AUTEKOLOGI DAN STUDI POPULASI
Myristica teijsmannii Miq. (Myristicaceae)
DI CAGAR ALAM PULAU SEMPU, JAWA TIMUR
ROSNIATI APRIANI RISNA
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
BOGOR
2009
AUTEKOLOGI DAN STUDI POPULASI
Myristica teijsmannii Miq. (Myristicaceae)
DI CAGAR ALAM PULAU SEMPU, JAWA TIMUR
ROSNIATI APRIANI RISNA
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains pada
Program Studi Biologi
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
BOGOR
2009
PERNYATAAN MENGENAI TESIS
DAN SUMBER INFORMASI
Dengan ini saya menyatakan bahwa tesis ”Autekologi dan Studi Populasi
Myristica teijsmannii Miq. (Myristicaceae) di Cagar Alam Pulau Sempu, Jawa
Timur” adalah karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apapun kepada perguruan tinggi manapun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan Daftar Pustaka di bagian akhir tesis ini.
Bogor, Februari 2009
Rosniati Apriani Risna
ABSTRACT
ROSNIATI APRIANI RISNA. Autecology and Population Study of Myristica
teijsmannii Miq. (Myristicaceae) in Sempu Island Nature Reserve, East Java.
Under supervision of DEDE SETIADI and DIDIK WIDYATMOKO.
In an attempt to determine ecological characteristics and current population status of Myristica teijsmannii Miq., an endangered species under the IUCN, the autecology and population study of the species was conducted in six localities within Sempu Island Nature Reserve, East Java. Vegetation analysis was employed to determine vegetation and population structure, as well as interspecific association within communities. Both physical and chemical analysis of soil and topographical observation were carried out to identify environmental factors with the greatest influence on the presence of the species. The study revealed that M. teijsmannii distributed very frequently with the highest tree density of 14 individuals/ha covering a total of 82 individuals within 6 ha area studied with a clumped distribution pattern throughout the island, and was associated with 12 tree species. Overall population was found in a good structure showing domination of younger stages and decreasing density as plants grow. Individual numbers of tree, pole and sapling stages were found significantly correlated with sand content whilst abundance parameter of the sapling stage showed a strong correlation with carbon, nitrogen and K contents. The species favored flat areas (0-8% slope) with north-west aspect preference, suggesting that the species was reasonably semi-shade tolerant. This study also identified some threats to the populations and discussed some implications of the biological and habitat preferences of the species to develop a proper conservation strategy. Keywords: Myristica teijsmannii, autecology, population study, Sempu Island
RINGKASAN
ROSNIATI A. RISNA. Autekologi dan Studi Populasi Myristica teijsmannii Miq. (Myristicaceae) di Cagar Alam Pulau Sempu, Jawa Timur. Dibimbing oleh DEDE SETIADI dan DIDIK WIDYATMOKO.
Myristicaceae merupakan famili khas tropis dengan Myristica sebagai
genus terbesar. Di antara 175 spesies Myristica di dunia 9 spesies merupakan spesies asli Indonesia bahkan beberapa termasuk endemik, langka dan dilindungi undang-undang. Myristica teijsmannii Miq. atau dikenal dengan nama Durenan, Palan, Kosar atau Kayu Resep merupakan salah satu spesies yang termasuk ke dalam kategori endangered atau genting menurut IUCN dengan kriteria EN B1+2C. Spesies ini memiliki penyebaran jarang dan baru dilaporkan ditemukan di Jawa Timur, yaitu di kawasan Pacitan - Gunung Kawi, Gunung Wilis, Gunung Anjasmoro dan Pulau Sempu. Data kuantitatif status populasi serta aspek-aspek ekologis, kebutuhan dan interaksi ekologis M. teijsmannii dengan habitatnya (autekologi) belum diketahui. Berdasarkan hal tersebut, maka penelitian ini dilakukan untuk mempelajari populasi, karakteristik habitat, interaksi dan kebutuhan-kebutuhan ekologis M. teijsmannii dengan memfokuskan penelitian di Pulau Sempu yang berstatus sebagai cagar alam. Studi ini dilakukan di enam lokasi yang secara estimasi visual mewakili keragaman floristik dan kondisi lingkungan Cagar Alam, yaitu Telaga Lele, Telaga Sat, Teluk Semut, Air Tawar, Gua Macan, dan Waru-waru.
Berdasarkan hasil analisis vegetasi diketahui bahwa tegakan pohon
M. teijsmannii menunjukkan nilai dominasi relative dan kerapatan tertinggi di
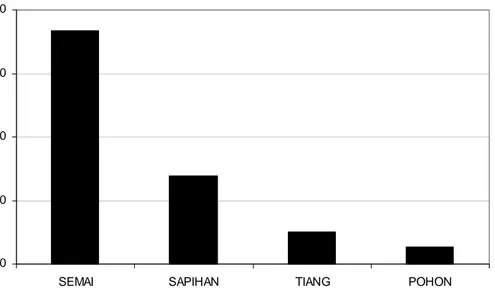
lokasi penelitian, berturut-turut sebesar 11,36% dan 13,7 individu/ha dengan indeks nilai penting sebesar 27,91% yang menunjukkan dominasi M. teijsmannii. Struktur populasinya secara keseluruhan didominasi oleh fase semai, sedangkan fase-fase yang lebih dewasa memperlihatkan kecenderungan jumlah individu yang semakin menurun dengan fase pohon memiliki proporsi terendah. Setiap lokasi penelitian memperlihatkan variasi dalam struktur populasi M. teijsmannii. Waru-waru memiliki jumlah total individu terbanyak (59 individu) sedangkan di Telaga Lele hanya ditemukan 5 individu. Di Waru-waru, Gua Macan dan Teluk Semut nampak jelas dominasi fase semai dengan kecenderungan menurun pada populasi fase lebih dewasa. Di Telaga Lele dan Telaga Sat tidak ditemukan satu pun individu semai dalam plot penelitian. Hal ini menunjukkan bahwa rekruitmen semai tidak merata di setiap lokasi, berkaitan dengan faktor lingkungan yang mempengaruhinya. Dilihat dari segi gangguan manusia, kawasan Teluk Semut secara umum mendapat ancaman yang lebih besar karena hutan di kawasan ini merupakan jalur utama bagi tujuan wisata. Jalan setapak semakin melebar membuat gap di hutan juga semakin lebar sehingga dapat mengakibatkan efek tepi (edge effect) yang lebih meluas ke dalam hutan sehingga dapat merugikan pertumbuhan populasi M. teijsmannii.
