TINGKAT SUPEROVULASI PADA BEBERAPA BANGSA SAPI
DENGAN SUMBER FOLLICLE STIMULATING HORMONE
(FSH) YANG BERBEDA
SKRIPSI
DHEDY PRASETYO
DEPARTEMEN ILMU PRODUKSI DAN TEKNOLOGI PETERNAKAN FAKULTAS PETERNAKAN
RINGKASAN
DHEDY PRASETYO D14054326. Tingkat Superovulasi pada Beberapa Bangsa Sapi dengan Sumber Follicle Stimulating Hormone (FSH) yang Berbeda. Skripsi. Departemen Ilmu Produksi dan Teknologi Peternakan, Fakultas Peternakan, Institut Pertanian Bogor.
Pembimbing Utama : Prof. Dr. Ir. Cece Sumantri, M.Agr.Sc. Pembimbing Anggota : Muhammad Imron, S.Pt. M.Si.
Permintaan daging dan susu sapi akan terus meningkat seiring dengan pertumbuhan penduduk. Namun peningkatan tersebut tidak sebanding dengan perkembangan populasi sapi potong dan sapi perah. Upaya peningkatan populasi ternak khususnya ternak sapi dapat dilakukan dengan mengembangkan bioteknologi di bidang peternakan yang salah satunya adalah sistem transfer embrio. Penelitian ini bertujuan untuk mempelajari pengaruh pemberian variasi sumber Follicle Stimulating Hormone (FSH) terhadap tingkat superovulasi pada beberapa bangsa sapi yang meliputi Response Rate, total Corpus Luteum (CL), total embrio dan ovum terkoleksi serta Recovery Rate.
Penelitian ini telah dilaksanakan selama tiga bulan, yaitu mulai bulan April 2011 hingga bulan Juni 2011. Penelitian dilaksanakan di Laboratorium Balai Embrio Ternak (BET) yang terletak di Desa Cipelang, Kecamatan Cijeruk, Kabupaten Bogor. Penelitian ini dilakukan dengan menggunakan data sekunder produksi embrio yang diperoleh dari BET Cipelang. Data tersebut berupa catatan produksi embrio secara in vivo selama tahun 2009 sampai tahun 2010. Data tersebut meliputi semen yang digunakan, FSH yang digunakan dalam superovulasi, jumlah CL, jumlah embrio grade A, B, C, D, dan ovum tidak dibuahi atau Unfertilized (UF). Ternak sapi donor yang digunakan dalam penelitian ini sebanyak 90 ekor sapi, terdiri atas 29 ekor sapi Friesian Holstein (FH), 23 ekor sapi Simmental, 27 ekor sapi Limmousin dan 11 sapi Angus. Data yang didapatkan diolah dengan metode Rancangan Acak Lengkap (RAL) pola faktorial 3x4 dengan 2 faktor yaitu sumber FSH (Folltropin-V, Opti-Stim dan Ovagen) dan bangsa sapi (FH, Simmental, Limousin dan Angus).
Berdasarkan hasil penelitian menunjukkan bahwa bangsa sapi memberikan pengaruh nyata (p<0,05) terhadap Response Rate, total CL dan total embrio dan ovum terkoleksi serta Recovery Rate. Sumber FSH tidak berpengaruh nyata pada Response Rate, total CL, rasio dari CL, total embrio dan Recovery Rate.
ii ABSTRACT
Superovulation Level in Some Cattle Breeds with Different Source of Follicle Stimulating Hormone (FSH)
Prasetyo, D., C. Sumantri, and M. Imron
Demand for beef and milk will continue to increase along with population growth. But the increases of demand are not proportional to the population growth of beef and dairy cattle. Attempt to improve livestock population especially cattle population can be done by developing biotechnology of animal science such embryo transfer system. This study aimed to evaluate the effects of variations source Follicle Stimulating Hormone (FSH) to superovulation in cattle which includes Response Rate, total of Corpus Luteum (CL), CL ratio, total of embryos and Recovery Rate.
This research has been carried out for three months, from April 2011 to June 2011. Research conducted at the Balai Embrio Ternak (BET) Laboratory located in the Cipelang Village, Cijeruk district, Bogor. The research was conducted using data of embryos production obtained from BET Cipelang. The data contains in vivo embryo production during 2009 to 2010. The data include semen used, FSH used in superovulation, total of CL, total of embryos grade A, B, C, D, and Unfertilized (UF). Ninety cows were used in the research, consisting of twenty-nine Holstein Friesian (HF), twenty-three Simmental, twenty-seven Limousin and eleven Angus. The data obtained is processed by the method of Randomized Block Design (RBD) factorial 3x5 pattern with 2 factors consist FSH source (Folltropin-V, Opti-Stim and Ovagen) and cattle breeds (HF, Simmental, Limousin and Angus).
Based on the research, data showed that breeds of cattle gives significantly effect (p<0,05) to the Response Rate, Recovery Rate, total of CL and total of embryo
and ovum collected. The source of FSH didn’t gives significantly effect on Response Rate, total of CL, ratio of CL, total of embryos and Recovery Rate.
iii
TINGKAT SUPEROVULASI PADA BEBERAPA BANGSA SAPI
DENGAN SUMBER FOLLICLE STIMULATING HORMONE
(FSH) YANG BERBEDA
DHEDY PRASETYO
D14054326
Skripsi ini merupakan salah satu syarat untuk memperoleh Sarjana Peternakan pada
Fakultas Peternakan Institut Pertanian Bogor
DEPARTEMEN ILMU PRODUKSI DAN TEKNOLOGI PETERNAKAN FAKULTAS PETERNAKAN
v RIWAYAT HIDUP
Penulis dilahirkan pada tanggal 29 Mei 1987 di Gresik, Jawa Timur. Penulis adalah anak pertama dari pasangan Bapak Mat Sari dan Ibu Suciyanti. Pendidikan di Taman Kanak-kanak diselesaikan pada tahun 1993 di TK Dharma Wanita Petrokimia Gresik. Pendidikan dasar diselesaikan pada tahun 1999 di SDN 1 Kebomas, Gresik. Pendidikan lanjutan menengah pertama diselesaikan pada tahun 2002 di SMPN 4 Gresik dan pendidikan lanjutan menengah atas diselesaikan pada tahun 2005 di SMAN 1 Gresik, Jawa Timur.
vi KATA PENGANTAR
Puji syukur Penulis panjatkan kehadirat Allah SWT atas limpahan, hidayah dan karunia-Nya sehingga Penulis dapat menyelesaikan studi hingga tugas akhir penyusunan skripsi ini. Shalawat dan salam semoga senantiasa tercurahkan kepada Nabi Muhammad SAW.
Skripsi dengan judul “Tingkat Superovulasi pada Beberapa Bangsa Sapi dengan Sumber Follicle Stimulating Homone (FSH) yang Berbeda” ini disusun
sebagai salah satu syarat untuk memperoleh gelar Sarjana Peternakan, Fakultas Peternakan, Institut Pertanian Bogor. Penelitian ini bertujuan untuk meneliti pengaruh pemberian variasi sumber FSH terhadap tingkat superovulasi pada berbagai bangsa sapi yang meliputi respon superovulasi, jumlah CL, jumlah total embrio dan ovum terkoleksi dan Recovery Rate, yang dapat dijadikan sebagai acuan pemberian FSH yang tepat dalam program superovulasi.
Penulis menyadari sepenuhnya bahwa dalam penulisan skripsi ini masih jauh dari kata sempurna, oleh sebab itu Penulis memohon maaf apabila terdapat banyak kekurangan. Ucapan terima kasih tidak lupa Penulis sampaikan kepada semua pihak yang turut membantu penyusunan skripsi ini, hanya Allah Yang Maha Pemurah dan Penyayang yang akan membalasnya. Semoga skripsi ini dapat bermanfaat bagi dunia pendidikan dan memberikan sumbangan yang berarti bagi kemajuan dunia peternakan di Indonesia. Amin.
Bogor, Januari 2012
DAFTAR ISI
Follicle Stimulating Hormone (FSH) ... 5
Seleksi Betina Donor ... 6
Keadaan Umum Lokasi Penelitian ... 20
viii
Tingkat Ovulasi ... 21
Produksi Embrio ... 23
Recovery Rate ... 25
KESIMPULAN DAN SARAN ... 27
Kesimpulan ... 27
Saran ... 27
UCAPAN TERIMAKASIH ... 28
DAFTAR PUSTAKA ... 29
DAFTAR TABEL
Nomor Halaman
1 Tahap Perkembangan Embrio ...11
2 Kualitas Embrio ...13
3 Respon Sapi Terhadap Superovulasi ...21
4 Persentase Corpus Luteum Hasil Superovulasi ...22
5 Rataan Corpus Luteum Hasil Superovulasi ...23
6 Rataan Jumlah Embrio dan Ovum Terkoleksi ...24
2 DAFTAR GAMBAR
Nomor Halaman
1. Tahap Perkembangan Embrio...……….. 10
2. (a) FH (b) Simmental (c) Limousin (d) Angus...……….. 15
3. (a) Folltropin-V (b) Ovagen (c) Optistim... 16
3 DAFTAR LAMPIRAN
Nomor Halaman
1. Analisis Sidik Ragam Respon Sapi Terhadap Superovulasi... 33 2. Uji t-Student Jumlah Corpus Luteum (CL) pada Ovarium Kanan dan
Kiri ………...
33
1 PENDAHULUAN
Latar Belakang
Permintaan daging dan susu sapi akan terus meningkat seiring dengan pertumbuhan penduduk dan peningkatan kesejahteran masyarakat. Kesadaran masyarakat akan pentingnya protein hewani juga menjadi penyebab peningkatan permintaan daging dan susu sapi. Namun peningkatan tersebut tidak sebanding dengan perkembangan populasi sapi potong dan sapi perah. Saat ini terdapat kecenderungan yang menunjukkan semakin lebarnya kesenjangan antara laju permintaan dan laju penawaran. Permasalahan utama di dalam upaya pemenuhan kebutuhan protein hewani nasional adalah ketidakmampuan sektor produksi domestik untuk mengimbangi laju pertumbuhan konsumsi.
