30
IV. HASIL DAN PEMBAHASAN
A. PRODUKSI INOKULUM
1. Karakteristik Substrat Inokulum
Substrat yang digunakan terdiri dari onggok (ampas tapioka), bekatul, bungkil kacang tanah dan ampas tahu. Substrat tersebut dipilih dikarenakan mudah untuk diperoleh dan merupakan hasil samping dari industri pertanian.
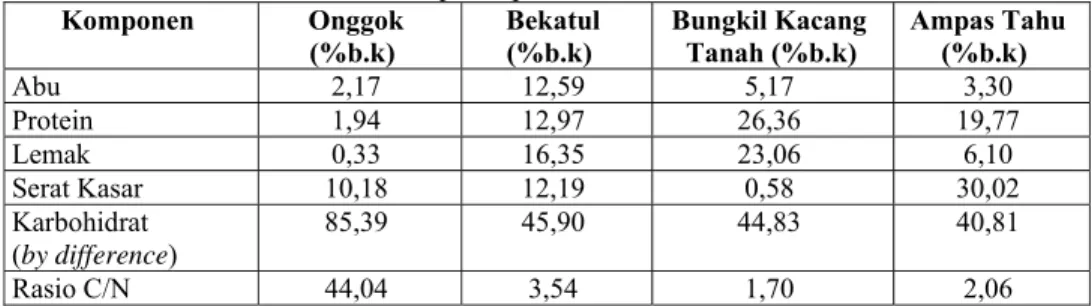
Substrat yang digunakan harus dikecilkan ukurannya sampai sebesar 40 mesh dan digunakan keadaan steril. Hal ini dilakukan agar kapang dapat tumbuh secara merata dan tidak adanya mikroba lain yang tumbuh di dalam inokulum kapang tersebut. Selanjutnya substrat tersebut dianalisis komponen proksimat untuk menentukan jumlah nitrogen dan karbon di dalam media tersebut. Jumlah karbon diambil dengan melakukan pendekatan analisa karbohidrat (by difference). Jumlah nitrogen diambil dengan melakukan pendekatan analisa protein kasar. Nisbah karbon dan protein ini digunakan untuk menentukan perbandingan nilai C/N yang terbaik untuk pembuatan inokulum. Pada Tabel 2, diketahui bahwa media yang sesuai untuk pertumbuhan kapang mengandung karbon sebanyak 40-63% (b.k) dan nitrogen sebanyak 7-10% (b.k). Berdasarkan data tersebut diketahui bahwa kandungan C/N adalah antara 4-9. Hasil analisa proksimat onggok, bekatul, bungkil kacang tanah dan ampas tahu dapat dilihat pada Tabel 4.
Tabel 4. Komposisi proksimat substrat inokulum Komponen Onggok
(%b.k) Bekatul (%b.k) Bungkil Kacang Tanah (%b.k) Ampas Tahu (%b.k)
Abu 2,17 12,59 5,17 3,30 Protein 1,94 12,97 26,36 19,77 Lemak 0,33 16,35 23,06 6,10 Serat Kasar 10,18 12,19 0,58 30,02 Karbohidrat (by difference) 85,39 45,90 44,83 40,81 Rasio C/N 44,04 3,54 1,70 2,06
Berdasarkan analisa proksimat tersebut, diperlukan kombinasi substrat untuk membuat media dengan nisbah C/N = 5 bagi pembuatan inokulum kapang. Hal ini dikarenakan untuk membuat inokulum dibutuhkan kandungan nitrogen yang cukup serta kandungan karbon yang tidak terlalu tinggi. Hal ini diperkuat oleh Peppler (1979), yang menyatakan bahwa untuk pertumbuhan kapang, nitrogen sangat dibutuhkan sebagai stimulan dalam pertumbuhan kapang sehingga dihasilkan spora yang banyak, sedangkan karbon dibutuhkan sebagai sumber energi dengan cara melakukan proses pemecahan karbohidrat yang ada pada bahan sehingga diperoleh energi.
Substrat onggok digunakan sebagai komponen substrat utama sumber karbon, sedangkan sumber protein diperoleh dari bekatul, bungkil kacang tanah dan ampas tahu. Substrat tersebut dikombinasikan dengan berdasarkan penghitungan yang dilakukan secara subtitusi yang dapat dilihat pada Lampiran 2. Berdasarkan hasil perhitungan diketahui bahwa perbandingan onggok dengan bekatul adalah 1,86 : 7,43, perbandingan onggok dengan bungkil kacang tanah adalah 4,02 : 3,50 , serta perbandingan onggok dengan ampas tahu adalah 3,61 : 4,71. Hasil perhitungan untuk setiap jenis kombinasi substrat dapat dilihat pada Tabel 5.
31
Tabel 5. Hasil perhitungan komposisi campuran substrat inokulum dengan rasio C/N = 5/1Komponen Onggok+bekatul (%b.k) Onggok+Bungkil Kacang Tanah (%b.k) Onggok+Ampas Tahu (%b.k) Persentase bobot 1,86 : 7,43 4,02 : 3,50 3,61: 4,71 Abu 10,50 3,56 2,81 Protein 10,76 13,31 12,03 Lemak 13,14 10,91 3,60 Serat Kasar 11,78 5,71 21,41 Karbohidrat (by difference) 53,81 66,51 60,16
2. Pembuatan Inokulum
Inokulum merupakan bahan dalam bentuk padat maupun cair yang mengandung spora atau konidia yang sengaja ditambahkan pada substrat. Tujuan dibuatnya spora kapang dalam bentuk inokulum adalah untuk mempermudah penggunaan kapang tersebut dan membuat kapang menjadi tahan lama untuk dapat digunakan langsung saat diperlukan.
Produksi inokulum dilakukan dengan menggunakan media yang telah dihitung kombinasinya. Sebelum diinokulasi, media tersebut harus ditambahkan air untuk memberikan kondisi pertumbuhan yang sesuai untuk kapang. Selain itu, substrat juga harus disterilkan untuk menghindari tumbuhnya kapang lain.
Media substrat yang telah siap, dimasukkan ke dalam wadah plastik (baskom) yang telah disterilkan dan dimasukkan ke dalam ruangan steril. Kapang yang telah disegarkan dengan menggunakan media agar miring PDA diinokulasikan ke dalam media dan ditutup dengan menggunakan kertas buram. Kertas buram digunakan karena kapang membutuhkan oksigen untuk melakukan metabolisme dalam pertumbuhannya, namun untuk menghindari kontaminasi maka wadah ditutup dengan menggunakan bahan yang tidak terlalu rapat seperti kertas buram.
Media yang telah diinokulasikan dengan kapang tersebut kemudian diinkubasikan selama tiga hari untuk mendapatkan pertumbuhan kapang pada fase awal stasioner. Fase ini dipilih karena spora kapang berada pada kondisi maksimal. Menurut Gandjar (2006), kapang Trichoderma viride dan Rhizopus oryzae cenderung mengalami fase stationer setelah diinkubasi selama 3 hari.
Setelah pemanenan inokulum kapang selanjutnya dilakukan pengeringan inokulum tersebut pada suhu 50°C selama satu hari. Suhu pengeringan ini tidak boleh terlalu tinggi dikarenakan dapat mematikan spora kapang. Pengeringan dilakukan agar aktivitas metabolisme kapang menjadi terhenti dikarenakan tidak memiliki kondisi kadar air yang sesuai bagi kapang untuk melakukan metabolisme. Menurut Fardiaz (1989), air sangat mempengaruhi pertumbuhan kapang dikarenakan air berfungsi sebagai reaktan dan struktur organ dari kapang.
