EKSPLORASI GEN
GROWTH HORMONE
EXON
3 PADA
KAMBING PERANAKAN ETAWAH (PE), SAANEN
DAN PESA MELALUI TEKNIK PCR-SSCP
SKRIPSI __ PAULINA YUNIARSIH
DEPARTEMEN ILMU PRODUKSI DAN TEKNOLOGI PETERNAKAN FAKULTAS PETERNAKAN
RINGKASAN
PAULINA YUNIARSIH. D14070043. 2011. Eksplorasi Gen Growth Hormone Exon 3 pada Kambing Peranakan Etawah (PE), Saanen dan PESA Melalui Teknik PCR-SSCP. Skripsi. Departemen Ilmu Produksi dan Teknologi Peternakan, Fakultas Peternakan, Institut Pertanian Bogor.
Pembimbing Utama : Dr. Jakaria, S.Pt., M.Si. Pembimbing Anggota : Prof Dr. Ir. Muladno, MSA
Ternak kambing memiliki keunggulan sebagai ternak yang memiliki potensi produktivitas yang cukup tinggi. Sehingga kambing memiliki peran penting sebagai sumber daya genetik ternak. Jumlah populasi kambing dan konsumsi daging di Indonesia yang masih rendah. Upaya peningkatan mutu genetik ternak kambing dapat dilakukan dengan seleksi dan persilangan. Pengukuran potensi ternak dapat diamati melalui sifat pertumbuhan. Pertumbuhan merupakan sifat yang dikendalikan banyak gen. Salah satu gen penting yang mempengaruhi pertumbuhan ternak kambing adalah gen growth hormone (GH) yang disekresikan oleh kelenjar
pituitary. Keragaman gen GH dapat diidentifikasi melalui teknik single-strand conformation polymorphism (SSCP). Penelitian ini bertujuan untuk mengetahui keragaman gen GH exon 3 pada kambing Peranakan Etawah (PE), Saanen dan PESA (Persilangan PE dan Saanen) melalui teknik PCR-SSCP.
ABSTRACT
Exploration of Exon 3 Growth Hormone Gene on
Etawah Grade (EG), Saanen and Their Crossbred Goat using PCR-SSCP Technique
Yuniarsih, P., Jakaria, and Muladno
This research was conducted to identify genetic polymorphism at the exon 3 growth hormone gene in three goat breeds. Polymorphisms at exon 3 growth hormone gene was identified by single strand conformational polymorphism polymerase chain reaction (SSCP-PCR) method. The DNA of 234 goat used were Etawah Grade (98 samples), Saanen (92 samples) and their crossbred (44 samples) in Cariu, Ciapus, Sukajaya, Cijeruk, Balitnak and Sukabumi. The PCR-SSCP method was performed at 250 V for 8 hours using 12% of acrylamide concentration. The result showed that the annealing temperature is 60 0C. The PCR product was 157 bp (base pair). The result SSCP method found four conformational patterns. The genotype frequency in exon 3 are AA (0,205), AB (0,856), AC (0,163) and BC (0,045). Beside that it was found three allele (allele A, B, and C). The highest frequencies were allele A (0,602) and B (0,443) at Saanen, Etawah Grade and their crossbred goat. The highest genotype frequency was AB at three goat breeds. The highest heterozygosity was found in Etawah Grade, Saanen and their crossbred goat (0,938). Exon 3 GH gene on three breeds goat have high polymorphism in six population.
EKSPLORASI GEN
GROWTH HORMONE
EXON
3 PADA
KAMBING PERANAKAN ETAWAH (PE), SAANEN
DAN PESA MELALUI TEKNIK PCR-SSCP
PAULINA YUNIARSIH D14070043
Skripsi ini merupakan salah satu syarat untuk memperoleh gelar Sarjana Peternakan pada
Fakultas Peternakan Institut Pertanian Bogor
DEPARTEMEN ILMU PRODUKSI DAN TEKNOLOGI PETERNAKAN FAKULTAS PETERNAKAN
Judul : Eksplorasi Gen Growth Hormone Exon 3 pada Kambing Peranakan Etawah (PE), Saanen Dan PESA Melalui Teknik PCR-SSCP
Nama : Paulina Yuniarsih NIM : D14070043
Menyetujui,
Pembimbing Utama, Pembimbing Anggota,
Dr. Jakaria, S.Pt. M.Si. Prof. Dr. Ir. Muladno, MSA NIP: 19660105 199303 1 001 NIP: 19610824 198703 1 001
Mengetahui, Ketua Departemen
Ilmu Produksi dan Teknologi Peternakan
Prof. Dr. Ir. Cece Sumantri, M. Agr. Sc. NIP: 19591212 198603 1 004
RIWAYAT HIDUP
Penulis dilahirkan pada tanggal 22 Juni 1989 di Tanjungkarang, Bandar Lampung. Penulis adalah anak ketiga dari empat bersaudara dari pasangan Bapak Ignasius. Jasmin dan Ibu Samirah.
Penulis mengawali pendidikan dasar pada tahun 1995 di Sekolah Dasar Fransiskus Tanjungkarang dan diselesaikan pada tahun 2001. Pendidikan lanjutan tingkat pertama dimulai pada tahun 2001 dan diselesaikan pada tahun 2004 di Sekolah Menengah Pertama Fransiskus Tanjungkarang. Di tingkat menengah atas, penulis dapat menyelesaikan pendidikan di Sekolah Menengah Atas Negeri 9 Bandar Lampung pada tahun 2007. Pada tahun yang sama penulis diterima sebagai mahasiswa Institut Pertanian Bogor pada tahun 2007 melalui jalur USMI (Undangan Seleksi Masuk IPB) pada Program Studi Produksi dan Teknologi Peternakan, Departemen Ilmu Produksi dan Teknologi Peternakan, Fakultas Peternakan IPB dan diselesaikan pada tahun 2011.
Selama mengikuti pendidikan di perguruan tinggi, penulis aktif di Unit Kegiatan Mahasiswa (UKM) Taekwondo sebagai Pelatih Tahun 2007-2010. Penulis aktif di Tim Pendamping IPB dan Asisten Praktikum Mata Kuliah Teknik Pengolahan Susu dan Genetika Ternak. Penulis aktif di Himpunan Mahasiswa Produksi Ternak (HIMAPROTER) Fakultas Peternakan IPB menjadi pengurus periode 2009-2010. Penulis tergabung keanggotaan dalam Animal Breeding Science
KATA PENGANTAR
Puji dan syukur penulis panjatkan kepada Tuhan Yang Maha Esa atas berkat dan rahmatNya sehingga penulis dapat menyelesaikan penelitian dan skripsi ini. Skripsi ini berjudul Eksplorasi Gen Growth Hormone exon 3 pada Kambing Peranakan Etawah (PE), Saanen dan PESA (Persilangan PE dan Saanen) melalui teknik PCR-SSCP.
Penelitian ini dilakukan untuk mengekplorasi gen growth hormone pada ternak kambing. Hal ini perlu dilakukan karena informasi genetik mengenai ternak kambing masih sangat jarang. Khususnya ternak kambing perah seperti kambing PE, Saanen dan PESA yang memiliki potensi produksi susu.
Hasil yang diperoleh dari penelitian ini berupa keragaman genetik yang dapat digunakan sebagai penciri DNA sebagai langkah awal untuk pembentukan bibit yang unggul. Penulis berharap, adanya penulisan skripsi ini dapat menjadi inspirasi bagi mahasiswa yang lain untuk lebih berani mengeksplorasi sumber daya genetik ternak kambing. Semoga skripsi ini dapat berguna dan bermanfaat untuk peternak Indonesia.
Bogor, April 2011
DAFTAR ISI
Kambing Persilangan PE dan Saanen (PESA) ... 6
Keragaman Genetik ... 7
Gen Growth Hormone (GH) ... 7
Keragaman Gen Growth Hormone ... 9
PCR-SSCP ... 10
Pendeteksian Keragaman Gen GH dengan Metode PCR-SSCP ... 15
Frekuensi Genotipe dan Alel ... 19
Nilai Heterozigositas ... 22
KESIMPULAN ... 24
Kesimpulan ... 24
Saran ... 24
UCAPAN TERIMA KASIH ... 25
DAFTAR PUSTAKA ... 26
DAFTAR TABEL
Nomor Halaman
1. Hasil Frekuensi Genotipe Gen GH Kambing Algarvia dengan
Teknik SSCP……… 9
2. Frekuensi Genotipe Gen GH Exon 3 pada Kambing PE, Saanen
DAFTAR GAMBAR
Nomor Halaman
1. Rekonstruksi Struktur Gen GH Berdasarkan Sekuens gen GH di
GenBank (Kode Akses D00476)……… 8 2. Fragmen Gen GH Capra hircus didasarkan pada Sekuens Gen GH
di GenBank (Kode Akses D00476)……… 10 3. Sekuens Primer Didasarkan pada Sekuens Gen GH exon 3 Kambing 13
4. Hasil Visualisasi Pita Gen GH pada Kambing Algarvia melalui
Teknik PCR-SSCP……….. 16
5. Hasil Amplifikasi Gen GH Exon 3 melalui PCR pada Gel
Poliakrilamida 6%... 17 6. Hasil Visualisasi Pita Gen GH Exon 3 melalui Teknik
1 PENDAHULUAN
Latar Belakang
Kambing merupakan ternak ruminansia kecil yang telah berkembang cukup luas di masyarakat Indonesia. Ternak kambing memiliki keunggulan yang menjadikannya sebagai ternak yang memiliki potensi produktivitas tinggi. Pemanfaatan kambing digunakan untuk produksi daging dan susu. Pemanfaatan yang lain adalah produksi kulit dan bulu sebagai hasil ikutan ternak. Kambing mampu beradaptasi pada lingkungan dengan hijauan yang terbatas sehingga tahan terhadap beberapa penyakit, dapat beranak sepanjang tahun, bersifat prolifik. Sehingga kambing memiliki peran penting sebagai sumber daya genetik ternak.
Jumlah populasi kambing di Indonesia masih tergolong rendah. Hal ini didukung oleh data Direktorat Jenderal Peternakan (2009) menyatakan bahwa populasi kambing di Indonesia pada tahun 2009 berjumlah 15.655.740 ekor. Kendala lain yang dihadapi adalah sistem pemeliharaan yang buruk. Pemanfaatan kambing secara genetik belum diteliti secara optimal. Banyak bangsa kambing di Indonesia yang belum dapat dikarakterisasi dan sebagian jumlah populasi mendekati punah. Kambing sebagai sumber daya genetik ternak belum dieksplorasi potensi keragaman genetik untuk dimanfaatkan sebagai sumber peningkatan mutu genetik sebagai penghasil susu dan daging. Upaya peningkatan mutu genetik ternak dapat dilakukan dengan seleksi dan persilangan.