Upaya peningkatan populasi ternak khususnya ternak sapi dapat dilakukan dengan mengembangkan bioteknologi di bidang peternakan yang salah satu diantaranya adalah sistem transfer embrio. Transfer embrio merupakan suatu metode perkawinan yang dilakukan dengan cara memproduksi banyak embrio pada seekor betina unggul, kemudian diimplantasikan pada banyak resipien sampai anak tersebut dilahirkan. Ternak sapi merupakan hewan monotokus, sehingga untuk memperoleh sejumlah embrio yang memadai untuk ditransfer ke resipien perlu dilakukan superovulasi pada sapi donor.
Superovulasi bertujuan memperbanyak oosit yang diovulasikan dengan menggunakan hormon gonadotropin eksogen seperti Follicle Stimulating Hormone (FSH) dengan cara penyuntikan hormon secara terus-menerus selama empat hari dengan dosis menurun. Pemberian hormon tersebut dengan dosis tertentu akan menstimulasi proses pertumbuhan, perkembangan, pematangan dan ovulasi dari sejumlah besar folikel pada ternak sapi.
Tujuan
2 Manfaat
3 TINJAUAN PUSTAKA
Siklus Estrus Sapi Betina
Sistem reproduksi sapi betina lebih kompleks daripada hewan jantan, karena terdiri atas beberapa organ yang memiliki peran dan fungsi masing- masing. Ovarium merupakan dua organ kecil yang terletak di ruang abdominal dengan fungsi utama adalah untuk menghasilkan ovum sekaligus sebagai tempat terjadinya proses oogenesis (proses produksi sel telur). Tugas lain dari ovarium adalah menghasilkan estrogen dan progesteron dimana kedua hormon ini memiliki peran penting dalam siklus reproduksi betina (Hafez dan Hafez, 2000). Partodiharjo (1982) menambahkan, ternak sapi bersifat poliestrus dan memperlihatkan berahi secara periodik sepanjang tahun. Estrus berasal dari kata latin oestrus yang dikenal dengan istilah berahi yaitu satu periode secara psikologis maupun fisiologis pada hewan betina yang bersedia menerima pejantan untuk kopulasi.
Toliehere (1985) menerangkan bahwa tanda-tanda berahi pada sapi adalah sapi betina menjadi tidak tenang, kurang nafsu makan, menguak, berkelana mencari pejantan, mencoba menaiki betina lain dan diam jika dinaiki sapi lain, selain itu vulva sapi tersebut terlihat membengkak, memerah, hangat dan penuh dengan lendir. Saat hewan betina mengalami estrus, serviks akan membuka sehingga sperma bisa masuk. Serviks berhubungan dengan vagina yang merupakan organ mirip pipa atau selongsong (sheath-like organ) dan berfungsi sebagai saluran kelahiran agar fetus dapat keluar dari uterus induk. Bagian paling luar dari saluran reproduksi betina adalah vulva yang sekaligus merupakan akhir dari saluran urinari (Herren, 2000).
Folikulogenesis
4 tumbuh dan berkembang sementara semua folikel subordinat lainnya mengalami atresia (Evans et al., 2004).
Menurut Fortune (1994) perkembangan folikel (folikulogenesis) dimulai dari proses rekrutmen, mekanisme seleksi dan akhirnya sampai pada satu titik folikel dihadapkan pada dua pilihan, terus berkembang kemudian berovulasi atau berhenti berkembang dan mengalami atresia atau mati Istilah rekrutmen identik dengan proses pertumbuhan folikel pada saat sebagian besar folikel lain mengalami atresia dan masing-masing folikel berusaha untuk mencapai tahap ovulasi. Rekrutmen tidak berlangsung secara acak, melainkan dalam kelompok folikel mereka akan terus berkembang apabila terjadi peningkatan konsentrasi Follicle Stimulating Hormone (FSH) dalam sistem sirkulasi.
Menurut Bo et al. (1995), perkembangan folikel di dalam ovarium merupakan proses yang berkesinambungan dan tidak hanya melibatkan satu folikel selama siklus, tetapi sekelompok folikel sehingga dianalogikan sebagai gelombang folikel. Gelombang folikel didefinisikan sebagai perkembangan folikel dengan diameter 4-5 mm dalam jumlah besar secara serentak yang diikuti dengan mekanisme seleksi, perkembangan menjadi folikel dominan dan penekanan atau supresi terhadap perkembangan folikel subordinat.
Proses perkembangan folikel hanya melibatkan beberapa folikel yang berkembang dan berhasil diovulasikan, sedangkan sebagian besar di antaranya akan mengalami atresia sebelum mencapai tahap ovulasi. Atresia dapat muncul pada setiap proses perkembangan folikel, namun frekuensi atresia tidak selalu terjadi dan tidak terdistribusi merata sepanjang proses folikulogenesis (Fortune, 1994). Triwulanningsih et al. (2001) menambahkan bahwa atresia dapat disebabkan degenerasi sel-sel kumuius, degenerasi oosit, peredaran darah yang memberi nutrisi ke oosit berkurang dan faktor penghambat dari folikel dominan terhadap folikel lainnya (folikel subordinat).
5 (supresi) terhadap folikel subordinat. Pada ternak, setiap gelombang terdiri atas kelompok folikel (15 folikel). Kemudian mereka berkompetisi (mekanisme seleksi) sehingga menghasilkan folikel dominan dan menekan perkembangan folikel lain (Bo et al., 1995). Folikulogenesis ovarium merupakan sistem yang kompleks dari morfologis dan peristiwa biokimia yang mengatur pertumbuhan serta diferensiasi dari folikel primordial ke tahap ovulasi (folikel ovulasi tunggal pada sapi) dan pelepasan oosit. Peningkatan folikulogenesis terkait dengan perubahan pada produksi dari faktor pertumbuhan dan hormon ovarium tetapi tidak dalam sekresi gonadotropin (Echternkamp, 2000).
Follicle Stimulating Hormon (FSH) dan Luteinizing Hormon (LH)
Gonadotropin adalah kelompok hormon yang bekerja pada gonad, misalnya FSH dan LH yang berperan dalam menginduksi perkembangan folikel ovari dan stimulasi ovulasi (Triwulanningsih et al., 2001). Eyestone dan Boer (1993) menjelaskan bahwa FSH berfungsi merangsang pertumbuhan folikel dalam ovari, proses pematangan Oosit dan perkembangan embrio secara dini, tetapi kurang berperan untuk perkembangan selanjutnya. Untuk meningkatkan pematangan folikel dalam jumlah besar (gelombang), diberikan perlakuan hormon gonadotropin selama fase luteal siklus estrus (Armstrong, 1993).
Toelihere (1985) menyatakan bahwa hormon utama yang digunakan pada superovulasi adalah hormon gonadotropin. yaitu FSH dan LH. FSH merupakan hormon gonadotropin dengan unsur glikopeptida yang memiliki reseptor pada sel granulosa folikel yang berfungsi menstimulasi pertumbuhan folikel, sehingga sangat diperlukan dalam proses superovulasi. Kaiin dan Tappa (2006) menambahkan hormon yang umum digunakan untuk menginduksi superovulasi pada sapi adalah Follicle Stimulating Hormone (FSH) yang berasal dari hipofisa. FSH merupakan hormon glikoprotein yang mempunyai waktu paruh pendek, sehingga memerlukan pemberian secara berulang untuk merangsang aktivitas folikel secara lebih efisien.
6 kualitas embrio yang kurang memenuhi klasifikasi yang telah ditentukan (Yusuf et al., 1993). Waktu paruh PMSG yang panjang menyebabkan terus terjadi stimulasi pembentukan folikel baru, meskipun ovulasi sudah selesai sehingga dari folikel yang terbentuk akan menghasilkan estrogen dengan kadar cukup tinggi yang pada akhirnya akan mengganggu transpor dan daya tahan hidup embrio (Mustofa, 1999). Baik FSH maupun PMSG telah banyak digunakan dalam teknik-teknik tertentu (misalnya produksi embrio) untuk menginduksi superovulasi (Hunter, 1995). Respon imunologi terhadap pemberian injeksi berulang hormon gonadotropin dapat membatasi kemampuan respon sapi donor terhadap superovulasi. PMSG dan FSH merupakan hormon protein sehingga sangat potensial menginduksi reaksi anafilaksis. Hal ini menandakan bahwa injeksi berulang dapat merangsang pembentukan anti- gonadotropin yang dapat mengurangi respon selanjutnya terhadap hormon gonadotropin endogen (Seidel dan Elsden, 1985).
Seleksi Betina Donor
7 Sinkronisasi Estrus
Sinkronisasi estrus adalah pencocokan siklus estrus dari ternak donor dan penerima dengan injeksi prostaglandin (PGF2) untuk merangsang estrus (Kunkel,
1998). Preparat Prostaglandin F2α (PGF2α) dikenal sebagai agen luteolitik yang dapat menyamakan siklus estrus dalam waktu yang bersamaan, sedangkan hCG dapat menginduksi ovulasi, sehingga pemberian hCG pada pertengahan estrus dapat merangsang pelepasan ovum dalam waktu yang lebih seragam. Dengan demikian perkembangan folikel dapat diamati sehingga dapat ditentukan waktu inseminasi yang lebih tepat (Arifiantini et al., 2010).
Superovulasi
Ovulasi adalah proses pemecahan folikel de graaf yang terjadi sewaktu ovum dilepaskan dari ovarium. Tingkatan ovarium adalah primer, sekunder dan tersier dan folikel de graaf. LH menyebabkan pengendoran dinding folikel sehingga lapisan-lapisan pecah dan melepaskan ovum dan cairan folikel, sesudah ovulasi terbentuk Corpus luteum di dalam folikel yang telah pecah dan mulai mensekresikan progesterone. Hewan-hewan betina dewasa yang disuntikan hormon gonadotropin dapat menghasilkan 20-100 ova pada satu estrus. FSH menggertak pematangan beberapa folikel, sedangkan LH menyebabkan ovulasi hal ini disebut superovulasi (Toelihere, 1985). Dalam program Transfer Embrio (TE), untuk merangsang ovulasi ganda (multiple ovulation), maka diberikan hormon superovulasi sehingga diperoleh 12-15 sel telur dalam satu kali ovulasi (Herren, 2000).
