SKRIPSI
MEMPELAJARI PEMURNIAN ENZIM KITOSANASE TERMOSTABIL DARI ISOLAT Bacillus licheniformis MB-2
ASAL TOMPASO, MANADO, SULAWESI UTARA
Oleh :
DIAN RACHMADANI F24103049
2007
MEMPELAJARI PEMURNIAN ENZIM KITOSANASE TERMOSTABIL DARI ISOLAT Bacillus licheniformis MB-2
ASAL TOMPASO, MANADO, SULAWESI UTARA
Oleh :
DIAN RACHMADANI F24103049
SKRIPSI
Sebagai salah satu syarat untuk memperoleh gelar SARJANA TEKNOLOGI PERTANIAN Pada Departemen Ilmu dan Teknologi Pangan
Fakultas Teknologi Pertanian Institut Pertanian Bogor
2007
DEPARTEMEN ILMU DAN TEKNOLOGI PANGAN FAKULTAS TEKNOLOGI PERTANIAN
INSTITUT PERTANIAN BOGOR FAKULTAS TEKNOLOGI PERTANIAN
MEMPELAJARI PEMURNIAN ENZIM KITOSANASE TERMOSTABIL DARI ISOLAT Bacillus licheniformis MB-2
ASAL TOMPASO, MANADO, SULAWESI UTARA
SKRIPSI
Sebagai salah satu syarat untuk memperoleh gelar SARJANA TEKNOLOGI PERTANIAN Pada Departemen Ilmu dan Teknologi Pangan
Fakultas Teknologi Pertanian Institut Pertanian Bogor
oleh
DIAN RACHMADANI F24103049
Dilahirkan pada tanggal 18 Juni 1985 Di Bogor, Jawa Barat
Tanggal lulus : 8 Agustus 2007
Menyetujui Bogor, Agustus 2007
Prof. Dr. Ir. Maggy T. Suhartono Dosen Pembimbing
Dian Rachmadani. F24103049. Mempelajari Pemurnian Enzim Kitosanase Termostabil Dari Isolat Bacillus licheniformis MB-2 Asal Tompaso, Manado, Sulawesi Utara. Di bawah bimbingan : Prof. Dr. Ir. Maggy Thenawijaya Suhartono
ABSTRAK
Enzim kitosanase adalah enzim yang dapat menghidrolisis kitosan menjadi oligomernya. Enzim ini dapat dihasilkan oleh bakteri, fungi, dan tanaman. Kegunaan kitosan dan turunannya dalam berbagai bidang mendorong eksplorasi enzim pendegradasi kitin dan kitosan dalam jumlah besar. Penelitian ini bertujuan untuk mempelajari produksi dan pemurnian enzim kitosanase menggunakan kromatografi filtrasi gel dari isolat Bacillus licheniformis MB-2 asal Tompaso, Manado, Sulawesi Utara serta melakukan karakterisasi beberapa sifat enzim kitosanase yang dihasilkan.
Isolat Bacillus licheniformis MB-2 diinokulasi ke dalam media thermus padat (1.0% koloidal kitosan, 0.7% K2HPO4, 0.3% KH2PO4, 0.5% MgSO4, 0.25%
ekstrak khamir, 0.25% casiton, 1.5% bacto agar, dan 0.4% gelrite) lalu diinkubasi pada 550C. Setelah 5 hari terbentuk areal bening di sekitar koloni bakteri. Selanjutnya, koloni bakteri tersebut diinokulasi ke dalam media thermus cair (0.4% koloidal kitosan, 0.7% K2HPO4, 0.3% KH2PO4, 0.5% MgSO4, 0.25%
ekstrak khamir, dan 0.25% casiton) lalu diinkubasi pada shaker waterbath suhu 550C selama 24 jam dengan kecepatan agitasi 120 rpm dan digunakan sebagai kultur starter. Enzim kitosanase diproduksi dengan memasukkan kultur starter ke dalam media thermus cair lalu diinkubasi pada shaker waterbath suhu 550C selama 7 hari dengan kecepatan agitasi 120 rpm. Pada akhir inkubasi, sel bakteri dan sisa-sisa media yang tidak larut dipisahkan dengan sentrifugasi (40C, 20 menit, 10000 rpm). Supernatan enzim bebas sel yang diperoleh kemudian diendapkan dengan ammonium sulfat 80%. Endapan protein dipisahkan dengan sentrifugasi (40C, 20 menit, 10000 rpm) lalu dilarutkan dalam 0.05 M buffer fosfat pH 6 sehingga diperoleh enzim kasar (crude enzyme). Dialisis dilakukan terhadap crude enzyme untuk menghilangkan molekul garam. Enzim hasil dialisis kemudian di-freeze dry dan selanjutnya digunakan untuk pemurnian. Pemurnian dilakukan dengan teknik kromatografi filtrasi gel menggunakan matriks Sephadex G-100. Kolom disetimbangkan dan dielusi dengan 0.05 M buffer fosfat pH 6.
Aktivitas enzim kitosanase diuji dengan menentukan jumlah gula pereduksi (D-glukosamin) yang terbentuk karena reaksi hidrolisis substrat soluble chitosan oleh enzim kitosanase. Pengujian dilakukan dengan teknik spektrofotometri pada panjang gelombang 420 nm. Satu unit aktivitas enzim kitosanase didefinisikan sebagai jumlah enzim yang memproduksi 1 µmol gula pereduksi (glukosamin) per menit pada kondisi tertentu.
700C adalah 0.0003/menit dan 0.0024/menit pada 900C. Waktu paruh (t1/2) pada
700C adalah 2310.5 menit dan 288.8 menit pada 900C. Energi deaktivasi (Ea) fraksi 44 adalah 25.823 kkal/gmol.0K. Pengujian pengaruh pH terhadap stabilitas enzim menunjukkan bahwa enzim murni fraksi 44 stabil pada pH 6 selama 2 jam dengan suhu inkubasi 700C (98%).
Berdasarkan hasil SDS-PAGE terhadap crude enzyme diketahui ada 5 pita protein dengan berat molekul 61.5, 48.5, 40.6, 28.5, dan 19.9 kDa. Hasil analisis terhadap enzim murni menunjukkan bahwa enzim murni fraksi 44 memiliki berat molekul sebesar 75.4 kDa.
i KATA PENGANTAR
Puji dan syukur penulis panjatkan pada Allah SWT karena atas ridho-Nya, penulisan skripsi ini dapat selesai tepat pada waktunya. Skripsi dengan judul Mempelajari Pemurnian Enzim Kitosanase Termostabil Dari Isolat Bacillus licheniformis MB-2 Asal Tompaso, Manado, Sulawesi Utara disusun berdasarkan
penelitian yang telah dilaksanakan pada bulan November 2006-Juni 2007 di Laboratorium Mikrobiologi dan Biokimia PAU-IPB.
Pada kesempatan ini, penulis mengucapkan terima kasih kepada :
1. Prof. Dr. Ir. Maggy T. Suhartono sebagai dosen pembimbing yang telah memberikan bimbingan dan dukungan kepada penulis selama melakukan tugas akhir sampai terselesaikannya skripsi ini
2. Ir. Sutrisno Koswara, MSi. selaku dosen penguji yang telah bersedia menguji pada sidang sarjana penulis, mengkoreksi, dan memberikan masukan dalam penyusunan skripsi ini
3. Dr. Sukarno selaku dosen penguji yang telah bersedia menguji pada sidang sarjana penulis, mengkoreksi, dan memberikan masukan dalam penyusunan skripsi ini
4. Keluarga tercinta yang telah memberikan keceriaan, dukungan, dan doa pada penulis. Specially for my mother ”thank you very much”
5. My best friend forever, Ira Susanti yang selalu memberikan dukungan dan mendengarkan setiap keluh kesah penulis
6. Evanda Puspita yang selalu ada dalam susah dan senang selama penulis melakukan penelitian
7. Kardita (Rucit, Anis, Oca, Irma, Dini, Iin, Indah, Wati, Abdy, Riska) yang memberikan dukungan kepada penulis
8. Teman-temanku (Rina, Tuti, Jengye, Andiny, Dion) atas dukungan dan keceriaan selama ini
9. Adit, Fajar, Doni, Eka, Rika, Aji atas bantuannya selama penulis melakukan penelitian
11.Teman-teman TPG 40 atas dukungan, kenangan indah, dan keceriaan selama di ITP
12.Ikhsan Novembrianto atas segala bantuan dan dukungannya selama ini kepada penulis
Penulis menyadari bahwa skripsi ini masih banyak kekurangannya. Oleh karena itu, penulis mengharapkan kritik dan saran dari pembaca. Semoga skripsi ini bermanfaat bagi pihak-pihak yang membutuhkan.
