KARAKTERISASI DAN KONSTRUKSI VEKTOR EKSPRESI
GEN SMALL HEAT SHOCK PROTEIN (sHSP) Lactobacillus
plantarum SEBAGAI ALTERNATIF PENANDA SELEKSI
FOOD GRADE DI Lactococcus lactis
HASLIA MARGARETA
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA*
Dengan ini saya menyatakan bahwa tesis berjudul Karakterisasi dan Konstruksi Vektor Ekspresi Gen small Heat Shock Protein (sHSP) Lactobacillus plantarum sebagai Alternatif Penanda Seleksi Food Grade di Lactococcus lactis adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor dan LIPI.
RINGKASAN
HASLIA MARGARETA. Karakterisasi dan Konstruksi Vektor Ekspresi Gen small Heat Shock Protein (sHSP) Lactobacillus plantarum sebagai Alternatif Penanda Seleksi Food Grade di Lactococcus lactis. Dibimbing oleh UTUT WIDYASTUTI dan APON ZAENAL MUSTOPA.
Bakteri asam laktat dikenal dengan bakteri yang memiliki status Generallly Recogninize As Safe (GRAS) yaitu mikroba yang aman dan tidak beresiko terhadap kesehatan. BAL umumnya digunakan pada industri fermentasi makanan. Lactobacillus plantarum U10 adalah salah satu BAL yang telah diisolasi dari makanan fermentasi tradisional Indonesia, yaitu Tempoyak. BAL mempunyai sifat yang sensitif terhadap kondisi lingkungan, ketika digunakan sebagai starter komersial atau ketika produk fermentasi sedang diproses, diangkut dan disimpan. Strategi yang digunakan untuk meningkatkan ketahanan BAL terhadap tekanan lingkungan adalah dengan mengembangkan food grade vector dengan marka seleksi gen small Heat Shock Protein (sHSP). Penelitian ini bertujuan untuk mengkarakterisasi gen small Heat Shock Protein (sHSP) asal L. plantarum U10 yang diisolasi dari makanan tradisional Indonesia
“Tempoyak”. Selain itu penelitian ini juga bertujuan untuk mengklon promotor slpA dan gen sHSP ke dalam vektor kloning pGEM-T dan konstruksi gen fusi antara promotor slpA dan gen sHSP ke dalam vektor ekspresi pNZ8148, serta mengintroduksikannya ke dalam strain BAL jenis L. lactis NZ3900.
Penelitian ini terdiri dari tiga tahapan, yaitu karakterisasi, kloning dan konstruksi. karakterisasi dimulai dengan perlakuan kejut panas pada L. plantarum U10 dan SDS-PAGE. Selanjutnya dilakukan uji aktivitas Chaperon, isolasi total RNA dan analisis Reverse Transcriptase PCR. Tahapan kloning dimulai dari amplifikasi gen sHSP dari DNA kromosom L. plantarum U10 dan promotor slpA dari isolat L. acidophilus C9-9, kemudian mengkloningnya ke dalam vektor kloning pGEM-Teasy. Selanjutnya vektor kloning pGEM-T slpA digunakan sebagai sumber untuk memperoleh promotor slpA dan pGEM-T sHSP sebagai sumber untuk gen sHSP. Konstruksi vektor food grade dilakukan dengan memfusikan promotor dan gen terlebih dahulu. Fusi keduanya dilakukan dengan teknik PCR overlapping sehingga menghasilkan fusi antara promotor slpA dan gen sHSP. Fusi dipotong dengan menggunakan enzim restriksi SalI-BglII. Vektor ekspresi pNZ8148 juga dipotong dengan enzim yang sama pada daerah kloramfenikol, dan kemudian diligasikan dengan fusi slpA-sHSP. Tahapan terakhir adalah mengintroduksikan hasil ligasi ke inang L. lactis NZ3900 menggunakan teknik elektroporasi, dan dilakukan konfirmasi dengan PCR Koloni.
Fragmen promotor slpA berukuran 192 pb berhasil teramplifikasi dari L. acidhopilus C9-9. Fragmen gen sHSP berukuran 423 pb berhasil teramplifikasi dari kromosom L. plantarum U10. Hasil analisis sekuen menunjukkan bahwa promotor slpA memiliki tingkat kemiripan mendekati 100% dengan L. acidophilus ATCC 4356 referensi. Analisis sekuen gen sHSP juga menunjukkan tingkat kemiripan tinggi 100% dengan L. plantarum WCFS1 referensi. Fusi antara promotor slpA dan gen sHSP berhasil termplifikasi dengan ukuran 615 pb dengan teknik PCR overlapping. Fusi gen tersebut berhasil dimasukkan ke dalam vektor ekspresi dengan mengganti bagian kloramfenikol yang telah dipotong terlebih dahulu dengan enzim restriksi yang sama yaitu SalI dan BglII sehingga terbentuk pNZ8148-slpA-sHSP dan diintroduksikan ke inang L. lactis NZ3900. Konfirmasi vektor rekombinan dilakukan dengan PCR koloni dengan penggunakan primer dari gen fusi slpA dan sHSP dan PCR koloni dengan menggunakan primer promotor dan terminator pNZ8148.
SUMMARY
HASLIA MARGARETA. Characterization and Construction Of Lactobacillus plantarum small Heat Shock Protein (sHSP) Gene Expression Vector As Alternative Food Grade Selection Marker in Lactococcus lactis. Supervised by UTUT WIDYASTUTI and APON ZAENAL MUSTOPA.
Lactic acid bacteria are known generally recognize as safe (GRAS) that microbes are safe, without risk on health. LAB is commonly used in food fermentation industry. Lactobacillus plantarum U10 is one of LAB has been isolated from Indonesian traditional fermented food, namely Tempoyak. LAB are sensitive to environmental conditions when commercial starters or fermentation products are being processed, transported and strored. The startegy used to increase resistance of LAB to environmental stress is to develop food grade vektor with selection marker small heat shock protein (sHSP) gene. This study aimed to characterize the genes small Heat Shock Protein (sHSP) of L. plantarum U10 isolated from Indonesian traditional food "Tempoyak". In addition , this study also aims to clone a promoter slpA and sHSP gene into the cloning vector pGEM-T and the construction of a fusion gene between the promoter slpA and sHSP gene into the expression vector pNZ8148, and introduction into strain L. lactis NZ3900.
This study consisted of three phases, namely the characterization, cloning and construction. Characterization starting with heat shock treatment on L. plantarum U10 and SDS-PAGE. The next step is Chaperon activity assay, total RNA isolation and Reverse-Transcriptase PCR analysis. Cloning starts with amplification of sHSP gene from chromosomal DNA of L. plantarum U10 and Promoter slpA from isolate L. acidhophilus C9-9, then cloned into cloning vector pGEM-Teasy. Furthermore the cloning vector pGEM-T slpA used as a source for obtain the slpA promoter and pGEM-T sHSP as a source for sHSP genes. The construction of food grade vector is done with promoter and genes merge first. The fusion between slpA promoter and sHSP genes by overlapping PCR technique. The result of fusion then digested with restriction enzymes SalI-BglII. Expression vector pNZ8148 also digested with same enzyme in chlorampenicol area, and then ligated with fusion slpA-sHSP. The last step is introduce the result of ligation to host L. lactis NZ3900 using electroporation technique, and confirmed by PCR colonies.
The characterization of small heat shock protein (sHSP) from tempoyak-originated Lactobacillus plantarum was investigated. The heat adaptive response proteins were ranging from 18 kDa to 51 kDa. The Intercellular Protein (IP) fraction of heat shocked-L.plantarum U10 exhibited chaperone like activity by the ability to prevent loss of proteinase K activity from denaturation. Furthermore, The sHSP gene that related to the predicted sHSP ±18 kDa protein were successfully identified by PCR method and this gene has 423 pb size. Moreover, the sHSP ±18 kDa was indeed up-regulated after L. plantarum U10 treated by heat shocking as proven by Reverse Transcriptase-PCR. This result suggested that sHSP protein ±18 kDa in our study may confers a survival advantage on Lactobacillus plantarum and capable of protecting the cell against under temperature stress.
the chromosome of L. plantarum U10. Results of sequence analysis showed that the promoter slpA have a similarity rate approaching 100 % with L. acidophilus ATCC 4356 reference. Analysis of gene sequence similarity sHSP also show high rate 100% with L. plantarum WCFS1 reference. Fusion between the slpA promoter and sHSP gene successfully termplifikasi with the size 615 pb PCR overlapping. The fusion gene was successfully inserted into the expression vector by replacing the chlorampenicol that has been cut in advance with the same restriction enzyme that SalI and BglII to form pNZ8148-slpA-sHSP and introduced into the host L. lactis NZ3900. Confirmation recombinant vector is done by colony PCR with the use of gene fusion primer slpA and SHSP and colony PCR using a primer promoter and terminator pNZ8148.
© Hak Cipta Milik IPB dan LIPI, Tahun 2015
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB dan LIPI.
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains
pada
Program Studi Bioteknologi
KARAKTERISASI DAN KONSTRUKSI VEKTOR EKSPRESI
GEN SMALL HEAT SHOCK PROTEIN (sHSP) Lactobacillus
plantarum SEBAGAI ALTERNATIF PENANDA SELEKSI
FOOD GRADE DI Lactococcus lactis
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
BOGOR 2015
PRAKATA
Puji dan syukur penulis panjatkan kepada Allah subhanahu wa ta’ala atas segala karunia-Nya sehingga karya ilmiah ini berhasil diselesaikan. Karya ilmiah ini berjudul Karakterisasi dan Konstruksi Vektor Ekspresi Gen small Heat Shock Protein (sHSP) Lactobacillus plantarum sebagai Alternatif Penanda Seleksi Food Grade di Lactococcus lactis yang telah dilaksanakan sejak bulan November 2014 sampai Juni 2015.
