MARKA SNAP BERBASIS FRAGMEN GENOMIK GEN SAD
SEBAGAI INDIKATOR KANDUNGAN ASAM LEMAK TIDAK JENUH
KELAPA SAWIT
RISMAYANTI
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA
Dengan ini saya menyatakan bahwa tesis berjudul “Marka SNAP berbasis Fragmen Gen SAD sebagai Indikator Kandungan Asam Lemak Tidak Jenuh Kelapa Sawit”, adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
SUMMARY
RISMAYANTI. Marka SNAP Base on SAD Gene Fragment for Unsaturated Fatty Acid Indicator of Oil Palm. Supervised by SUDARSONO as chairman, NURUL KHUMAIDA and WAHYU PURBOWASITO as member of advisory committee.
The oil palm is a major vegetable oil producing plant which has the highest yields per hectare of all oil crops. It is necessary to improved, high-yield genetic material with better oil quality to meet the increase global demand for healthy and cheap vegetable oil. The SNPs marka is one of technology, instead of convensional breeding technique that can be used to strenghten breeding program to get oil palm genetic material with high quality oil. The quality of oil affected by unsaturated fatty acids content in the oil. Oleic acid is main component of monounsaturated fatty acids that well known it is good for health, stabil and able to reduce heart disease risk. Stearoyl Acyl-carrier-protein Desaturase (SAD) is a key enzyme for oleic acid biosynthesis. This enzyme play an important role in determining composition of unsaturated fatty acids in oil palm (Elaeis sp.).
In this experiment, the genomic fragment of SAD isolated using SAD spesific primer (SAD1 and SAD2) from 5 sources of oil palm variety and used to identified SNPs (single nucleotide polymorphisms) loci. There were 13 loci of SNPs identified from SAD genomic fragments and they used to disain 26 pairs of oligonucleotide primers to generate 13 SNAP markas loci. The 13 SNAP marker were used to evaluate 50 oil palm accessions consisted of E. guineensis and E. oleifera. Out of the 13 SNAP marka loci analyzed, six (6) loci (2 in the intron, 3 in the exon and 1 in 3’UTRs part of SAD genomic fragment) were polymorphic for E. guineensis and E. oleifera acessions. Since E. oleifera is known to have different (high) unsaturated fatty acids compotition in its fruit than those of E. guineensis, these SNAP marka loci might be used to predict genetic variations and oleic acid biosynthetic activities which lead to different composition of unsaturated fatty acid content among segregated progenies derived from hybridization of E. oleifera and E. guineensis.
(SAD) sebagai Indikator Kandungan Asam Lemak Tidak Jenuh pada Kelapa Sawit”.
RINGKASAN
RISMAYANTI. Marka SNAP berbasis Fragmen Gen SAD sebagai Indikator Kandungan Asam Lemak Tidak Jenuh Kelapa Sawit. Dibimbing oleh SUDARSONO sebagai ketua, NURUL KHUMAIDA dan WAHYU PURBOWASITO sebagai anggota komisi pembimbing.
Kelapa sawit merupakan salah satu tanaman penghasil minyak nabati utama dengan produktifitas per hektar tertinggi dibandingkan semua tanaman penghasil minyak lainnya. Kebutuhan terhadap kualitas minyak nabati yang sehat dengan harga yang murah menjadi tren pada perkembangan industri minyak dan asam lemak secara global, diharapkan kelapa sawit dapat memenuhi kebutuhan tersebut. Oleh karena itu perlu dikembangkan materi genetik dengan produktifitas yang tinggi dan kualitas minyak yang baik, sebagai sumber bahan pangan maupun non pangan. Salah satu cara mempercepat mendapatkan tanaman unggul yang memenuhi kriteria yang diinginkan adalah dengan mengembangkan marka molekuler yang dapat mengindikasikan kualitas minyak yang dihasilkan tanaman kelapa sawit sejak dini.
SNPs (Single Nucleotide Polimorphisms) merupakan salah satu teknologi marka molekuler, selain pemuliaan konvensional yang dapat dimanfaatkan untuk memperkuat dan mempercepat program pemuliaan kelapa sawit.Kualitas minyak dipengaruhi oleh kandungan asam lemak tidak jenuh pada minyak. Asam oleat merupakan salah satu komponen utama asam lemak tidak jenuh tunggal yang baik untuk kesehatan, bersifat stabil dan mampu mengurangi resiko serangan penyakit jantung. Pembentukan dan kandungan asam oleat serta asam lemak tidak jenuh pada kelapa sawit dipengaruhi oleh gen kunci stearoyl ACP desaturase (SAD). Gen ini memainkan peran penting dalam menentukan komposisi asam lemak tidak jenuh pada kelapa sawit (Elaeis sp.). Dalam penelitian ini, fragmen gen SAD diisolasi dari genom sumber keragaman kelapa sawit untuk mengidentifikasi dan meanganalisis lokus SNP yang digunakan sebagai dasar pengembangan marka SNAP (Single Nucleotide Amplified Polimorphisme).
Dua pasang primer gen SAD spesifik berhasil didisain untuk mengisolasi fragmen gen SAD dari genom sumber keragaman kelapa sawit, yakni primer SAD1 dan SAD2. Fragmen gen SAD telah berhasil diisolasi dan diamplifikasi dari DNA genom 5 aksesi kelapa sawit yang terdiri atas satu aksesi spesies E.oleifera (open polinated) dan 4 aksesi dari spesies E.guineensis var tenera, Dura, Pisifera, dan Virescens.
Fragmen gen SAD yang diisolasi dari berbagai sumber keragaman kelapa sawit mempunyai tingkat homologi 99% dengan gen SAD yang terdeposit di NCBI (National Center for Biotechnology Information). Primer SAD1 berhasil mengamplifikasi semua sampel dengan ukuran 1100 bp dengan satu intron ukuran 507 bp. Primer SAD2 menghasilkan urutan basa dengan ukuran 600 bp. Total ukuran exon yang teramplifikasi berkisar antara 969-1032 bp. Hasil identifikasi dan analisis sekuens SAD menghasilkan 13 lokus SNPs gen SAD, yang kemudian dikembangkan untuk mendapatkan 26 pasang marka SNAP.
3’UTRs), yang dapat digunakan untuk menduga variasi aktivitas biosintesis dan komposisi asam lemak tidak jenuh dalam buah kelapa sawit umumnya dan kandungan asam oleat pada khususnya. Ini adalah hasil penelitian pertama yang menunjukkan gabungan lokus SNPs pada exon, intron dan 3’UTRs dapat mengindikasikan kandungan asam lemak tidak jenuh pada tanaman penghasil minyak nabati.
Hasil penelitian ini dapat dimanfaatkan oleh industri benih kelapa sawit untuk mendapatkan induk superior dengan kandungan asam oleat yang tinggi, meningkatkan efisiensi seleksi dan mengurangi jumlah generasi silang balik pada backcross E.oleifera dan E.guineesis, mengindentifikasi tanaman tenera yang mengalami mutasi alami pada gen SAD dengan komposisi kandungan asam lemak spesifik yang dapat diperbanyak dengan teknik kultur jaringan sehingga didapatkan tanaman klon tenera unggul alami tanpa melalui transformasi atau bukan GMO (Genetically Modified Organism). Dengan demikian dapat mempercepat program pemuliaan kelapa sawit untuk mendapatkan tanaman dengan kandungan asam oleat tinggi, yang baik untuk kesehatan dan memenuhi persyaratan internasional sebagai sumber bahan bioedisel. Hasil penelitian ini telah terdaftar di Direktorat Jenderal Hak Kekayaan Intelektual, Kementerian Hukum dan Hak Azazi Manusia Republik Indonesia untuk dipatenkan sejak 11 April 2014, nomor P00201402143 dengan judul Penanda SNAP (Single Nucleotide Amplified Polimorphism) Berbasis DNA Sekuens dari Fragmen Genomik Gen Stearoyl Acyl carrier protein Desaturase (SAD) sebagai Indikator Kandungan Asam Lemak Tidak Jenuh pada Kelapa Sawit.
© Hak Cipta Milik IPB, Tahun 2014
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains
pada
Program Studi Pemuliaan dan Bioteknologi Tanaman
MARKA SNAP BERBASIS FRAGMEN GENOMIK GEN SAD
SEBAGAI INDIKATOR KANDUNGAN ASAM LEMAK TIDAK
JENUH KELAPA SAWIT
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
Judul Tesis : Marka SNAP Berbasis Fragmen Gen SAD sebagai Indikator Kandungan Asam Lemak Tidak Jenuh Kelapa Sawit
Nama : Rismayanti NRP : A 253110051
Disetujui oleh Komisi Pembimbing
Prof Dr Ir Sudarsono MSc Ketua
Dr Ir Nurul Khumaida, MS Anggota
Dr Wahyu Purbowasito, MSc Anggota
Diketahui oleh
Ketua Program Studi
Pemuliaan dan Bioteknologi Tanaman
Dr Ir Yudiwanti Wahyu EK, MS
Dekan Sekolah Pascasarjana
Dr Ir Dahrul Syah, MScAgr
PRAKATA
Puji dan syukur penulis panjatkan kepada Allah swt atas segala karunia-Nya tesis ini berhasil diselesaikan. Tesis ini disusun berdasarkan penelitian dengan judul “Marka SNAP Berbasis Fragmen Gen SAD sebagai Indikator Kandungan Asam Lemak Tidak Jenuh Kelapa Sawit”.
Terima kasih penulis sampaikan kepada Prof Dr Ir Sudarsono MSc selaku ketua komisi pembimbing, Dr Ir Nurul Khumaida MS dan Dr Wahyu Purbowasito MSc sebagai anggota komisi pembimbing, yang telah banyak memberi saran dan bimbingan dalam penelitian dan penyusunan tesis ini. Disamping itu, penghargaan penulis sampaikan kepada Kepala Balai Pengkajian Bioteknologi, Badan Pengkajian dan Penerapan Teknologi dan seluruh staff serta Bapak Satyoso dan staf PT. Astra Agro Lestari, serta seluruh tim PMB Lab, IPB. Ungkapan terima kasih juga disampaikan kepada suami, papa (almarhum), mama, kakak dan adik-adik serta seluruh keluarga, atas segala doa, dukungan dan kasih sayangnya.
Penulis berharap semoga tesis ini dapat memberikan manfaat bagi kemajuan ilmu pemuliaan dan biologi molekuler tanaman penghasil minyak, khususnya perkembangan tanaman kelapa sawit. Penulis menyadari masih banyak kekurangan, besar harapan penulis atas saran dan kritik yang membangun untuk kesempurnaan tesis ini.
