BAHANAJAR (Hand Out)
BIOKIMIA
Disusun oleh:
Prof. Dr. agr. Moh.Amin, M.Si
Dr. Betty Lukiati, M.S
Siti Imroatul Maslikah, S.Si, M.Si
Balqis, S.Pd, M.Si
JURUSAN BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
UNIVERSITAS NEGERI MALANG
i
DAFTAR ISI
1. BAB I Pendahuluan ...1
2. BAB IIBiomolekul Dalam Organisasi Molekuler Sel dan
Materi Genetik ...7
3. BAB III Asam Amino dan Protein ...15
4. BAB IV Enzim ...30
5. BAB V Struktur dan Fungsi Karbohidrat ...44
6. BAB VIMetabolisme Karbohidrat ...58
7. BAB VIIMetabolisme Lipid ... 78
1
BAB I
PENDAHULUAN
A. KOMPETENSI
1. mampu menjelaskan pengertian biokima
2. mampu menjelaskan tujuan yang dipelajari dalam biokimia 3. mampu menjelaskan tiga bidang bahasan dalam Biokimia 4. mampu menjelaskan perkembangan ilmu biokimia
5. mampu menjelaskan kegunaan ilmu biokimia dalam kehidupan sehari-hari
B. ELEMEN KOMPETENSI
1. mendeskripsikan pengertian biokima
2. mendeskripsikan tujuan yang dipelajari dalam biokimia 3. mendeskripsikan tiga bidang bahasan dalam Biokimia 4. mendeskripsikan perkembangan ilmu biokimia
5. mendeskripsikan kegunaan ilmu biokimia dalam kehidupan sehari-hari
C. MATERI
Pengertian Biokimia
Selamat datang biokimia! Anda sudah menemukan aspek dari subjek ini dengan pengalaman sebelumnya dalam ilmu kimia dan biologi. Ahli biokimia menggunakan hukum-hukum dasar ilmu kimia, biologi, dan ilmu fisika untuk menjelaskan proses hidup sel. Sungguhpun istilah biokimia telah menjadi hal yang biasa di dalam bahasa kita, suatu definisi yang penuh arti ringkas adalah sulit. Definisi yang paling sederhana adalah " ilmu kimia sel hidup. Biokimia mempunyai komponen biologi dan ilmu kimia dan ahli biokimia harus baik berpengalaman dalam kedua-duanya. Dengan demikian Biokimia adalah Sains Biologi, yang memanfaatkan Hukum-hukum Fisika dan Kimia untuk menjelaskan proses kehidupan. Untuk itu perlu dijelakan ciri-ciri makhluk hidup dibandingkan dengan makhluk tidak hidup diantaranya adalah:
1) Mempunyai susunan yang kompleks, tetapi terorganisir dengan sangat rapi.
2) Mampu mempertahankan ‘keteraturan’ dirinya di dalam lingkungan yang semakin
tidak teratur (Hukum Thermodinamika II).
2
Ketiga ciri utama di atas dimiliki secara melekat oleh organisme. Kekurangan dari satu sifat akan meniadakan ciri sebagai organisme.
Tujuan Biokimia
Mengacu pada uraian pengertian Biokimia di atas, keseluruhan tujuan biokimia adalah mendeskripsikan proses hidup di tingkatan molekul. Bahkan sel yang terkecil-pun berisi beribu-ribu bahan-kimia anorganik maupun organik, banyak di antara mereka molekul besar disebut makromolekul. Semua proses biologi yang mencakup visi, diesti (pencernaan), pemikiran, gerakan, imunitas, dan kondisi-kondisi penyakit diakibatkan oleh perilaku molekul. Oleh karena itu, dalam rangka mendeskripsikan proses-proses ini, pertama-tama harus mempunyai suatu pengetahuan struktur yang kimia molekul yang berperan. Kedua, harus mempunyai suatu pemahaman fungsi biologi molekular. Dengan demikian tujuan Biokimia adalah mendiskripsikan struktur, organisasi dan fungsi zat hidup pada tingkat molekul.
Dengan tujuan ini banyak masalah yang ingin dipecahkan antara lain:
Bagaimana komponen-komponen organisme mengorganisir diri dalam menyusun
struktur “supramolekul”: sel jaringan organisme?
Bagaimana organisme mengekstrak energi dari lingkungan untuk
mempertahankan hidupnya?
Bagaimana organisme menyimpan dan menyalurkan informasi genetik dengan
sangat akurat?
Reaksi kimia apa saja yang menyebabkan / menyertai proses reproduksi, penuaan
dan kematian suatu sel/ organisme?
Bagaimana reaksi-reaksi kimia di dalam sel dikendalikan?
3
Gambar 1.1. Biokimia alam menunjukkan konsep penting dan hubungannya
Biokimia dibagi oleh beberapa ilmuwan hidup ke dalam dua tingkatan :
1. Conformational: menemukan struktur yang kimia dan susunan tiga dimensi biomolecules.
2. Informational: penjelasan suatu bahasa untuk komunikasi di dalam sel dan organisma.
Kekayaan yang unik suatu sel ditentukan oleh gen yang dinyatakan oleh sel itu. Informasi genetik dalam wujud DNA dikodekan untuk menghasilkan protein yang adalah molekul sel fungsional dan struktural yang utama.
Tiga Bidang Bahasan Biokimia:
Mengacu pada deskripsi tujuan Biokimia maka secara ringkas bidang bahasannya meliputi:
1) Struktur dan Fungsi Biomolekul:
Membahas struktur kimia komponen-komponen penyusun organisme serta hubungan antara struktur tersebut dengan fungsi biologisnya.
2) Metabolisme
Membahas keseluruhan reaksi-reaksi kimia yang terjadi di dalam organisme. 3) Penyimpanan dan Aliran Informasi Genetik
4
Sejarah Biokimia Sejarah awal biokimia
Masyarakat pada awal peradaban Mesir, Negeri China, India, Roma, dan di tempat lain tidak memahami prinsip dasar biokimia tentang membakar roti, peragian jus buah, atau perawatan penyakit dengan tumbuhan dan zat-zat dari hewan. Bagaimanapun, ketiadaan pengetahuan tidak mencegah kenikmatan mereka dari proses biokimia ini. Studi awal dalam biologi yang terkonsentrasi pada perawatan penyakit dan pencapaian kesehatan yang baik, telah berakar kuat dalam filosofi dan agama.
Masyarakat Cina pada abad yang keempat sebelum Masehi percaya bahwa manusia berisi lima unsur: air, api, kayu, metal dan bumi. Ketidakseimbangan susunan unsur-unsur ini menyebabkan sakit/penyakit. Pada abad ketujuh setelah Masehi, dokter Cina menemukan bahwa rabun ayam itu bisa diperlakukan pada hati babi dan domba. Ahli biokimia modern dan dokter mengetahui bahwa rabun ayam disebabkan oleh kekurangan vitamin A, yaitu suatu zat kimia yang berada di dalam hati.
Awal sejarah Yunani, termasuk Plato telah mencoba untuk menjelaskan bahwa tubuh dalam kaitan dengan teori kosmologi dan stres diet berguna untuk perawatan penyakit. Istilah Yunani untuk pencernaan, pepsin, sebagai suatu kata yang menandakan panas dalam, adalah asal dari kata pepsin yaitu suatu enzim pencernaan.
Biokimia modern
5
Gambar 1.2. Dua perspektif biokimia, yaitu ilmu pengetahuan fisika - kimia dan biologi
Aplikasi Biokima
Biokimia sebagai suatu disiplin ilmu pengetahuan terutama telah membantu perkembangan di bidang pertanian, kedokteran, pangan, farmasi dan lain-lain.
1) Di bidang kedokteran, biokimia telah berperan dalam deteksi penyakit dan diagnosis serta memonitor respons-respons terhadap bermacam perlakuan. Contohnya pada diagnosis penyakit hati dapat dimonitor dengan pengukuran aktivitas enzim transaminase (dengan tes laboratorium SGPT (serum glutamic pyruvic transaminase), SGOT (serum glutamic oxaloacetic transaminase) dan kadar bilirubin.
SGPT adalah sebuah enzim yang biasanya hadir di sel-sel hati dan jantung.SGPT
dilepaskan ke dalam darah ketika hati atau jantung rusak.Juga disebut alanin aminotransferase (ALT).
SGOT, kini disebut AST (aspartat aminotransferase), adalah enzim yang biasanya
hadir di sel-sel hati dan jantung. SGOT dilepaskan ke dalam darah ketika hati atau jantung rusak. Tingkat SGOT meningkat bila ada kerusakan hati (misalnya, dari virus hepatitis) atau penyakit jantung. Beberapa obat juga dapat meningkatkan kadar SGOT.
Bilirubin adalah pigmen kekuningan yang dilepaskan ketika sel-sel darah merah
6
bilirubin dalam darah (hiperbilirubinemia) dapat mengindikasikan kerusakan hati, dan dapat menyebabkan sakit kuning (menguningnya kulit dan putih mata), tinja berwarna.
2) Di bidang farmasi dan toksikologi, obat-obatan umumnya mengganggu jalur metabolisme spesifik.
Contoh:
antibiotik penisilin dan sejenisnya dapat membunuh bakteri dengan cara
menghambat enzim yang berperan dalam sintesis polisakarida penting penyusun dinding sel bakteri. Sebaliknya pada sel hewan tidak memerlukan sintesis senyawa ini sehingga dapat digunakan untuk pengobatan.