Terima kasih penulis ucapkan kepada Ibu Dr Ir Utut Widyastuti, MSi selaku pembimbing utama dan Bapak Dr Apon Zaenal Mustopa, MSi selaku anggota pembimbing atas bimbingan dalam penyusunan karya ilmiah ini. Ucapan terima kasih juga disampaikan kepada Dr Ir Harsi Dewantari Kusumaningrum yang telah memberikan masukan dalam memperkaya tulisan ini. Penulis juga mengucapkan terima kasih kepada Dr Ir Bambang Sunarko sebagai Kepala Pusat Penelitian Bioteknologi LIPI yang telah memberikan izin kepada penulis untuk melakukan penelitian di Pusat Penelitian Bioteknologi LIPI. Penulis juga mengucapkan terima kasih kepada Ayahanda Sarman, AmdPd dan Ibunda Miharsyawati, AmdPd atas doa dan kasih sayangnya, serta keempat saudara saya Yenni Afrida, SPt, Dwi Wahyu Gandadi Putra, Amd, Seppi Triani, SH dan Lisa Puspita Sari, SKM yang selalu memberikan semangat dan kasih sayang kepada saya. Tidak lupa saya ucapkan terima kasih kepada teman-teman dan rekan-rekan staf di Laboratorium Bioteknologi-LIPI, dan teman-teman Bioteknologi 2013-2014 yang membantu penulis. Selain itu, ucapan terima kasih juga saya haturkan kepada Badan Litbang Pertanian, Kementerian Pertanian yang telah membiayai penelitian ini melalui skema Kerjasama Kemitraan Penelitian dan Pengembangan Pertanian Nasional (KKP3N) 2014-2015.
Penyusunan karya ilmiah ini tentunya tidak terlepas dari kekurangan. Oleh karena itu, penulis mengharapkan adanya saran dan kritik yang membangun untuk menyempurnakan penulisan karya ilmiah ini. Semoga karya ilmiah ini bermanfaat.
DAFTAR ISI
DAFTAR TABEL xii
DAFTAR GAMBAR xii
DAFTAR LAMPIRAN xii
1 PENDAHULUAN
Latar Belakang 1
Tujuan Penelitian 2
2 TINJAUAN PUSTAKA 3
Lactobacillus plantarum 3
Small Heat Shock Protein 4
Penanda Seleksi 5
Lactococcus lactis 6
3 METODE 7
Tempat dan Waktu Penelitian 7
Bahan 7
Prosedur Kerja 9
4 HASIL DAN PEMBAHASAN 13
Pengaruh Perlakuan Kejut Panas pada Profil Ekspresi Protein
L.plantarum U10 13
Ekspresi Gen dari Kejut Panas L. plantarum U10 dan Uji Aktivitas
Chaperon 14
Konstruksi Vektor Kloning pGEM-T slpA dan pGEM-T 16 Konstruksi Vektor Ekspresi dengan Fusi Promotor slpA
dan Gen sHSP 20
5 SIMPULAN DAN SARAN 23
Simpulan 23
Saran 23
DAFTAR PUSTAKA 24
LAMPIRAN 28
DAFTAR TABEL
1 Primer yang digunakan untuk amplifikasi gen 7
2 Plasmid dan strain bakteri 9
3 Kondisi reaksi PCR 11
4 Profile ekspresi protein intraseluler L. plantarum U10 setelah perlakuan
kejut panas. 14
DAFTAR GAMBAR
1 Scaning Electron Micrograph dari L. plantarum WCFS1 3 2 Mekanisme Pengaturan Heat Shock Response dan HSF-1 5 3 Vektor kloning pGEM-Teasy dan vektor ekspresi pNZ8148 7
4 Diagram Alir Penelitian 8
5 Profile ekspresi L. plantarum protein intraseluler setelah perlakuan kejut panas. 9 Hasil transformasi E. coli TOP 10 dengan pGEM-slpA. 17 10 Hasil PCR Koloni dan PCR Plasmid E. coli TOP 10 dengan
pGEM-slpA.
17 11 Hasil amplifikasi PCR promotor sHSP dan Peta vektor kloning
rekombinan pGEM-sHSP
18 12 Hasil transformasi E. coli TOP 10 dengan pGEM-sHSP. 18 13 Hasil PCR Koloni dan PCR Plasmid E. coli TOP 10 dengan
pGEM-sHSP
19 14 Hasil analisis sekuen promotor surface layer protein A 19 15 Hasil analisis sekuensing gen penyandi small Heat Shock Protein 20 16 Skema prosedur kontsruksi pNZ8148 slpA_sHSP 21 17 Hasil amplifikasi fusi slpA dan sHSP dan Transformasi
1 Komposisi media pertumbuhan de Man, Rogosa, Sharpe (MRS) 30
2 Komposisi larutan elektroforesis SDS-PAGE 31
3 Metode Isolasi Total RNA 32
4 Komposisi media pertumbuhan bakteri Luria-Bertani (LB) 33
5 Komposisi media pertumbuhan M17 34
6 Zona Bening Uji Aktivitas Chaperon 35
7 Hasil pengurutan nukleotida promotor pGEM-T slpA 36
1
PENDAHULUAN
Latar Belakang
Bakteri asam laktat dikenal dengan bakteri yang memiliki status Generallly Recogninize As Safe (GRAS) yaitu mikroba yang aman dan tidak beresiko terhadap kesehatan. BAL umumnya digunakan pada industri pangan dan farmasi. Dalam industri pangan bakteri asam laktat telah digunakan secara luas sebagai kultur starter untuk beragam fermentasi daging, susu, sayuran, buah, dan roti. Selain itu BAL juga digunakan sebagai pengawet produk pangan, karena BAL mampu menghasilkan senyawa antimikroba yang disebut bakteriosin. Pada industri farmasi, bakteri asam laktat digunakan sebagai probiotik dan untuk produksi vaksin. BAL merupakan sel inang yang aman terhadap pangan. Bakteri asam laktat digunakan dalam berbagai macam industri fermentasi makanan dan telah diketahui memberikan efek yang menguntungkan bagi kesehatan manusia. Metabolit yang dihasilkan oleh BAL dapat secara efektif mengontrol pertumbuhan bakteri patogen dan pembusuk. BAL sebagai food grade microorganism dikatakan aman terutama dalam pangan karena sifatnya tidak menghasilkan racun bahkan beberapa jenis diantaranya berguna bagi kesehatan.
Sel inang yang aman dan telah memiliki status GRAS yang sering digunakan dalam mengekspresikan protein rekombinan adalah Lactococcus lactis (Mierau et al. 2005). Selain aman, penggunaan L. lactis juga dikarenakan inang tersebut mudah ditangani, telah dikarakterisasi sebagai mikroorganisme yang baik untuk industri dan mampu mensekresikan protein rekombinan pada media pertumbuhannya sehingga produk yang dihasilkan bebas dari endotoksin. Terdapat banyak penelitian yang telah dikembangkan dengan menggunakan inang L. lactis misalnya produksi protease, amilase dan bakteriosin sendiri (Martin et al. 2007; Liang et al. 2010; Jorgensen et al. 2013; Wu et al. 2013 dan Lages et al. 2015).
BAL mempunyai sifat yang sensitif terhadap kondisi lingkungan, ketika digunakan sebagai starter komersial atau ketika produk fermentasi sedang diproses, diangkut dan disimpan (Tian et al. 2012). Strategi yang digunakan untuk meningkatkan ketahanan BAL terhadap tekanan lingkungan adalah dengan mengembangkan food grade vector dengan marka seleksi gen small Heat Shock Protein (sHSP).
2
ini tidak diinginkan dalam pembuatan produk makanan dan pakan (Diekely et al. 1995). Saat ini, telah banyak digunakan penanda seleksi untuk membangun food grade vector seperti β-galaktosidase dari S. thermophilus (Herman et al. 1985), gen sHSP (El Demardash et al. 2003; Spano et al. 2005), gen yang terkait auxotrophy (Bron et al. 2002), bakteriosin (Mills et al. 2002; Takala et al. 2002), dan jalur metabolik baru (Boucher et al. 2002) telah digunakan dalam konstruksi food grade vector sebagai marka seleksi.
Lactobacillus plantarum dengan kode isolat U10 pada penelitian ini telah diisiolasi dan dikarakterisasi dari makanan tradisioanal Indonesia yaitu tempoyak yang merupakan makanan fermentasi dari buah durian. L. plantarum ini menunjukkan aktivitas antibakteri yang sangat baik terhadap bakteri patogen dan galur ini menunjukkan keuntungan yang akan diterapkan dalam bidang medis sebagai antibakteri alami (Urnemi et al. 2010). Studi lain tentang L. plantarum sudah dilakukan oleh Spano (2005) yaitu dengan mengklon dan mengkarakterisasi gen small heat shock protein (sHSP) dari L. plantarum yang diisolasi dari Wine. Gen sHSP memiliki peranan terhadap stres lingkungan pada L. plantarum dan menurut El Demardash et al. 2003 gen sHSP dari plasmid S. thermophilus merupakan marka seleksi yang ideal digunakan karena gen sHSP dapat meningkatkan ketahanan terhadap tekanan lingkungan.
Heat shock protein merupakan suatu protein yang dihasilkan karena adanya stres lingkungan. Gen sHSP merupakan gen yang berperan dalam proteksi terhadap beberapa jenis stres dengan berat molekul protein berkisar 11-42 kDa dan banyak terdapat pada prokariot dan eukariot (Tian et al. 2012). Heat shock protein terekspresi karena adanya stres atau disebut juga gen yang sifatnya inducible. Studi promotor yang akan digunakan yaitu promotor surface layer protein A (slpA) telah dilakukan oleh McCracken (1999) dari tipe liar Lactobacillus acidophilus ATCC 4356 ke dalam plasmid pNZ272 yang merupakan promotor kuat.
Dari penjelasan di atas untuk meningkatkan ketahanan dari BAL yang bersifat sensitif terhadap stress lingkungan adalah dengan konstruksi vektor ekspresi yang membawa gen small heat shock protein atau disebut juga sHSP yang dapat meningkatkan stres resinten dari sel inang.