Bogor, Nofember 2014
DAFTAR ISI
DAFTAR TABEL DAFTAR GAMBAR DAFTAR LAMPIRAN xii xii xiii 1 PENDAHULUANLatar Belakang Tujuan Penelitian Manfaat Penelitian
Ruang Lingkup Penelitian
1 3 3 4 2 TINJAUAN PUSTIDAKA
Kelapa Sawit
Metabolisme Asam Lemak dan Gen SAD Pemuliaan Kelapa Sawit
Marka SNAP Berdasarkan SNP
5 8 10 10 3 ISOLASI FRAGMEN GEN STEAROYL ACP DESATURASE
(SAD) ASAL 5 SUMBER KERAGAMAN KELAPA SAWIT Pendahuluan
Bahan dan Metode Hasil dan Pembahasan Simpulan
14 15 17 22 4 IDENTIFIKASI DAN ANALISIS SNP BERDASARKAN
KERAGAMAN NUKLEOTIDA GEN SAD KELAPA SAWIT Pendahuluan
Bahan dan Metode Hasil dan Pembahasan Simpulan
25 27 28 34
5 APLIKASI MARKA SNAP UNTUK MEMPREDIKSI
KANDUNGAN ASAM OLEAT (ASAM LEMAK TIDAK JENUH) PADA KELAPA SAWIT
Pendahuluan Bahan dan Metode Hasil dan Pembahasan Simpulan
37 39 41 48 6 PEMBAHASAN UMUM
xii
DAFTAR TABEL
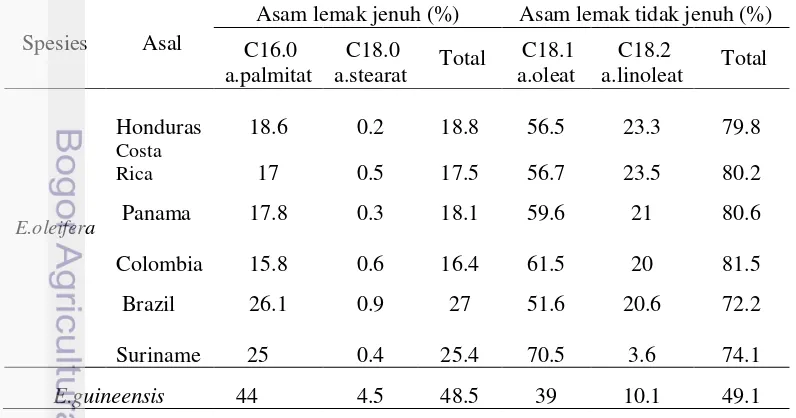
1 Komposisi asam lemak jenuh dan asam lemak tidak jenuh pada minyak sawit dari buah kelapa sawit spesies Elaeis guineensis
dan sejumlah aksesi E. oleifera. 14
2 Dua pasang primer SAD spesifik untuk mengamplifikasi genomik
fragmen gen SAD. 18
3 Hasil isolasi 5 sumber keragaman kelapa sawit 19 4 Contoh tingkat kemiripan sequense sampel (Tenera) dengan
informasi sequense SAD yang terdeposit di Bank gen, NCBI 21 5 Identifikasi situs SNPs berdasarkan hasil pensejajaran (multiple
allignment) DNA sequense fragmen genomik gen SAD dari spesies Elaeis guineensis var Tenera, Dura, Pisifera, Virescens dan spesies E. oleifera serta dari tiga aksesi NCBI (aksesi
U68756 - E. guineensis var Tenera, aksesi FJ940768 - E. oleifera, EU057621 - E. oleifera) yang dijadikan sebagai
lokus untuk disain primer SNPs. 30
6 DNA sekuens dari 26 pasang spesifik primer yang digunakan untuk mengamplifikasi marka single nucleotide amplified
polymorphism (SNAP) dari genomik DNA kelapa sawit. 32 7
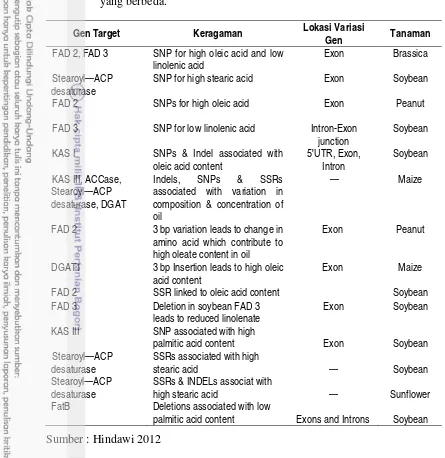
Variasi asam lemak dan gen pada pathway biosintesis TAG yang berasosiasi dengan tingginya kandungan minyak pada spesies
tanaman yang berbeda. 38
8 Interpretasi hasil analisis dua aksesi kelapa sawit spesies Elaeis guineensis Tenera 06 dan Tenera 08 dengan pasangan primer yang merepresentasikan sejumlah lokus SNPs sebagaimana yang dihasilkan
dalam elektroferogram pada Gambar 13. 42
9 Lokus marka SNAP yang dapat memprediksi kandungan asam
lemak tidak jenuh 45
10 Lokus SNPs diferential yang mampu membedakan SAD asal
E. oleifera dan asal E. guineensis dan berpotensi untuk memprediksi
kandungan asam lemak tidak jenuh kelapa sawit. 46
DAFTAR GAMBAR
1 Bagan alir penelitian 4
2 Ragam tanaman kelapa sawit. (A) Elaeis guineensis Nigrescens, (B) E. guineensis Virescens, (C) E. oleifera. (D) Buah E.guineensis var. Tenera (Nigrescens), (E) Buah E. guineensis var. Virescens, (F) Buah E.
guineensis var. Dura (Nigrescens), dan (G) Buah E. guineensis var.
Pisifera (Nigrescens). 6
xiii
3 Produk minyak kelapa sawit 7
4 Biosintesis asam lemak pada tanaman 8
5 Dua skema pathway sistesis membran glycerolipid pada daun
Arabidopsis. 9
6 Disain Primer SAD spesifik, SAD 1 terdiri atas pasangan primer Forward (F1) dan Revers 1 (R1), SAD 2 terdiri atas pasangan primer Forward (F2) dan Revers 1 (R2)
17 7 Bahan Isolasi DNA (a) Daun Pisifera dari kultur jaringan embrio,
(b) Daun tomabak Tenera berasal dari lapangan 19
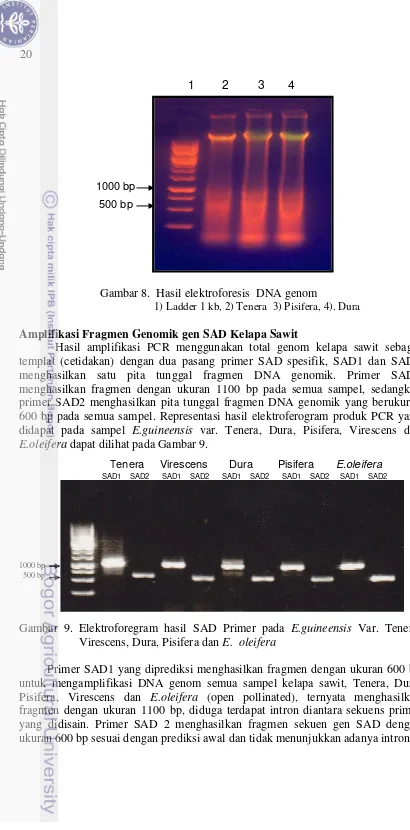
8 Hasil elektroforesis isolasi DNA 20
9
Elektoforegram hasil SAD primer pada E.guineensis Var. Tenera,
Virescens, Dura, Pisifera dan E. oleifera 20 10 Panjang sekuens yang dihasilkan oleh primer SAD1 dan SAD2.
(a) Ukuran Gen SAD aksesi U68756, (b) Hasil amplifikasi Primer
SAD 1 dan SAD 2 pada dua daerah yang berbeda. 21
11 Kandungan dan kualitas minyak nabati 25
12 Siklus biosintesis pembentukan asam lemak pada plastid tanaman 26
13 Reaksi katalisasi gen SAD 26
14 Identifikasi SNPs dari hasil pensejajaran urutan basa pada intron 29 15 Pensejajaran prediksi asam amino aksesi sampel dengan asam
amino yang terdeposit dalam bank gen NCBI E.guineensis var
Tenera aksesi U68756, E.oleifera FJ940768 dan RU57621. 32
16 Bahan isolasi genom aksesi kelapa sawit 41
17 Elektroferogram aksesi 8 dan aksesi 06 41
18
Dendogram hasil amplifikasi 26 pasang SNAP primer pada 50
aksesi kelapa sawit. 43
19 Dua lokus SNPs gen SAD yang tergolong sebagai SNPs non- synonimous spesifik untuk spesies Elaeis oleifera. 44 20 Dendogram 50 aksesi kelapa sawit dengan pada 6 locus SNPs
yang mengindikasikan biosintesis kandungan asam oleat 47
DAFTAR LAMPIRAN
1 Lokasi disain primer gen SAD spesifik (SAD1 dan SAD2) 56
xii
3 Pensejajaran sekuens sampel (exon, intron dan 3’UTRs) 61
4 Pensejajaran 5 sampel dengan aksesi rujukan NCBI 67
5 Asal bahan tanaman 50 aksesi kelapa sawit 75
6 Pendaftaran paten dan nomor paten 76
BAB I
PENDAHULUAN
Latar Belakang
Kelapa sawit (Elaeis guineensis Jacq.) merupakan tanaman penghasil minyak nabati dengan kemampuan produksi paling tinggi dibandingkan dengan sumber minyak nabati lainnya. Minyak kelapa sawit dapat dimanfaat sebagai sumber bahan pangan, bahan baku industri maupun sebagai alternatif bioenergi (Tan 2009). Produktifitas kelapa sawit per hektar per tahun sepuluh kali lebih tinggi daripada tanaman bunga matahari dan tiga kali lebih tinggi dibandingkan dengan tanaman kelapa, dengan potensi hasil bisa mencapai 18-20 ton (Cocchi et al. 2009). Indonesia sejak tahun 2007 merupakan produsen utama minyak sawit dunia, yang memproduksi lebih dari 44% dan menjadi negara konsumen kelapa sawit terbesar juga sejak tahun 2013 (Hariyadi 2010; USDA 2014).
Dua spesies tanaman kelapa sawit yang dikenal luas adalah Elaeis guineensis, yang berasal dari Afrika Barat dan Elaeis oleifera yang berasal dari Amerika Selatan (Corley dan Tinker 2003). E. guineensis telah dibudidayakan secara luas dan komersial sebagai tanaman penghasil minyak nabati, sedangkan pengembangan E. oleifera masih relatif terbatas.
Biosintesis minyak pada buah kelapa sawit tidak terlepas dari proses biosintesis asam lemak pada mesokarpa dan inti biji (kernel). Minyak yang berasal dari mesokarpa dan kernel mempunyai komposisi yang berbeda-beda dan tergantung pada proses pembentukan dan akumulasi minyak selama perkembangan buah kelapa sawit (Basri et al. 2004). Asam oleat merupakan salah satu komponen utama asam lemak tidak jenuh tunggal pada minyak sawit yang baik untuk kesehatan. Asam oleat bersifat stabil dan diketahui mampu mengurangi resiko serangan penyakit jantung. Selain itu, kandungan asam lemak tidak jenuh juga penting dalam menunjang industri oleochemical dan nutraceutical (Sundram et al. 2003; Georgios et al. 2005; Masani dan Parveez 2008). Terdapat korelasi positif antara tingginya Kandungan asam lemak tidak jenuh dengan tingginya nilai iodine pada minyak sawit, nilai iodine E. oleifera 80 sedangkan Elaeis guineensis 50 (Rajanaidu et al. 2000). Nilai Iodine adalah banyaknya Iodine (gram) yang dibutuhkan untuk menetralkan ikatan rangkap yang terdapat pada 100 gram minyak (Zulkarnain et al. 2011). Kandungan asam oleat dan nilai iodine yang tinggi akan mempengaruhi tingkat liquiditas minyak kelapa sawit, sehingga minyak sawit tidak akan mudah membeku walaupun berada pada suhu yang rendah. Karakteristik ini penting apabila minyak sawit akan dimanfaatkan sebagai sumber energi atau biofuel (Choo dan Cheah 2000).