Bogor, Agustus 2007
RIWAYAT HIDUP
DAFTAR ISI
KATA PENGANTAR ... i
DAFTAR ISI ... iii
DAFTAR GAMBAR ... v
DAFTAR TABEL ... vi
DAFTAR LAMPIRAN ... vii
I. PENDAHULUAN ... 1
A. Latar Belakang ... 1
B. Tujuan Penelitian ... 2
C. Manfaat Penelitian ... 2
II. TINJAUAN PUSTAKA ... 3
A. Kitin ... 3
B. Kitosan ... 3
C. Enzim Kitosanase ... 6
D. Bacillus licheniformis MB-2 ... 7
E. Pemurnian Enzim ... 8
F. Elektroforesis SDS-PAGE ... 13
III.METODOLOGI PENELITIAN ... 17
A. Bahan dan Alat ... 17
B. Metode ... 17
1. Pembuatan Tepung Kitosan ... 17
2. Pembuatan Koloidal Kitosan ... 18
3. Penyegaran Kultur ... 18
4. Pembuatan Kultur Starter ... 18
5. Produksi Enzim Kitosanase... 18
6. Dialisis ... 19
7. Freeze Dry ... 19
8. Pemurnian Enzim Kitosanase ... 20
9. Elektroforesis SDS-PAGE ... 20
10.Karakterisasi Enzim Kitosanase ... 22
iv
12.Pengukuran Konsentrasi Protein ... 24
IV.HASIL DAN PEMBAHASAN ... 25
A. Produksi Enzim Kitosanase... 25
B. Presipitasi Protein... 27
C. Dialisis ... 28
D. Pemurnian Enzim Kitosanase ... 29
E. Karakterisasi Enzim Kitosanase ... 31
F. Penentuan Berat Molekul Enzim Kitosanase ... 38
V. KESIMPULAN DAN SARAN ... 40
A. Kesimpulan ... 40
B. Saran ... 40
DAFTAR PUSTAKA ... 42
SKRIPSI
MEMPELAJARI PEMURNIAN ENZIM KITOSANASE TERMOSTABIL DARI ISOLAT Bacillus licheniformis MB-2
ASAL TOMPASO, MANADO, SULAWESI UTARA
Oleh :
DIAN RACHMADANI F24103049
2007
MEMPELAJARI PEMURNIAN ENZIM KITOSANASE TERMOSTABIL DARI ISOLAT Bacillus licheniformis MB-2
ASAL TOMPASO, MANADO, SULAWESI UTARA
Oleh :
DIAN RACHMADANI F24103049
SKRIPSI
Sebagai salah satu syarat untuk memperoleh gelar SARJANA TEKNOLOGI PERTANIAN Pada Departemen Ilmu dan Teknologi Pangan
Fakultas Teknologi Pertanian Institut Pertanian Bogor
2007
DEPARTEMEN ILMU DAN TEKNOLOGI PANGAN FAKULTAS TEKNOLOGI PERTANIAN
INSTITUT PERTANIAN BOGOR FAKULTAS TEKNOLOGI PERTANIAN
MEMPELAJARI PEMURNIAN ENZIM KITOSANASE TERMOSTABIL DARI ISOLAT Bacillus licheniformis MB-2
ASAL TOMPASO, MANADO, SULAWESI UTARA
SKRIPSI
Sebagai salah satu syarat untuk memperoleh gelar SARJANA TEKNOLOGI PERTANIAN Pada Departemen Ilmu dan Teknologi Pangan
Fakultas Teknologi Pertanian Institut Pertanian Bogor
oleh
DIAN RACHMADANI F24103049
Dilahirkan pada tanggal 18 Juni 1985 Di Bogor, Jawa Barat
Tanggal lulus : 8 Agustus 2007
Menyetujui Bogor, Agustus 2007
Prof. Dr. Ir. Maggy T. Suhartono Dosen Pembimbing
Dian Rachmadani. F24103049. Mempelajari Pemurnian Enzim Kitosanase Termostabil Dari Isolat Bacillus licheniformis MB-2 Asal Tompaso, Manado, Sulawesi Utara. Di bawah bimbingan : Prof. Dr. Ir. Maggy Thenawijaya Suhartono
ABSTRAK
Enzim kitosanase adalah enzim yang dapat menghidrolisis kitosan menjadi oligomernya. Enzim ini dapat dihasilkan oleh bakteri, fungi, dan tanaman. Kegunaan kitosan dan turunannya dalam berbagai bidang mendorong eksplorasi enzim pendegradasi kitin dan kitosan dalam jumlah besar. Penelitian ini bertujuan untuk mempelajari produksi dan pemurnian enzim kitosanase menggunakan kromatografi filtrasi gel dari isolat Bacillus licheniformis MB-2 asal Tompaso, Manado, Sulawesi Utara serta melakukan karakterisasi beberapa sifat enzim kitosanase yang dihasilkan.
Isolat Bacillus licheniformis MB-2 diinokulasi ke dalam media thermus padat (1.0% koloidal kitosan, 0.7% K2HPO4, 0.3% KH2PO4, 0.5% MgSO4, 0.25%
ekstrak khamir, 0.25% casiton, 1.5% bacto agar, dan 0.4% gelrite) lalu diinkubasi pada 550C. Setelah 5 hari terbentuk areal bening di sekitar koloni bakteri. Selanjutnya, koloni bakteri tersebut diinokulasi ke dalam media thermus cair (0.4% koloidal kitosan, 0.7% K2HPO4, 0.3% KH2PO4, 0.5% MgSO4, 0.25%
ekstrak khamir, dan 0.25% casiton) lalu diinkubasi pada shaker waterbath suhu 550C selama 24 jam dengan kecepatan agitasi 120 rpm dan digunakan sebagai kultur starter. Enzim kitosanase diproduksi dengan memasukkan kultur starter ke dalam media thermus cair lalu diinkubasi pada shaker waterbath suhu 550C selama 7 hari dengan kecepatan agitasi 120 rpm. Pada akhir inkubasi, sel bakteri dan sisa-sisa media yang tidak larut dipisahkan dengan sentrifugasi (40C, 20 menit, 10000 rpm). Supernatan enzim bebas sel yang diperoleh kemudian diendapkan dengan ammonium sulfat 80%. Endapan protein dipisahkan dengan sentrifugasi (40C, 20 menit, 10000 rpm) lalu dilarutkan dalam 0.05 M buffer fosfat pH 6 sehingga diperoleh enzim kasar (crude enzyme). Dialisis dilakukan terhadap crude enzyme untuk menghilangkan molekul garam. Enzim hasil dialisis kemudian di-freeze dry dan selanjutnya digunakan untuk pemurnian. Pemurnian dilakukan dengan teknik kromatografi filtrasi gel menggunakan matriks Sephadex G-100. Kolom disetimbangkan dan dielusi dengan 0.05 M buffer fosfat pH 6.
Aktivitas enzim kitosanase diuji dengan menentukan jumlah gula pereduksi (D-glukosamin) yang terbentuk karena reaksi hidrolisis substrat soluble chitosan oleh enzim kitosanase. Pengujian dilakukan dengan teknik spektrofotometri pada panjang gelombang 420 nm. Satu unit aktivitas enzim kitosanase didefinisikan sebagai jumlah enzim yang memproduksi 1 µmol gula pereduksi (glukosamin) per menit pada kondisi tertentu.
700C adalah 0.0003/menit dan 0.0024/menit pada 900C. Waktu paruh (t1/2) pada
700C adalah 2310.5 menit dan 288.8 menit pada 900C. Energi deaktivasi (Ea) fraksi 44 adalah 25.823 kkal/gmol.0K. Pengujian pengaruh pH terhadap stabilitas enzim menunjukkan bahwa enzim murni fraksi 44 stabil pada pH 6 selama 2 jam dengan suhu inkubasi 700C (98%).
Berdasarkan hasil SDS-PAGE terhadap crude enzyme diketahui ada 5 pita protein dengan berat molekul 61.5, 48.5, 40.6, 28.5, dan 19.9 kDa. Hasil analisis terhadap enzim murni menunjukkan bahwa enzim murni fraksi 44 memiliki berat molekul sebesar 75.4 kDa.
i KATA PENGANTAR
Puji dan syukur penulis panjatkan pada Allah SWT karena atas ridho-Nya, penulisan skripsi ini dapat selesai tepat pada waktunya. Skripsi dengan judul Mempelajari Pemurnian Enzim Kitosanase Termostabil Dari Isolat Bacillus licheniformis MB-2 Asal Tompaso, Manado, Sulawesi Utara disusun berdasarkan
penelitian yang telah dilaksanakan pada bulan November 2006-Juni 2007 di Laboratorium Mikrobiologi dan Biokimia PAU-IPB.
Pada kesempatan ini, penulis mengucapkan terima kasih kepada :
1. Prof. Dr. Ir. Maggy T. Suhartono sebagai dosen pembimbing yang telah memberikan bimbingan dan dukungan kepada penulis selama melakukan tugas akhir sampai terselesaikannya skripsi ini
2. Ir. Sutrisno Koswara, MSi. selaku dosen penguji yang telah bersedia menguji pada sidang sarjana penulis, mengkoreksi, dan memberikan masukan dalam penyusunan skripsi ini
3. Dr. Sukarno selaku dosen penguji yang telah bersedia menguji pada sidang sarjana penulis, mengkoreksi, dan memberikan masukan dalam penyusunan skripsi ini
4. Keluarga tercinta yang telah memberikan keceriaan, dukungan, dan doa pada penulis. Specially for my mother ”thank you very much”
5. My best friend forever, Ira Susanti yang selalu memberikan dukungan dan mendengarkan setiap keluh kesah penulis
6. Evanda Puspita yang selalu ada dalam susah dan senang selama penulis melakukan penelitian
7. Kardita (Rucit, Anis, Oca, Irma, Dini, Iin, Indah, Wati, Abdy, Riska) yang memberikan dukungan kepada penulis
8. Teman-temanku (Rina, Tuti, Jengye, Andiny, Dion) atas dukungan dan keceriaan selama ini
9. Adit, Fajar, Doni, Eka, Rika, Aji atas bantuannya selama penulis melakukan penelitian
11.Teman-teman TPG 40 atas dukungan, kenangan indah, dan keceriaan selama di ITP
12.Ikhsan Novembrianto atas segala bantuan dan dukungannya selama ini kepada penulis
Penulis menyadari bahwa skripsi ini masih banyak kekurangannya. Oleh karena itu, penulis mengharapkan kritik dan saran dari pembaca. Semoga skripsi ini bermanfaat bagi pihak-pihak yang membutuhkan.