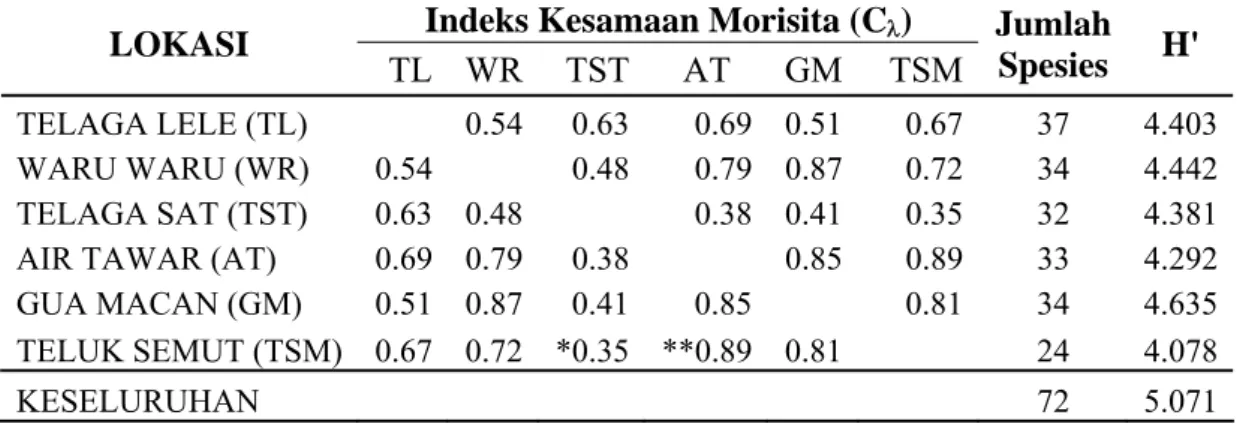
M. teijsmannii menyebar secara mengelompok dengan indeks penyebaran
Morisita (Ip) 0,51 dan berasosiasi positif dengan 12 spesies pohon pada tingkat asosiasi yang bervariasi. Tingkat asosiasi yang cukup tinggi berdasarkan Jaccard Index ditunjukkan oleh Pterospermum javanicum (0,529), Cryptocarya ferrea (0,500), Orophea hexandra (0,455), dan Aglaia elliptica (0,467). Agen dispersal bijinya adalah lutung Jawa (Trachypitecus auratus) dan kera ekor panjang (Macaca fascicularis). Keberadaan kedua spesies tersebut perlu dilindungi agar kelangsungan populasi M. teijsmannii juga dapat terjaga.
Hasil analisis korelasi antara parameter kemelimpahan spesies dan faktor fisika serta kimia tanah menunjukkan bahwa jumlah individu fase sapihan, tiang dan pohon M. teijsmannii berkorelasi kuat pada tanah yang memiliki kandungan pasir lebih tinggi. Ini menjadi argumen mengapa Waru-waru dikoloni individu
M. teijsmannii lebih banyak daripada lokasi lainnya karena lokasi ini memiliki
kandungan pasir lebih tinggi dibandingkan dengan lokasi lainnya. Fase sapihan memiliki korelasi kuat tidak saja dengan kandungan pasir tetapi juga dengan kandungan C, N dan K. Ini menunjukkan faktor kesuburan tanah (rasio C dan N) menentukan seedling recruitment dan seedling establishment menuju fase sapihan yang lebih dewasa.
Secara umum pH di lokasi penelitian bersifat netral (6,0 – 7,0) dengan rataan suhu 24-260C dan kelembaban tanah 45-86%. Suhu udara pada pengukuran sekitar pukul 08.00-14.00 tercatat antara 22-300C dengan kisaran kelembaban udara 53-92%. Teluk Semut memperlihatkan nilai kelembaban udara yang relatif tinggi, antara lain disebabkan oleh kondisi hutan di kawasan tersebut yang relatif terang dan memiliki lebih banyak gap kanopi. Kondisi ini kemungkinan menjadi salah satu penyebab kesintasan semai di kawasan ini sangat rendah.
Pengujian statistik untuk model distribusi populasi M. teijsmannii dengan menggunakan metode Generalized Linear Model (GLM) terhadap pengaruh faktor topografis memberikan ketepatan model atau deviansi sebesar 1,10 yaitu berkategori baik dengan nilai signifikan pada lereng dan arah lereng. Hasil ini menunjukkan bahwa distribusi populasi M. teijsmannii ditentukan oleh lereng dan arah lereng tetapi tidak oleh ketinggian. Kehadiran individu M. teijsmannii secara signifikan dijumpai lebih banyak pada kawasan datar dengan kemiringan 0-8%, sedangkan pada kemiringan agak curam (15-25%) dijumpai populasi yang rendah.
M. teijsmannii ditemukan pada seluruh kisaran kelas ketinggian di CA Pulau
Sempu, tepatnya pada 25–86 m dpl sehingga variabel ketinggian lokasi tidak memberikan perbedaan yang berarti secara statistik terhadap kehadiran individu. Jumlah individu dalam plot pengamatan cenderung semakin banyak pada arah lereng yang tidak mengarah langsung ke sebelah timur, yaitu pada utara-barat. Sebaliknya pada aspek yang langsung mengarah ke timur, jumlah individu ditemukan semakin sedikit. Jumlah individu terbanyak ditemukan pada lereng yang tidak memperoleh sinar matahari sepanjang hari. Fenomena ini mendukung dugaan bahwa spesies ini memiliki sifat semi toleran.
Pihak pengelola Resort CA Pulau Sempu perlu menekan dampak aktivitas pengunjung terhadap keaslian dan kealamian habitat di kawasan ini. Kawasan Teluk Semut perlu mendapat pengawasan dan pengelolaan yang lebih baik karena ada ketidakseimbangan populasi M. teijsmannii sebagai tumbuhan langka Jawa
Timur. Selain itu, lokasi ini juga merupakan jalur jelajah lutung sebagai agen dispersal biji, dan merupakan habitat bagi macan tutul (hewan yang juga dilindungi) seperti yang teramati dalam penelitian.
@ Hak Cipta milik IPB, tahun 2009 Hak cipta dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumber.
a. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik atau tinjauan suatu masalah.
b. Dilarang mengumumkan dan memperbanyak sebagian atau seluruh karya tulis dalam bentuk apapun tanpa izin IPB.