Inokulum yang telah kering disimpan dalam plastik yang telah diberi klep serta disimpan dalam stoples yang tahan terhadap uap air dari luar. Kemasan ini digunakan untuk menjaga agar kadar air di dalam bahan tidak mengalami perubahan serta untuk mempermudah dalam pengujian viabilitas spora. Jenis plastik yang digunakan adalah jenis polietilen. Menurut Sacharow dan Griffin (1970), plastik jenis polietilen memiliki derajat kerapatan yang baik dan mempunyai ketebalan 0,001 sampai 0,01 inchi. Dikarenakan hal inilah maka plastik polietilen baik digunakan untuk menjaga kondisi suhu dan kelembapan inokulum. Selain itu jenis plastik ini juga mudah diperoleh di pasaran sehingga dapat memudahkan masyarakat dalam penggunaannya. Penyimpanan dilakukan dalam kondisi suhu ruang (28-32°C).
B. P
p t k t1
PERUBAH
Pengu pengamatan se terdiri atas pen kapang tersebu terhadap perub1. Kadar A
H dan intera kadar air. kadar air b L Menurut B mikoba ya berpengaru Perubahan dilihat pad Gambar 3 1 1 2 Kad ar Air ( % ) 1 1 2 Kad ar Air (% )HAN KUALI
ujian terhadap elama dua bula ngujian peruba ut. Hasil sidik r bahan parameteAir
Hasil analisis ra aksi antara jeni
Selanjutnya h berbeda nyata p Lama penyimp Buckle et al., ang menghasilk uh nyata terh n kadar air pa da Gambar 3. . Perubahan k penyimpana -5 0 5 0 0 1 Umur S -5 0 5 0 0 1 Umur S
ITAS INOK
kualitas inoku an, sehingga dip ahan kadar airragam (Lampi er kualitas inok
agam (Lampira is substrat dan hasil uji lanjut pada semua pe panan mengaki (1985), pening kan air. Berdas hadap peruba ada inokulum
kadar air inokul an 2 3 4 Simpan Inokul 2 3 4 Simpan Inoku
KULUM SE
ulum dilakukan peroleh sembil dan viabilitas ran 3), menunj kulum selama p an 3) menunju n lama penyim t Duncan terha erlakuan kompo ibatkan kadar gkatan kadar a sarkan data ana ahan paramete Rhizopus oryz lum Rhizopus o 5 6 7 lum (Minggu) 5 6 7 ulum (Minggu)ELAMA PEN
n setiap semin lan titik penguj spora dari keti jukan bahwa je penyimpanan.ukan bahwa jen mpanan member adap komposis osisi substrat.
air di dalam air dapat terjad alisis ragam m er kadar air yzae dan Trich
oryzae (a) dan 7 8 ) 7 8 )
NYIMPAN
nggu sekali de jian. Pengujian iga jenis substr enis kapang benis substrat, lam rikan pengaruh si substrat men bahan cender di karena adan menunjukan bah bahan selama hoderma viride Trichoderma v Bekatul + O Kacang Tan Ampas Tahu Bekatul +On Kacang Tan Ampas Tahu
3
AN
ngan melakuk n yang dilakuk rat dan dua jen erpengaruh nya ma penyimpan h nyata terhad nunjukan bahw rung bertamba nya metabolism hwa jenis kapan a penyimpana e tersebut dap viride (b) selam Onggok nah + Onggok u + Onggok nggok nah + Onggok u +Onggok32
kan kan nis ata nan dap wa ah. me ng an. pat ma33
Berdasarkan data pada Lampiran 3.A dapat diketahui bahwa kadar air awal inokulum Rhizopus oryzae lebih tinggi bila dibandingkan dengan kadar air awal inokulum Trichoderma viride. Hal ini dikarenakan struktur kapang R.oryzae yang memiliki miselium yang cenderung lebih rapat bila dibandingkan dengan kapang T.viride, sehingga mengakibatkan tingginya kandungan air dalam inokulum (Dwidjoseputro, 1978).Perubahan kadar air pada inokulum kapang Rhizopus oryzae cenderung lebih stabil bila dibandingkan dengan perubahan kadar air pada inokulum kapang Trichoderma viride. Pada inokulum kapang Rhizopus oryzae yang menggunakan substrat bekatul+onggok terjadi perubahan dari 10,49% menjadi 10,18%, pada inokulum yang menggunakan substrat bungkil kacang tanah+onggok terjadi perubahan dari 16,12% menjadi 17,08% dan pada inokulum ampas tahu+onggok terjadi perubahan dari 12,39% menjadi 12,36%. Pada inokulum kapang Trichoderma viride yang menggunakan substrat bekatul+onggok terjadi kenaikan dari 7,35% menjadi 9,05%, pada inokulum yang menggunakan substrat bungkil kacang tanah+onggok terjadi kenaikan dari 8,92% menjadi 9,95% dan pada inokulum yang menggunakan substrat ampas tahu+onggok terjadi kenaikan dari 10,20% menjadi 12,51%.
Berdasarkan data peningkatan kadar air tersebut, inokulum kapang Rhizopus oryzae cenderung lebih stabil bila dibandingkan dengan inokulum kapang Trichoderma viride. Adanya perbedaan dari peningkatan kadar air pada inokulum kedua kapang tersebut ditentukan dari sifat substrat yang digunakan serta jenis dari kapang yang digunakan. Bahan yang bersifat higroskopis, cenderung lebih cepat menyerap uap air bila dibandingkan dengan bahan yang tidak mempunyai sifat higroskopis. Ketiga jenis substrat kombinasi yang digunakan mempunyai sifat higroskopis yang relatif sama, oleh sebab itu perubahan kadar air pada inokulum lebih cenderung ditentukan oleh jenis kapang yang digunakan. Kapang Rhizopus oryzae memiliki miselium yang tidak bersekat-sekat dan cenderung lebih rapat bila dibandingkan dengan kapang Trichoderma viride. Hal inilah yang mengakibatkan uap air yang ada di udara menjadi sulit masuk ke dalam inokulum, sehingga mengakibatkan inokulum kapang Rhizopus oryzae cenderung lebih stabil bila dibandingkan dengan inokulum kapang Trichoderma viride.
Peningkatan kadar air juga dapat diakibatkan oleh adanya penetrasi uap air dari lingkungan yang masuk ke dalam bahan. Kenaikan kadar air yang terjadi tidak terlalu tinggi dikarenakan penyimpanan dilakukan pada tempat yang cenderung kering dan menggunakan kemasan polietilen yang tahan terhadap uap air dari luar. Hal ini sesuai dengan pernyataan Sacharow dan Griffin (1970), plastik kemasan polietilen mempunyai daya proteksi yang baik terhadap uap air akan tetapi kurang baik terhadap gas-gas yang lain seperti oksigen.
Inokulum kering bersifat higroskopis, sehingga mudah menyerap air yang ada di udara. Semakin kecil kadar air inokulum dibandingkan dengan kadar air ruangan penyimpanan, mengakibatkan semakin banyak uap air yang menyerap masuk ke dalam bahan untuk menyeimbangkan kelembaban di dalam bahan dengan kelembaban yang ada di luar kemasan. Adanya perbedaan kelembaban di dalam bahan dengan lingkungan di luar dapat mengakibatkan terjadinya penetrasi uap air ke dalam bahan (Buckle et al., 1985).