Bogor, Agustus 2007
RIWAYAT HIDUP
DAFTAR ISI
KATA PENGANTAR ... i
DAFTAR ISI ... iii
DAFTAR GAMBAR ... v
DAFTAR TABEL ... vi
DAFTAR LAMPIRAN ... vii
I. PENDAHULUAN ... 1
A. Latar Belakang ... 1
B. Tujuan Penelitian ... 2
C. Manfaat Penelitian ... 2
II. TINJAUAN PUSTAKA ... 3
A. Kitin ... 3
B. Kitosan ... 3
C. Enzim Kitosanase ... 6
D. Bacillus licheniformis MB-2 ... 7
E. Pemurnian Enzim ... 8
F. Elektroforesis SDS-PAGE ... 13
III.METODOLOGI PENELITIAN ... 17
A. Bahan dan Alat ... 17
B. Metode ... 17
1. Pembuatan Tepung Kitosan ... 17
2. Pembuatan Koloidal Kitosan ... 18
3. Penyegaran Kultur ... 18
4. Pembuatan Kultur Starter ... 18
5. Produksi Enzim Kitosanase... 18
6. Dialisis ... 19
7. Freeze Dry ... 19
8. Pemurnian Enzim Kitosanase ... 20
9. Elektroforesis SDS-PAGE ... 20
10.Karakterisasi Enzim Kitosanase ... 22
iv
12.Pengukuran Konsentrasi Protein ... 24
IV.HASIL DAN PEMBAHASAN ... 25
A. Produksi Enzim Kitosanase... 25
B. Presipitasi Protein... 27
C. Dialisis ... 28
D. Pemurnian Enzim Kitosanase ... 29
E. Karakterisasi Enzim Kitosanase ... 31
F. Penentuan Berat Molekul Enzim Kitosanase ... 38
V. KESIMPULAN DAN SARAN ... 40
A. Kesimpulan ... 40
B. Saran ... 40
DAFTAR PUSTAKA ... 42
DAFTAR GAMBAR
Gambar 1. Struktur kitin ... 3
Gambar 2. Struktur kitosan ... 4
Gambar 3. Prinsip pemisahan protein menggunakan filtrasi gel ... 13
Gambar 4. Aktivitas kitosanase MB-2 pada media thermus padat ... 26
Gambar 5. Hasil pemurnian kitosanase menggunakan filtrasi gel ... 30
Gambar 6. Pengaruh suhu terhadap aktivitas crude enzyme ... 31
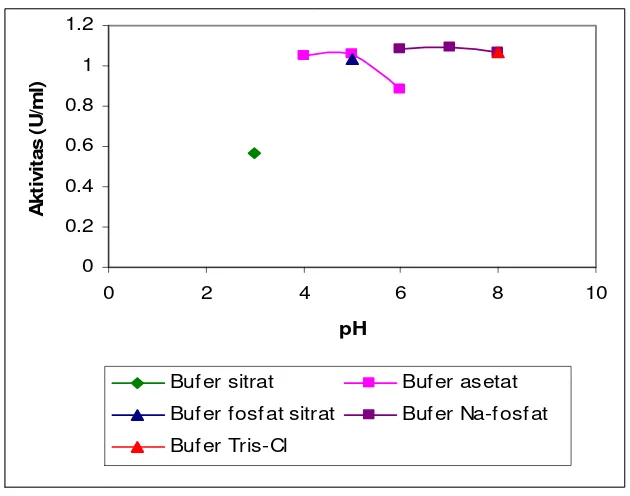
Gambar 7. Pengaruh pH terhadap aktivitas crude enzyme ... 33
Gambar 8. Pengaruh suhu terhadap aktivitas enzim murni ... 34
Gambar 9. Pengaruh pH terhadap aktivitas enzim murni ... 35
Gambar 10. Hubungan ln aktivitas enzim terhadap waktu pemanasan ... 36
Gambar 11. Hubungan ln k terhadap suhu pemanasan ... 37
Gambar 12. Stabilitas enzim murni terhadap panas ... 37
Gambar 13. Stabilitas enzim murni terhadap pH ... 38
vi DAFTAR TABEL
Tabel 1. Morfologi dan karakteristik biokimia B. licheniformis MB-2 ... 8
Tabel 2. Komposisi gel SDS-PAGE ... 20 Tabel 3. Pengukuran aktivitas enzim kitosanase ... 23
DAFTAR LAMPIRAN
Lampiran 1. Pembuatan soluble chitosan ... 47
Lampiran 2. Kurva standar glukosamin ... 48 Lampiran 3. Kurva standar BSA ... 49
Lampiran 4. Kurva standar SDS-PAGE... 50 Lampiran 5. Aktivitas kitosanase pada supernatan, crude enzyme, dialisis ... 51 Lampiran 6. Konsentrasi protein pada supernatan, crude enzyme, dialisis ... 52 Lampiran 7. Aktivitas kitosanase hasil pemurnian filtrasi gel ... 53 Lampiran 8. Konsentrasi protein pada enzim hasil pemurnian ... 59 Lampiran 9. Pengaruh suhu terhadap aktivitas crude enzyme ... 61 Lampiran 10. Pengaruh pH terhadap aktivitas crude enzyme ... 62 Lampiran 11. Pengaruh suhu terhadap aktivitas enzim murni ... 63 Lampiran 12. Pengaruh pH terhadap aktivitas enzim murni ... 64 Lampiran 13. Stabilitas enzim murni terhadap pemanasan 700C ... 65 Lampiran 14. Stabilitas enzim murni terhadap pemanasan 900C ... 66 Lampiran 15. Stabilitas enzim murni pada pH 6 suhu inkubasi 700C ... 67
1
BAB I PENDAHULUAN
A. Latar Belakang
Sebagai negara yang memiliki kekayaan hayati tinggi, Indonesia
merupakan salah satu habitat bagi mikroorganisme termofilik penghasil enzim
kitosanase. Mikroorganisme termofilik banyak ditemukan di daerah-daerah
ekstrim seperti kawasan sumber air panas, daerah geothermal, dan sebagainya.
Kawasan-kawasan tersebut merupakan wilayah Indonesia yang belum banyak
digali potensinya.
Isolasi oleh Suhartono (2002) dilakukan dengan mengambil sampel yang
berasal dari 4 lokasi yang berbeda di Sulawesi Utara terutama dalam hal
keasamannya. Tompaso mewakili daerah agak asam (pH 4-7), Lahendong
mewakili daerah asam (pH 1-4), Karimenga mewakili daerah netral (pH 7),
dan Likupang mewakili daerah agak basa (pH 8). Bakteri termofilik berhasil
dikulturkan dari sampel asal Tompaso dan Lahendong. Tompaso merupakan
daerah sumber air panas, sedangkan Lahendong merupakan daerah
geothermal. Di antara 484 isolat bakteri termofilik asal Tompaso, hanya 41
koloni saja (sekitar 10%) yang menunjukkan aktivitas kitinolitik. Di antara
isolat bakteri kitinolitik tersebut, isolat Bacillus licheniformis MB-2
merupakan isolat dengan indeks kitinolitik yang tinggi (lebih dari 1.5), yaitu
2.25 (Welan, 2001). Isolat Bacillus licheniformis MB-2 selanjutnya digunakan
untuk memproduksi enzim kitosanase ekstraseluler.
Enzim kitosanase adalah enzim yang dapat menghidrolisis kitosan, yaitu
suatu polisakarida rantai lurus yang tersusun atas residu D-glukosamin
berikatan β-(1,4). Reaksi ini akan menghasilkan oligomer kitosan
(kitooligosakarida). Perkembangan aplikasi komersial kitosan meningkat
secara cepat dalam tahun-tahun belakangan ini. Kitosan berguna sebagai
wound healing, surgical suture, contact lenses, penurun kolesterol, fiber dan
penurun berat badan, preservative, kosmetik, penghilangan ion metal pada
limbah cair, pemurnian air, dan sebagainya. Kitosan memiliki berat molekul
Pengembangan aplikasi oligomer kitosan pada saat ini dikarenakan sifatnya
yang dapat larut dalam air dan memiliki sifat fisiologis yang lebih baik
dibandingkan kitosan sehingga aplikasinya lebih luas (Goosen, 1997).
Manfaat yang diperoleh dari oligomer kitosan antara lain mengikat kelebihan
lemak dan menghambat penyerapan lemak, menurunkan LDL dan
meningkatkan HDL, anti kanker, anti bakteri, menurunkan gula darah,
mengontrol tekanan darah, mencegah konstipasi, meningkatkan penyerapan
kalsium, prebiotic health food, dan sebagainya. Wahyuni (2006) melaporkan
bahwa enzim kitosanase dari Bacillus licheniformis MB-2 dapat digunakan
untuk memproduksi oligomer kitosan yang memiliki aktivitas terhadap sel
limfosit dan sel kanker. Aktivitas anti mikroba oligomer kitosan telah
dilaporkan oleh Meidina (2005) yang dihasilkan dengan mengunakan enzim
kitosanase dari Bacillus licheniformis MB-2.
Mikroorganisme termofilik berpotensi sebagai penghasil enzim
termostabil. Pada beberapa dekade, enzim termostabil telah lama diminati oleh
kalangan industri. Kebutuhan akan enzim termostabil didorong karena enzim
tersebut memiliki nilai ekonomis yang tinggi antara lain reaksi akan
berlangsung pada suhu tinggi sehingga mengurangi kontaminasi bakteri
mesofilik dan laju reaksi lebih cepat sehingga mengurangi biaya produksi
(Suhartono, 1989).
B. Tujuan Penelitian
Penelitian ini bertujuan untuk mempelajari produksi dan pemurnian enzim
kitosanase menggunakan kromatografi filtrasi gel dari isolat Bacillus
licheniformis MB-2 asal Tompaso, Manado, Sulawesi Utara serta melakukan
beberapa karakterisasi terhadap enzim kitosanase yang dihasilkan.
C. Manfaat Penelitian
Setelah dilakukan penelitian ini diharapkan diperoleh informasi yang
cukup mengenai produksi, pemurnian dengan filtrasi gel, dan karakteristik
enzim kitosanase guna pemanfaatan enzim untuk memproduksi oligomer
3
BAB II
TINJAUAN PUSTAKA
A. Kitin
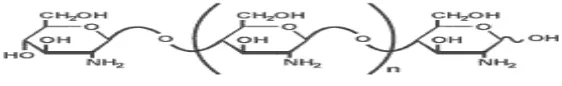
Kitin merupakan biopolimer rantai lurus yang disusun oleh
monomer-monomer N-asetilglukosamin yang dihubungkan oleh ikatan β-(1,4). Kitin
merupakan biomassa yang berlimpah di alam, yaitu nomor dua setelah
selulosa. Kitin mempunyai struktur kimia yang mirip dengan selulosa, yaitu
suatu polisakarida yang disusun oleh molekul-molekul gula sederhana yang
identik. Struktur kitin dapat dilihat pada gambar 1. Kitin ditemukan sebagai
komponen utama pada kulit kepiting; udang; atau kelompok kerang-kerangan
(crustacea), serangga, dan dinding sel beberapa mikroorganisme (Goosen,
1997). Kitin paling banyak terdapat pada kulit golongan crustacea dengan
kandungan kitin mencapai 40-60% (Angka dan Suhartono, 2002).
Gambar 1. Struktur kitin
Kitin tidak larut dalam air, asam anorganik encer, asam organik, dan
pelarut organik. Namun, kitin dapat larut dalam N,N-dimetilasetamida yang
mengandung LiCl2, asam pekat seperti H2SO4, asam fosfat, dan asam format
anhidrida (Goosen, 1997). Kitin dapat dimanfaatkan sebagai anti fungi untuk
melindungi tanaman dari serangan fungi dan anti bakteri terhadap beberapa
patogen (Shahidi et al., 1999).
B. Kitosan
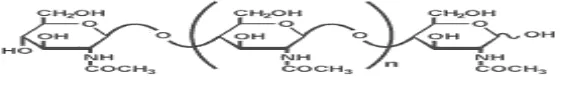
Kitosan merupakan biopolimer yang tersusun atas D-glukosamin dengan
ikatan glikosidik 1→4. Struktur kitosan dapat dilihat pada gambar 2. Kitosan
untuk penggunaan komersial dan penelitian diperoleh dari proses deasetilasi
dari fungi golongan Zygomycetes (Miyoshi et al., 1992). Kitin dan kitosan
memiliki sifat ramah lingkungan dan tidak beracun.
Gambar 2. Struktur kitosan
Kitosan yang merupakan turunan dari kitin dapat diperoleh dari deasetilasi
sempurna atau sebagian. Deasetilasi kitin akan menghilangkan gugus asetil
dan menyisakan gugus amin yang bermuatan positif sehingga kitosan bersifat
polikationik. Biopolimer ini disusun oleh 2 jenis gula amino, yaitu glukosamin
(2-amino-2-deoksi-D-glukosa, 70-80%) dan N-asetilglukosamin
(2-asetamino-2-deoksi-D-glukosa, 20-30%) (Goosen, 1997).
