i
DAMPAK DOMESTIKASI
ZINGIBERACEAE
DI TAMAN
NASIONAL GUNUNG HALIMUN SALAK TERHADAP
KERAGAMAN CENDAWAN ENDOFITNYA
IVAN PERMANA PUTRA
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA*
Dengan ini saya menyatakan bahwa tesis berjudul Dampak Domestikasi
Zingiberaceae
di Taman Nasional Gunung Halimun Salak terhadap Keragaman
Cendawan Endofitnya adalah benar karya saya dengan arahan dari komisi
pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana
pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan
maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan
dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut
Pertanian Bogor.
Bogor, Juli 2014
Ivan Permana Putra
RINGKASAN
IVAN PERMANA PUTRA. Dampak Domestikasi Zingiberaceae di Taman Nasional Gunung Halimun Salak Terhadap Keragaman Cendawan Endofitnya. Dibimbing oleh GAYUH RAHAYU dan IMAN HIDAYAT.
Taman Nasional Gunung Halimun Salak (TNGHS) merupakan salah satu hutan tropis yang tersisa di Pulau Jawa. TNGHS memiliki keragaman
Zingiberaceae liar yang relatif tinggi. Dua spesies diantaranya yakni Alpinia malaccensis (AM) dan Horstendia conica (HC), telah didomestikasi sebagai tanaman pekarangan di sekitar area hutan sebagai tanaman obat. Dampak domestikasi tanaman terhadap cendawan patogen sering dilaporkan, namun dampak domestikasi tanaman terhadap cendawan endofit jarang diteliti. Oleh sebab itu penelitian ini bertujuan untuk menganalisis dampak dari domestikasi
Zingiberaceae di TNGHS terhadap keragaman cendawan endofit yang berasosiasi dengan dua spesies Zingiberaceae tersebut.
Keragaman cendawan endofit asal Zingiberaceae liar dan domestikasi dianalisis berdasarkan cendawan yang dapat dikulturkan. Masing-masing satu individu tanaman diambil dari rumpun AM dan HC liar serta rumpun AM dan HC domestikasi terpilih. Setiap tanaman terdiri atas akar, rimpang, batang dan daun. Semua bagian tanaman disterilisasi permukaan. Potongan-potongan setiap bagian tanaman ditanam pada media MEA (Malt Extract Agar) half strength dan diinkubasi pada suhu kamar. Cendawan yang tumbuh dari bagian-bagian tanaman tersebut diisolasi dan dimurnikan. Identifikasi hingga ke tingkat spesies dilakukan dengan pendekatan morfologi serta molekuler berdasarkan sekuen ITS rDNAnya.
Secara morfologi, koleksi biakan cendawan endofit ini dapat dibagi ke dalam 17 morfotipe. Setelah diidentifikasi secara molekuler morfotipe ini terdiri atas 19 spesies cendawan yaitu Arthrinium malaysianum, Aspergillus flavipes, As. sydowii, Chaetomium globosum, Cladosporium sp., Cl. oxysporum, Colletotrichum aff. boninense-complex, Co. aff. cliviae-complex, Co. aff.
gloeosporioides-complex, Diaporthe sp., D. anacardii, D. gardenia, Exophiala
sp., E. lecanii-corni, Guignardia mangiferae, Ochroconis gallopava, Penicillium citrinum, Pyricularia costina, dan Sydowiellaceae tidak bersporulasi. Semua cendawan endofit pada Zingiberaceae tergolong Ascomycota, Pezizomycotina
yang tersebar ke dalam 3 kelas yaitu: Dothideomycetes, Eurotiomycetes, dan
Sordariomycetes.
iii
Domestikasi Zingiberaceae mendorong hilangnya spesies cendawan endofit dari inang liarnya atau sebaliknya mendorong munculnya spesies cendawan endofit yang baru ditemukan pada inang yang dibudidayakan.
Aspergillus flavipes, As. sydowii, Cl. oxysporum, Diaporthe sp. dan Py. costina
ditemukan pada inang liarnya dan tidak ditemukan lagi pada inang domestikasi.
Arthrinium malaysianum, Ch. globosum, Co. aff. cliviae-complex, D. gardenia, dan Sydowiellaceae tak bersporulasi hanya ditemukan pada inang domestikasi. Sementara itu, beberapa spesies endofit yaitu Co. aff. boninense-complex,
Cladosporium sp., Co. aff. gloeosporioides-complex, dan E. lecanii-corni,
tampaknya tidak dipengaruhi oleh domestikasi.
Penelitian ini mengindikasikan adanya dampak domestikasi Zingiberaceae
terhadap keragaman dan keberadaan cendawan endofitnya. Keterbatasan dalam jumlah dan frekuensi pengambilan contoh serta pemilihan metode isolasi menyebabkan hasil penelitian ini hanya dapat menjadi informasi awal mengenai dampak domestikasi terhadap keragaman endofit terutama pada Zingibercaeae.
Oleh sebab itu penelitian ini masih perlu diperkuat dengan penambahan jumlah sampel serta analisis komunitas dengan pendekatan metagenomik agar hasil yang diperoleh lebih akurat.
Kata kunci: Domestikasi, cendawan endofit , keragaman, Zingiberaceae
SUMMARY
IVAN PERMANA PUTRA. The Impact of Zingiberaceae Domestication at Mount Halimun Salak National Park to Their Fungal Endophytes Diversity. Supervised by GAYUH RAHAYU and IMAN HIDAYAT.
Mount Halimun-Salak National Park (MHSNP) is one of tropical forest remnants in Java island. The National Park has been recognized with relatively high diversity of wild Zingiberaceae. Of that Zingiberaceae two species, namely
Alpinia malaccensis (AM) and Horstendia conica (HC), were domesticated as garden plants in the surrounding area of the forest for medicinal use. The impact of plant domestication to the pathogenic fungi has been reported. However, the impact of Zingiberaceae domestication in MHSNP to their fungal endophytes is never been studied yet. Therefore, diversity of fungal endophytes associated with these two species of Zingiberaceae grown in natural forest and those cultivated in the garden is reported here.
Fungal endophyte diversity in the wild and domesticated AM andHC was analyzed based on those that are culturable. One healthy individual plant of each species were collected from selected wild and domesticated AM dan HC. Each plant consisted of leaves, stem, root and rhizome. All these plant organs were surface sterilised. Several pieces of each organs were planted on half strength MEA (Malt Extract Agar) medium and incubated at room temperature. Fungi that grew out from the plant organs were isolated and purified. Identification to species level used morphological approach, and molecular approaches using ITS rDNA sequence.
Based on morphological observation to the colony, the fungal endophyte collections can be classified into 17 morphotypes. These morphotypes represented nineteen species of fungi, namely Arthrinium malaysianum, Aspergillus flavipes, As. sydowii, Chaetomium globosum, Cladosporium sp., Cladosporium oxysporum, Colletotrichum aff. boninense-complex, Co. aff. cliviae-complex, Co. aff.
gloeosporioides-complex, Diaporthe sp., D. anacardii, D. gardenia, Exophiala
sp., E. lecanii-corni, Guignardia mangiferae, Ochroconis gallopava, Penicillium citrinum, Pyricularia costina, and one isolates that tentatively identified as unsporulated Sydowiellaceae. These endophytic fungi belong to ascomycetes, in either Dothideomycetes, Eurotiomycetes, or Sordariomycetes.
Domestication affected the diversity and occurrance of the fungal endophytes. Fungal endophytes diversity in cultivated host was relatively lower than those in wild host, and this is in contrast to their occurrances. Domesticated
Zingiberaceae grow in rather open area and the wild Zingiberaceae that grow under forest canopy that cause their environment has less light intensity and more humid. Some endophytic fungi showed environmental preferences. The fungal endophytes community composition varied depending on their host species. Besides, their distribution were also depended on their micro-habitat. Fungal endophytes species were generally found in certain micro-habitat. Leaves was the most preferred micro-habitat, and followed by root, stem and rhizome.
v
host, but were not found in the cultivated host. Arthrinium malaysianum, Ch. globosum, Co. aff. cliviae-complex, D. gardenia, and unsporulated
Sydowiellaceae were newly emerging endophytes in the cultivated host. Some fungus, Co. aff. boninense-complex, Co. Aff. boninense-complex, Cladosporium
sp., Co. aff. gloeosporioides-complex, and E. lecanii-corni were not affected by domestication.
This research indicated that Zingiberaceae domestication affected the diversity and occurance of the fungal endophytes. The limitation number of samples and sampling frequency as well as in isolation method has restricted the the information obtained from this research. In order to get more accurate information, this research should therefore be strengthened with more samples and using metagenomic approach.