2
mempunyai kandungan asam lemak tidak jenuh yang berkisar antara 40-60%. Sebaliknya, minyak sawit dari E. oleifera dilaporkan mempunyai kandungan asam lemah tidak jenuh yang lebih tinggi, yaitu berkisar antara 70-80% (Rajanaidu et al. 2000). E. oleifera memiliki karakter unggul yang dapat dimanfaatkan untuk perbaikan genetik spesies E. guineensis yang telah dibudidayakan secara komersial, melalui pemuliaan tanaman guna menghasilkan varietas E. guineensis baru dengan karakter unggul kandungan asam lemak tidak jenuh yang tinggi (Subhash dan Farida 2012).
Proses pembentukan asam lemak tidak jenuh berhubungan dengan aktifitas enzim stearoyl ACP (acyl carier protein) desaturase (SAD) yang merupakan gen kunci pada pembentukan asam oleat. Mesokarpa buah kelapa sawit mengandung enzim SAD yang aktif dan sebagian besar enzim SAD efektif membentuk asam oleat (Parveez 2004). Sejumlah gen kunci yang berperanan dalam pembentukan asam lemak pada tanaman secara umum, juga mempunyai fungsi yang sama dalam biosintesis asam lemak pada buah kelapa sawit (Basri et al. 2004).
Gen SAD merupakan gen penyandi enzim stearoyl ACP desaturase, yang merupakan lintasan gen awal untuk pembentukan asam lemak tidak jenuh pada tanaman. Gen SAD menyandi protein yang berfungsi dalam biosintesis asam lemak tidak jenuh (asam oleat dan asam linoleat). Karakterisasi dan purifikasi gen penting tersebut pada kelapa sawit telah dilakukan (Ravigadevi et al. 2000), namun studi yang mempelajari keragaman DNA sekuens dari gen SAD antar varietas dan spesies kelapa sawit yang mempunyai perbedaan kandungan dan komposisi asam lemak tidak jenuh belum terdokumentasi dengan baik.
Mesokarpa kelapa sawit sangat aktif dan efektif merubah hampir semua stearoyl ACP menjadi oleoyl-ACP dalam pembentukan asam oleat pada lintasan bisosintesis asam lemak pada kelapa sawit, sehingga untuk meningkatkan kandungan asam oleat pada kelapa sawit dibutuhkan peningkatan aktifitas gen SAD. Mesokarpa E.oleifera mengandung komposisi asam lemak tidak jenuh (asam oleat) lebih tinggi dibandingkan E.guineensis, mempelajari dan membandingkan aktifitas gen SAD pada kedua spesies ini akan membantu mengetahui kunci utama yang menyebabkan perbedaan komposisi asam lemak tidak jenuh pada kedua tanaman ini (Siti et al. 2001).
3 fragmen gen tertentu paling banyak digunakan karena langsung berhubungan dengan fungsi gen target yang diinginkan, bersifat stabil, dapat diulang dan mampu menghasilkan marka dengan resolusi yang bagus (Syvanen 2001).
Marka molekuler SNPs pada kelapa sawit telah dikembangkan untuk mendeteksi perbedaan sifat terkait dengan warna buah dan ketebalan cangkang. Marka SNPs yang telah dikembangkan tersebut mampu memprediksi warna buah dan ketebalan cangkang dengan tingkat akurasi yang tinggi (Ooi et al. 2011). Pada tanaman Brassica yang juga penghasil minyak nabati, perubahan gen FAD2 dan FAD3 yang berbasis SNPs dapat digunakan sebagai marka molekuler (US Pat.No.2006/0248611 A1) (Xueyi et al. 2006).
Tujuan Penelitian
Tujuan umum : Mendapatkan marka molekuler SNAP berbasis SNP yang dapat digunakan untuk memprediksi kandungan asam lemak tidak jenuh kelapa sawit.
Tujuan khusus :
(1) Mengisolasi dan mengetahui runutan nukleotida fragmen gen SAD dari sumber keragaman kelapa sawit.
(2) Mempelajari dan mengidentifikasi keragaman nukleotida gen SAD dalam rangka pengembangan Marka molekuler berbasis SNPs.
(3) Mengembangkan marka SNAP berbasis SNPs berdasarkan keragaman nukleotida gen SAD yang dapat mengindikasikan kandungan asam lemak tidak jenuh dan asam oleat khususnya pada kelapa sawit.
Hipotesis
(1) Terdapat keragaman runutan nukleotida fragmen gen SAD dari spesies kelapa sawit E. guinensis dan E. Oleifera.
(2) Marka SNAP berbasis single nucleotide polymorphisms (SNPs) dapat diperoleh berdasarkan keragaman gen SAD pada spesies kelapa sawit yang dapat mengindikasikan keragaman dan kandungan asam oleat (asam lemak tidak jenuh) pada kelapa sawit
Manfaat Penelitian
4
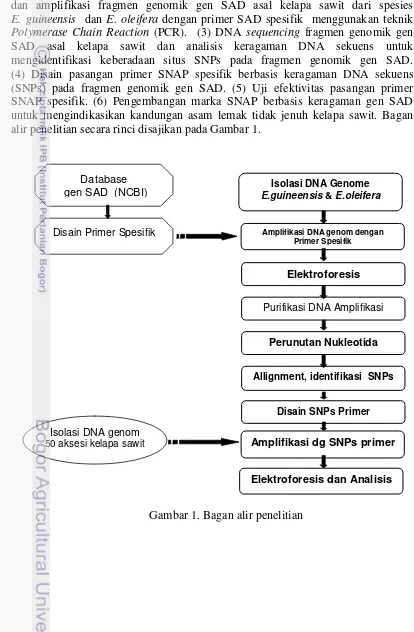
Ruang Lingkup Penelitian
Penelitian ini mempunyai ruang lingkup yang saling berkaitan menjadi satu kesatuan untuk mencapai tujuan penelitian yang melingkupi: (1) Penelusuran data aksesi DNA sekuens gen SAD asal kelapa sawit dan menggunakannya untuk mendisain SAD spesifik primer. (2) Isolasi DNA genom dari daun kelapa sawit
dan amplifikasi fragmen genomik gen SAD asal kelapa sawit dari spesies E. guineensis dan E. oleifera dengan primer SAD spesifik menggunakan teknik
Polymerase Chain Reaction (PCR). (3) DNA sequencing fragmen genomik gen SAD asal kelapa sawit dan analisis keragaman DNA sekuens untuk
mengidentifikasi keberadaan situs SNPs pada fragmen genomik gen SAD. (4) Disain pasangan primer SNAP spesifik berbasis keragaman DNA sekuens
(SNPs) pada fragmen genomik gen SAD. (5) Uji efektivitas pasangan primer SNAP spesifik. (6) Pengembangan marka SNAP berbasis keragaman gen SAD untuk mengindikasikan kandungan asam lemak tidak jenuh kelapa sawit. Bagan alir penelitian secara rinci disajikan pada Gambar 1.
Gambar 1. Bagan alir penelitian
Amplifikasi dg SNPs primer
Elektroforesis dan Analisis Amplifikasi DNA genom dengan
Primer Spesifik
Elektroforesis
Allignment, identifikasi SNPs
Disain SNPs Primer Isolasi DNA Genome E.guineensis & E.oleifera
Purifikasi DNA Amplifikasi
Perunutan Nukleotida Database
gen SAD (NCBI)
Isolasi DNA genom 50 aksesi kelapa sawit
5 BAB II
TINJAUAN PUSTAKA
Kelapa Sawit
Kelapa sawit sebagai tanaman penghasil minyak nabati mempunyai peran strategis dalam perekonomian nasional. Komoditas kelapa sawit berperan dalam menyerap tenaga kerja, sumber pendapatan dan devisa bagi negara, stimulator pertumbuhan pusat-pusat ekonomi baru di pedesaan, serta sebagai sumber pangan dan energi yang penting bagi Indonesia (Warta 2008).
Genus Elaeis terdiri atas tiga spesies yakni Elaeis guineeensis, Elaeis oleifera dan Elaeis odora yang dikenal juga dengan nama Barcella odora (Corley dan Tinker 2003). Namun, yang banyak dikenal dan dimanfaatkan adalah E.guineensis dan E.oleifera. Kelapa sawit adalah tanaman bergenom diploid yang memiliki 32 kromosom dengan 16 pasang kromosom homolog. Elaeis berasal dari kata bahasa Yunani elaion dengan arti minyak, sedangkan guineensis didasarkan kepada asalnya yakni Guinea (pantai Barat Afrika), penamaan ini diberikan oleh Jacquin (1973) ( Corley dan Tinker 2003; Lubis 1992).
Elaeis oleifera atau Elaeis melanococca berasal dari Amerika Selatan, merupakan spesies penting sebagai sumber plasma nutfah yang dapat dimanfaatkan untuk perbaikan karakter kelapa sawit komersial (E.guineensis). Kanopi E.oleifera relatif kecil dengan pertumbuhan tinggi tanaman hanya 20 cm/tahun, sedangkan E.guineensis mencapai 45 cm/tahun, sehingga dapat ditanam lebih banyak perhektarnya dan lebih mudah untuk melakukan pemanenan. Perbandingan bunga jantan dan buang betina cukup tinggi, dengan tandan bunga jantan yang sedikit sekali. Disamping itu E.oleifera memiliki kandungan asam lemak tidak jenuh yang lebih tinggi (70-83%) dari pada E.guineensis (40-60%) (Lubis 1992; Pamin 1998).
Introduksi kelapa sawit ke Indonesia dimulai pada tahun 1848 dengan penanaman 4 bibit tanaman kelapa sawit di Kebun raya Bogor. Dari keempat bibit tersebut dua bibit diintroduksi dari Bourbon atau Mauritius pada bulan Februari 1848, dan dua bibit lain diintroduksi dari Amsterdam pada bulan Maret 1948. Kemudian dari 4 bibit ini kelapa sawit menyebar menjadi tanaman komersial seperti saat ini. Awalnya kelapa sawit ditanam sebagai komoditi perkebunan dilakukan pada tahun 1911 yang dibangun oleh M.Adrien Hallet berkebangsaan Belgia dengan menanam tanaman kelapa sawit di Perkebunan Sungai Liput (Aceh) dan Pulu Raja (Asahan) (Pamin 1998).
Taksonomi tanaman kelapa sawit adalah : Divisi : Tracheophy
Subdivisi : Pteropsida Kelas : Angiospermae Subkelas : Monocotyledoneae Ordo : Cocoideae
Familia : Palmae Sub Famili : Cocoidae Genus : Elaeis
6
Gambar 2. Ragam tanaman kelapa sawit.(A) Elaeis guineensis Nigrescens, (B) E. guineensis Virescens, (C) E. oleifera. (D) Buah E.guineensis var.
Tenera (Nigrescens), (E) Buah E. guineensis var. Virescens, (F) Buah E.
guineensis var. Dura (Nigrescens), dan (G) Buah E. guineensis var. Pisifera (Nigrescens).