Superovulasi dapat diinduksi secara buatan melalui pemberian hormon gonadotropin eksogen (berasal dari luar tubuh), misalnya FSH dan PMSG. Pemberian hormon tersebut dengan dosis tertentu akan menstimulasi proses pertumbuhan, perkembangan, pematangan dan ovulasi dari sejumlah besar folikel pada ternak sapi. Betina donor diinjeksikan setiap hari dengan FSH (Herren, 2000) yang dapat berasal dari ekstrak hipofise babi dan domba (Wheeler dan Bowen, 1989) atau dari esktrak hipofise sapi (Wilson, 1992). Donor tertentu memerlukan penambahan LH selain FSH, namun umumnya preparat FSH yang dijual sudah ditambahkan LH (Wright, 1987).
8 agar siklus estrus berjalan normal dan teratur sehingga perlakuan superovulasi dapat sinkron dengan pola hormonal hewan secara normal. Jika siklus estrus abnormal, maka perlakuan superovulasi mungkin mengalami kegagalan Wilson (1992).
Inseminasi Buatan
Setelah berhasil memilih hewan donor berkualitas tinggi, kunci keberhasilan Transfer Embrio (TE) selanjutnya terletak pada inseminasi dengan semen yang berasal dari sapi jantan bibit unggul (Davis, 2004). Setelah perlakuan superovulasi, perlu dilakukan pengamatan terhadap tanda-tanda estrus pada sapi donor sehingga dapat dijadikan acuan untuk menentukan waktu inseminasi yang tepat dan IB dilakukan, penting untuk memeriksa berahi dan inseminasi 10 sampai 20 jam setelah awal berahi (Inseminasi dilakukan ketika sejumlah besar telur dilepaskan dari ovarium, ovulasi dapat terjadi selama 24 jam atau lebih). Pada saat telur terakhir dilepaskan dari ovarium mungkin tidak ada cukup sperma untuk membuahi. Sangat penting bahwa prosedur IB harus dilakukan secara tepat dan hanya dilakukan oleh teknisi IB yang sangat berpengalaman (Lewis, 1996).
9 dapat digunakan semen segar dengan konsentrasi 10-50 juta sperma motil atau semen beku yang mengandung 30 juta sperma motil dan diberikan dengan dosis ganda. Grimes (2008) menyarankan agar inseminasi dilakukan 1-3 kali selama dan setelah estrus dengan interval yang sama.
Koleksi Embrio
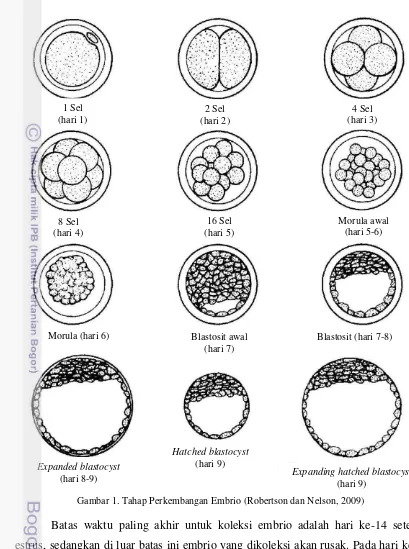
Panen atau koleksi embrio pada sapi donor dilakukan pada hari ke-7 sampai hari ke-8 setelah berahi saat sebagian besar embrio sudah memasuki ujung cornua uteri pada masa itu. Embrio akan berkembang sekitar satu minggu, kemudian embrio dipanen pada tahap morula sampai blastocyst (Grimes, 2008). Embrio dikoleksi antara hari ke-6 dan ke-8 setelah estrus (Herren, 2000). Pemanenan embrio tidak dilakukan lebih awal karena dapat menurunkan efisiensi koleksi embrio dengan metode non bedah. Sebelum hari ke-4, hampir semua embrio terletak di dalam oviduk yang dipisahkan dari uterus oleh utero-tubal junction. Struktur ini berfungsi sebagai katup (valve) yang dapat mengatur masuknya sperma dari uterus menuju oviduk sehingga fertilisasi terjadi tepat waktu dan mengatur transpor embrio ke arah sebaliknya. Embrio akan ditranspor menuju uterus pada hari ke-4 sampai hari ke-5 setelah estrus, melalui kontraksi ritmik pada dinding oviduk dan relaksasi dari otot pada dinding utero-tubal junction sehingga tingkat keberhasilan koleksi embrio akan lebih tinggi pada hari ke-6 dan seterusnya daripada hari ke-4. Kadang-kadang beberapa embrio masih ditemukan dalam oviduk pada sapi yang disuperovulasi sampai hari ke-10 (Seidel dan Elsden, 1989).
10
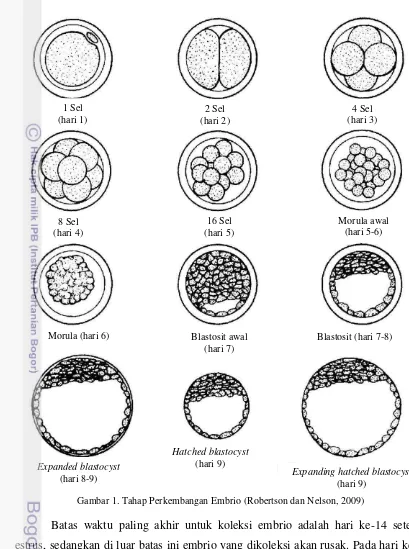
Gambar 1. Tahap Perkembangan Embrio (Robertson dan Nelson, 2009)
Batas waktu paling akhir untuk koleksi embrio adalah hari ke-14 setelah estrus, sedangkan di luar batas ini embrio yang dikoleksi akan rusak. Pada hari ke-9, embrio mulai keluar dari zona pellusida dan koleksi embrio tidak disarankan pada tahap ini karena kesulitan dalam mengidentifikasi embrio ketika ia sudah keluar dari zona pellusida. Hampir semua embrio telah keluar pada hari ke-11, pada saat diameter embrio meningkat secara drastis. Hari ke-12 sampai ke-13, embrio mulai memanjang (elongate) dan tampak seperti bola (American football). Hari ke-14 sampai ke-15, bentuk embrio sangat panjang seperti spaghetti. Hari ke-18 sampai
Morula (hari 6) Blastosit awal
11 hari ke-19, embrio hampir memenuhi cornua uteri. Koleksi embrio mungkin dapat dilakukan pada hari ke-17 dengan teknik non bedah, tetapi potensi terjadinya kerusakan/cacat pada embrio sangat besar sejak hari ke-14 (Seidel dan Elsden, 1989). Tahap perkembangan embrio dapat dilihat pada Tabel 1.
Tabel 1. Tahap Perkembangan Embrio Tahap Deskripsi
1 Belum dibuahi
2 2-12 sel
3 Morula awal
4 Morula
5 Blastosit awal
6 Blastosit
7 Expanded blastocyst
8 Hatched blastocyst
9 Expanded hatched blastocyst
Sumber: Wright (2009)
12
Corpus Luteum (CL)
Corpus Luteum (CL) merupakan benda yang terbentuk pada tempat ovum diovulasikan dan dijadikan patokan untuk mendeteksi berapa jumlah ovum yang diovulasikan oleh seekor sapi (Adriani et al., 2009). Setelah terjadi ovulasi maka pada situs pelepasan oosit akan terbentuk Corpus Luteum (CL). Selama awal fase luteal (metestrus). CL dibentuk dari sel-sel luteal. Pada pertengahan fase luteal (diestrus) sel-sel luteal menghasilkan sejumlah besar progesteron. Selama akhir fase luteal, CL dilisiskan PGF2α yang dihasilkan endometrium uterus. Lisis CL diikuti dengan penurunan kadar progesteron. sehingga mekanisme umpan balik negatif progesteron pada hypotalamus hilang, mengakibatkan peningkatan GNRH yang menandakan dimulai fase folikular. Ukuran CL pada hari ke 3-5 mulai meningkat sampai maksimal disertai dengan peningkatan produksi progesteron sampai kadar maksimal sekitar hari ke-10 (Senger, 1999).
Menurut Amiridis et al. (2006), CL tersusun atas sel-sel luteal yang berperan menghasilkan progesteron. Konsitensi atau kekenyalan badan CL sangat ditentukan jumlah sel-sel luteal dan vaskuralisasi darah kebagian tersebut. Demikian juga kemampuan CL memproduksi progesteron tergantung pada tingkat vaskularisasi pada lapisan seluler. Fungsi CL yang rendah (sintesis dan sekresi progesteron sedikit) diyakini akan menjadi penyebab penting kegagalan reproduksi dan ketidakmampuan uterus dalam mendukung perkembangan embrio dini. Senger (1999) menambahkan, dalam satu siklus estrus CL harus mengalami lisis agar fase folikular dimulai. Ovulasi tidak dapat terjadi dalam kondisi saat kadar progesteron dominan. Luteolisis berarti disintengrasi atau dekomposisi dari CL yang terjadi 2-3 hari pada akhir fase luteal. Dua hormon yang berperan penting dalam lisis CL yaitu oxytocin yang dihasilkan CL dan hormon PGF2α yang dihasilkan endometrium uterus.
Klasifikasi Embrio
13 ditransfer ke resipien yang tepat, dimana embrio dengan kualitas terbaik ditransfer ke respien yang paling baik pula (Seidel dan Elsden, 1985).
15 METODE
Lokasi dan Waktu
Penelitian ini dilaksanakan melalui magang selama tiga bulan, yaitu mulai bulan April 2011 hingga bulan Juni 2011. Penelitian dilaksanakan di Laboratorium Balai Embrio Ternak (BET) yang terletak di Desa Cipelang, Kecamatan Cijeruk, Kabupaten Bogor.
Materi
Penelitian ini dilakukan dengan menggunakan data sekunder produksi embrio yang diperoleh dari Balai Embrio Ternak (BET), Cipelang, Bogor. Data tersebut berupa catatan produksi embrio secara in vivo selama tahun 2009-2010. Data tersebut meliputi tanggal superovulasi, kode dan jenis ternak donor, kode semen yang digunakan, merk dagang hormon superovulasi yang digunakan, jumlah Corpus Luteum (CL), jumlah embrio grade A, B, C, dan D serta jumlah embrio layak transfer dan jumlah ovum tidak dibuahi atau Unfertilized (UF). Ternak sapi donor yang digunakan dalam penelitian ini sebanyak 90 ekor sapi, terdiri atas 29 ekor sapi FH, 23 ekor sapi Simmental, 27 ekor sapi Limmousin dan 11 ekor sapi Angus. Sebagian besar sapi disuperovulasi lebih dari sekali dengan jarak antar superovulasi sekitar tiga bulan sehingga didapatkan data superovulasi sebanyak 223 data. Data yang diperoleh kemudian dimasukkan dalam program Microsoft Access Database 2010. Pakan yang diberikan kepada sapi donor adalah rumput dan konsentrat. Gambaran sapi donor yang digunakan dapat dilihat pada Gambar 2.