Perkembangan ilmu genetika molekuler telah membuka peluang untuk mengetahui tingkat keragaman genetik pada tingkat DNA yang dapat digunakan untuk mengetahui potensi genetik suatu ternak. Teknologi DNA dapat menjadi dasar untuk penentuan genotipe gen-gen yang bernilai ekonomis yang diperlukan sebagai bibit yang unggul. Data produksi dan molekuler ternak kambing masih terbatas sehingga hal ini menjadi tantangan di teknologi molekuler untuk mengeksplorasi.
2 protein, lipid dan karbohidrat. Pendeteksian gen GH pada ternak kambing penting dilakukan untuk mengetahui keragaman gen tersebut karena diduga terkait dengan sifat-sifat yang bernilai ekonomis dapat dijadikan sebagai penciri genetik.
Keragaman gen dapat diidentifikasi dengan dua metode yaitu metode
restriction fragment length polymorphism (RFLP) dan metode single-strand conformation polymorphism (SSCP). Kedua teknik tersebut dapat dimanfaatkan untuk mengidentifikasi karakteristik gen-gen yang penting untuk pertumbuhan pada ternak. Teknik PCR-SSCP merupakan teknik yang mudah dan efisien untuk mengidentifikasi variasi urutan nukleotida pada fragmen gen DNA. Keterbatasan terhadap informasi keragaman genentik kambing sehingga teknik ini digunakan untuk mengidentifikasi gen GH pada ternak kambing.
Tujuan
3 TINJAUAN PUSTAKA
Klasifikasi Kambing
Kambing diklasifikasikam ke dalam kingdom Animalia; phylum Chordata; subphylum Vertebrata; class Mammalia; ordo Artiodactyla; sub-ordo Ruminantia;
family Bovidae; sub family Caprinae dan genus Capra (Mileski dan Myers 2004). Kambing memiliki 60 kromosom (30 pasang kromosom) yang terdiri atas 29 pasang kromosom autosom dan sepasang kromosom kelamin (Gall, 1981).
Kambing merupakan hewan yang didomestifikasi oleh manusia untuk produksi daging, susu dan kulit. Penyebaran kambing sangat luas dan hampir menyebar di seluruh dunia. Hal ini disebabkan daya adaptasi yang baik dari kambing terhadap berbagai iklim dan kemampuan bertahan hidup pada daerah dengan hijauan terbatas (Gall, 1981). Kambing adalah hewan ternak yang sanggup hidup di daerah kering dan pakan hijauan pakan yang terbatas, serta mampu memanfaatkan hijauan pakan secara efisien (Devendra dan Burns, 1994). Kambing dapat dikelompokkan berdasarkan kegunaannya, yaitu kambing penghasil daging, susu, dan bulu (mohair). Di samping itu, ada pula beberapa bangsa kambing yang tergolong tipe dwiguna (dual purpose), seperti bangsa kambing PE yang tergolong tipe daging dan susu (Heriyadi, 2004).
Kambing Lokal Indonesia
Kambing lokal merupakan kambing asli yang berasal dari Indonesia. Kambing lokal ini termasuk dalam kambing tipe pedaging. Kambing lokal Indonesia terdiri dari kambing Kacang, kambing Marica, kambing Samosir, kambing Muara, kambing Kosta, kambing Gembrong, kambing Peranakan Etawah (PE).
Kambing Kacang memiliki ciri bulu pendek dan berwarna tunggal (putih, hitam dan cokelat). Tanduk berbentuk pedang lengkung ke atas dan ke belakang. Pada umumnya telinga kambing pendek dan tegak. Kambing Kacang memiliki leher yang pendek dan punggungnya melengkung sedikit lebih tinggi dari pada bahunya. (Devendra dan Burns, 1983). Kambing jantan dan betina dewasa memiliki bobot kurang lebih 25 dan 20 kg (Devendra dan Burns, 1983; Pamungkas et al., 2009).
4 (Pamungkas et al., 2009). Kambing ini memiliki potensi genetik yang mampu beradaptasi baik di daerah agroekosistem lahan kering di daerah tanah bebatuan dengan curah hujan sepanjang tahun sangat rendah. Ciri yang paling khas pada kambing ini adalah telinganya tegak dan relatif kecil pendek dibanding telinga kambing kacang. Tanduk pendek, kecil, kelihatan lincah dan agresif (Pamungkas et al., 2009).
Kambing Samosir merupakan kambing yang dipelihara oleh masyarakat di Pulau Samosir, di tengah Danau Toba, Kabupaten Samosir, Provinsi Sumatera Utara (Pamungkas et al., 2009). Kambing ini dapat beradaptasi dengan kondisi ekosistem lahan kering dan berbatu-batu. Ciri khas yang paling menonjol adalah warna bulu putih yang sangat dominan, warna tanduk dan kuku agak keputihan (Pamungkas et al., 2009).
Kambing Muara berasal dari daerah Kecamatan Muara, Kabupaten Tapanuli Utara, Propinsi Sumatera Utara. Kambing ini memiliki penampilan yang gagah dan tubuh kompak. Selain itu, kambing ini memiliki warna bulu yang bervariasi yaitu warna bulu cokelat kemerahan, putih dan bulu hitam. Rata-rata bobot badan induk kambing adalah 49,4 kg dan pejantan dewasa 68,3 kg (Pamungkas et al., 2009).
Kambing Kosta banyak dikembangkan di Jakarta dan Propinsi Banten. Kambing Kosta merupakan persilangan kambing Kacang dan kambing Khasmir (Pamungkas et al., 2009). Kambing ini memiliki bentuk tubuh sedang, tanduk pendek, bulu pendek, hidung rata dan ditemukan melengkung. Warna tubuh kambing Kosta adalah cokelat tua sampai hitam (Pamungkas et al., 2009).
Kambing Gembrong berasal dari daerah timur Pulau Bali terutama di Kabupaten Karangasem. Ciri khas yang dimiliki kambing ini adalah rambut panjang mencapai 15-25 cm. Rambut ini terdapat pada bagian kepala sampai menutupi muka dan telinga. Warna tubuh dominan kambing gembrong pada umumnya putih (61,5%), sebagian berwarna cokelat muda (23,08%) dan cokelat (15,38%) (Pamungkas et al., 2009).
Kambing Peranakan Etawah (PE)
5 Timur dan pesisir utara Jawa Barat. Kambing PE telah berkembang dengan baik dan diterima masyarakat (Heriyadi, 2004).
Pemeliharaan kambing PE dapat menghasilkan daging dan susu (kambing tipe dwiguna). Kambing PE betina memiliki kemampuan menghasilkan susu yang baik. Kambing PE betina rata-rata dapat menghasilkan susu 1,2 liter per ekor per hari selama fase 70 hari pertama laktasi (Balai Penelitian Ternak, 2001). Kambing PE di Indonesia mampu menghasilkan susu 2-3 liter per ekor per hari dengan masa laktasi lebih dari 150 hari (Direktorat Jenderal Peternakan, 2003). Kambing PE memiliki karakteristik tubuh yang besar dengan bobot badan jantan dan betina mencapai 90 dan 60 kg (Direktorat Jenderal Peternakan, 2003).
Ciri-ciri spesifik kambing PE antara lain bentuk hidung benguk, panjang telinga 25-30 cm menggantung ke bawah dan sedikit kaku, tanduk melengkung, warna bulu bervariasi, kuping, kaki dan bulu yang panjang, memiliki ambing besar, dan produksi susu tinggi (Direktorat Jenderal Peternakan, 2003). Kambing PE memiliki ukuran tubuh yang relatif tinggi (65-86 cm), ramping dan relatif besar jika dibandingkan kambing Kacang
Kambing PE dapat beranak 3 kali dalam 2 tahun dengan rataan jumlah sekelahiran 1-3 ekor (Balai Penelitian Ternak, 2001). Rataan bobot lahir kambing PE kelahiran tunggal betina dan jantan masing-masing sebesar 3,2 dan 3,7 kg (Setiadi dan Sutama, 1997).
Kambing Saanen
Kambing Saanen berasal dari Swiss Barat. Kambing Saanen adalah kambing perah yang baik, memberikan penampilan yang baik, disesuaikan terhadap lingkungan subtropik dan sangat peka terhadap sinar matahari yang kuat. Kambing ini sangat peka terhadap cahaya sehingga pemeliharaan kambing Saanen di daerah tropis menggunakan naungan (Devendra dan McLeroy, 1982).
6 bertanduk (Greenwood, 1997). Kambing Saanen memiliki bentuk tubuh perah yang bagus dan ambing yang berkembang sangat baik (Devendra dan McLeroy, 1982).
Kambing Saanen memiliki rata-rata produksi susu 216 kg dengan panjang laktasi 275 hari (Gall, 1981). Menurut Devendra dan Burns (1994) rata-rata produksi susu kambing Saanen di daerah tropis mencapai 1-3 kg per ekor per hari, di daerah
temperate prduksi susu dapat mencapai lima kg per ekor per hari. Kambing Saanen mempunyai rata-rata produksi susu tertinggi dibandingkan dengan bangsa kambing perah manapun sehingga bangsa kambing ini telah dimasukkan ke banyak negara (Devendra dan Burns, 1994).
Kambing Saanen mempunyai bobot dewasa kelamin sekitar 50-70 kg dan tinggi betina dan jantan sekitar 81 dan 94 cm (Devendra dan Burns, 1994). Rataan berat badan kambing betina dan jantan berturut-turut adalah 65 dan 75 kg (Devendra dan McLeroy, 1982). Jumlah anak lahir seperindukan adalah 1,80 ekor (Devendra dan Burns, 1994).
Kambing Persilangan PE dan Saanen (PESA)
Kambing PESA merupakan kambing hasil persilangan antara kambing PE betina dengan kambing Saanen jantan. Wahyuarman (2001) melaporkan hasil persilangan PE dan Saanen memiliki keunggulan bobot lahir, bobot sapih dan produksi susu yang melebihi tetua PE masing-masing sebesar 0,22%; 5,47% dan 2,87%.
7 Keragaman Genetik
Genotipe hewan merupakan sebuah pendekatan yang berguna untuk menggambarkan prinsip-prinsip genetika dan penerapan langsung dalam hal pewarisan sifat. Hukum Hardy-Weinberg menyatakan bahwa frekuensi genotipe suatu populasi yang cukup besar akan selalu dalam keadaan seimbang bila tidak ditemukan seleksi, migrasi, mutasi dan genetic drift. Sifat-sifat ditemukan dalam keragaman genetik dalam spesies dan bangsa atau galur dalam masing-masing spesies. Genetika dipandang dari segi populasi, terutama frekuensi gen dengan efek yang diinginkan (Warwick et al., 1990).