Kelapa sawit diklasifikasikan berdasarkan berbagai hal yang dapat dibedakan atas tipe buah, bentuk buah, tebal cangkang dan warna buah. Berdasarkan ketebalan cangkang buah ada tiga tipe tanaman kelapa sawit, yakni: tipe Dura, tipe Pisifera, dan tipe Tenera (Basri et al. 2004). Salah satu ciri E. guineensis tipe Dura adalah mempunyai cangkang biji yang tebal dan sabut atau mesokarpa yang tebal. Kelapa Sawit tipe Dura pada umumnya digunakan sebagai induk betina dalam pemuliaan kelapa sawit. Tipe Pisifera mempunyai cangkang biji tipis yang digunakan sebagai induk jantan dalam pemuliaan kelapa sawit (Latiff 2000). Kelapa sawit tipe Tenera merupakan kelapa sawit yang dibudidayakan secara komersial untuk menghasilkan minyak. Karakteristik buah Tenera mempunyai ketebalan cangkang biji sedang dan sabut yang tebal sehingga
A
B
C
D
E
7 banyak mengandung minyak (Latiff 2000; Basri et al. 2004). Kelapa sawit Tenera pada umumnya merupakan hibrida dari persilangan antara tetua betina Dura dan tetua jantan Pisifera (Hafiz dan Rashid 2011), keragaman kelapa sawit seperti terlihat pada Gambar 2.
Berdasarkan warna buah, kelapa sawit dapat dibedakan atas varietas nigrescens, virescens dan Albescens. Varietas nigrescens, buahnya berwarna violet sampai hitam waktu muda dan menjadi merah-kuning (orange) sesudah matang, dengan warna buah yang hampir sama baik yang masih muda ataupun yang sudah masak fisiologis sehingga sulit untuk membedakan perkembangan buahnya. Kelapa sawit dengan warna buah kehijauan saat muda dan buah tua berwarna kekuningan dikenal sebagai Virescens (Latiff 2000). Akibat perubahan warna tersebut memudahkan proses pemanenan tandan buah karena tandan buah yang siap panen akan mempunyai warna buah yang berbeda dengan tandan buah yang masih muda (Hafiz dan Rashid 2011). Buah albescens berwarna kuning pucat, tembus cahaya karena mengandung sedikit protein (Lubis 1992).
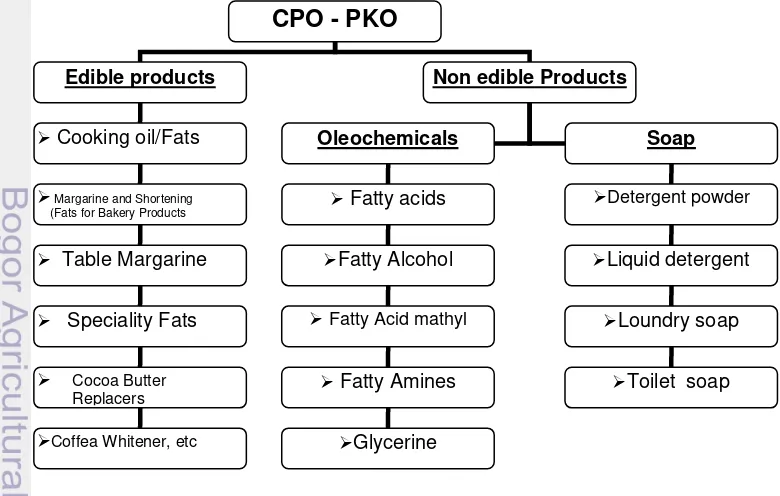
Komposisi minyak nabati secara umum terdiri atas trigliserida asam lemak yang bisa mencapai 95%, asam lemak bebas (Free Fatty Acid), monogliserida dan digliserida, serta beberapa komponen lain seperti phosphoglyserida, vitamin, mineral dan sulfur. Minyak kelapa sawit terdiri atas dua jenis yang berasal dari bagian buah yang berbeda yakni CPO (crude palm oil) yang berasal dari daging buah, dan Palm Kernel Oil (PKO) yang berasal dari inti biji kelapa sawit. Minyak sawit digunakan sebagai bahan pangan dan industri setelah melalui proses penyulingan, penjernihan, dan penghilangan bau (Refined, Bleached and Deodorized Palm Oil, RBDPO). CPO dapat diuraikan untuk memproduksi minyak sawit cair (RBD Olein) dan minyak sawit padat (RBD Stearin). RBD olein digunakan untuk minyak goreng, sedangkan RBD stearin digunakan untuk margarine dan shortening, serta sebagai bahan baku industri sabun dan deterjen. Produk minyak kelapa sawit disajikan pada Gambar 3.
Gambar 3. Produk minyak kelapa sawit
CPO - PKO
Edible products Non edible Products
Oleochemicals Soap
Detergent powder
Fatty acids Cooking oil/Fats
Margarine and Shortening (Fats for Bakery Products
Fatty Alcohol
Fatty Acid mathyl
Fatty Amines Table Margarine
Speciality Fats
Cocoa Butter Replacers
Coffea Whitener, etc
Liquid detergent
Loundry soap
Toilet soap
8
Produk turunan kelapa sawit (CPO dan PKO) dapat dibagi atas produk pangan dan non pangan. Produk pangan umumnya digunakan untuk minyak goreng, mentega dan cocoa butter, sedangkan non pangan dimanfaatkan sebagai bahan baku sabun, oleochemical dan sumber energi alternatif yang dikenal dengan biodiesel. Komitmen negara-negara di dunia untuk menggunakan biodiesel dan bioethanol sebagai sumber energi, menjadi peluang untuk perkembangan industri kelapa sawit, disamping itu diversifikasi produk dan pengembangan produk baru dengan nilai tambah diharapkan berlanjut sehingga memperkuat industri kelapa sawit (Pamin 1998; Tan 2009; Bangun 1998).
Minyak kelapa sawit relatif cepat diterima oleh pasar domestik dan dunia, dari awal sampai sekarang fokus perhatian perkembangan industri sejak tahun 1968 salah satunya adalah Indonesia dapat menjadi pemasok utama minyak nabati utama dunia. Hal ini tidak terlepas dari beragam manfaat dan kegunaaan minyak kelapa sawit, produksi yang semakin meningkat, aspek nutrisi dan harga yang kompetitif (Pamin 1998). Di pasar pangan dunia, minyak sawit bisa ditemukan sebagai komponen pada satu dari setiap 10 produk pangan yang diperdagangkan (Hariyadi 2010).
Metabolisme Asam Lemak dan Gen SAD (Stearoyl ACP Desaturase)
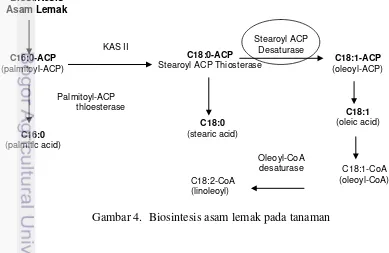
Pembentukan minyak pada kelapa sawit tidak terlepas dari metabolisme pembentukan asam lemak pada mesokarpa dan inti biji kelapa sawit. Gen-gen kunci dalam pembentukan asam lemak secara umum pada tanaman juga terdapat pada metabolisme pembentukan kelapa sawit seperti pada Gambar 4. Gen ß-ketoacyl ACP Syntetase II (KAS II) dan Stearoyl ACP Desaturase merupakan bagian dari gen-gen kunci yang penting pada biosinthesis pembentukan asam lemak pada tanaman. Gen SAD berperan dalam merubah C18:0-ACP menjadi C18:1-ACP untuk pembentukan asam oleat. Karakterisasi dan purifikasi gen tersebut pada kelapa sawit telah dilakukan (Ravigadevi et al. 2000), namun keragaman gen-gen tersebut pada varietas dan spesies kelapa sawit belum tereksplorasi dengan baik.
Gambar 4. Biosintesis asam lemak pada tanaman
C18:0-ACP
Stearoyl ACP Thiosterase
9
Kandungan dan komposisi asam lemak yang berbeda pada E.guineensis (40-60%) dan E.oleifera (70-80%) mengindikasikan adanya keragaman gen pada pembentukan asam lemak kelapa sawit diantara keragaman gen SAD (Rajanaidu et al. 2000). Keragaman ini dapat diidentifikasi dengan memanfaatkan teknologi molekular untuk mendapatkan tanaman kelapa sawit unggul.
Kandungan asam oleat tinggi baik untuk kesehatan terutama dalam mengurangi resiko penyakit jantung. Tingginya kandungan asam oleat bersamaan diindikasikan dengan tingginya nilai iodine. Kedua faktor ini akan mempengaruhi tingkat liquiditas minyak kelapa sawit, dimana minyak tidak akan mudah membeku walaupun berada pada suhu yang rendah. Hal ini penting apabila minyak kelapa sawit dimanfaatkan sebagai sumber energi, biofuel.
Gambar 5. Dua alur biosintesis asam lemak pada daun Arabidopsis
10
Pemuliaan Kelapa Sawit
Pemuliaan tanaman kelapa sawit merupakan salah satu contoh program pemuliaan yang berhasil, dimana dari 4 bibit yang di tanam di Kebun Raya Bogor tahun 1848 berkembang menjadi tanaman penghasil minyak nabati terbesar di dunia pada saat ini. Pemuliaan tanaman merupakan suatu cabang dari ilmu pertanian yang fokus pada manipulasi keturunan tanaman untuk mengembangkan jenis tanaman baru yang dapat dimanfaatkan oleh manusia (Pamin 1998)
Keberhasilan pelaksanaan pemuliaan kelapa sawit sangat bergantung pada ketersediaan variasi sumber genetik. Indonesia memiliki keragaman genetik kelapa sawit yang tidak luas dan hanya berada dalam kisaran segregasi dari bahan genetik yang sempit seperti Deli Dura dan turunan Tenera/Pisifera yang berkerabat dekat (Warta 2008). Oleh karena itu ekploitasi sumber genetik yang berbeda melalui kegiatan introduksi dan eksplorasi ke pusat-pusat keragaman genetik kelapa sawit di Afrika dan Amerika Selatan penting bagi perkembangan industri kelapa sawit. Persilangan dan seleksi pada pemuliaan kelapa sawit memakan waktu yang cukup lama, demikian pula untuk melakukan crossing guna pemanfaatan karakter yang baik dari famili liar. Dengan adanya perkembangan teknologi Marka Asisted Selection (MAS), dapat mempersingkat waktu yang dibutuhkan, perkembangan marka molekuler sangat membantu pemuliaan kelapa sawit.
Marka molekuler sebagai salah satu hasil dari perkembangan ilmu pengetahuan dan teknologi di bidang bioteknologi memberikan dampak positif untuk perkembangan pemuliaan tanaman. Teknologi marka molekuler pada tanaman berkembang sejalan dengan makin banyaknya pilihan marka DNA yaitu: 1) marka yang berdasarkan pada hibridisasi DNA seperti restriction fragment length polymorphism (RFLP), 2) marka yang berdasarkan pada reaksi rantai polimerase yaitu polymerase chain reaction (PCR) dengan menggunakan sekuens-sekuens nukleotida sebagai primer, seperti randomly amplified polymorphic DNA (RAPD) dan amplified fragment lengthpolymorphism (AFLP), 3) marka yang berdasarkan pada PCR dengan menggunakanprimer yang menggabungkan sekuens komplementer spesifik dalam DNA target, seperti sekuens tagged sites (STS), sekuens characterized amplified regions (SCARs), simple sekuens repeats (SSRs) atau mikrosatelit, dan single nucleotide polymorphisms (SNPs) (Azrai 2006).