Proses deasetilasi kitin menjadi kitosan dapat dilakukan secara kimia
maupun enzimatis. Kedua reaksi tersebut bertujuan untuk menghilangkan
gugus asetil yang terdapat pada kitin. Proses deasetilasi secara kimia
dilakukan dengan mengkombinasikan perlakuan panas (60-1400C) dan
perendaman dalam larutan alkali (NaOH 30-50%). Perendaman dalam larutan
alkali dilakukan terhadap kitin dalam bentuk tepung. Penepungan dilakukan
agar proses deasetilasi dapat berlangsung lebih cepat dan sempurna karena
semakin luasnya permukaan yang dapat diakses oleh larutan alkali.
Konsentrasi NaOH akan mempengaruhi laju deasetilasi. Semakin tinggi
konsentrasi NaOH, laju deasetilasi akan semakin cepat. Menurut Kolodziejska
et al. (2000), kitin yang direndam dalam NaOH 50% lalu dipanaskan pada
1050C selama 1 jam akan menghasilkan derajat deasetilasi sebesar 87%. Pada
umumnya, derajat deasetilasi kitosan berkisar antara 70-90% tergantung
metode yang digunakan (Goosen, 1997). Derajat deasetilasi akan meningkat
dengan meningkatnya suhu atau konsentrasi NaOH. Proses deasetilasi secara
kimia dalam banyak hal tidak menguntungkan karena prosesnya tidak mudah
dikendalikan, tidak ramah lingkungan, dan kitosan yang dihasilkan memiliki
5 Proses deasetilasi secara enzimatis dilakukan dengan menggunakan enzim
kitin deasetilase (EC 3.5.1.41). Enzim kitin deasetilase diperoleh dari Mucor
rouxii (Kafetzopoulos et al., 1993), Escherichia coli (Tokoyasu et al., 1999),
dan Colletotrichum lindemuthianum (Tokuyasu et al., 1996). Berdasarkan
hasil isolasi dan karakterisasi oleh Suhartono et al. (2002) diketahui bahwa
bakteri termofilik Bacillus sp. K29-14 dari kawah Kamojang mempunyai
aktivitas enzim kitin deasetilase. Dari kelompok khamir, ekstrak kasar
Saccharomyces cerevisiae diketahui mempunyai aktivitas enzim kitin
deasetilase. Selain itu, enzim kitin deasetilase juga diperoleh dari kapang
Absidia coerula dan Aspergillus nidulans (Tsigos et al., 2000). Proses
deasetilasi secara enzimatis yang telah dilakukan hanya memberikan derajat
deasetilasi yang rendah di bawah standar yang biasa ditetapkan untuk hasil
deasetilasi secara kimia. Proses deasetilasi menggunakan kombinasi perlakuan
secara kimia dan enzimatis yang dilaporkan oleh Emmawati (2004) dan
Rochima (2005) merupakan alternatif proses deasetilasi yang lebih baik.
Proses pembuatan kitosan mempengaruhi kualitas dan sifat dari kitosan.
Kualitas dan sifat dari kitosan ditentukan dari derajat deasetilasi, berat
molekul, dan viskositas. Derajat deasetilasi kitosan ditentukan dengan cara
menghitung kandungan amin bebas dari kitosan. Berat molekul kitin
umumnya > 106 Dalton, namun berat molekul kitosan komersial berkisar
antara 0.1-1.2 x 106 Dalton. Metode viskometri merupakan metode yang
sederhana dan cepat untuk menentukan berat molekul kitosan. Viskositas
kitosan dalam larutan dipengaruhi oleh derajat polimer deasetilasi, berat
molekul, konsentrasi, kekuatan ion, pH, dan suhu. Jika suhu meningkat, maka
viskositas kitosan akan menurun. Perubahan pH pada larutan kitosan akan
memberikan hasil yang berbeda tergantung dari tipe asam yang digunakan.
Jika perubahan pH menggunakan asam asetat, maka viskositas kitosan akan
meningkat dengan semakin turunnya pH. Kitosan tidak larut dalam air, alkali,
dan pelarut organik, tetapi kitosan larut dalam asam organik ketika pH larutan
di bawah 6. Asam asetat dan asam formik merupakan jenis asam yang banyak
Hidrolisis kitosan menjadi oligomer kitosan dapat dilakukan secara kimia
maupun enzimatis. Hidrolisis kitosan melalui proses kimia dilakukan dengan
menggunakan larutan asam kuat. Proses ini menghasilkan banyak kerugian
seperti pada prosedur kimia lainnya, yaitu sulit dikontrol, membutuhkan
proses lebih lanjut, menghasilkan produk samping, dan memiliki derajat
polimerisasi yang rendah. Derajat polimerisasi menunjukkan panjang rantai
oligomer yang dihasilkan dari proses hidrolisis. Panjang rantai oligomer
kitosan menentukan berat molekul oligomer tersebut. Oligomer dengan derajat
polimerisasi tinggi (pentamer sampai heptamer) memiliki sifat fungsional
yang lebih baik dibandingkan dengan oligomer dengan derajat polimerisasi
rendah. Oligomer yang dihasilkan dengan hidrolisis asam adalah monomer
sampai trimer dan sebagian kecil tetramer sampai heptamer (Jeon dan Kim,
2000).
Hidrolisis kitosan melalui proses enzimatis merupakan cara yang lebih
baik untuk memperoleh oligomer kitosan dengan derajat polimerisasi tinggi.
Hal ini dikarenakan proses enzimatis bersifat spesifik. Oligomer yang
dihasilkan melalui proses enzimatis memiliki derajat polimerisasi yang tinggi
dan sedikit monomer glukosamin (Jeon dan Kim, 2000). Kitosan dapat
dihidrolisis oleh berbagai enzim. Enzim komersial seperti lipase, selulase, dan
pektinase dapat digunakan untuk menghidrolisis kitosan (Tsai et al., 2000).
Pantaleone et al. (1992) dan Brine et al. (1992) melaporkan glikanase,
protease, lipase, dan tannase yang diperoleh dari bakteri, fungi, mamalia, dan
tanaman dapat digunakan untuk menghidrolisis kitosan. Muzzarelli et al.
(1995a, 1995b) menggunakan papain dan lipase untuk menghidrolisis kitosan.
Penggunaan enzim-enzim tersebut memerlukan konsentrasi enzim yang relatif
tinggi. Enzim lainnya yang dapat digunakan untuk menghidrolisis kitosan
adalah enzim kitosanase. Penggunaan enzim kitosanase menunjukkan aktivitas
yang cukup baik pada konsentrasi enzim yang kecil (Jeon dan Kim, 2000).
C. Enzim Kitosanase
Enzim kitosanase merupakan enzim yang dapat memotong ikatan β-(1,4)
7 digolongkan menjadi 3 kelas berdasarkan spesifik pemotongannya, yaitu (1)
enzim yang memotong pada ikatan GlcN-GlcN dan GlcNAc-GlcN.
Contohnya pada enzim kitosanase dari Streptomyces sp. N174, (2) enzim yang
memotong hanya pada ikatan GlcN-GlcN. Contohnya pada enzim kitosanase
dari Bacillus sp. no. 7-M, (3) enzim yang memotong pada ikatan GlcN-GlcN
dan GlcN-GlcNAc. Contohnya pada enzim kitosanase dari Bacillus circulans
MH-K1 (Saito et al., 1999; Fukamizo dan Brzezinski, 1997).
Enzim kitosanase dapat diperoleh dari bakteri, fungi, dan tanaman.
Sebagian besar bakteri dan fungi menghasilkan enzim kitosanase
ekstraseluler. Enzim kitosanase dari bakteri dapat diperoleh dari Burkholderia
gladioli strain CHB101 (Shimosaka et al., 2000), Matsuebacter
chitosanotabidus 3001 (Park et al., 1999), Myxobacter sp. AL-1 (Reyes dan
Corona, 1997), Enterobacter sp. G-1 (Yamasaki et al., 1993), Bacillus sp.
PI-7S (Seino et al., 1991), Streptomyces sp. N174 (Somashekar dan Joseph,
1996), Acinetobacter sp. strain CHB101 (Shimosaka et al., 1995),
Amycolatopsis sp. CsO-2 (Okajima et al., 1994), Bacillus sp. CK4 (Yoon et
al., 2001), Psedomonas sp. H-14 (Yoshihara et al., 1992), Bacillus cereus S1
(Kurakake et al., 2000), dan Actynomycetes (Brzezinski et al., 1997). Enzim
kitosanase dari fungi dapat diperoleh dari Fusarium solani f. sp. phaseoli
(Shimosaka et al., 1993), dan Aspergillus sp. Y2K (Cheng dan Li, 2000).
Enzim kitosanase dari tanaman dapat diperoleh dari Cucumis sativus, Citrus
sinensis, dan barley (Somashekar dan Joseph, 1996).
D. Bacillus licheniformis MB-2
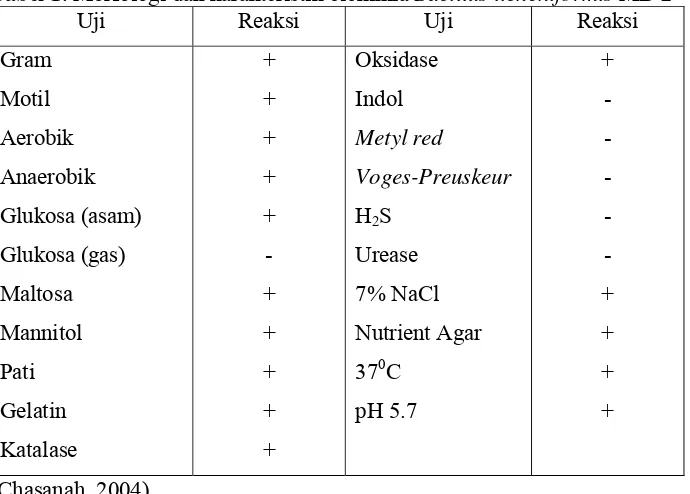
Bacillus licheniformis MB-2 merupakan bakteri Gram positif, berbentuk
batang, bersifat motil, dan dapat tumbuh pada kondisi aerobik maupun
anaerobik. Spora dari bakteri ini berbentuk oval. Bacillus licheniformis MB-2
dapat menggunakan glukosa, maltosa, mannitol, pati, dan gelatin sebagai
sumber karbon. Bakteri ini memberikan hasil yang positif pada uji katalase
dan oksidase, namun memberikan hasil yang negatif pada uji indol, metyl red,
Tabel 1. Morfologi dan karakteristik biokimia Bacillus licheniformis MB-2
Uji Reaksi Uji Reaksi
Gram Motil Aerobik Anaerobik Glukosa (asam) Glukosa (gas) Maltosa Mannitol Pati Gelatin Katalase + + + + + - + + + + + Oksidase Indol Metyl red Voges-Preuskeur
H2S
Urease
7% NaCl
Nutrient Agar
370C
pH 5.7 + - - - - - + + + + (Chasanah, 2004)
E. Pemurnian Enzim 1. Presipitasi Protein
Penambahan senyawa yang hanya menggumpalkan protein dan tidak
menggumpalkan bahan lain akan memisahkan dan lebih memurnikan
protein yang dihasilkan. Tahap ini diistilahkan dengan presipitasi
(Suhartono et al., 1992). Menurut Chaplin dan Bucke (1990), presipitasi
protein merupakan metode yang berguna untuk pemekatan protein dan
sering dilakukan pada tahap awal dari pemurnian enzim.