2. Viabilitas Spora
Spora kapang tidak dapat tumbuh di dalam bahan inokulum yang sudah dikeringkan. Hal ini diperkuat oleh Dwidjoseputro (1978), yaitu dalam keadaan nutrisi yang optimum, kapang membiak dengan cepat sekali, namun apabila terjadi kekeringan, dapat mengakibatkan faktor pembatas bagi pertumbuhan kapang. Hal inilah yang mengakibatkan selama penyimpanan spora kapang tidak terjadi kenaikan jumlah spora di dalam inokulum tersebut.
Hasil analisis ragam viabilitas spora menunjukan bahwa jenis substrat, lama penyimpanan dan interaksi antara jenis substrat dengan lama penyimpanan memberikan pengaruh nyata pada viabilitas spora (Lampiran 3.B). Hasil dari uji lanjut Duncan diketahui bahwa viabilitas spora pada substrat kombinasi antara bekatul dengan onggok tidak berbeda nyata dengan substrat kombinasi antara ampas tahu dengan onggok, sedangkan untuk substrat kombinasi bungkil kacang tanah dan onggok berbeda nyata dengan kedua bahan yang lain. Hal ini menandakan bahwa viabilitas spora pada penggunaan substrat kombinasi bekatul dengan onggok tidak berbeda jauh dengan substrat kombinasi pada ampas tahu dan onggok pada kedua jenis kapang selulolitik yang digunakan.
34
Menurut Gaman dan Sherrington (1981), spora kapang merupakan bentuk germinasi dari kapang yang merupakan alat untuk bereproduksi pada kapang secara aseksual. Spora untuk setiap kapang selulolitik memiliki kemampuan yang berbeda-beda untuk dapat bertahan hidup pada setiap media yang digunakan. Hal inilah yang mengakibatkan terjadinya perbedaan jumlah spora yang dihasilkan pada setiap media.Perbedaan pertumbuhan spora juga dapat diakibatkan oleh adanya faktor kematangan spora pada saat proses pembuatan inokulum. Menurut Bilgrami dan Verma (1978), spora kapang selama mengalami periode dorman melakukan penyempurnaan proses pematangan. Apabila spora tersebut telah matang maka periode dorman digunakan sebagai periode menjalani istirahat sebelum bergerminasi. Hal inilah yang mengakibatkan spora yang sudah matang akan lebih cepat bergerminasi bila dibandingkan dengan spora yang belum cukup matang. Spora yang belum cukup matang akan melakukan penyempurnaan kematangannya dengan menambah waktu dormannya. Hal inilah yang juga mengakibatkan terjadinya perbedaan jumlah spora yang dihasilkan oleh setiap kapang terhadap media inokulum yang digunakan.
Spora yang telah matang juga memiliki kelebihan bila dibandingkan dengan spora yang belum terlalu matang. Hal ini diperkuat oleh Ilyas (2007), yaitu secara morfologis dan fisiologis spora yang sudah matang akan memiliki ketahanan yang lebih terhadap perubahan kondisi lingkungan. Hal inilah yang mengakibatkan perubahan viabilitas spora bagi spora yang viabilitas awalnya tertinggi akan menjadi lebih dapat bertahan lama apabila dibandingkan dengan yang viabilitas awalnya rendah.
Berdasarkan hasil analisis ragam (Lampiran 3) menunjukan bahwa jenis kapang berpengaruh nyata terhadap perubahan parameter viabilitas spora pada inokulum selama penyimpanan. Hasil analisis viabilitas spora pada kedua jenis kapang menunjukan terjadinya penurunan jumlah spora. Hal ini dikarenakan adanya perubahan komponen nutrisi di dalam substrat serta adanya kondisi lingkungan yang mempengaruhi inokulum tersebut. Hal ini diperkuat oleh Frazier (1992), spora mikroba dapat berkurang kemampuannya untuk bergerminasi dikarenakan kondisi lingkungan yang tidak sesuai. Menurut Fardiaz (1989), faktor yang mempengaruhi pertumbuhan mikroba tersebut terdiri atas perubahan kadar air, oksigen, suhu, pH, menurunnya kandungan nutrien di dalam bahan serta adanya komponen anti mikroba di dalam bahan. Hal ini yang mengakibatkan semakin lama inokulum disimpan, maka akan semakin berkurang jumlah spora yang ada di dalam inokulum tersebut.
Berdasarkan data hasil pengamatan viabilitas spora selama dua bulan untuk kapang R.oryzae (Lampiran 3B), diketahui bahwa semakin lama penyimpanan inokulum mengakibatkan spora dalam berbagai substrat mengalami penurunan. Data viabilitas spora dapat dilihat pada Gambar 4. Pada substrat bekatul+onggok mengalami penurunan dari 106,13 x 107 menjadi 24,49 x 107 (spora/g bobot kering) atau mengalami penurunan sebesar 7,06%. Pada substrat bungkil kacang tanah+onggok mengalami penurunan dari 21,46 x 107 menjadi 1,21 x 107 (spora/g bobot kering) atau mengalami penurunan sebesar 15,01%. Pada substrat ampas tahu+onggok mengalami penurunan dari 92,45x 107 menjadi 32,00 x 107 (spora/g bobot kering) atau mengalami penurunan sebesar 6,66%.
Berdasarkan data hasil pengamatan viabilitas spora selama dua bulan untuk kapang T.viride, diketahui bahwa semakin lama penyimpanan inokulum mengakibatkan spora dalam berbagai substrat juga mengalami penurunan. Pada substrat bekatul+onggok mengalami penurunan dari 75,01 x 107 menjadi 13,74 x 107 (spora/g bobot kering) atau mengalami penurunan sebesar 8,3%. Pada substrat bungkil kacang tanah+onggok mengalami penurunan dari 11,53 x 107 menjadi 1,11 x 107 (spora/g bobot kering) atau mengalami penurunan sebesar 12,61%. Pada substrat ampas tahu+onggok mengalami penurunan dari 60,69 x 107 menjadi 12,00 x 107 (spora/g bobot kering) atau mengalami penurunan sebesar 8,01%.