Frekuensi gen merupakan istilah yang digunakan untuk menunjukkan proporsi dari semua lokus untuk pasangan gen atau rangkaian alel ganda dalam suatu populasi. Frekuensi gen dari perbedaan-perbedaan itu sangat beragam dari bangsa-bangsa dan antar galur (Warwick et al., 1990). Frekuensi gen yang timbul dipengaruhi oleh seleksi, mutasi gen, pencampuran dua populasi yang frekuensi gen berbeda, silang dalam (inbreeding), silang luar (outbreeding) dan genetic drift.
Ekspresi gen dapat mempengaruhi sifat yang yang muncul. Fenotipik yang muncul dapat dipengaruhi oleh variasi gen pada arah dan besar respon terhadap perubahan lingkungan (Noor, 2008). Fenotipik yang bersifat ekonomis merupakan sifat kuantitatif yang dikontrol oleh banyak gen dan masing-masing gen memberikan sedikit kontribusi pada sifat tersebut (Noor, 2008). Gen semacam ini disebut dengan gen mayor yang terletak pada lokus sifat kuantitatif atau quantitative traits loci
(QTL). Gen mayor yang dapat digunakan sebagai kandidat dalam program Marker Assisted Selection (MAS) jika gen tersebut mempunyai fungsi dan pengaruh biologis yang nyata terhadap sifat kuantitatif (Diyono, 2009).
Gen Growth Hormone (GH)
Gen growth hormone (GH) dikenal sebagai somatotropin yaitu terdiri atas 22.000-dalton hormon polipeptida rantai tunggal. Gen ini disintesis dan disekresi sel somatotrof pada lobus anterior pituitary. Gen GH pada vertebrata terdiri atas rantai polipeptida tunggal 190 atau 191 asam amino yang terdiri atas jembatan dua sulfida di antara sistein pada posisi 53-164 dan 181-89 (Paladini et al., 1983).
8
et al., 2001). Gen GH pada hewan yang sedang tumbuh, berguna untuk meningkatkan efisiensi produksi, pengurangan deposisi lemak, merangsang pertumbuhan otot, meningkatkan efisiensi penggunaan pakan, meningkatkan pertumbuhan organ dan meningkatkan pertumbuhan tulang (Etherton dan Bauman, 1998). Pada ternak ruminansia, gen GH berperan dalam pengaturan perkembangan kelenjar mamae (Akers, 2006).
Gen GH dijadikan sebagai salah satu kandidat gen yang dapat digunakan sebagai penanda genetik dalam program seleksi ternak (Malveiro et al., 2001). Gen-gen lain yang berpengaruh terhadap pertumbuhan yaitu Gen-gen Growth Hormone Releasing Hormone (GHRH), Somatotropin Releasing-Inhibitor Factor (SRIF),
Pituitary-Specific Positive Transcription Factor 1 (PIT-1) (Brunsch et al., 2002), dan
insulin-like growth factor-1 (IGF-1) (Hartman, 2000).
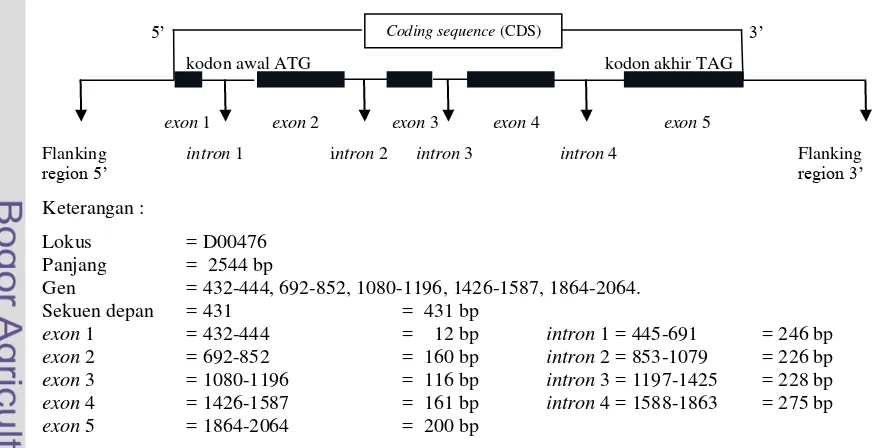
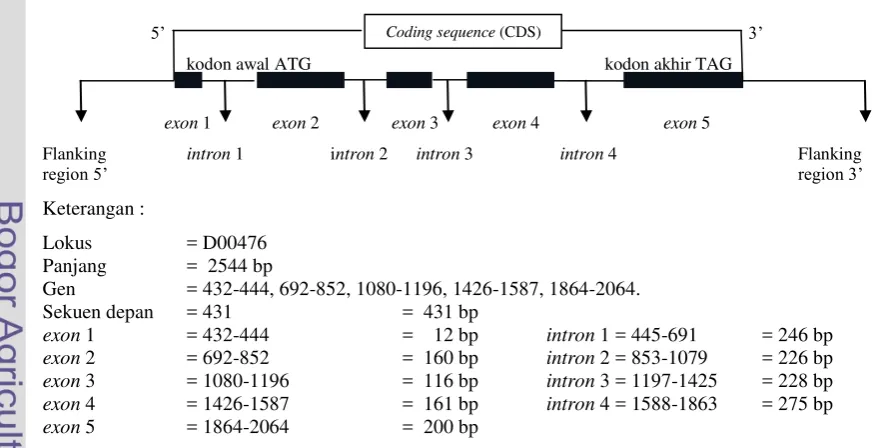
Gen GH memiliki kode yang terdiri atas empat intron dan lima exon pada semua spesies (Khan, 2009). Struktur gen GH dapat dilihat pada Gambar 1. Exon
adalah pengkode protein sementara intron merupakan spacer internal antara pengkode protein, pada saat transkripsi bagian intron hilang (splicing), sehingga proses translasi berjalan dengan baik (Jakaria, 2008). Sekuens gen GH pada Capra hircus berjumlah 2544 base pair (bp) yang dapat dilihat pada Gambar 2 (Malveiro et al., 2001).
Gen = 432-444, 692-852, 1080-1196, 1426-1587, 1864-2064. Sekuen depan = 431 = 431 bp
Gambar 1. Rekonstruksi Struktur Gen GH Berdasarkan Sekuens gen GH di GenBank
(Kode Akses D00476)
9 Keragaman Gen Growth Hormone (GH)
Identifikasi keragaman gen GH dapat dihubungkan antara sifat produksi susu dan polimorfisme gen GH kambing (Malveiro et al., 2001; Marques et al., 2003). Marques et al. (2003) melaporkan bahwa sampel DNA kambing Serrana yang dianalisis dengan teknik PCR-SSCP memiliki perbandingan yang tinggi pada polimorfisme genetik terutama gen GH. Dua bentuk konformasi dideteksi pada exon
1 dan 2, enam pola pada exon 3, sepuluh pola pada exon 4 dan lima pola pada exon 5. Yao et al. (1996) melaporkan bahwa ada dua polimorfisme yaitu T → C pada intron
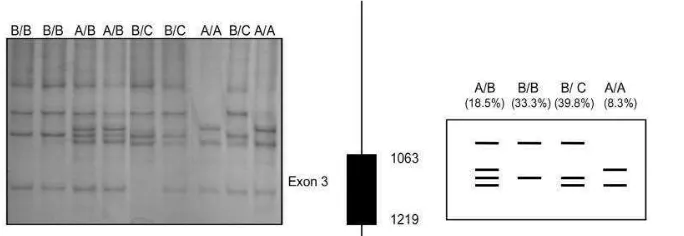
3 dan A → C pada exon 5 dengan menggunakan metode PCR-SSCP. Sedangkan Malveiro et al. (2001) melaporkan bahwa ada keragaman gen GH kambing melalui polimorfisme SSCP didapatkan dua bentuk konformasi pada exon 1 dan 2, empat pola pada exon 3, enam pola pada exon 4 dan lima pola pada exon 5. Pada exon 4 dan 5 gen GH ternak kambing Algarvia memiliki produksi, protein dan lemak susu tertinggi. Menurut Marques et al. (2003) pada exon 4 memiliki potensi produksi susu tinggi sehingga gen GH digunakan sebagai Marker Assisted Selection (MAS). Keragaman haploid gen GH│HaeIII pada kambing Boer berpengaruh terhadap bobot lahir, bobot sapih, pertambahan bobot badan per hari sebelum sapih dan bobot pada umur 11 bulan (Hua, 2009). Hasil frekuensi genotipe kambing Algarvia melalui teknik SSCP disajikan pada Tabel 1.