Presipitasi protein dapat dilakukan dengan beberapa cara antara lain
perubahan pH, penambahan garam, dan penambahan pelarut organik.
Protein akan mengendap jika pH larutan berada pada pH isoelektrik
protein. Garam yang digunakan dalam presipitasi protein dapat berupa
ammonium sulfat, sodium sulfat, dan sebagainya tergantung kepada jenis
protein. Konsentrasi garam yang ditambahkan adalah konsentrasi jenuhnya
(Suhartono et al., 1992). Pemekatan protein dengan menambahkan
ammonium sulfat ke dalam larutan enzim merupakan cara yang banyak
9 Pemekatan protein dengan penambahan ammonium sulfat ke dalam
larutan enzim kitosanase dilakukan pada kejenuhan 30-90%. Hasil yang
diperoleh adalah tingkat kejenuhan 80% merupakan konsentrasi yang
optimum untuk pemekatan protein Bacillus coagulans LH 28.38 (Haliza,
2003). Pemekatan protein Matsuebacter chitosanotabidus 3001 (Park et
al., 1999) dan Enterobacter sp. G-1 (Yamasaki et al., 1993) menggunakan
ammonium sulfat pada tingkat kejenuhan 70%. Pemekatan protein
Acinetobacter sp. strain CHB101 (Shimosaka et al., 1995), Burkholderia
gladioli strain CHB101 (Shimosaka et al., 2000), dan Fusarium solani f.
sp. phaseoli (Shimosaka et al., 1993) menggunakan ammonium sulfat
pada tingkat kejenuhan 80%. Pemekatan protein Bacillus cereus S1
(Kurakake et al., 2000) dan Psedomonas sp. H-14 (Yoshihara et al., 1992)
menggunakan ammonium sulfat pada tingkat kejenuhan 90%. Pemekatan
protein Amycolatopsis sp. CsO-2 (Okajima et al., 1994) dan Bacillus sp.
strain CK4 (Yoon et al., 2001) menggunakan ammonium sulfat bertingkat
dari 30-80%. Menurut Chaplin dan Bucke (1990), beberapa enzim tidak
dapat tahan dalam presipitasi dengan ammonium sulfat. Pemekatan protein
Myxobacter sp. AL-1 menggunakan ZnCl2 (Reyes dan Corona, 1997).
Pemekatan protein Aspergillus sp. Y2K menggunakan evaporator vakum
pada 400C (Cheng dan Li, 2000).
Beberapa keuntungan menggunakan ammonium sulfat antara lain
mudah larut, tidak toksik, murah, dan stabilitasnya terhadap enzim karena
tidak mempengaruhi struktur protein (Webb dan Dixon, 1979). Selain
keuntungan yang diperoleh, penggunaan ammonium sulfat juga
menimbulkan kerugian antara lain konsentrasi garam yang tertinggal
dalam produk tinggi, kurang efisien dalam menghilangkan impuritis, dan
ammonium sulfat tidak bersifat buffer sehingga dapat membebaskan
ammonia yang mengakibatkan kemungkinan penambahan nilai pH
(Suhartono et al., 1992).
Ion garam yang ditambahkan akan mempengaruhi kelarutan protein.
Pada konsentrasi rendah, ion-ion ini akan melingkungi molekul-molekul
melarut. Peristiwa ini disebut salting in. Pada konsentrasi tinggi, terjadi
peningkatan muatan listrik di sekitar protein yang akan menarik mantel air
dari koloid protein. Interaksi hidrofobik di antara sesama molekul protein
pada suasana ionik tinggi akan menurunkan kelarutan protein. Peristiwa
ini disebut salting out (Suhartono et al., 1992).
Penambahan pelarut organik akan menurunkan konstanta dielektrik
dari medium dan akibatnya akan menurunkan kelarutan protein. Beberapa
presipitan yang umum digunakan adalah metanol, etanol, isopropanol, dan
aseton. Penambahan pelarut organik dapat memperbesar kemungkinan
terjadinya denaturasi protein terutama pada suhu yang tinggi. Oleh karena
itu, presipitasi protein menggunakan pelarut organik dilakukan pada suhu
yang rendah (di bawah 00C). Kerugian lain dalam penggunaan pelarut
organik adalah sifatnya yang mudah terbakar dan harganya mahal
(Suhartono et al., 1992).
Etanol merupakan pelarut yang ideal karena adanya keseimbangan
antara efek melarutkan dan sifat hidrofilik yang cukup untuk mengurangi
denaturasi protein. Isopropanol telah digunakan untuk presipitasi berbagai
protein ekstraseluler. Isopropanol memiliki sifat mudah terbakar yang
lebih rendah, tetapi cenderung bersifat hidrofobik. Penggunaan aseton
telah terbukti dapat mempertahankan stabilitas beberapa protein.
Penambahan aseton harus dilakukan secara perlahan-lahan biasanya
melalui sisi samping wadah (Suhartono et al., 1992).
Beberapa protein diendapkan dengan menggunakan suatu polimer
seperti polietilen glikol (PEG). Polimer ini seperti penambahan garam dan
pelarut organik akan menarik molekul air sehingga protein tereksklusi dan
bersatu membentuk gumpalan endapan. Keuntungan penggunaan PEG
antara lain tidak toksik, tidak mudah terbakar, tidak membutuhkan suhu
yang rendah karena polimer ini memberikan efek stabilisasi terhadap
protein, dan mudah diperoleh dengan harga yang relatif murah. Polimer ini
dapat ditambahkan sampai konsentrasi 50% (b/v) dan presipitasi protein
11
2. Kromatografi
Kromatografi menunjukkan proses pemisahan berdasarkan distribusi
diferensial dari komponen sampel di antara 2 fase, yaitu fase stasioner dan
fase bergerak. Komponen yang memiliki interaksi yang kuat dengan fase
stasioner akan bergerak lebih lambat dibandingkan komponen yang
memiliki interaksi yang kuat dengan fase bergerak sehingga terjadi
resolusi (Harris dan Angal, 1989).
Kromatografi kolom merupakan sistem pengaliran suatu fluida
melalui kolom yang mengandung matriks bahan pengisi dan substanta
yang ingin dipisahkan menjadi beberapa komponen dengan adanya
perbedaan daya ikat terhadap bahan pengisi. Pemurnian enzim dengan
kromatografi kolom merupakan cara yang paling efektif dibandingkan cara
pemisahan yang lain (Suhartono et al., 1992).
Berdasarkan mekanisme kerjanya, kromatografi kolom dapat
dikelompokkan menjadi 4 metode, yaitu kromatografi pertukaran ion,
kromatografi affinitas, kromatografi filtrasi gel, dan kromatografi interaksi
hidrofobik (Walsh, 2002). Kromatografi pertukaran ion merupakan
metode pemisahan protein di mana pemisahan terjadi secara selektif
berdasarkan perbedaan muatan dengan ion yang terikat pada matriks. Pada
kromatografi pertukaran ion, larutan enzim diberi muatan tergantung pada
pH, struktur, dan titik isoelektrik enzim. Jika pH larutan enzim di bawah
titik isoelektrik, maka enzim akan bermuatan positif dan kromatografi
yang akan dilakukan adalah kromatografi pertukaran kation. Jika pH
larutan enzim di atas titik isoelektrik, maka enzim akan bermuatan negatif
dan kromatografi yang akan dilakukan adalah kromatografi pertukaran
anion. Contoh bahan pertukaran kation adalah CM selulosa, SE selulosa,
FE selulosa, Dowex 50, dan Aberlite IRC 50. Contoh bahan pertukaran
anion adalah DEAE selulosa, DEAE sephadex, dan TEAE selulosa (Harris
dan Angal, 1989). Kromatografi pertukaran ion paling baik dipergunakan
pada tahap awal pemurnian. Kromatografi ini dapat menampung volume
yang besar karena kapasitas tinggi. Kecepatan dan pemisahannya pun
Kromatografi affinitas merupakan metode pemisahan protein karena
adanya interaksi spesifik antara komponen sampel dengan ligan yang
terikat pada matriks. Pada prinsipnya, suatu ligan yang terikat kovalen
pada matriks yang tidak larut air akan menyerap salah satu atau beberapa
komponen dari campuran. Komponen yang diserap harus memiliki
affinitas yang spesifik terhadap ligan tersebut. Komponen yang tidak
memiliki affinitas akan melaju terus. Beberapa bahan baik organik
maupun anorganik dapat digunakan sebagai matriks. Bahan organik yang
sering dipakai antara lain karbohidrat (selulosa, agarose, pati) dan
poliakrilamida. Bahan anorganik yang dapat dipakai antara lain gelas
(poros dan solid) dan koloid silika. Kromatografi affinitas biasanya
dipergunakan pada tahap menjelang akhir pemurnian (Suhartono et al.,
1992).
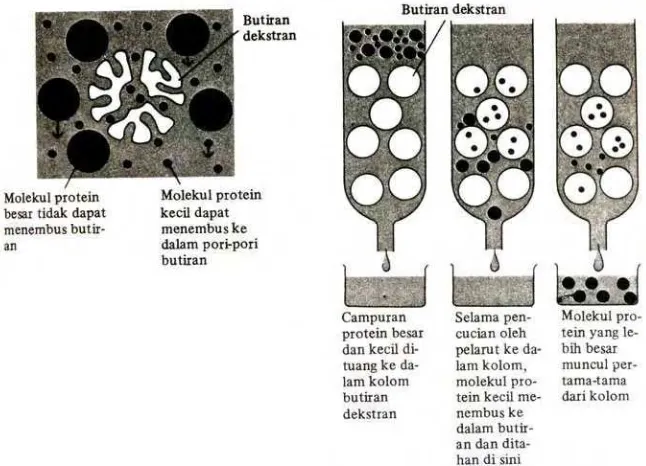
Kromatografi filtrasi gel merupakan metode pemisahan protein
berdasarkan perbedaan ukuran molekul. Proses pemisahan menggunakan
matriks gel berpori yang dipak di dalam kolom dan dikelilingi oleh solven.