Berdasarkan data hasil pengamatan viabilitas spora R.oryzae dan T.viride terhadap media (Lampiran 3B), diperoleh hasil yang tidak berpengaruh signifikan antara substrat bekatul+onggok dengan ampas tahu+onggok, sedangkan dengan substrat bungkil kacang tanah+onggok terdapat perbedaan yang signifikan bila dibandingkan dengan kedua substrat yang lain. Hal ini dikarenakan perbandingan media yang digunakan ini dilakukan berdasarkan persamaan nisbah C/N yang diambil dari pendekatan karbohidrat by difference dan pendekatan protein kasar tanpa memperhatikan perbandingan komposisi zat kimia lain pembentuk substrat inokulum. Substrat inokulum bekatul+onggok dan ampas tahu+onggok memiliki komposisi serat kasar yang tinggi dan kadar lemak yang rendah bila dibandingkan dengan substrat inokulum bungkil kacang
tanah+ong inokulum Gambar 4 Ju dikarenaka lemak yan spora mik bahwa mi terlalu ba pembentuk tahu+ongg inokulum K pertumbuh karbohidra kasar yang terhadap k kapang di bekatul+on 1 Ju ml ah S p or a (% ) 1 Juml ah Sp ora (% )
ggok. Hal inil tersebut. 4. Perubahan v penyimpana umlah spora ya an adanya kan ng tinggi dapat kroba tersebut. inyak diperluk anyak dapat m kan sel mikrob gok menghasi bungkil kacan Kandungan se han kapang. M at kompleks ya g tinggi di dala kapang dalam b i dalam substr nggok. 80 85 90 95 00 0 1 Umur 80 85 90 95 00 0 1 U
lah yang mem
viabilitas spora an
ang sangat ren ndungan lemak t menghambat . Hal ini diper kan dalam pem mengakibatkan
ba. Hal ini men lkan sel spor g tanah+onggo erat kasar di Menurut Mayna ang hanya dapa am bahan akan
bentuk karbon. rat ampas tahu
2 3 4 r Simpan Inok 2 3 4 Umur Simpan mpengaruhi pe a Rhizopus ory
ndah pada subst k yang tinggi pertumbuhan b rkuat oleh Mc mbuatan medi minyak menu ngakibatkan su ra yang lebih ok. dalam substr ard dan Loosli at dicerna oleh n dihidrolisis ol . Hal inilah yan u+onggok leb 4 5 6 7 kulum (Minggu 4 5 6 7 n Inokulum (M erbedaan juml
yzae (a) dan T
trat kombinasi pada substrat bagi mikroba s cNeil dan Harv ia, namun kan
utupi substrat ubstrat inokulu tinggi bila d rat dapat me i (1956), serat h mikroba, teru
leh kapang unt ng mengakibat bih kecil bila
7 8 u) 7 8 Minggu) ah spora yang Trichoderma v bungkil kacan t tersebut. Ad serta mempeng vey (2008), y ndungan jumla sehingga dap um bekatul+on dibandingkan emberikan pen kasar mengan utama kapang. K tuk memberika tkan persentase dibandingkan Bekatul + O Bungkil Ka Onggok Ampas Tah (b Bekatul + O Bungkil Ka Onggok Ampas Tah
3
g ada di dala viride (b) selam ng tanah+ongg anya kandung garuhi jumlah s ang mengatak ah minyak yan pat menghamb nggok dan amp dengan substr ngaruh terhad ndung kompon Kandungan ser an sumber ener e kematian spo dengan substr Onggok acang Tanah + hu + Onggok b) Onggok acang Tanah + hu + Onggok35
am ma ok gan sel kan ng bat pas rat dap nen rat rgi ora rat36
Berdasarkan data hasil pengamatan viabilitas spora inokulum yang telah disimpan selama dua bulan, diketahui bahwa substrat yang menghasilkan spora terbanyak adalah substrat bekatul+onggok. Hal ini dikarenakan pada media ini mengandung nutrien yang baik untuk mendukung pertumbuhan spora kapang. Hal ini diperkuat oleh Rachman (1989), yaitu media untuk membentuk spora kapang pada media padat yang baik adalah dengan menggunakan media dari biji-bijian seperti barley, bekatul dan jagung. Setyowati (2006) juga mengatakan bahwa bekatul merupakan media yang baik untuk pertumbuhan mikroba dikarenakan mempunyai kandungan karbohidrat, protein, lemak, dan vitamin yang cukup banyak, sehingga cocok untuk nutrisi bagi pertumbuhan mikroba. Nursalim dan Razali (2007), juga menambahkan bahwa bekatul memiliki kandungan gizi yang tinggi dikarenakan mengandung mineral yang terdiri atas kalsium (Ca), magnesium (Mg), mangan (Mn), besi (Fe), kalium (K) dan seng (Zn). Hal inilah yang mengakibatkan bekatul menjadi media terbaik untuk menghasilkan inokulum spora kapang Rhizopus oryzae dan Trichoderma viride.Berdasarkan data hasil pengamatan viabilitas spora inokulum R.oryzae juga diperoleh bahwa bila dibandingkan antara substrat bekatul+onggok dengan substrat ampas tahu+onggok terdapat perbedaan yang tidak signifikan. Selain itu juga penurunan viabilitas terendah didapatkan pada substrat ampas tahu+onggok. Hal ini dikarenakan substrat ampas tahu+onggok mengandung kadar serat kasar yang tinggi sehingga dapat menjadi media pengisi bagi spora inokulum untuk dapat bertahan hidup.
Jumlah spora yang terdapat di dalam inokulum T.viride dan R.oryzae masih menunjukan hasil yang baik meskipun telah dilakukan penyimpanan selama dua bulan. Hal ini dilihat dari jumlah spora yang masih tinggi dan penurunan viabilitas spora yang tidak terlalu tinggi. Menurut Lindajati (1983) kandungan spora inokulum kapang yang terbaik adalah berkisar antara 107 – 108 spora/g. Hal ini sesuai dengan hasil penelitian yang dilakukan terhadap viabilitas spora T. viride dan R. oryzae yang berkisar antara 107 -108 spora/g, sehingga dapat dikatakan bahwa inokulum masih dalam keadaan baik untuk digunakan meskipun telah disimpan selama dua bulan.
C. APLIKASI INOKULUM UNTUK HIDROLISIS TONGKOL JAGUNG
1. Karakterisasi Tongkol Jagung
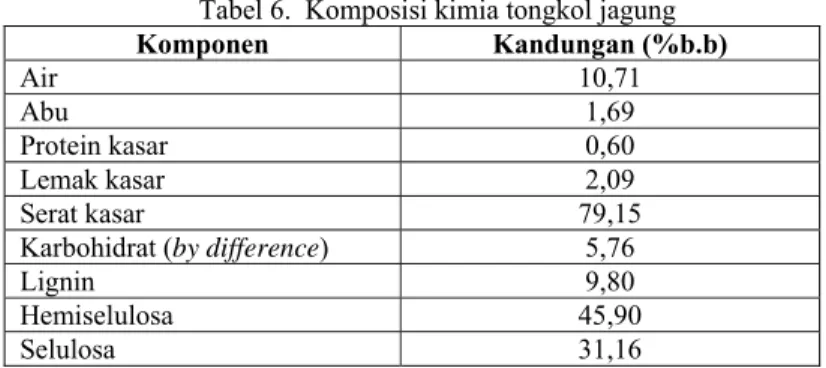
Berdasarkan hasil uji proksimat terhadap tongkol jagung yang digunakan sebagai media dalam penelitian ini mengandung komposisi yang disajikan dalam Tabel 6. Menurut Prihatman (2000), kandungan protein serta serat kasar yang dapat memenuhi kebutuhan ternak ruminansia masing-masing sebesar 8 % dan 15%. Jika dilihat dari data hasil uji proksimat tongkol jagung yang diperoleh, maka perlu dilakukan peningkatan kandungan protein dan penurunan kandungan serat kasar di dalam tongkol jagung tersebut sehingga dapat menjadi bahan pakan ternak yang baik untuk ternak ruminansia.
