ortogonal kontras
memperlihatkan konsentrasi NH3 yang sangat nyata (P<0,01) pada rumput gajah
(10,8±2,9 mM) dan jerami padi (9,5±2,7 mM) dibandingkan dengan serat sawit (4,7±1,7 mM), sedangkan konsentrasi NH3 rumput gajah berbeda (P<0,05) dengan
33 Penelitian yang dilakukan oleh Sulistiani (2005) dengan menggunakan isolat murni dari masing-masing bakteri, rumput gajah memproduksi NH3 sebesar
8,67±0,30 mM, jerami padi 8,05±0,33 mM dan serat sawit 8,26±0,52 mM. Hal ini menunjukan bahwa produksi NH3 yang lebih optimal adalah dengan mengkulturkan
bakteri rayap dengan bakteri rumen. Hasil ini juga didukung oleh Sopandi (2007) yang mengkulturkan bakteri rayap dengan bakteri rayap yang memperoleh hasil yang lebih kecil yaitu rumput gajah 6,61±2,85 mM, jerami padi 8,30±3,66 mM dan serat sawit 3,95±0,95 mM.
Tabel 3. Konsentrasi NH3 oleh Isolat Bakteri Rayap dan Rumen
Keterangan : 1) T1 (A + SE 511), T2 (A + SE 512), T3 (A + SE 513), T4 (D + SE 511), T5 (D + SE
Rumput gajah mempunyai nilai yang paling tinggi karena kandungan proteinnya paling besar. Kandungan protein rumput gajah, jerami padi dan serat sawit berturut-turut 8,69% ; 4,15% dan 5,93% (Sutardi, 1981; Agustin, 1991). Haryanto dan Djayanegara (1993) menyatakan bahwa konsentrasi NH3 dalam rumen
dipengaruhi oleh beberapa faktor yaitu jenis makanan, kelarutan protein, tingkat degradasi protein dan kadar protein dalam ransum. Selain itu, kandungan lignin yang tinggi pada jerami padi dan serat sawit menyebabkan bakteri sulit untuk mendegradasi zat-zat makanan yang terdapat pada isi sel. Menurut Sopandi (2007), kandungan lignin rumput gajah, jerami padi dan serat sawit berturut-turut 10,51% ;
Kombinasi Bakteri
Pakan Sumber Serat
Rataan ± Sd Rumput Gajah Jerami Padi Serat Sawit