AZT (azido dideoksi timidin) adalah senyawa menggantikan Timin berperan untuk memblokir sintesis DNA oleh virus (HIV)
6-Merkapto Purinadalah senyawa yang berperan menghambat sintesis DNA
pada pembiakan sel leukemia
Isoprotenol adalah senyawa kimia yang menyerupai hormon ephineprin/
adrenalin sehingga menghambat rangsangan oleh hormon itu
7
BAB II
BIOMOLEKUL DALAM ORGANISASI MOLEKULER SEL DAN MATERI GENETIK
A. KOMPETENSI
1. mampu menjelaskan hierarki organisasi molekuler sel 2. mampu menjelaskan pembentukan makromolekul
3. mampu menjelaskan keterkaitan pembentukan makromolekul penting dalam sel
B. ELEMEN KOMPETENSI
2. mendeskripsikan hierarki organisasi molekuler sel 3. mendeskripsikan pembentukan makromolekul
4. mendeskripsikan keterkaitan pembentukan makromolekul penting dalam sel
C. MATERI
Biomolekul organisme hidup disusun dalam suatu hirarhi menurut kekompleksan molekul yang meningkat. Semua biomolekul organik diturunkan dari prekursor yang sangat sederhana, berbobot molekul rendah dan terdapat di lingkungan, terutama karbon dioksida, air dan nitrogen atmosfer. Prekursor-prekursor ini melalui serangkaian intermediet-intermediet metabolik diubah oleh benda hidup menjadi biomolekul building block, senyawa organik yang berbobot molekul lebih besar. Molekul-molekul building block ini selanjutnya saling berikatan kovalen satu dengan lainnya membentuk makromolekul sel yang relatif berbobot molekul lebih tinggi. Jadi, asam amino merupakan building block dari protein, nukleotida merupakan building block dari asam nukleat, monosakarida merupakan building block dari kebanyakan lipid.
8
ikatan nonkovalen tersebut besar, maka kompleks supramolekul seperti ribosom stabil pada kondisi biologis. Lagi pula karena penggabungan interaksi nonkovalen makromolekul ke dalam kompleks supramolekul sangat spesifik, sehingga menghasilkan ketepatan geometri atau komplementer diantara bagian-bagian komponennya. Misalnya. Struktur tiga dimensi ribosom sangat teratur dan spesifik, sesuai dengan fungsi kompleksnya dalam translasi informasi genetik ke dalam Struktur protein.
Akhimya, pada level tertinggi organisasi dalam hirarhi struktur sel, berbagai kompleks supramolekul dan sistem bergabung lebih lanjut menjadi organel - inti, mitokondria dan kloroplas, dan menjadi Struktur intrasel dan "inclusion" - lisosom "microbodies" dan vacuola.
Tabel 2.1 Hierarki organisasi molekuler sel
Sel
Organel Inti, mitokondria, kloroplas, badan golgi Kumpulan Supramolekul
(bobot molekul 106-109
Ribosom, kompleks enzim, sistem kontaktil, mikrotubul
Makromolekul (103-109) Asam nukleat, protein, lipid, polisakarida Building blok (bobot molekul
100-350)
Nukleotida, asam amino, monosakarida, asam lemak, gliserol
Intermediet metabolik (bobot molekul 50-250)
Piruvat, sitrat, malat, gliseraldehid-3 fospat
Prekusor dari lingkungan (bobot molekul 14-88)
Karbondioksida, amonia, air, nitrogen
D. MATERI GENETIK
9
Pada prokariot, karena tidak memiliki membran inti, maka materi genetik (kromosom) berada di sitoplasma sehingga proses metabolismenya berbeda dengan dengan eukariot.Eukaryot materi genetik terdapat di nukleus atau inti sel dan sebagian kecil di mitokondria (DNA mitokondria), dan pada sel tumbuhan terdapat di kloroplas. Untuk memahami struktut materi genetik di prokariot dan eukariot, maka perhatikan gambar di bawah ini.
B
Gambar 2.1. A: sel prokariot (materi genetik berupa nukleoid, tanpa dikemas di dalam membran inti; B: sel eukariot (materi genetik terkemas di dalam membran inti).
Untuk mempermudah memahami istilah dan esensi materi genetik, maka fokus bahasan ini lebih ke struktur materi genetik eukariotik karena organisasinya lebih kompleks dibandingkan dengan prokariotik.Untuk materi genetik prokariotik akan dibahas lebih detail di kajian genetika. Perhatikan gambar 2.2 di bawah ini.
Gambar 2. Organisasi materi genetik eukariot
2
atas struktur protein histon-non histon dan rantai DNA.
Istilah yang berikutnya adalah gen, yaitu satu segmen DNA yang menjalankan fungsi untuk mengkode suatu ekspresi (mengkode suatu sifat). Ekspresi ini di dalam istilah umum dalam bentuk protein, karena gen mengkode urutan asam amino. Untuk lebih memahami hal ini akan dibahas secara ringkas tentang struktur asam nukleat (DNA/RNA), gen dan kode genetik.
1. Struktur DNA dan RNA
Data difrasiX-ray Nampak bahwa DNA adalah berbentuk Kristal.Sesuai yang diusulkan oleh James Watson dan Francis Crick pertama kali pada tahun 1953.Oleh Watson dan Francis Crick diusulkan struktur molekul DNA merupakan double heliksdari rantai polinukleotida.Model ini menunjukkan bahwa DNA merupakan model berbentuk 2 helik dengan complementer antiparalel, yang berhubungan melalui ikatan hidrogen hal ini karena basa-basa purin dan pirimidin mengandung gugus fungsi yang memungkinkan terjadinya ikatan hidrogen.Hal ini sangat menentukan struktur asam nukleat. Secara eksperimen diketahui bahwa pada molekul DNA konsentrasi Adenin (A) adalah sebanding dengan Timin (T) sedangkan konsentrasi Sitosin (C) sebanding dengan Guanin (G). Hal ini adalah bahwa A berpasangan dengan T, sedangkan G berpasangan dengan C. berdasarkan hal ini menurut Watson dan Crick, bahwa ikatan G dan C melibatkan 3 ikatan hydrogen, sedangkan antara a dan T mengandung 2 ikatan hydrogen. Hal ini membuat pasangan basa G-C lebih stabil dibandingkan pasangan basa A-T.
DNA terdiri dari 2 utas benang polipeptida yang saling berpilin (double helix=berpilin ganda).Materi genetik, DNA, selalu dalam keadaan aktif. Aktivitas ini berhubungan dengan ekspresi gen dan juga aktivitas tambahan seperti replikasi, perbaikan dan rekombinasi.
2
Sekarang terbukti bahwa molekul DNA dengan urutan nukleotidanya merupakan penentu Struktur kimia dasar kehidupan, yaitu protein-protein, khususnya enzim-enzim. Diketahui pula bahwa DNA menyusun gen yang terletak pada tempat-tempat tertentu dalam kromosom; dan materi genetik dapat diwariskan dari satu generasi ke generasi berikutnya.Seutas polipeptida tersusun atas rangkaian nukleotida.Setiap nukleotida tersusun atas:
1. Gugusan gula deoksiribosa,
2. Gugusan asam fosfat yang terikat pada atom karbon (C) nomor 5 dari gula, 3. Gugusan basa nitrogen yang terikat pada atom C nomor 1 gula.
Struktur DNA digambarkan seperti berikut ini.
Gambar 2.3.(a) DNA Heliks Ganda (b) Struktur Kimia DNA Sumber: Campbell (2002:303)
RNA
3
Susunan kimiawi RNA terdiri dari gula pentose (D-ribosa), molekul gugusan pospat dan basa nitrogen yang terdiri dari purin (adenin dan guanin), dan pirimidin (sitosin dan urasil). Dapat dibedakan menjadi RNA genetik dan RNA non genetic. Benang RNA merupakan benang polipeptida tunggal yang tersusun atas:
1. Gugusan gula yang terdiri dari gula ribosa,
2. Gugusan asam fosfat yang terikat pada atom C nomor 5 dari gula, 3. Gugusan basa nitrogen yang terikat pada atom C nomor 1 dari gula. Basa nitrogen RNA tersusun dari:
a. Basa purin, yaitu Adenin (A) dan Guanin (G), b. Basa pirimidin, yaitu Urasil (U) dan Sitosin (C),
a. RNA Genetik
Dapat berbentuk pita tunggal dan dapat pula berbentuk pita ganda tak terpilin virus tumbuhan (misalnya virus mozaik tembakau TMV, virus burik kuning pada
padi, dsb)
Virus hewan (misalnya virus influenza, virus penyebab luka kaki dan mulut pada
sapi, virus poliomyetis, bakteriophag, dsb).
b. RNA Non Genetik
mRNA (pembawa kode genetik), dicetak oleh DNA dalam suatu proses yang
disebut transkripsi. Membawa informasi yang menentukan urutan asam amino protein dari DNA ke ribosom.Disintesis di dalam nukleus melalui transkripsi oleh DNA pencetak, RNA-m mengandung kodon.RNA-m yang panjang dan berfungsi menyusun beberapa rantai polipeptida disebut RNA-m polisistronik.RNA-m di dalam sel eukarotik lebih stabil dibandingkan di dalam sel prokariotik.
tRNA (Menerjemahkan sandi genetika ke dalam urutan basa polipeptida),
dicetak oleh DNA dan menempati sitoplasma. Berperan sebagai molekul adaptor dalam sintesis protein, mentranslasi kodon-kodon RNAm menjadi asam amino. tRNA berfungsi:membawa asam amino yang akan disintesis menjadi polipeptida ke dalam ribosom, menempatkan asam amino di tempat yang tepat pada rantai polipeptida, mendatangi kodon pada RNA-m sambil membawa asam amino.
rRNA (sebagai adaptor), dicetak oleh DNA dan menempati di dalam ribosom.