Tabel 1. Hasil Frekuensi Genotipe Gen GH Kambing Algarvia dengan Teknik SSCP
Exon Jumlah Pola Frekuensi Genotipe (%)
1 2 M (97,2); N (2,8)
2 2 A/B (75,9); B/B (24,1)
3 4 A/A (8,3); B/B (33,3); A/B (18,5); B/C (39,8)
4 6 A/A (13,9); B/B(27,8); C/C(35,2); D/D (5,6); E/E (14,8); F/F2,8)
5 5 A/A (2,8); B/B (27,8); A/B (14,8); B/C (44,4); A/C (10,2)
10
1 gggattttct gacccaggga ttaaacctga gtctcctgca tttgcagctc gattctttat 61 ggctgagcca cctgggaagc ccattcgttt ctgctacctc ccccttaaaa agaaaaccta 121 tggggtgggc tctcaagctg agaccctgtg tgtacagccc tcaggctggt ggcagtggag 181 aggggatgat gatgagcctg ggggacatga ccccagagaa ggaacgggaa caggatgagt 241 gagaggaggt tctaaattat ccattagcac aggctgccag tggtccttgc ataaatgtat 301 agagcacaca ggtgggggga aagggagaga gaagaagcca gggtataaaa agggcccagc 361 agagaccaat tccaggatcc caggacccag ttcaccagac gactcagggt cctgctgaca 421 gctcaccaac tatgatggct gcaggtaagc tcacaaaaat cccctccatt agcgtgtcct 481 aagggggtga tgcgggagaa ctgccgatgg atgtgtccac agctttgggt tttagggctt 541 ctgaatgcga acataggtat ctgcacccag acatttggcc aagtttgaaa tgttctcagt 601 ccctggaggg aagggcaggc gggggctggc aggagatcag gcatccagct ctctgggccc 661 ctccgtcgcg gccctcctgg tctctcccta gggccccgga cgtccctgct cctggctttc 721 accctgctct gcctgccctg gactcaggtg gtgggcgcct tcccagccat gtccttgtcc 781 ggcctgtttg ccaacgctgt gctccgggct cagcacctgc atcaactggc tgctgacacc 841 ttcaaagagt ttgtaagctc cccagagatg tgtcctagag gtggggaggc aggaaggggt 901 gaatccgcac cccctccaca caatgggagg gaactgagga cctcagtggt attttatcca 961 agtaaggatg tggtcagggg agtagaaatg ggggtgtgtg gggtggggag ggttccgaat 1021 aaggcagtga ggggaaccac acaccagctt agacccgggt gggtgtgttc tccccccagg 1081 agcgcaccta catcccggag ggacagagat actccatcca gaacacccag gttgccttct 1141 gcttctccga aaccatcccg gcccccacgg gcaagaatga ggcccagcag aaatcagtga 1201 gtggccacct aggaccgagg agcaggggac ctccttcatc ttaagtaggc tgccccagct 1261 ctctgcaccg ggcctggggt ggcgttctcc ctgaggtggc agagggtgtt ggatggcagt 1321 ggaggatgat ggttggtggt ggtggcagga ggtcctcggg cagaggccga ccttgcaggg 1381 ctgccccgag cccggggcac ccaccaacca cccatctgcc agcaggactt ggagctgctt 1441 cgcatctcac tgctccttat ccagtcgtgg cttgggcccc tgcagttcct cagcagagtc 1501 ttcaccaaca gcctggtgtt tggcacctcg gaccgtgtct atgagaagct gaaggacctg 1561 gaggaaggca tcctggcgct gatgcgggtg aggatggcgt tgttgggtcc cttccatgct 1621 gggggccatg cccaccctct cctggcttag ccaggagaac acacgtgggc tgggggagag 1681 agatccctgc tctctctctc tctttctagc agcccagtct tgacccagga gaaacctctt 1741 cccgttttga aacctccttc ctcgcccttc tccaagccta taggggaggg tggaaaatgg 1801 agcgggcagg agggagccgc tcctgagggc cttcggcctc tctgtctctc cctcccttgg 1861 caggagctgg aagatgttac cccccgggct gggcagatcc tcaagcagac ctatgacaaa 1921 tttgacacaa acatgcggag tgacgacgcg ctgctgaaga actacggtct gctctcctgc 1981 ttccggaagg acctgcacaa gacggagacg tacctgaggg tcatgaagtg tcgccgcttc 2041 ggggaggcga gctgcgcgtt ctagttgcca gccatctgtt gttacccctc cccgtgcctt 2101 cctagaccct ggaaggtgcc actccagtgc ccactgtcct ttcctaataa agcgaggaaa 2161 ttgcatcaca ttgtctgagt aggtgtcatt ctattctagg gggtggggtc aggcaggata 2221 gcgagaggga ggattgggaa gacaatagca gggatgctgt gggctctatg ggtacccagg 2281 tgctgaataa ttgacccggt tcttcctggg ccagaaggaa gcaggcacat ccccttctct 2341 gtgacacacc cggtcctcgc ccctggtcct tagttccagc cccactcata ggacactcat 2401 agctcaggag ggctctgcct tcagtcccac ccgctaaagt gcttggagcg gtttctcctt 2461 ccctcatcag cccaccaaac caaacctagc ctccaagagt gggaagaaat taaagcaaga 2521 caggctatga agtacagagg gaga
Gambar 2. Fragmen Gen GH Capra hircus didasarkan pada Sekuens Gen GH di
GenBank (Kode Akses D00476)
Polymerase Chain Reaction-Single Strand Conformation Polymorphism (PCR-SSCP)
11 nukleotida akan mempengaruhi bentuk (conformation) dari fragmen DNA untai tunggal (Bastos et al., 2001) dan laju migrasi pada saat elektroforesis (Orita et al.,
1989; Baroso et al,. 1999) walaupun perbedaan hanya satu nukleotida saja (Nataraj et al., 1999). Fragmen DNA untai tunggal yang mengalami perubahan pada susunan nukleotida akan membentuk suatu konformasi tiga dimensi yang kompleks dan berbeda fragmen DNA yang tidak mengalami perubahan (normal). Konformasi yang berbeda akan mempengaruhi laju migrasi dalam gel poliakrilamida sehingga diidentifikasi keragaman.
Faktor-faktor yang mempengaruhi sensitifitas SSCP yaitu: (1) komposisi gel, ukuran fragmen DNA, konsentrasi DNA, kandungan basa G dan C dalam fragmen DNA (Nataraj et al., 1999); (2) komposisi bufer (termasuk kekuatan ion dan derajat keasaman), bufer aditif seperti gliserol dan suhu saat elektroforesis (Sheffield et al., 1993 dan Nataraj et al., 1999); (3) persentase akrilamida dan rasio bis (Sheffield et al., 1993); (4) lokasi mutasi pada fragmen DNA (Baroso et al., 1999); (5) kelebihan dNTP dan primer dalam reaksi PCR (Cai and Touitou, 1994).
Metode analisis SSCP meliputi beberapa tahapan yaitu amplifikasi DNA target menggunakan primer melalui PCR, tahap denaturasi DNA produk PCR pada suhu 94 0C yang diikuti pendinginan untuk mencegah pre-annealing dari untaian DNA, penambahan formamida dye dan tahap elektroforesis dalam gel poliakrilamida (Nataraj et al., 1999; Hidayat et al., 2010).
Beberapa kelebihan SSCP dibanding metode lain yaitu: (1) sederhana dan tidak memerlukan peralatan yang rumit dan khusus (Bastos et al., 2001; Nataraj et al., 1999); (2) dapat mendeteksi mutasi pada fragmen DNA (Baroso et al., 1999) sehingga dapat dibedakan dengan yang normal (Nataraj et al., 1999); (3) visualisasi tidak perlu menggunakan bahan radioaktif (Nataraj et al., 1999); (4) dapat dikerjakan di laboratorium biasa dan tidak terlalu mahal (Bastos et al., 2001).
12 MATERI DAN METODE
Lokasi dan Waktu
Penelitian dilaksanakan mulai September 2010 sampai Januari 2011. Penelitian dilaksanakan di Laboratorium Genetika dan Molekuler Ternak, Bagian Pemuliaan dan Genetika Ternak, Departemen Ilmu Produksi dan Teknologi Peternakan, Fakultas Peternakan, Institut Pertanian Bogor.
Materi Sampel Darah dan Ekstraksi DNA
Sampel darah kambing disediakan sebanyak 234 sampel yang berasal dari kambing PE, Saanen dan PESA (Persilangan PE dan Saanen) pada lokasi berbeda. Sampel tersebut terdiri atas kambing PE berasal dari populasi Ciapus (20 sampel), Cariu (28 sampel) dan Sukajaya (50 sampel). Kambing Saanen berasal dari populasi Cijeruk (21 sampel), Cariu (31 sampel) dan Sukabumi (40 sampel). Kambing PESA berasal dari populasi Cariu (25 sampel) dan Balitnak (19 sampel).
Peternakan kambing perah Cariu memelihara kambing Peranakan Etawah, Saanen dan PESA dimana kandang berbentuk panggung. Bahan pakan yang diberikan berupa hijauan alam dan konsentrat yaitu ampas bir, ampas tahu dan dedak. Produksi susu harian 1-1,5 liter per hari per ekor. Bangsa kambing perah yang banyak dipelihara di Peternakan Cariu adalah Peranakan Etawah dengan sistem pemeliharaan intensif. Pakan yang diberikan berupa hijauan dan konsentrat berupa kulit ari kacang kedelai dan konsentrat. Rataan produksi susu kambing 0,8-1 liter per hari per ekor. Pemeliharaan ternak kambing perah di daerah Cijeruk adalah kambing Saanen dan PESA. Pemeliharaan ternak berbentuk kandang panggung. Produksi susu yang dihasilkan 2-4 liter per hari per ekor. Sampel kambing perah asal Balai Penelitian Ternak yang digunakan adalah kambing PESA. Produksi susu rata-rata 1 liter per hari per ekor (Nasution, 2010).
13 Primer GH
Primer merupakan molekul oligonukleotida yang berukuran pendek (18-24 base pair) dan menempel pada DNA cetakan di tempat yang spesifik. Primer yang digunakan dalam penelitian berdasarkan Malveiro et al. (2001) yaitu gen GH exon 3 sebagai berikut forward 5’-GTG TGT TCT CCC CCC AGG AG-3’ dan reverse 5’ -CTC GGT CCT AGG TGG CCA CT-3’. Primer dapat menempel gen GH dengan panjang produk 157 bp yang disajikan pada Gambar 3.
1021 aaggcagtga ggggaaccac acaccagctt agacccgggt gggtgtgttc tccccccagg
1081 agcgcaccta catcccggag ggacagagat actccatcca gaacacccag gttgccttct
1141 gcttctccga aaccatcccg gcccccacgg gcaagaatga ggcccagcag aaatcagtga
1201 gtggccacct aggaccgagg agcaggggac ctccttcatc ttaagtaggc tgccccagct
Keterangan : huruf tebal dan garis bawah merupakan situs primer
Gambar 3. Sekuens Primer Didasarkan pada Sekuens Gen GH Exon 3 pada Kambing Gel Poliakrilamida 12% untuk Polymerase Chain Reaction-Single-Strand Conformation Polymorphism (PCR-SSCP)
Komponen Gel Poliakrilamida 12% terdiri atas 10 ml larutan 30% akrilamida (acrylamide : bisacrylamide = 29:1); 2,5 ml larutan 5 x TBE (tris boric acid-EDTA); 15 µl TEMED (N,N,N’,N’-tetramethylethylenediamine) dan 150 µl 10% APS (ammonium peroxodisulfat). Alat-alat yang digunakan adalah plat kaca cetakan gel berukuran 20 x 20 cm, pipet makro dan mikro.
Pewarnaan Perak
Bahan-bahan yang digunakan dalam pewarnaan perak adalah amonia, AgNO3, NaOH, 10 N NaOH, formaldehid dan asam asetat. Alat yang digunakan adalah nampan dan water bath shaker.