Key words: Domestication, diversity, fungalendophytes, Zingiberaceae
© Hak Cipta Milik IPB, Tahun 2014
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB
7
Tesis
sebagai salah satu syarat untuk memperoleh gelar
Magister Sains
pada
Program Studi Mikrobiologi
DAMPAK DOMESTIKASI
ZINGIBERACEAE
DI TAMAN
NASIONAL GUNUNG HALIMUN SALAK TERHADAP
KERAGAMAN CENDAWAN ENDOFITNYA
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
BOGOR
2014
9
Judul Tesis : Dampak Domestikasi
Zingiberaceae
di Taman Nasional Gunung Halimun Salak
Terhadap Keragaman Cendawan Endofitnya
Nama
: Ivan Permana Putra
NIM
: G351114011
Disetujui oleh
Komisi Pembimbing
Dr Gayuh Rahayu
Ketua
Iman Hidayat PhD
Anggota
Diketahui oleh
Ketua Program Studi Mikrobiologi
Prof Dr Anja Meryandini, MS
Dekan Sekolah Pascasarjana
Dr Ir Dahrul Syah, MSc Agr
Tanggal Ujian:
Tanggal Lulus:
PRAKATA
Alhamdulillahirobbil’alamin, puji syukur penulis panjatkan ke hadirat Allah SWT yang
telah memberikan rahmat dan karunia sehingga penulis dapat menyelesaikan karya ilmiah ini.
Tema yang dipilih dalam penelitian ini adalah mengenai Dampak Domestikasi
Zingiberaceae
di
Taman Nasional Gunung Halimun Salak terhadap Keragaman Cendawan Endofitnya. Penelitian
ini dilaksanakan mulai bulan Februari hingga November 2013 di Laboratorium Mikologi
Departemen Biologi, Institut Pertanian Bogor.
Penulis mengucapkan terima kasih kepada Dr Gayuh Rahayu dan Iman Hidayat PhD,
selaku pembimbing atas arahan dan bimbingan yang diberikan dalam pelaksanaan penelitian dan
penyusunan karya ilmiah ini. Penulis juga mengucapkan terimakasih kepada komite beasiswa
LPDP KEMENKEU atas bantuan dana penelitian yang diberikan sehingga penelitian ini
diselesaikan. Terima kasih yang sebesar-besarnya kepada keluarga tercinta Ayahanda Wahyu
Setyawan, Ibunda Hartini, Kakak Alfian Firdaus Setiawan, Kakak Wulan, Adek Agnes Putri
Wahyu Ningtyas, Keponakan Gelzie Adiva Naira dan Keluarga Besar atas do’a, dukungan, serta
kasih sayang yang diberikan. Terima kasih juga kepada Bu Tatik, Bu Nani, Bu Emi, Dr Sri
Listiyowati, Dr Nampiah Sukarno, Prof Dr Okky S Dharmaputra, Dr Lisdar I Sudirman, Pak
Kus, Sepri, Aldian, Mas Eris, Bu Mala dan teman-teman Lab Mikologi IPB atas semua
kebersamaan dan motivasi yang telah diberikan.
Semoga karya ilmiah ini dapat bermanfaat.
Bogor, Juli 2014
11
DAFTAR ISI
DAFTAR TABEL
xiv
DAFTAR GAMBAR
xiv
DAFTAR LAMPIRAN
xiv
1 PENDAHULUAN
Latar Belakang
1
Perumusan Masalah
2
Tujuan Penelitian
2
Manfaat Penelitian
2
2 METODE
Waktu dan Tempat Penelitian
2
Pengambilan Sampel Tanaman
2
Isolasi Cendawan Endofit
2
Pengelompokan Berdasarkan Morfotipe
2
Identifikasi Molekuler
3
Ekstraksi DNA, Amplifikasi, dan Sekuensing
3
Analisis Filogenetik
3
Analisis Keragaman dan Pengelompokan Komunitas
3
3 HASIL DAN PEMBAHASAN
Keragaman Cendawan Endofit Asal
Zingiberaceae
4
Arthrinium malaysianum
4
Aspergillus
spp.
6
Chaetomium globosum
7
Cladosporium
spp.
7
Colletotrichum
spp.
8
Diaporthe
spp.
9
Exophiala
spp.
10
Guignardia
mangiferae
10
Ochroconis gallopava
11
Penicillium citrinum
11
Pyricularia costina
12
Sydowiellaceae
13
Dampak Domestikasi Terhadap Keragaman Endofit Asal
Zingiberaceae
13
4 SIMPULAN DAN SARAN
Simpulan
18
Saran
18
DAFTAR PUSTAKA
18
LAMPIRAN
22
RIWAYAT HIDUP
31
DAFTAR TABEL
1
Parameter lingkungan sampel
Zingiberaceae
16
2
Keragaman endofit pada habitat yang berbeda
16
3
Daftar spesies endofit yang muncul, hilang, dan baru ditemukan
pada
Zingiberaceae
17
DAFTAR GAMBAR
1
Pohon filogenetik endofit asal
Zingiberaceae
5
2
Arthrinum
asal
Zingiberaceae
6
3
Aspergillus
asal
Zingiberaceae
6
4
Chaetomium
asal
Zingiberaceae
7
5
Cladosporium
asal
Zingiberaceae
8
6
Koloni
Colletotrichum
spp. pada PDA
8
7
Colletotrichum
asal
Zingiberaceae
8
8
Diaporthe
asal
Zingiberaceae
9
9
Exophiala
asal
Zingiberaceae
10
10
Guignardia
asal
Zingiberaceae
10
11
Ochroconis
asal
Zingiberaceae
11
12
Penicillium
asal
Zingiberaceae
12
13
Pyricularia
asal
Zingiberaceae
12
14
Sydowiellaceae
asal
Zingiberaceae
13
15
Frekuensi keberadaan endofit pada berbagai ekosistem
14
16
Analisis klustering endofit asal
Zingiberaceae
16
DAFTAR LAMPIRAN
1
Pohon filogenetik
Arthrinium
23
2
Pohon filogenetik
Aspergillus
23
3
Pohon filogenetik
Chaetomium
24
4
Pohon filogenetik
Cladosporium
25
5
Pohon filogenetik spesies kompleks dari genus
Colletotrichum
26
6
Pohon filogenetik
Diaporthe
spp.
28
7
Pohon filogenetik
Exophiala
29
8
Pohon filogenetik
Guignardia
29
9
Pohon filogenetik
Ochroconis
30
10
Pohon filogenetik
Penicillium
30
11
Pohon filogenetik
Pyricularia
30
1 PENDAHULUAN
Latar Belakang
Aktivitas antropogenik seperti domestikasi tanaman dapat menyebabkan perubahan kompleks pada morfologi dan fisiologi tanaman yang membedakannya dari karakter asli moyangnya (Hancock 2005). Secara langsung ataupun tidak, proses domestikasi memberikan pengaruh yang besar terhadap keragaman tanaman dan organisme di sekitarnya. Selama proses domestikasi berlangsung terdapat beberapa gen yang berubah dan menimbulkan variasi genetik pada tanaman yang didomestikasi. Domestikasi juga menyebabkan perubahan ekologi pada tanaman. Diferensiasi genetik antara tanaman liar dan budidaya juga mengarah kepada perubahan habitat mikroorganisme simbion yang menyebabkan perubahan komposisinya (Lebarbenchon et al. 2008; Stukenbrock dan McDonald 2008). Munkacsi et al. (2008) menyatakan bahwa beberapa cendawan patogen memiliki kemampuan untuk mengikuti tanaman inang selama proses domestikasi berlangsung melalui mekanisme host-tracking.
Setiap tumbuhan secara alami berasosiasi dengan mikroorganisme di lingkungannya. Pengaruh domestikasi tanaman (inang) terhadap komunitas mikroorganisme yang berasosiasi dengan tanaman itu seringkali kurang diperhatikan. Salah satunya ialah cendawan endofit. Cendawan endofit hidup di dalam jaringan inang tanpa menyebabkan gejala sakit ataupun kerugian lainnya. Asosiasi antara tumbuhan dan cendawan endofit bersifat mutualisme. Cendawan endofit memberikan beberapa keuntungan terhadap inangnya diantaranya: produksi faktor tumbuh, resistensi terhadap kekeringan dan hama, serta sumber metabolit sekunder yang bermanfaat bagi pertumbuhan inangnya (Schulz dan Boyle 2005).
Salah satu tanaman yang sering didomestikasi di Indonesia ialah
Zingiberaceae. Tanaman ini telah digunakan sebagai obat-obatan selama ratusan tahun. Priyadi et al. (2010) mengkonfirmasi bahwa setidaknya terdapat 10 spesies
Zingiberaceae liar di Taman Nasional Gunung Halimun Salak (TNGHS) dan beberapa diantaranya telah didomestikasi oleh penduduk setempat. Penelitian sebelumnya menunjukkan bahwa Zingiberaceae merupakan inang bagi cendawan. Lebih dari 170 spesies dan 17 genus cendawan endofit, saprob, dan patogen asal
Zingiberaceae telah dideskripsikan (Bussaban et al. 2002; Hyde et al. 2007). Pengaruh domestikasi Zingiberaceae terhadap keragaman endofit belum pernah dilaporkan sebelumnya.