4
merupakan penyusun ribosom.Ribosom adalah organel sel yang berfungsi untuk mensintesis polipeptida.RNA-r belum diketahui strukturnya secara jelas. RNA-r berfungsi sebagai adaptor atau penyelaras pada proses sintesis polipeptida.
2. Gen dan Kode genetik
Gen tertentu membawa informasi yang dibutuhkan untuk membuat protein dan informasi itulah yang disebut sebagaikode genetik (kodon). Dengan kata lain, kode genetik adalah cara pengkodean urutan nukleotida pada DNA atau RNA untuk menentukan urutan asam amino pada saat sintesis protein. Informasi pada kode genetik ditentukan oleh basa nitrogen pada rantai DNA yang akan menentukan susunan asam amino (Gurungeblog, 2008, Levine, tanpa tahun). Kode genetik dapat dilihat pada tabel 1 di bawah ini.
Tabel 1. memperlihatkan "kamus" lengkap dari kode genetika. Kode itu mengandung U dan bukan T dalam triplet-triplet. Ini disebabkan oleh fakta bahwa para ahli genetika memandang triplet-triplet yang dibawa oleh molekul-molekul ARNd sebagai komponen-komponen kode genetika dan bukan sebagai-rangkatan dalam DNA sendiri. Hal ini didapatkan dari penelitian terhadap ARNd yang menghasilkan pemecahan kode. Tiap triplet yang mewakili informasi bagi suatu asam amino tertentu dinyatakan sebagai kodon.
5
dalam rantai polipeptida (Stansfield, dkk., 2006). Penetapan triplet kodon didasarkan atas 20 asam amino penyusun protein dan empat macam basa penyusunnya, sehingga secara matematik apabila menggunakan 2 basa hanya menghasilkan 16 (42) kode genetik tidak cukup mewakili 20 asam amino dan 64 (43) kode genetik apabila menggunakan 3 basa. Penggunaan triplet kodon sebagai kombinasi kode genetik menyebabkan satu macam asam amino dapat disandi oleh lebih dari satu triplet kodon, sebagai contoh leusin dapat dikode oleh 6 kodon berbeda pada mRNA. Keadaan ini disebut kode genetikbersifat degenerate (Brown, 2002; Stansfield, dkk., 2008; Pai, 2005; Levine, tanpa tahun). Akibat dari degenerasi kode ini, banyak perubahan (mutasi) dapat terjadi pada sebuah gen tanpa mempunyai efek terhadap komposisi asam amino pada produk gen tersebut. Perubahan tersebut disebut mutasi bisu (silent) (Stansfield dkk., 2008).
Pada tahun 1968 Nirenberg, Khorana dan Holley menerima hadiah Nobel untuk penelitian mereka yang sukses menciptakan kode-kode genetik yang hingga sekarang kita kenal.Seperti kita ketahui asam amino dikenal ada 20 macam.Bagaimana 4 basa nitrogen ini dapat mengkode 20 macam asam amino yang diperlukan untuk mengontrol semua aktivitas sel?
Para peneliti melakukan penelitian pada bakteri E. Coli. Mula-mula digunakan basa nitrogen singlet maka diperoleh 4 asam amino saja yang dapat diterjemahkan padahal ke 20 asam amino ini harus diterjemahkan semua agar protein yang dihasilkan dapat digunakan, kemudian para ilmuwan mencoba lagi dengan kodon duplet dan untuk menterjemahkan 16 asam amino ini pun belum cukup juga. Kemudian dicoba dengan triplet dan dapat menterjemahkan 64 asam amino. Hal ini wajar sekalipun melebihi 20 asam amino toh dari 64 asam amino yang diterjemahkan ada yang memiliki fungsi yang sama diantaranya (kodon asam asparat (GAU dan GAS) sama dengan asam asam tirosin (UAU, UAS) dan sama juga dengan triptopan (UGG). Bahkan ini sangat menguntungkan pada proses pembentukkan protein karena dapat menggantikan asam amino yang kemungkinan rusak. Selain itu, dari 20 asam amino diantaranya ada yang berfungsi sebagai agen pemotong gen atau tidak dapat bersambung lagi dengan doubel helix. Asam amino yang berfungsi sebagai agen pemotong gen diantaranya (UAA, UAG, UGA). Beberapa sifat dari kode triplet diantaranya:
6
SS memperinci prolin, (SSU, SSS, SSA dan SSG) semua kodon yang diawali dengan AS memperinci treosin (ASU, ASS, ASA, ASG).
2. Tidak tumpang tindih, artinya tidak satu basa tunggalpun yang dapat mengambil bagian dalam pembentukan lebih dari satu kodon, sehingga 64 itu berbeda-beda nukleotidanya.
3. Kode genetik dapat mempunyai dua arti yaitu kodon yang sama dapat memperinci lebih dari satu asam amino.
4. Kode genetik itu ternyata universal. Ini berarti, bahwa kode bagi suatu asam amino tertentu pada bakteri juga memberi kode bagi asam amino yang sama dalam organisma-organisma kompleks. Akhir-akhir ini, sifat universal dari kode telah dimanfaatkan sebaik-baiknya dalam teknik rekayasa genetika, yang memungkinkan para ilmuwan memasukkan DNA manusia ke dalam bakteri. Gen-gen manusia itu ditranskripsi, dan protein-protein manusia disintesa oleh bakteri (Elrod, dkk., 2007; Gurungeblog, 2008).
Kebanyakan organisme mempunyai kode genetik yang sama (kode genetik bersifat universal). Kecuali pada genom mitokondria yang menggunakan kode genetik
tidak standard.Kodon 5′-UGA-3′ yang biasanya mengkode untuk terminasi namun pada
mitokondria manusia, 5′-UGA-3′ mengkode untuk triptofan.Penggantian kodon terminasi juga di temukan pada genom nuklear eukariotik tingkat rendah.Kodon
terminasi 5′-UGA-3′ berubah menjadi selenosistein yang tetap digunakan sebagai
kodon terminasi. Pembeda antara selenosistein dengan kodon 5′-UGA-3′ adalah adanya hairpin loop pada mRNA di daerah downstream kodon selenosistein pada
prokariot dan pada daerah 3′ yang tidak ditranslasi (misalnya bagian mRNA setelah kodon terminasi) pada eukariot. Dua contoh diatas meruntuhkan pendapat bahwa kode genetik bersifat universal karena pada kenyataanya kode genetik bersifat tidak universal (Brown, 2002).
7
bioteknologi.Terdapat beberapa pengecualian mengenai keuniversalan kode genetik ini, yaitu sistem translasi di mana beberapa kodon berbeda dari kodon standar.Contoh utamanya ditemukan dalam eukariot bersel tunggal tertentu, seperti paramecium. Contoh lain ditemukan dalam mitokondria dan kloroplas tertentu, yang mentranskripsi dan mentranslasi gen yang dibawa oleh DNA yang sedikit jumlahnya.
Tiap triplet yang mewakili informasi bagi suatu asam amino tertentu dinyatakan sebagai kodon.Kode genetika bersifat degeneratif dikarenakan 18 dan 20 macam asam amino ditentukan oleh lebih dari satu kodon, yang disebut kodon sinonimus.Hanya metionin dan triptofan yang memiliki kodon tunggal.Kodon
sinonimus tidak ditempatkan secara acak, tetapi dikelompokkan.Kodon sinonimus memiliki perbedaan pada urutan basa ketiga (Gurungeblog, 2008).
8
Pustaka
Brown, T. A. 2002. DNA in Genomes, 2nd ed. (Online),
(http://www.ncbi.nlm.nih.gov/books/bv.fcgi?rid=genomes.section.5818, diakses tanggal 26 Oktober 2009).
Campbell, N. A., Jane, B. R., & Lawrence, G. M. 2002.Biologi Edisi Kelima Jilid 1. Jakarta: Erlangga.
Elrod, S. & W. Stansfield. 2007. Genetika Edisi Keempat. Jakarta: Erlangga. Gurungeblog. 2008. Kode Genetik.(Online),
(http://gurungeblog.wordpress.com/2008/11/15/kode-genetik/, diakses pada tanggal 26 Oktober 2009).
Levine, L. Tanpa tahun.Biologi of The Gene.
Murakami, Kazuo dan Mizan, 2007.Keajaiban Kode Genetik. (Online), http://amanah-land.blogspot.com/2007/08/keajaiban-kode-genetik.html, diakses pada tanggal 26 Oktober 2009).
Pai, A.C. 1985. Dasar-Dasar Genetika Edisi Kedua.Diterjemahkan oleh Muchidin Apandi.Foundation Of Genetics. Jakarta: Erlangga.