16
(a) (b)
(c) (d)
Gambar 2. (a) FH, (b) Limousin, (c) Simmental, (d) Angus
(a) (b) (c)
17 Prosedur
Pengumpulan Data
Penelitian ini dilakukan dengan cara mengumpulkan dan menyeleksi data produksi embrio di BET yang dilakukan secara rutin. Program superovulasi di BET dilakukan dengan dua kali penyuntikan hormon FSH perhari yaitu pagi dan sore secara intra muscular pada hari ke-10 sampai hari ke-13 setelah estrus. Hormon Prostaglandin PGF2α diberikan tiga hari setelah awal pemberian hormon gonadotropin yang berfungsi untuk meregresikan corpus luteum, sehingga 2-3 hari setelah penyuntikan hormon PGF2α sapi akan berahi. Palpasi rektal pada hari ke-7 setelah IB, sehingga diperoleh data jumlah corpus luteum pada ovarium kiri dan kanan. Panen atau koleksi embrio pada sapi donor dilakukan pada hari ke-7 sampai hari ke-8 setelah berahi.
Peubah yang Diamati
Peubah yang diamati dari tingkat superovulasi ternak donor adalah sebagai berikut:
1) Response Rate, yaitu perbandingan jumlah ternak donor yang respon terhadap jumlah ternak yang disuperovulasi
ternak yang disupero ulasi (ekor) ternak donor respon (ekor)
2) Persentase corpus luteum pada ovarium kiri dan kanan; 3) Jumlah total corpus luteum;
4) Jumlah total embrio dan ovum terkoleksi;
5) Recovery Rate yaitu perbandingan jumlah embrio dan ovum terkoleksi terhadap jumlah corpus luteum.
embrio dan o um ( F) orpus luteum
Analisis Data
18
S12 = nilai varian kelompok pertama
S22 = nilai varian kelompok kedua Rancangan Acak Lengkap (RAL) dengan perlakuan perbedaan bangsa terhadap response rate. Menurut Steel dan Torrie (1993) model matematika yang digunakan adalah :
Yij = µ + Bi + ij
Keterangan:
Yij = respon percobaan karena pengaruh perlakuan bangsa ke-i
μ = rataan umum hasil percobaan Bj = perlakuan bangsa ke-i
19 Rancangan Acak Lengkap (RAL) pola faktorial 3x4 dengan 2 faktor yaitu sumber FSH (Folltropin-V, Opti-Stim dan Ovagen) dan bangsa sapi (FH, Simmental, Limousin dan Angus) digunakan dalam penelitian ini untuk melihat parameter rataan jumlah corpus luteum, embrio dan ovum terkoleksi. Model matematikanya menurut Steel dan Torrie (1993) sebagai berikut:
i k μ i ( )i εi k
Keterangan :
Yijk = respon percobaan karena pengaruh perlakuan dosis FSH ke-i dan perlakuan
bangsa ke-j dan ulangan ke-k μ = rataan umum hasil percobaan αi = perlakuan sumber FSH ke-i
j = perlakuan bangsa ke-j
(α)ij = interaksi antar perlakuan sumber FSH ke-i dan perlakuan bangsa ke-j
εijk = pengaruh kesalahan percobaan perlakuan sumber FSH ke-i dan perlakuan
bangsa ke-j pada ulangan ke-k.
20 HASIL DAN PEMBAHASAN
Keadaan Umum Lokasi Penelitian
Penelitian ini dilaksanakan di Laboratorium Balai Embrio Ternak (BET) yang terletak di Desa Cipelang, Kecamatan Cijeruk, Kabupaten Bogor. Topografi lokasi ini berada di punggung sebelah timur gunung Salak dengan kemiringan 8-400 dan ketinggian 600-1,350m dpl. Lingkungan lokasi penelitian ini mempunyai temperatur 18-22°C, kelembaban 70-80% dan curah hujan 3,222 mm per tahun. Menurut Abidin (2006) lingkungan yang baik untuk sapi adalah mempunyai temperatur optimal dengan kisaran suhu 10-270C, curah hujan 800-1.500 mm pertahun, sehingga lokasi penelitian ini cocok untuk pertumbuhan dan reproduksi sapi. Gambaran lokasi penelitian dapat dilihat pada Gambar 4.
(a) (b)
Gambar 4. (a) Luar kandang, (b) Dalam kandang
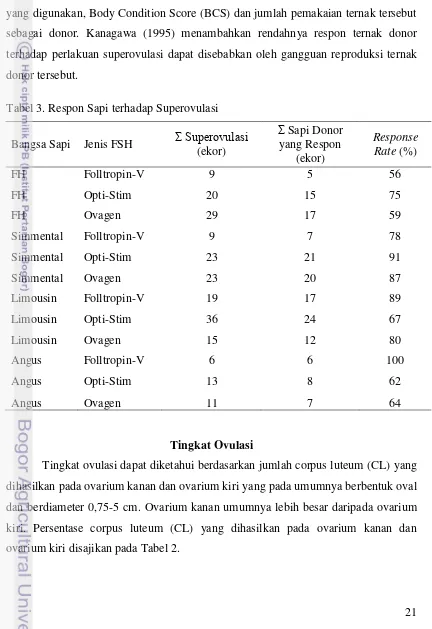
Respon Sapi terhadap Superovulasi
21 superovulasi yaitu sebesar 100%. Analisis sidik ragam respon sapi terhadap superovulasi dapat dilihat pada Lampiran 1.
Menurut Muawanah (2000) beberapa faktor yang mempengaruhi respon ternak donor terhadap superovulasi antara lain faktor umur ternak donor, dosis FSH yang digunakan, Body Condition Score (BCS) dan jumlah pemakaian ternak tersebut sebagai donor. Kanagawa (1995) menambahkan rendahnya respon ternak donor terhadap perlakuan superovulasi dapat disebabkan oleh gangguan reproduksi ternak donor tersebut.
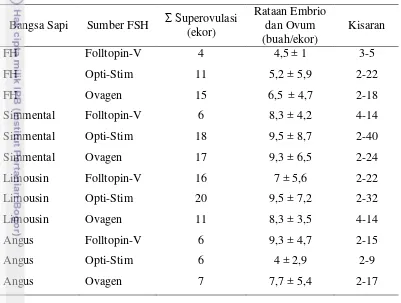
Tabel 3. Respon Sapi terhadap Superovulasi
Bangsa Sapi Jenis FSH upero ulasi
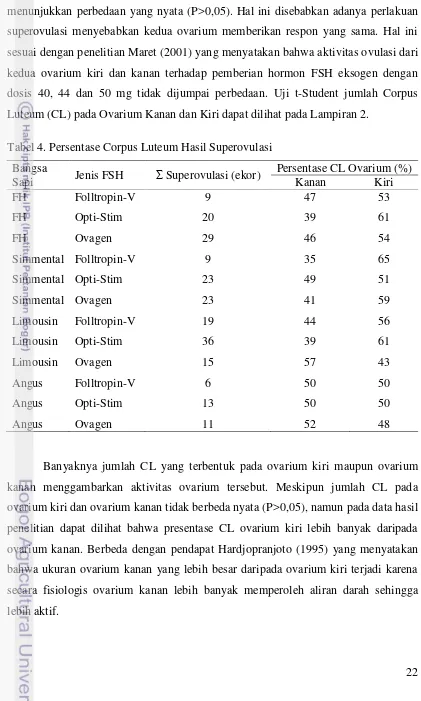
22 Berdasarkan uji statistik jumlah CL sebelah kanan dan kiri tidak menunjukkan perbedaan yang nyata (P>0,05). Hal ini disebabkan adanya perlakuan superovulasi menyebabkan kedua ovarium memberikan respon yang sama. Hal ini sesuai dengan penelitian Maret (2001) yang menyatakan bahwa aktivitas ovulasi dari kedua ovarium kiri dan kanan terhadap pemberian hormon FSH eksogen dengan dosis 40, 44 dan 50 mg tidak dijumpai perbedaan. Uji t-Student jumlah Corpus Luteum (CL) pada Ovarium Kanan dan Kiri dapat dilihat pada Lampiran 2.
Tabel 4. Persentase Corpus Luteum Hasil Superovulasi Bangsa
Sapi Jenis FSH Superovulasi (ekor)
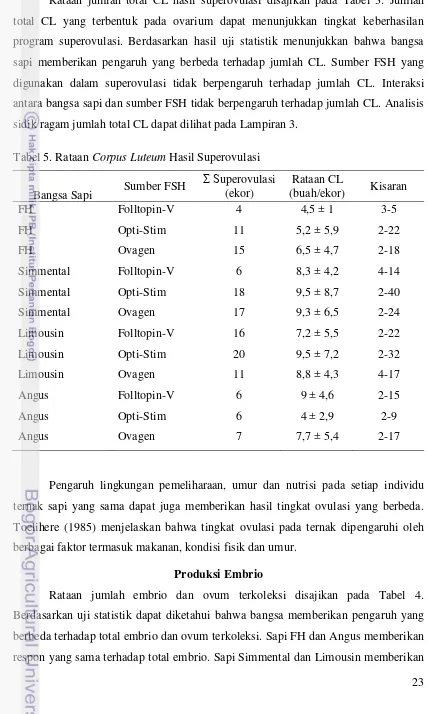
23 Rataan jumlah total CL hasil superovulasi disajikan pada Tabel 3. Jumlah total CL yang terbentuk pada ovarium dapat menunjukkan tingkat keberhasilan program superovulasi. Berdasarkan hasil uji statistik menunjukkan bahwa bangsa sapi memberikan pengaruh yang berbeda terhadap jumlah CL. Sumber FSH yang digunakan dalam superovulasi tidak berpengaruh terhadap jumlah CL. Interaksi antara bangsa sapi dan sumber FSH tidak berpengaruh terhadap jumlah CL. Analisis sidik ragam jumlah total CL dapat dilihat pada Lampiran 3.