Informasi mengenai perbandingan keragaman cendawan simbion serta keragamannya pada inang dengan karakteristik lingkungan yang berbeda jarang dilaporkan. Pengetahuan mengenai dampak aktivitas antropogenik terhadap keragaman hayati merupakan salah satu aspek yang perlu diperhatikan untuk menekan terjadinya gangguan selama eksploitasi tanaman, sehingga informasi tentang keberadaan endofit serta fungsinya terhadap inang pada kondisi lingkungan yang berbeda menjadi penting. Oleh sebab itu hasil penelitian ini diharapkan dapat memberikan gambaran mengenai ragam cendawan endofit pada tanaman liar dan domestikasinya dan menjadi rujukan dalam strategi domestikasi
Zingiberaceae di Indonesia.
Tujuan Penelitian
Penelitian ini bertujuan untuk:
1. Mengetahui keragaman cendawan endofit asal Zingiberaceae di TNGHS.
2. Menganalisis dampak domestikasi terhadap cendawan endofit asal
Zingiberaceae di TNGHS.
Manfaat Penelitian
Manfaat dari penelitian yaitu memberikan informasi mengenai keragaman endofit asal Zingibeaceae dan merupakan informasi awal yang diharapkan dapat menjadi rujukan dalam strategi domestikasi tanaman Zingiberaceae di Indonesia.
2 METODE
Waktu dan Tempat Penelitian
Penelitian ini dilaksanakan selama 10 bulan (Februari-November 2013) bertempat di Laboratorium Mikologi, Departemen Biologi, Institut Pertanian Bogor.
Pengambilan Sampel Tanaman
Zingiberaceae yang digunakan dalam penelitian ini ialah Alpinia malaccensis (nama lokal: Raja Gowah) dan Hornstedtia conica (nama lokal: Pinding Hijau). Sampel dipilih dari rumpun liar dari kedua spesies Zingiberaceae
yang sehat (tidak menunjukkan gejala sakit) di kawasan TNGHS maupun rumpun hasil budidaya yang sehat di desa Citalahab, perbatasan perkebunan teh Nirmala dan kawasan TNGHS, Jawa Barat pada bulan Februari 2012. Satu individu tanaman terdiri dari akar, rimpang, batang, dan daun diambil dari satu rumpun
Zingiberaceae terpilih. Sampel dicuci dan ditempatkan di dalam plastik ziplock, kemudian diletakkan di dalam box pendingin untuk dibawa ke laboratorium.
Isolasi Cendawan Endofit
Isolasi endofit dilakukan dengan mengacu pada Okane et al. (2008). Sampel dipotong-potong dengan ukuran 0.5 x 0.5 cm2. Sebanyak 12 potong sampel (4 potong/cawan) dari setiap bagian tanaman diletakkan pada dalam media
Malt Extract Agar (MEA) half-strengh dan diinkubasi pada suhu 25˚C selama 1 bulan. Ujung hifa dari setiap koloni yang tumbuh diisolasi dan dimurnikan. Koloni yang murni ditumbuhkan pada media Low Carbon Agar (LCA) dan kemudian Potato Dextrose Agar (PDA).
Pengelompokan Berdasarkan Morfotipe
3
elevasi, tekstur, tipe miselia, tepian, densitas, serta diameter. Biakan murni dari setiap morfotipe diidentifikasi secara morfologi sampai tingkat genus.
Identifikasi Molekuler
Minimal satu biakan dipilih untuk mewakili satu morfotipe dalam identifikasi ke tingkat spesies. Identifikasi sampai ke tingkat spesies dilakukan dengan pendekatan molekuler menggunakan daerah ITS, dan EF jika diperlukan.
Ekstraksi DNA, Amplifikasi, dan Sekuensing
Genom DNA diisolasi dari miselia yang ditumbuhkan pada media PDB selama 7 hari pada suhu ruang. Miselia dipanen dengan menggunakan spatula dan dimasukkan ke dalam tabung mikro berukuran 2 ml. Ekstraksi dilakukan dengan menggunakan Phytopure DNA extraction kit (GE). Setiap tabung berisi 25 μl yang terdiri dari 10 μl nuclease free water, Go taq green mastermixTM 12.5 μl, ITS5 (5′-GGAAGTAAAAGTCGTAACAAGG-3′) dan ITS4 (5′ -TCCTCCGCT-TATTGATATGC-3′) (White et al. 1990) masing-masing 0.5 μl, 0.5 μl DMSO, dan1 μl DNA target. Amplifikasi dilakukan dengan kondisi PCR yaitu predanaturasi 95oC selama 90 detik, selanjutnya diikuti oleh 35 siklus, denaturasi (94°C, 30 detik), annealing (52°C, 30 detik) extension (72°C, 90 detik), dan final extension 72oC selama 5 menit. Amplifikasi untuk primer EF,EF1-728F (5′ -CATCGAGAAGTTCGAGAAGG-3′), EF1-986R (5′ -TACTTGAAGGAACCC-TTACC3′) dilakukan dengan komposisi dan protokol yang sama kecuali untuk komposisi primer. Hasil amplifikasi kemudian dielektroforesis pada gel agarose dengan konsentrasi 1 % dan direndam dalam etidium bromida selama ± 30 menit dan divisualisasikan dibawah sinar UV (Bio-Rad UV Transilluminator 2000). Hasil PCR dikirim ke 1st Base (Malaysia) untuk disekuensing.
Analisis Filogenetik
Sekuen yang diperoleh dibandingkan dengan sekuen dari database the National Center for Biotechnology Information (NCBI) (www.ncbi.nlm.nih.gov) melalui program BLASTn (Basic Local Alignment Search Tool for nucleotide) untuk memperoleh nama genus terdekat. Sekuens homolog diambil dan disejajarkan dengan sekuen yang sedang dianalisa menggunakan program MEGA 5 (Tamura et al. 2011). Sekuen rujukan juga menyertakan publikasi dari genus terkait. Publikasi yang digunakan adalah : Asgari dan Zare (2011), Brensch et al.
(2012), Busssaban et al. (2005), Cannon et al. (2012), Crous dan Groenewald (2013), Glienke et al. (2011), Gomes et al. (2013), Houbraken et al. (2010), Kruys dan Castlebury (2012), Martin-Shanchez et al. (2012), Seyedmousavi et al. (2011), Suh et al. (2012), dan Thompson et al. (2011). Pohon filogenetik dibangun dengan metode Maximum Likelihood (ML) menggunakan program MEGA 5 dengan 1000 ulangan bootstrap (BS) serta analisis Bayesian (untuk
Analisis Keragaman dan Pengelompokan Komunitas
Keragaman dinyatakan dalam frekuensi keberadaan endofit (satu isolat dianggap sebagai satu individu) pada ekosistem dan organ yang berbeda. Frekuensi keberadaan dihitung dengan persamaan = x 100% Keragaman juga dinyatakan dalam indeks keragaman Shannon-Wiener (H’). Indeks Shannon-Wiener dengan menggunakan rumus sebagai berikut: H= -Ʃpi. ln pi, dimana pi( kelimpahanrelatif) = ∑
∑ . Keberadaan
endofit pada setiap organ dan ekosistem inang dikelompokkan dengan program PASW (Predictive Analytics Software) versi 18.
3 HASIL DAN PEMBAHASAN
Keragaman Cendawan Endofit Asal Zingiberaceae
Jumlah total koloni dan periode inkubasi yang diperlukan cendawan endofit untuk muncul dari segmen bervariasi. Sebagian besar organ tanaman yang diisolasi mengandung satu koloni cendawan, sementara sebagian lainnya bebas cendawan atau memiliki lebih dari satu koloni. Setelah purifikasi, sebanyak 148 isolat cendawan endofit berhasil diisolasi dari 192 organ tanaman yang diinkubasi selama 1-3 bulan. Koloni pertama muncul setelah 5 hari inkubasi, koloni yang paling lambat muncul setelah 3 bulan inkubasi, namun sebagian besar koloni muncul setelah 5-14 hari inkubasi.
Semua endofit yang berhasil diisolasi dikelompokkan ke dalam 17 morfotipe berdasarkan penampakan koloninya. Semua isolat endofit pada
Zingiberaceae tergolong Ascomycota, Pezizomycotina yang tersebar ke dalam 3 kelas yaitu: Sordariomycetes, Eurotyomycetes, dan Dothideomycetes (Gambar 1).