Tujuan Penelitian
Penelitian ini bertujuan untuk mengkarakterisasi gen small Heat Shock Protein (sHSP) asal L. plantarum U10 yang diisolasi dari makanan tradisional
Indonesia “Tempoyak”. Selain itu penelitian ini juga bertujuan untuk mengklon promotor slpA dan gen sHSP ke dalam vektor kloning pGEM-T dan konstruksi gen fusi antara promotor slpA dan gen sHSP ke dalam vektor ekspresi pNZ8148, serta mengintroduksikannya ke dalam strain BAL jenis L. lactis NZ3900.
3
2
TINJAUAN PUSTAKA
Lactobacillus plantarum
Bakteri asam laktat (BAL) adalah kelompok bakteri Gram positif, tidak membentuk spora, katalase negatif, tanpa sitokrom, bersifat anaerob, dan oksidase positif, toleran asam, asam laktat sebagai produk utama fermentasi karbohidrat. Umumnya bakteri asam laktat tumbuh pada habitat yang kaya nutrisi, seperti produk makanan, susu, daging, dan sayuran. Bakteri asam laktat merupakan bakteri yang banyak terlibat dalam hasil fermentasi pangan terutama yang melibatkan proses fermentasi spontan seperti bekasam (fermentasi daging sapi), dadih (fermentasi susu kerbau), tape ketan (fermentasi beras ketan), dan tempoyak (fermentasi buah durian).
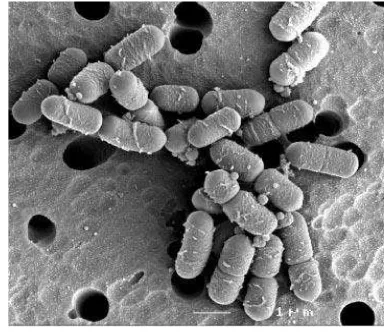
Lactobacillus plantarum merupakan salah satu jenis dari BAL yang juga termasuk dalam bakteri Gram positif yang berbentuk kokus atau batang, tidak menghasikan spora dan katalase, resisten terhadap kondisi lingkungan yang asam, tumbuh optimum secara anaerobik aerotoleran pada suhu 30 °C (mesofilik) atau 42 °C (termofilik) dengan kisaran pH optimum 4.0-4.5(Gambar 1). Lactobacillus plantarum banyak diisolasi dari makanan fermentasi. L. plantarum bersifat heterofermentatif fakultatif, dimana proses fermentasi karbohidrat pada umumnya melalui jalur sintesa fosfoketolase. Fermentasi pentosa (xylosa dan ribosa) akan menghasilkan piruvat dan asetil fosfat yang nantinya akan dikonversi menjadi laktat dan asetat. Selain itu, heksosa (glukosa, fruktosa dan mannosa) juga akan dikonversi menjadi laktat, CO2 dan etanol (Mayo et al. 2010; Todorov dan Franco 2010; Hammes dan Vogel 1995). Selain itu pula, L. plantarum juga telah dikembangkan sebagai produk probiotik, salah satu strain yang telah dikomersialkan adalah L. plantarum 299v (Siezen dan van Hylckama Vlieg 2011).
Gambar 1 Scaning Electron Micrograph dari L. plantarum WCFS1 (Bron et al. 2004) Penerbit American society for Microbiology.
4
BAL karena menyediakan kompleks nutrisi yang mendukung pertumbuhan BAL. Jenis mikroorganisme BAL pada pangan tempoyak diantaranya adalah L. plantarum, L. sakei dan L. corynebacterium (Yuliana dan Garcia 2009). Penelitian ini menggunakan Lactobacillus plantarum U10 yang berhasil diisolasi dari
“Tempoyak” makanan fermentasi dari buah durian (Urnemi et al. 2010).
small Heat Shock Protein
Protein merupakan kompenen utama dalam metabolisme suatu sel. Pada suatu organisme terdapat perbedaan ekspresi protein dari jaringan satu dengan jaringan lain tergantung kondisi lingkungan. Heat Shock Protein (HSP) merupakan protein yang dihasilkan karena adanya tekanan lingkungan. HSP juga merupakan suatu molekular chaperon yang berfungsi untuk melindungi protein lain dari agregasi, melonggarkan protein yang beragregasi, membantu pelipatan protein baru atau pelipatan kembali protein yang rusak, mendegradasi protein yang rusak cukup parah dan dalam kasus kerusakan yang sangat berat, mengasingkan protein yang rusak menjadi agregat yang lebih besar (Felix et al. 2009).
HSP dihasilkan karena adanya Heat shock response (HSR). HSR adalah suatu respon untuk menginduksi gen-gen yang mengkode molekular chaperon, protease dan protein-protein lain yang penting dalam mekanisme pertahanan dan pemulihan terhadap seluler yang berhubungan dengan terjadinya misfolded protein. HSR merupakan suatu tanggapan sel terhadap berbagai macam gangguan, baik yang bersifat fisiologik maupun yang berasal dari lingkungan (Wiesterheide et al. 2005).
Klasifikasi kelas-kelas HSP dilakukan berdasarkan ukuran molekul dan fungsinya. Ada subkelas HSP100, HSP90, HSP70, HSP60, HSP40 dan small heat shock protein (sHSP). Angka yang mengikuti kata HSP menunjukkan berat molekulnya, contoh: angka 100 menunjukkan berat molekul dari HSP, yakni 100 kDa. sHSP adalah sub-kelas dari HSP yang mempunyai karakter berat molekular yang rendah (9-40 kDa) (Lelj et al. 2006).
HSR (Heat Shock Response) adalah reaksi sel dari organisme terhadap kenaikan suhu (heat shock atau heat stress). Heat stress yang berat dapat mengakibatkan kerusakan dan kematian sel, sedangkan dosis subletal dari heat stress akan memicu reaksi seluler yang disebut heat shock response (Schoffl dan Reindl 1998). HSR diatur pada tingkat transkripsi oleh suatu mekanisme yang melibatkan heat shock transciption factor (HSF).
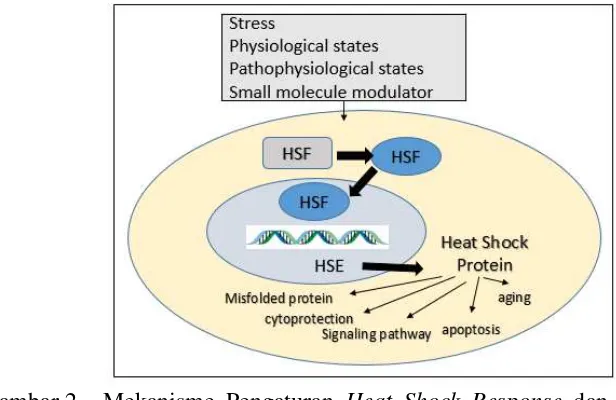
5 Raf/ERK, jalur protein kinase yang diaktivasi mitogen yang responsif pada faktor pertumbuhan, dan berakibat pada inhibisi aktivitas HSF1 (Felix et al. 2009 dan Schoffl dan Reindl 1998).
Gambar 2 Mekanisme Pengaturan Heat Shock Response dan HSF-1 (Felix et al. 2009).
Ciri khususnya small heat shock protein (sHSP) adalah memiliki urutan asam amino 80-100 residu yang berperan pada proses stabilisasi. Kelompok HSP ini memiliki ukuran yang relatif rendah berkisar 11-42 kDa. sHSP merupakan gen yang berperan dalam proteksi terhadap beberapa jenis stres dan banyak terdapat pada prokariot dan eukariot (Tian et al. 2012). sHSP diinduksi oleh berbagai tekanan, seperti tekanan suhu dan pH yang rendah. Protein ini dapat meningkatkan
ketahanan BAL terhadap tekanan suhu dan pH (O’Sullivan et al. 1999 dan Solow et al. 2000). sHSP terdiri dari kelompok yang diinduksi oleh tekanan dari molekul chaperon yang dapat mengikat protein terdenaturasi. sHSP berfungsi mencegah agregasi dan mempertahankan protein dari kesalahan pelipatan di bawah kondisi tekanan (Helsbeck et al. 2002 dan Narberhaus et al. 2002). Gen sHSP dari plasmid S. thermophilus merupakan marka seleksi yang ideal digunakan karena gen sHSP dapat meningkatkan ketahanan terhadap tekanan lingkungan (El Demardhash et al. 2003)
Penanda Seleksi
6
pengembangan gen seleksi baru untuk mengatasi permasalahan tersebut telah dilakukan yaitu dengan mengembangkan food grade vector.
Food grade vector merupakan suatu vektor yang tidak berbahaya dan dapat digunakan dalam industri makanan (Song et al. 2012). Vektor ini berasal dari mikroorganisme yang tidak membahayakan manusia.Transformasi pada bakteri menggunakan plasmid sebagai pembawa gen atau vektor yang akan dimasukkan ke sel inang. Plasmid adalah DNA ekstrakromosomal yang umumnya berbentuk sirkular dan secara alami dapat dijumpai pada bakteri dan beberapa jenis yeast uniselular seperti Saccharomyces cereviseae. Plasmid berukuram mulai dari 1000 base pair (bp) sampai 1000 kilo base pair (Kbp), dengan jumlah per sel bervariasi dari satu sampai ribuan salinan (copy) molekul. Plasmid umumnya merupakan elemen genetik yang dapat dipindahkan dari satu individu sel ke individu lainnya melalui konjugasi.
Plasmid memiliki peran yang sentral dalam rekayasa genetika sebagai vektor untuk pengklonan dan ekspresi gen. Sebagai vektor yang ideal, plasmid memiliki ori (origin of replication) sebagai titik awal replikasi untuk perbanyakannya di dalam sel inang. Plasmid juga memiliki daerah multiple cloning sites (MCS) sebagai tempat penyisipan segmen DNA atau gen yang akan diklon atau diekspresikan. Plasmid memiliki gen penanda seleksi, misalnya gen resistensi antibiotik yang berguna untuk seleksi klon. Food grade vector sudah banyak dikembangkan, pada penelitian ini penanda seleksi yang digunakan adalah gen sHSP untuk mengganti gen resisten antibiotik kloramfenikol pada plasmid pNZ8148.