Tabel 5. Rataan Corpus Luteum Hasil Superovulasi
Bangsa Sapi Sumber FSH
Pengaruh lingkungan pemeliharaan, umur dan nutrisi pada setiap individu ternak sapi yang sama dapat juga memberikan hasil tingkat ovulasi yang berbeda. Toelihere (1985) menjelaskan bahwa tingkat ovulasi pada ternak dipengaruhi oleh berbagai faktor termasuk makanan, kondisi fisik dan umur.
Produksi Embrio
24 respon yang sama terhadap total embrio dan ovum terkoleksi. Sedangkan sapi FH dengan Simmental memberikan respon yang berbeda terhadap total embrio dan ovum terkoleksi. Sumber FSH tidak berpengaruh terhadap total embrio dan ovum terkoleksi. Analisis sidik ragam jumlah embrio dan ovum terkoleksi dapat dilihat pada Lampiran 4.
Tabel 6. Rataan Jumlah Embrio dan Ovum Terkoleksi Hasil Superovulasi
Bangsa Sapi Sumber FSH uperovulasi
Faktor-faktor seperti sumber dan kondisi sperma, kualitas oosit yang diperoleh, kondisi alat reproduksi sapi betina, nutrisi pakan, ketrampilan inseminator, lingkungan pemeliharaan dan jadwal pengkoleksian embrio yang tepat dapat juga mempengaruhi pembuahan dan perkembangan ovum. Seidel dan Elsden (1989) menjelaskan bahwa Koleksi dengan metode tanpa pembedahan melalui serviks dilakukan pada hari ke-7 atau ke-8 setelah estrus, koleksi pada hari ke-7 akan menghasilkan embrio stadium kompak morula dan blatosit awal sedangkan pada hari ke-8 embrio mencapai stadium blatosit penuh.
25 hilang, tidak dibuahi atau tidak terkembang. Lebih lanjut lagi dikatakan bahwa koleksi embrio tanpa pembedahan memungkinkan adanya sekitar 10% embrio yang tidak berhasil dibilas karena masih berada di oviduk.
Recovery Rate
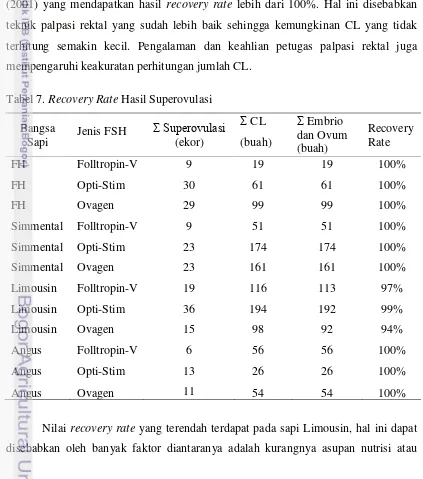
Respon sapi terhadap superovulasi, yang ditandai dengan jumlah CL berkorelasi positif dengan jumlah embrio yang dihasilkan. Berdasarkan Tabel 5 menunjukkan nilai recovery rate yang berada pada kisaran 80%. Hanya ada satu nilai recovery rate yang lebih dari 100% yaitu pada sapi Angus yang disuperovulasi dengan Folltropin-V. Hasil ini berbeda dengan penelitian Suradi (2004) dan Maret (2001) yang mendapatkan hasil recovery rate lebih dari 100%. Hal ini disebabkan teknik palpasi rektal yang sudah lebih baik sehingga kemungkinan CL yang tidak terhitung semakin kecil. Pengalaman dan keahlian petugas palpasi rektal juga mempengaruhi keakuratan perhitungan jumlah CL.
Tabel 7. Recovery Rate Hasil Superovulasi
Bangsa
27 KESIMPULAN DAN SARAN
Kesimpulan
Perlakuan pemberian sumber FSH yang berbeda tidak menunjukkan pengaruh yang nyata terhadap tingkat superovulasi. Sapi yang disuperovulasi dengan hormon Folltropin-V menghasilkan rataan jumlah CL, embrio dan ovum yang lebih tinggi daripada sapi yang disuperovulasi dengan hormon lainnya. Bangsa sapi berpengaruh nyata terhadap nilai response rate,recovery rate, rataan jumlah CL dan rataan jumlah embrio. Sapi yang menghasilkan jumlah total CL, jumlah embrio dan ovum terkoleksi adalah Simmental. Hal ini diduga karena daya adaptasi sapi Simmental terhadap iklim dan lingkungan lebih baik dibanding sapi lainnya.
Saran
28 UCAPAN TERIMAKASIH
Puji syukur penulis panjatkan kehadirat Allah SWT atas limpahan, hidayah dan karunia-Nya sehingga Penulis dapat menyelesaikan studi hingga tugas akhir penelitian dan penyusunan skripsi ini. Shalawat dan salam semoga senantiasa tercurahkan kepada Nabi Muhammad SAW. Penulis menyampaikan terimakasih sebesar-besarnya kepada Prof. Dr. Ir. Cece Sumantri, M.Agr.Sc. sebagai Dosen Pembimbing Utama dan Muhammad Imron, S.Pt. M.Si. sebagai Pembimbing Anggota yang banyak memberikan masukan, saran dan pengarahan sehingga penulis dapat menyelesaikan skripsi ini. Terima kasih kepada Ir. Rini Herlina Mulyono, M.Si. dan Dr. Despal, S.Pt. M.Agr.Sc sebagai dosen penguji ujian lisan yang memberikan banyak masukan dan koreksi terhadap skripsi ini. Terima kasih kepada Dr. Ir. Henny Nuraini, M.Si. selaku Pembimbing Akademik yang telah memberi pengarahan mulai awal hingga akhir perkuliahan. Terimakasih pula penulis ucapkan kepada seluruh staf pengajar Fakultas Peternakan yang telah membagi ilmu pengetahuan dan pengalaman selama menyelesaikan pendidikan di Fakultas Peternakan IPB.
Tidak lupa penulis mengucapkan terimakasih kepada Ibunda Suciyanti dan Ayahanda Mat Sari selaku orang tua penulis atas dukungan, doa, kasih sayang, bantuan moril dan materil yang selalu diberikan hingga penulis dapat menyelesaikan tugas akhir. Terimakasih kepada adinda Dwi Fatmala Sari dan Gala Prima Dana atas motivasi yang diberikan. Terimakasih kepada Kepala Balai Embrio Ternak (BET) Cipelang dan staff (Bu Lela dan Pak Darlin) serta teman penelitian Aidil Marsan yang banyak membantu penelitian. Terimakasih penulis ucapkan kepada Mayagita Yunidar yang telah memberikan inspirasi dan dukungan dalam menyelesaikan tugas akhir. Terimakasih kepada teman-teman kost Wisma Wijayakusuma (Adit, Iwan, Panda dan Dadang). Terimakasih penulis ucapkan juga kepada Fachri, Erli, Hida, Revan, Lia, Andwie dan seluruh teman-teman IPTP 42 yang tidak dapat disebutkan satu per satu atas persahabatannya. Semoga skripsi ini dapat bermanfaat.
Bogor, Januari 2012
29 DAFTAR PUSTAKA
Abidin, Z. 2006. Penggemukan Sapi Potong. Agromedia Pustaka, Tangerang.
Adriani, B. Rosidi & Depison. 2009. Penggunaan follicle stimulating hormone dan pregnant mare serum gonadotropin untuk superovulasi pada sapi persilangan brahman. Med. Pet. 32: 163-170.
Amiridis, G. S., T. Tsiligianni & N. C. Rawling. 1994. Follicular waves and circulating gonadotropins in 8-month-old prepubertal heifer. J. Reprod. Fertil. 100 : 27-33.
Arifiantini, R. I., B. Purwatara, T. L. Yusuf, D. Sajuthi & Amrozi. 2010. Angka konsepsi hasil inseminasi semen cair versus semen beku pada kuda yang disinkronisasi estrus dan ovulasi. Med. Pet. 33: 1-5.
Armstrong, D.T. 1993. Recent advances in superovulation of cattle. Theriogenology 39: 7-24.
Betteridge, K. J. 1980. Producere and Result Obtainable in Cattle. In: A. D. Morrow (Ed.). Current Theraphy in Theriogenology. 2nded. W. B. Saunders and Co., Philadelphia.
Bo, G. A., G. P. Adams, R. A. Pierson & P. J. Mapietoft. 1995. Exogenous control of follicular wave emergence in cattle.Theriogenology 43: 31-40.
Davis, R. L. 2004. Embryo transfer in beef cattle. http://www.davis-rairdan.com/embryo-transfer.htm.[3 April 2011].
Echternkamp, S. E. 2000. Endocrinology of Increased Ovarian Folliculogenesis in Cattle Selected for Twin Births. USDA, Nebraska.
Evans, E. C. O., J. L. H. Ireland, M. E. Winn, P. Lonergan, G. W. Smith, P. M. Coussens & J. J. Ireland. 2004. Identification of genes involved apoptosis and dominant follicle development during follicular waves in cattle. J. Biol. Repr. 70: 1475-1484.
Eyestone, W. H. & H. A. Boer. 1993. FSH enhance development potential at bovine oocytes mature in chemically defined medium. Theriogenology 39: 216. Fortune, J. E. 1994. Ovarian follicular growth and development in mammals. J. Biol.
Repr. 50: 225-232.
Greve, T., H. Callesen, P. Hyttel, R. Hoier & R. Assey. 1995. Effects of exogenous gonadotropins on oocyte and embryo quality in cattle. Theriogenology 43: 41-50.
Grimes, J. F. 2008. Utilization of embryo transfer in beef cattle. http://ohioline. osu.edu/anr-fact/pdf/ANR_17_08.pdf. [12 Desember 2011].
Hafez, E. S. E. & B. Hafez. 2000. Reproduction in Farm Animal. 7th ed. Lippincott Williams & Wilkins, Piladelphia.
30 Herren, R. 2000. The Science of Animal Agriculture. 2nd ed. Delmar Thomson
Learning, Albany.