Judul Tesis : Autekologi dan Studi Populasi Myristica teijsmannii Miq. (Myristicaceae) di Cagar Alam Pulau Sempu, Jawa Timur Nama : Rosniati Apriani Risna
NIM : G351060501
Disetujui Komisi Pembimbing
Prof. Dr. Dede Setiadi, MS. Dr. Didik Widyatmoko, M.For.Sc
Ketua Anggota
Diketahui
Ketua Program Studi Biologi Dekan Sekolah Pascasarjana
Dr. Dedy Duryadi Solihin, D.E.A. Prof. Dr. Ir. Khairil A. Notodiputro
PRAKATA
Puji dan syukur penulis panjatkan kepada Allah SWT atas segala karuniaNya sehingga tesis ini berhasil diselesaikan. Tema yang dipilih dalam penelitian yang dilaksanakan sejak bulan Mei 2007 ini adalah autekologi tumbuhan langka dan endemik, dengan judul Autekologi dan Studi Populasi
Myristica teijsmannii Miq. (Myristicaceae) di Cagar Alam Pulau Sempu, Jawa
Timur.
Tema ini dipilih sesuai dengan kecintaan terhadap bidang ekologi dan ketertarikan penulis pada famili Myristicaceae (pala-palaan) yang merupakan salah satu famili khas kawasan tropis. Anggota famili ini banyak yang merupakan spesies asli Indonesia, sekaligus banyak pula di antaranya yang dikategorikan rentan (vulnerable) terhadap kepunahan bahkan genting (endangered) menurut versi IUCN. Penelitian yang dilakukan ini juga selaras dengan pekerjaan penulis dalam bidang reintroduksi tumbuhan langka di Pusat Konservasi Tumbuhan, Kebun Raya Bogor.
Terima kasih penulis ucapkan kepada Lembaga Ilmu Pengetahuan Indonesia (LIPI) yang telah memberikan beasiswa serta Departemen Pendidikan Nasional, Yayasan Konservasi Flora dan Yayasan Damandiri atas bantuan dana penelitiannya sehingga penelitian ini dapat dilakukan dan ditulis menjadi sebuah tesis. Penghargaan setinggi-tingginya disampaikan pada Bapak Prof. Dr. Dede Setiadi, MS. dan Dr. Didik Widyatmoko, M.For.Sc selaku pembimbing, serta Bapak Suhirman, PhD. yang telah banyak memberi masukan bagi penelitian ini. Penulis juga menghaturkan terima kasih kepada para teknisi Kebun Raya (Cecep Suryana, Dwinarko, Matrani dan Ruspandi) yang banyak membantu penelitian di Cagar Alam Pulau Sempu dan mengidentifikasi spesimen herbarium di Herbarium Bogoriense, rekan-rekan di INetPC serta Didit Okta Pribadi, MS. atas dukungan dan saran-saran mereka dalam analisis dan penulisan. Bantuan dari para staf resort Cagar Alam Pulau Sempu Wilayah BKSDA II Jatim, para asisten dosen mata kuliah GIS di Fakultas Kehutanan dan Ekologi Lansekap di PPLH juga sangat penulis hargai.
Ucapan terima kasih juga disampaikan kepada suami tercinta, Tata M. Syaid, putra tersayang Naufal Oktafiandri Nursaid, serta kepada Ibu, Bapak, Mimih, Bapa, Ade, Ida dan seluruh keluarga besar atas segala motivasi, do’a dan kasih sayang mereka.
Semoga karya ilmiah ini bermanfaat, terutama bagi program konservasi
M. teijsmannii dan membantu pencapaian target Strategi Global Konservasi
Tumbuhan.
Bogor, Februari 2009
RIWAYAT HIDUP
enulis dilahirkan di Bandung pada tanggal 17 April 1974 dari pasangan M. Rosjid Abdurachman M.Sc dan Nia Suniarsih, sebagai anak sulung dari dua bersaudara.
P
Setelah tamat Sekolah Menengah Atas di SMA Negeri 3 Bandung pada tahun 1992, penulis melanjutkan studi S1 di Jurusan Biologi FMIPA Universitas Padjadjaran Bandung hingga tahun 1997 lulus dengan predikat cum laude.
Tahun 1997 – 2001 penulis bekerja sebagai asisten peneliti di sebuah yayasan swasta yang bergerak dalam konservasi flora dan bertugas melakukan penelitian, menangani database dan koleksi Polyporaceae.
Pada tahun 2000 penulis diterima bekerja sebagai peneliti di UPT Kebun Raya Bogor, Lembaga Ilmu Pengetahuan Indonesia (sekarang bernama Pusat Konservasi Tumbuhan Kebun Raya Bogor). Mulai tahun 2002 penulis dikhususkan bekerja di bagian reintroduksi tumbuhan langka di bawah bidang Konservasi Ex-situ di PKT Kebun Raya Bogor.
Sejak tahun 2004 penulis merangkap sebagai volunteer di sebuah gugus tugas PKT Kebun Raya Bogor, yaitu Indonesian Network for Plant Conservation (INetPC), suatu jaringan dan wadah komunikasi bagi aktivis konservasi tumbuhan yang beranggotakan individu dan organisasi dalam dan luar negeri. Penulis di bawah koordinatornya bertanggung jawab terhadap keberlangsungan kegiatan rutin INetPC dan menjadi salah satu editor majalah Eksplorasi, majalah bilingual triwulanan yang diterbitkan INetPC.
Beberapa konferensi internasional telah penulis ikuti sebagai pembicara. Tahun 2000 penulis mendapatkan small grant for young scientist dari Hong Kong University untuk mempresentasikan penelitiannya dalam Asian Mycological Congress (AMC) 2000. Kemudian tahun 2003 penulis menjadi presenter makalah ilmiah dalam International Botanic Gardens Conference di Bedugul, Bali.
Pada tahun 2005 penulis mendapatkan kesempatan mengikuti Applied Plant Conservation Internship Training Program di Denver Botanic Gardens, Colorado, AS selama 10 minggu. Setahun berselang, penulis berhasil mendapatkan beasiswa dari LIPI untuk melanjutkan studi S2 di Program Studi Biologi, Sub program Ekologi, Sekolah Pascasarjana Institut Pertanian Bogor.