Lactococcus lactis
Strain Lactococcus lactis telah banyak diisolasi dari berbagai sumber dan telah berhasil disekuensing. Salah satu karakteristik genetik dari L. lactis yaitu memiliki kandungan basa nitrogen guanin (G) dan sitosin (C) yang relatif rendah yaitu sekitar 35% (Cavanagh et al. 2015). Hal ini membuat L. lactis telah banyak dimanfaatkan dalam beberapa bidang bioteknologi seperti dalam ekspresi antigen bakteri dan virus sebagai pengembangan vaksin oral yang aman, produksi hormon sitokinin dan agen terapeutik lainnya, serta pilot production pengembangan produk farmasetika (Mierau 2005).
7
3
METODE
Tempat dan Waktu Penelitian
Penelitian ini dilaksanakan pada bulan November 2014 - Juni 2015 di Laboratorium Aplikasi Rekayasa Genetika dan Desain Protein, Pusat Penelitian Bioteknologi LIPI, Cibinong, Bogor.
Bahan
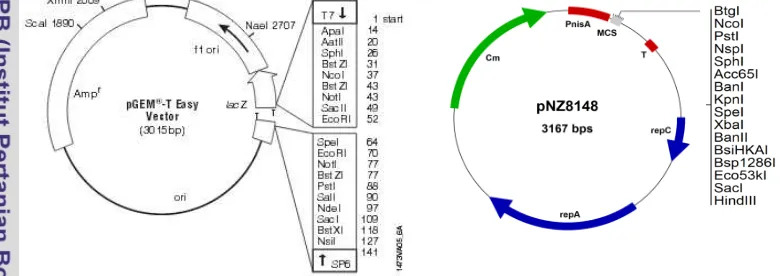
Bahan yang digunakan pada penelitian ini adalah media tumbuh bakteri Luria-Bertani (LB), M17B glukosa, MRS (de Man, Rogosa, Sharpe), enzim restriksi, enzim taq polimerase, enzim ligase, DNA ladder 1 kb (Thermo) dan 100 pb (Kappa), ampisilin, kloramfenikol. Plasmid yang digunakan adalah pGEMT (vektor kloning) dan pNZ8148 yang digunakan sebagai vektor ekspresi (Gambar 3). Beberapa pasang primer, serta beberapa vektor dan strain bakteri yang digunakan dalam penelitian ini dapat dilihat pada Tabel 1 dan Tabel 2
Gambar 3 Peta Plasmid pGEM-Teasy dan peta plasmid pNZ8148 Tabel 1. Primer yang digunakan untuk amplifikasi gen
Primer Enzim
8
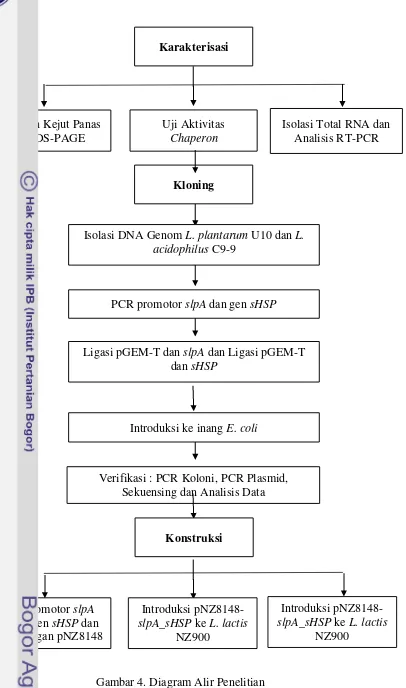
Gambar 4. Diagram Alir Penelitian Karakterisasi
Perlakuan Kejut Panas dan SDS-PAGE
Uji Aktivitas Chaperon
Isolasi Total RNA dan Analisis RT-PCR
Verifikasi : PCR Koloni, PCR Plasmid, Sekuensing dan Analisis Data
Kloning
Isolasi DNA Genom L. plantarum U10 dan L. acidophilus C9-9
PCR promotor slpA dan gen sHSP
Ligasi pGEM-T dan slpA dan Ligasi pGEM-T dan sHSP
Introduksi ke inang E. coli
Konstruksi
Fusi promotor slpA dengan gen sHSP dan ligasi dengan pNZ8148
Introduksi pNZ8148-slpA_sHSP ke L. lactis
NZ900
Introduksi pNZ8148-slpA_sHSP ke L. lactis
9
Tabel 2. Plasmid dan strain bakteri
Prosedur Kerja
Perlakuan Kejut Panas dan SDS-PAGE
Perlakuan kejut panas dilakukan dengan menumbuhkan L. plantarumU10 dalam 5 ml media MRS (de Man, Rogosa, Sharpe) dan diinkubasi pada 37 ºC semalaman. Selanjutnya, L. plantarum U10 ditumbuhan dalam media 100 ml MRS. Sel dipanen dengan sentrifugasi pada 10000 x g ketika OD 600 adalah ~0,6. Pelet yang didapatkan diresuspensi dengan media 20 ml MRS baru. Perlakuan kejutan panas dilakukan pada 42 ⁰C : (a) Kontrol (tanpa heat shock) , (b) heat shock 30 menit , (c) heat shock 45 menit. Setelah perlakuan, sampel dibiarkan pada suhu ruang selama 20 menit. Tingkat kelangsungan hidup sel dipantau dengan menghitung CFU pada agar setelah inkubasi pada 37 ºC semalaman. Perlakuan berdasarkan Delmas et al. 2001 dan Guzzo et al. 1997 dengan modifikasi. SDS-PAGE (Lampiran 2) dilakukan dengan meresuspen sampel yang telah diberi perlakuan dengan buffer sonikasi dan disonikasi selama 15 detik, kemudian diulang sebanyak 20 kali. Ekstrak sampel yang sudah disonikasi diisolasi dengan sentrifugasi pada 17000 x g, 4 ºC selama 20 menit. Supernatan dipindahkan ke tabung steril kemudian pelet dianalisis dengan SDS –PAGE.
Uji Aktivitas Chaperon
Uji aktivitas chaperon dilakukan mengikuti metode dari Collada et al. 1997, Kim et al. 1998 dengan kombinasi. Sebanyak 0,25 gram agarose dilarutkan dalam 50 mM Tris-HCl pH 7,4 dan kemudian dipanaskan. Larutan agarose kemudian ditambahkan 2,5 ml 2% gelatin, diaduk sampai tercampur rata. Sebanyak 25 ml larutan agar dituangkan ke cawan petri, dibiarkan mengeras dan bentuk sumuran pada agar. Perlakuan terdiri dari beberapa kombinasi untuk masing-masing sampel: sampel 1; proteinase K denaturasi selama 15 menit pada 100⁰C dicampur dengan IP (protein intraselular L. plantarum U10) (1:1, w/w), kemudian sampel 2; proteinase K tanpa denaturasi dicampur dengan IP (1:1, w/w), sampel 3; IP tanpa denaturasi, sampel 4; proteinase K (4 mg/uL) denaturasi selama 15 menit pada 100 ⁰C, sampel 5; proteinase K (8 mg/uL) tanpa denaturasi. Sampel diinkubasi selama 24 jam pada 37 ºC. Aktivitas chaperon terdeteksi oleh aktivitas Proteinase K dan ditunjukkan dengan pembentukan zona bening.
Strain dan plasmid Karakteristik Sumber atau acuan Strain Isolat asal ayam, sumber slpA
10
Isolasi Total RNA dan Analisis RT-PCR
Total RNA dari L. plantarum U10 diperoleh dengan menumbuhkan L. plantarum U10 dalam medium MRS selama 13 jam pada 30 ºC. Setelah itu, sel dipindahkan ke dalam media MRS segar kemudian diinkubasi pada kondisi yang sama sampai OD 600 mencapai 0,6. Sampel dipanen dengan sentrifugasi pada 13000 x g. Perlakuan kejut panas dilakukan dengan menambahkan 1 mL MRS segar pada pelet dan kemudian dipanaskan di 42 ºC selama 15 menit dan kontrol tanpa perlakuan heat shock, kemudian dibiarkan pada suhu ruang selama 20 menit. Langkah selanjutnya isolasi RNA mengikuti panduan Ribo Pure Bakteri Kit Ambion (Lampiran 3). Kualitas sampel RNA diperiksa pada 1,2% agrose gel, dan konsentrasi ditentukan secara spektrofotometri pada 260 nm. Sekitar 150 ng RNA total yang digunakan dalam volume akhir 25 uL untuk sampel RT-PCR. Program RT-PCR adalah sebagai berikut: 45 ºC, 30 menit (reaksi reverse transcriptase), 94 ºC, 5 menit; 94 ºC, 30 detik (denaturasi); 52 ºC, 1 menit (anealing); 72 ºC, 1 menit; 72 ºC, 5 menit (ekstensi). Fragmen PCR kemudian divisualisasikan pada gel agarosa 1,2%.
Isolasi DNA Genom L. plantarum U10 dan L. acidophilus C9-9
Genom L. plantarum U10 dan L. acidophilus C9-9 diisolasi dengan menggunakan metode Mustopa dan Fatimah (2014). L. plantarum U10 dan L. acidophilus C9-9 ditumbuhkan di dalam media 5 mL MRS (de Man, Rogosa, Sharpe) (Lampiran 1) dipanen pada fase stasioner dengan sentrifugasi pada kecepatan 11000 x g selama 10 menit. Pelet yang didapatkan diresuspen dengan 540 µL bufer Tris-EDTA yang mengandung 5 mg/mL lisozim, kemudian diinkubasi selama 1 jam pada suhu 37 ⁰C. Proses lisis sel dilanjutkan dengan menambahkan 200 µL 10% sodium dedocyl sulfate, 100 µL 5M NaCl, dan 80 µL 10% CTAB kemudian diinkubasi selama 30 menit pada suhu 68 ⁰C. Campuran tersebut ditambahkan kloroform dengan perbandingan 1:1, kemudian disentrifugasi pada kecepatan 23000 x g selama 10 menit. Sentrifugasi menghasilkan tiga fase larutan dimana fase paling atas merupakan cairan bening yang mengandung nukleotida untuk kemudian dipresipitasi dengan isopropanol. Pelet atau DNA yang diendapkan disentrifugasi dan dikering-anginkan untuk menghilangkan sisa alkohol. DNA dilarutkan dengan nuclease-free water yang telah mengandung 0.1 mg/mL RNAse. DNA disimpan pada suhu -20 ⁰C.