Hunter, R. H. F. 1995.Fisiologi dan Teknologi Reproduksi Hewan Betina Domestik. Penerbit ITB, Bandung.
Kaiin, E. M. & B. Tappa. 2006. Induksi superovulasi dengan kombinasi CIDR, hormon FSH dan hCG pada induk sapi potong. Med. Pet. 29: 141-146.
Kaiin, E. M., S. Said & B. Tappa. 2008. Kelahiran anak sapi hasil fertilisasi secara in vitro dengan sperma hasil pemisahan. Med. Pet. 31: 22-28.
Kanagawa, H., I. Shimamora & N. Saito. 1995. Manual of Bovine Embryo Transfer. Japan Livestock Technology Association, Tokyo.
Kunkel, J. R. 1998. Embryo transfer. www.wvu.edu/agexten/forglvst/Dairy/dirm26 .pdf. [12 Desember 2011].
Lewis, I. 1996. Conventional Embryo Transfer. In: I. Lewis, J. Owens, S. McClintoch dan M. Trevean (Eds.). Cattle Breeding Technology. Genetics Australia, Australia.
Maret, D. 2001. Pengaruh dosis follicle stimulating hormone (FSH) dan body condition score (BCS) terhadap superovulasi sapi perah fries holstein (FH). Skripsi. Fakultas Peternakan, Institut Pertanian Bogor, Bogor.
Muawanah. 2000. Superovulasi pada sapi perah fries holland (FH) dengan pemberian dosis FSH yang berbeda. Skripsi. Fakultas Peternakan, Institut Pertanian Bogor, Bogor.
Mustofa, I. 1999. Pengaruh pengunduran waktu penyuntikan hCG terhadap hasil superovulasi pada sapi perah. J. Med. Vet. 15: 242-247.
Partodiharjo, S. 1982. Ilmu Reproduksi Hewan. Mutiara, Jakarta.
Robertson, I. & R. E. Nelson. 2009. Certification and Identification of Embryos. In: D. A. Stringfellow & M. D. Givens (Eds.). Manuals of the International Embryo Transfer Society. 4th ed. International Embryo Transfer Society, Illionis.
Seidel, G. E. Jr. 1981. Superovulation and embryo transfer in cattle. Sci. 211: 351-358.
Seidel, G. E. & R. P. Elsden. 1985.Procedures for Recovery, Bisection, Freezing and Transfer of Bovine Embryos. Colorado State Univ, Colorado.
Seidel, G. E. & R. P. Elsden. 1989.Embryo Transfer in Dairy Cattle. WD Hoard & Sons, Colorado.
Senger, P. L. 1999. Pathways to Pregnancy and Parturition. Current Concept Inc., Washington.
Steel, R. G. D. & R. A. Torrie. 1993. Prinsip Prosedur Statistika. Terjemahan: B. Sumantri. Edisi ke-2. PT Gramedia Pustaka Utama, Jakarta.
31 Toelihere, M. R. 1985. Fisiologi Reproduksi pada Ternak. Penerbit Angkasa,
Bandung.
Triwulanningsih, E., M. R. Toelihere, J. J. Rutledge, T. L. Yusuf, B. Purwantara & K. Djuyanto. 2001. Produksi embrio in vitro dengan modifikasi waktu dan hormon gonadotropin selama pematangan oosit. JITV 6: 179.
Wheeler, M. B. & R. A. Bowen. 1989. Endocrinology and Superovulation. In: Bovine Embryo Transfer : 1989 Short Course Proceedings. Colorado State Univ, Colorado.
Wilson, R. 1992. Embryo Transfer in Cattle. http://www.cruachan.com.au/embryo_ transfer.htm. [3 April 2011].
Wright, J. M. 2009. Photographic Illustrations of Embryo Developmental Stage and Quality Codes. In: D. A. Stringfellow & M. D. Givens (Eds.). Manuals of the International Embryo Transfer Society. 4th ed. International Embryo Transfer Society, Illionis.
Wright, R. 1987. Present Status of and Prospects for Embryo Transfer in the United States. In: Technical Meeting on Embryo Transfer and Animal Production. National Academy Press, Washington DC.
33 Lampiran 1. Analisis Sidik Ragam Respon Sapi Terhadap Superovulasi
SK DB JK KT F Hitung P
Pada analisis sidik ragam di atas, kesimpulan yang diperoleh sebagai berikut: 1. Pengaruh bangsa
H0: α1 α2 … α4=0 H1: minimal ada satu i dimana αi≠
P-value (0,008) α ( , 1) maka terima H0. Hal ini menunjukkan bahwa
bangsa sangat berpengaruh terhadap response rate. Untuk mengetahui bangsa mana yang memberi pengaruh yang berbeda terhadap response rate maka perlu melakukan uji lanjut Duncan.
Duncana,,b,,c
34 3. Pengaruh interaksi antara bangsa dan hormon
H0: (αβ)11 (αβ)12 …. (αβ)43=0 H1: minimal ada satu (αβ)ij≠
P-value (0,363) > α ( , 5) maka terima H0. Hal ini menunjukkan bahwa
interaksi antara bangsa dan hormon tidak berpengaruh terhadap response rate sehingga tidak perlu uji lanjut Duncan.
Lampiran 2. Uji t-Student Jumlah Corpus Luteum (CL) pada Ovarium Kanan dan Kiri
Lampiran 3. Analisis Sidik Ragam Jumlah Corpus Luteum Hasil Superovulasi
SK DB JK KT F Hitung P
Pada analisis sidik ragam di atas, kesimpulan yang diperoleh sebagai berikut: 1. Pengaruh bangsa
H0: α1 α2 … α4=0 H1: minimal ada satu i dimana αi≠
P-value (0,032) < α ( , 5) maka tolak H0. Hal ini menunjukkan bahwa
35
hormon tidak berpengaruh terhadap jumlah CL sehingga tidak perlu uji lanjut Duncan.
3. Pengaruh interaksi antara bangsa dan hormon
H0 : (αβ)11 (αβ)12 …. (αβ)43=0 H1: minimal ada satu (αβ)ij≠
P-value (0,372) > α ( ,05) maka terima H0. Hal ini menunjukkan bahwa
interaksi antara bangsa dan hormon tidak berpengaruh terhadap jumlah CL.
Lampiran 4. Analisis Sidik Ragam Jumlah Embrio dan Ovum Hasil Superovulasi
SK DB JK KT F Hitung P
Pada analisis sidik ragam di atas, kesimpulan yang diperoleh sebagai berikut: 1. Pengaruh bangsa
H0: α1 α2 … α4=0 H1: minimal ada satu i dimana αi≠
P-value (0,041) < α ( , 5) maka tolak H0. Hal ini menunjukkan bahwa
36
hormon tidak berpengaruh terhadap jumlah embrio dan ovum terkoleksi sehingga tidak perlu uji lanjut Duncan.
3. Pengaruh interaksi antara bangsa dan hormon
H0 : (αβ)11 (αβ)12 …. (αβ)43=0 H1: minimal ada satu (αβ)ij≠
P-value (0,305) > α ( ,05) maka terima H0. Hal ini menunjukkan bahwa
interaksi antara bangsa dan hormon tidak berpengaruh terhadap jumlah embrio dan ovum terkoleksi.
Lampiran 5. Analisis Sidik Ragam Recovery Rate
SK DB JK KT F Hitung P
Pada analisis sidik ragam di atas, kesimpulan yang diperoleh sebagai berikut: 1. Pengaruh bangsa
H0: α1 α2 … α4=0 H1: minimal ada satu i dimana αi≠
P-value (0,141) α ( , 5) maka terima H0. Hal ini menunjukkan bahwa bangsa tidak
37 2. Pengaruh hormon
H : β1 β2 β3=0 H1 : ada minimal satu dimana βj≠
P-value (0,853) > α ( , 5) maka terima H0. Hal ini menunjukkan bahwa hormon
tidak berpengaruh terhadap recovery rate sehingga tidak perlu uji lanjut Duncan. 3. Pengaruh interaksi antara bangsa dan hormon
H0: (αβ)11 (αβ)12 …. (αβ)43=0 H1: minimal ada satu (αβ)ij≠
P-value (0,972) > α ( , 5) maka terima H0. Hal ini menunjukkan bahwa interaksi
TINGKAT SUPEROVULASI PADA BEBERAPA BANGSA SAPI
DENGAN SUMBER FOLLICLE STIMULATING HORMONE
(FSH) YANG BERBEDA
SKRIPSI
DHEDY PRASETYO
DEPARTEMEN ILMU PRODUKSI DAN TEKNOLOGI PETERNAKAN FAKULTAS PETERNAKAN
ii ABSTRACT
Superovulation Level in Some Cattle Breeds with Different Source of Follicle Stimulating Hormone (FSH)
Prasetyo, D., C. Sumantri, and M. Imron
Demand for beef and milk will continue to increase along with population growth. But the increases of demand are not proportional to the population growth of beef and dairy cattle. Attempt to improve livestock population especially cattle population can be done by developing biotechnology of animal science such embryo transfer system. This study aimed to evaluate the effects of variations source Follicle Stimulating Hormone (FSH) to superovulation in cattle which includes Response Rate, total of Corpus Luteum (CL), CL ratio, total of embryos and Recovery Rate.
This research has been carried out for three months, from April 2011 to June 2011. Research conducted at the Balai Embrio Ternak (BET) Laboratory located in the Cipelang Village, Cijeruk district, Bogor. The research was conducted using data of embryos production obtained from BET Cipelang. The data contains in vivo embryo production during 2009 to 2010. The data include semen used, FSH used in superovulation, total of CL, total of embryos grade A, B, C, D, and Unfertilized (UF). Ninety cows were used in the research, consisting of twenty-nine Holstein Friesian (HF), twenty-three Simmental, twenty-seven Limousin and eleven Angus. The data obtained is processed by the method of Randomized Block Design (RBD) factorial 3x5 pattern with 2 factors consist FSH source (Folltropin-V, Opti-Stim and Ovagen) and cattle breeds (HF, Simmental, Limousin and Angus).
Based on the research, data showed that breeds of cattle gives significantly effect (p<0,05) to the Response Rate, Recovery Rate, total of CL and total of embryo
and ovum collected. The source of FSH didn’t gives significantly effect on Response Rate, total of CL, ratio of CL, total of embryos and Recovery Rate.