DAFTAR ISI
Halaman
DAFTAR TABEL ...iv
DAFTAR GAMBAR ... v
DAFTAR LAMPIRAN ...vi
I. PENDAHULUAN ... 1
1.1 Latar Belakang ... 1
1.2 Tujuan Penelitian ... 4
1.3 Manfaat Penelitian ... 4
II. TINJAUAN PUSTAKA ... 5
2.1. Myristica teijsmannii Miq. ... 5
2.1.1. Nomenklatur ... 5
2.1.2. Nama-nama Lokal ... 5
2.1.3. Morfologi ... 5
2.1.4. Klasifikasi ... 6
2.1.5. Distribusi dan Ekologi ... 6
2.1.6. Pertumbuhan dan Perkembangan ... 7
2.1.7. Pola Penyebaran ... 8
2.1.8. Asosiasi Interspesifik ... 8
2.1.9. Status Kelangkaan dan Konservasi ... 10
2.1.10. Aspek Pemanfaatan ... 11
2.2. Kategori Kelangkaan ... 11
2.3. Istilah-istilah yang digunakan ... 15
III. METODOLOGI ... 17
3.1. Waktu dan Tempat Penelitian ... 17
3.2. Bahan dan Alat ... 17
3.3. Metode Penelitian ... 18
3.3.1. Autekologi M. teijsmannii ... 18
3.3.2. Pengambilan Data Karakteristik Habitat ... 18
3.3.2.1. Penentuan Lokasi Penelitian ... 18
3.3.2.2. Pengukuran di Lapangan ... 19 i
3.3.2.3. Pengambilan Contoh Tanah ... 20
3.3.2.4. Analisis Contoh Tanah ... 20
3.4. Analisis Data ... 21
3.4.1. Autekologi dan Populasi M. teijsmannii serta Struktur Komunitas ... 21
3.4.1.1. Kerapatan ... 22
3.4.1.2. Frekuensi ... 22
3.4.1.3. Dominasi ... 22
3.4.1.4. Indeks Nilai Penting ... 23
3.4.1.5. Indeks Keragaman ... 23
3.4.1.6. Indeks Kesamaan Komunitas ... 23
3.4.1.7. Pola Penyebaran ... 25
3.4.1.8. Peta Penyebaran ... 26
3.4.1.9. Asosiasi Interspesifik ... 28
3.4.2. Karakteristik Habitat M. teijsmannii berdasarkan Variabel Lingkungan ... 28
3.4.3. Interaksi M. teijsmannii dengan Variabel Lingkungan ... 28
3.4.4. Diagram Alir Penelitian ... 30
IV. KONDISI UMUM KAWASAN ... 31
4.1. Letak Geografis, Batas–batas Administratif dan Status Kawasan .. 31
4.2. Kondisi Iklim ... 33
4.3. Geologis dan Hidrologis ... 33
4.4. Kondisi Biologis ... 34
4.5. Aksesibilitas ... 35
V. HASIL DAN PEMBAHASAN... 37
5.1. Struktur Komunitas Vegetasi di Cagar Alam Pulau Sempu ... 37
5.2. Autekologi M. teijsmannii ... 38
5.2.1. Struktur Populasi ... 38
5.2.2. Pola Penyebaran ... 43
5.2.3. Pertumbuhan dan Perkembangan ... 44
5.2.4. Reproduksi ... 46
5.2.5. Interaksi dengan Komponen Biologis ... 48
5.2.5.1. Asosiasi Interspesifik ... 48
5.2.5.2. Predasi dan Agen Dispersal Biji ... 48
5.2.6. Karakteristik Habitat ... 52
5.2.6.1. Faktor Edafik ... 52
5.2.6.2. Faktor Klimatik ... 60
5.2.6.3. Faktor Topografis ... 60
5.3. Aspek Konservasi M. teijsmannii ... 62
5.3.1. Implikasi Karakter dan Interaksi Biologis M. teijsmannii terhadap Konservasi ... 62
5.3.2. Implikasi Karakter dan Preferensi Habitat M. teijsmannii terhadap Konservasi ... 64
5.3.3. Status Kelangkaan ... 65
5.3.4. Potensi dan Ancaman ... 67
VI. KESIMPULAN DAN SARAN... 70
6.1. Kesimpulan ... 70
6.2. Saran – saran ... 71
DAFTAR PUSTAKA ... 72
LAMPIRAN – LAMPIRAN ... 76
DAFTAR TABEL
Halaman 1 Asosiasi di antara spesies akibat adanya interaksi antar spesies dan
proses ekologis ... 9
2 Kriteria status kelangkaan untuk kategori kritis, genting dan rentan ... 14
3 Kriteria penilaian Sifat Kimia Tanah ... 21
4 Matriks data kehadiran dan ketidakhadiran dari S spesies dalam N unit sampling ... 26
5 Tabel kontingensi 2 x 2 untuk asosiasi spesies ... 28
6 Klasifikasi variabel-variabel yang digunakan dalam uji statistik... 29
7 Data curah hujan CA Pulau Sempu tahun 2002-2006... 33
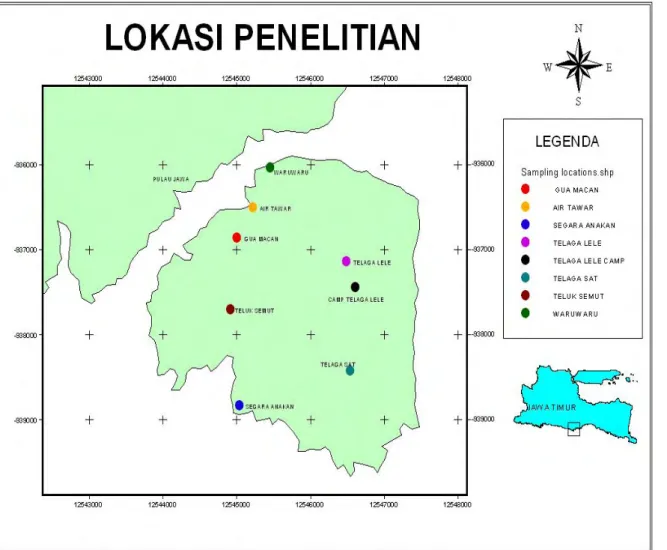
8 Posisi dan kondisi lokasi penelitian ... 36
9 Indeks kesamaan, jumlah spesies dan indeks keragaman pohon di lokasi penelitian Cagar Alam Pulau Sempu ... 37
10 Parameter kemelimpahan pohon M. teijsmannii di lokasi penelitian... 41
11 Hasil uji asosiasi interspesifik antara M. teijsmannii dengan 14 spesies di CA Pulau Sempu berdasarkan tes chi-square... 49
12 Hasil analisis sifat fisika dan kimia tanah di lokasi penelitian ... 55
13 Nilai korelasi antara parameter kemelimpahan M. teijsmannii dan variabel edafik di lokasi penelitian... 59
14 Data iklim mikro pada tanah dan udara di lokasi penelitian ... 60
15 Perbandingan variabel topografi dengan kehadiran dan ketidakhadiran M. teijsmannii di CA Pulau Sempu ... 61
16 Ringkasan hasil pengujian generalized linear model (GLM) untuk distribusi populasi M. teijsmannii ... 61
17 Kemelimpahan M. teijsmannii dan karakteristik lingkungannya ... 63
18 Potensi dan ancaman terhadap kawasan konservasi CAPS dan keberadaan M. teijsmannii ... 69
DAFTAR GAMBAR
Halaman
1 Skema pembuatan plot pada sampling ... 19
2 Skema plot bersarang pada plot 20 x 20 m ... 19
3 Diagram alir penelitian ... 30
4 Peta lokasi penelitian di CA Pulau Sempu ... 31
5 Perbatasan CA Pulau Sempu dan Pulau Jawa dilihat dari Selat Sempu .... 32
6 Batas selatan CA Pulau Sempu dengan Samudera Indonesia ... 32
7 Struktur populasi M. teijsmannii di seluruh lokasi penelitian dalam kawasan CA Pulau Sempu berdasarkan diameter setinggi dada ... 38
8 Struktur populasi M. teijsmannii di setiap lokasi penelitian ... 39
9 Peta distribusi populasi M. teijsmannii di CA Pulau Sempu, Jawa Timur .. 42
10 Habitus M. teijsmannii ... 46
11 Sisa buah M. teijsmannii yang dimakan primata ... 51
12 Beberapa variabel tanah yang berkorelasi nyata dengan kemelimpahan M. teijsmannii ... 58
DAFTAR LAMPIRAN
Halaman 1 Daftar spesies yang ditemukan dalam plot pengamatan di Cagar
Alam Pulau Sempu ... 76 2 Morfologi Myristica teijsmannii ... 79 3 Gambaran kondisi umum CA Pulau Sempu ... 80 4 Hasil analisis vegetasi di lokasi penelitian dalam kawasan CA Pulau
Sempu ... 83 5 Hasil analisis vegetasi pada enam lokasi penelitian di CA Pulau
Sempu ... 84 6 Hasil uji chi-square untuk pengujian asosiasi interspesifik spesies
berpasangan ... 90
1
I.