Pemanfaatan marka molekuler juga telah diterapkan pada tanaman kelapa sawit, akan tetapi untuk deteksi dini kualitas minyak kelapa sawit belum tereksplorasi dengan baik. Marka molekuler dapat dikembangkan berbasis keragaman DNA sekuens yang ada dan dapat dimanfaatkan untuk membantu percepatan pengembangan varietas tanaman kelapa sawit unggul di masa yang akan datang, guna menghemat waktu, tenaga dan biaya dalam pemuliaan kelapa sawit. Pengembangan marka molekuler berbasis SNPs diharapkan dapat dilakukan untuk meningkatkan kualitas minyak kelapa sawit, guna memenuhi beragam kebutuhan terhadap beragam komposisi kandungan asam lemak dalam industri yang menggunakan minyak kelapa sawit.
Marka SNAP berbasis SNP
11 berfungsi sebagai pembeda pada populasi (Ruangchai et al. 2011). Marka SNAP dikembangkan berdasarkan keberadaan lokus SNP (single nucleotide polymorphism). SNP merupakan perbedaan basa tunggal diantara fragmen DNA yang sama yang disebabkan oleh perbedaan basa pada lokus yang sama maupun karena terjadinya indel (insertion and deletion) sebagai akibat dari keragaman DNA sekuens pada genom tanaman. Satu fragmen DNA yang identik, tetapi berasal dari individu yang berbeda, seringkali juga mempunyai keragaman dalam DNA sekuensnya. Evaluasi marka SNAP secara tidak langsung dapat membantu pemulia tanaman guna menseleksi galur-galur hasil pemuliaan tanaman dengan daya hasil tinggi atau yang mempunyai keunggulan tertentu.
12
BAB III
ISOLASI FRAGMEN GEN STEAROYL ACP DESATURASE
(SAD) ASAL 5 SUMBER KERAGAMAN KELAPA SAWIT
Abstrak
Kelapa sawit merupakan tanaman penghasil minyak nabati dengan produktifitas per hektar tertinggi dibandingkan tanaman penghasil minyak lainnya. Asam oleat merupakan salah satu komponen asam lemak tidak jenuh yang baik untuk kesehatan dan terdapat pada tanaman penghasil minyak nabati dan mempengaruhi kualitas minyak. Kandungan asam oleat (asam lemak tidak jenuh) pada kelapa sawit komersil (Elaeis guineensis) lebih rendah dibandingkan minyak yangdihasilkan Elaeis Oleifera, yang merupakan spesies kelapa sawit yang belum termanfaatkan secara optimal. Pembentukan asam oleat dipengaruhi oleh gen kunci stearoyl acp desaturase, diduga perbedaan komposisi asam oleat pada E. guineensis dan E.oleifera disebabkan oleh perpedaan basa penyusun gen stearoyl acp desaturase. Pada penelitian ini dilakukan isolasi fragmen gen SAD menggunakan dua primer spesisfik SAD (SAD1 dan SAD2) dari 5 sumber keragaman kelapa sawit yang berasal dari dua spesies kelapa sawit, E.guineensis var Dura, Pisifera, Tenera, Virescens dan spesies E.oleifera (open pollinated), untuk melihat perbedaan basa penyusun gen stearoyl acp desaturase. Hasil sequense pada fragmen gen SAD menunjukkan kemiripan yang tinggi (99%) antar kelima sampel dan dengan sequense SAD aksesi E.guineensis U68756, AF 507965 dan aksesi E.oleifera FJ940768, EU057621 yang terdeposit pada NCBI. Primer SAD1 yang menghasilkan fragmen dengan ukuran 1100 bp dan primer SAD2 yang menghasilkan fragmen gen SAD dengan ukuran 600 bp pada kelima sampel yang diisolasi.
13
STEAROYL ACP DESATURASE (SAD) GENE FRAGMENT
ISOLATION FROM 5 DIFFERSITY SOURCE OF OIL PALM
Abstract
Oil palm is a vegetable oil crops with the highest productivity per hectare than other oil crops. Oleic acid is a component of the unsaturated fatty acids which have good for human health (lowering blood LDL, Low Density Lipoprotein) and influense to the quality of the vegetable oil. The composition of oleic acid content (unsaturated fatty acids) in commercial oil palm (Elaeis guineensis) is lower that other oil palm spesies (Elaeis oleifera), which is not utilize optimally yet. Oleic acid formation on vegetable oil influence by the key enzym stearoyl acp desaturase (SAD). The oleic acid composition different between E. guineensis and E.oleifera may caused by the difference bases constituent stearoyl acp desaturase of both spesies. This research conduvted to isolate of SAD gene fragments was isolate using two spesifik SAD primers (SAD1 and SAD2) from 5 sources of oil palm diversity derived from two species of oil palm, E.guineensis var Dura, Pisifera, Tenera, virescens and species E.oleifera (open-pollinated). The SAD fragment sequense showed high similarity (99%) to each others sample source and oil palm SAD sequense acsesion E.guineensis U68756, AF 507965 and E.oleifera acsesion FJ940768, EU057621 of NCBI. The SAD 1 produce SAD sequense fragments 1100 bp and the SAD2 primer 600 bp in all five isolated samples.
14
Pendahuluan
Kualitas Minyak Kelapa Sawit
Kebutuhan terhadap kualitas minyak nabati yang sehat dengan harga yang murah menjadi tren pada perkembangan industri minyak dan asam lemak secara global. Kelapa sawit sebagai salah satu tanaman penghasil minyak utama dengan produktifitas per hektar tertinggi dibandingkan semua tanaman penghasil minyak lainnya, diharapkan dapat memenuhi kebutuhan tersebut. Oleh karena itu perlu dikembangkan materi genetik kelapa sawit dengan hasil tinggi dan kualitas minyak yang baik guna memenuhi kriteria sebagai bahan pangan maupun non pangan. Salah satu cara mempercepat mendapatkan tanaman unggul kelapa sawit, yang memenuhi kriteria diinginkan adalah dengan mengembangkan marka molekuler yang dapat mengidentifikasi kualitas minyak yang dihasilkan.
[image:31.595.82.480.511.720.2]Mesokarpa buah kelapa sawit merupakan sumber penghasil minyak terbesar pada tanaman kelapa sawit. Perbaikan kualitas minyak yang dihasilkan mesokarpa penting dilakukan untuk perkembangan industri kelapa sawit pada masa sekarang dan yang akan datang. Kualitas minyak berkaitan erat dengan biosintesis pembentukan asam lemak dan perbandingan komposisi asam lemak jenuh dan asam lemak tidak jenuh yang dihasilkan tanaman, kualitas minyak berkorelasi dengan kandungan asam lemak tidak jenuh, semakin tinggi kandungan asam lemak tidak jenuh (asam oleat) kualitas minyak juga semakin tinggi. Lintasan pembentukan asam lemak tidak jenuh pada kelapa sawit diawali oleh aktifitas gen SAD (Gambar 4). Ini merupakan gen kunci yang mempengaruhi pembentukan asam lemak tidak jenuh pada kelapa sawit, baik asam oleat yang merupakan monounsaturated fatty acid (MUFA) maupun asam linoleat dan linolenat (PUFA). Tabel 1. Komposisi asam lemak jenuh dan asam lemak tidak jenuh pada minyak
sawit spesies Elaeis guineensis dan sejumlah aksesi plasma nutfah E. oleifera.
Spesies Asal
Asam lemak jenuh (%) Asam lemak tidak jenuh (%)
C16.0 a.palmitat
C18.0
a.stearat Total
C18.1 a.oleat
C18.2
a.linoleat Total
E.oleifera
Honduras 18.6 0.2 18.8 56.5 23.3 79.8
Costa
Rica 17 0.5 17.5 56.7 23.5 80.2
Panama 17.8 0.3 18.1 59.6 21 80.6
Colombia 15.8 0.6 16.4 61.5 20 81.5
Brazil 26.1 0.9 27 51.6 20.6 72.2
Suriname 25 0.4 25.4 70.5 3.6 74.1
E.guineensis 44 4.5 48.5 39 10.1 49.1
15
Kandungan dan komposisi asam lemak yang berbeda pada E.guineensis dan E.oleifera (Tabel 1) mengindikasikan adanya perbedaan gen penyandi enzim yang berperanan dalam lintasan biosintesis pada pembentukan asam lemak pada genom kedua tanaman tersebut. Perbedaan ini dapat diidentifikasi dengan mengevaluasi keragaman DNA sekuens dari masing-masing gen terkait menggunakan teknologi molekuler.
Tujuan Penelitian
Penelitian ini bertujuan untuk mendapatkan primer gen SAD spesifik dan fragmen gen SAD yang berasal dari 5 sumber keragaman genom kelapa sawit spesies E.guineensis dan E.oleifera.
Bahan dan Metode
Waktu dan Tempat
Penelitian dilaksanakan pada bulan November 2012 sampai Mei 2013, di Laboratorium Teknologi Gen dan Kultur Jaringan Balai Pengkajian Bioteknologi BPPT Serpong, Tangerang, Banten.
Bahan dan Alat
Bahan tanaman yang digunakan adalah daun kelapa sawit Elaeis guinensis, Tenera, Dura, Pisifera, Virescens dan Elaeis oleifera. Bahan lain yang digunakan meliputi liquid nitrogen, buffer CTAB (cetyltrimetilammonium bromide) yang terdiri atas CTAB 2 %, 100 mM Tris (pH8), 20mM EDTA (pH8), 1.4 M NaCl, dan dH2O, kloroform - isoamil alkohol (24 : 1), agarose, etanol absolut, 5x
phusion HF buffer, 10 mM dNTP, DMSO, Phusion Hot start II DNA Polymerase. DNA fragments Extraction kit (Genaid). Loading dye, Gelred, ladder 1 kb, pasangan Primer SAD, SAD1 dan SAD2 forward dan revers 10µL.
Alat yang digunakan meliputi mesin PCR, Elektroforesis tank, Laminar Air Flow Cabinet, sentrifus, Water bath, mesin nanodrop, timbangan analitik, oven, mikrowave, pinset, bunsen, petri, mikropipet, shaker, tabung eppendorf, erlenmeyer, ice keeper, mortar, spatula, Tabung Liquid Nitrogen dan pipet tips. Alat gelas yang digunakan labu erlenmeyer 250 ml, tabung penyimpan bahan 200 mL, dan cawan petri.
Pelaksanaan Penelitian Persiapan
16
diintroduksikan oleh PT. AAL. Bahan kimia untuk isolasi DNA, PCR, dan elekstroforesis disiapkan di laboratorium Teknologi Gen.