Sampel yang mengandung campuran molekul besar dan kecil dilewatkan
ke dalam kolom. Molekul protein yang lebih kecil dapat menembus ke
dalam pori-pori butiran dan karenanya tertahan selama aliran ke bawah
kolom. Molekul protein yang besar tidak dapat menembus ke dalam
pori-pori butiran dan melewati kolom lebih cepat. Molekul protein yang
berukuran menengah akan mengalir ke bawah pada kecepatan antara
tergantung pada tingkat kemampuan menembus butiran. Pemisahan
protein menggunakan kromatografi filtrasi gel dapat dilihat pada gambar
3. Kromatografi filtrasi gel disebut juga dengan saringan molekul
(Lehninger, 1982).
Salah satu bahan yang penting sebagai gel adalah dekstran (polimer
gula yang biasanya larut dalam air) yang telah mengalami reaksi cross
linkage dengan bantuan epiklorohidrin. Hasil yang didapat adalah dekstran
yang menjadi tidak larut dalam air, tetapi masih dapat menyerap
molekul-molekul di dalam molekul-molekulnya sendiri. Gel dekstran disebut juga dengan
13 asam lemah. Selain itu, Sephadex juga stabil pada kisaran pH 1-10
sehingga hal yang paling utama dalam memilih buffer adalah kestabilan
molekul yang dikehendaki dalam buffer yang dipilih. Kromatografi filtrasi
gel baik dipergunakan pada tahap akhir pemurnian. Kromatografi ini
memiliki kapasitas terbatas dan kecepatan fraksinasi rendah (Suhartono et
al., 1992). Sebagian besar pemurnian enzim kitosanase menggunakan
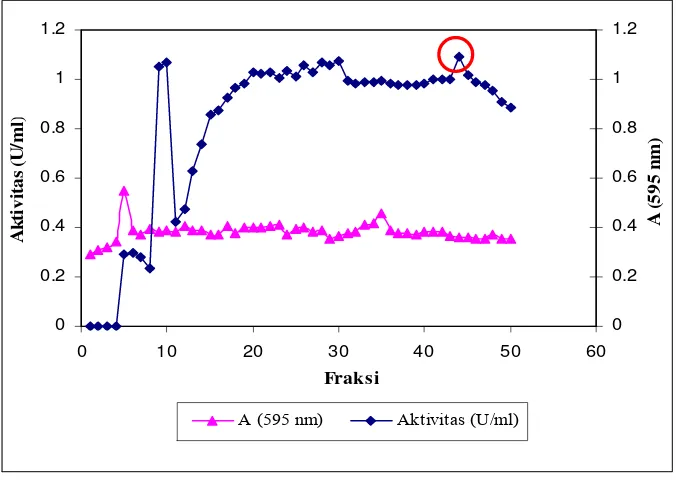
[image:36.612.168.491.226.459.2]kromatografi filtrasi gel dan kromatografi pertukaran ion (Haliza, 2003).
Gambar 3. Prinsip pemisahan protein menggunakan filtrasi gel
Kromatografi interaksi hidrofobik merupakan metode pemisahan
protein karena perbedaan polaritas antar molekul protein pada kekuatan
ion tertentu. Peningkatan kekuatan ionik larutan karena penambahan
garam netral seperti ammonium sulfat atau sodium klorida akan
meningkatkan interaksi hidrofobik protein. Protein diikat oleh matriks
yang bersifat non polar. Protein dibebaskan dari ikatan ini dengan
menggunakan eluen yang polaritasnya diturunkan (Walsh, 2002).
F. Elektroforesis SDS-PAGE
Elektroforesis adalah teknik pemisahan fraksi-fraksi zat berdasarkan
perbedaan ukuran, bentuk, muatan, atau sifat kimia molekul. Elektroforesis
dapat digunakan untuk menentukan berat molekul protein, mendeteksi
kemurnian dan kerusakan protein, menetapkan titik isoelektrik protein, dan
memisahkan spesies-spesies molekuler yang berbeda secara kuantitatif dan
kualitatif (Boyer, 1986).
Protein memiliki gugus-gugus yang dapat terionisasi sehingga membentuk
gugus bermuatan pada pH tertentu baik sebagai anion maupun kation sehingga
dapat dipisahkan dengan teknik elektoforesis. Protein akan bermuatan
tergantung pada pH larutan dan titik isoelektrik. Pada titik isoelektrik, protein
tidak akan bergerak di bawah pengaruh medan listrik. Jika pH larutan di
bawah pI, protein bergerak sebagai kation dan kecepatannya naik dengan
turunnya pH lalu kation ini akan bergerak ke elektroda negatif. Jika pH larutan
di atas pI, protein bergerak sebagai anion dan kecepatannya naik dengan
meningkatnya pH lalu anion ini akan bergerak ke elektroda positif (Edelstein
dan Bollag, 1991).
Media yang digunakan untuk elektroforesis harus dapat mengurangi atau
mencegah terjadinya konveksi dan tidak bereaksi dengan sampel atau
menghambat pergerakan partikel sebagai akibat adanya ikatan antara sampel
dengan matriks. Keberhasilan pemisahan protein dengan elektroforesis
dipengaruhi oleh beberapa faktor antara lain buffer, suhu, waktu, dan arus
listrik yang digunakan. Buffer berfungsi untuk mempertahankan pH baik
dalam chamber maupun gel serta pembawa muatan listrik (Nur dan
Adijuwana, 1988). Pada umumnya, buffer yang digunakan bersifat sedikit
alkalis dengan pH sekitar 8-9. Pada pH tersebut, sebagian besar protein
bermuatan negatif dan bergerak ke kutub anoda yang biasanya terletak pada
bagian dasar gel. Hal ini mencegah kemungkinan protein untuk bergerak ke
atas dari sistem elektroforesis (Scopes, 1987). Elektroforesis dilakukan pada
suhu dingin (0-40C) untuk mengurangi kehilangan aktivitas protein akibat
denaturasi dan kerusakan akibat proteolisis (Edelstein dan Bollag, 1991).
Selain itu, kenaikan suhu yang tinggi selama elektroforesis dapat
menyebabkan pemisahan yang kurang baik. Waktu yang diperlukan untuk
15 tinggi arus listrik yang digunakan, maka semakin pendek waktu yang
dibutuhkan, tetapi suhu akan meningkat (Nur dan Adijuwana, 1988).
Menurut Harris dan Angal (1989), elektroforesis gel poliakrilamida
(PAGE) adalah metode yang paling banyak digunakan karena memiliki
kapasitas pemisahan yang tinggi. Salah satu metode PAGE yang umumnya
digunakan adalah SDS-PAGE (Sodium Dodecyl Sulphate Polyacrilamide Gel
Electrophoresis).
Menurut Nur dan Adijuwana (1988), radikal-radikal bebas yang terbentuk
dari ammonium persulfat (APS) akan bereaksi dengan akrilamida di mana
terjadi penyimpanan radikal bebas di dalam molekul akrilamida sehingga
terbentuk akrilamida aktif. Akrilamida aktif ini akan bereaksi dengan cara
yang sama dengan akrilamida yang lain sehingga dihasilkan suatu rantai
polimer yang panjang. Gel poliakrilamida terbentuk oleh adanya polimerisasi
dari monomer akrilamida dan pembentukan ikatan silang kovalen antar rantai
panjang akrilamida. Cross linking agent yang digunakan adalah
N,N’-metilen-bis-akrilamida. Kompleks APS-TEMED mengkatalisa pembentukan radikal
bebas dari persulfat yang selanjutnya berfungsi sebagai inisiator polimerisasi.
Lembaran elektroforesis gel poliakrilamida terdiri atas 2 bagian, yaitu gel
penahan (stacking gel) dan gel pemisah (separating gel). Stacking gel terletak
pada bagian atas dari separating gel. Stacking gel berfungsi sebagai tempat
dicetaknya sumur untuk memasukkan sampel. Selain itu, stacking gel
berfungsi untuk mengkonsentrasikan sampel membentuk pita yang tajam
sebelum memasuki separating gel. Separating gel berfungsi sebagai tempat
terjadinya pemisahan protein (Nur dan Adijuwana, 1988).
Penambahan SDS pada gel poliakrilamida menghasilkan SDS-PAGE yang
digunakan untuk sampel yang terdenaturasi. SDS adalah detergen anionik
yang bersama dengan β-merkaptoetanol dan pemanasan akan merusak struktur
tiga dimensi protein dengan cara memecah ikatan disulfida yang selanjutnya
tereduksi menjadi gugus sulfidril. Protein yang terdenaturasi sempurna akan
mengikat SDS dalam jumlah yang setara dengan berat molekul protein
tersebut. Setiap 1.4 gram SDS dapat mengikat 1 gram protein (Harris dan
mempunyai mobilitas yang lebih kecil. Adanya gliserol dalam buffer sampel
berfungsi untuk meningkatkan berat jenis larutan sampel sehingga dapat
masuk dengan mudah ketika diinjeksi ke dalam sumur gel. Buffer sampel juga
mengandung pewarna bromphenol blue yang berfungsi untuk membantu
memonitor jalannya elektroforesis. Jika pewarna telah mencapai bagian bawah
gel, maka elektroforesis dihentikan (Wilson dan Walker, 2000).
Sebelum dilakukan prosedur pewarnaan, terlebih dahulu dilakukan proses
fiksasi terhadap gel hasil elektroforesis. Proses fiksasi bertujuan untuk
mengendapkan dan mengimobilisasi pita-pita protein pada gel dan
menghilangkan komponen-komponen non protein yang dapat mengganggu
pewarnaan seperti SDS. Proses fiksasi dilakukan dengan menggunakan
campuran metanol-asam asetat-air, formaldehid, TCA, atau asam perklorat
5-10% (Scopes, 1987).
Pita-pita protein hasil pemisahan dengan elektoforesis diperjelas dengan
teknik pewarnaan. Syarat dari pewarna yang digunakan untuk staining adalah
dapat berikatan kuat dengan protein, tetapi tidak bereaksi dengan gel. Ikatan
antara pewarna dan gel adalah ikatan non kovalen sehingga mudah dilepaskan
dengan pencucian secara intensif (Scopes, 1987). Proses penghilangan warna
(destaining) dilakukan dengan merendam gel dalam larutan peluntur sampai
diperoleh latar belakang yag relatif jernih sehingga pita-pita protein yang
dihasilkan dapat diamati dengan jelas (Nur dan Adijuwana, 1988).