Kloning pGEM-T slpA dan pGEM-T sHSP
11 terligasi dengan vektor pGEM-T kemudian diintroduksikan ke dalam E.coli TOP 10 (Sambrook et al. 2002). Vektor rekombinan pGEM-T_slpA dan pGEM-T_sHSP dikonfirmasi dengan menggunakan PCR koloni, PCR plasmid dan sekuensing. Karakteristik promotor slpA dan gen sHSP dilihat melalui homologi sekuen nukleotida dan asam amino yang analisis menggunakan BLAST (http://blast.ncbi.nlm.nih.gov/Blast.cgi) dan Bioedit versi 7.0.9.0 (Tom Hall). Tabel 3 Kondisi reaksi PCR
Konstruksi Vektor Ekspresi dengan Fusi Promotor slpA dan Gen sHSP Fusi gen dilakukan dengan merancang primer overlapping antara promotor slpA dan gen penyandi sHSP. Proses PCR dilakukan tiga tahap, tahap pertama yaitu amplifikasi promotor slpA dari template pGEM-T_slpA. Tahap kedua yaitu amplifikasi gen sHSP template pGEM-T_sHSP. Tahap ketiga yaitu amplifikasi fusi antara promotor slpA dan gen sHSP. Gen yang telah didapatkan kemudian dimurnikan dengan QIAquick® Gel Extraction Kit. Gen yang telah murni kemudian dipotong menggunakan enzim restriksi, sebanyak 5-15 ug DNA plasmid, 2 µL larutan penyangga reaksi 10x (sesuai enzim yang digunakan), 1 µL enzim restriksi (10 unit/µL) dan ddH2O hingga volume larutan menjadi 20 µL kemudian diinkubasi pada suhu 37 ⁰C selama 2 jam. Enzim restriksi yang digunakan adalah SalI-BglII untuk gen fusi, selain itu plasmid pNZ8148 juga dipotong menggunakan enzim yang sama untuk memotong bagian kloramfenikolnya. Gen maupun plasmid yang telah terpotong kemudian dimurnikan kembali sebelum dilakukan ligasi. Ligasi antara insert dan vektor dengan perbandingan 3:1 hingga volume akhir 15 µl dilakukan dengan menggunakan T4 DNA ligase (NEB) dan diinkubasi pada suhu 40C selama overnight.
Introduksi Hasil Ligasi ke dalam E.coli dan L.lactis
Transformasi E. coli dilakukan dengan metode Heat shock (Sambrook et al. 2001). Pembuatan sel kompeten dilakukan dengan menumbuhkan E. coli TOP 10 di dalam 5 ml LB (Lampiran 4) pada inkubator goyang kecepatan 150 rpm pada suhu 37 oC selama semalam. Selanjutnya 1 mL dari kultur yang diinkubasi selama semalam dikultur kembali di dalam 100 mL LB pada suhu 37 oC pada inkubator goyang dengan kecepatan 150 rpm. Pemanenan dilakukan saat OD600 ~0,4. Sentrifugasi dilakukan pada kecepatan 2700 x g pada suhu 4 ºC selama 10 menit. Supernatan hasil sentrifugasi dipisahkan dari pelet, sebanyak 30 mL larutan CaCl2-MgCl2 (80 mM MgCl2, 20 mM CaCl2) ditambahkan ke dalam pelet hingga teresuspensi. Sentrifugasi kembali pada kecepatan 2700 x g pada suhu 4 ºC selama 10 menit. Supernatan hasil sentrifugasi dipisahkan dari pelet. Setelah tidak ada lagi sisa supernatan, maka dilakukan penambahan 2 mL 0,1 M CaCl2.
12
Selanjutnya sel kompeten ini ditransformasi. Transformasi dilakukan dengan mencampurkan 100 µ L sel kompeten dengan 5 µ L hasil ligasi. Kemudian campuran tersebut diinkubasi di dalam es selama 30 menit, disusul dengan dipanaskan di dalam water bath pada 42 oC selama 90 detik. Setelah pemanasan selesai, dengan segera di pindahkan kedalam es dan diinkubasi selama2 menit. Sebanyak 400 µ L LB ditambahkan ke dalam eppendof dan diinkubasi pada suhu 37 oC selama 2 jam. Proses selanjutnya adalah penumbuhan hasil transformasi ke media LB agar yang telah ditambah 100 µg/mL ampisilin sebagai penanda seleksi. Penumbuhan hasil transformasi dilakukan dengan teknik surface plate atau spread plate. Sebanyak 100 µ L hasil transformasi ditumbuhkan di atas medium menggunakan triangle dryglass. Sel kompeten juga ditumbuhkan di LB agar yang mengandung ampisilin sebagai kontrol positif dan tanpa ampisilin sebagai kontrol negatif. Sel kompeten yang ditumbuhkan sebanyak 50 µ L. Selanjutnya hasil penumbuhan tersebut diinkubasi pada suhu 37 oC selama 16 jam. PCR koloni dilakukan terhadap transforman yang tumbuh. Koloni tunggal yang mengandung plasmid dengan sisipan gen target ditumbuhkan pada media LB untuk kemudian diisolasi plasmid rekombinan yang membawa gen target. Isolasi plasmid pada E. coli dilakukan dengan menggunakan GenJET Plasmid Miniprep Kit (Thermo Scientific). Plasmid yang didapatkan selanjutnya digunakan sebagai template dalam PCR dan disekuensing untuk mengkonfirmasi gen yang telah disisipkan ke dalam plasmid.
13
4
HASIL DAN PEMBAHASAN
Pengaruh Perlakuan Kejut Panas pada Profil Ekspresi Protein L. plantarum U10
Karakterisasi gen small heat shock protein (sHSP) dilakukan dengan memberikan perlakuan kejut panas pada L. plantarum U10. Tujuan dari perlakuan ini adalah untuk melihat keragaman dan fungsi protein heat shock pada pertumbuhan dan kelangsungan hidup L. plantarum U10. Bakteri L. plantarum U10 diberi perlakuan dengan inkubasi pada suhu yang berbeda yaitu 42 ⁰C (Delmas et al. 2001 dan Guzzo et al. 1997). Pengaruh dari kejut panas (42 ⁰C) pada ekspresi protein diamati selama 30 menit dan 45 menit. Hasil pengamatan menunjukkan bahwa ekspresi protein dinyatakan dengan berat molekul yang beragam (18,16 kDa, 34 kDa, 40,48 kDa dan 51,93 kDa). Hasil SDS menunjukkan bahwa lama waktu induksi perlakuan panas berpengaruh terhadap ekspresi protein. Waktu induksi perlakuan panas yang lebih lama (45 menit) menunjukkan hasil ekpresi protein berkurang apa bila dibandingkan dengan waktu induksi 30 menit, hal itu disebabkan karena perbedaan pada waktu degradasi protein dalam sel (Gambar 5).
Gambar 5 Profil ekspresi protein intraseluler L. Plantarum U10 setelah perlakuan kejut panas. Marker protein (kDa). Kontrol L. plantarum tanpa perlakuan. Bakteri diinkubasi di 42 ⁰C selama 30 menit dan 45 menit Pada penelitian ini ditemukan beberapa protein dari L. plantarum U10 setelah diberikan perlakuan suhu menjadi 42 ⁰C dengan massa protein berkisar 18,16 kDa sampai dengan 51,93 kDa, yang diprediksi sebagai kelompok heat shock protein. Guzzo et al. 1997 melaporkan bahwa protein dari intraselular sel L. oenos memiliki berat molekul yang beragam (75 kDa, 66 kDa, 64 kDa, 24 kDa, 18 kDa dan 14,5 kDa) setelah diberi perlakuan panas dengan suhu 42 ⁰C, OD 0,4. Protein dengan berat molekul 18 kDa menunjukkan ekspresi yang paling tinggi di bawah tekanan panas dan asam. small Heat shock Protein (sHSP) dengan berat molekul 18 kDa memiliki peranan penting dalam adaptasi dan kelangsungan hidup bakteri asam laktat terhadap tekanan lingkungan.
14
tidak ada perbedaan yang signifikan antara kelompok perlakuan panas (3.3x1010 CFU uL/1 selama 30 menit induksi, dan 3.2x1010 CFU uL/1 selama 45 menit induksi) apabila dibandingkan dengan kelompok kontrol (3x1010 CFU uL/1). Hal ini disebabkan karena suhu perlakuan yang diberikan tidak berbeda jauh dengan suhu kontrol. Delmas et al. 2001 melaporkan bahwa pengaruh tekanan suhu 45 ⁰C sampai 60 ⁰C terhadap sHSP Lo18 Oenococcus oeni dapat meningkatkan ketahanan hidup bakteri Oenococcu oeni dibandingkan dengan kontrol (30 ⁰C). Hasil pada penelitian ini menunjukkan bahwa ekspresi protein meningkat pada L. plantarum U10 adalah karena efek perlakuan stres panas.
Tabel 4 Profil ekspresi protein intraseluler L. plantarum U10 setelah perlakuan kejut panas. Populasi bakteri dihitung sebelum dan sesudah perlakuan kejut panas dan ditampilkan sebagai CFU/ml
Ekspresi Gen dari Kejut Panas L. plantarum U10 dan Uji Aktivitas Chaperon
Lactobacillus plantarum U10 merupakan bakteri asam laktat yang diisolasi dari Tempoyak. Fiacco et al. 2007 menyatakan bahwa peranan sHSP dalam adaptasi dan kelangsungan hidup BAL di bawah kondisi tekanan tertentu, seperti stres abiotik panas, dingin, dan etanol sangat berdampak pada tingkat ekspresi dari mRNA sHSP dan juga kuantitas produksi sHSP. Hasil karakterisasi menunjukkan adanya protein dengan ukuran 18.16 kDa yang diperkirakan merupakan protein sHSP. Selain itu, konfirmasi dilakukan dengan Reverse transcriptase-PCR pada tingkat ekspresi mRNA sHSP. RNA hasil isolasi dari L. plantarum U10 kemudian dijadikan cetakan untuk RT-PCR. Ekspresi dari gen penyandi sHSP dari L. plantarum U10 diekspresikan setelah suhu lingkungan pertumbuhan sel menjadi 42 ⁰C. Hasil RT-PCR menunjukkan terdapat pita dengan ukuran 423 pb (sHSP) dan tidak ada pita yang berhubungan dengan sHSP pada kontrol. Kontrol pada perlakuan ini tanpa kejut panas (Gambar 6). Hasil ini menunjukkan gen protein heat shock diekpresikan di bawah lingkungan stres panas.