Isolasi DNA Genom
Isolasi DNA daun dilakukan berdasarkan metode CTAB Doyle and Doyle (1990) yang dimodifikasi. Sampel daun (0.5 – 1 gr) digerus dengan bantuan nitrogen cair, kemudian dimasukkan ke dalam tabung eppendorf dan ditambahkan 700 µl bufer ekstraksi. Campuran tersebut divorteks lalu diinkubasi dalam water bath selama 30 menit pada suhu 65oC. Setelah dibiarkan dingin hingga suhu kamar, campuran tersebut ditambahkan dengan 700 µL kloroform - isoamil alkohol (CIA) (24:1). Campuran disentrifugasi selama 10 menit dengan kecepatan 12.000 rpm pada suhu 4oC, supernatan dimasukkan ke dalam tabung eppendorf baru. Prosedur penambahan CIA diulangi sampai dua kali dengan tujuan untuk mendapatkan hasil isolasi yang murni. Pelet DNA dicuci dengan menambahkan etanol absolut dua kali volume sampel, kemudian disentrifugasi 10 menit dengan 12.000 rpm pada suhu 40C, kedalam pelet ditambahkan etanol 70% dan disentrifus 12.000 rpm pada suhu 40C selama 5 menit, supernatan dibuang kemudian pellet dikeringkan. Endapan DNA yang telah kering dilarutkan dalam 50 µl bufer TE (10 mM Tris-HCl pH 8,0; 1 mM EDTA pH 8,0). Pengecekan kualitas dan kuantitas DNA genom dilakukan dengan elektroforesis (1% gel agarose) dan nanodrop. Disain Primer Spesifik
Disain primer spesifik dilakukan dengan mengakses data sekuenss gen Stearoyl ACP Desaturase dari aksesi tanaman, Elaeis oleifera, Elaeis guineensis, yang terdeposit di NCBIyang terdiri atas U68756 (E.guineensis var Tenera) yang merupakan fragmen gen SAD kelapa sawit terpanjang yang terdapat di NCBI, FJ940768 (E.oleifera), EU057621 (E.oleifera, klon). Daerah yang sama diidentifikasi menggunakan Clustalw Bioedit, sedangkan primer dirancang menggunakan software primer3plus.
Amplifikasi DNA Genom dengan Primer Spesifik
Sampel DNA yang akan digunakan sebagai template PCR dicek konsentrasi dan kemurniannya terlebih dahulu dengan nanodrop, kemurnian DNA yang digunakan pada panjang gelompang 260/280 berkisar antara 1,8-2,0. Total volume mixed PCR adalah 20µL yang terdiri atas : 4 µL 5x Phusion HF buffer , 0.4 µL 10 µM dNTPs, masing-masing 1 µL Primer Forward dan Revers 10 µM, 0.2 µL Phusion Hot Start II DNA polymerase, 100 ng template DNA dan sisanya dH20. Reaksi PCR
dilakukan dengan 30 siklus pada suhu 98ºC selama 30 detik, 98ºC selama 5 detik, 52ºC selama 30 detik, 72ºC selama 15 detik, dan 72ºC selama 5 menit.
Elektroforesis
17 PCR dinyatidakan positif apabila terlihat adanya produk yang spesifik sesuai ukuran primer.
Perunutan nukleotida dan analisis sekuen produk PCR
Produk PCR (single band) dipurifikasi menggunakan PCR DNA fragments Extraction kit (Genaid kit). Hasil purifikasi digunakan sebagai bahan untuk perunutan nukleotida menggunakan mesin DNA sequenser otomatis AB 3130 Genetic Analyzer DNA. Hasil runutan DNA dianalisis menggunakan perangkat lunak BLAST 2.0, yang tersedia secara online di situs NCBI (http://blast.ncbi.nlm.nih.gov/ Blast.cgi) untuk mengetahui identitas fragmen genomic DNA yang didapatkan serta tingkat kemiripannya dengan gen SAD tanaman lain.
Hasil dan Pembahasan
Disain Primer SAD Spesifik
Disain primer dilakukan untuk membuat untaian basa nukleotida yang akan menempel dan mengapit daerah untaian DNA tertentu dari total genom yang ada pada kromosom. Pengapitan daerah untaian DNA diperlukan untuk melakukan perbanyakan (amplifikasi) hanya pada daerah untaian DNA tertentu sehingga lebih mudah dalam melakukan analisis. Keberhasilan melakukan amplifikasi daerah untai DNA sangat dipengaruhi oleh kecocokan primer terhadap kondisi DNA target.
Disain primer gen SAD spesifik dilakukan dengan mengakses data sekuens gen SAD kedua spesies kelapa sawit dari NCBI aksesi U68756 (E.guineensis var Tenera), FJ940768 (E.oleifera), EU057621 (E.oleifera). Daerah yang sama diidentifikasi dengan Clustalw Bioedit, sedangkan primer dirancang dengan software primer3plus untuk dua daerah dengan urutan basa yang berbeda (Gambar 6) sehingga dihasilkan dua pasang primer, SAD 1 dan SAD 2 seperti pada Tabel 2, lokasi primer berdasarkan aksesi U68756 terdapat pada Lampiran 1.
Gambar 6. Disain Primer SAD spesifik, SAD 1 terdiri atas pasangan primer Forward (F1) dan Revers 1 (R1), SAD 2 terdiri atas pasangan primer Forward (F2) dan Revers 1 (R2)
Optimasi suhu annealing dilakukan untuk mendapatkan suhu optimal guna mengamplifikasi gen SAD dari genom kelapa sawit. Dari hasil optimasi diperoleh, suhu optimal untuk primer SAD1 guna mengamplifikasi genom kelapa sawit adalah 52oC, sedangkan suhu optimal untuk primer SAD2 adalah 50oC. Asmini et al. (2010) menambahkan selain suhu annealing (penempelan) dan konsentrasi ion Mg, spesifisitas primer merupakan salah satu faktor penting dalam amplifikasi suatu fragmen DNA.
Gen SAD : Ukuran 1715 bp pada aksesi U68756
18
Tabel 2. Dua pasang primer SAD spesifik untuk mengamplifikasi genomik fragmen gen SAD.
Primer Forward Reverse Suhu Annealing
(oC)
SAD1 ATGCTCAACACCCTTGATGG GGCATAGTCCTTGGCTGTGT 52
SAD2 CCTGCCCATCTGATGTATGA CGAGCAAGAACTAGGTTCCA 50
Primer SAD didisain untuk mendapatkan spesifisitas primer untuk gen SAD kelapa sawit, dimana gen SAD merupakan gen kunci pada lintasan pembentukan asam lemak tidak jenuh pada kelapa sawit, karena merupakan gen awal yang terbentuk ke arah lintasan pembentuk asam lemak tidak jenuh (diawali dengan pembentukan asam oleat oleh gen SAD). Biosintesis asam lemak kelapa sawit dikendalikan oleh gen-gen kunci penyandi enzim yang berperanan dalam pembentukan asam lemak pada tanaman secara umum. Biosintesis asam lemak menghasilkam asam palmitat (palmitic acid) dan asam stearat (stearic acid) yang merupakan produk asam lemak jenuh, sedangkan asam oleat (oleic acid) dan asam lonoleat serta asam linolenat merupakan produk asam lemak tidak jenuh. Proses pembentukan asam lemak tidak jenuh berhubungan dengan aktifitas gen SAD pada buah kelapa sawit, ini merupakan gen kunci yang berperanan penting dalam proses biosintesis asam lemak tidak jenuh (Basri et al. 2004).
Isolasi DNA Genom dari Daun Kelapa Sawit
DNA genom yang diisolasi berasal dari tanaman E. guineensis var. Tenera, Dura, Pisifera (Nigrescens), dan E. guineensis var. Tenera (Virescens) serta E. Oleifera untuk mendapatkan informasi sekuens gen SAD. Bahan isolasi DNA yang digunakan dapat digolongkan atas dua yakni dari daun muda yang berasal lapangan dan dari daun serta tahap multiplikasi embrio asal kultur jaringan (Gambar 7). Tenera berasal dari daun muda tanaman di lapangan, sedangkan untuk tanaman Dura, Pisifera, Virescens dan E.oleifera berasal dari daun kultur jaringan embrio.
19
Gambar 7. Bahan Isolasi DNA, a). Daun hasil kultur jaringan, b) daun tombak dari lapangan.
Isolasi DNA menunjukan hasil yang baik, dilihat dari hasil uji kualitatif dan kuantitatif yang telah dilakukan. Hasil isolasi DNA secara kualitatif memperlihatkan intensitas pita hasil elektroforesis yang tampak jelas (Gambar 8). Adanya pita pada bagian awal proses migrasi menunjukkan bahwa DNA yang dihasilkan adalah DNA total. Hasil kuantitatif terdapat pada Tabel 3 dimana konsentrasi DNA yang didapat memadai dengan kemurnian bervariasi berkisar dari 1.8 – 2.0 kecuali virescens dengan kemurnian 1.72, namun ini masih dapat ditolerir karena menghasilkan produk amplifikasi yang bagus (Gambar 9).
Tabel 3. Hasil isolasi 5 sampel sumber keragaman kelapa sawit NO Sumber Keragaman ng/µL 260/280
1 Tenera 932 1.94
2 Dura 2443 1.86
3 Pisifera 130 1.83
4 Virescens 172 1.72
5 E.oleifera 658 2.06
DNA yang diisolasi dari tanaman seringkali terkontaminasi oleh polisakarida dan metabolit sekunder seperti tanin, pigmen, alkaloid dan flavonoid (Safaruddin et al. 2011). Pengukuran konsentrasi DNA dengan spektrofotometer dilakukan pada panjang gelombang 260nm, sedangkan protein diukur pada panjang gelombang 280. Kemurnian larutan DNA dihitung melalui perbandingan A260 nm dengan A280
nm. Batas kemurnian yang biasa dipakai dalam analisis molekuler pada rasio A260/A280 adalah 1,8-2,0 (Sambrook et al. 1989).
20
Gambar 8. Hasil elektroforesis DNA genom
1) Ladder 1 kb, 2) Tenera 3) Pisifera, 4). Dura Amplifikasi Fragmen Genomik gen SAD Kelapa Sawit
Hasil amplifikasi PCR menggunakan total genom kelapa sawit sebagai templat (cetidakan) dengan dua pasang primer SAD spesifik, SAD1 dan SAD2 menghasilkan satu pita tunggal fragmen DNA genomik. Primer SAD1 menghasilkan fragmen dengan ukuran 1100 bp pada semua sampel, sedangkan primer SAD2 menghasilkan pita tunggal fragmen DNA genomik yang berukuran 600 bp pada semua sampel. Representasi hasil elektroferogram produk PCR yang didapat pada sampel E.guineensis var. Tenera, Dura, Pisifera, Virescens dan E.oleifera dapat dilihat pada Gambar 9.
[image:37.595.66.476.11.841.2]1000 bp 500 bp
Gambar 9. Elektroforegram hasil SAD Primer pada E.guineensis Var. Tenera, Virescens, Dura, Pisifera dan E. oleifera
Primer SAD1 yang diprediksi menghasilkan fragmen dengan ukuran 600 bp untuk mengamplifikasi DNA genom semua sampel kelapa sawit, Tenera, Dura, Pisifera, Virescens dan E.oleifera (open pollinated), ternyata menghasilkan fragmen dengan ukuran 1100 bp, diduga terdapat intron diantara sekuens primer yang didisain. Primer SAD 2 menghasilkan fragmen sekuen gen SAD dengan ukuran 600 bp sesuai dengan prediksi awal dan tidak menunjukkan adanya intron.
1 2 3 4
1000 bp
500 bp
SAD1 SAD2 SAD1 SAD2 SAD1 SAD2 SAD1 SAD2 SAD1 SAD2
21
Gambar 9. Panjang sekuens yang dihasilkan oleh primer SAD 1 dan SAD 2. (a) Ukuran Gen SAD aksesi U68756, (b) Hasil amplifikasi Primer SAD 1 dan SAD 2 pada dua daerah yang berbeda.