Tabel 6. Komposisi kimia tongkol jagung
Komponen Kandungan (%b.b) Air 10,71 Abu 1,69 Protein kasar 0,60 Lemak kasar 2,09 Serat kasar 79,15
Karbohidrat (by difference) 5,76
Lignin 9,80 Hemiselulosa 45,90 Selulosa 31,16
37
Menurut Prihatman (2000), kandungan protein serta serat kasar yang dapat memenuhi kebutuhan ternak ruminansia masing-masing sebesar 8% dan 15%. Jika dilihat dari data hasil uji proksimat tongkol jagung yang diperoleh, maka perlu dilakukan peningkatan kandungan protein dan penurunan kandungan serat kasar di dalam tongkol jagung tersebut sehingga dapat menjadi bahan pakan ternak yang baik untuk ternak ruminansia.Komponen protein yang rendah perlu ditingkatkan dengan cara menambahkan inokulum kapang ke dalam tongkol jagung. Hal ini dikarenakan inokulum kapang mengandung komponen protein yang tinggi sebagai komponen pembentuk sel kapang tersebut. Semakin banyak sel kapang yang terbentuk maka akan meningkatkan kandungan protein di dalam bahan tersebut.
Komponen serat yang tinggi dapat mengakibatkan tongkol jagung menjadi sulit dicerna oleh ternak. Komponen serat kasar yang terdapat di dalam tongkol jagung merupakan komponen serat lignoselulosa yang terdiri atas komponen lignin, hemiselulosa dan selulosa dalam struktur yang kompak. Kapang selulotik mempunyai kemampuan untuk menghidrolisis selulosa yang ada di dalam bahan menjadi gula-gula sederhana. Oleh sebab itu, adanya hidrolisis menggunakan kapang selulolitik akan menurunkan kadar serat kasar pada tongkol jagung. Kelemahan dari kultivasi tongkol jagung menggunakan kapang selulolitik adalah adanya kandungan lignin di dalam tongkol jagung yang masih cukup tinggi dikarenakan lignin sulit terhidrolisis oleh kapang selulolitik, sehingga untuk mengoptimalkan hidrolisis tongkol jagung perlu dilakukan pendegradasian kadar lignin di dalam bahan.
2. Hidrolisis Tongkol Jagung Menggunakan Inokulum Terpilih
Inokulum kapang yang memiliki jumlah spora tertinggi dari setiap media yang digunakan pada kapang T.viride dan R.oryzae merupakan inokulum yang menggunakan substrat kombinasi bekatul+onggok. Hasil analisa kadar air dan viabilitas spora inokulum yang terbaik dapat dilihat pada Lampiran 6. Inokulum yang terbaik kemudian dikultivasikan ke dalam tongkol jagung untuk menghidrolisis tongkol jagung. Kultivasi dilakukan pada minggu keempat dan minggu kedelapan dari lama penyimpanan yang telah dilakukan.
Tongkol jagung yang akan dihidrolisis harus dikeringkan terlebih dahulu agar dapat diberikan perlakuan pengecilan ukuran terhadap tongkol jagung tersebut. Pengecilan ukuran dilakukan untuk menghasilkan luas permukaan yang besar sebagai tempat tumbuhnya kapang selulolitik, serta untuk memotong rantai amorf pada lapisan struktur lignoselulosa pada bahan sehingga mempermudah dalam proses hidrolisis. Hal ini diperkuat oleh Meryandini (2007), hemiselulosa maupun lignin akan mengganggu aktivitas enzim selulase yang hanya spesifik memotong ikatan β-1,4-glikosidik pada selulosa. Oleh sebab itu untuk meningkatkan luas permukaan substrat maka tongkol jagung harus diperkecil ukurannya.
Tongkol jagung yang telah berukuran 40 mesh tersebut ditambahkan dengan air dengan perbandingan tongkol jagung : air adalah 2 : 3. Penambahan air ini dilakukan untuk membuat kadar air di dalam bahan menjadi sekitar 60 - 68% agar kapang dapat tumbuh dengan baik pada media tersebut. Hal ini diperkuat oleh Fardiaz (1989), bahwa jika kadar air bahan yang digunakan untuk kultivasi dibawah 62% maka akan menyebabkan pertumbuhan kapang menjadi terhambat.
Tongkol jagung yang telah mengandung kadar air sekitar 60-68% disterilkan dengan menggunakan autoklaf. Ruangan tempat inokulasi dan inkubator ruang kultivasi perlu disterilkan juga agar tidak terjadi kontaminasi mikroba lain yang dapat mengganggu proses hidrolisis terhadap tongkol jagung. Menurut Riadi (2007), sterilisasi merupakan suatu proses yang bertujuan untuk membunuh mikroba yang mengakibatkan tidak terdapatnya mikroba di dalam bahan maupun area tersebut.
Inokulasi spora ke dalam media tongkol jagung dilakukan dengan menggunakan perbandingan bobot inokulum sebanyak 15% dari bobot tongkol jagung dalam setiap media inokulasi. Hal ini diperkuat oleh Fakhrudin (1995) dalam penelitiannya untuk memproduksi enzim selulosa menggunakan substrat campuran tandan kosong dan sabut kelapa sawit. Perlakuan pengadukan perlu dilakukan setelah spora diinokulasikan ke dalam media tongkol jagung. Pengadukan diperlukan untuk memperbesar kontak antara enzim selulase dan komponen selulosa sehingga dapat meningkatkan aktivitas enzim selulase secara merata (Meryandini, 2007). Kemudian media yang telah berisi spora ditutup dengan menggunakan kertas buram agar oksigen dapat tetap masuk (Arbianto, 1980) dan disimpan di dalam inkubator.
38
Kultivasi dilakukan selama sembilan hari. Hal ini berdasarkan penelitian sebelumnya diketahui bahwa kedua jenis kapang tersebut dapat menurunkan kadar serat tertinggi pada kultivasi yang dilakukan selama sembilan hari. Untuk mendapatkan hasil yang optimal, selama kultivasi perlu dilakukan penjagaan terhadap kondisi lingkungan sekitarnya. Hal ini dikarenakan menurut Arbianto (1980), faktor lingkungan seperti perubahan kandungan nutrien bahan, kandungan air bahan, pH, kelembaban, ketersediaan oksigen serta aerasi perlu dijaga agar aktivitas hidrolisis yang dilakukan kapang dapat terjadi secara optimal. Suhu yang digunakan selama proses inkubasi adalah suhu ruang, yaitu berkisar antara 25 – 37 ºC. Hal ini dikarenakan suhu optimal untuk kapang Trichoderma viride adalah 32-35°C (Enari, 1983), sedangkan suhu yang optimal untuk kapang Rhizopus oryzae adalah sekitar 25 - 37ºC (Steinkraus et al., 1960).3. Karakteristik Tongkol Jagung Hasil Kultivasi
Setelah dilakukan kultivasi selama 9 hari, maka dilakukan pemanenan terhadap tongkol jagung tersebut. Pemanenan dilakukan dengan cara pengeringan pada suhu 50°C selama satu hari agar mikroba tidak dapat tumbuh dan spora menjadi dorman. Hasil dari kultivasi itu kemudian dilakukan pengujian terhadap parameter perubahan-perubahan komposisi kimia di dalam tongkol jagung dengan menggunakan analisa proksimat. Hasil analisa parameter perubahan komposisi kimia di dalam tongkol jagung sebelum dan setelah kultivasi dapat dilihat pada Tabel 7.