9
BAB III
ASAM AMINO DAN PROTEIN
I. ASAM AMINO
A. KOMPETENSI:
1. mampu menjelaskan ciri-ciri asam amino 2. mampu menjelaskan reaksi kimia asam amino 3. mampu menjelaskan klasifikasi asam amino 4. mampu menjelaskan klasifikasi protein
5. mampu menjelaskan protein dalam fungsi biologi 6. mampu menjelaskan cara pemisahan protein
B. ELEMEN KOMPETENSI
1. mendeskripsikan dua level yang dipelajari dalam biokimia 2. mendeskripsikan perkembangan ilmu biokimia
3. mendeskripsikan kegunaan ilmu biokimia dalam kehidupan sehari-hari 4. mendeskripsikan fungsi protein dalam kehidupan
5. mampu menjelaskan fungsi protein dalam biologi dan kehidupan 6. mampu menjelaskan cara pemisahan protein
C. MATERI
1. Ciri-ciri Asam Amino
Asam amino adalah senyawa penyusun protein.Asam amino mempunyai satu gugus karboksil dan satu gugus amino.Pada umumnya gugus amino terikat pada
posisi dari gugus karboksil.Struktur:satu atom C sentral yang mengikat secara kovalent:
gugus amino,
gugus karboksil,
satu atom H dan
rantai samping (gugus R)COOH
10
Asam amino adalah suatu molekul organik dengan sedikitnya mengandung satu gugus karboksil (asam organik) dan paling sedikit satu gugus amino (basa organik). Dengan penggunaan definisi ini, beratus-ratus asam amino telah diketahui kehadirannya pada sel tumbuhan dan sel hewan. Di dalam bab ini, kita memfokuskan pada kelompok asam amino (20 jenis) yang dikode untuk pembentukan protein. Satuan asam amino adalah suatu struktur umum yang terdiri atas karbon (C) pusat (karbon alfa) yang dikelilingi oleh suatu hidrogen, gugus karboksil, gugus amino, dan suatu rantai sisi R. Oleh karena penempatan gugus-gugus fungsional ini ada di sekitar
α-carbon pusat, maka mereka sering disebut allfa-asam amino. Sifat alami rantai sisi, yang dapat bervariasi dari suatu atom hidrogen sederhana ke sistem kompleks, menentukan reaktivitas biologi dan kimia yang unik dari tiap asam amino.
Tabel 3.1 Asam Amino Essensial dan Nonesssensial
Essential Nonessential
Argininea Methionineb Alanine Glutamine
Histidine Phenylalaninec Aspartate Glycine
Isoleucine Threonine Asparagine Proline
Leucine Tryptophan Cysteine Serine
Lysine Valine Glutamate Tyrosine
11
kebutuhannya.Asam amino karnitin juga bersifat "setengah esensial" dan sering diberikan untuk kepentingan pengobatan.
Penamaan asam amino, didasarkan pada struktur D – gliseraldehid jika gugus NH3+ terletak disebelah kanan diberi awalan D, jika NH3+ dikiri diberi awalan L.
Semua asam amino yang ada di alam dalam protein mempunyai konfigurasi L. Ada beberapa asam amino yang penting dalam struktur dan metabolisme mempunyai konfigurasi D, yaitu asam D-alanin dan D-glutamat yang merupakan komponen penyusun dinding sel bakteri tertentu.Penulisan asam amino (20 asam amino yang
umum) dapat disingkat dengan 3 huruf.Misal : Serine Ser, Glysin gly.
Karbon alfa yang tetrahedral dalam tiap-tiap asam amino, kecuali glisin (R = H), mempunyai empat gugus atom yang berbeda yang terikat padanya (karbon alfa), karenanya ini merupakan suatu chiralcenter (chiral memusat). Konsekuensi penting dari pengaturan ini adalah keberadaan dua nonsuperimposable stereoisomers (enantiomers), yang disebut D- dan L- asam amino (Gambar ....). D dan L isometri adalah gambaran cermin dari satu sama lain. Tiap stereoisomer menunjukkan keaktifan optis, mereka berputar pada sudut plane-polarized light. Sudut putar adalah kebalikan untuk keduanya. Walaupun kedua isometri D dan L asam amino ada secara alami, namun hanya L isometri yang digunakan sebagai pembangun blok protein. Peran kritis yang dimainkan oleh isometri L- asam amino pada fungsi protein sedang dimulai untuk dipelajari dan dipahami. Ahli biokimia menemukan peningkatan sejumlah D- asam amino aspartat di dalam protein yang terakumulasi di dalam gigi dan lensa mata manusia seiring dengan bertambahnya umur. Kenyatannya, pada protein penyimpan, konversi asam amino L-aspartat ke asam amino D-aspartat (racemization
12
Gambar 3.1 D dan L enantiomer asam amino fenilalanin. Catatan bahwa kedua struktur ada dalam bayangan cermin, garis putus-putus menandai adanya ikatan gugus dan menandai adanya ikatan ke karbon alfa
Ke 20 alfa asam amino bentuk murni adalah putih, kristal, padat dengan high-melting (daya leleh tinggi) . Zat ini adalah larut dalam air dan dalam bahan pelarut organik seperti aseton, cloroform, dan eter tidak dapat larut. Larutan yang mengandung asam amino dapat menghantarkan arus elektrik. Ciri-ciri yang dimiliki seperti diuraikan di atas diperkirakan dari senyawa ionik atau garam, tetapi corak seperti tidak dapat ditunjukkan di susunan rumus yang umum seperti pada Gambar 8.1 dan 8.2. Pada pH fisiologis ( sekitar 7.4), asam amino bersifat ionik dipolar (Gambar 8.3); karena mempunyai suatu muatan positif dan muatan negatif pada molekul yang sama. Bentuk ini kadang-kadang disebut ’zwitterions’ dari istilah Jerman yang berarti "inner salt".
Seperti ditunjukkan pada reaksi di bawah, gugus karboksil melepaskan proton pada pKa 2.3 dan gugus NH pada pKa 9.7. Diasumsikan bahwa rantai sisi R tidak bermuatan dan muatan pada bentuk zwitterion (struktur B) asam amino (pada pH netral) adalah nol. Pemahaman terhadap ciri-ciri dasar asam amino adalah sangat penting, sebab dapat meramalkan bentuk ionik utama dari suatu asam amino pada sejumlah nilai pH. Prediksi terhadap muatan elektrik asam amino menjadi sangat penting ketika kita mendiskusikan struktur dan fungsi protein.
Klasifikasi asam amino
Klasifikasi ini didasarkan atas sifat gugus R-nya, dibedakan atas 4 macam asam amino: bersifat non polar alifatik, aromatik, polar tidak bermuatan dan polar bermuatan. Struktur lengkap keempat macam asam amino dapat dilihat pada gambar di bawah. Asam amino diklasifikasikan berdasarkan polaritas rantai samping (R).
1. Asam Amino dengan Rantai Sisi Nonpolar dan Alifatik
a. Glisin dan Prolin
13
sedikit fleksibilitas dalam struktur protein. Atom nitrogen melekat secara kovalen ke rantai sisi, membentuk sebuah einem yang kaku.
b. Alanin dan asam amino rantai bercabang
Alanin dan asam amino rantai bercabang (valin, leusin, dan isoleusin) memilikirari sisi alifatik, nonpolar, dan berukuran besar. Alanin mengandung sebuah gugus metil. Valin, leusin, dan isoleusin memiliki rantai bercabang yang sangat hidrofobik.
Gambar 3.4Asam amino dengan rantai sisi nonpolar dan alifatik
2. Asam Amino dengan Rantai Sisi Aromatik
14
Gambar 3.5 Asam amino dengan rantai sisi aromatik
3. Asam Amino dengan Rantai Sisi Polar dan Tidak Bermuatan
Gugus asam amino bersifat polar, tetapi tidak bermuatan. Asam amino ini bersifat hidrofilik (menyukai air) dan dalam larutan encer sering ditemukan di permukaan protein globular tempat dimana asam tersebut berinteraksi dengan air. Rantai sisi serin dan treonin mengandung gugus amida dari asam amino aspartat dan glutamate. Keduanya polar karena adanya karbonil dan atom nitrogen di gugus amidanya- Sistein dan metionin relative polar karena keduanya masing-masing mengandung sebuah atom sulfur. Namun, keduanya lebih hidrofobik daripada asam amino lain dalam kelas ini.
Adanya gugus hidroksil pada serin dan treonin dan gugus amida padaasparagindan glutamin memungkinkan asam-asam amino tersebut membentuk ikatan hidrogen satu sama lain, dengan air, atau dengan senyawa polar lain yang berikatan dengan protein. Walaupun tirosin bersifat hidrofobik, gugus hidroksimya memungkinkan asam amino ini membentuk ikatan hidrogen.
15
Gambar 3.6Asam amino dengan rantai sisi polar dan tidak bermuatan
4. Asam Amino dengan Rantai Sisi Bermuatan
Pada pH faali, 5 asam amino memiliki muatan pada rantai sisi mereka. Dua asam amino lainnya tidak bermuatan, tetapi menjadi bermuatan negatif pada rentang pH yang lebih tinggi, Gugus yang bermuatan positif atau negatif dapat berperan dalam interaksi elektrostatik dengan gugus yang bermuatan berlawanan pada asam amino atau pada senyawa lain
a. Aspartat dan Glutamat, Asam Amino Bermuatan Negatif
Asam aspartat dan glutamat mengandung asam karboksilat pada rantai sisi mereka sehingga disebut asam amino asam (acidic). Pada pH faali, proton mengalami disosiasi, dan rantai sisi membawa muatan negatif. Bentuk asam amino yang bermuatan negatif disebut aspartat dan glutamat.
Nilai pKa untuk gugus asam pada rantai sisi asam amino tersebut adalah sekitar 4 (Tabel 8.2). Apabila pH sama dengan pKa. spesies yang mengalami protonisasi(tidak bermuatan) jumlahnya setara dengan jumlah spesies bermuatan negacif, yaitu, jumlah molekul dengan muatan nol pada rantai sisi setara dengan jumlah molekul yang bermuatan negatif. Pada pH kurang dari 4, jumlah molekul yang membawa sebuah proton dan, dengan demikian, tidak bermuatan lebih besar daripada jumlah molekul yang bermuatan negatif. Pada pH di atas 4, molekul yang predominan telah kehilangan sebuah proton, sehingga bermuatan negatif.
b. Arginin, Lisin, dan Histidin, Asam Amino Bermuatan Positif
16
mengandung sebuah gugus amino pada karbon ε (epsilon) (pKa ~ 10,5). Oleh karena itu, asam-asam amino bermuatan positif pada pH fisiologis (~7,4). Histidin memiliki sebuah cincin imida (pKa ~ 6). Dalam protein, pKa gugus imidazol terletak antara 6 dan 7. Oleh karena itu ini menitrasi dalam rentang pH fisiologis, dan perubahan kecil pada pH atau lingkungan lokal dapat mengubah muatan. Sifat ini memungkinkan histidin berperan penting dalam fungsi protein.
c. Sistein dan Tirosin
Rantai sisi sistein (pKa ~ 8,4) dan tirosin (pKa ~ 10,5) sebagian besar mengalami protonisasi di bawah pKa-nya dan tidak memiliki muatan.. Di atas pKa, proton mengalami disosiasi, dan rantai sisi kemudian membawa muatan negatif. Oleh karena itu, dalam protein asam amino ini biasanya tidak bermuatan pada pH fisiologis.