Gambar 6 Analisis cDNA sHSP L. plantarum U10 1: sampel yang diberi perlakuan panas dan 2: sampel tanpa perlakuan panas, M: 100 pb Kappa universal DNA ladder. Pita yang sesuai dengan mRNA sHSP ditunjukkan oleh panah kuning.
Perlakuan CFU/mL
Kontrol 3.0x1010 CFU/mL
15
Protein Heat Shock diketahui mengandung aktivitas chaperon dimana protein ini dalam kondisi stres akan membantu dalam mencegah protein intraselular (IP) dari mekanisme degradasi seluler (Ehrnsperger et al. 1997, Veinger et al. 1998, Lee and Vierling 2000). Pada penelitian ini dilakukan metode sederhana berdasarkan modifikasi Collada et al. 1997 dan Kim et al. 1998 untuk mendeteksi aktivitas chaperon dalam IP L. plantarum menggunakan uji aktivitas enzim proteolitik pada agarose. Metode ini berdasarkan aksi perlindungan yang diduga protein chaperon terhadap degradasi enzim proteolitik karena perlakuan panas dengan pengukuran aktivitas residu enzim setelah perlakuan panas pada protein pendamping yang mengandung sampel. Uji aktivitas chaperon dilakukan untuk mengkonfirmasi keterlibatan gen sHSP pada L. plantarum U10 terhadap ketahanan stres panas. Uji aktivitas chaperon pada fraksi protein interselular (IP) dari L. plantarum U10 dengan mengamati aktivitas proteinase K. Hasil fraksi IP menunjukkan aktivitas chaperon (Gambar 7). Sampel 1 menunjukkan adanya aktivitas setelah denaturasi (1.3±0.11 cm) (Lampiran 6), sedangkan sampel 4 proteinase K sendiri tanpa IP kehilangan aktivitas proteinase K setelah denaturasi (0.73±0.11 cm). Proteinase K sampel 5 tanpa denaturasi memiliki aktivitas yang tinggi (3.36±0.11 cm), sedangkan pada sampel 3 (IP didenaturasi) tidak memiliki aktivitas proteinase. Hasil ini menunjukkan bahwa resistensi panas L. plantarum U10 dimungkinkan karena aktivitas cheperon di dalam selnya.
Gambar 7 Uji aktivitas chaperon. Sampel 1: proteinase K denaturasi dicampur dengan IP, 2: proteinase K tanpa denaturasi dicampur dengan IP, 3: IP tanpa denaturasi, 4: proteinase K denaturasi, 5: proteinase K tanpa denaturasi.
16
Konstruksi Vektor Kloning pGEM-T slpA dan pGEM-T sHSP
Surface layer protein A memiliki bentuk kristal pada permukaan sel dan telah diisolasi lebih dari 300 spesies yang berbeda termasuk bakteri Gram positif dan Gram negatif. slpA terdiri dari subunit protein tunggal atau glikoprotein dengan berbagai ukuran 40-200 kDa. Promotor surface layer protein A (slpA) merupakan salah satu promotor kuat yang telah diisolasi dari tipe liar Lactobacillus acidophilus oleh McCracken 1999. Pada studi ini promotor slpA diisolasi dari dari isolat Lactobacillus acidophilus dengan kode isolat C9-9. Promotor slpA merupakan salah satu promotor konstitutif yang umum digunakan dalam ekspresi BAL dari Lactobacillus brevis, dimana telah diketahui dapat meningkatkan level ekspresi sebesar 28% terhadap protein amino peptidase N (Diep et al. 2009).
Konstruksi vektor kloning dilakukan dengan isolasi genom dari L. plantarum U10 dan L. acidophilus C-9-9. L. plantarum U10 digunakan sebagai sumber untuk gen sHSP dan L. acidophilus C-9-9 sebagai sumber promotor slpA. Promotor slpA telah berhasil diamplifikasi dengan menggunnakan primer spesifik promotor slpA dengan menggunakan metode PCR. Hasil amplifikasi promotor slpA menggunakan primer slpA-F dan primer slpA-R menunjukkan bahwa ukuran amplikon adalah sebesar 192 pb (Gambar 8a). Gen promotor slpA tersebut telah berhasil diligasikan dengan plasmid pGEM-Teasy (Gambar 8b) dan ditransformasikan pada E.coli TOP10.
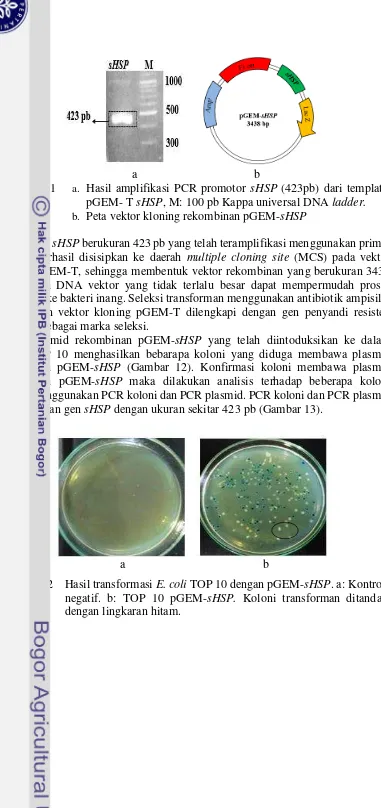
a b
Gambar 8 a. Hasil amplifikasi PCR promotor slpA (192pb) dari template pGEM- T slpA, M: 100 pb Kappa universal DNA ladder.
b. Peta vektor kloning rekombinan pGEM-slpA
17
a b
a b
Gen small Heat Shock Protein (sHSP) yang merupakan protein yang berperan dalam stres panas. Studi tentang peranan gen sHSP terhadap tekanan panas pada L. plantarum telah dilakukan. Gen sHSP dari L. plantarum (wine) telah diklon ke vektor pGEM-T (promega) dan diintroduksikan ke E. coli JM109 (Spano et al. 2005). Gen sHSP pada penelitian ini diisolasi dari isolat Lactobacillus plantarum dengan kode isolat U10. Heat Shock Protein terutama dengan massa molekul ±18 kDa telah dipelajari secara intensif karena memiliki peranan yang penting dalam pertumbuhan mikroorganisme, adaptasi dan kelangsungan hidup dalam respon terhadap stres lingkungan (Sugimoto et al. 2008, Tsakalidou dan Papadimitriou 2011, De Angelis dan Gobetti 2011). Kelompok protein ini dalam industri makanan menunjukkan dampak yang signifikan terhadap peningkatan BAL terutama ketika digunakan sebagai strain pemula (Carvalho et al. 2004, Ricciardi et al. 2012).
Hasil amplifikasi gen sHSP juga dilakukan menggunakan primer sHSP-F dan primer sHSP-R menunjukkan bahwa ukuran amplikon adalah sebesar 423 pb (Gambar 9a). Gen sHSP tersebut juga telah berhasil diligasikan dengan plasmid pGEM-Teasy (Gambar 9b) dan ditransformasikan pada E.coli TOP10.
Gambar 9 Hasil transformasi E. coli TOP 10 dengan pGEM-slpA. a: Kontrol negatif. b: TOP 10 pGEM-slpA. Koloni transforman ditandai dengan lingkaran hitam.
Gambar 10 a. Hasil PCR Koloni E. coli TOP 10 dengan pGEM-slpA. M: 100 pb Kappa universal DNA ladder , 1, 2, 3, 4 dan 5 koloni rekombinan.
18
a b
Gambar 11 a. Hasil amplifikasi PCR promotor sHSP (423pb) dari template pGEM- T sHSP, M: 100 pb Kappa universal DNA ladder.
b. Peta vektor kloning rekombinan pGEM-sHSP
Gen sHSP berukuran 423 pb yang telah teramplifikasi menggunakan primer spesifik berhasil disisipkan ke daerah multiple cloning site (MCS) pada vektor kloning pGEM-T, sehingga membentuk vektor rekombinan yang berukuran 3438 pb. Ukuran DNA vektor yang tidak terlalu besar dapat mempermudah proses introduksi ke bakteri inang. Seleksi transforman menggunakan antibiotik ampisilin dikarenakan vektor kloning pGEM-T dilengkapi dengan gen penyandi resisten ampisilin sebagai marka seleksi.
Plasmid rekombinan pGEM-sHSP yang telah diintoduksikan ke dalam E.coli TOP 10 menghasilkan bebarapa koloni yang diduga membawa plasmid rekombinan pGEM-sHSP (Gambar 12). Konfirmasi koloni membawa plasmid rekombinan pGEM-sHSP maka dilakukan analisis terhadap beberapa koloni dengan menggunakan PCR koloni dan PCR plasmid. PCR koloni dan PCR plasmid menghasilkan gen sHSP dengan ukuran sekitar 423 pb (Gambar 13).
a b
19
a b
Kromatografi sekuensing promotor slpA dan gen sHSP di E. coli TOP 10 (Lampiran 7) selanjutnya digunakan untuk analisis sekuen. Hasil analisis sekuen menunjukkan Promotor slpA berukuran 192 pb memiliki similarity yang tinggi (99%) apabila dibandingkan dengan sekuen referensi L. acidhopilus ATCC 4356 (Gambar 14). Hasil BLAST menunjukkan urutan nukleotida sHSP dari L. plantarum U10 memiliki tingkat homolog yang tinggi (100%) dengan sHSP dari L. plantarum WCFS1 referensi (Gambar 15). Hasil analisis sekuen menunjukkan tersebut menunjukkan bahwa hasil kloning ini dapat digunakan sebagai sumber untuk mendapatkan promotor slpA dan sekuen gen penyandi small Heat Shock Protein (sHSP).