Tabel 7. Karakteristik kimia tongkol jagung hasil kultivasi Komponen
Inokulum Kapang
Rhizopus oryzae Trichoderma viride Umur Simpan (minggu)
4 8 4 8 Air (%b.b) 7,83a 5,18b 7,85a 5,18b Abu (%b.k) 3,65a 3,98a 3,23a 3,99b Protein (%b.k) 6,43a 6,41a 5,51a 5,31a Lemak (%b.k) 3,96a 3,70a 2,37a 2,21a Serat Kasar (%b.k) 45,76a 47,06a 47,08a 47,48a Total Gula (ppm) 91,4a 100,27a 110,52a 114,69a Gula Pereduksi (ppm) 41,03a 31,77a 50,04a 50,5a Derajat Polimerisasi 2,23a 3,16a 2,19a 2,30a Keterangan:
Angka-angka yang diikuti huruf yang sama pada baris yang sama tidak berpengaruh nyata pada taraf 5% menurut uji pembeda T. Perbandingan hanya dibandingkan antar sesama jenis inokulum kapang.
a. Kadar Air
Berdasarkan hasil analisis ragam kadar air (Lampiran 5.A dan Tabel 7) menunjukkan bahwa lama penyimpanan setiap jenis kapang berpengaruh nyata terhadap perubahan kadar air tongkol jagung pada setiap jenis kapang. Kadar air tongkol jagung yang dikultivasi dengan inokulum kapang yang disimpan selama 4 minggu lebih besar bila dibandingkan dengan inokulum kapang yang disimpan selama 8 minggu.
Perbedaan kadar air terjadi akibat proses hidrolisis komponen selulosa substrat oleh enzim selulase yang dihasilkan kapang. Menurut Pelczar et al. (1974), untuk melakukan proses hidrolisis komponen selulosa, kapang membutuhkan air dalam jumlah yang besar. Air tersebut dapat diperoleh dari media tempat tumbuhnya kapang tersebut. Hal inilah yang mengakibatkan kadar air yang dikonsumsi oleh kapang yang disimpan selama 4 minggu lebih besar dibandingkan dengan kapang yang disimpan selama 8 minggu dikarenakan jumlah spora kapang yang semakin berkurang.
39
Pengaruh lain yang mengakibatkan penurunan kadar air tongkol jagung adalah lamanya penyimpanan tongkol jagung tersebut. Semakin lama tongkol jagung disimpan maka dapat mengakibatkan penurunan kadar air di dalam tongkol jagung. Hal ini diperkuat oleh Aprianie (2009), dalam penelitiannya tentang penurunan kadar air pada tongkol jagung didapatkan hasil bahwa apabila tongkol jagung disimpan dalam waktu tertentu akan mengakibatkan penurunan kadar air. Hal ini dikarenakan tongkol jagung tidak bersifat higroskopis, sehingga pada suhu kamar, penguapan akan berlangsung cukup cepat dikarenakan bahan akan mengeluarkan kadar air bebas yang dikandungnya untuk mempertahankan kelembaban dan mencegah kehilangan air yang lebih besar. Hal inilah yang mengakibatkan kadar air tongkol jagung yang telah dikultivasi dengan inokulum yang disimpan selama 8 minggu lebih rendah bila dibandingkan dengan inokulum yang disimpan selama 4 minggu, begitu pula dengan tongkol jagung yang belum dikultivasi.b. Kadar Abu
Berdasarkan hasil analisis ragam kadar abu (Lampiran 5.B dan Tabel 7) menunjukkan bahwa lama penyimpanan inokulum kapang tidak berpengaruh nyata terhadap kadar abu tongkol jagung pada inokulum kapang Rhizopus oryzae sedangkan pada inokulum kapang Trichoderma viride lama penyimpanan inokulum kapang berpengaruh nyata dengan kadar abu tongkol jagung. Hal ini menandakan adanya penurunan kuantitas spora kapang pada inokulum Rhizopus oryzae tidak berpengaruh nyata terhadap kadar abu tongkol jagung sebaliknya pada inokulum Trichoderma viride adanya penurunan kuantitas spora kapang memberikan pengaruh nyata pada penurunan kadar abu tongkol jagung.
Berdasarkan Lampiran 7 diketahui bahwa kadar abu tongkol jagung pada awal sebelum dikultivasi adalah sebesar 3,18%. Adanya penambahan spora kapang mengakibatkan terjadinya peningkatan kadar abu pada tongkol jagung. Peningkatan yang terjadi adalah sebesar 0,47% pada inokulum Rhizopus oryzae yang disimpan selama 4 minggu dan sebesar 0,8% pada inokulum Rhizopus oryzae yang disimpan selama 8 minggu. Peningkatan juga terjadi pada inkulum inokulum Trichoderma viride yaitu sebesar 0,05% pada inokulum yang disimpan selama 4 minggu dan sebesar 0,81% pada inokulum yang disimpan selama 8 minggu.
Adanya penambahan inokulum kapang ke dalam tongkol jagung mengakibatkan terjadinya kenaikan kadar abu pada tongkol jagung. Hal ini dikarenakan menurut Cowan dan Talaro (2009), struktur sel mikroba terdiri atas DNA, nukleus, plasmid dan mitokondria, dinding sel serta unsur pembentuk tubuh lainnya. Unsur-unsur tersebut mengandung mineral yang digunakan untuk menyusun setiap komponen sel di dalam tubuh mikroba. Hal inilah yang mengakibatkan di dalam spora kapang terdapat kandungan mineral yang akan meningkatkan kadar abu di dalam tongkol jagung yang telah dikultivasi.
c. Kadar Protein
Hasil analisis ragam kadar protein (Lampiran 5.C dan Tabel 7) menunjukkan bahwa lama penyimpanan inokulum kapang tidak berpengaruh nyata terhadap kadar protein tongkol jagung pada setiap jenis kapang. Hal ini menandakan adanya penurunan jumlah spora kapang tidak mempengaruhi kadar protein tongkol jagung yang dihasilkan.
Berdasarkan Lampiran 7 diketahui bahwa kadar protein tongkol jagung pada awal sebelum dikultivasi adalah sebesar 2,18%. Adanya penambahan spora kapang mengakibatkan terjadinya peningkatan kadar protein pada tongkol jagung. Peningkatan yang terjadi adalah sebesar 4,25% pada inokulum Rhizopus oryzae yang disimpan selama 4 minggu dan sebesar 4,29% pada inokulum Rhizopus oryzae yang disimpan selama 8 minggu. Peningkatan juga terjadi pada inkulum inokulum Trichoderma viride yaitu sebesar 3,33% pada inokulum yang disimpan selama 4 minggu dan sebesar 3,13% pada inokulum yang disimpan selama 8 minggu.
40
Adanya penambahan substrat bekatul+onggok akan mengakibatkan peningkatan kadar protein pada tongkol jagung menjadi sebesar 2,18% yang dapat dilihat pada Lampiran 7. Selain dikarenakan adanya penambahan substrat bekatul+onggok, kenaikan kadar protein ini juga disebabkan karena adanya peningkatan biomassa kapang. Biomassa kapang mengandung nitrogen sebesar 7 - 10% (Stanbury dan Whitaker, 1984) dan protein sebesar 14 - 44% bobot kering (Griffin, 1981).Kenaikan kadar protein di dalam tongkol jagung dengan kultivasi menggunakan inokulum kapang Rhizopus oryzae lebih tinggi bila dibandingkan dengan menggunakan inokulum kapang Trichoderma viride. Hal ini dikarenakan menurut Gandjar (2006), setiap kapang memiliki kemampuan untuk menghasilkan enzim protease yang dapat menghidrolisis protein menjadi asam amino yang dibutuhkan oleh kapang untuk pertumbuhan. Kapang yang memiliki kemampuan untuk menghasilkan enzim protease yang cukup baik adalah kapang Rhizopus oryzae. Menurut Sumanti (2005), adanya proses perombakan protein pada substrat menjadi asam-asam amino akan meningkatkan komposisi nitrogen terlarut di dalam bahan. Hal inilah yang mengakibatkan kapang Rhizopus oryzae menghasilkan protein yang lebih baik bila dibandingkan dengan kapang Trichoderma viride.