Gambar 3.7Asam amino dengan rantai sisi bermuatan
Metabolisme asam amino
Asam amino yg terbentuk di usus akan diabsorpsi dan dibawa oleh peredaran darah ke dalam sel-sel tubuh. Metabolisme asam amino di dalam sel
katabolisme :
1. katabolisme nitrogen asam amino urea
2. katabolime kerangka karbon asam amino senyawa amfibolik anabolisme sintesis protein
ada 20 macam asam amino dasar: 10 macam adalah asam amino esensial
Sifat Asam Amino
1. Larut dalam air dan tidak larut dalam pelarut non polar seperti eter, aseton, dan kloroform.
17
Karena atom C pusat mengikat empat gugus yang berbeda, maka asam amino—kecuali glisin—memiliki isomer optik: l dan d.
Pada umumnya, asam amino alami yang dihasilkan eukariota merupakan tipe l meskipun beberapa siput laut menghasilkan tipe d. Dindingselbakteri banyak mengandung asam amino tipe d.
3. Zwitter-ion/ Ion Amfoter
Karena asam amino memiliki gugus aktif amina dan karboksil (berupa asam karboksilat) sekaligus, zat ini dapat dianggap sebagai asam dan basa (walaupun pH alaminya biasanya dipengaruhi oleh gugus-R yang dimiliki). Amfoter yang berarti mempunyai sifat basa karena adanya gugus NH2 dan mempunyai sifat asam karena adanya gugus COOH. Dengan demikian dalam larutan ataupun dalam cairan darah asam amino merupakan molekul yang tidak bermuatan.Pada pH tertentu yang disebut titikisolistrik, gugus amina pada asam amino menjadi bermuatan positif (terprotonasi, –NH3+), sedangkan gugus karboksilnya menjadi bermuatan negatif (terdeprotonasi, –COO-).Titik isolistrik ini spesifik bergantung pada jenis asam aminonya.Dalam keadaan demikian, asam amino tersebut dikatakan berbentuk zwitter-ion.
Zwitter-ion dapat diekstrak dari larutan asam amino sebagai struktur kristal putih yang bertitik lebur tinggi karena sifat dipolarnya. Kebanyakan asam amino bebas berada dalam bentuk zwitter-ion pada pH netral maupun pH fisiologis yang dekat netral.
Polipeptida
Ikatan peptida yakni rantai pendek dari dua atau lebih asam amino yang dihubungkan oleh ikatan kovalen.Sel dapat merangkai ke 20 asam amino dalam berbagai kombinasi dan urutan sehingga dapat membuat produk yang sangat bervariasi. Dua α asam amino dapat berikatan bersama membentuk suatu amida atau ikatan peptida. Berdasarkan konvensi ikatan peptida ditulis dengan asam amino yg mempunyai NH3+ bebas (sebelah kiri) dan as.Amino dg gugus COO- bebas (sebelah
18
Gambar 3.8 Reaksi pembetukan ikatan peptida
Reaksi kimiawi di atas menggambarkan hilangnya suatu molekul air, membawa bersama-sama gugus karboksil suatu asam amino dengan amino kelompok lain untuk menghasilkan suatu dipeptide. Reaksi dapat diulangi dengan penambahan asam amino untuk bergabung untuk membentuk suatu tripeptide, tetrapeptide, pentapeptide, dan seterusnya. Masing-masing asam amino asam, ketika bergabung membentuk polypeptide dikenal sebagai suatu residu. Peptides dengan dua hingga sepuluh residu asam amino pada umumnya dinamai oleh awalan ilmu kimia yang umum untuk angka-angka (di-, tri-, tetra-, penta-, hexa-, hepta-, octa-, nona-, dan decapeptide). Produk dengan 10 hingga 100 asam amino disebut polypeptids, sedangkan mereka yang mempunyai lebih dari 100 asam amino disebut protein.
2. PROTEIN
Protein adalah biopolimer yang terdiri dari banyak satuan asam Amino yg dihubungkan oleh ikatan peptide. Beberapa protein merupakan komponen utama dalam jaringan struktur (otot, rambut, kuku, kulit).Kata protein berasal dari bahasa
Mesir “proteus” yang terjemahan kasarnya berarti “yang utama”.Protein adalah sumber
asam-asam amino yang mengandung unsur-unsur C, H, O, dan N ada pula yang mengandung unsur S dan P.Protein tersusun dari beberapa asam amino yang saling berikatan (mempunyai lebih dari 100 asam amino disebut protein).
Kebanyakan peptides dan protein yang diisolasi dari sel dan jaringan tersusun antara 2 hingga 2000 asam amino. Diasumsikan rata-rata bobot molekul dari semua amino adalah 110, bobot molekular kebanyakan peptide dan rantai protein berkisar dari 220 hingga 220.000, walaupun banyak yang lebih besar telah ditemukan.
19
Gambar 3.9Suatu peptide pendek yang menunjukkan jumlah dan arah. Residu ujung amino terminal, selalu digambarkan di sebelah kiri dimulai dengan nomor 1. Struktur pentapeptide ditulis Gly-Glu-Val-Ser-Lys. Panah menandai adanya ikatan peptide.
Klasifikasi Protein
Berdasarkan komposisi
Protein dibagi menjadi dua kelompok utama, yaitu: 1. Protein sederhana
2. Protein konjugasi
Protein sederhana adalah protein yang pada hidrolisihanya menghasilkan asam-asam amino. Kelompok protein ini umumnya mengandung kurang lebih 50% karbon, 7% hidrogen, 23% oksigen, 16% nitrogen dan 0 - 3% sulfur. Kelompok protein konjugasi adalah protein yang pada hidrolisis tidak hanya menghasilkan asam-asam amino, tetapi juga komponen organik dan komponen anorganik lain, yang disebut gugus prostetik dari protein. Berdasarkan sifat kimiia gugus prostetiknya. protein konjugasi dapat dikelompokan menjadi nuleoprotein, lipoprotein. karena mengandung gugus prostetik asam nukleat dan lipid. Juga dikenal fosfoprotein, metalloprotein dan glucoprotein.
Berdasarkan konformasi
Protein dibagi menjadi tiga kelompok utama, yaitu: 1. protein serat,
2. protein globular,
3. protein dengan konformasi antara protein serat dan protein globuar.
20
Sebaliknya dalam protein globular, rantai-rantai polipeptida melipat ke dalam menjadi bentuk globular atau bola yang kompak. Kebanyakan protein globular larut dalam sistem air. Umumnya protein ini mempunyai fungsi yang mobil dan dinamik di dalam sei. Dari sebanyak 2.000 enzim-enzim yang dikenal, sejumlah hormon, serum, albumin dan hemoglobin, semuanya termasuk püten globular. Protein kelompok tiga, protein dengan konformasi antara protein serat dan protein globular, struktumya berbentuk seperti batang menyerupai protein serat tetapi menunjukkan sifat larut dalam larutan garam, seperti protein globular. Contoh myosin yang merupakan komponen dalam otot, fibrionogen merupakan prekursor fibrin yang berperan dalam pembekuan darah.
Berdasarkan strukturnya Protein dibagi menjadi 4 yaitu:
1. Struktur primer adalah struktur rantai polipeptida linier, yang terjadi karena ikatan peptida antara residu asam amino dengan residu asam amino lainnya. Atau urutan asam amino dalam suatu molekul protein. Contoh : Lys-Asp-Gly-Ala-Ala-Glu-Ser-Gly. Struktur primer suatu protein terjadi karena penggabungan "head to tail" satu residu asam amino dengan asam amino berikutnya, melalui eliminasi molekul air dari gugus karboksilat residu asam amino dan gugus a-amino dari residu asam amino berikutnya.
Gambar 3.10 Struktur primer protein
21
pula bahwa 4 atom dari gugus peptida dan 2 atom karbon a terletak dalam satu bidang, sedemikian mpa sehingga atom 0 dari gugus karbonil dan atom H dari gugus -NH- berada pada posisi trans. Penataan planar ini merupakan hasil stabilisasi resonansi ikatan peptida. Dari penemuan ini, tulang punggung rantai polipeptida dapat digambarkan sebagai serangkaian bidang-bidang yang relatif kaku, yang dipisahkan satu dengan lainnya oleh gugus metilen (-CHR-). Dalam tulang punggung rantai polipeptida, satu pertiga dari semua ikatan-ikatan tunggal adalah ikatan-ikatan C-N yang tidak dapat berotasi karena sifat ikatan rangkapnya yang mempengaruhi kekakuan ("constrains") sejumlah konformasi rantai-rantai polipeptida. Struktur sekunder suatu protein distabilkan oleh ikatan hidrogen. Ikatan hidrogen dapat terjadi antara atom H dari gugus -NH- residu asam amino dengan atom O gugus karbonil residu asam amino berikutnya, atau atom O gugus karbonil asam amino ketiga dan seterusnya.