Prosedur Pengambilan Sampel Darah
Sampel darah diambil melalui pembuluh vena jugularis menggunakan tabung
14 Ekstraksi DNA
Ekstraksi DNA dilakukan menggunakan sampel darah mengikuti metode
phenol-chloroform (Sambrook et al., 1989). Sampel darah dipindahkan sebanyak 200 µl ke dalam tabung 1,5 ml. Sampel ditambahkan aquadest atau TE sebanyak 1000 µl kemudian divorteks dan disentrifugasi kecepatan 8000 rpm selama lima menit. Sampel darah akan terbentuk dua lapisan. Lapisan supernatan dibuang dan ditambahkan TE sebanyak 1000 µl. Hal yang sama diulangi yaitu sampel divorteks dan disentrifugasi kecepatan 8000 rpm selama lima menit. Sampel akan terbentuk dua lapisan lagi. Lapisan atas yang disebut supernatan dibuang dan ditambahkan SDS 10% sebanyak 40 µl, STE 400 µl, proteinase K 10 µl. Sampel diinkubasi pada suhu 55 0C selama dua jam. Lalu sampel ditambahkan larutan fenol, CIAA, 5 M NaCl berturut-turut 400 µl, 400 µl, dan 40 µl. Sampel dikocok pelan selama satu jam lalu disentrifugasi kecepatan 12000 rpm selama lima menit. DNA bening yang berada di tabung 1,5 ml dipindahkan ke tabung baru sebanyak 400 µl dan ditambahkan EtoH absolute sebanyak 800 µl dan 5 M NaCl sebanyak 40 µl. Sampel DNA dibekukan selama over night. Setelah itu, sampel DNA disentrifuse kecepatan 12000 rpm selama lima menit. Supernatan yang muncul dibuang dan didiamkan dalam keadaan terbuka sampai alkohol hilang. Sampel DNA ditambahkan TE 80% sebanyak 100 µl. Sampel DNA disimpan dalam freezer sampai siap digunakan. Amplifikasi DNA
Gen GH exon 3 diamplifikasi oleh PCR menggunakan primer forward dan
15 Pendeteksian Keragaman Gen GH melalui Teknik Polymerase Chain Rreaction Single-Strand Conformation Polymorphism (PCR-SSCP)
Pendeteksian gen GH exon 3 berdasarkan Malveiro et al. (2001) dilakukan menggunakan 10 µl produk PCR yang ditambahkan larutan formamida dye ( 95% formamida;10 mM NaOH; 0,05% xylene cyanol dan 0,05% bromofenol blue) hingga mencapai 20 µl. Campuran tersebut didenaturasi pada suhu 95 0C selama lima menit dengan tujuan membuat fragmen DNA untai ganda menjadi untai tunggal. Setelah itu, tabung tersebut segera didinginkan pada suhu 0 0C selama tiga menit. Larutan
formamida dye berfungsi untuk mencegah penempelan kembali antara DNA untai tunggal. Sehingga didapatkan DNA untai tunggal yang dapat dideteksi keragaman bentuk dalam gel poliakrilamida 12%. Sampel sebanyak 5 µl dimasukkan ke dalam gel poliakrilamida 12% untuk mendeteksi konformasi untai DNA. Sampel DNA dielektroforesis pada gel poliakrilamida pada tegangan 250 volt selama delapan jam pada suhu 4 0C. Gel poliakrilamida 12% dikeluarkan dari kaca dan dilakukan proses pewarnaan perak.
Pewarnaan Perak
Pewarnaan perak dilakukan menggunakan metode Byun et al. (2009) yang telah dimodifikasi. Tahap awal, gel direndam dalam larutan A (200 ml air destilasi; 0,23 g AgNO3; 80 µl N NaOH dan 800 µl ammonia) selama delapan menit sambil digoyang menggunakan water bath shaker. Kemudian larutan dibuang dan gel dibilas dengan menggunakan air destilasi.
Pemunculan pita diperoleh dengan merendam gel dalam larutan B (200 ml air destilata, 6 g NaOH, 200 µl formaldehid) sambil dipanaskan. Setelah itu, larutan B dibuang dan gel direndam dalam 100 ml asam asetat untuk menghentikan reduksi perak.
Penentuan Genotipe
16 Gambar 4. Hasil Visualisasi Pita Gen GH Exon 3 pada Kambing Algarvia Melalui
Teknik PCR-SSCP (Malveiro et al., 2001) Analisis Data Frekuensi Alel dan Genotipe
Hasil genotipe dilakukan perhitungan frekuensi alel dan genotipe. Perhitungan frekuensi alel sebagai berikut (Nei dan Kumar, 2000) sebagai berikut:
Frekuensi genotipe dapat dihitung dengan persamaan sebagai berikut (Nei dan Kumar, 2000) :
Keterangan :
Xi = frekuensi alel ke-i Xii = frekuensi genotipe ke-i nii = jumlah individu bergenotipe ii nij = jumlah individu bergenotipe ij N = jumlah total sampel
Derajat Heterozigositas
Keragaman genetik dilakukan melalui perhitungan nilai heterozigositas pengamatan (Ho) (Weir, 1996) :
Keterangan :
Ho = frekuensi heterozigositas pengamatan
17 HASIL DAN PEMBAHASAN
Amplifikasi Gen GH
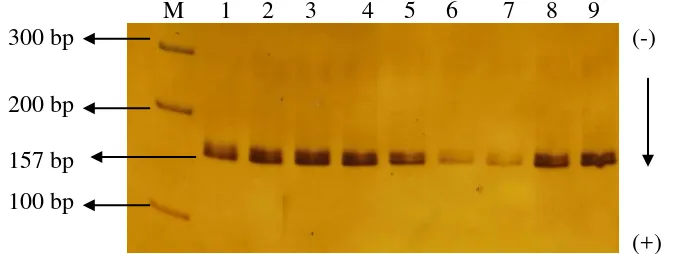
Gen GH exon 3 pada kambing PE, Saanen, dan PESA (Persilangan PE dan Saanen) berhasil diamplifikasi menggunakan metode PCR (Polymerase Chain Reaction). Panjang fragmen gen GH berukuran 2544 bp (GenBank Kode Akses D00476). Hasil amplifikasi gen GH exon 3 pada kambing adalah berukuran 157 bp. Proses amplifikasi gen GH exon 3 menggunakan pasangan primer Malveiro et al. (2001). Proses amplifikasi sangat dipengaruhi kondisi penempelan primer pada DNA target, bahan pereaksi PCR dan kondisi mesin thermal cycler. Gambar 5 memperlihatkan hasil amplifikasi gen GH exon 3 kambing.
M 1 2 3 4 5 6 7 8 9
300 bp (-)
200 bp 157 bp
100 bp
(+)
Gambar 6. Hasil Amplifikasi Gen GH Exon 3 Melalui Teknik PCR pada Gel Poliakrilamida 6% (M : marker 100 bp DNA)
Keberhasilan amplifikasi gen GH exon 3 dapat ditentukan oleh penempelan primer. Suhu yang digunakan agar primer dapat menempel pada fragment DNA target 60 0C. Suhu penempelan primer (annealing) sesuai dengan suhu yang digunakan dalam penelitian Malveiro et al. (2001). Suhu annealing menjadi penting dalam proses amplifikasi, hal ini dikarenakan proses penggandaan DNA baru dimulai dari primer. Suhu annealing adalah suhu yang membuat pasangan primer menempel dengan pasangan (komplemen) pada fragmen DNA target pada saat proses PCR dilakukan.
18 ditentukan oleh konsentrasi sampel DNA, taq polymerase, dinukleotida, ion Mg, bufer dan primer (Muladno, 2002).
Pendeteksian Keragaman Gen GH
Hasil pendeteksian keragaman GH exon 3 kambing melalui teknik PCR-SSCP dan gambaran dari diagram elektroforesis (zymogram) dari masing-masing genotipe kambing dapat dilihat pada Gambar 7 dan 8.
1 2 3 4 5 6 7 8
(-)
pita
target
(+)
AB BC AA AB AB AA AB AC
Gambar 7. Hasil Visualisasi Pita Gen GH Exon 3 Kambing Melalui Teknik PCR-SSCP pada Gel Poliakrilamida 12 %.
AA CC BB AB AC BC
Gambar 8. Diagram Elektroforesis (Zymogram) Pita Gen GH Exon 3 Pendeteksian keragaman gen GH exon 3 dilakukan menggunakan teknik
19 nukleotida akan mempengaruhi bentuk (conformation) dari fragmen DNA untai tunggal (Bastos et al., 2001).
Hasil pendekteksian ditemukan empat macam genotipe yaitu genotipe AA, AB, AC dan BC. Munculnya empat macam genotipe dibedakan berdasarkan jumlah pita yang muncul pada gel poliakrilamida 12%. Penentuan keempat macam genotipe didasarkan pada Malveiro et al. (2001). Genotipe AA merupakan genotipe homozigot yang muncul dua pita. Genotipe AB terbentuk karena dua genotipe homozigot (genotipe AA dan BB) bergabung menjadi heterozigot (muncul empat pita). Genotipe AC terbentuk karena dua genotipe homozigot (genotipe AA dan CC) yang bergabung menjadi heterozigot (muncul tiga pita). Genotipe CC tidak muncul dalam gel poliakrilamida 12% tetapi diasumsikan berdasarkan Malveiro et al. (2001) bahwa genotipe CC adalah satu pita. Genotipe BC terbentuk karena dua genotipe homozigot (genotipe BB dan CC) bergabung menjadi heterozigot (muncul tiga pita). Genotipe BB tidak muncul dalam gel sehingga genotipe ini diasumsikan dua pita sesuai dengan Malveiro et al. (2001). Menurut Bastos et al. (2001) jumlah maksimum pita yang muncul dari satu individu diploid adalah empat pita. Hasil visualisasi pada semua sampel DNA menunjukkan jumlah pita yang muncul yaitu dua, tiga dan empat pita.
Penemuan empat macam genotipe dapat dihasilkan tiga macam alel yaitu alel A, B dan C. Pendeteksian keragaman gen GH exon 3 yang ditemukan, didukung oleh Malveiro et al. (2001) yang menemukan dua macam genotipe homozigot (AA dan BB) dan dua pola genotipe heterozigot (AB dan BC) pada gen GH exon 3 dari 108 kambing Algarvia. Hasil pendeteksian yang dilakukan pada penelitian, ditemukan genotipe AC yang bersifat heterozigot dimana genotipe ini tidak ditemukan pada penelitian Malveiro et al. (2001). Genotipe AC diduga dapat mempengaruhi produksi susu kambing.