PENDAHULUAN
1.1 Latar Belakang
Myristicaceae atau famili pala-palaan adalah kelompok tumbuhan yang
populer dan penting di Indonesia, baik bagi ekonomi maupun ilmu pengetahuan. Famili ini menjadi kelompok tumbuhan yang penting secara ekonomi setelah diperkenalkannya Myristica fragrans atau Pala dari Indonesia ke dunia perdagangan internasional oleh Belanda pada abad ke-16. Selain itu, Myristicaceae memiliki banyak kegunaan yang telah dimanfaatkan secara luas oleh masyarakat lokal maupun untuk diperjualbelikan ke luar negeri, yaitu sebagai bahan makanan, minyak sayur dan lemak, rempah-rempah dan bumbu masak, obat, pewarna dan sumber kayu untuk furnitur dan bahan bangunan (Heyne 1987; Jansen et al. 1993).
Myristicaceae adalah famili khas kawasan tropis beranggotakan 16 genera
dengan genus Myristica sebagai genus terbesar (Heywood 1993). Dari 175 spesies
Myristica di dunia yang tersebar dari India bagian selatan dan Indochina sampai
kawasan Malesia hingga Australia bagian utara dan di Pasifik bagian timur sampai Fiji, 9 spesies diantaranya merupakan spesies asli Indonesia bahkan beberapa termasuk endemik (Arrijani 2005) serta langka dan dilindungi undang-undang (Mogea et al. 2001; Nurdjito dan Maryanto 2001).
Salah satu spesies anggota Myristicaceae yang termasuk ke dalam kategori
endangered atau genting menurut the International Union for the Conservation of
Nature and Natural Resources atau IUCN (IUCN 2006) adalah Myristica
teijsmannii Miq. atau dikenal dengan nama Durenan, Palan, Kosar atau Kayu
Resep di daerah Jawa Timur. Spesies ini penyebarannya jarang dan baru dilaporkan ditemukan di Jawa Timur, yaitu di hutan tropis campuran dataran rendah dan submontana pada ketinggian 50-1000 m dpl (Heyne 1987; de Wilde 2000). Berdasarkan studi herbarium di Herbarium Bogoriense pada tahun 2006, didukung informasi dari ahli Myristicaceae, W.J.J.O de Wilde, M. teijsmannii Miq. baru ditemukan di kawasan Gunung Wilis, Gunung Anjasmoro dan Pulau Sempu, seluruhnya di Jawa Timur. Namun informasi mengenai aspek ekologi, kebutuhan dan interaksi ekologis dari spesies ini dengan habitat atau lingkungannya
2 (autekologi) masih sangat terbatas, termasuk status populasi terkini, preferensi habitat, interaksi dengan komponen abiotik, asosiasi, serta penyebaran maupun program-program konservasinya.
Konservasi spesies penting dilakukan karena pada dasarnya kita sebagai umat manusia memiliki tanggung jawab moral dan etika untuk peduli dan memelihara kehidupan di bumi. Beberapa alasan lain pentingnya mengkonservasi spesies adalah karena spesies dapat memberikan manfaat bagi manusia (antara lain bahan obat, sumber makanan, bahan bakar, bahan bangunan), organismenya berperan untuk ekosistem sebagai penyangga sistem kehidupan di bumi, serta spesies merupakan salah satu tolok ukur atau standar tentang pemanfaatan kekayaan bumi secara berkelanjutan (Kunin & Lawton 1996). Pemanfaatan sumberdaya di bumi yang tidak berkelanjutan karena eksploitasi berlebihan, konversi habitat untuk pemukiman, industri dan pertanian, antara lain akan bermuara pada kelangkaan spesies, yang dapat berdampak buruk tidak saja pada lingkungannya tapi juga terhadap spesies lain di mana mereka berinteraksi. IUCN (2000) mempublikasikan 590 spesies tumbuhan Indonesia terancam kepunahan dan tidak ada satu spesies pun mendapat perencanaan program recovery atau strategi konservasi spesies sehingga perhatian terhadap spesies terancam kepunahan tersebut begitu penting untuk menghasilkan pendekatan konservasi spesies yang efektif (Widyatmoko 2001). Menurut Widyatmoko (2001), konservasi berbasis spesies akan efektif dengan melakukan identifikasi kebutuhan-kebutuhan habitat dan ekologi, status populasi, viabilitas populasi, asosiasi, distribusi, jumlah lokasi dan area minimum di mana spesies dapat dikonservasi, serta aspek-aspek biologis penting lainnya yang dapat menjadi penyebab kelangkaan spesies. Oleh karena itu konservasi spesies harus lebih diperhatikan serta perlu dijadikan suatu aksi global, dan sebagai langkah awalnya adalah menginventarisasi dan meneliti status populasi ataupun status kelangkaan spesies pada skala lokal dan nasional.