Identifikasi Sequense SAD
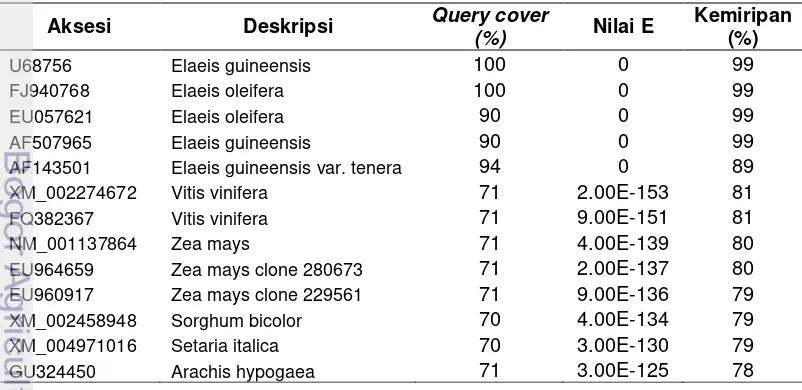
[image:38.595.108.509.500.695.2]Hasil analisis BLAST runutan DNA fragmen gen SAD dari 5 sampel keragaman kelapa sawit (Lampiran 2) yang berhasil diisolasi memiliki tingkat kemiripan 99% dengan sequense SAD kelapa sawit E.guineensis aksesi U68576 dan AF507965 serta E.oleifera aksesi AF940768 dan EU057621 (Tabel 4). Kemiripan dengan Vitis vinifera 80%, sedangkan dengan tanaman jagung (Zea mays) tingkat kemiripannya 79-80%. Hal ini mengindikasikan bahwa fragmen genomik DNA yang dihasilkan dalam penelitian ini adalah benar sebagai fragmen DNA genomik dari gen SAD asal kelapa sawit.
Tabel 4. Contoh tingkat kemiripan sequense sampel (Tenera) dengan informasi sequense SAD yang terdeposit di Bank gen, NCBI
Aksesi Deskripsi Query cover
(%) Nilai E
Kemiripan (%)
U68756 Elaeis guineensis 100 0 99
FJ940768 Elaeis oleifera 100 0 99
EU057621 Elaeis oleifera 90 0 99
AF507965 Elaeis guineensis 90 0 99
AF143501 Elaeis guineensis var. tenera 94 0 89
XM_002274672 Vitis vinifera 71 2.00E-153 81
FQ382367 Vitis vinifera 71 9.00E-151 81
NM_001137864 Zea mays 71 4.00E-139 80
EU964659 Zea mays clone 280673 71 2.00E-137 80
EU960917 Zea mays clone 229561 71 9.00E-136 79
XM_002458948 Sorghum bicolor 70 4.00E-134 79
XM_004971016 Setaria italica 70 3.00E-130 79
GU324450 Arachis hypogaea 71 3.00E-125 78
F1 F2 R1 R2
(A) Gen SAD : Ukuran 1715 bp pada aksesi U68756
(B) Prediksi produk PCR dengan primer spesifik SAD:
I ntron(507 bp)
SAD 1 (1100 bp)
22
Simpulan
23
BAB IV
IDENTIFIKASI DAN ANALISIS SNP BERDASARKAN
KERAGAMAN NUKLEOTIDA GEN SAD KELAPA SAWIT
Abstrak
Gen Stearoyl ACP Desaturase (SAD) merupakan gen kunci yang mempengaruhi pembentukan asam oleat pada minyak kelapa sawit. Elaeis guineensis dan Elaeis oleifera adalah dua spesies kelapa sawit yang memiliki komposisi kandungan asam oleat (asam lemak tidak jenuh) yang berbeda, komposisi asam oleat E.oleifera lebih tinggi dibandingkan E.guineensis. Penelitian ini dilakukan untuk mengidentifikasi dan menganalisis perbedaan susunan sekuens gen SAD kelapa sawit dan mendisain SNP marka berdasarkan lokus SNP yang teridentifikasi. Analisa nilai Tajima lima sumber keragaman gen SAD kelapa sawit adalah -1.283062, nilai negatif Tajima menandakan keragaman sequense gen SAD kelapa sawit rendah. Hasil identifikasi SNP pada fragmen genome gen SAD terdapat 13 lokus SNP yang digunakan untuk mendisain 26 pasang primer oligonukleotida untuk mengembangkan 13 lokus marka SNAP.
24
SNP IDENTIFICATION AND ANALYSIS OF OIL PALM SAD
(Stearoyl ACP Desaturase) GENE DIFFERSITY
Abstrak
The Stearoyl ACP desaturase (SAD) is a key enzyme that inffluence to oleic acid formation in palm oil. Oil palm have two spesies Elaeis guineensis and Elaeis oleifera that have different oleic acid (unsaturated fatty acids) composition. This experiments conducted to indentify the loci difference of bases constituent stearoyl acp desaturase and design of SNP markers based on SNP loci identified. The Tajima value of SAD of five sources of oil palm differcity is -1.283062, Tajima negative value indicated that the diversity of SAD sequense is narrow in oil palm. There were 13 SNP loci identify from oil palm SAD fragments genome and use to design 26 oligonucleotide primer pairs to develop the 13 loci markers SNAP.
25 Pendahuluan
Lipid atau lemak merupakan suatu senyawa biomolekul yang larut dalam pelarut organik seperti eter, kloroform dan benzen, tetapi tidak larut dalam air. Minyak merupakan salah satu sumber lipid/lemak alami yang terdapat pada tanaman. Tanaman sebagai sumber penghasil lemak yang signifikan karena banyak spesies tanaman menyimpan asam lemak dalam bentuk triasilgliserol pada biji (Jay et al. 2002). Kelapa sawit merupakan salah satu tanaman penyimpan dan penghasil asam lemak yang paling tinggi dibandingkan tanaman penghasil minyak lainnya.
Kandungan asam oleat yang tinggi pada minyak konsumsi, baik untuk kesehatan karena dapat menurunkan kadar kolesterol LDL (Low Density Lipoprotein) dalam darah, ini dapat membantu mengurangi resiko penyakit jantung (Gambar 11). Tingginya kandungan asam oleat (asam lemak tidak jenuh) mengindikasikan tingginya nilai iodine pada minyak. Kedua faktor ini akan mempengaruhi tingkat liquiditas minyak kelapa sawit, minyak tidak akan mudah membeku walaupun berada pada suhu yang rendah. Hal ini penting apabila minyak kelapa sawit dimanfaatkan sebagai sumber energi, biofuel (Choo and Cheah 2000).
Gambar 11. Kandungan dan kualitas minyak nabati Sumber : Alberto HSHO Nutrisun
26
Gambar 12. Siklus biosintesis pembentukan asam lemak pada plastid tanaman (Umi et al. 2002)
Gen SAD pada Lintasan Pembentukan Asam Lemak. Stearoyl-ACP
Desaturase
18:0-ACP 18 :1-ACP (Strearoyl-ACP) + O2 (Oleoyl-ACP) + ferredoxin
Gambar 13. Reaksi katalisasi gen SAD Sumber : Deborah et al. 1992
Gen SAD (∆9 18:0ACP desaturase) terdapat pada jaringan daun dan biji tanaman. Gen SAD berperan dalam pembentukan asam oleat dengan menambahkan satu ikatan rangkap dari 18:0 ACP menjadi 18:1-ACP. Semua enzim acyl-ACP desaturase menggunakan ferredoxin (Gambar 13) sebagai kofactor elektron donor, yang dilakukan dalam bentuk fotosintesis (David et al. 2000). Penentuan lokus genomik gen sintesis asam lemak pada kelapa sawit akan membantu memahami regulasi biosintesis asam lemak tanaman (Katayoon et al. 2001). Hal ini dapat dilakukan dengan memanfaatkan teknologi marka molekuler SNPs.
SNPs (Single Nucleotide Polymorphisms)
27 SNPs (Single Nucleotide Polymorpism) pada warna buah kelapa sawit (Ooi et al. 2011)
SNPs adalah pasangan basa tunggal pada genom yang berbeda antara satu individu dengan individu yang lainnya. SNPs adalah variasi DNA (deoxsi ribonucleic acid) yang terjadi ketika pada suatu lokasi yang bersesuaian hanya satu nukleotid yaitu A (adenin), C (sitosin), G (guanin), T (timin) dalam genom yang berbeda. Misalnya, urutan pasangan basa TAGATA dan TGGATA berbeda pada nukleotide kedua (Setiawan, 2008)
Tujuan
Penelitian ini bertujuan untuk mengidentifikasi SNPs pada sekuens gen SAD dari 5 sumber keragaman kelapa sawit. Hasil identifikasi digunakan sebagai dasar untuk mendisain SNPs primer.
Bahan dan Metode
Waktu dan Tempat
Penelitian dilaksanakan pada bulan April 2013 sampai Juni 2013, di Laboratorium Teknologi Gen Balai Pengkajian Bioteknologi BPPT Serpong, Tangerang, Banten.
Bahan dan Alat
Penelitian ini menggunakan data sekuen SAD yang dihasilkan dari percobaan sebelumnya. Data sekuen SAD yang digunakan dalam penelitian ini adalah seguense SAD dari spesies E.guineesis dan E.oleifera. Sekuens SAD spesies E. guineesis berasal dari varietas : 1. Dura, 2. Pisifera, 3. Tenera, 4. Virescens dan 5. spesies E.oleifera
Bahan lain yang digunakan adalah agarose, etanol absolut, 5x phusion HF buffer, 10 mM dNTP, DMSO, Phusion Hot start II DNA Polymerase. DNA fragments Extraction kit (Genaid). Loading dye, Gelred, ladder 1 kb, pasangan primer SAD, SAD1 dan SAD2 10µM
Alat yang digunakan meliputi mesin PCR, mesin DNA sekuensr, Elektroforesis tank, sentrifus, Water bath, mesin nanodrop, oven, mikrowave, pinset, bunsen, petri, mikropipet, shaker, tabung eppendorf, erlenmeyer, ice keeper, dan pipet tips. Alat gelas yang digunakan labu erlenmeyer 250 ml, tabung penyimpan bahan 200 mL.
Allignment dan Identifikasi Posisi SNPs
28
Disain SNPs Primer
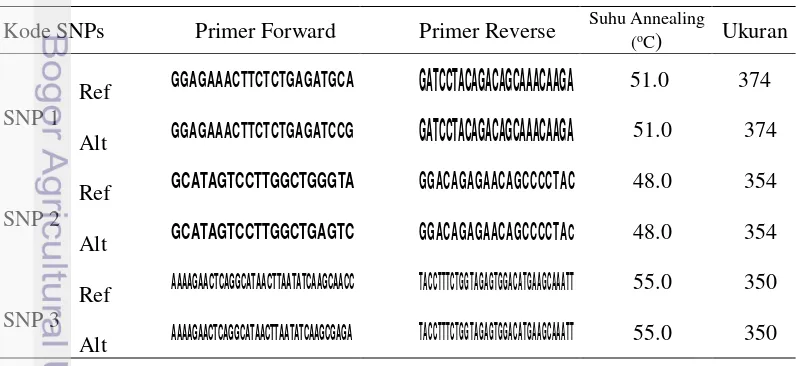
Primer spesifik yang berhubungan dengan bagian SNPs yang sudah teridentifikasi, didisain menggunakan Web SNAPER http://ausubellab.mgh. harvard.edu/ program SNAP (Single Nucleotide Amplified Polymorphisms).