Gambar 3.11 Struktur skunder protein
3. Struktur tersier adaiah Struktur sekunder yang membelok-belok dan melipat-lipat
ke dalam tiga dimensi membeniuk protein globularyang kompak.
Gambar 3.12Struktur tersier protein
22
Gambar 3.13 struktur kuarter protein
Fungsi Protein dalam Fungsi Biologi
1. Protein Regulator / Bioregulator( Hormon) 2. Biokatalisator ( Enzim )
3. Protein Transport: Hb, Alb, Lipoprotein,Transferin, protein integral membran 4. Protein Kontraktil: aktin dan miosin
5. Protein Struktural: kolagen,tubulin,keratin,glikoprotein
6. Protein Pelindung dan pertahanan: Ig, interferon, perforin,IL, fibrinogen 7. Protein Reseptor
Protein menurut klasifikasi berdasarkan peran biologinya dibedakan atas: 1. Enzim
2. Protein struktural 3. Protein imun
4. Protein transport dan cadangan 5. Protein regulator dan reseptor 6. Kontraksi otot dan mobilitas
Denaturasi Protein
23
1. suhu yang tinggi
2. keasaman (perubahan pH yg ekstrim) 3. zat kimia tertentu (urea, deterjen) 4. karena pengaruh mekanik (guncangan) 5. penyinaran/ radiasi UV
6. konsentrasi ion hidrogen yg tinggi
24
BAB IV ENZIM
A. KOMPETENSI:
1. mampu menjelaskan struktur enzim
2. mampu menjelaskan nomenklatur dan klasifikasi enzim 3. mampu menjelaskan kinetika enzim
B. ELEMEN KOMPETENSI
1. mendeskripsikan struktur enzim
2. mendeskripsikan nomenklatur dan klasifikasi enzim 3. mendeskripsikan kinetika enzim
C. MATERI
Enzim merupakan senyawa organik bermolekul besar yang berfungsi untuk mempercepat jalannya reaksi metabolisme di dalam tubuh (organisme) tanpa mempengaruhi keseimbangan reaksi.Enzim adalah suatu zat organik yang dapat mempengaruhi berbagai reaksi kimia yang terjadi dalam suatu makhluk hidup. Enzim berperan untuk mengkatalisis proses kimia (biokimia) dalam makhluk hidup atau dalam sistem biologi. Enzim berfungsi sebagai biokatalisator,artinya mempercepat jalannya reaksi dengan cara menurunkan energi aktivasi. Energi aktivasi adalah energi yang diperlukan untuk mengaktifkan suatu reaktan sehingga dapat bereaksi untuk membentuk senyawa lain.
Tanpa adanya enzim biasanya reaksi kimia akan berlangsung sangat lambat, bahkan mungkin tidak dapat terjadi. Reaksi yang dibantu dengan katalis enzim laju reaksinya lebih tinggi 106-1012 kali.Kerja enzim sangat spesifik baik jenis maupun substratnya, artinya satu enzim hanya menjalankan satu fungsi saja.Misalnya adalah
enzim α-Amylase yang bekerja spesifik dalam mulut, enzim ini terdapat bersama
dengan air liur (saliva), enzim α-Amylase berperan dalam melakukan hidrolisis awal makanan terutama yang mengandung pati.Enzim tidak ikut bereaksi, struktur enzim tidak berubah baik sebelum dan sesudah reaksi tetap.
25
oleh suhu.Temperatur yang tinggi (lebih dari 40°C) dapat menyebabkan kerja enzim tidak aktif (non-aktif) bahkan kerusakan enzim (denaturasi). Faktor-faktor yang dapat mempengaruhi fungsi enzim diantaranya adalah:
1. Suhu
Suhu dapat mempengaruhi reaksi kimia, maka reaksi menggunakan katalis enzim dapat dipengaruhi oleh suhu. Di samping itu, karena enzim adalah suatu protein maka kenaikan suhu dapat menyebabkan denaturasi dan bagian aktif enzim akan terganggu sehingga konsentrasi dan kecepatan enzim berkurang. Suhu yang tinggi (>40 oC) akan mengakibatkan hilangnya fungsi kerja enzim karena mengalami denaturasi. Enzim bekerja secara optimal pada suhu sekitar 30-40 oC, suhu > 60 oC sudah mulai mengalami denaturasi dan akan terdenaturasi sempurna pada suhu 90-100 oC. Kenaikan suhu 10˚C (sampai 40˚C), kecepatan reaksi naik 2 x lipatnya
dan reaksi terhambat dan berhenti pada 60˚C.
2. Ph
Umumnya enzim efektifitas maksimum pada pH optimum, yang lazimnya berkisar antara pH 4,5-8.0. Pada pH yang terlalu tinggi atau terlalu rendah umumnya enzim menjadi non aktif secara irreversibel karena mengalami denaturasi protein.setiap enzim mempunyai pH optimum utk bekerja. contoh : pepsin pH 2, amylase pH 7.0
3. Konsentrasi enzim
Seperti pada katalis lain, kecepatan suatu reaksi yang menggunakan enzim tergantung pada konsentrasi enzim tersebut. Pada suatu konsentrasi substrat tertentu, kecepatan reaksi bertambah dengan bertambahnya konsentrasi enzim. 4. Konsentrasi substrat
Hasil eksperimen menunjukkan bahwa dengan konsentrasi substrat akan menaikkan kecepat reaksi. Akan tetapi, pada batas tertentu tidak terjadi kecepatan reaksi, walaupun konsenrasi substrat diperbesar.
5. Pengaruh aktifator
26
6. Zat-zat penghambat/inhibitor
Hambatan atau inhibisi suatu reaksi akan berpengaruh terhadap penggabungan substrat pada bagian aktif yang mengalami hambatan. Aktifitas enzim dapat dikontrol sesuai dengan kebutuhan organisme itu sendiri. Contoh: enzim yg mengkatalisis reaksi pertama pada suatu siklus biosintesis biasanya di hambat oleh produk akhirnya (feedback inhibition).Beberapa enzim disintesis dalam bentuk tidak aktif dan akan diaktifkan oleh kondisi dan waktu yang sesuai (misalnya pepsinogen dirubah menjadi pepsin) disebut sebagai enzim allosterik, dan prekursor yang tidak aktif disebut sebagaizymogen.
SUSUNAN ENZIM
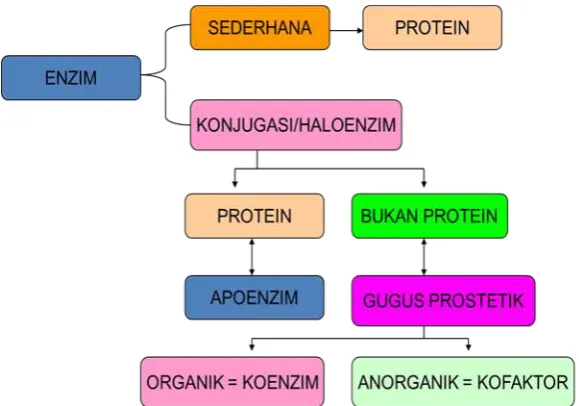
[image:38.595.151.441.405.608.2]Komponen utama enzim adalah protein. Protein yang sifatnya fungsional, bukan protein struktural dan tidak semua protein bertindak sebagai enzim. Berikut ini adalah bagan susunan enzim.
Gambar 4.1 Bagan Susunan Enzim
27
membuktikan bahwa enzim adalah protein. Selanjutnya dalam kurun waktu 1930 sampai 1936, J. Northrop berhasil mengkristalkan pepsin, tripsin dan kimotripsin.Sampai kini telah dikenal ribuan enzim.
Walaupun kebanyakan enzim yang berkaitan dengan metabolisme sel telah diidentifikasi, tetapi masih banyak persoalan yang perlu dipecahkan, termasuk kontrol genetik pada biosintesis enzim, pengendalian mekanisme aktivitas enzim secara molekular dan peran enzim bentuk ganda dalam perkembangan dan diferensiasi. Bagian-bagian enzim adalah:
1. Holoenzim
2. Apoenzim/ apoprotein 3. Gugus prostetik 4. Koenzim
5. Kofaktor
Secara keseluruhan enzim (holoenzim) memiliki dua bagian utama yaitu bagian protein (apoenzim) dan bagian non protein (proteolitik).Apoenzim merupakan suatu polipeptida yang memiliki struktur kuartener atau struktur tersier dengan urutan dan komposisi asam amino tertentu dan rantai polipeptida tersebut distabilkan oleh ikatan kimia yang terjadi dari gugus samping yang terdapat dalam asam aminonya.Ikatan yang terjadi adalah ikatan kimia sulfida, ikatan hidrogen, ikatan elektrostatik, ikatan non polar dan ikatan van der Waals.Gugus prostetik berfungsi sebagai pengaktivasi enzim sehingga reaksi lebih mudah berlangsung.