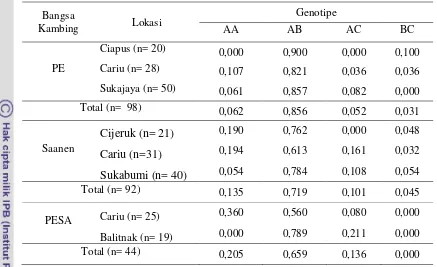
Frekuensi Genotipe dan Alel
20 Tabel 2. Frekuensi Genotipe Gen GH Exon 3 pada Kambing PE, Saanen dan PESA
Bangsa
Keterangan : n = jumlah individu
Hasil frekuensi genotipe pada bangsa kambing PE memiliki genotipe AB (0,856) tinggi, sedangkan genotipe AA (0,062), AC (0,052), dan BC (0,031) rendah. Pada bangsa kambing Saanen frekuensi AB (0,719) tinggi juga, kemudian AA poliakrilamida. Hasil penelitian ini berbeda dengan yang didapatkan oleh Malveiro et al. (2001) yang melaporkan frekuensi genotipe tertinggi pada genotipe BC (0,398), BB (0,333), AB (0,185) dan AA (0,083 ). Perbedaan frekuensi genotipe fragmen GH
exon 3 antara kambing PE, Saanen, dan PESA disebabkan oleh perbedaan bangsa ternak.
21 Tabel 3. Frekuensi Alel Gen GH Exon 3 pada Kambing PE, Saanen dan PESA
Bangsa Kambing Lokasi Alel
A B C
PE
Ciapus (n= 20) 0,450 0,500 0,050
Cariu (n= 28) 0,536 0,429 0,036
Sukajaya (n= 50) 0,531 0,429 0,041
Total (n= 98) 0,515 0,443 0,041
Saanen
Cijeruk (n= 21) 0,571 0,405 0,024
Cariu (n=31) 0,581 0,323 0,097
Sukabumi (n= 40) 0,500 0,419 0,081
Total (n= 92) 0,545 0,382 0,073
PESA Cariu (n= 25) 0,680 0,280 0,040
Balitnak (n= 19) 0,500 0,395 0,105
Total (n= 44) 0,602 0,330 0,068
Keterangan : n = jumlah individu
Pada kambing PE adalah alel A dan B tinggi. Frekuensi alel A sebesar 0,531 dan alel B sebesar 0,443. Frekuensi alel pada bangsa kambing Saanen memiliki alel A dan B tinggi. Frekuensi alel A dan B sebesar 0,545 dan 0,382. Frekuensi alel pada kambing PESA yaitu alel A dan B tinggi sebesar 0,602 dan 0,330. Sedangkan frekuensi alel C pada ketiga bangsa kambing dihasilkan alel yang rendah. Perbedaan frekeunsi alel dan genotipe disebabkan oleh bangsa ternak.
Hasil frekuensi genotipe (Tabel 2) dan alel (Tabel 3) fragmen gen GH exon
22 Nilai Heterozigositas
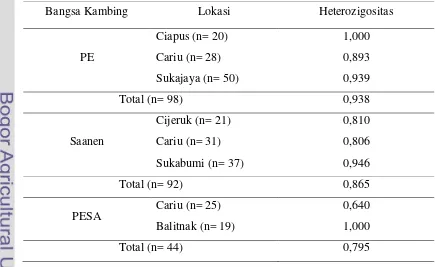
Derajat heterozigositas merupakan rataan persentase lokus heterozigositas tiap individu atau rataan persentase individu heterozigot dalam populasi. Nilai heterozigositas merupakan cara yang paling tepat untuk mengukur keragaman genetik suatu populasi (Nei, 1987). Nilai heterozigositas pengamatan pada fragmen gen GH exon 3 kambing PE, Saanen dan PESA disajikan pada Tabel 4.
Hasil analisis menunjukkan nilai heterozigositas pengamatan tertinggi pada kambing PE di lokasi Ciapus, Cariu dan Sukajaya merupakan nilai heterozigositas yang tinggi sebesar 1,000; 0,893; 0,939. Lokasi Cariu memiliki nilai heterozigositas tertinggi sebesar 1,000. Artinya di populasi sebanyak 20 individu memiliki genotipe heterozigot 100%. Nilai heterozigotas pada kambing Saanen di lokasi Cijeruk, Cariu dan Sukabumi memiliki nilai yang tinggi sebesar 0,810; 0,806; 0,946. Nilai heterozigotas pada kambing PESA di populasi Cariu dan Balitnak memiliki nilai sebesar 0,640 dan 1,000. Nilai heterozigositas sebesar 1,000 pada populasi Sukabumi artinya dalam populasi sebanyak 19 sampel memiliki genotipe heterozigot 100%. Nilai heterozigositas dipengaruhi oleh jumlah sampel yang terbatas, jumlah alel, dan frekuensi alel. Nilai Heterozigositas yang tinggi (1,000) menandakan bahwa genotipe AB dominan terhadap genotipe yang lain pada suatu lokasi.
Tabel 4. Nilai Heterozigositas Pengamatan pada Fragmen Gen GH
Bangsa Kambing Lokasi Heterozigositas
PE
Ciapus (n= 20) 1,000
Cariu (n= 28) 0,893
Sukajaya (n= 50) 0,939
Total (n= 98) 0,938
Saanen
Cijeruk (n= 21) 0,810
Cariu (n= 31) 0,806
Sukabumi (n= 37) 0,946
Total (n= 92) 0,865
PESA Cariu (n= 25) 0,640
Balitnak (n= 19) 1,000
23 Hasil analisis nilai heterosigositas pengamatan (Tabel 4) menunjukkan bahwa ketiga bangsa kambing memiliki keragaman yang tinggi. Hal ini ditandai dengan nilai heterozigositas yang tinggi pada kambing PE (0,938), kambing Saanen (0,865) dan kambing PESA (0,795). Semakin tinggi derajat heterozigositas suatu populasi maka daya tahan hidup populasi tersebut akan semakin tinggi seiring dengan menurunnya derajat heterozigositas akibat silang dalam dan fragmentasi populasi maka sebagian besar alel resesif yang bersifat letal akan semakin meningkat frekuensi (Avise, 1994). Nilai heterozigositas pengamatan dapat digunakan sebagai salah satu cara untuk menduga nilai koefisien biak dalam (inbreeding) pada suatu kelompok ternak (Hartl dan Clark, 1997). Nilai heterozigositas memiliki peranan penting untuk mengetahui tingkat polimorfisme suatu alel serta prospek populasi di masa yang akan datang (Falconer and Mackay, 1996).
Keragaman gen ternak kambing perah dapat dimanfaatkan untuk sifat produksi. Pemanfaatan dengan menghubungkan keragaman gen GH dengan sifat produksi susu dan pertumbuhan bobot badan ternak kambing pada lingkungan yang sama. Marques et al. (2003) melaporkan bahwa sampel DNA kambing Serrana yang dianalisis dengan teknik PCR-SSCP memiliki perbandingan yang tinggi pada polimorfisme genetik terutama gen GH. Dua bentuk konformasi dideteksi pada exon
24 KESIMPULAN DAN SARAN
Kesimpulan
Hasil penelitian gen GH exon 3 dapat disimpulkan bahwa sampel DNA kambing PE, Saanen dan PESA dengan teknik PCR-SSCP bersifat beragam. Pada fragmen gen GH exon 3 ditemukan empat macam genotipe yaitu AA, AB, AC dan BC. Tiga macam alel ditemukan yaitu alel A, B dan C. Kambing PE, Saanen dan PESA di lokasi Cariu, Ciapus, Sukajaya, Sukabumi, Cijeruk dan Balitnak memiliki nilai heterozigositas yang tinggi. Analisis tersebut menunjukkan bahwa gen GH exon
3 pada ketiga bangsa kambing di lokasi berbeda memiliki polimorfisme atau keragaman yang tinggi.
Saran
25 UCAPAN TERIMA KASIH
Penulis memanjatkan syukur kepada Tuhan atas segala berkat dan rahmatnya sehingga penulis dapat menyelesaikan penulisan skripsi ini. Penulis mengucapkan terima kasih kepada Dr. Jakaria, S.Pt., M.Si. dan Prof. Dr. Ir. Muladno, MSA selaku pembimbing atas segala bimbingan, perhatian, motivasi, arahan dan nasihat yang diberikan. Penulis juga mengucapkan terima kasih kepada Ir. Rini H. Mulyono, MSi dan Dr. Ir. Yuli Retnani, MSc selaku penguji yang telah memberikan kritik dan saran. Untuk Ir. Maman Duldjaman, MS selaku pembimbing akademik yang telah memberikan nasehat. Penulis mengucapkan terima kasih kepada kedua orang tua tercinta yang tidak pernah berhenti memberikan kasih sayang, doa dan dukungan. Penulis tidak akan pernah mampu membalas kasih sayang dan pengorbanan yang telah diberikan. Kepada Maria, Daniel dan Katrin yang selalu memberikan semangat dan motivasi bagi penulis. Kepada Bapak Yahya Sudarma (alm), penulis mengucapkan banyak terima kasih atas bantuan, perhatian, kasih sayang selama delapan tahun sudah menjadi orang tua asuh bagi penulis.
Penulis juga mengucapkan terima kasih kepada Prof. Cece yang memberikan saran dan kritik. Kak Erick, Kak Restu, Kak Surya, Pak Ihsan, Pak Andi yang tergabung dalam ABGSCi yang telah membantu saya dalam menjalankan penelitian ini dan memberikan pengalaman yang berharga bagi penulis.
Untuk teman seperjuangan di Lab. Genetika dan Molekuler Ternak, Ferdy, Irine, Lenny, Priskila, Annisa, Wieke, Gabby, Dhini, Tifanny, Gina, Desy, Rahmah, Dian Dinar dan Erwin yang tergabung dalam kelompok kurcaci. Tidak lupa juga penulis mengucapkan terima kasih kepada Keluarga IPTP 44. Selain itu, ucapan terima kasih ini untuk teman-teman Pendamping IPB terutama Alfons dan Densus 08 (Lucia, Anton, Lisa, Eny, Adian, Imanta, Gregorio, Ariyanti, Aprella, Dika, Chissy, Brury, Sari, Bambang, Ayu, Izak dan Leo). Akhirnya penulis mengharapkan semoga karya ini bermanfaat bagi upaya pembangunan peternakan di Indonesia.
Bogor, April 2011
26 DAFTAR PUSTAKA
Agung, P. P. 2009. Identifikasi keragaman gen miostatin dan pengaruhnya terhadap sifat pertumbuhan domba lokal Indonesia.Tesis. Sekolah Pascasarjana, Institut Pertanian Bogor, Bogor.
Akers, R. M. 2006. Major advances associated with hormone and growth factor regulation of mammary growth and lavtation in dairy cows. J. Dairy Sci. 89: 1222-1234.
Avise JC. 1994. Moleculer Markers Natural History Evolution. Washington: Chapman and Hall Inc.
Balai Penelitian Ternak. 2001. Kambing PE penghasil daging sekaligus susu. warta penelitian dan pengembangan pertanian. Vol. 23 no. 4.