Adanya peningkatan kadar protein pada tongkol jagung akan mendukung kegunaan tongkol jagung sebagai pakan ternak. Menurut Prihatman (2000), kandungan protein yang dapat memenuhi kebutuhan ternak ruminansia adalah sebesar 8 %. Jika dilihat dari kenaikan kadar protein di dalam tongkol jagung yang telah dikultivasi, maka diketahui bahwa tongkol jagung tersebut masih belum layak digunakan sebagai pakan ternak tunggal. Penggunaan tongkol jagung yang telah dikultivasi sebagai pakan ternak perlu ditambahkan dengan sumber protein tambahan, sehingga dapat mencukupi kebutuhan protein pada ternak.
d. Kadar Lemak
Berdasarkan analisis ragam kadar lemak (Lampiran 5.D dan Tabel 7) menunjukkan bahwa lama penyimpanan inokulum kapang tidak berpengaruh nyata terhadap kadar lemak tongkol jagung pada setiap jenis kapang. Hal ini menandakan adanya penurunan jumlah spora kapang tidak mempengaruhi kadar lemak tongkol jagung yang dihasilkan.
Berdasarkan Lampiran 7 diketahui bahwa kadar lemak tongkol jagung pada awal sebelum dikultivasi adalah sebesar 3,97%. Adanya penambahan spora kapang mengakibatkan terjadinya penurunan kadar lemak pada tongkol jagung. Penurunan yang terjadi adalah sebesar 0,01% pada inokulum Rhizopus oryzae yang disimpan selama 4 minggu dan sebesar 0,27% pada inokulum Rhizopus oryzae yang disimpan selama 8 minggu. Penurunan juga terjadi pada inkulum inokulum Trichoderma viride yaitu sebesar 1,59% pada inokulum yang disimpan selama 4 minggu dan sebesar 1,75% pada inokulum yang disimpan selama 8 minggu.
Penurunan kadar lemak pada tongkol jagung terjadi dikarenakan kapang dapat menghasilkan lipase, sehingga dapat mendegradasi lemak untuk menjadi sumber karbon (Gandjar, 2006). Menurut Rapp dan Backhaus (1992), kapang melakukan degradasi lemak dengan cara mensekresikan enzim lipase ke bahan untuk mengubah lemak tersebut sebelum dimasukkan ke dalam sel. Proses degradasi lemak tersebut dilakukan dengan cara pemutusan ikatan gliserol pada triasilgliserol menjadi diasilgliserol, monogliserol serta asam lemak dan gliserol (Ratledge dan Tan, 1990). Hal inilah yang mengakibatkan kapang dapat mendegradasi lapisan lemak yang ada di dalam bahan untuk sumber energi meskipun tidak terlalu besar.
e. Kadar Serat Kasar
Berdasarkan analisis ragam kadar serat kasar (Lampiran 5.E dan Tabel 7) menunjukkan bahwa lama penyimpanan inokulum kapang tidak berpengaruh nyata terhadap kadar serat kasar tongkol jagung pada setiap jenis kapang. Hal ini menandakan adanya
41
penurunan jumlah spora kapang tidak mempengaruhi kadar serat kasar tongkol jagung yang dihasilkan.Berdasarkan Lampiran 7 diketahui bahwa kadar serat kasar tongkol jagung pada awal sebelum dikultivasi adalah sebesar 77,11%. Adanya penambahan spora kapang mengakibatkan terjadinya penurunan kadar serat kasar pada tongkol jagung. Penurunan yang terjadi adalah sebesar 31,35% pada inokulum Rhizopus oryzae yang disimpan selama 4 minggu dan sebesar 30,05% pada inokulum Rhizopus oryzae yang disimpan selama 8 minggu. Penurunan juga terjadi pada inkulum inokulum Trichoderma viride yaitu sebesar 30,03% pada inokulum yang disimpan selama 4 minggu dan sebesar 29,63% pada inokulum yang disimpan selama 8 minggu.
Serat kasar terdiri atas lignin, selulosa dan hemiselulosa yang saling berikatan yang disebut sebagai lignoselulosa. Ikatan yang sangat kompleks ini mengakibatkan kapang selulolitik tidak dapat melakukan hidrolisis terhadap selulosa dan hemiselulosa di dalam bahan dengan sempurna (Orth et al., 1993). Adanya penurunan kadar serat tongkol jagung yang kurang maksimal dikarenakan ada beberapa bagian dalam tongkol jagung yang berikatan dengan lignin sehingga menjadi ikatan lignoselulosa.
Kapang selulolitik dapat menghidrolisis serat di dalam bahan, serat yang dapat dihidrolisis adalah selulosa dan hemiselulosa. Menurut Perez et al. (2002), selulosa merupakan polimer glukosa dengan ikatan β-1,4 glukosida dalam rantai lurus. Selulosa dapat dihidrolisis dengan menggunakan enzim selulase. Selulase merupakan enzim yang termasuk dalam kelompok enzim yang terdiri atas endoglukanase, eksoglukanase dan β-glukosidase (Maheshwari, 2005). Endoglukanase merupakan enzim yang dapat menghidrolisis daerah amorf selulosa secara acak. Enzim ini dapat membentuk oligosakarida dengan panjang rantai yang berbeda-beda dan membentuk ujung rantai non-pereduksi (Sinegani dan Emtiazi, 2006). Eksoglukanase merupakan komponen enzim yang produk hidrolisis utamanya adalah selobiosa. Enzim ini memecah selulosa dengan cara menghilangkan ujung akhir gugus selobiosa pada rantai selulosa (Raja dan Shafiq-Ur-Rehman, 2008). β-glukosidase merupakan enzim yang dapat memutuskan unit glukosa secara spesifik dari ujung nonpereduksi dari selo-oligosakarida (Sinegani dan Emtiazi, 2006). Hal inilah yang mengakibatkan selulosa dapat dihidrolisis oleh kapang selulolitik menjadi glukosa dan gula-gula sederhana.
Hemiselulosa tersusun atas ikatan β 1-4 pentosa yang berikatan dengan ikatan hidrogen terhadap selulosa dan ikatan kovalen terhadap lignin (Maheshwari, 2005). Hemiselulosa merupakan polisakarida yang mempunyai tingkat degradasi lebih baik bila dibandingkan selulosa dan lignin (Suparjo, 2008). Menurut Tjokroadikoesoemo (1986), Hemiselulosa merupakan polimer dari sejumlah sakarida yang berbeda-beda, yaitu: D-xilosa, L-arabinosa, D-galaktosa, D-glukosa, dan D-glukoronat. Berbeda dengan selulosa, hemiselulosa berbentuk amorf. Hal ini mengakibatkan hemiselulosa di dalam bahan lebih mudah dihidrolisis oleh kapang selulolitik. Adanya serat yang dihidrolisis mengakibatkan glukosa dan gula-gula sederhana di dalam bahan bertambah.
