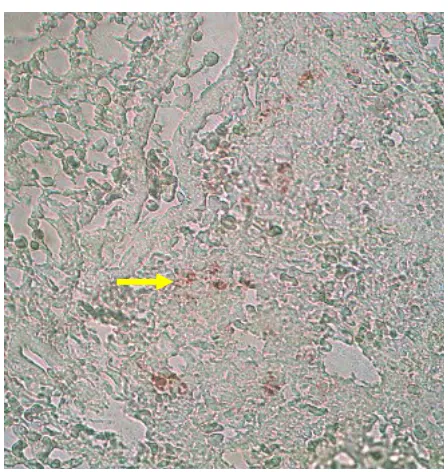
Distribusi Virus Avian Influenza (H5N1) Pada Jaringan Tubuh Itik Dengan Metode Imunohistokimia
Teks penuh
Gambar
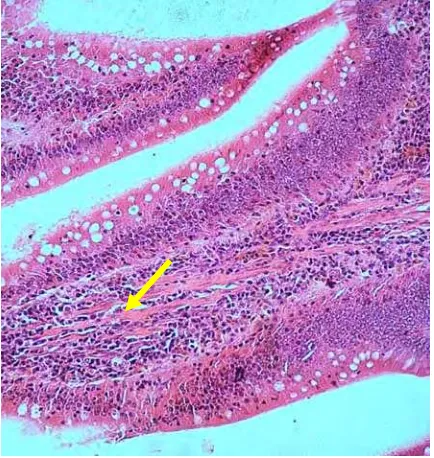

Dokumen terkait
Meskipun file ini dapat dilihat oleh pemakai, file kunci lisensi ini relative susah untuk dimanipulasi untuk merubah kondisi file dari status uji coba menjadi tidak
• Lawatan ke tapak bina di Ayer Baloi, Pontian bersama dengan Kak Shiqin (Quantity Surveyor) untuk membuat valuevation bersama Encik Hanafi selaku jurutera tapak
yang signifikan antara kelompok umur dan lokasi terhadap kemampuan membaca dalam bahasa Fataluku berarti bahwa dari reponden yang berumur lebih muda, ialah responden
Bab ketiga ini akan dijelaskan metode penelitiannya, antara lain tempat dan waktu pelaksanaan penelitian, bahan dan alat yang digunakan saat penelitian, prosedur
Berdasarkan kesimpulan di atas, maka penulis memberikan saran seba- gai berikut: (1) Bagi guru atau calon pe- neliti yang tertarik untuk menerapkan penelitian
,enar! tentu berbeda. 2rang akan lebih merasa terpuaskan dengan kenyang karena sepiring nasi dan lauk dibanding kenyang karena segelas air putih. Disitulah letak unsur atau
Seperti halnya pada palung cermin parabola, transfer cairan panas atau uap dipanaskan dalam receiver (menara yang mampu mengkonsentrasikan energi
Adapun tujuan dari penelitian ini adalah untuk mengetahui apakah hasil belajar siswa yang diajarkan dengan menggunakan metode ekspositori berbasis peta pikiran