Gambar 13 a. Hasil PCR Koloni E. coli TOP 10 dengan pGEM-sHSP. M: 100 pb Kappa universal DNA ladder, 1,2,4: koloni rekombinan. 3: koloni bukan rekombinan.
b. PCR Plasmid E. coli TOP 10 dengan pGEM-sHSP. M: 100 pb Kappa universal DNA ladder.
20
Konstruksi Vektor Ekspresi dengan Fusi Promotor slpA dan Gen sHSP
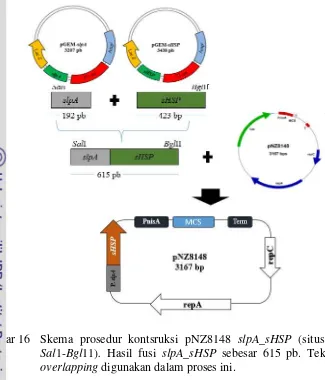
Metode yang digunakan untuk memfusikan suatu gen dengan promoter lain terdapat beberapa metode. Metode tersebut antara lain dengan menggunakan linker dan PCR overlapping. Tian et al. 2012 melakukan konstruksi plasmid dengan menggunakan PCR overlapping dari fragmen yang mengandung promotor 32 dan gen penyandi sHSP. Pada penelitian konstruksi vektor rekombinan dilakukan dengan menggunakan PCR overlapping dari fragmen promotor slpA dan gen penyandi sHSP (Gambar 16).
Konstruksi vektor ekspresi dilakukan dengan PCR promotor slpA dari pGEM-slpA dan PCR gen sHSP dari pGEM-sHSP. Hasil PCR dijadikan sebagai template untuk PCR Fusi dengan teknik overlapping. PCR overlapping dilakukan dengan mendesain primer spesifik untuk fusi promotor slpA dan gen sHSP. Pada penelitian ini primer slpA-sHSP-F1 dengan situs restriksi SalI, primer slpA-sHSP-F2 dan R-2 dengan bagian tumpang tindih (overlap), yaitu urutan nukleotida merupakan bagian dari promotor slpA dan gen sHSP, dan primer slpA-sHSP-R1 dengan situs restriksi BglII.
21
Gambar 16 Skema prosedur kontsruksi pNZ8148 slpA_sHSP (situs restriksi Sal1-Bgl11). Hasil fusi slpA_sHSP sebesar 615 pb. Teknik PCR overlapping digunakan dalam proses ini.
Promotor slpA dengan ukuran 192 pb dan untuk gen penyandi sHSP berukuran 423 pb. Hasil fusi antara promotor slpA dan gen sHSP didapatkan ukuran 615 pb (Gambar 17a). Dari gambar tersebut terlihat bahwa hasil amplifikasi fusi memiliki ukuran yang lebih tinggi apabila dibandingkan dengan masing-masing fragmen. Hal ini menunjukkan bahwa fusi antara promotor slpA dan gen penyandi sHSP telah berhasil diamplifikasi dengan teknik PCR Overlapping. Food grade vector dibentuk dengan mengganti bagian antibiotik dari vektor ekspresi yang digunakan. Pada penelitian ini vektor ekspresi yang digunakan adalah pNZ8148 dengan menghapus bagian gen yang menyandikan pendegradasi kloramfenikol.
Fusi antara promotor slpA dan gen penyandi sHSP kemudian disisipkan pada vektor pNZ8148 pada bagian kloramfenikol yang telah dipotong dengan enzim restriksi Sal1-BglII sehingga terbentuk plasmid pNZ8148 rekombinan pNZ8148-slpA_sHSP. Plasmid rekombinan kemudian diintroduksikan ke inang L. lactis (Gambar 17b). Bakteri L. lactis NZ3900 merupakan turunan dari L. lactis MG1363 yang diketahui tidak memiliki gen penghasil nisin (Kunji et al. 2003)
22
sehingga membentuk pori pada dinding/membran sel yang dapat tersisipi DNA asing (Rattanachaikunsopon dan Phumkachorn 2009). Penambahan sukrosa dan glisin dapat membantu proses transformasi menjadi lebih efisien. Selain sukrosa sebagai penstabil kondisi osmotik, glisin yang ditambahkan pada media pertumbuhan BAL diketahui dapat melemahkan membran sel bakteri inang dengan cara menghambat pembentukan ikatan silang yang akan memperkuat membran sel tersebut (Heravi et al. 2012). Dalam penelitian ini, digunakan sebanyak 2.5% glisin untuk melemahkan membran sel bakteri inang. Penggunaan glisin yang terlalu tinggi (lebih dari 8%) akan berdampak pada menurunnya efisiensi transformasi, dikarenakan memicu terjadinya autolisis bakteri (Kim et al. 2005).
a b
Hasil transformasi menunjukkan tingkat efisiensi transformasi yang rendah. Hal ini dapat dikarenakan oleh karakteristik dari gen replikasi vektor pNZ8148. Vektor ekspresi pNZ8148 diketahui tidak memiliki tingkat replikasi/penggandaan vektor yang cukup tinggi, akan tetapi hal ini diimbangi dengan adanya dua titik awal replikasi (repA dan repC) yang mudah dikenali inang baik E. coli maupun L. lactis (de Ruyter et al. 1996). Koloni transforman yang tumbuh dijadikan cetakan PCR koloni.
Hasil PCR koloni menggunakan primer F/R gen fusi slpA-sHSP (Gambar 18) pada L. lactis rekombinan menunjukkan bahwa dari empat koloni tranforman yang tumbuh, hanya dua koloni yang positif membawa sisipan fusi promotor slpA dan gen sHSP. Dari gambar tersebut terlihat bahwa L. lactis rekombinan memiliki ukuran yang sama dengan kontrol positif gen fusi pGEM-slpA-sHSP (615 pb). Hal ini dapat disimpulkan bahwa fusi gen telah berhasil disisipkan ke dalam vektor pNZ8148 dan diintroduksikan ke dalam L. lactis. Selain itu konfirmasi vektor ekspresi pNZ8148 yang berhasil diintroduksikan ke L. lactis NZ3900 juga dilakukan dengan PCR koloni menggunakan primer promotor NisA dan teminator pNZ8148 (Gambar 19) pada L. lactis rekombinan. Hasil PCR menunjukkan bahwa pNZ8148 berhasil diintroduksikan ke dalam inang L. lactis dengan ukuran promotor sampai terminator yang sama dengan kontrol positif. Kontrol positif pada PCR dengan menggunakan plasmid pNZ8148.
Gambar 17 a. Hasil amplifikasi fusi slpA dan sHSP. M: 100 pb Kappa universal DNA ladder. 1: hasil amplifikasi promotor slpA (192 pb) , 2: hasil amplifikasi gen penyandi sHSP (423 pb) dan 3: fusi slpA dan sHSP (615 pb)
23
Gambar 18 Hasil PCR koloni menggunakan primer F/R gen fusi pada L. lactis rekombinan. 1 dan 3: L. lactis rekombinan fusi slpA-sHSP, 5: kontrol positif (fusi). M: 100 pb Kappa universal DNA ladder.
Gambar 19 Hasil PCR koloni menggunakan primer F/R Pro NisA dan Term pNZ8148. 1: kontrol positif (pNZ8148), 2 dan 4: L. lactis pNZ8148. M: 100 pb Kappa universal DNA ladder.
24
5
SIMPULAN DAN SARAN
Simpulan
Karakterisasi terhadap gen small Heat Shock Protein (sHSP) pada isolat L. plantarum U10 yang diisolasi dari makanan tradisional Indonesia Tempoyak dengan berat molekul 18,16 kDa. Protein ini diekspresikan dengan perlakuan kejut panas pada perubahan suhu menjadi 42 ⁰C. Promotor slpA yang berukuran 192 pb dengan gen sHSP yang berukuran 423 pb telah berhasil dikloning ke vektor pGEM-T. Promotor slpA dengan ukuran 192 pb juga telah berhasil difusikan dengan gen sHSP yang berukuran 423 pb sehingga diperoleh ukuran fusi 615 pb. Fusi promotor slpA dan gen sHSP kemudian dikloning ke dalam vektor pNZ8148 dan diekspresikan pada inang L.lactis NZ3900.
Saran
25
DAFTAR PUSTAKA
Boucher I, Parrot M, Gaudreau H, Moineau S. 2002. Novel food-grade plasmid vector based on melibiose fermentation for the genetic engineering of Lactococcus lactis. Appl Environ Microbiol. 68:6152-6161.
Bron PA, Benchimol MG, Lambert J, Palumbo E, Deghorain M, Delcour J, De Vos WM, Kleerebezem M, Hols P. 2002. Use of the alr gene as a food-grade selection marker in lactic acid bacteria. Appl Environ Microbiol. 68:5663-5670.
Carvalho AS, Silva J, Ho P, Teixeira P, Malcata FX, Gibbs P, 2004. Relevant factors for the preparation of freeze dried lactic acid bacteria. Int Dairy J.14:835–847.doi: 10.1016/j.idairyj. 2004.02.001.
Cavanagh D, Fitzgerald GF, McAuliffe O. 2015. From field to fermentation the origins of Lactococcus lactis and its domestication to dairy environment. Food Microbiol. 47:45-61.
Collada C, Gomez L, Casado R, Aragoncillo C. 1997. Purification and invitro chaperone activity of a class I small heat-shock protein abundant chestnut seeds. Plant Physiol. 115:71–77.
De Angelis M, Gobbetti M. 2011. Stress responses of Lactobacilli. Springer New York. 23:219–249.
de Ruyter PG, Kuipers OP, de Vos WM. 1996. Controlled gene expression system for Lactococcus lactis with the food grade inducer nisin. Appl Environ Microbiol. 62:3662-3667.
Delmas, Francoise, Fabrice P, Coucheney, Charles D, Jean G. 2001. Biochemical and physiological studies of the small heat shock protein Lo18 from the lacic acid bacterium Oenococcus oeni. Microbiol Biotechnology. 3(4):601-610. Dickely. 1995. Isolation of L. Lactis nonsense suppressors and construction of a
food grade cloning vector. Mol Microbiol.15:839-847.