Di Indonesia sendiri M. teijsmannii tidak digolongkan sebagai spesies langka Indonesia (Mogea et al. 2001) maupun jenis hayati yang dilindungi perundang-undangan Indonesia dalam Peraturan Pemerintah No. 8 Tahun 1999 (Nurdjito dan Maryanto 2001). Perbedaan status kelangkaan M. teijsmannii antara yang ditetapkan IUCN dengan PP No. 8 Tahun 1999 tersebut perlu dikaji kembali
3 dengan melakukan studi populasi. Studi seperti ini penting untuk mengetahui status populasi terkini dari spesies terancam kepunahan sehingga hasilnya akan memberikan informasi dasar bagi upaya-upaya konservasi spesies maupun konservasi kawasan di mana spesies tersebut hidup, termasuk di dalamnya kebijakan-kebijakan untuk perlindungan, pelestarian dan pemanfaatannya.
Penelitian mengenai aspek ekologi, biologi konservasi dan penyebaran populasi M. teijsmannii juga perlu dilakukan karena spesies ini masih tergolong liar, memiliki nilai manfaat namun belum dibudidayakan dan ditemukan di habitat hutan tropis dataran rendah yang memiliki gangguan serta ancaman yang tinggi dari perusakan hutan akibat aktivitas manusia. Berdasarkan data spesimen herbarium, jumlah populasi spesies ini terbatas pada 5 lokasi di Jawa Timur dengan distribusi jarang dan kemungkinan termasuk spesies endemik kawasan ini. Oleh karena alasan-alasan tersebut, penelitian autekologi M. teijsmannii ini perlu menjadi salah satu prioritas dalam konservasi spesies terancam kepunahan. Menurut Given (1994), penelitian dengan fokus berbasis pada satu spesies perlu menjadi prioritas riset dalam konservasi karena dapat menyediakan dasar-dasar ilmiah untuk perlindungan, pengelolaan, propagasi dan reintroduksi spesies terancam kepunahan. Hal ini antara lain mengarah pada tujuan utama untuk mendapatkan pemahaman dari proses kepunahan spesies maupun persistensi serta viabilitas populasinya.
Studi mengenai status populasi spesies terancam kepunahan juga merupakan langkah penting yang mendukung implementasi Target 2 Strategi Global Konservasi Tumbuhan atau Global Strategy for Plant Conservation (GSPC). Target kedua tersebut adalah penilaian awal mengenai status konservasi dari seluruh spesies tumbuhan yang telah diketahui, baik pada level nasional, regional maupun internasional yang ditargetkan dapat tercapai pada tahun 2010 (CBD 2002).
Cagar Alam Pulau Sempu (CAPS) merupakan kawasan yang tepat untuk dijadikan lokasi penelitian autekologi dan pola penyebaran M. teijsmannii dalam komunitas hutan tropis dataran rendah. Kawasan konservasi tersebut memiliki batas ekologi yang jelas karena berupa pulau kecil yang dikelilingi selat dan samudera serta tidak berpenduduk. Vegetasi, kekayaan hayati lain dan kondisi
4 lingkungannya relatif masih terjaga (Goni et al. 1997). Selain itu, penelitian autekologi spesies tumbuhan belum pernah dilaporkan dari kawasan ini. Pentingnya penelitian-penelitian flora dan fauna di kawasan konservasi dapat menentukan jumlah spesies atau keragaman spesies di dalamnya yang dapat dilindungi dari kepunahan dalam integritas dan stabilitas kawasan yang terjamin.
1.2 Tujuan Penelitian
Tujuan dari penelitian ini adalah untuk mengetahui struktur dan status populasi M. teijsmannii Miq. di Cagar Alam Pulau Sempu berdasarkan data kuantitatif, serta menentukan karakteristik habitat dan interaksinya dengan kondisi lingkungan untuk mendapatkan peta distribusi geografis dari populasinya di Pulau Sempu. Selain itu penelitian ini bertujuan mengidentifikasi ancaman bagi penurunan populasinya di lokasi penelitian untuk mendapatkan informasi bagi strategi konservasi yang tepat bagi spesies langka ini.
1.3 Manfaat Penelitian
Hasil penelitian diharapkan dapat memberikan informasi dasar bagi kebijakan dalam pengelolaan kawasan konservasi dan strategi konservasi spesies
M. teijsmannii Miq. dalam pembangunan berkelanjutan sehingga dapat
menyelamatkan spesies langka tersebut dari kepunahan sekaligus bermanfaat bagi masyarakat dan pemerintah daerah setempat. Penelitian ini juga dapat memberikan data kuantitatif untuk memastikan kategori kelangkaan sehingga memberikan nilai konservasi bagi P. Sempu serta mendukung implementasi dari Target 2 GSPC pada level nasional.
5
II. TINJAUAN
PUSTAKA
2.1 Myristica teijsmannii Miq.
2.1.1 Nomenklatur
Myristica teijsmannii Miq. pertama kali dideskripsikan pada publikasi
tahun 1858 berdasarkan type Teijsmann s.n. dari Jawa. M. teijsmannii Miq. memiliki beberapa sinonim yaitu M. teysmanni dari type yang sama, dan M.
hyposticta Miq. dari type Horsfield s.n. dari Jawa (de Wilde 2000).
2.1.2 Nama-nama Lokal
Di Jawa Timur, yaitu di kawasan-kawasan ditemukannya spesies ini di alam, Myristica teijsmannii dikenal dengan berbagai nama. Masyarakat kawasan Gunung Wilis menyebutnya dengan Kayu Resep yang berarti kayu haid karena kayunya yang mengeluarkan getah berwarna merah seperti darah. Di kawasan lain di Jawa Timur, spesies ini juga dikenal dengan nama Durenan, Kosar, Palan atau Kayu Palan.
2.1.3 Morfologi
Myristica teijsmannii merupakan pohon dengan ketinggian 10 – 20 m.
Batangnya seringkali memiliki akar udara atau akar lutut, kulit batangnya coklat kemerahan dengan getah berwarna merah coklat. Ranting berbentuk sedikit bersegi, berwarna kekuningan atau coklat oranye yang ditutupi rambut pendek 0.1 mm. Susunan daunnya tunggal, berbentuk lonjong-bulat atau bulat memanjang berukuran 10-30(-34) x 2.5–9 cm, dengan pangkal membulat atau agak meruncing dan ujung yang meruncing. Permukaan bagian atas berwarna coklat olive sedangkan permukaan bagian bawah berwarna coklat keabu-abuan dengan rambut yang menyebar menyerupai sisik. Pada daun tidak terdapat kelenjar, tulang daun utamanya rata di bagian atas, dengan pertulangan lateral berjumlah 14 – 19 buah di setiap sisinya, membentuk sudut 45(-60)0 terhadap ibu tulang daun. Tangkai daun berukuran 10-20(-30) x 1.5-3mm sedangkan tunas daunnya 8-12 x 2.5 mm dengan rambut yang rapat setinggi 0.2 mm.