Barroso, A., S. Dunner, & J. Canon. 1999. Technical note : use of PCR-single strand conformation polymorphism analysis for detection of bovine β-casein variants A1, A2, A3 and B. J. Anim Sci.77 : 2629-2632.
Bastos, E., A. Crvador, J. Azevedo, & H. G. Pinto. 2001. Single strand conformation polymorphism (SSCP) detection in six genes in Portuguese indigenous sheep breed “Churra da Terra Quente”. Biotechnol. Agron. Soc. Environ. 5(1): 7-15.
Brunsch, C., I. Strenstein., P. Reinecke & J. Bieniek. 2002. Analysis of Associations of Pit-1 genotypes with growh, meat quality and carcass composition traits in pigs. J. Appl. Genet. 43 (1): 85-91.
Byun, S.O., Q. Fang, H. Zhou, & J. G. H. Hickford. 2009. An effective method for silver-staining DNA in large numbers of polyacrylamide gels. Analytical Biochemistry 385: 174–175.
Cai, Q.Q., & I. Touitou. 1994. Excess PCR primers may dramatically affect SSCP efficiency. Nucleic Acid Res. 21: 3909-3910.
Devendra, C., & G. B. McLeroy. 1982. Goat and Sheep Production in The Tropics. Intermediate Tropical Agriculture Series. Longman, London and New York. Devendra, C., & M. Burn. 1983. Goat Production in The Tropics. Commonwealth
Agricultural Bureaux, London.
Devendra, C., & M. Burn. 1994. Produksi Kambing di Daerah Tropis. Terjemahan : Harya Putra. Penerbit Institut Teknologi Bandung, Bandung.
Direktorat Jenderal Peternakan. 2009. Statistik Peternakan 2009. Jakarta: Direktorat Jenderal Peternakan Departemen Pertanian RI.
27 Direktorat Jenderal Bina Produksi Peternakan. Departemen Pertanian RI, Jakarta.
Diyono, R. 2009. Karakteristik ukuran tubuh dan polimorfisme gen GH, GHRH dan Pit-1 pada populasi kerbau di Banten. Tesis. Sekolah Pascasarjana, Institut Pertanian Bogor, Bogor.
Entherton, T. D. & D. E. Bauman. 1998. Biology of somatotropin in growth and lactation of domestic animals. Physical Rev., 78: 745-761.
Falconer DS & Mackay TFC. 1996. Introduction to Quantitative Genetics. Fourth Ed. Longman Inc. New York.
Gall, C. 1981. Goat Production. Academic Press Inc., New York.
Garrett, A., G. Rincon, J. F. Medrano, M. A. Elzo, G. A. Silver, & M. G. Thomas. 2008. Promoter region of bovine growth hormone receptor gene : Single nucleotide polymorphism discovery in cattle and association with performance in Brangus bulls. J. Anim. Sci. 86 : 3315-3323.
Greenwood, P. 1997. Goat Breed Saanen. Agfact A7.3.4.2nd Edition.
Hartl D. L. & A. G. Clark. 1997. Principle of Population Genetic. Sinauer Associates, Sunderland, MA.
Hartman, M. L. 2000. Physiological Regulations of Growth Hormone Secretion. 2nd Ed. Cambridge Unversity Press, Cambrige.
Heriyadi, D. 2004. Standarisasi Mutu Bibit Kambing PE. Kerja Sama Antara Dinas Peternakan Provinsi Jawa Barat dengan Fakultas Peternakan Universitas Padjadjaran, Bandung.
Hua, G. H. 2009. Polymorphism of the growth hormone gene and its association with growth traits in Boer goat bucks. Meat Sci 81: 391-395.
Hidayat, T., & A. Pancoro. 2010. Single strand conformation polymorphism method for initial detection DNA sequences homogeneity. J. Biosci. 17 : 50-52. Khan, M. A. 2009. Genetic analysis of caprine growth hormone gene and production
of recombinant caprine growth hormone.Thesis. School of Biological Sciences. University of The Punjab, Pakistan.
28 Mileski, A., & P. Myers. 2004. Capra hircus, Animal Diversity Web. http://animaldiversity.ummz.umich.edu/site/accounts/information/Capra_hirc us.html. [27 Juli 2010].
Muladno. 2002. Seputar Teknologi Rekayasa Genetika. Pustaka Wirausaha Muda, Bogor.
Nasution, A. 2010. Keragaman kualitas susu dan keju dari susu kambing peranakan etawah (PE), saanen dan persilangannya. Tesis. Sekolah Pascasarjana, Institut Pertanian Bogor, Bogor.
Nataraj, A. J., I.,O. Glander, N. Kusukawa, & W. E. Highsmith.1999. Single-strand conformation polymorphism and heteroduplex analysis gor gel-based mutation detection. WILEY-VCH Verlag GmnH. Wienheim.
Nei, M. 1987. Molecular Evalutionery Genetics. Columbia University Press. New York.
Nei, M. & S. Kumar. 2000. Molecular Evolution and Phylogenetics. Oxford University Press, New York.
Noor, R. R. 2008.Genetika Ternak. Penebar Swadaya, Jakarta.
Orita, M., Y.Suzuki, T. Sekiya, & K. Hayashi.1989. Rapid and sensitive detection of point mutations and DNA polymorphisms using the polymerase chain reaction. Genomics 5: 874-879.
Paladini, A. C., Pena, C. & Poskus, E. 1983. Molecular biology of growth hormone. CRC. Crit. Rev. Biochem. 15 : 25-26.
Pamungkas, F. A., A. Batubara, M. Doloksaribu, & E. Sihite. 2009. Potensi beberapa plasma nutfah kambing lokal Indonesia. Pusat Penelitian dan Pengembangan Peternakan, Sumatera Utara.
Ruhimat A. 2003. Produktivitas kambing persilangan Peranakan Etawah betina dengan kambing Saanen jantan (PESA) di PT Fajar Taurus Dairy Farm skripsi. Bogor: Fakultas Peternakan, Institut Pertanian Bogor.
Sambrook, J., E. F. Fritsch & T. Manuatis.1989.Molecular Cloning, A Laboratory Manual. Third Editio. Cold Sring Harbour Lab. Press, New York.
Sheffield, V. C., J. S. Beck, A. E. Kwitek, D.W. Sandstorm & E. M. Stone. 1993. The sensitivity of single-strand conformation polymorphism for detection of single base substitution. Genomics. 16:325-332.
Setiadi, B., & I.K. Sutama.1997. Kambing PE, Kambing Perah Indonesia. Balai Penelitian Ternak, Bogor.
29 Warwick, E. J., J. M. Astuti, & W. Hardjosubroto.1990. Pemuliaan Ternak. Gadjah
Mada University Press, Yogyakarta.
Weir, B.S. 1996. Genetic Data Analysis : Method for Discrete Population Genetic Data. Second ed. Sinauer Associates. Sunderland, MA USA.
Yao, J., Aggrey, S. E. Zadworny, D., Hayes, & J.F. Kuhnlein. 1996. Sequence variations in the bovine GH gene characterization by SSCP analysis and their association with milk production traits in Holstein. Genetics 144: 1809-1816.
31 Lampiran 1. Informasi Sekuens Gen GH pada Ternak Kambing (Capra hircus)
LOCUS GOTGHRA 2544 bp DNA hilinear MAM 12-JUL-2007
DEFINITION Capra hircus gene for growth hormone, complete cds. ACCESSION D00476
Mammalia; Eutheria; Laurasiatheria; Cetartiodactyla; Ruminantia; Pecora; Bovidae; Caprinae; Capra.
REFERENCE 1 (bases 1 to 2544)
AUTHORS Kioka,N., Manabe,E., Abe,M., Hashi,H., Yato,M., Okuno,M., Yamano,Y., Sakai,H., Komano,T., Utsumi,K. and Iritani,A. TITLE Cloning and sequencing of goat growth hormone gene JOURNAL Agric. Biol. Chem. 53, 1583-1587 (1989)
32
1 gggattttct gacccaggga ttaaacctga gtctcctgca tttgcagctc gattctttat 61 ggctgagcca cctgggaagc ccattcgttt ctgctacctc ccccttaaaa agaaaaccta 121 tggggtgggc tctcaagctg agaccctgtg tgtacagccc tcaggctggt ggcagtggag 181 aggggatgat gatgagcctg ggggacatga ccccagagaa ggaacgggaa caggatgagt 241 gagaggaggt tctaaattat ccattagcac aggctgccag tggtccttgc ataaatgtat 301 agagcacaca ggtgggggga aagggagaga gaagaagcca gggtataaaa agggcccagc 361 agagaccaat tccaggatcc caggacccag ttcaccagac gactcagggt cctgctgaca 421 gctcaccaac tatgatggct gcaggtaagc tcacaaaaat cccctccatt agcgtgtcct 481 aagggggtga tgcgggagaa ctgccgatgg atgtgtccac agctttgggt tttagggctt 541 ctgaatgcga acataggtat ctgcacccag acatttggcc aagtttgaaa tgttctcagt 601 ccctggaggg aagggcaggc gggggctggc aggagatcag gcatccagct ctctgggccc 661 ctccgtcgcg gccctcctgg tctctcccta gggccccgga cgtccctgct cctggctttc 721 accctgctct gcctgccctg gactcaggtg gtgggcgcct tcccagccat gtccttgtcc 781 ggcctgtttg ccaacgctgt gctccgggct cagcacctgc atcaactggc tgctgacacc 841 ttcaaagagt ttgtaagctc cccagagatg tgtcctagag gtggggaggc aggaaggggt 901 gaatccgcac cccctccaca caatgggagg gaactgagga cctcagtggt attttatcca 961 agtaaggatg tggtcagggg agtagaaatg ggggtgtgtg gggtggggag ggttccgaat 1021 aaggcagtga ggggaaccac acaccagctt agacccgggt gggtgtgttc tccccccagg 1081 agcgcaccta catcccggag ggacagagat actccatcca gaacacccag gttgccttct 1141 gcttctccga aaccatcccg gcccccacgg gcaagaatga ggcccagcag aaatcagtga 1201 gtggccacct aggaccgagg agcaggggac ctccttcatc ttaagtaggc tgccccagct 1261 ctctgcaccg ggcctggggt ggcgttctcc ctgaggtggc agagggtgtt ggatggcagt 1321 ggaggatgat ggttggtggt ggtggcagga ggtcctcggg cagaggccga ccttgcaggg 1381 ctgccccgag cccggggcac ccaccaacca cccatctgcc agcaggactt ggagctgctt 1441 cgcatctcac tgctccttat ccagtcgtgg cttgggcccc tgcagttcct cagcagagtc 1501 ttcaccaaca gcctggtgtt tggcacctcg gaccgtgtct atgagaagct gaaggacctg 1561 gaggaaggca tcctggcgct gatgcgggtg aggatggcgt tgttgggtcc cttccatgct 1621 gggggccatg cccaccctct cctggcttag ccaggagaac acacgtgggc tgggggagag 1681 agatccctgc tctctctctc tctttctagc agcccagtct tgacccagga gaaacctctt 1741 cccgttttga aacctccttc ctcgcccttc tccaagccta taggggaggg tggaaaatgg 1801 agcgggcagg agggagccgc tcctgagggc cttcggcctc tctgtctctc cctcccttgg 1861 caggagctgg aagatgttac cccccgggct gggcagatcc tcaagcagac ctatgacaaa 1921 tttgacacaa acatgcggag tgacgacgcg ctgctgaaga actacggtct gctctcctgc 1981 ttccggaagg acctgcacaa gacggagacg tacctgaggg tcatgaagtg tcgccgcttc 2041 ggggaggcga gctgcgcgtt ctagttgcca gccatctgtt gttacccctc cccgtgcctt 2101 cctagaccct ggaaggtgcc actccagtgc ccactgtcct ttcctaataa agcgaggaaa 2161 ttgcatcaca ttgtctgagt aggtgtcatt ctattctagg gggtggggtc aggcaggata 2221 gcgagaggga ggattgggaa gacaatagca gggatgctgt gggctctatg ggtacccagg 2281 tgctgaataa ttgacccggt tcttcctggg ccagaaggaa gcaggcacat ccccttctct 2341 gtgacacacc cggtcctcgc ccctggtcct tagttccagc cccactcata ggacactcat 2401 agctcaggag ggctctgcct tcagtcccac ccgctaaagt gcttggagcg gtttctcctt 2461 ccctcatcag cccaccaaac caaacctagc ctccaagagt gggaagaaat taaagcaaga 2521 caggctatga agtacagagg gaga
EKSPLORASI GEN
GROWTH HORMONE
EXON
3 PADA
KAMBING PERANAKAN ETAWAH (PE), SAANEN
DAN PESA MELALUI TEKNIK PCR-SSCP
SKRIPSI __ PAULINA YUNIARSIH
DEPARTEMEN ILMU PRODUKSI DAN TEKNOLOGI PETERNAKAN FAKULTAS PETERNAKAN
RINGKASAN
PAULINA YUNIARSIH. D14070043. 2011. Eksplorasi Gen Growth Hormone Exon 3 pada Kambing Peranakan Etawah (PE), Saanen dan PESA Melalui Teknik PCR-SSCP. Skripsi. Departemen Ilmu Produksi dan Teknologi Peternakan, Fakultas Peternakan, Institut Pertanian Bogor.