Penurunan serat kasar pada tongkol jagung yang dikultivasi dengan kapang Trichoderma viride serta tongkol jagung yang dikultivasi dengan kapang Rhizopus oryzae tidak terlalu berbeda dikarenakan kedua jenis kapang tersebut hanya dapat menghidrolisis sebagian dari serat tongkol jagung akibat adanya ikatan lignoselulosa. Hidrolisis yang tidak sempurna ini mengakibatkan terjadinya kesulitan dalam membedakan kemampuan kedua jenis kapang selulotik dalam menghidrolisis serat kasar di dalam tongkol jagung.
f. Total Gula
Berdasarkan analisis ragam total gula (Lampiran 5.F dan Tabel 7) menunjukkan bahwa lama penyimpanan inokulum kapang tidak berpengaruh nyata terhadap total gula tongkol jagung pada setiap jenis kapang. Hal ini menandakan adanya penurunan jumlah spora kapang tidak mempengaruhi total gula tongkol jagung yang dihasilkan.
Total gula di dalam tongkol jagung yang dikultivasi dengan menggunakan inokulum kapang Rhizopus oryzae yang disimpan selama 4 minggu dan 8 minggu mengalami penurunan sebesar 53,6 ppm dan 44,73 ppm, sedangkan kultivasi dengan menggunakan inokulum kapang Trichoderma viride yang disimpan selama 4 minggu dan 8 minggu
42
mengalami penurunan sebesar 34,48 ppm dan 30,31 ppm. Hal ini dikarenakan kapang mengkonsumsi gula yang ada di dalam substrat, sehingga berkurangnya jumlah spora kapang mengakibatkan penurunan total gula dalam tongkol jagung menjadi makin rendah. Hal ini diperkuat oleh Gandjar (2006) yang mengatakan bahwa kapang membutuhkan nutrisi-nutrisi untuk pertumbuhannya, seperti gula, lemak, protein dan zat-zat kimia lainnya yang diambil dari substrat.Total gula pada tongkol jagung yang dikultivasi dengan kapang Trichoderma viride lebih besar bila dibandingkan dengan kapang Rhizopus oryzae. Hal ini dikarenakan dalam pengukuran total gula semua gula sederhana, oligosakarida dan turunannya dapat bereaksi dengan fenol dalam asam sulfat pekat, sehingga hemiselulosa, selulosa dan turunannya ikut bereaksi dengan fenol, sehingga menghasilkan total gula yang lebih tinggi. Hal ini dapat dilihat dari lebih besarnya kandungan serat kasar tongkol jagung yang dihidrolisis kapang Trichoderma viride dibandingkan dengan tongkol jagung yang dihidrolisis kapang Rhizopus oryzae.
g. Gula Pereduksi
Berdasarkan analisis ragam gula pereduksi (Lampiran 5.G dan Tabel 7) menunjukkan bahwa lama penyimpanan inokulum kapang tidak berpengaruh nyata terhadap kadar gula pereduksi tongkol jagung pada setiap jenis kapang. Hal ini menandakan adanya penurunan jumlah spora kapang tidak mempengaruhi kadar gula pereduksi tongkol jagung yang dihasilkan.
Kenaikan gula pereduksi pada tongkol jagung yang dikultivasi dengan kapang Rhizopus oryzae adalah sebesar 34,04 ppm pada inokulum yang disimpan selama empat minggu dan sebesar 24,77 ppm pada inokulum yang disimpan selama delapan minggu. Kenaikan gula pereduksi juga terjadi pada tongkol jagung yang dikultivasi dengan kapang Trichoderma viride yaitu sebesar 43,04 ppm pada inokulum yang disimpan selama empat minggu dan sebesar 43,5 ppm pada inokulum yang disimpan selama delapan minggu.
Kenaikan kadar gula pereduksi di dalam tongkol jagung yang telah dikultivasi dikarenakan adanya hidrolisis terhadap karbohidrat kompleks yang terdapat di dalam bahan menjadi glukosa dan gula-gula sederhana lainnya. Gula pereduksi terdiri atas gula-gula sederhana seperti monosakarida (glukosa, fuktosa, galaktosa) dan disakarida (laktosa dan maltosa). Hal inilah yang mengakibatkan terjadinya kenaikan kadar gula pereduksi di dalam tongkol jagung yang telah dikultivasi. Kenaikan gula pereduksi di dalam tongkol jagung yang telah dikultivasi, juga dapat dilihat dari turunnya kadar serat kasar pada tongkol jagung tersebut. Hal ini dikarenakan serat kasar terdiri atas karbohidrat kompleks, sehingga karbohidrat tersebut terhidrolisis menjadi gula-gula sederhana.
Kenaikan gula pereduksi pada tongkol jagung yang dihidrolisis dengan kapang Trichoderma viride lebih tinggi dibandingkan dengan kapang Rhizopus oryzae. Hal ini dikarenakan kapang Trichoderma viride memiliki kemampuan yang lebih besar dalam menghidrolisis kandungan polisakarida di dalam substrat menjadi gula sederhana. Hal ini diperkuat oleh Mandels (1982) yang mengatakan bahwa Trichoderma viride dapat menghasilkan enzim kompleks selulase yang kemampuan untuk memecah selulosa menjadi glukosa sehingga mudah dicerna oleh ternak.
h. Derajat Polimerisasi
Berdasarkan analisis ragam derajat polimerisasi (Lampiran 5.H dan Tabel 7) menunjukkan bahwa lama penyimpanan inokulum kapang tidak berpengaruh nyata terhadap derajat polimerisasi tongkol jagung pada setiap jenis kapang. Hal ini menandakan adanya penurunan jumlah spora kapang tidak mempengaruhi derajat polimerisasi tongkol jagung yang dihasilkan.
Tongkol jagung yang dikultivasi dengan menggunakan inokulum kapang Rhizopus oryzae terjadi penurunan DP menjadi 2,23 pada inokulum yang disimpan selama empat hari
43
dan sebesar 3,16 pada inokulum yang disimpan selama delapan hari. Penurunan DP juga terjadi pada tongkol jagung yang dikultivasi dengan menggunakan inokulum kapang Trichoderma viride yaitu menjadi sebesar 2,19 pada inokulum yang disimpan selama empat hari dan sebesar 2,30 pada inokulum yang disimpan selama delapan hari.Penurunan derajat polimerisasi di dalam tongkol jagung menjadi antara 2 sampai 20 menunjukkan dihasilkannya gula dalam bentuk oligosakarida (Deman, 1997). Penurunan ini diakibatkan adanya perubahan selulosa dan hemiselulosa menjadi gula sederhana akibat enzim selulase yang dihasilkan oleh inokulum kapang. Pembentukan gula oligosakarida yang dihasilkan memberikan pengaruh terhadap peningkatan nilai nutrisional di dalam tongkol jagung. Hal ini dikarenakan menurut Van Der Kapm et al. (2004), oligosakarida digunakan sebagai alternatif pengganti antibiotik dalam usaha peningkatan produksi ternak dikarenakan berfungsi sebagai senyawa prebiotik. Prebiotik merupakan zat yang digunakan oleh mikroba yang untuk sumber makanan bagi pertumbuhan mikroba tersebut. Menurut Fahey et al. (2004), prebiotik yang berbasis oligosakarida dapat membantu pertumbuhan populasi mikroba yang ada di dalam saluran pencernaan pada ternak, sehingga saluran pencernaan pada ternak dapat berjalan dengan baik.