6 Perbungaan M. teijsmannii terletak di antara daun, kadang-kadang dekat pucuk lateral, berwarna coklat kekuningan, bertangkai pendek, tidak bercabang dan berambut sangat pendek (0.1-0.2 mm) serta memiliki braktea. Bunga jantan berukuran lebih besar daripada bunga betina. Pada bunga jantan panjang pedicel 4-6 mm dengan tunas yang bulat lonjong dan brakteola berbentuk ginjal. Jumlah bunga jantan pada setiap tangkai perbungaan berjumlah (5-)10-20. Pada bunga betina, panjang pedicel hanya 2 mm, tunasnya bulat dengan ovarium bulat berukuran 2.5-3.5 x 2-2.5mm. Tidak ada informasi mengenai tipe perbungaannya, namun de Wilde (2000) melaporkan bahwa salah satu spesimen M. teijsmannii di British Museum memperlihatkan bunga jantan dan betina pada perbungaan yang sama (dioecious).
Buahnya berwarna coklat, berjumlah 1-2 pada setiap perbuahannya, sedikit duduk dengan tangkai sepanjang 5 mm, berbentuk membulat dengan diameter 3-4.5 cm. Buahnya berambut rapat sangat pendek (0.1 mm) dengan warna karat. Daging buah memiliki ketebalan 4-8 mm, berair dan mengeluarkan aroma khas buah pala. Aril berwarna oranye pada buah masak, dan tidak saling menutupi. Bijinya lonjong pendek berukuran 2.5-3 cm.
2.1.4 Klasifikasi
Klasifikasi M. teijsmannii Miq. menurut Takhtajan (1997) adalah sebagai berikut: Divisi : Magnoliophyta Kelas : Magnoliopsida Ordo : Myristicales Famili : Myristicaceae Genus : Myristica
Spesies : Myristica teijsmannii
2.1.5 Distribusi dan Ekologi
M. teijsmannii Miq. merupakan spesies asli Indonesia (Arrijani 2005).
Berdasarkan studi herbarium yang dilakukan di Herbarium Bogoriense, Pusat Penelitian Biologi – LIPI, M. teijsmannii Miq. baru ditemukan di Jawa Timur, seperti yang dilaporkan de Wilde (2000) yaitu di Desa Parang, Gunung Wilis di
7 daerah Kediri. Seluruh spesimen M. teijsmannii Miq. di National Herbarium the Netherland di Leiden dilaporkan berasal dari Jawa Timur dan di antaranya P. Sempu (de Wilde WJJO 5 Maret 2007, komunikasi pribadi). Backer dan van den Brink (1963) menyatakan M. teijsmannii ditemukan dari Pacitan hingga Gunung Kawi dan Gunung Anjasmoro di Jawa Timur. Berdasarkan spesimen herbarium, beberapa lokasi lain yang merupakan tempat ditemukannya spesies tersebut dari tahun 1898 hingga 1956 adalah Gunung Wilis dan Desa Parang di Kediri, Gunung Anjasmoro serta P. Sempu. Dalam dekade terakhir, belum pernah dilaporkan ditemukannya populasi M. teijsmannii di kawasan lain. Berdasarkan data herbarium dan informasi literatur tersebut, ada kemungkinan M. teijsmannii Miq. bukan hanya asal Indonesia tapi juga kemungkinan besar merupakan spesies endemik Jawa Timur. Sampai saat ini M. teijsmannii dilaporkan jarang ditemukan, hidup di hutan tropis campuran dataran rendah dan submontana Jawa pada ketinggian 50-1000 m di atas permukaan laut (de Wilde 2000). Informasi ekologis lainnya pun sangat terbatas dan belum pernah dilaporkan sampai saat ini.
2.1.6 Pertumbuhan dan Perkembangan
Tidak banyak informasi mengenai pertumbuhan dan perkembangan
M. teijsmannii, baik pada periode germinasi maupun fase dewasanya. Namun pada
umumnya karakteristik genus Myristica memperlihatkan pertumbuhan yang cenderung lambat pada fase germinasi. Sebagai contoh, M. crassa memiliki periode germinasi 46-104 hari dan M. malaccensis 47-125 hari (Ng 1992).
Pada umumnya spesies-spesies yang termasuk famili Myristicaceae menunjukkan pertumbuhan yang intermittent atau diskontinyu sehingga membentuk percabangan yang diskontinyu. Bagian apikal akan beristirahat setelah inisiasi cabang dan selama periode ini aktivitas pertumbuhan dialihkan ke percabangan (Ng 1992). Susunan percabangannya plagiotropik dan diidentifikasi memiliki model arsitektur Massart (Ng 1992; Keller 1996).
Masa berbunga M. teijsmannii diketahui pada bulan Oktober hingga Mei, sedangkan masa berbuahnya dimulai sekitar bulan Mei hingga Juni (de Wilde 2000). Usia reproduktifnya belum pernah diteliti, demikian juga kemampuan perekrutan semai (seedling recruitment) maupun cara dispersi bijinya.
8
2.1.7 Pola Penyebaran
Individu-individu dalam populasi dapat menyebar menurut tiga pola yaitu acak, seragam dan mengelompok (Krebs 2002; Odum 1994; Ludwig & Reynolds 1988). Banyak populasi tumbuhan (maupun hewan) penyebarannya bersifat mengelompok di alam, dan hanya sedikit yang populasinya menyebar dalam pola yang teratur (Krebs 2002). Pola penyebaran populasi ini perlu diketahui secara objektif melalui metode yang tepat agar dapat menjelaskan sifat populasi secara biologis. Metode yang sering digunakan untuk mengetahui pola penyebaran adalah indeks penyebaran atau dispersal index. Indeks kesamaan yang paling banyak diterapkan dalam bidang ekologi adalah indeks Morisita karena hampir tidak dipengaruhi oleh ukuran sampel (Krebs 2002). Indeks ini dinilai merupakan metode pengukuran terbaik untuk kesamaan dalam penerapan ekologi (Wolda 1982 dalam Krebs 2002).
Informasi mengenai pola penyebaran M. teijsmannii akan memiliki peranan penting dalam menentukan extent of occurrence atau daerah sebaran geografisnya serta area of occupancy atau luas daerah yang ditempatinya di kawasan Pulau Jawa, sehubungan dengan keberadaannya yang kemungkinan besar tergolong endemik. Kedua kriteria tersebut menjadi faktor penting dan direkomendasikan oleh IUCN dalam menentukan kategori kelangkaan spesies langka (IUCN 2008).