Gugus prostetik biasanya berasal dari senyawa-senyawa organik yang disebut kofaktor sebagai contoh ion Fe2+, Zn2+, Cu2+. Sedangkan gugus prostetik dari dari molekul organik kompleks disebut dengan koenzim, contohnya NADH, FADH dan CoASH. Dalam reaksi biokimia, beberapa enzim diaktifkan oleh koenzim maupun ion logam sekaligus, sebaliknya ada juga enzim yang diaktifkan oleh koenzim saja atau ion logam saja.Seperti halnya protein lain, enzim memiliki BM antara 12,000 – 1 juta kd. Beberapa enzim tidak membutuhkan molekul kimiawi lain untuk aktifitasnya, beberapa membutuhkan kofaktor / koenzim. Kofaktor adalah ion-ion inorganik yg dibutuhkan enzim untuk melakukan fungsinya, sedang koenzim adalah molekul organik (komplek) yang dibutuhkan enzim untuk melakukan fungsinya. Berikut ini adalah contoh koenzim:
28
4. Cytokrom: cytokrom a, a3, b, b6, c, dan f 5. Plastoquinon, plastosianin, feredoksin
6. ATP: senyawa organik berenergi tinggi, mengandung 3 gugus P dan adenin ribose
Sifat utama enzim ada 3 yaitu: 1. Kemampuan katalitiknya 2. Spesifisitas
3. Kemampuan untuk diatur (regulasi)
Sifat enzim: enzim dibentuk dalam protoplasma sel, enzim beraktifitas di dalam sel tempat sintesisnya (disebut endoenzim) maupun di tempat yang lain diluar tempat sintesisnya (disebut eksoenzim) dan sebagian besar enzim bersifat seperti beikut ini:
1. Enzim bersifat koloid, luas permukaan besar, bersifat hidrofil
2. Dapat bereaksi dengan senyawa asam maupun basa, kation maupun anion 3. Enzim sangat peka terhadap faktor-faktor yang menyebabkan denaturasi
protein misalnya suhu, pH dll
4. Enzim dapat dipacu maupun dihambat aktifitasnya
5. Enzim merupakan biokatalisator yang dalam jumlah sedikit memacu laju reaksi tanpa merubah keseimbangan reaksi
6. Enzim tidak ikut terlibat dalam reaksi, struktur enzim tetap baik sebelum maupun setelah reaksi berlangsung
7. Enzim bermolekul besar 8. Enzim bersifat khas/spesifik
NOMENKLATUR DAN PENGGOLONGAN ENZIM
sehari-29
hari, satu nama sistematik yang mengidentifikasi reaksi yang dikatalisisnya, dan satu nomor golongan yang dipakai apabila diperiukan identifikasi suatu enzim yang akurat,
misainya dalam majalah penelitia
n intemasional, abstrak atau indeks. Contoh
reaksi enzimatik :
ATP + kreatin ADP + fosfokreatin
Nama rekomendasi enzim, yang biasa digunakan adaiah kreatin
kinasenama sistematik, berdasarkan reaksi yang dikatalisisnya adaiah ATP:
kreatin fosfotransferase.Nomor golongannya adaiah EC.2.7.3.2.dimana EC
menyatakan "Enzyme Comission", dengan digit pertama (2) menunjukkan
nama golongan (transferase), digit ke dua (7) menunjukkan nama sub golongan
(fosfotransferase), digit ke tiga (3) menunjukkan nama sub sub golongan
(fosfotransferase dengan satu gugus nitrogen sebagai aseptor) dan digit ke
empat (2) menunjukkan kreatin kinase. Di sini penamaan hanya ditulis dalam
dua digit, untuk penamaan empat digit dapat lihat referensi lain.
Mis:
1.1.1.1 Alkohol : NAD Oksidoreduktase
= alkohol dehidrogenase
1.
INFORMASI TAMBAHAN
Mis:
1.1.1.37 L-MALAT : NAD OKSIDOREDUKTASE
(decarboxylating)
L-MALAT + NAD
+
PIRUVAT + CO
2+ NADH + H
+
1.1.1.37 L-MALAT : NAD OKSIDOREDUKTASE
L-MALAT + NAD
+
OKSALOASETAT + NADH + H
+2. NOMOR KODE SISTEMATIK
-D-GLUKOSA
Heksokinasi/Glukokinase
-D-GLUKOSA 6-P
ATP
ADP
30
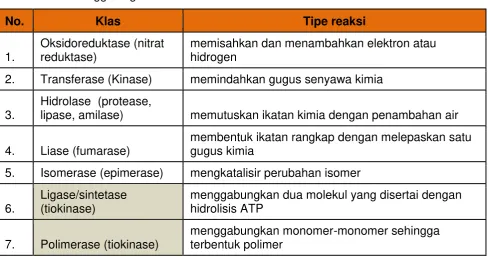
[image:42.595.66.557.180.438.2]Tabel di bawah ini menunjukkan bahwa semua enzim yang diketahui
dapat diklasifikasikan ke dalam 6 kategori (ada yang berpendapat digolongkan
menjadi 7 golongan) dasar perbedaanya terletak pada Tabel di bawah ini.
Tabel 4.1 Penggolongan Enzim
No.
Klas
Tipe reaksi
1.
Oksidoreduktase (nitrat
reduktase)
memisahkan dan menambahkan elektron atau
hidrogen
2.
Transferase (Kinase)
memindahkan gugus senyawa kimia
3.
Hidrolase (protease,
lipase, amilase)
memutuskan ikatan kimia dengan penambahan air
4.
Liase (fumarase)
membentuk ikatan rangkap dengan melepaskan satu
gugus kimia
5.
Isomerase (epimerase)
mengkatalisir perubahan isomer
6.
Ligase/sintetase
(tiokinase)
menggabungkan dua molekul yang disertai dengan
hidrolisis ATP
7.
Polimerase (tiokinase)
menggabungkan monomer-monomer sehingga
terbentuk polimer
Ikatan Substrat
–
Enzim
Sisi aktif Enzim
Katalisis enzimatik dimulai dengan adanya kombinasi dari suatu molekul
enzim dengan suatu molekul substrat untuk membentuk suatu kompleks ( E+ S
= E). Molekul substrate umumnya lebih kecil dibanding enzim mengikat pada
suatu daerah spesifik di dalam enzim yang disebut daerah sisi aktif (active site).
Gagasan untuk daerah sisi aktif sebagai suatu konsep yang muncul dari
pengamatan kinetika Michaelis-Menten dan studi struktur protein. Sisi aktif
adalah suatu tempat atau celah di dalam struktur tiga dimensi enzim dimana
peristiwa katalitis terjadi.
31
2. Sisi aktif merupakan daerah yang sangat kecil pada struktur 3 dimensi
enzim.
3. Sisi aktif berisi asam amino
4. Sisi aktif ini berikatan dengan substrat secara lemah dalam ikatan non
kovalen dan dalam interaksi reversibel. Interaksi ini seperti ditunjukkan gambar
4.2.
Gambar4.2
Sisi aktif enzim dan asam amino yang terlibat
Pada tahun 1890, Emil Fisher menggambarkan model ikatan ES seperti ikatan
antara kunci dengan anak kunci dan model inducet-fit sebagaimana
diilustrasikan pada gambar 4.2.
Gambar 4.3
Model ikatan antara substrat
–
enzim (a) lock and key model,
(b) inducet-fit model
32
mengaktipkan ke arah reaksi. Ketika substrat sepenuhnya berikatan dengan
sisi aktif, terbentuk status transisi untuk suatu reaksi (Gambar 4.3).
Penghambatan Reaksi Enzimatis
Penghambatan aktifitas enzim ada dua tipe:
1. Kompetitif: zat penghambat mempunyai struktur yang mirip dengan substrat
sehingga dapat bergabung dengan sisi aktif enzim. Terjadi kompetisi antara
substrat dengan inhibitor untuk bergabung dengan sisi aktif enzim (
misal feed
back effect
). Inhibitor bersaing dgn substrat untuk terikat pd sisi aktif. Biasanya
inhibitor berupa senyawa yg menyerupai substratnya, & mengikat enzim
membentuk
komplek
EI,
karena
terikat
secara
reversible
maka
penghambatannya bias, yaitu ketika ditambah substrat maka penghambatan
berkurang.
2. Non kompetitif: zat penghambat menyebabkan struktur enzim rusak
sehingga sisi aktifnya tidak cocok lagi dengan substrat. Inhibitor terikat pada sisi
lain dari enzim (bkn sisi aktif)jadi tidak memblok pembtkan enzim-substrat
komplek. Enzim menjadi tidak aktif ketika inhibitor terikat walau enzim mengikat
substrat
Inhibitor mengurangi konsentrasi enzim yang aktif, sehingga mempengaruhi
Vmax
–
nya.
KINETIKA ENZIM
a. Persamaan Michaelis
–
Menten
Walaupun enzim mempunyai ciri katalisator organik dan anorganik,
mereka mempunyai ciri kinetik unik yang memedakannya dari katalisator lain.
Satu pengamatan awal pada pengaruh konsentrasi substrat yang tidak biasa
pada tingkat reaksi enzyme-catalyzed itu. Tingkat reaksi enzyme-catalyzed
dipelajari dengan pencampuran substrate dan enzim dalam suatu larutan
buffer yang sesuai (untuk memelihara agar pH konstan) dan pada suatu
33
Tingkat awal ( Vo, kecepatan awal) ditentukan selama beberapa menit awal
reaksi dengan mengukur baik pengurangan konsentrasi komponen reaktan
maupun peningkatan konsentrasi produk.
Satu set tabung disiapkan, masing-masing berisi suatu larutan
penyangga/bufeer dengan meningkatkan jumlah substrat. Suatu jumlah enzim
yang konstan ditambahkan untuk masing-masing tabung, rerata reaksi diukur,
dan suatu grafik rerata reaksi terhadap konsentrasi substrat dapat
direkonstruksi.Pada konsentrasi substrat yang rendah, rata-rata reaksi awal
meningkat seiring dengan meningkatnya konsentrasi substrat sebagaimana
yang diharapkan. Pada konsentrasi substrat yang lebih lebih tinggi, peningkatan
rerata reaksi menjadi lebih sedikit sampai suatu titik yang dapat dicapai dimana
rerata reaksi menjadi konstan tak peduli berapa banyak substrat yang ada.