Amplifikasi Primer SNPs pada genom asal disain primer
Primer SNPs digunakan pada genom awal yang jadi bahan untuk mendisain SNPs primer yang terdiri atas 5 sumber keragaman kelapa sawit yakni spesies E.oleifera, dan spesies E.guinenesis var dura, pisifera, tenera, dan virescence dengan PCR. Total volume mixed PCR adalah 20µL yang terdiri atas : 4 µL 5x Phusion HF buffer, 0.4 µL 10 µM dNTPs, masing-masing 1 µL Primer Forward dan Revers 10 µM, 0.2 µL Phusion Hot Start II DNA polymerase, 100 ng template DNA dan sisanya dH20. Reaksi PCR dilakukan dengan 30 siklus pada suhu 98ºC
selama 30 detik, 98ºC selama 5 detik, 52ºC selama 30 detik, 72ºC selama 15 detik, dan 72ºC selama 5 menit.
Hasil dan Pembahasan
Analisis Urutan Basa Gen Stearoyl ACP Desaturase (SAD)
Berdasarkan hasil runutan DNA, fragmen genomik DNA yang diamplifikasi menggunakan pasangan primer SAD1 berukuran 1,100 bp. Hasil analisis juga menunjukkan keberadaan intron dengan panjang 507 bp (Gambar 14) pada fragmen DNA genomik yang diamplifikasi. Hasil amplifikasi dengan pasangan primer SAD2 menghasilkan fragmen sekuens SAD dengan ukuran 600 bp yang terdiri atas exon dan 3’UTRs (3-Untranslated Regions). Analisis runutan DNA hasil amplifikasi kedua pasang primer SAD1 dan SAD2 menunjukkan dua bagian ekson dengan total ukuran antara 969 - 1032 bp. Satu ekson pada posisi upstream dari intron, teridentifikasi mempunyai ukuran 148 bp, sedangkan satu ekson yang berada downstream dari intron, teridentifikasi mempunyai ukuran antara 821 - 884 bp. Bagian 3 UTRs yang teridentifikasi mempunyai ukuran 282 - 590 bp (Lampiran 2). Pensejajaran hasil sekuens fragmen genom 5 sampel sumber keragaman kelapa sawit bagian exon, intron dan 3 UTRs terdapat pada (Lampiran 3). Analisa nilai tajima menghasilkan nilai -1.283062, nilai negatif pada analisa ini menunjukkan keragaman sequense SAD kelapa sawit rendah, terbukti dengan hasil BLAST, semua sampel menunjukkan tingkat kemiripan yang tinggi satu sama lain (99%).
Analisa urutan nukleotida fragmen intron (noncoding) gen SAD menunjukkan adanya 5 situs SNPs. Pada gen SAD kelapa sawit ditemukan lebih banyak SNPS ditemukan pada intron dari pada exon (coding) perpanjang basa. SNPs dua kali lipat lebih banyak terdapat pada urutan basa noncoding dari pada daerah coding tanaman Bunga Matahari (Judith et al. 2007). SNPs yang terdapat pada daerah noncoding, dapat mempengaruhi tingkat ekspresi gen, stabilitas RNA dan menghasilkan varian kuantitatif (Zang dan Zhang 2005).
29 ....|....| ....|....| ....|....| ....|....| ....|....| ....|....|
5 15 25 35 45 55 Oleifera GTAAGTTTAT CATGACAGTC TAGTATTTGC TCAATTTCTT TAATTGATAT TTTCCTTGTG
Tenera GTAAGTTTAT CATGACAGTC TAGTATTTGC TCAATTTCTT TAATTGATAT TTTCCTTGTG Pisifera GTAAGTTTAT CATGACAGTC TAGTATTTGC TCAATTTCTT TAATTGATAT TTTCCTTGTG Dura GTAAGTTTAT CATGACAGTC TAGTATTTGC TCAATTTCTT TAATTGATAT TTTCCTTGTG Virescens GTAAGTTTAT CATGACAGTC TAGTATTTGC TCAATTTCTT TAATTGATAT TTTCCTTGTG ....|....| ....|....| ....|....| ....|....| ....|....| ....|....|
65 75 85 95 105 115 Oleifera ATTATAGCAT CATCAAATTC TGATAAGAGT GAGGGTGCAT TTTGCTTGCC CGAACAAGCT
Tenera ATTATAGCAT CATCAAATTC TGATAAGAGT GAGGGTGCAT TTTGCTTGCC CGAACAAGCT Pisifera ATTATAGCAT CATCAAATTC TGATAAGAGT GAGGGTGCAT TTTGCTTGCC CGAACAAGCT Dura ATTATAGCAT CATCAAATTC TGATAAGAGT GAGGGTGCAT TTTGCTTGCC CGAACAAGCT Virescens ATTATAGCAT CATCAAATTC TGATAAGAGT GAGGGTGCAT TTTGCTTGCC CGAACAAGCT ....|....| ....|....| ....|....| ....|....| ....|....| ....|....|
125 135 145 155 165 175 Oleifera TCAAAGGTCG CCAATCATGG AGAAACTTCT CTGAGATACA GTTGTTATCT CAAAATATGG
Tenera TCAAAGGTCG CCAATCATGG AGAAACTTCT CTGAGATACA GTTGTTATCT CAAAATATGG Pisifera TCAAAGGTCG CCAATCATGG AGAAACTTCT CTGAGATACG GTTGTTATCT CAAAATATGG Dura TCAAAGGTCG CCAATCATGG AGAAACTTCT CTGAGATACA GTTGTTATCT CAAAATATGG Virescens TCAAAGGTCG CCAATCATGG AGAAACTTCT CTGAGATACA GTTGTTATCT CAAAATATGG
....|....| ....|....| ....|....| ....|....| ....|....| ....|....|
185 195 205 215 225 235 Oleifera GCTTTTTTTT CTTTTATCTT TTTTTATTTT GAATAATTGG AGCAACTGCA AGTTACTTCA
Tenera GCTTTTTTTT CTTTTATCTT TTTTTATTTT GAATAATTGG AGCAACTGCA AGTTACTTCA Pisifera GCTTTTTTTT CTTTTATCTT TTTTTATTTT GAATAATTGG AGCAACTGCA AGTTACTTCA Dura GCTTTTTTTT CTGTTATCTT TTTTTATTTT GAATAATTGG AGCAACTGCA AGTTACTTCA Virescens GCTTTTTTTT CTTTTATCTT TTTTTATTTT GAATAATTGG AGCAACTGCA AGTTACTTCA
....|....| ....|....| ....|....| ....|....| ....|....| ....|....|
245 255 265 275 285 295 Oleifera AAAATAGCTT GCTTGATATT AAGTTATGCC TGAGTTCTTT TATGTGTTCT TTTAGGTTTC
Tenera AAAATAGCTT GCTTGATATT AAGTTATGCC TGAGTTCTTT TATGTGTTCT TTTAGGTTTC Pisifera AAAATATCTT GCTTGATATT AAGTTATGCC TGAGTTCTTT TATGTGTTCT TTTAGGTTTC Dura AAAATAGCTT GCTTGATATT AAGTTATGCC TGAGTTCTTT TATGTGTTCT TTTAGGTTTC Virescens AAAATAGCTT GCTTGATATT AAGTTATGCC TGAGTTCTTT TATGTGTTCT TTTAGGTTTC
....|....| ....|....| ....|....| ....|....| ....|....| ....|....|
305 315 325 335 345 355 Oleifera TTTTTAGCCC ATTATTGGCT TAGTGTTTTA TTATCTGGAT CAAACATCTT AGGATAGACT
Tenera TTTTTAGCCC ATTATTGGCT TAGTGTTTTA TTATCTGGAT CAAACATCTT AGGATAGACT Pisifera TTTTTAGCCC ATTATTGGCT TAGTGTTTTA TTATCTGGAT CAAACATCTT AGGATAGACT Dura TTTTTAGCCC ATTATTGGCT TAGTGTTTTA TTATCTGGAT CAAACATCTT AGGATAGACT Virescens TTTTTAGCCC ATTATTGGCT TAGTGTTTTA TTATCTGGAT CAAACATCTT AGGATAGACT
....|....| ....|....| ....|....| ....|....| ....|....| ....|....|
365 375 385 395 405 415 Oleifera ATTTTGTATC TGTTAACAAA TAGTTTGAAG CAGTGATTGC TTCCATAATT TGTTCAGCCG
Tenera ATTTTGTATC TGTTAACAAA TAGTTTGAAG CACTGATTGC TTCCATAATT TGTTCAGCCG Pisifera ATTTTGTATC TGTTAACAAA TAGTTTGAAG CAGTGATTGC TTCCATAATT TGTTCAGCCG Dura ATTTTGTATC TGTTAACAAA TAGTTTGAAG CAGTGATTGC TTCCATAATT TGTTCAGCCG Virescens ATTTTGTATC TGTTAACAAA TAGTTTGAAG CACTGATTGC TTCCATAATT TGTTCAGCCG
....|....| ....|....| ....|....| ....|....| ....|....| ....|....|
425 435 445 455 465 475 Oleifera ATGCTGGAGA TTGCTTGAAT TCATGGTATT TTTACTTGAT TGAATAGGTC ACATCAATTA
Tenera ATGCTGGAGA TTGCTTGAAT TCATGGTATT TTTACTTGAT TGAATATGTC ACATCAATTA Pisifera ATGCTGGAGA TTGCTTGAAT TCATGGTATT TTTACTTGAT TGAATATGTC ACATCAATTA Dura ATGCTGGAGA TTGCTTGAAT TCATGGTATT TTTACTTGAT TGAATATGTC ACATCAATTA Virescens ATGCTGGAGA TTGCTTGAAT TCATGGTATT TTTACTTGAT TGAATAGGTC ACATCAATTA
....|....| ....|....| ....|... 485 495 505 Oleifera ATATCACTTT CTTGTTTGCT GTCTGTAG Tenera ATATCACTTT CTTGTTTGCT GTCTGTAG Pisifera ATATCACTTT CTTGTTTGCT GTCTGTAG Dura ATATCACTTT CTTGTTTGCT GTCTGTAG Virescens ATATCACTTT CTTGTTTGCT GTCTGTAG
30
Intron telah terbukti meningkatkan efisiensi transkripsi banyak gen pada berbagai organisme. Sebagai contoh, satu studi awal pada tikus menunjukkan bahwa transgen tanpa intron menunjukkan transkripsi 10-100 kali lebih rendah dibandingkan yang mengandung intron. Demikian pula dengan efisiensi transkripsi pengkodean gen pada Drosophila, dimana alkohol dehidrogenase berkurang setelah penghapusan intron. Salah satu cara intron dapat mempengaruhi transkripsi adalah dengan bertindak sebagai penerima elemen regulasi transkripsi (Brinster 1988; Mackenzie et al. 1996; Helve et al. 2003).
Pada fragmen exon, untuk mendisain primer SNPs dilakukan penambahan 3 informasi sequense SAD dari aksesi kelapa sawit yang terdapat di NCBI (E.guineensis-Tenera aksesi U68756, E.oleifera aksesi EU057621 dan FJ940768) Lampiran 4. Hasil identifikasi memperlihatkan bahawa SNPs pada daerah exon gen SAD kelapa sawit yg dapat merubah asam amino (Non-synonymous) di 6 lokus SNPs, sedangkan 2 SNPs tidak merubah asam amino (synonymous) (Tabel 5). Mutasi titik di kodon pada posisi