Endofit tersebut merupakan fase anamorf dari Askomiset. Hasil ini mengkonfirmasi penelitian sebelumnya yang dilakukan oleh Davies et al. (2003), Rodriguez et al. (2009), dan Khan et al. (2010). Mereka melaporkan bahwa sebagian besar cendawan endofit merupakan anggota Askomiset. Davies et al. (2003) menyatakan bahwa Askomiset mampu berasosiasi dengan lumut. Sementara itu Khan et al. (2010) juga menambahkan bahwa semua cendawan endofit asal Solanaceae tergolong ke dalam Askomisetdan anamorfnya.
Hampir semua morfotipe cendawan endofit telah diidentifikasi sampai dengan spesies, kecuali satu morfotipe yang tidak bersporulasi hanya dapat diidentifikasi sampai dengan tingkat famili. Adapun cendawan-cendawan endofit yang ditemukan pada penelitian ini adalah sebagai berikut:
Arthrinium malaysianum
Satu galur endofit asal akar H. conica (HCD D6) diidentifikasi sebagai A. malaysianum, karena galur ini berada dalam satu clade dengan A. malaysianum
5
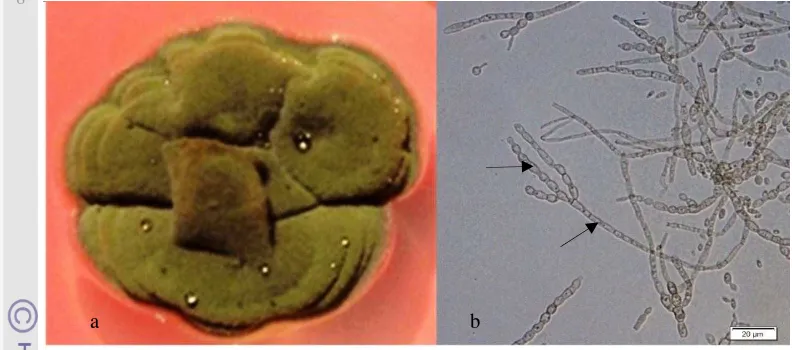
Gambar 2 Arthrinum malaysianum asal Zingiberaceae a. koloni pada PDA umur 30 hari danb. konidia
Arthrinium pernah dilaporkan sebagai endofit pada rerumputan (Crous and Groenewald 2013), tanaman Juncus trifidus (Chlebicki 2009), dan liken (He dan Zhang 2012). Arthrinium malaysianum belum pernah dilaporkan hidup sebagai endofit, sehingga laporan ini merupakan laporan pertama A. malaysianum sebagai endofit.
Aspergillus spp.
Dua galur Aspergillus (HCL R2 dan AML B6) endofit Zingiberaceae pada penelitian ini dikelompokkan ke dalam dua morfotipe. Morfotipe pertama memiliki ciri koloni yang menutupi media PDA setelah 14 hari inkubasi dengan bentuk sirkuler. Koloni bertepung putih, dan tepiannya rata (Gambar 3a). Berbeda dengan morfotipe 1, koloni morfotipe 2 berwarna jingga (Gambar 3b). Kedua morfotipe ini memiliki konidiofor tidak bersekat dan hialin, vesikel bulat hingga lonjong, fialid uniseriat serta konidianya bulat (Gambar 3c). Analisis molekuler menunjukkan bahwa morfotipe 1 berada dalam satu clade dengan galur rujukan
As. flavipes isolat NRRL 295 dengan nilaiBS 0.789dan morfotipe 2 berada dalam satu clade dengan galur As. sydowii isolat NRRL 254 rujukan (BS = 0.60) (Gambar Lampiran 2), sehingga morfotipe 1 adalah As. flavipes dan morfotipe 2 adalah As. sydowii.
Beberapa spesies Aspergillus telah dilaporkan hidup sebagai endofit berbagai tanaman. Aspergillus sydowii pernah dilaporkan sebagai endofit pada pada Gorgonia spp. oleh Toledo-Hernandez et al. (2008) dan Tinospora cordifolia
asal India oleh Mishra et al. (2012).
a b
Gambar 3 Koloni dan ciri mikroskopis Aspergillus spp., a. koloni As. flavipes pada PDA, b. koloni As. sydowii pada PDA, c. konidiofor, vesikel, fialid dan konidia
c
a b
konidiofor vesikel fialid
7
Chaetomium globosum
Hanya satu isolat (HCD B1) yang secara molekuler diidentifikasi sebagai
Ch. globosum, karena isolat tersebut berada dalam satu clade dengan galur rujukan Ch. globosum CgD9HN3768 (Gambar Lampiran 3) dengan nilai BS yang kuat (0.998). Morfotipe galur ini dicirikan dengan koloni yang menutupi media PDA setelah 14 hari inkubasi, koloni berwarna putih dengan tepian koloni jingga dan rata (Gambar 4a). Askospora berwarna coklat gelap, bersel tunggal, oval melonjong dengan cekungan di bagian tengahnya (Gambar 4b). Askoma berambut dan berwarna kecoklatan (Gambar 4c).
Penelitian ini merupakan laporan pertama mengenai keberadaan
Chaetomium endofit dari Zingiberaceae di Indonesia. Sebelumnya, Khan et al. (2010) juga menemukan Ch. bostrycodes endofit namun pada tanaman obat
Withania somnifera.
Gambar 4 Chaetomium globosum asal Zingiberaceae a. koloni pada media PDA umur 30 hari; b. konidia; c. peritesium.
Cladosporium spp.
Semua galur Cladosporium yang berhasil diisolasi pada penelitian ini dikelompokkan menjadi satu morfotipe. Seluruh spesies Cladosporium pada penelitian ini menutupi media PDA setelah 30 hari inkubasi. Koloninya kompak, convex, dan berwarna kehijauan (Gambar 5a). Konidia berantai, lurus atau sedikit melengkung (Gambar 5b), bagian terminal atau interkalar elips, gelendong, atau agak bulat dengan bekas luka di kedua ujung konidia. Dengan analisis molekuler, morfotipe ini dapat dibagi ke dalam 2 spesies. Sebanyak 12 isolat diidentifikasi sebagai Cladosporium sp. yang memiliki kekerabatan dengan Cl. dominicanum
dan Cl. sphaerospermum, dan satu isolat (AMD R5) yang berada dalam satu
clade dengan Cl. oxysporum HSW-16 rujukan dengan nilai BS (0.881).
Beberapa spesies Cladosporium dilaporkan hidup sebagai endofit pada berbagai tanaman. Di Indonesia, Cl. oxysporum pernah dilaporkan sebagai endofit pada tanaman pacar cina (Aglaia odorata) (Sugijanto dan Dora 2012). Sebelumnya, Paul dan Yu (2008) melaporkan bahwa Cl. oxysporum dan Cl. sphaerospermum adalah cendawan endofit pada daun jarum Pinus sp. di Korea.
a b c
Colletotrichum spp.
Seluruh galur Colletotrichum endofit Zingiberacae terbagi ke dalam tiga morfotipe. Secara umum, keseluruhan koloni menutupi media PDA setelah 30 hari inkubasi, koloni bertipikal aerial, seperti beludru, permukaan koloni datar dengan pola pertumbuhan melingkar berwarna abu-abu hingga putih (Gambar 7) dengan permukaan bawah koloni berwarna jingga. Sel konidiogen hialin dan berdinding halus, konidiofor dibentuk langsung pada hifa dan berwarna abu pucat hingga coklat (Gambar 6a). Konidia hialin, aseptat dan silindris pada kedua ujungnya (Gambar 6b). Apressoria sederhana berbentuk bundar hingga elips (Gambar 6c).
a b
Gambar 5 Cladosporium sp. asal Zingiberaceae a. koloni pada PDA, b. konidiofor dan rantai konidia.
a
b
c
a
b
Gambar 6 Ciri mikroskopik Colletotrichum gloeosporioides a. aservulus b. konidia c. apressoria.
9
Sebanyak satu galur Colletotrichum asal Zingiberaceae termasuk dalam
clade Co. cliviae (sister clade dari clade orbiculare), 14 galur termasuk dalam
clade Co. Gloeosporioides, serta 12 galur Co. boninense dengan nilai BS yang lemah (Gambar Lampiran 5), sehingga secara tentatif galur-galur tersebut dinyatakan memiliki kedekatan dengan Co. aff. cliviae, Co. aff. gloeosporioides, dan Co aff. boninense. Bussaban et al. (2001) melaporkan Co. gloeosporioides
sebagai endofit pada Zingiberaceae liar di Thailand. Penelitian ini menunjukkan bahwa Colletotrichum juga merupakan endofit asal Zingiberaceae liar dan domestikasi.
Diaporthe spp.