Pembimbing Utama : Dr. Jakaria, S.Pt., M.Si. Pembimbing Anggota : Prof Dr. Ir. Muladno, MSA
Ternak kambing memiliki keunggulan sebagai ternak yang memiliki potensi produktivitas yang cukup tinggi. Sehingga kambing memiliki peran penting sebagai sumber daya genetik ternak. Jumlah populasi kambing dan konsumsi daging di Indonesia yang masih rendah. Upaya peningkatan mutu genetik ternak kambing dapat dilakukan dengan seleksi dan persilangan. Pengukuran potensi ternak dapat diamati melalui sifat pertumbuhan. Pertumbuhan merupakan sifat yang dikendalikan banyak gen. Salah satu gen penting yang mempengaruhi pertumbuhan ternak kambing adalah gen growth hormone (GH) yang disekresikan oleh kelenjar
pituitary. Keragaman gen GH dapat diidentifikasi melalui teknik single-strand conformation polymorphism (SSCP). Penelitian ini bertujuan untuk mengetahui keragaman gen GH exon 3 pada kambing Peranakan Etawah (PE), Saanen dan PESA (Persilangan PE dan Saanen) melalui teknik PCR-SSCP.
ABSTRACT
Exploration of Exon 3 Growth Hormone Gene on
Etawah Grade (EG), Saanen and Their Crossbred Goat using PCR-SSCP Technique
Yuniarsih, P., Jakaria, and Muladno
This research was conducted to identify genetic polymorphism at the exon 3 growth hormone gene in three goat breeds. Polymorphisms at exon 3 growth hormone gene was identified by single strand conformational polymorphism polymerase chain reaction (SSCP-PCR) method. The DNA of 234 goat used were Etawah Grade (98 samples), Saanen (92 samples) and their crossbred (44 samples) in Cariu, Ciapus, Sukajaya, Cijeruk, Balitnak and Sukabumi. The PCR-SSCP method was performed at 250 V for 8 hours using 12% of acrylamide concentration. The result showed that the annealing temperature is 60 0C. The PCR product was 157 bp (base pair). The result SSCP method found four conformational patterns. The genotype frequency in exon 3 are AA (0,205), AB (0,856), AC (0,163) and BC (0,045). Beside that it was found three allele (allele A, B, and C). The highest frequencies were allele A (0,602) and B (0,443) at Saanen, Etawah Grade and their crossbred goat. The highest genotype frequency was AB at three goat breeds. The highest heterozygosity was found in Etawah Grade, Saanen and their crossbred goat (0,938). Exon 3 GH gene on three breeds goat have high polymorphism in six population.
1 PENDAHULUAN
Latar Belakang
Kambing merupakan ternak ruminansia kecil yang telah berkembang cukup luas di masyarakat Indonesia. Ternak kambing memiliki keunggulan yang menjadikannya sebagai ternak yang memiliki potensi produktivitas tinggi. Pemanfaatan kambing digunakan untuk produksi daging dan susu. Pemanfaatan yang lain adalah produksi kulit dan bulu sebagai hasil ikutan ternak. Kambing mampu beradaptasi pada lingkungan dengan hijauan yang terbatas sehingga tahan terhadap beberapa penyakit, dapat beranak sepanjang tahun, bersifat prolifik. Sehingga kambing memiliki peran penting sebagai sumber daya genetik ternak.
Jumlah populasi kambing di Indonesia masih tergolong rendah. Hal ini didukung oleh data Direktorat Jenderal Peternakan (2009) menyatakan bahwa populasi kambing di Indonesia pada tahun 2009 berjumlah 15.655.740 ekor. Kendala lain yang dihadapi adalah sistem pemeliharaan yang buruk. Pemanfaatan kambing secara genetik belum diteliti secara optimal. Banyak bangsa kambing di Indonesia yang belum dapat dikarakterisasi dan sebagian jumlah populasi mendekati punah. Kambing sebagai sumber daya genetik ternak belum dieksplorasi potensi keragaman genetik untuk dimanfaatkan sebagai sumber peningkatan mutu genetik sebagai penghasil susu dan daging. Upaya peningkatan mutu genetik ternak dapat dilakukan dengan seleksi dan persilangan.
Perkembangan ilmu genetika molekuler telah membuka peluang untuk mengetahui tingkat keragaman genetik pada tingkat DNA yang dapat digunakan untuk mengetahui potensi genetik suatu ternak. Teknologi DNA dapat menjadi dasar untuk penentuan genotipe gen-gen yang bernilai ekonomis yang diperlukan sebagai bibit yang unggul. Data produksi dan molekuler ternak kambing masih terbatas sehingga hal ini menjadi tantangan di teknologi molekuler untuk mengeksplorasi.
2 protein, lipid dan karbohidrat. Pendeteksian gen GH pada ternak kambing penting dilakukan untuk mengetahui keragaman gen tersebut karena diduga terkait dengan sifat-sifat yang bernilai ekonomis dapat dijadikan sebagai penciri genetik.
Keragaman gen dapat diidentifikasi dengan dua metode yaitu metode
restriction fragment length polymorphism (RFLP) dan metode single-strand conformation polymorphism (SSCP). Kedua teknik tersebut dapat dimanfaatkan untuk mengidentifikasi karakteristik gen-gen yang penting untuk pertumbuhan pada ternak. Teknik PCR-SSCP merupakan teknik yang mudah dan efisien untuk mengidentifikasi variasi urutan nukleotida pada fragmen gen DNA. Keterbatasan terhadap informasi keragaman genentik kambing sehingga teknik ini digunakan untuk mengidentifikasi gen GH pada ternak kambing.
Tujuan
3 TINJAUAN PUSTAKA
Klasifikasi Kambing
Kambing diklasifikasikam ke dalam kingdom Animalia; phylum Chordata; subphylum Vertebrata; class Mammalia; ordo Artiodactyla; sub-ordo Ruminantia;
family Bovidae; sub family Caprinae dan genus Capra (Mileski dan Myers 2004). Kambing memiliki 60 kromosom (30 pasang kromosom) yang terdiri atas 29 pasang kromosom autosom dan sepasang kromosom kelamin (Gall, 1981).
Kambing merupakan hewan yang didomestifikasi oleh manusia untuk produksi daging, susu dan kulit. Penyebaran kambing sangat luas dan hampir menyebar di seluruh dunia. Hal ini disebabkan daya adaptasi yang baik dari kambing terhadap berbagai iklim dan kemampuan bertahan hidup pada daerah dengan hijauan terbatas (Gall, 1981). Kambing adalah hewan ternak yang sanggup hidup di daerah kering dan pakan hijauan pakan yang terbatas, serta mampu memanfaatkan hijauan pakan secara efisien (Devendra dan Burns, 1994). Kambing dapat dikelompokkan berdasarkan kegunaannya, yaitu kambing penghasil daging, susu, dan bulu (mohair). Di samping itu, ada pula beberapa bangsa kambing yang tergolong tipe dwiguna (dual purpose), seperti bangsa kambing PE yang tergolong tipe daging dan susu (Heriyadi, 2004).
Kambing Lokal Indonesia
Kambing lokal merupakan kambing asli yang berasal dari Indonesia. Kambing lokal ini termasuk dalam kambing tipe pedaging. Kambing lokal Indonesia terdiri dari kambing Kacang, kambing Marica, kambing Samosir, kambing Muara, kambing Kosta, kambing Gembrong, kambing Peranakan Etawah (PE).
Kambing Kacang memiliki ciri bulu pendek dan berwarna tunggal (putih, hitam dan cokelat). Tanduk berbentuk pedang lengkung ke atas dan ke belakang. Pada umumnya telinga kambing pendek dan tegak. Kambing Kacang memiliki leher yang pendek dan punggungnya melengkung sedikit lebih tinggi dari pada bahunya. (Devendra dan Burns, 1983). Kambing jantan dan betina dewasa memiliki bobot kurang lebih 25 dan 20 kg (Devendra dan Burns, 1983; Pamungkas et al., 2009).