Kurvanya adalah berupa hiperbolik.Kita menggambarkan rerata yang konstan
sebagai kecepatan yang maksimum atau Vrnax.Perilaku katalitis ini seperti
diamati untuk kebanyakan enzim, dapat dideskripsikan oleh suatu efek substrat
jenuh.
Gambar 4.4
Dengan Menggunakan kurva Michaelis-Menten untuk
menaksir Vmax Dan KM. V max diperkirakan dari grafik di titik-titik
dimana kecepatan reaksi tidak lagi meningkatkan konsentrasi
substrat. KM diukur pada sumbu [S].KM menggambarkan
konsentrasi substrat yang menghasilkan suatu kecepatan 1/2 V
max.
[image:45.595.203.399.416.570.2]34
suatu persamaan matematik untuk menyatakan bentuk kurva hiperbolik dan
untuk mengkalkulasi rerata konstan.Michaelis dan Menten mengusulkan
molekul enzim sebagai E dan molekul substrat adalah S dan mampu
berkombinasi membentuk suatu kompleks yang reversibel.
k1 k3
E + S
═ ES
═ E + P
k2 k4
Istilah k1, k2, k3, dan k4 menggambarkan konstanta untuk tahap-tahap
individual. Ada dua kemungkinan kompleks ES: (1) dapat berbalik menjadi
enzim dan substrate yang bebas, atau ( 2) dapat diikuti dengan suatu reaksi
dapat dibalik untuk membentuk produk (P) dan enzim yang bebas. Reaksi di
atas adalah urutan reaksi minimal yang diperlukan untuk menjelaskan kerja
enzim. Suatu versi reaksi yang rumit menunjukkan suatu kompleks
enzyme-product telah diusulkan tetapi memerlukan analisis matematik dan tidak secara
signifikan pemahaman kita terhadap fungsi enzim:
E+S = ES = EP = E + P
Persamaan dasar yang diturunkan oleh Michaelis dan Menten untuk
menjelaskan reaksi katalis enzim adalah sbb:
vo =
[
]
]
max[
S
KM
S
V
ket:
vo adalah kecepatan awal yang disebabkan oleh konsentrasi subsrat
Vmax adalah kecepatan maximum
KM adalah konstanta Michaelis.
(Sebagian dari Anda mungkin telah mengenali persamaan ini sebagai
persamaan matematik untuk suatu hiperbola) Michaelis dan Menten membuat
beberapa asumsi untuk mempermudah menurunkan rumus ini.Mereka memilih
untuk mengabaikan reaksi yang berbalik produk P dan enzim bebas terhadap
kompleks ES (yang digambarkan oleh
k4di dalam urutan reaksi.Reaksi ini
35
hanya mengukur hanya initial rate (rerata/konstanta awal ketika reaksi yang
diwakili oleh k4 adalah sangat-sangat lambat ( pada umumnya selama
beberapa menit yang awal). Asumsi lain yang perlu diusulkan di dalam
penurunan persamaan adalah bahwa kompleks ES adalah suatu steady-state
intermediate. Yaitu setelah pencampuran E dan S, suatu ES tertentu telah
terbentuk dengan cepat dan konsentrasinya relatif konstan sebab produk
dihasilkan dengan kecepatan yang sama dengan pemecahannya.
Dua konstanta penting dalam persamaan Michaelis-Menten, KM dan
Vrnax, memerlukan uraian lebih lanjut. Konstanta Michaelis, KM, dinyatakan
secara matematik sebagai berikut.
KM =
1 3 2 k k k Karena sukar untuk memperoleh suatu pemahaman KM yang riil dalam
hal ini, kita akan menggambarkan tetapan Michaelis dalam terminologi yang
berbeda. Jika suatu analisis diselesaikan untuk KM dalam konstanta
Michaelis-Menten, ditemukan untuk mendapatkan unit yang sama sebagai konsentrasi
substrat ( S]. Ini berimplikasi terhadap hubungan antara KM dan [S].Apa yang
terjadi terhadap persamaan Michaelis-Menten jika nilai untuk KM sama dengan
nilai [S]?
Vo
=
] [ ] [ ] max[ S S S V
=
] [ 2 ] max[ S S V=
2 max Vdimana KM = [S]
36
Informasi lanjut tentang KM dapat dijelaskan dari kasus khusus ketika k2
sangat banyak dibandingkan dengan k3. Di bawah kondisi ini k3 adalah tidak
penting dan KM digambarkan sebagai:
KM =
3 2
k k
Dimana k2>>> k3.
Dalam format ini, Anda akan mengenali KM sebagai konstanta disosiasi untuk
yang kompleks:
k1
ES
=
E + S
k2
b. Persamaan Lineweaver
–
Burk
Pada tahun 1934, Hans Lineweaver dan Dean Burk melaporkan suatu
metoda untuk mengubah persamaan Michaelis-Menten ke dalam suatu format
yang memungkinkan untuk analisa grafis.
Persamaan Lineweaver
–
Burk memungkinkan satu data percobaan di
dalam suatu format garis lurus.
vo
1
=
max
V KM
.
] [
1
S
+
max1
37
Gambar 4.5
Dengan menggunakan persamaan dan kurva
[image:49.595.234.386.87.235.2]38
BABV
STRUKTUR DAN FUNGSI KARBOHIDRAT
A.KOMPETENSI:
1. menjelaskan fungsi karbohidrat 2. menjelaskan struktur karbohidrat
3. menjelaskan reaksi-reaksi pada karbohidrat
B. ELEMEN KOMPETENSI
1. mendeskripsikan fungsi karbohidrat 2. mendeskripsikan struktur karbohidrat
3. mendeskripsikan reaksi-reaksi pada karbohidrat
C. MATERI
Karbohidrat merupakan bagian yang sangat berlimpah dari tumbuhan dan hewan,. senyawa ini mempunyai gugus fungsi aldehid atau keton serta kelompok hidroksil yang banyak. Zat ini berfungsi sebagai sumber energi, selain itu ada beberapa fungsi lain dari karbohidrat adalah:
1. Sebagai bahan utama penyusun dinding sel pada tumbuhan dan hewan,serta eksoskeleton pada artropoda.
2. Monosakarida ribose dan dioksiribosa sebagai komponen asam nukleat.
3. Karbohidrat yang berikatan kovalen dengan protein lipid permukaan sel berfungsi sebagai marker untuk pengenalan secara molekuler dengan biomolekul yang lain.
Karbohidrat dikelompokkan menjadi. Monosakarida, disakarida, trisakarida, tetrasakarida, dan polisakarida.
a). Monosakarida
Karbohidrat sederhana adalah monosakarida, senyawa dengan satu unit aldehid atau keton tunggal dan banyak hidroksil yang memiliki formula empirik (CH2O)n.
39
Monosakarida dengan n=5, 6, dan 7 disebut pentosa, heksosa, dan hepsosa, sementara jumlahatom karbon bertambah maka jumlah kemungkinan isomer juga bertambah.Gambar 4.2 menggambarkan hubungan stereokimia antara D-aldosa dari n=3 sampai n=6. D-ribosa dengan 5 atom karbon adalah satu aldopentosa, merupakan komponen dari RNA. Beberapa monosakarida penting dalam kelompok D-aldosa adalah: D-glukosa, D-manosa, dan D-galaktosa merupakan D-aldosa yang melimpah. D-Manosa dan D-galaktosa berbeda secara stereokimia dengan D-glukosa hanya pada tempat gugus fungsionalnya (manosa pad C2, galaktosa pada C4). Manosa dan galaktosa sebagai epimer dari glukosa.
[image:51.595.121.357.425.656.2]Hubungan stereokimia antara D-seri untuk ketosa disajikan dalam gambar 4.3. Semua senyawa mempunyai satu oksigen karbonil pada atom C2, dan memiliki satu gugus hidroksil pada setiap atom karbon yang lain.Ketosa yang umum adalah D-fruktosa, ketosa dengan jumlah n=7 adalah D-sedoheptulosa. Jika 5-karbon aldosa adalah D-ribosa, maka 5-karbon ketosa adalah D-ribulosa. Keduanya mempunyai gugus fungsional yang berbeda maka ribulosa merupakan isomer dari ribose. Nama dan klasifikasi untuk monosakarida yang umum dialam adalah sebagai berikut:
Tabel 5.1: Klasifikasi dan nama beberapa monosakarida:
40
Gambar 5.1: Monosakarida yang mengandung 4 karbon. Orientasi stereokimia (D atau L) ditentukan pada atom karbon nomor 3. Ke tiga D-tetrosa mempunyai konfigurasi absolut pada C3
[image:52.595.104.500.306.626.2]41
[image:53.595.133.437.84.654.2]42
b). Stuktur siklik karbohidrat
Gambar 5.1, 5.2 dan 5.3 menunjukkan struktur rantai lurus dari karbohidrat atau yang dikenal dengan proyeksi Fischer. Struktur cincin dibentuk melalui reaksi antara aldehid atau keton pada satu ujung dari molekul dengan gugus hidroksil pada ujung yang lain.
[image:54.595.90.447.218.556.2]Reaksi antara satu aldehid dengan satu hidroksil (alcohol) untuk membentuk satu hemiasetal adalah sebagai berikut:
Gambar 5.4. Pembentukan rantai siklik dari bentuk rantai terbuka D-ribosa.
43
Gambar 5.5: Struktur siklik dari D-glukosa membentuk 2 siklik hemiasetal
44
Gambar 5.6. Struktur siklik dari D-fruktosa membentuk 2 siklik hemiketal
β-D- fruktofuranosa dan α-D-fruktofuranosa
c). Reaksi dari glukosa dengan monosakarida yang lain 1.Reaksi oksidasi reduksi