Semua Diaporthe spp. asal Zingiberaceae terbagi ke dalam empat morfotipe. Setiap morfotipe ditunjukkan oleh pertumbuhan koloni dengan pola sirkuler yang berbeda-beda (Gambar 8a). Konidiomata berwarna hitam gelap (gambar 8b). Konidiofor dan konidiogennya berwarna hialin hingga kecoklatan. Konidia alfa hialin, oval, hingga silindris (Gambar 8d) sedangkan konidia beta aseptat, hialin, melengkung, dan lonjong (Gambar 8c). Berdasarkan analisis molekuler kombinasi gen ITS dan EF, isolat-isolat Diaporthe yang didapatkan pada penelitian ini terbagi ke dalam dua spesies yaitu D. anacardii CBS 72097 (BS = 0.99) dan D. Gardenia CBS 28856 (BS = 1) dan dua isolat yang belum dapat diidentifikasi sampai dengan spesies (Gambar Lampiran 6).
Diaporthe spp. telah dilaporkan sebagai patogen, endofit, dan saprob dengan kisaran inang yang luas (Gomes et al. 2013). Bussaban et al. (2001) berhasil mengisolasi Diaporthe spp. pada Zingiberaceae di Thailand. Hasil penelitian ini menguatkan informasi bahwa Diaporthe spp. adalah endofit
Zingiberaceae yang persebarannya di daerah Asia Tenggara.
a
b
d c
Exophiala spp.
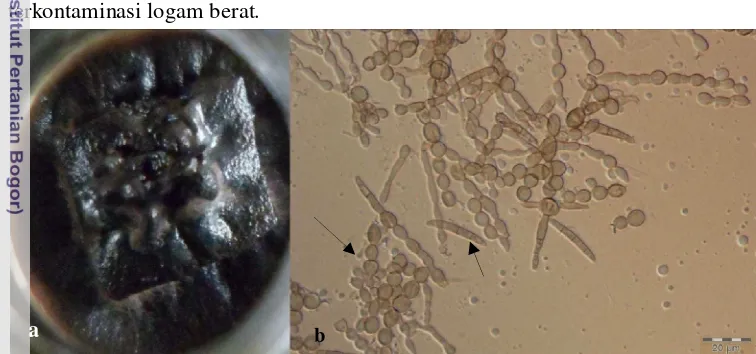
Galur Exophiala yang diperoleh pada penelitian ini dikelompokkan ke dalam satu morfotipe. Koloninya tumbuh lambat dan menutupi media PDA setelah 90 hari inkubasi dengan bentuk koloni yang tidak beraturan (Gambar 9a). Koloni kompak, convex, berwarna hitam gelap, dengan tepian koloni rata. Konidia terdiri dari dua tipe yakni bulat dan fusiform bersekat (Gambar 9b), keduanya hialin dan dapat berada dalam satu untaian konidia. Analisis molekuler menunjukkan bahwa enam isolat yang diisolasi dari akar (5 isolat) dan daun (1 isolat) teridentifikasi sebagai Exophiala spp. Dua isolat berada dalam satu clade
dengan E. lecanii-corni PW2642 dengan nilai BS sebesar 0.621 (Gambar Lampiran 7), namun empat isolat lainnya berada dalam satu clade yang terpisah dari clade strain-strain rujukan sehingga perlu dianalisis lebih lanjut untuk identifikasi hingga ke tingkat spesies.
Exophiala merupakan DSE (Dark Septate Endophyte) yang peranannya sebagai cendawan endofit masih sedikit dipelajari. Kelompok cendawan ini dikenali dengan cirinya yang mengandung pigmen hitam gelap (melanin) pada koloninya. Li et at. (2011) melaporkan bahwa E. pisciphila berperan sebagai endofit yang membantu pertumbuhan Zea mays pada lingkungan yang terkontaminasi logam berat.
Guignardiamangiferae
Tiga galur Guignardia yang diperoleh dikelompokkan ke dalam satu morfotipe. Koloninya tumbuh lambat, memiliki bentuk yang tidak teratur, mengeras seperti batu, cembung, berwarna hitam kehijauan, dan tepian koloni bergelombang (Gambar 10a). Konidiofor umumnya pendek dan sering tidak terlihat, konidium bersel satu, berbentuk oval, dan hialin (Gambar 10b). Berdasarkan analisis molekuler tiga galur morfotipe, galur-galur ini berada satu
clade dengan G. mangiferae IMI 260576 dengan nilai BS sebesar 0.512 (Gambar Lampiran 8).
Guignardia dikenal sebagai endofit dan patogen pada berbagai tanaman selain Zingiberacae ataupun sebagai saprob (Okane et al. 2003; Silva et al. 2008; Huang et al. 2009; Wulandari et al. 2009). Meskipun demikian sampai saat ini laporan mengenai Guignardia sebagai endofit pada Zingiberaceae belum ditemukan.
a b
11
Ochroconis gallopava
Koloni dari kapang ini dikelompokkan dalam satu morfotipe dan menutupi media PDA setelah 14 hari inkubasi dengan tipe koloni sirkuler (Gambar 11a). Koloni bertipikal aerial, membatu, convex, berwana hijau keabuan, dan tepian koloni yang bergelombang. Konidiofor bercabang dengan konidia kecoklatan yang berantai pada ujungnya (Gambar 11b).
Analisis molekuler sekuen ITS menunjukkan bahwa dua isolat asal kedua species Zingiberaceae yang diteliti berada dalam satu clade dengan Ochroconis gallopava IFM 52605 dengan nilai BS yang kuat (0,893) (Gambar Lampiran 9).
Ochroconis pertama kali dilaporkan sebagai endofit oleh Ho et al. (2012) pada
Zanthophylum wutaiense (Rutaceae) di Taiwan. Di Indonesia, adanya Ochroconis
endofit pada Zingiberaceae pertama kali dilaporkan.
Gambar 11 Ochroconis gallopava asal Zingiberaceae a. koloni pada PDA b. konidiofor (tanda panah) dengan rantai konidia di ujung konidiofornya
Penicillium citrinum
Tiga galur Penicillium yang diperoleh dikelompokkan ke dalam satu morfotipe. Koloni menutupi seluruh media PDA setelah 10 hari inkubasi dengan bentuk koloni sirkuler, berwarna hijau keabuan, dan tepian koloni bergelombang (Gambar 12a). Konidiofor hialin dengan percabangan 3 pada ujungnya. Konidia globoid, hialin hingga kehijauan, serta bergerombol (Gambar 12b).
Analisis molekuler sekuen ITS menunjukkan dua galur berada dalam satu
clade dengan strain P. citrinum GL-7 dengan nilai BS sebesar 0.992, sedangkan satu galur lainnya adalah Penicillium aff. citrinum GL-7 yang berada dalam satu clade dengan P. citrinum GL-7 rujukan dengan nilai BS 0.662 (Gambar Lampiran 10). Penicillium citrinum sebelumnya telah dilaporkan sebagai endofit pada akar kopi (Posada et al. 2007) dan akar Ixeris repenes (Khan et al. 2008).
a
a b
Gambar 10 Guignardia mangiferae asal Zingiberaceae, a. koloni pada PDA dan b. konidia b
Endofit yang didapatkan pada penelitian ini berasal dari akar dua spesies Zingibecaeae.
Gambar 12 Penicillium citrinum asal Zingiberaceae a. koloni pada PDA b. konodiofor dan konidia
Pyricularia costina
Pyricularia telah sering dilaporkan sebagai patogen tanaman pada berbagai komoditas penting pertanian. Namun hanya Bussaban et al. (2001) yang pernah melaporkan Py. costina sebagai endofit dari Zingiberaceae di Thailand. Pada penelitian ini, Pyricularia berhasil dikoleksi dan analisis molekuler dengan gen ITS-nya menunjukkan bahwa isolat yang didapatkan adalah Py. costina
CMUZE0003 dengan nilai BS sebesar 0.765 (Gambar Lampiran 13).
Semua galur Py. costina berada pada satu morfotipe dan menutupi media PDA setelah 30 hari inkubasi, koloni cembung dengan bagian atas mengeras, warna keabuan hingga krem (Gambar 13a). Konidia bersekat, piriform hingga lonjong, serta hialin (Gambar 13c).
Gambar 13 Pyricularia asal Zingiberaceae a. koloni pada PDA, b. konidia. a
b
konidiofor
konidia
a b
13
Sydowiellaceae tidak bersporulasi
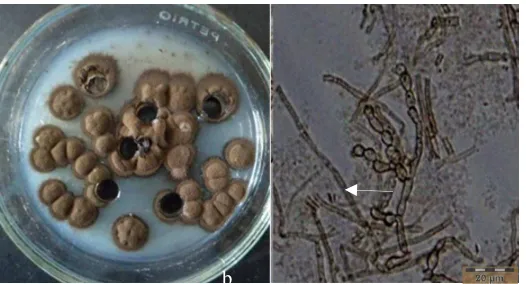
Pada penelitian ini koloni Sydowiellaceae hanya memiliki satu morfotipe. Koloni menutupi media setelah 14 hari inkubasi, seperti beludru, convex, warna krem, dan tepian koloni rata (Gambar 14), dan tidak bersporulasi. Pada saat BLASTn terhadap database nukleotida di NCBI, sekuen dari isolat HCD D10 ini memiliki kesamaan yang tertinggi dengan Prosthecium piriforme dan Sydowiella fenestrans, anggota dari Sydowiellaceae yaitu berturut-turut 89% dan 91%. Pada saat analisis filogenetik dengan anggota Sydowiellaceae (Gambar Lampiran 12), galur HCD 10 berada di luar clade strain rujukan sehingga identifikasi hanya bisa dilakukan hingga level famili.