Diep DB, Havarstein LS, Nes IF. 1996. Characterization of the locus responsible for the bacteriosin production in Lactobacillus plantarum C11. J Bacteriol. 178:4472-4483.
Donkor ES, Bishop CJ, Antonio M, Wren M, Hanage WM. 2011. High levels of recombination among Streptococcus pnemoniae isolates from the Gambia. Microbiol. 3(9):602-615.
El Demerdash HAM, Heller KJ, Geis A. 2003. Application of the sHSP gene, encoding a small heat shock protein, as a food-grade selection marker for lactic acid bacteria. Appl Environ Microbiol. 69: 4408-4412.
El Demerdash HAM, Oxmann J, Heller KJ, Geis, A. 2006. Yoghurt fermentation at elevated temperatures by strains of Streptococcus thermophilus expressing a small heat shock protein application of a two-plasmid system for constructing food-grade strains of Streptococcus thermophilus. Biotechnol J.69:398-404.
Ehrnsperger M, Gräber S, Gaestel M, and Buchner, J. 1997. Binding of non-native protein to HSP25 during heats hock creates a reservoir of folding intermediates for reactivation. EMBO J. 16: 221-229.
26
Guzzo J, Delmas F, Pieere F, Jobbin MP, Samyn B, Van Beeumen J, Cavin JF, Divies C. 1997. A small heat shock protein from Leuconoctoc oenos Iinduced by multiple stresses and during stationary growth phase. Microbiol.24:393-396.doi:10.1046/j.1472-765x.1997.00042.
Hammes WP, Vogel RF. 1995. The genus Lactobacillus. Di dalam: Wood BJN, editor. The Genera of Lactic Acid Bacteria. New York (US): Chapman & Hall. 4(3):19-54.
Haslbeck, M. (2002). sHSP and their role in the chaperone network. Cell Mol Life Sci. 59:1649-1657.
Herman RE, McKay LL. 1985. Isolation and partial characterization of plasmid DNA from Streptococcus thermophilus. Appl Environ Microbiol. 50:1103-1106.
Johansen E. 2003. Challenges when transferring technology from Lactococcus laboratory strain to industrial strains. Appl Biotechnol. 2(1): 112-116. Jorgensen CM, Madsen SM, Vrang A, Hansen OC, Johnsen MG. 2013.
Recombinant expression of Laceyella sacchari thermitase in Lactococcus lactis. Protein Expression and Purification. 92:148–155.
Kim, Rosalind, Kyeong Kyu Kim, Hisao Yokata, Sung-Hou Kim. 1998. Small heat shock protein of Methanococcus jannaschii a hyperthermophile. Proc Natl Acad Sci USA. 95:9129-9133.
Kim I, Rasmus L, Annette K, Mette P, Junge. 2002. A food grade cloning system for Industrial Strains of Lactococcus lactis. Microbiol. 8(12):1253-1258. Kunji ERS, Slotboom D, Poolman B. 2003. Lactococcus lactis as host for over
production of functional membrne proteins. Biochim Biophys Acta. 1610: 97-108.
LagesAC, Mustopa AZ, L Sukmarini, Suharsono. 2015. Cloning and Expression of Plantaricin W Produced by Lactobacillus plantarum U10 Isolate from
‘Tempoyak’ Indonesian Fermented Food as Immunity Protein in Lactococcus lactis. Appl Biochem Biotechnol. 38:126-135.doi :10.1007/s12010-015-1786-9.
Le Roux NJ, Steel SJ, Louw N. 1997. Variable section and Error rate estimation in discriminant analysis. J Statis Comput Simul. 99:195-219.
Lee GJ, dan Vierling E. 2000. A small Heat Shock Protein cooperates with Heat Shock Protein 70 systems toreactivate a heat-denatured protein. Plant Physiol. 122:189-198.doi: org/10.1104/pp.122.1.189.
Lelj Garolla B, Mauk AG. 20O6. Self-association and chaperone activity of HSP27 are thermally activaled. J Biol Chem. 281:8169-74.
Liang X, Sun Z, Zhong J, Zhang Q, Huan L. 2010. Adverse effect of nisin resistance protein on nisin induced expression system in Lactococcus lactis. Microbiol Research. 165:458-465.
Martín M, Gutiérrez J, Criado R, Herranz C, Cintas LM, Hernández PE. 2007. Cloning, production and expression of the bacteriocin enterocin A produced by Enterococcus faecium PLBC21 in Lactococcus lactis. Appl Microbiol Biotechnol. 76:667–675
27 McCracken Andrea, Peter Timms. 1999. Eficiency of transcription from promoter sequence variants in Lactobacilus is both strain and context dependent. J Bacteriol. 181:6569-6572.
Mierau I, Olieman K, Mond J dan Smid EJ. 2005. Optimization of the Lactococcus lactis nisin-controlled gene expression system NICE for industrial applications. Microbial Cell Factories. 68:705-717.
Mills S, Coffey A, O’Sullivan L, Stokes D, Hill C, Fitzgerald GF, Ross RP. 2002. Use of lacticin 481 to facilitate delivery of the bacteriophage resistance plasmid, pCBG104 to cheese starters. J Appl Microbiol. 92: 238-246.
Morelli L, FK Vogensen, AV. Wright. 2004. Genetics of lactic acid bacteria. Dalam: Salminen S, A. Wright, A. Ouwehand (Eds.). 2004. Lactic acid bacteria: Microbiological and functional aspects. 3rd ed. Marcel Dekker, Inc New York: xiii.
Mustopa AZ. Fatimah. 2014. Diversity of lactic acid bacteria isolated from Indonesian traditional fermented food. Microbiol Indones. 8:48-57.
Narberhaus, F. (2002). Crystallin-type heat shock proteins: socializing mini-chaperones in the context of a multichaperone network. Microbiol Mol Biol Rev. 66:64-93.
O'Sullivan T,Van Sinderen D, Fitzgerald G. 1999. Structural and functional analysis of pCI65st, a 6.5kb plasmid from Streptococcus thermophilus NDI-6. Microbiol. 145:127-134.
Rattanachaikunsopon P, Phumkhachorn P. 2009. Glass bead transformation method for gram-positive bacteria. Braz J Microbiol. 40: 923-926
Sambrook J, Fritsch EF, Maniatis T. 2001. Molecular cloning: a laboratory manual. New York: Cold spring harbor laboratory.
SchoffI Prendl, Reindl. 1998. A Regulation of the heat-shock response. Plant Physio. 17:35-41
Siezen RJ, Van Hylckama, V lieg. 2011. Genomic diversity and versatility of Lactobacillus plantarum, a natural metabolic engineer. Microb Cell Fact. 10(1):1-13.
Solow BT, Somkuti GA. (2000). Comparison of low-molecular weight heat stress proteins encoded on plasmids in different strains of Streptococcus thermophilus. Curr Microbiol. 41:177-181.
Spano G, Beneduce L, Perrotta C, Massa S. 2005. Cloning and characterization of the HSP 18.55 gene, a new member of the small heat shock genes family isolated from wine Lactobacillus plantarum. Research Microbiol. 156:219-224.doi: 10.1016/j.resmic.2004.09.014.
Sugimoto, Shinya, Abdullah AM, Kenji, S. 2008. Molecular chaperon in lactic acid bacteria physiological consequences and biochemical properties. J Bioscience and Bioengineering. 106: 324-336.doi:10.1263/jbb.106.324. Takala M, Saris PE. 2002. A food-grade cloning vector for lactic acid bacteria based
on the nisin immunity gene. Appl Microbiol Biotechnol.59:467-471.
28
Tsakalidou E, Papadimitriou K. 2011. Stress responses of lactic acid bacteria. Springer New York. 28:973-984.
Urnemi, A Zaenal M, M Ridwan. 2010. Potensi bakteri asam laktat dari lempok durian dalam menghasilkan bakteriosin sebagai biopreservatif pangan. Di dalam: Hernaman I, Tanuwiria UH, Hendronoto A, Yurmiati LH, Sulistyati M, Hidayati YA, Herlina L, Indrijani H, Sujana E, Putranto WS, Islami RZ, Widiawati Y, Sofjan O, Syamsul JA, editor. Sistem Produksi Berbasis Ekosistem Lokal. Seminar Nasional Peternakan Berkelanjutan; 2010 November 4; Bandung, Indonesia. Bandung (ID): Universitas Padjajaran. 679-685.
Veinger L, Diamant S, Buchner J, and Goloubinoff P. 1998. The small heat shock protein IpbB from Escherichia coli stabilizes stress-denatured proteins for subsequent refolding by a multichaperone network. J Biol Chem. 273: 11032-11037.
Westerheide SD, Morimoto RI. 2005. Heat shock response modulators as therapeutic tools for diseases of protein conformation. J Biol Chem. 280:33097-100.
Wu C, Juan Zhang J, Du G, Chen J. 2013. Heterologous expression of Lactobacillus casei RecO improved the multiple stress tolerance and lactic acid production in Lactococcus lactis NZ9000 during salt stress. Bioresource Tech. 143: 238– 241.
29
30
Lampiran 1. Komposisi media pertumbuhan de Man, Rogosa, Sharpe (MRS) (Oxoid)
Komposisi Media Jumlah
Peptom 10 g/L
Bubuk ‘Lab-Lemco’ 8 g/L
Ekstrak Khamir 4 g/L
Sorbitan mono-oleat 1 g/L
Di-potasium hidrogen fosfat 2 g/L
Sodium asetat 5 g/L
Tri-ammonium sitrat 2 g/L
Magnesium sulfat 0.2 g/L
Mangan sulfat 0.05 g/L
31 Lampiran 2. Komposisi larutan elektroforesis SDS-PAGE
Medium dan larutan-larutan
Bahan-bahan Jumlah
Larutan gel separating 10% H2O 6.25 ml
1.5 M Tris-HCl pH 8.8 containing 0.4% SDS
3.75 ml
30% Akrilamid 5 ml
10% amonium Persulfat 0.05 ml
TEMED 0.01 ml
10% amonium Persulfat 0.025 ml