1 PENDAHULUAN
Latar Belakang
Permintaan daging dan susu sapi akan terus meningkat seiring dengan pertumbuhan penduduk dan peningkatan kesejahteran masyarakat. Kesadaran masyarakat akan pentingnya protein hewani juga menjadi penyebab peningkatan permintaan daging dan susu sapi. Namun peningkatan tersebut tidak sebanding dengan perkembangan populasi sapi potong dan sapi perah. Saat ini terdapat kecenderungan yang menunjukkan semakin lebarnya kesenjangan antara laju permintaan dan laju penawaran. Permasalahan utama di dalam upaya pemenuhan kebutuhan protein hewani nasional adalah ketidakmampuan sektor produksi domestik untuk mengimbangi laju pertumbuhan konsumsi.
Upaya peningkatan populasi ternak khususnya ternak sapi dapat dilakukan dengan mengembangkan bioteknologi di bidang peternakan yang salah satu diantaranya adalah sistem transfer embrio. Transfer embrio merupakan suatu metode perkawinan yang dilakukan dengan cara memproduksi banyak embrio pada seekor betina unggul, kemudian diimplantasikan pada banyak resipien sampai anak tersebut dilahirkan. Ternak sapi merupakan hewan monotokus, sehingga untuk memperoleh sejumlah embrio yang memadai untuk ditransfer ke resipien perlu dilakukan superovulasi pada sapi donor.
Superovulasi bertujuan memperbanyak oosit yang diovulasikan dengan menggunakan hormon gonadotropin eksogen seperti Follicle Stimulating Hormone (FSH) dengan cara penyuntikan hormon secara terus-menerus selama empat hari dengan dosis menurun. Pemberian hormon tersebut dengan dosis tertentu akan menstimulasi proses pertumbuhan, perkembangan, pematangan dan ovulasi dari sejumlah besar folikel pada ternak sapi.
Tujuan
2 Manfaat
3 TINJAUAN PUSTAKA
Siklus Estrus Sapi Betina
Sistem reproduksi sapi betina lebih kompleks daripada hewan jantan, karena terdiri atas beberapa organ yang memiliki peran dan fungsi masing- masing. Ovarium merupakan dua organ kecil yang terletak di ruang abdominal dengan fungsi utama adalah untuk menghasilkan ovum sekaligus sebagai tempat terjadinya proses oogenesis (proses produksi sel telur). Tugas lain dari ovarium adalah menghasilkan estrogen dan progesteron dimana kedua hormon ini memiliki peran penting dalam siklus reproduksi betina (Hafez dan Hafez, 2000). Partodiharjo (1982) menambahkan, ternak sapi bersifat poliestrus dan memperlihatkan berahi secara periodik sepanjang tahun. Estrus berasal dari kata latin oestrus yang dikenal dengan istilah berahi yaitu satu periode secara psikologis maupun fisiologis pada hewan betina yang bersedia menerima pejantan untuk kopulasi.
Toliehere (1985) menerangkan bahwa tanda-tanda berahi pada sapi adalah sapi betina menjadi tidak tenang, kurang nafsu makan, menguak, berkelana mencari pejantan, mencoba menaiki betina lain dan diam jika dinaiki sapi lain, selain itu vulva sapi tersebut terlihat membengkak, memerah, hangat dan penuh dengan lendir. Saat hewan betina mengalami estrus, serviks akan membuka sehingga sperma bisa masuk. Serviks berhubungan dengan vagina yang merupakan organ mirip pipa atau selongsong (sheath-like organ) dan berfungsi sebagai saluran kelahiran agar fetus dapat keluar dari uterus induk. Bagian paling luar dari saluran reproduksi betina adalah vulva yang sekaligus merupakan akhir dari saluran urinari (Herren, 2000).
Folikulogenesis
4 tumbuh dan berkembang sementara semua folikel subordinat lainnya mengalami atresia (Evans et al., 2004).
Menurut Fortune (1994) perkembangan folikel (folikulogenesis) dimulai dari proses rekrutmen, mekanisme seleksi dan akhirnya sampai pada satu titik folikel dihadapkan pada dua pilihan, terus berkembang kemudian berovulasi atau berhenti berkembang dan mengalami atresia atau mati Istilah rekrutmen identik dengan proses pertumbuhan folikel pada saat sebagian besar folikel lain mengalami atresia dan masing-masing folikel berusaha untuk mencapai tahap ovulasi. Rekrutmen tidak berlangsung secara acak, melainkan dalam kelompok folikel mereka akan terus berkembang apabila terjadi peningkatan konsentrasi Follicle Stimulating Hormone (FSH) dalam sistem sirkulasi.
Menurut Bo et al. (1995), perkembangan folikel di dalam ovarium merupakan proses yang berkesinambungan dan tidak hanya melibatkan satu folikel selama siklus, tetapi sekelompok folikel sehingga dianalogikan sebagai gelombang folikel. Gelombang folikel didefinisikan sebagai perkembangan folikel dengan diameter 4-5 mm dalam jumlah besar secara serentak yang diikuti dengan mekanisme seleksi, perkembangan menjadi folikel dominan dan penekanan atau supresi terhadap perkembangan folikel subordinat.
Proses perkembangan folikel hanya melibatkan beberapa folikel yang berkembang dan berhasil diovulasikan, sedangkan sebagian besar di antaranya akan mengalami atresia sebelum mencapai tahap ovulasi. Atresia dapat muncul pada setiap proses perkembangan folikel, namun frekuensi atresia tidak selalu terjadi dan tidak terdistribusi merata sepanjang proses folikulogenesis (Fortune, 1994). Triwulanningsih et al. (2001) menambahkan bahwa atresia dapat disebabkan degenerasi sel-sel kumuius, degenerasi oosit, peredaran darah yang memberi nutrisi ke oosit berkurang dan faktor penghambat dari folikel dominan terhadap folikel lainnya (folikel subordinat).
5 (supresi) terhadap folikel subordinat. Pada ternak, setiap gelombang terdiri atas kelompok folikel (15 folikel). Kemudian mereka berkompetisi (mekanisme seleksi) sehingga menghasilkan folikel dominan dan menekan perkembangan folikel lain (Bo et al., 1995). Folikulogenesis ovarium merupakan sistem yang kompleks dari morfologis dan peristiwa biokimia yang mengatur pertumbuhan serta diferensiasi dari folikel primordial ke tahap ovulasi (folikel ovulasi tunggal pada sapi) dan pelepasan oosit. Peningkatan folikulogenesis terkait dengan perubahan pada produksi dari faktor pertumbuhan dan hormon ovarium tetapi tidak dalam sekresi gonadotropin (Echternkamp, 2000).
Follicle Stimulating Hormon (FSH) dan Luteinizing Hormon (LH)
Gonadotropin adalah kelompok hormon yang bekerja pada gonad, misalnya FSH dan LH yang berperan dalam menginduksi perkembangan folikel ovari dan stimulasi ovulasi (Triwulanningsih et al., 2001). Eyestone dan Boer (1993) menjelaskan bahwa FSH berfungsi merangsang pertumbuhan folikel dalam ovari, proses pematangan Oosit dan perkembangan embrio secara dini, tetapi kurang berperan untuk perkembangan selanjutnya. Untuk meningkatkan pematangan folikel dalam jumlah besar (gelombang), diberikan perlakuan hormon gonadotropin selama fase luteal siklus estrus (Armstrong, 1993).
Toelihere (1985) menyatakan bahwa hormon utama yang digunakan pada superovulasi adalah hormon gonadotropin. yaitu FSH dan LH. FSH merupakan hormon gonadotropin dengan unsur glikopeptida yang memiliki reseptor pada sel granulosa folikel yang berfungsi menstimulasi pertumbuhan folikel, sehingga sangat diperlukan dalam proses superovulasi. Kaiin dan Tappa (2006) menambahkan hormon yang umum digunakan untuk menginduksi superovulasi pada sapi adalah Follicle Stimulating Hormone (FSH) yang berasal dari hipofisa. FSH merupakan hormon glikoprotein yang mempunyai waktu paruh pendek, sehingga memerlukan pemberian secara berulang untuk merangsang aktivitas folikel secara lebih efisien.
6 kualitas embrio yang kurang memenuhi klasifikasi yang telah ditentukan (Yusuf et al., 1993). Waktu paruh PMSG yang panjang menyebabkan terus terjadi stimulasi pembentukan folikel baru, meskipun ovulasi sudah selesai sehingga dari folikel yang terbentuk akan menghasilkan estrogen dengan kadar cukup tinggi yang pada akhirnya akan mengganggu transpor dan daya tahan hidup embrio (Mustofa, 1999). Baik FSH maupun PMSG telah banyak digunakan dalam teknik-teknik tertentu (misalnya produksi embrio) untuk menginduksi superovulasi (Hunter, 1995). Respon imunologi terhadap pemberian injeksi berulang hormon gonadotropin dapat membatasi kemampuan respon sapi donor terhadap superovulasi. PMSG dan FSH merupakan hormon protein sehingga sangat potensial menginduksi reaksi anafilaksis. Hal ini menandakan bahwa injeksi berulang dapat merangsang pembentukan anti- gonadotropin yang dapat mengurangi respon selanjutnya terhadap hormon gonadotropin endogen (Seidel dan Elsden, 1985).
Seleksi Betina Donor
7 Sinkronisasi Estrus
Sinkronisasi estrus adalah pencocokan siklus estrus dari ternak donor dan penerima dengan injeksi prostaglandin (PGF2) untuk merangsang estrus (Kunkel,
1998). Preparat Prostaglandin F2α (PGF2α) dikenal sebagai agen luteolitik yang dapat menyamakan siklus estrus dalam waktu yang bersamaan, sedangkan hCG dapat menginduksi ovulasi, sehingga pemberian hCG pada pertengahan estrus dapat merangsang pelepasan ovum dalam waktu yang lebih seragam. Dengan demikian perkembangan folikel dapat diamati sehingga dapat ditentukan waktu inseminasi yang lebih tepat (Arifiantini et al., 2010).