Gambar 14 Koloni Sydowiellaceae tidak bersporulasi asal Zingiberaceae pada PDA umur 30 hari.
Dampak Domestikasi Terhadap Keragaman Endofit Asal Zingiberaceae
Semua sampel tanaman yang diambil terlihat tumbuh dengan sehat dan segar. Hal ini menunjukkan bahwa kondisi kesehatan tanaman tidak dipengaruhi oleh kondisi lingkungannya. Sampel Zingiberaceae yang dikoleksi berasal dari 2 ekosistem yang berbeda yakni hutan alami dan lahan semi-pertanian (Tabel 1). Parameter lingkungan antara 2 ekosistem tersebut sangat berbeda, terutama pada intensitas cahaya dan kecepatan angin. Zingiberaceae yang diambil dari ekosistem alami tumbuh di bawah kanopi hutan, sementara sampel yang diambil dari lahan semi-pertanian berada pada daerah yang lebih terbuka. Namun pengaruh kedua faktor ini pada keanekaragaman cendawan endofit belum diketahui. Thomas et al. (1988) menyatakan bahwa kecepatan angin mempengaruhi kelembapan relatif pada suatu ekosistem dan mempengaruhi perkembangan hifa cendawan.
Walaupun semua anggota komunitas cendawan endofit asal Zingiberaceae
0.00 10.00 20.00 30.00 40.00 50.00 60.00 70.00 F re k u en si k eb er ad aa n ( % ) Ekosistem Ar. malaysianum As. flavipes As. Sydowii Ch. globosum Cl. oxysporum Cladosporium sp.
Co. boninense complex
Co. cliviae complex
Co. gloeosporioides complex
Diaporthe sp. D. anacardii D. gardenia Exophiala sp. E. lecanii-corni G. mangiferae O. gallopava P. citrinum Py. costina unsporulated Sydowiellaceae
15
terbesar (33,11%) diikuti oleh AMD (25,68%), HCL (22,30%), dan AML (18,92%). Hasil tersebut berbanding terbalik dengan index diversitas (Shanon-Wiener) yang didapatkan (Tabel 2). Secara umum, A. malaccensis (AM) memiliki jumlah dan jenis endofit yang lebih sedikit dibandingkan dengan H. conica (HC). Inang yang memiliki keragaman endofit tertinggi ialah AML (2,14) diikuti dengan AMD (1,90), HCL (1,67) dan HCD (1,53) (Tabel 2). Gerard et al. (2006) menyatakan bahwa lahan semi-pertanian/domestikasi dicirikan dengan sistem pertanian yang monokultur dengan keragaman gen yang rendah dan densitas inang yang padat. Data yang diperoleh dari penelitian ini menunjukkan bahwa beberapa endofit memiliki perbedaan dalam kemampuan untuk beradaptasi dengan lingkungan baru (lahan semi-pertanian). Rodriguez et al. (2009) mengkonfirmasi bahwa beberapa endofit memiliki kemampuan beradaptasi yang baik terhadap habitat baru sehingga menyebabkan baik endofit ataupun inangnya bisa bertahan dalam perubahan ekosistem yang terjadi.
Beberapa endofit kadang-kadang memiliki preferensi ekosistem.
Colletotrichum aff. boninense-complex, Cladosporium sp., dan E. lecanii-corni
mendominasi komunitas endofit pada semua jenis dan lingkungan inang (16,89% - 24,32%). Colletotrichum aff. gloeosporioides-complex (13,51%) juga sering ditemukan pada kedua jenis inang. Hal ini menunjukkan bahwa spesies-spesies tersebut tidak memiliki preferensi ekosistem. Sebaliknya, spesies cendawan lainnya merupakan endofit dengan tipe ekosistem spesifik, misal D. anacardii
hanya ditemukan pada AM, G. mangiferae serta P. citrinum hanya ditemukan pada HC. Suryanarayanan et al. (2000) serta Cannon dan Simmons (2002) sebelumnya telah mengkonfirmasi adanya spesifitas endofit terhadap inangnya. Bahkan, laporan terbaru menunjukkan bahwa endofit diduga tidak hanya memiliki spesifitas terhadap inang tetapi juga terhadap organ inang (Moricca et al. 2012). Kecenderungan ini dipengaruhi oleh interaksi antara inang dan ekosistem inang, serta interaksi kompetitif dari komunitas endofit. Interaksi tersebut mengakibatkan terbentuknya diversifikasi relung ekologi termasuk pemilihan inang dan organnya.
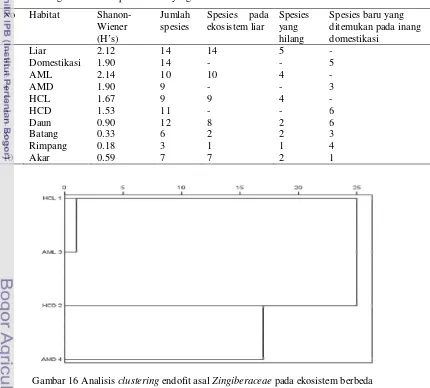
Komunitas cendawan pada HCL lebih dekat ke AML dibandingkan dengan HCD dan AMD (Gambar 16). Komunitas endofit pada AMD merupakan kelompok yang paling berbeda dibandingkan dengan endofit pada inang lainnya. Hal ini mengindikasikan bahwa tanaman domestikasi memiliki komunitas endofit yang lebih seragam dibandingkan dengan tanaman liar. Higgins et al. (2007) dan Sun et al. (2008) melaporkan bahwa komunitas endofit tampaknya dipengaruhi oleh inangnya. Wilbeforce et al. (2003) mengkonfirmasi bahwa aktivitas antropogenik seperti halnya pertanian monokultur berpotensi mengakibatkan perubahan keragaman mikroorganisme tanaman inang habitat liarnya.
Tabel 1 Parameter lingkungan sampel Zingiberaceae
No Parameter Inang* pada ekosistem berbeda
AML AMD HCL HCD
1 Posisi/Lintang S: 06° 44.293’
N:106º31.606’
S: 06° 44.333’ N:106º31.865’
S: 06° 44.293’ N:106º31.606’
S: 06° 44.327’ N:106º31.906’
2 Ketinggian (m DPL) 1104 1134 1104 1134
3 Kelembapan Relatif (%)
78 74 78 74
4 Suhu(◦C) 25 25 25 25
5 Cahaya (Lux) 773 1203 773 1946
6 Angin (MPH) 0 2.2 0 2.2
*AM: Alpinia malaccensis;HC: Horstendia conica;L= Liar/alami ; D= Domestikasi/dibudidayakan
Tabel 2 Keragaman endofit pada habitat yang berbeda
No Habitat
Shanon-Wiener (H’s)
Jumlah spesies
Spesies pada ekosistem liar
Spesies yang hilang
Spesies baru yang ditemukan pada inang domestikasi
1 Liar 2.12 14 14 5 -
2 Domestikasi 1.90 14 - - 5
3 AML 2.14 10 10 4 -
4 AMD 1.90 9 - - 3
5 HCL 1.67 9 9 4 -
6 HCD 1.53 11 - - 6
7 Daun 0.90 12 8 2 6
8 Batang 0.33 6 2 2 3
9 Rimpang 0.18 3 1 1 4
10 Akar 0.59 7 7 2 1
[image:30.612.103.533.263.651.2]17
ditentukan oleh jenis jaringan/organ yang ditempatinya dan perbedaan kemampuan untuk menggunakan substrat merupakan penyebab preferensi tersebut (Schardl et al. 2004). Colletotrichum aff. cliviae-complex, Diaporthe spp., G. mangiferae, dan Py. costina spesifik pada daun, 3 spesies (Arthrinium malaysianum , Cl. oxysporum,P. citrinum) dan satu spesies Sydowiellaceae yang tidak bersporulasi spesifik pada akar, 2 spesies pada batang (As. sydowii dan Ch. globosum), dan 1 spesies (As. flavipes) pada rimpang. Sementara itu cendawan endofit yang tidak spesifik organ yaitu: Cladosporium sp., Co. aff.
boninense-complex, Co. aff. gloeosporioides-complex, E. lecanii-corni, dan O. gallopava.
Colletotrichum aff. boninense-complex lebih menyukai daun (37,16%), dan cendawan ini juga ditemukan pada akar dan batang tetapi tidak pada rimpang. E. lecanii-corni ditemukan pada daun dan akar, sementara itu O. gallopava
ditemukan pada batang dan daun.