Berat molekul protein dapat diketahui dengan membandingkan mobilitas
relatif (Rf) protein dengan protein standar yang berat molekulnya sudah
diketahui. Rf protein merupakan perbandingan jarak antara titik awal ke pita
protein dengan titik awal ke titik akhir elektroforesis (Wilson dan Walker,
17
BAB III
METODOLOGI PENELITIAN
A. Bahan dan Alat
Isolat bakteri yang digunakan dalam penelitian ini adalah isolat Bacillus
licheniformis MB-2 yang merupakan koleksi Laboratorium Mikrobiologi dan
Biokimia PAU-IPB. Bahan-bahan yang digunakan antara lain tepung kitin
rajungan Cirebon, NaOH, HCl pekat, glass wool, akuades, koloidal kitosan,
K2HPO4, KH2PO4, MgSO4, ekstrak khamir, casiton, bacto agar, gelrite,
ammonium sulfat teknis, 0.05 M buffer fosfat pH 6, soluble chitosan,
glukosamin, pereaksi Schales, air bebas ion, BSA, Bradford reagent, kantung
dialisis, EDTA, NaHCO3, Sephadex G-100, buffer elektroforesis, buffer
sampel, larutan fiksasi, etanol, larutan enhancer, akuabides, larutan silver
nitrat, larutan destaining, Marker LMW, buffer sitrat, buffer asetat, buffer
fosfat-sitrat, buffer Na-fosfat, buffer tris-Cl, dan buffer universal.
Peralatan yang digunakan adalah peralatan yang terdapat pada
Laboratorium Mikrobiologi dan Biokimia PAU-IPB antara lain neraca
analitik, sudip, penangas air, oven, corong gelas, bunsen, laminar flow, jarum
ose, kapas, cawan petri, inkubator suhu 550C, pH meter, pH indikator,
autoclave, shaker waterbath suhu 550C, alat sentrifugasi dingin, magnetic
stirrer, eppendorf, tips, pipet mikro, alat vorteks, tabung reaksi,
spektrofotometer visible, freeze dryer, kolom kromatografi, alat vakum
(Millipore), alat pemampung fraksi (fraction collector), peralatan
elektoforesis, dan peralatan gelas lainnya.
B. Metode
1. Pembuatan Tepung Kitosan (Kolodziejska et al., 2000)
Tepung kitin rajungan Cirebon dicampurkan dengan larutan NaOH
50% dengan perbandingan 1:20 lalu dipanaskan pada 1000C selama 1 jam.
Setelah itu, dilakukan pencucian dengan air sampai mencapai pH netral.
Tepung tersebut kemudian dikeringkan dengan menggunakan oven suhu
2. Pembuatan Koloidal Kitosan (Arnold dan Solomon, 1986)
Tepung kitosan dicampurkan dengan HCl pekat dengan
perbandingan 1:20 lalu didiamkan selama semalam di cool room. Larutan
tersebut difiltrasi dengan glass wool lalu ditambahkan air dingin sebanyak
10 kali berat tepung kitosan. Filtrat yang diperoleh kemudian dinetralkan
pH-nya dengan larutan NaOH 12 N. Larutan tersebut disaring di cool
room lalu dibilas dengan 50 ml air dingin. Koloidal kitosan yang diperoleh
disimpan pada suhu dingin.
3. Penyegaran Kultur
Penyegaran kultur dilakukan dengan mengambil 1-2 ose isolat
Bacillus licheniformis MB-2 dari kultur persediaan gliserol lalu digoreskan
pada media thermus padat dan diinkubasi pada inkubator suhu 550C
selama 5 hari. Aktivitas enzim kitosanase ditandai dengan adanya areal
bening di sekitar koloni bakteri. Media thermus padat yang digunakan
adalah 1.0% koloidal kitosan, 0.7% K2HPO4, 0.3% KH2PO4, 0.5%
MgSO4, 0.25% ekstrak khamir, 0.25% casiton, 1.5% bacto agar, dan 0.4%
gelrite (pH 6) (Chasanah, 2004).
4. Pembuatan Kultur Starter
Pembuatan kultur starter dilakukan dengan menginokulasikan 1-2
ose koloni bakteri yang menunjukkan aktivitas enzim kitosanase ke dalam
150 ml media thermus cair lalu diinkubasi pada shaker waterbath suhu
550C selama 24 jam dengan kecepatan agitasi 120 rpm. Media thermus
cair yang digunakan berdasarkan pada Park et al. (1999), yaitu 0.4%
koloidal kitosan, 0.7% K2HPO4, 0.3% KH2PO4, 0.5% MgSO4, 0.25%
ekstrak khamir, dan 0.25% casiton (pH 7).
5. Produksi Enzim Kitosanase (Chasanah, 2004)
Kultur starter sebanyak 15 ml dimasukkan ke dalam 85 media
thermus cair lalu diinkubasi pada shaker waterbath suhu 550C selama 7
19 enzim kitosanase sama dengan media yang digunakan untuk pembuatan
kultur starter.
Setelah ditumbuhkan dalam media thermus cair selama 7 hari, sel
bakteri dan sisa-sisa media yang tidak larut dipisahkan dengan cara
sentifugasi pada 40C selama 20 menit dengan kecepatan 10000 rpm.
Supernatan bebas sel yang diperoleh kemudian ditambah ammonium sulfat
pada tingkat kejenuhan 80%. Penambahan ammonium sulfat dilakukan
secara perlahan-lahan pada suhu dingin sambil di-stirrer. Setelah itu,
supernatan disimpan selama semalam di cool room. Endapan yang berisi
enzim dan beberapa jenis protein lainnya dipisahkan dengan cara
sentrifugasi pada 40C selama 20 menit dengan kecepatan 10000 rpm.
Endapan tersebut kemudian dilarutkan dalam 0.05 M buffer fosfat pH 6
untuk menjadikan volume larutan sebanyak 8 ml. Larutan enzim yang
diperoleh disebut enzim kasar (crude enzyme).
6. Dialisis
Dialisis dilakukan dengan menggunakan kantung selofan yang
merupakan turunan membran selulosa dengan cutoff 12000 Dalton.
Kantung dialisis dipotong sesuai kebutuhan kemudian direbus dalam
larutan EDTA dan NaHCO3 selama 10 menit. Setelah dididihkan selama
10 menit, larutan tersebut dibuang dan kantung dialisis direbus kembali
dengan air bebas ion selama 10 menit sebanyak 2 kali. Proses dialisis
dilakukan dengan memasukkan crude enzyme sebanyak 4 ml terhadap
kantung tersebut. Setelah itu, kedua ujung kantung diikat dengan benang
kasur. Dengan posisi menggantung, kantung tersebut dimasukkan ke
dalam wadah yang berisi 0.025 M buffer fosfat pH 6. Selanjutnya, kantung
dialisis di-stirrer selama semalam di cool room.
7. Freeze Dry
Freeze dry bertujuan untuk meningkatkan konsentrasi protein dan
aktivitas enzim. Sebelum freeze dry dilakukan, enzim hasil dialisis
Enzim yang telah beku kemudian dimasukkan ke dalam tabung dan siap
untuk di-freeze dry. Freeze dry dilakukan sampai volume larutan enzim
yang tersisa ± 2/3 dari volume larutan enzim mula-mula (± 2 ml).
8. Pemurnian Enzim Kitosanase (Haliza, 2003)
Pemurnian enzim kitosanase dilakukan pada suhu dingin. Enzim
hasil freeze dry digunakan untuk pemurnian. Enzim ini dimurnikan dengan
menggunakan kromatografi filtrasi gel. Matriks yang digunakan untuk
kromatografi filtrasi gel adalah Sephadex G-100. Matriks ini terlebih
dahulu harus dikembangkan (swelling) sebelum digunakan. Swelling
dilakukan dengan merendam Sephadex G-100 dalam akuades pada suhu
dingin selama 3 hari. Setelah itu, gel tersebut divakum dengan
menggunakan Millipore untuk menghilangkan gelembung-gelembung
udara yang dapat mengganggu pemurnian. Kolom yang akan digunakan
untuk pemurnian harus dibilas terlebih dahulu menggunakan akuades dan
air bebas ion. Matriks dimasukkan secara perlahan ke dalam kolom sambil
disetimbangkan dengan 0.05 M buffer fosfat pH 6. Matriks tersebut
dibiarkan memadat selama semalam. Enzim hasil freeze dry sebanyak 2 ml
dimasukkan ke dalam kolom. Elusi dilakukan dengan buffer yang sama.
Fraksi dikumpulkan dengan menggunakan fraction collector setiap 100
drop. Setiap fraksi yang ditampung diukur aktivitas enzim kitosanase dan
konsentrasi protein.
9. Elektroforesis SDS-PAGE (Edelstein dan Bollag, 1991)
Persiapan awal yang perlu dilakukan dalam elektroforesis adalah
pembuatan gel. Metode yang digunakan dalam pembuatan gel adalah
metode Edelstein dan Bollag (1991). Komposisi gel SDS-PAGE dapat
[image:43.612.169.460.635.698.2]dilihat pada tabel 2.
Tabel 2. Komposisi gel SDS-PAGE
Bahan Separating gel (8%) Stacking gel (4%)
Larutan A
Larutan B
2.7 ml
2.5 ml
0.67 ml
21 Larutan C
Akuabides
APS 10%
TEMED
-
4.8 ml
50 µl
5.0 µl
1.25 ml
3.0 ml
50 µl
5.0 µl
Bahan untuk separating gel dicampur satu persatu dengan
memasukkan TEMED pada akhir campuran. Larutan tersebut diaduk dan
dipipet perlahan ke dalam plate kaca sampai 1.5 cm dari permukaan kaca
lalu didiamkan sekitar 15-20 menit. Dalam proses ini diusahakan agar
tidak terbentuk gelembung udara. Setelah gel memadat, campuran stacking
gel dipipet perlahan ke dalam plate kaca lalu dengan segera dimasukkan
sisir (10 sumur) sebagai tempat memasukkan sampel.
Sampel yang telah dipanaskan pada 1000C selama 3 menit
dicampurkan dengan buffer sampel lalu dilakukan loading sampel ke
dalam sumur sebanyak 12 µl. Berbeda halnya dengan sampel, Marker
yang di-loading ke dalam sumur sebanyak 10 µl. Sebelum running
dilakukan, buffer elektroforesis dimasukkan ke dalam chamber. Running
elektroforesis dilakukan pada 100 Volt, 50 mA dalam kondisi dingin.
Waktu yang diperlukan untuk running elektroforesis sekitar 1.5 jam.
Setelah pemisahan, gel dilepas dari plate kaca lalu direndam dalam
larutan fiksasi (25% metanol + 12% asam asetat) selama 1 jam.
Selanjutnya, gel tersebut direndam dalam larutan etanol 50% selama 20
menit dan larutan etanol 30% selama 2 x 20 menit. Setelah itu, gel tersebut
direndam dalam larutan enhancer (larutan Na2S2O3.5H2O) selama 1 menit.