Superovulasi
Ovulasi adalah proses pemecahan folikel de graaf yang terjadi sewaktu ovum dilepaskan dari ovarium. Tingkatan ovarium adalah primer, sekunder dan tersier dan folikel de graaf. LH menyebabkan pengendoran dinding folikel sehingga lapisan-lapisan pecah dan melepaskan ovum dan cairan folikel, sesudah ovulasi terbentuk Corpus luteum di dalam folikel yang telah pecah dan mulai mensekresikan progesterone. Hewan-hewan betina dewasa yang disuntikan hormon gonadotropin dapat menghasilkan 20-100 ova pada satu estrus. FSH menggertak pematangan beberapa folikel, sedangkan LH menyebabkan ovulasi hal ini disebut superovulasi (Toelihere, 1985). Dalam program Transfer Embrio (TE), untuk merangsang ovulasi ganda (multiple ovulation), maka diberikan hormon superovulasi sehingga diperoleh 12-15 sel telur dalam satu kali ovulasi (Herren, 2000).
Superovulasi dapat diinduksi secara buatan melalui pemberian hormon gonadotropin eksogen (berasal dari luar tubuh), misalnya FSH dan PMSG. Pemberian hormon tersebut dengan dosis tertentu akan menstimulasi proses pertumbuhan, perkembangan, pematangan dan ovulasi dari sejumlah besar folikel pada ternak sapi. Betina donor diinjeksikan setiap hari dengan FSH (Herren, 2000) yang dapat berasal dari ekstrak hipofise babi dan domba (Wheeler dan Bowen, 1989) atau dari esktrak hipofise sapi (Wilson, 1992). Donor tertentu memerlukan penambahan LH selain FSH, namun umumnya preparat FSH yang dijual sudah ditambahkan LH (Wright, 1987).
8 agar siklus estrus berjalan normal dan teratur sehingga perlakuan superovulasi dapat sinkron dengan pola hormonal hewan secara normal. Jika siklus estrus abnormal, maka perlakuan superovulasi mungkin mengalami kegagalan Wilson (1992).
Inseminasi Buatan
Setelah berhasil memilih hewan donor berkualitas tinggi, kunci keberhasilan Transfer Embrio (TE) selanjutnya terletak pada inseminasi dengan semen yang berasal dari sapi jantan bibit unggul (Davis, 2004). Setelah perlakuan superovulasi, perlu dilakukan pengamatan terhadap tanda-tanda estrus pada sapi donor sehingga dapat dijadikan acuan untuk menentukan waktu inseminasi yang tepat dan IB dilakukan, penting untuk memeriksa berahi dan inseminasi 10 sampai 20 jam setelah awal berahi (Inseminasi dilakukan ketika sejumlah besar telur dilepaskan dari ovarium, ovulasi dapat terjadi selama 24 jam atau lebih). Pada saat telur terakhir dilepaskan dari ovarium mungkin tidak ada cukup sperma untuk membuahi. Sangat penting bahwa prosedur IB harus dilakukan secara tepat dan hanya dilakukan oleh teknisi IB yang sangat berpengalaman (Lewis, 1996).
9 dapat digunakan semen segar dengan konsentrasi 10-50 juta sperma motil atau semen beku yang mengandung 30 juta sperma motil dan diberikan dengan dosis ganda. Grimes (2008) menyarankan agar inseminasi dilakukan 1-3 kali selama dan setelah estrus dengan interval yang sama.
Koleksi Embrio
Panen atau koleksi embrio pada sapi donor dilakukan pada hari ke-7 sampai hari ke-8 setelah berahi saat sebagian besar embrio sudah memasuki ujung cornua uteri pada masa itu. Embrio akan berkembang sekitar satu minggu, kemudian embrio dipanen pada tahap morula sampai blastocyst (Grimes, 2008). Embrio dikoleksi antara hari ke-6 dan ke-8 setelah estrus (Herren, 2000). Pemanenan embrio tidak dilakukan lebih awal karena dapat menurunkan efisiensi koleksi embrio dengan metode non bedah. Sebelum hari ke-4, hampir semua embrio terletak di dalam oviduk yang dipisahkan dari uterus oleh utero-tubal junction. Struktur ini berfungsi sebagai katup (valve) yang dapat mengatur masuknya sperma dari uterus menuju oviduk sehingga fertilisasi terjadi tepat waktu dan mengatur transpor embrio ke arah sebaliknya. Embrio akan ditranspor menuju uterus pada hari ke-4 sampai hari ke-5 setelah estrus, melalui kontraksi ritmik pada dinding oviduk dan relaksasi dari otot pada dinding utero-tubal junction sehingga tingkat keberhasilan koleksi embrio akan lebih tinggi pada hari ke-6 dan seterusnya daripada hari ke-4. Kadang-kadang beberapa embrio masih ditemukan dalam oviduk pada sapi yang disuperovulasi sampai hari ke-10 (Seidel dan Elsden, 1989).
10
Gambar 1. Tahap Perkembangan Embrio (Robertson dan Nelson, 2009)
Batas waktu paling akhir untuk koleksi embrio adalah hari ke-14 setelah estrus, sedangkan di luar batas ini embrio yang dikoleksi akan rusak. Pada hari ke-9, embrio mulai keluar dari zona pellusida dan koleksi embrio tidak disarankan pada tahap ini karena kesulitan dalam mengidentifikasi embrio ketika ia sudah keluar dari zona pellusida. Hampir semua embrio telah keluar pada hari ke-11, pada saat diameter embrio meningkat secara drastis. Hari ke-12 sampai ke-13, embrio mulai memanjang (elongate) dan tampak seperti bola (American football). Hari ke-14 sampai ke-15, bentuk embrio sangat panjang seperti spaghetti. Hari ke-18 sampai
Morula (hari 6) Blastosit awal
11 hari ke-19, embrio hampir memenuhi cornua uteri. Koleksi embrio mungkin dapat dilakukan pada hari ke-17 dengan teknik non bedah, tetapi potensi terjadinya kerusakan/cacat pada embrio sangat besar sejak hari ke-14 (Seidel dan Elsden, 1989). Tahap perkembangan embrio dapat dilihat pada Tabel 1.
Tabel 1. Tahap Perkembangan Embrio Tahap Deskripsi
1 Belum dibuahi
2 2-12 sel
3 Morula awal
4 Morula
5 Blastosit awal
6 Blastosit
7 Expanded blastocyst
8 Hatched blastocyst
9 Expanded hatched blastocyst
Sumber: Wright (2009)
12
Corpus Luteum (CL)
Corpus Luteum (CL) merupakan benda yang terbentuk pada tempat ovum diovulasikan dan dijadikan patokan untuk mendeteksi berapa jumlah ovum yang diovulasikan oleh seekor sapi (Adriani et al., 2009). Setelah terjadi ovulasi maka pada situs pelepasan oosit akan terbentuk Corpus Luteum (CL). Selama awal fase luteal (metestrus). CL dibentuk dari sel-sel luteal. Pada pertengahan fase luteal (diestrus) sel-sel luteal menghasilkan sejumlah besar progesteron. Selama akhir fase luteal, CL dilisiskan PGF2α yang dihasilkan endometrium uterus. Lisis CL diikuti dengan penurunan kadar progesteron. sehingga mekanisme umpan balik negatif progesteron pada hypotalamus hilang, mengakibatkan peningkatan GNRH yang menandakan dimulai fase folikular. Ukuran CL pada hari ke 3-5 mulai meningkat sampai maksimal disertai dengan peningkatan produksi progesteron sampai kadar maksimal sekitar hari ke-10 (Senger, 1999).
Menurut Amiridis et al. (2006), CL tersusun atas sel-sel luteal yang berperan menghasilkan progesteron. Konsitensi atau kekenyalan badan CL sangat ditentukan jumlah sel-sel luteal dan vaskuralisasi darah kebagian tersebut. Demikian juga kemampuan CL memproduksi progesteron tergantung pada tingkat vaskularisasi pada lapisan seluler. Fungsi CL yang rendah (sintesis dan sekresi progesteron sedikit) diyakini akan menjadi penyebab penting kegagalan reproduksi dan ketidakmampuan uterus dalam mendukung perkembangan embrio dini. Senger (1999) menambahkan, dalam satu siklus estrus CL harus mengalami lisis agar fase folikular dimulai. Ovulasi tidak dapat terjadi dalam kondisi saat kadar progesteron dominan. Luteolisis berarti disintengrasi atau dekomposisi dari CL yang terjadi 2-3 hari pada akhir fase luteal. Dua hormon yang berperan penting dalam lisis CL yaitu oxytocin yang dihasilkan CL dan hormon PGF2α yang dihasilkan endometrium uterus.
Klasifikasi Embrio
13 ditransfer ke resipien yang tepat, dimana embrio dengan kualitas terbaik ditransfer ke respien yang paling baik pula (Seidel dan Elsden, 1985).
15 METODE
Lokasi dan Waktu
Penelitian ini dilaksanakan melalui magang selama tiga bulan, yaitu mulai bulan April 2011 hingga bulan Juni 2011. Penelitian dilaksanakan di Laboratorium Balai Embrio Ternak (BET) yang terletak di Desa Cipelang, Kecamatan Cijeruk, Kabupaten Bogor.
Materi
Penelitian ini dilakukan dengan menggunakan data sekunder produksi embrio yang diperoleh dari Balai Embrio Ternak (BET), Cipelang, Bogor. Data tersebut berupa catatan produksi embrio secara in vivo selama tahun 2009-2010. Data tersebut meliputi tanggal superovulasi, kode dan jenis ternak donor, kode semen yang digunakan, merk dagang hormon superovulasi yang digunakan, jumlah Corpus Luteum (CL), jumlah embrio grade A, B, C, dan D serta jumlah embrio layak transfer dan jumlah ovum tidak dibuahi atau Unfertilized (UF). Ternak sapi donor yang digunakan dalam penelitian ini sebanyak 90 ekor sapi, terdiri atas 29 ekor sapi FH, 23 ekor sapi Simmental, 27 ekor sapi Limmousin dan 11 ekor sapi Angus. Sebagian besar sapi disuperovulasi lebih dari sekali dengan jarak antar superovulasi sekitar tiga bulan sehingga didapatkan data superovulasi sebanyak 223 data. Data yang diperoleh kemudian dimasukkan dalam program Microsoft Access Database 2010. Pakan yang diberikan kepada sapi donor adalah rumput dan konsentrat. Gambaran sapi donor yang digunakan dapat dilihat pada Gambar 2.