Penelitian ini membuktikan bahwa domestikasi Zingiberaceae mendorong hilangnya spesies cendawan endofit dari inang liarnya atau sebaliknya mendorong munculnya spesies cendawan endofit yang baru ditemukan pada inang yang dibudidayakan. Aspergillus flavipes, As. sydowii, Cl. oxysporum, Diaporthe sp. dan Py. costina ditemukan pada inang liarnya dan tidak ditemukan lagi pada inang domestikasi. Arthrinium malaysianum, Ch. globosum, Co. aff. cliviae-complex,
[image:31.612.100.544.427.626.2]D. gardenia, dan Sydowiellaceae tak bersporulasi hanya ditemukan pada inang domestikasi. Sementara itu, beberapa spesies endofit yaitu Co. boninense -complex, Cladosporium sp., Co. aff. gloeosporioides-complex, dan E. lecanii-corni, tampaknya tidak dipengaruhi oleh domestikasi. Secara umum, 5 dari 14 spesies hilang akibat domestikasi Zingiberaceae (Tabel 2; Tabel 3).
Tabel 3 Daftar spesies cendawan endofit* yang muncul, hilang, dan baru ditemukan pada Zingiberaceae
Habitat Spesies pada ekosistem liar Spesies yang hilang
Spesies baru yang ditemukan pada inang domestikasi
Liar Asf, Ass, Cl, Clo,Cob,Cog,
Dsp, Da, E, El, G, P, Pyc, Ogv
Asf, Ass, Clo, Dsp, Pyc
-
Domestikasi - - Arm, Ch, Coc, Dg, US
AM Ass, Cl, Clo,Cob, E, El, G, P, Pyc, Ogv
Ass, Clo, Dsp, Pyc Coc, Cog, Dg
HC Asf, Cl, Cob, Dsp, Da, E, El, P, Pyc
Dsp, E, El, Pyc Arm, Ch, Cog, Da, US, Ogv
Daun Cl, Cob,Cog, Dsp, Da, G,
Pyc
Dsp, Pyc Coc, Dg,E, El, US, Ogv
Batang Ass, Ogv Ass, Ogv Ch, Cl, Cob
Rimpang Asf Asf Ch, Cl,Cob, P
Akar Cl, Clo, Cob, Cog, E, El, P Cl, Clo Arm
Keterbatasan dalam jumlah dan frekuensi pengambilan sampel serta metode analisis komunitas menyebabkan perlunya kehati-hatian dalam interpretasi hasil penelitian. Meskipun demikian, penelitian ini dapat menjadi informasi awal mengenai dampak domestikasi terhadap keragaman endofit terutama pada
Zingibercaeae yang masih perlu diperkuat dengan penambahan jumlah sampel serta analisis komunitas dengan pendekatan metagenomik agar hasil yang diperoleh lebih akurat.
4 SIMPULAN DAN SARAN
Simpulan
Berdasarkan analisis cendawan yang dapat dikulturkan, domestikasi dua spesies Zingiberaceae di kawasan Taman Nasional Halimun Salak (Alpinia malaccensis dan Horstendia conica) mengindikasikan adanya dampak yang signifikan terhadap keragaman cendawan endofitnya.Tanaman domestikasi memiliki populasi endofit yang lebih tinggi dibandingkan dengan spesies yang sama pada ekosistem alaminya. Zingiberaceae liar memiliki keragaman cendawan endofit yang lebih tinggi dibandingkan dengan Zingiberaceae yang telah didomestikasi. Beberapa spesies cendawan hanya ditemukan pada tanaman yang didomestikasi dan beberapa lainnya hilang sebagai akibat domestikasi. Namun, domestikasi Zingiberaceae di TNHS tidak mempengaruhi keberadaan endofit dominan, Co. boninense.
Saran
Penelitian lebih lanjut dengan penambahan jumlah sampel Zingiberaceae
serta analisis molekuler cendawan endofit dengan tambahan gen diperlukan untuk mengkonfirmasi dampak domestikasi Zingiberaceae dan keragaman endofitnya.
DAFTAR PUSTAKA
Asgari B, Zare R. 2011. The genus Chaetomium in Iran, a phylogenetic study including six new species. Mycologia 103(4): 863-882.
Arnold AE, Lutzoni F. 2007. Diversity and host range of foliar fungal endophytes : are tropical leaves biodiversity hotspots?. Ecol 88(3):541-549.
Brensch K, Braun U, Groenewald JZ, Crous PW. 2012. The genus Cladosporium. Stud Mycol 72: 1-401.
Bussaban B, Lumyong S, Lumyong P, McKenzie EHC, Hyde KD. 2001. Endophytic fungi from Amomum siamese. Can J Microbiol. 47:943-948. Bussaban B, Lumyong P, McKenzie KHC, Hyde KD, Lumyong S. 2002. Index of
fungi described from Zingiberaceae. Mycotaxon 83:165-182.
Bussaban B, Lumyong S, Lumyong P, Seelanan T, Park DC, McKenzie EHC, Hyde KD. 2005. Molecular and morphological characterization of
Pyricularia and allied genera. Mycologia 97(5): 1002-1011.
Cannon PF, Simmons CM. 2002. Diversity and host preference of leaf endophytic fungi in the Iwokrama Forest Reserve, Guyana. Mycologia 94(2): 210-220. Cannon PF, Damm U, Johnston PR, Weir BS. 2012. Colletotrichum - current
19
Chlebicki A. 2009. Some endophytes of Juncus trifidus from Tatra Mts. in Poland. Acta Mycol 44(1): 11-17.
Crous PW, Groenewald JZ. 2013. A phylogenetic re-evaluation of Arthrinium.
IMA Fungus 4(1): 133-154.
Davies EC, Franklin JB, Shaw AJ, Vilgalys R. 2003. Endophytic Xylaria
(xylariaceae) among liverworts and angiosperms: phylogenetics, distribution, and symbiosis. Am J Bot 90(11): 1661-1667.
Gerard PR, Husson C, Pinon J, Frey P. 2006. Comparison of Genetic and Virulence Diversity of Melampsora larici-populina Populations on Wild and Cultivated Poplar and Influence of the Alternate Host. APS 96 (9) : 1027-1036.
Glienke C, Pereira OL, Stringari D, Fabris J, Kava-Cordeiro1 V, Galli-Terasawa L, Cunnington J, Shivas RG, Groenewald JZ, Crous PW. 2011. Endophytic and pathogenic Phyllosticta species, with reference to those associated with Citrus Black Spot. Persoonia 26: 47-56.
Gomes RR, Glienke C, Videira SIR, Lombard L, Groenewald JZ, Crous PW. 2013. Diaporthe: a genus of endophytic, saprobic and plant pathogenic fungi. Persoonia 31: 1-41.
Hancock JF. 2005. Contributions of domesticated plant studies to our understanding of plant evolution. Ann Bot 96: 953-963.
He Y, Zhang Z. 2012. Diversity of organism in the Usnea longissima lichen.
African J of Microbiol Res 6: 4797-4804.
Higgins KL, Arnold AE, Miadlikowska J, Sarvate SD, Lutzoni F. 2007. Phylogenetic relationships, host affinity, and geographic structure of boreal and arctic endophytes from three major plant lineages. Mol Phylogenet and Evol 42: 543-555.
Ho MY, Chung WC, Huang HC, Chung WH, Chung WH. 2012. Identification of endophytic fungi of medicinal herbs of lauraceae and Rutaceae with antimicrobial property. Taiwania 57(3) : 229-241.
Houbraken JAMP, Frisvad JC, Samson RA. 2010. Taxonomy of Penicillium citrinum and related species. Fungal Divers 44:117-133.
Huang WY, Cai YZ, Surveswaran S, Hyde KD, Corke H, Sun M. 2009. Molecular phylogenetic identification of endophytic fungi isolated from three Artemisia species. Fungal Divers 36: 69-88.
Hyde KD, Bussaban B, Paulus B, Crous PW, Lee S, McKenzie EHC, Photota W, Lumyong S. 2007. Diversity of saprobic microfungi. Biodivers Conserv
16:7-35.
Khan SA, Hamayun M, Yoon H, Kim HY, Suh SJ, Hwang SK, Kim JM, Lee IJ, Choo YS, Yoon UH, Kong WS, Lee BM, Kim J. 2008. Plant growth promotion and Penicillium citrinum. BMC Microbiol 8:231-241.
Khan R, Shahzad S, Choudhary MI, Khan SA, Ahmad A. 2010. Communities of endophytic fungi in medicinal Plant Withania somnifera. Pakistan J Bot
42(2): 1281-1287.
Kruys A, Castlebury LA. 2012. Molecular phylogeny of Sydowiellaceae-resolving the position of Cainiella. Mycologia 104(2): 419-426.