Gel kemudian dicuci dengan akuabides selama 3 x 20 menit. Setelah
dicuci dengan akuabides, gel direndam dalam larutan stainingsilver nitrat
(larutan AgNO3 + formaldehida 37%) selama 30 menit lalu dibilas cepat
dengan akuabides selama 2 x 20 detik. Kelebihan warna dihilangkan
dengan larutan destaining (larutan Na2CO3 + formaldehida 37%) sampai
diperoleh pita-pita protein yang jelas teramati dengan latar belakang relatif
10.Karakterisasi Enzim Kitosanase (Chasanah, 2004) a. Suhu Optimum
Aktivitas enzim dianalisis pada suhu inkubasi 37, 50, 60, 70, 80,
dan 900C untuk crude enzyme dan enzim murni.
b. pH Optimum
Aktivitas crude enzyme dianalisis pada 0.05 M buffer sitrat pH 3,
0.05 M buffer asetat pH 4-6, 0.05 M buffer fosfat-sitrat pH 5, 0.05 M
buffer Na-fosfat pH 6-8, dan 0.05 M buffer tris-Cl pH 8 pada suhu
optimum crude enzyme. Sedangkan, aktivitas enzim murni dianalisis
pada buffer universal pH 4-12 pada suhu optimum enzim murni.
c. Pengaruh Panas Terhadap Stabilitas Enzim
Pengujian pengaruh panas terhadap stabilitas enzim murni
dilakukan dengan cara memanaskan enzim (tanpa substrat dan buffer
enzim) pada 700C selama 0, 0.5, 1, 1.5, dan 2 jam serta pada 900C
selama 0, 1, dan 2 jam. Pengukuran pengaruh panas terhadap stabilitas
enzim dinyatakan dalam nilai k, t1/2, Ea (Toledo, 1991). Nilai k suatu
enzim adalah konstanta laju deaktivasi enzim.
ln C = -k (t) + ln C0 ... (1)
C0 = aktivitas enzim pada awal inkubasi (U/L)
t = waktu inkubasi (menit)
Nilai t1/2 suatu enzim adalah waktu inkubasi pada suhu tertentu yang
menyebabkan aktivitas enzim tinggal 50% dari aktivitas enzim semula.
t1/2 = -ln (0.5) ...(2) k
Energi aktivasi (Ea) dapat ditetapkan secara grafik berdasarkan
persamaan Arrhenius. Persamaan ini merupakan hubungan konstanta
laju deaktivasi terhadap suhu absolut. Ea merupakan slope dari ln k
terhadap 1/T.
k = A0 (e) –Ea/RT (persamaan Arrhenius)
ln k = -Ea . 1 + ln A0 ...(3) R T
k = konstanta laju deaktivasi
23 Ea = energi aktivasi (kkal/gmol.0K)
R = tetapan gas (1.987 kal/gmol.0K)
A0 = faktor frekuensi
d. Pengaruh pH Terhadap Stabilitas Enzim
Pengujian pengaruh pH terhadap stabilitas enzim murni dilakukan
dengan cara memanaskan enzim dalam buffer universal pH 6 tanpa
substrat pada 700C selama 0, 0.5, 1, 1.5, dan 2 jam.
11.Pengukuran Aktivitas Enzim Kitosanase (Yoon et al., 2000)
Prinsip pengukuran aktivitas enzim kitosanase didasarkan pada
perhitungan gula pereduksi yang diproduksi dalam hidrolisis soluble
chitosan dengan metode Schales modifikasi dan glukosamin digunakan
[image:46.612.167.506.332.624.2]sebagai standar (Imoto dan Yagashita, 1971).
Tabel 3. Pengukuran aktivitas enzim kitosanase Bahan Sampel
(µl)
Kontrol
(µl)
Blanko
(µl)
0.05 M buffer fosfat pH 6 100 100 -
Soluble chitosan 100 100 -
Enzim kitosanase 100 - -
Inkubasi pada 700C selama 30 menit
Freeze selama 15 menit
Campuran 200 133 -
Enzim kitosanase - 67 -
Air bebas ion 800 800 1000
Pereaksi Schales 1000 1000 1000
Didihkan selama 15 menit
Sentrifugasi pada 40C selama 10 menit dengan kecepatan 8000 rpm
Ukur absorbansi pada panjang gelombang 420 nm
Nilai absorbansi dari sampel, kontrol, dan blanko dimasukkan ke
dalam kurva standar glukosamin sehingga dapat ditentukan jumlah
glukosamin yang terkandung di dalam sampel. Selanjutnya, jumlah
aktivitas enzim. Satu unit aktivitas enzim kitosanase didefinisikan sebagai
jumlah enzim yang memproduksi 1 µmol gula pereduksi (glukosamin) per
menit pada kondisi tertentu. Aktivitas spesifik enzim ditentukan dengan
cara membagi unit aktivitas enzim dengan konsentrasi protein. Tingkat
kemurnian diperoleh dengan membagi aktivitas spesifik enzim pada satu
tahap dengan aktivitas spesifik enzim pada tahap sebelumnya.
Unit aktivitas enzim = 300 x Glc x 1 x 1000 x 1 ...(4) (U/ml) 200 BM 100 30
Keterangan :
300 = volume sampel hasil reaksi enzimatis (µl)
200 = volume sampel untuk reaksi Schales (µl)
Glc = jumlah glukosamin sampel (µg/ml)
BM = berat molekul glukosamin, yaitu 215.6 (gram/mol)
1000= faktor konversi dari µl ke ml
100 = volume larutan enzim atau larutan soluble chitosan (µl)
30 = waktu inkubasi (menit)
12.Pengukuran Konsentrasi Protein (Bradford, 1976)
Sampel protein sebanyak 100 µl ditambah dengan 2 ml Bradford
reagent. Campuran tersebut divorteks dan didiamkan pada suhu ruang
selama 5 menit. Protein akan diikat oleh Coomassie Briliant Blue G-250
yang terdapat pada Bradford reagent membentuk kompleks berwarna biru.
Absorbansi diukur pada panjang gelombang 595 nm. Konsentrasi protein
sampel dihitung berdasarkan kurva standar BSA.
25
BAB IV
HASIL DAN PEMBAHASAN
A. Produksi Enzim Kitosanase
Semua mikroorganisme memerlukan nutrien dasar sebagai sumber karbon,
nitrogen, dan faktor esensial pertumbuhan (mineral dan vitamin) untuk
menopang pertumbuhannya. Nutrien dasar tersebut di samping menyediakan
energi juga digunakan untuk pembentukan konstituen seluler. Oleh karena itu,
untuk mendapatkan hasil yang maksimum, media pertumbuhan yang
digunakan harus mengandung nutrien dasar tersebut (Wang et al., 1979).
Isolat Bacillus licheniformis MB-2 disegarkan terlebih dahulu agar
mencapai kondisi optimalnya. Isolat tersebut ditumbuhkan pada media
thermus padat dan diinkubasi pada inkubator suhu 550C. Inkubasi dalam suhu
ruang dilakukan selama ± 30 menit sebelum dimasukkan ke dalam inkubator
suhu 550C agar isolat tidak mengalami thermal shock akibat perubahan suhu
drastis. Media thermus padat yang digunakan terdiri atas 1.0% koloidal
kitosan, 0.7% K2HPO4, 0.3% KH2PO4, 0.5% MgSO4, 0.25% ekstrak khamir,
0.25% casiton, 1.5% bacto agar, dan 0.4% gelrite (Chasanah, 2004). Koloidal
kitosan digunakan sebagai substrat induser untuk memproduksi enzim
kitosanase. Hal ini karena sebagian besar mikroorganisme memproduksi
enzim tersebut secara induktif. K2HPO4 dan KH2PO4 digunakan sebagai
sumber fosfor yang diperlukan dalam sintesis asam nukleat, fosfolipid, dan
ATP. MgSO4 digunakan sebagai kofaktor enzim dan pertumbuhan. Ekstrak
khamir dan casiton digunakan sebagai sumber nitrogen untuk produksi enzim
kitosanase (Stanbury dan Whitaker, 1984).
Isolat Bacillus licheniformis MB-2 ditumbuhkan pada waktu optimumnya,
yaitu 5 hari. Selama waktu tersebut dihasilkan enzim dengan aktivitas
tertinggi. Waktu yang terlalu singkat akan menghasilkan enzim yang tidak
optimal akibat mikroba belum beradaptasi dengan lingkungannya. Waktu
yang terlalu lama akan menyebabkan enzim mengalami inhibisi akibat
menumpuknya produk reaksi enzim dengan substrat. Jumlah mikroba yang
banyak. Nutrien yang berbentuk polimer tidak dapat memasuki sel mikroba
karena ukuran fisiknya. Polimer ini biasanya dicerna terlebih dahulu oleh
enzim-enzim ekstraseluler yang disekresikan oleh mikroba (Suhartono, 1989).
Oleh karena itu, produksi enzim-enzim mikrobial memanfaatkan polimer ini
(salah satunya kitosan) sebagai substrat dan induser untuk menghasilkan
enzim kitosanase. Penggunaan kitosan sebagai sumber karbon biasanya tidak
langsung ditambahkan dalam bentuk serbuk ke dalam media, tetapi dalam
bentuk yang lebih memungkinkan dan lebih mudah untuk menerima penetrasi
enzim kitinolitik agar dapat diuraikan menjadi monomer-monomer
glukosamin yang dapat diangkut melalui membran sel dan dimetabolisme oleh
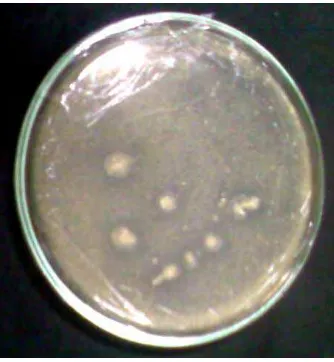
mikroba. Hidrolisis kitosan oleh enzim kitosanase pada media thermus padat
[image:49.612.245.412.312.493.2]akan menghasilkan areal bening di sekitar koloni bakteri (gambar 4).
Gambar 4. Aktivitas kitosanase MB-2 pada media thermus padat
Enzim kitosanase diproduksi menggunakan media thermus cair yang
mengandung 0.4% koloidal kitosan. Pembuatan kultur starter sebelum
produksi enzim bertujuan untuk memperbanyak sel yang seragam dengan
umur fase pertumbuhan yang sama (Kurakake et al., 2000). Kultur starter
yang ditambahkan ke dalam media thermus cair diinkubasi pada shaker
waterbath suhu 550C selama 7 hari dengan kecepatan agitasi 120 rpm. Dari
hasil karakterisasi yang dilakukan oleh Chasanah et al. (2000) diketahui
bahwa Bacillus licheniformis MB-2 yang ditumbuhkan pada media thermus
27 licheniformis MB-2 sama dengan Bacillus coagulans LH 28.38 yang
membutuhkan waktu 7 hari pada 550C (Haliza, 2003). Matsuebacter
chitosanotabidus 3001 (Park et al., 1999), Amycolatopsis sp. CsO-2 (Okajima
et al., 1994),