Li T, Liu MJ, Zhang XT, Zhang HB, Sha T, Zhao ZW. 2011. Improved tolerance of maize (Zea mays L.) to heavy metals by colonization of a dark septate endophyte (DSE) Exophiala pisciphila. Sci of The Tot Env 409 : 1069-1074.
Martin-Shanchez PM, Novakova A, Bastian F, Alabouvette C, Saiz-Jimenez C. 2012. Two new species of the genus Ochroconis, O. lascauxensis, and O. anomala isolated from black stains in Lascaux Cave, France. Fungal Biol
116: 574-589.
Mishra A, Surendra KG, Kumar A, Sharma VK, Verma SK, Kharwar RN, and Sieber TN. 2012. Season and Tissue Type Affect Fungal Endophyte Communities of the Indian Medicinal Plant Tinospora cordifolia More Strongly than Geographic Location. Microb Ecol 64(2): 388-398.
Moricca S, Ginetti B, Ragazzi A. 2012. Species and organ specificity in endophytes colonizing healthy and declining Mediterranean oaks.
Phytopathol Mediterr 51(3): 587-598.
Munkacsi AB, Stoxen S, May G. 2008. Ustilago maydis populations tracked maize through domestication and cultivation in the Americas. Proc Biol Sci 275 :1037-1046.
Okane I, Lumyong S, Nakagiri A, Ito T. 2003. Extensive host range of an endophytic fungus, Guignardia endophyllicola (anamorph: Phyllosticta capitalensis). Mycoscience 44: 353-363.
Okane I, Praset I, Kyoko T, Thomas L, Somsak S, Nigel HJ, Akira N, Wanchern P, Suzuki K. 2008. Study of endophytic Xylariaceae in Thailand: diversity and taxonomy inferred from rDNA sequence analyses with saprobes forming fruit bodies in the field. Mycoscience 49: 359-3772.
Paul NC, Yu SH. 2008. Two species of endophytic Cladosporium in pine trees in Korea. Mycobiology 36(4) : 211-216.
Posada F, Aime M, Peterson SW, Rehner SA, Vega F . 2007. Inoculation of coffee plants with the fungal entomopathogen Beauveria bassiana (Ascomycota: Hypocreales). Mycol Res 111:748-757.
Priyadi H, Gen T, Irma R, Bambang S, Wim IN, Ismail R. 2010. Five hundred plant species in Gunung Halimun Salak National Park, West Java. A checklist including Sundanese names, distribution and use. Bogor: CIFOR. Rodriguez RJ, White Jr JF, Arnold AE, Redman RS. 2009. Fungal endophytes:
diversity and functional roles. New Phytol 182: 314-330.
Saunders M, Glenn AE, Kohn LM. 2010. Exploring the evolutionary ecology of fungal endophytes in agricultural systems: using functional traits to reveal mechanisms in community processes. Evol Appl. 3: 525-537.
Schardl CL, Leuchtmann A, Spiering MJ. 2004. Symbioses of grasses with seed borne fungal endophytes. Annu Rev Plant Biol 55: 315-340.
Schulz B, Boyle C. 2005. The Endophytic continuum. Mycol Res109:661-668 Seyedmousavi S, Badali H, Chlebicki A, Zhao J, Prenafeta-Blod FX, De Hoog
GS. 2011. Exophiala sideris, a novel black yeast isolated from environments polluted with toxic alkyl benzenes and arsenic. Fungal Biol
115 : 1030-1037.
Silva M, Pereira OL, Braga IF, Leli SM. 2008. Leaf and pseudobulb diseases on
Bifrenaria harrisoniae (Orchidaceae) caused by Phyllosticta capitalensis
21
Stukenbrock EH, McDonald BA.2008. The origins of plant pathogens in agro-ecosystems. Ann Rev Phytopathol 46:75-100.
Sugijanto NE, Dorra BL. 2012. Aktivitas antimikroba fraksi-fraksi dari ekstrak jamur endofit Cladosporium oxysporum yang diisolasi dari Aglaia odorata lour. Ind J of Medicine 4(1): 2012.
Suh MK, Lee HC, Kim DM, Ha GY, Choi JS. 2012. Molecular phylogenetics of
Exophiala Species isolated from Korea. Ann Dermatol 24(3): 2012. Sun JQ, Guo LD, Zang W, Ping W, Chi DF, 2008. Diversity and ecological
distribution of endophytic fungi associated with medicinal plants. Sci China Ser C 51: 751-759.
Suryanarayanan TS, Senthilarasu G, Muruganandam V. 2000. Endophytic fungi from Cuscuta reflexa and its host plants. Fungal Divers 4: 117-123.
Tamura K, Peterson D, Peterson N, Stecher G, Nei M, Kumar S. 2011. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol Biol Evol
28:2731-2739.
Thomas CS, Marios JJ, English JT. 1988. The effect of wind speed, temperature, and relative humidity, on development of aerial mycelium and conidia of
Botrytis cinerea on grape. Phytopathology 78:260-265.
Thompson SM, Tan YP, Young AJ, Neate1 SM, Aitken EAB, Shivas RG. 2011. Stem cankers on sunflower (Helianthus annuus) in Australia reveal a complex of pathogenic Diaporthe (Phomopsis) species. Persoonia 27: 80-89.
Toledo-HernándezC, Zuluaga-Montero A, Bones-González, Rodríguez JA, Sabat AM, Bayman P. 2008. Fungi in healthy and diseased sea fans (Gorgonia ventalina): is Aspergillus sydowii always the pathogen? Coral Reefs
[Internet].[diunduh 2014 Jul 14]; 27 (2): 707-714. Tersedia pada : http://link.springer.com/article/10.1007/s00338-008-0387-2.
White TJ, Bruns T, Lee S, Taylor J. 1990. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In: Innis MA, Gelfand DH, Sninsky JJ, White TJ, editors. PCR Protocols: A Guide to Methods and Applications. New York: Academic Press. p 315-322.
Wilberforce EM, Boddyc L, Griffiths R, Griffith GW.2003.Agricultural management affects communities of culturable root-endophytic fungi in temperate grasslands. Soil Biol Biochem 35: 1143-1154.
23
Lampiran 1 Pohon filogenetik Arthrinium malaysianum dengan Guignardia citricarpa sebagai outgroup (clade ditunjukkan dengan garis tebal) sekuen ITS dibuat berdasarkan analisis ML dengan model Kimura2+G (1000x ulangan bootstrap). Hanya bootstrap bernilai lebih dari 50% yang ditampilkan.
25
Clade cliviae
27
Clade boninense Clade gleosporioides
Lampiran 6 Pohon filogenetik (A;B;C) Diaporthe sp., D. gardenia, dan D. anacardii (clade ditunjukkan dengan garis tebal) sekuen ITS+EF dibuat berdasarkan analisis Bayesian dengan 6x106 ulangan bootstrap. Hanya bootstrap bernilai lebih dari 50% yang ditampilkan.
A
29
Lampiran 7 Pohon filogenetik Exophiala spp. dengan Rhinocladiella aquaspersa sebagai outgroup (clade ditunjukkan dengan garis tebal) sekuen ITS dibuat berdasarkan analisis ML dengan model Kimura2+G (1000x ulangan bootstrap). Hanya bootstrap bernilai lebih dari 50% yang ditampilkan.
Lampiran 11 Pohon filogenetik Pyriculariacostina dengan Tumularia aquatic sebagai outgroup (clade ditunjukkan dengan garis tebal) sekuen ITS dibuat berdasarkan analisis ML dengan model Kimura2+G (1000x ulangan bootstrap). Hanya bootstrap bernilai lebih dari 50% yang ditampilkan.
Lampiran 12 Pohon filogenetik Sydowiellaceae dengan Pyricularia parasitica sebagai outgroup, analisis ML dengan model Kimura2+G dan 1000x ulangan bootstrap. Hanya bootstrap bernilai lebih Lampiran 9 Pohon filogenetik Ochroconis gallopava dengan Aspergillus tabanicus sebagai outgroup (clade
ditunjukkan dengan garis tebal) sekuen ITS dibuat berdasarkan analisis ML dengan model Kimura2+G (1000x ulangan bootstrap). Hanya bootstrap bernilai lebih dari 50% yang ditampilkan.
31
RIWAYAT HIDUP
Ivan Permana Putra lahir pada 10 September 1989 di Manggar-Belitung. Penulis merupakan anak kedua dari ayah Wahyu Setyawan dan Ibu Hartini. Pendidikan SMA diselesaikan tahun 2007 di SMAN I Pemali-Bangka, sementara pendidikan S1 ditempuh pada Jurusan Biologi IPB dan lulus pada tahun 2011 dengan predikat cum laude. Selama menyelesaikan studi S1 penulis aktif sebagai asisten praktikum berbagai mata kuliah, program kreatifitas mahasiswa (PKM), himpunan mahasiswa biologi (HIMABIO) IPB, serta unit kegiatan mahasiswa (UKM) sepak bola IPB.