2.1.8 Asosiasi Interspesifik
Hampir semua spesies tumbuhan tropis berada dalam interaksi kompleks atau asosiasi untuk melengkapi fase-fase dalam siklus kehidupannya (Widyatomoko 2001). Interaksi-interaksi species seperti ini sangat penting dalam ekologi suatu spesies sehingga studi untuk mengetahui adanya asosiasi spesies dapat memiliki implikasi ekologis yang penting (Ludwig & Reynolds 1988). Namun demikian asosiasi antar spesies dalam komunitas pada akhir tahun 1980-an masih menjadi sentral perdebatan (Fritz et al. 1987). Ada kelompok ekologis yang berpendapat bahwa spesies hidup individualistis di lingkungannya dan cenderung untuk berasosiasi secara acak, kelompok lain berpendapat bahwa spesies yang berkompetisi untuk sumber makanan akan berasosiasi negatif, sedangkan pendapat kelompok lain mengarah pada kecenderungan untuk berasosiasi positif karena
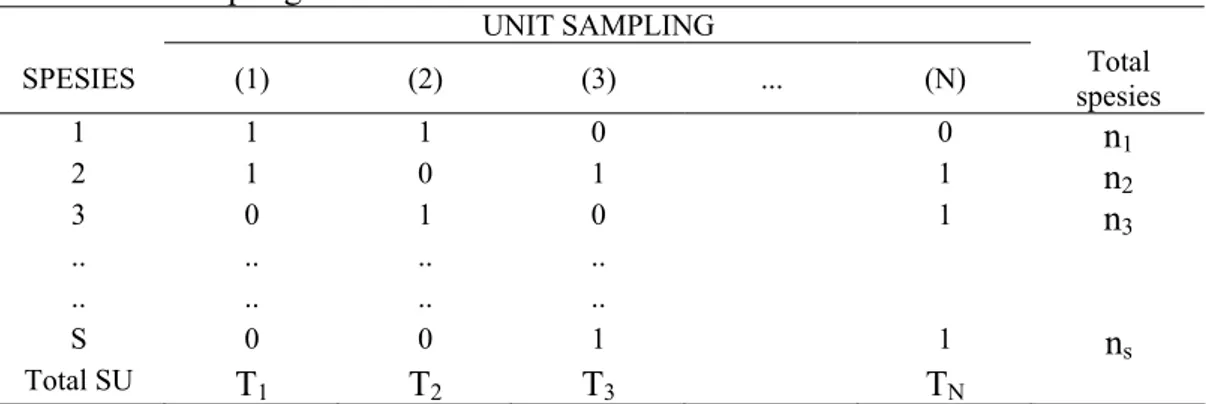
9 respons serupa terhadap variasi dalam kualitas ataupun kuantitas sumber makanannya. Beberapa proses ekologis dapat memperlihatkan asosiasi positif atau negatif antara dua spesies dalam suatu komunitas seperti diringkas dalam Tabel 1 (Schluter 1984, dalam Ludwig & Reynolds 1989).
Penelitian ekologi mengenai M. teijsmannii Miq. sejauh ini belum pernah dilaporkan, dan informasi mengenai spesies tumbuhan yang secara umum berasosiasi dengan spesies langka ini juga tidak didapatkan. Akan tetapi dengan melihat distribusi geografis spesies ini yang menempati hutan hujan dataran rendah pada ketinggian 0-1000 m dpl, maka ada kemungkinan spesies ini berasosiasi dengan anggota-anggota dari famili Dipterocarpaceae, Lauraceae, Myrtaceae, Ebenaceae, Moraceae atau Burseraceae, karena famili-famili tersebut banyak ditemukan dan menjadi karakteristik zona hutan hujan bawah (Indriyanto 2006). Prediksi ini perlu mendapatkan pembuktian melalui penelitian ekologi untuk mendapatkan data yang dapat dipertanggungjawabkan kebenarannya.
Tabel 1 Asosiasi di antara spesies akibat adanya interaksi antar spesies dan proses ekologis
Contoh proses ekologis Tipe interaksi
Positif Negatif Tidak ada Spesies memiliki respon yang sama
terhadap suplai sumberdaya yang terbatas
Spesies memiliki kebutuhan sumberdaya yang berbeda
Mutualisme Spesies satu dengan lainnya saling
mempertinggi daya survivalnya Sumberdaya diperebutkan dan dipergunakan secara eksklusif oleh spesies
Kompetisi Spesies berfluktuasi selaras dengan
terbatasnya sumberdaya Gangguan antar spesies menghasilkan keterasingan spesies lainnya Predasi Predator berfluktuasi selaras dengan
kepadatan mangsanya
Kepadatan predator yang tinggi menurunkan kepadatan mangsanya Sumber: Schluter (1984) dalam Ludwig & Reynolds (1988).
Data mengenai asosiasi interspesifik perlu dilakukan untuk M. teijsmannii untuk mendapatkan tambahan gambaran mengenai preferensi habitatnya, yang salah satunya dapat diimplikasikan pada pendugaan distribusi populasi di skala regional yang lebih luas. Dengan mengetahui pendugaan distribusi populasinya akan diperkirakan habitat-habitat yang sesuai bagi pertumbuhannya, yang dapat bermuara pada program konservasi spesies langka tersebut, antara lain program reintroduksi.
10 Ludwig dan Reynolds (1988) menyatakan bahwa penentuan asosiasi spesies melibatkan dua komponen yang berbeda: 1) uji statistik terhadap hipotesis bahwa dua spesies berasosiasi atau tidak, dan 2) pengukuran tingkat atau kekuatan asosiasi. Uji statistik untuk mendeteksi asosiasi spesies dilakukan dengan tes chi-square (Ludwig & Reynolds 1988) atau tes varians rasio (Schluter 1984, dalam Ludwig & Reynolds 1988). Tes varians rasio ini dapat diterapkan tidak hanya untuk asosiasi interspesifik namun juga untuk pola asosiasi intraspesifik (Fritz et
al. 1987). Pengukuran tingkat asosiasi ini dapat diketahui dengan berbagai metode
dalam bentuk indeks asosiasi, antara lain Ochiai Index, Dice Index, Jaccard Index (Ludwig & Reynolds 1988) dan Yule’s V Index (Slone & Croft 2001; Ludwig & Reynolds 1988).
2.1.9 Status Kelangkaan dan Konservasi
Pada daftar status kelangkaan tumbuhan versi IUCN (2006) terdapat 44 spesies Myristica yang tergolong rentan (vulnerable) hingga genting (endangered) dan 22 spesies di antaranya ditemukan di Indonesia, termasuk M. teijsmannii Miq.
M. teijsmannii Miq. oleh IUCN digolongkan ke dalam status endangered dengan
kriteria B1+2C berdasarkan penilaian kualitatif data herbarium oleh de Wilde pada tahun 1998. M. teijsmannii dinilai memiliki risiko kepunahan yang tinggi di alam karena hilangnya atau degradasi habitat sebagai penyebab utamanya. Degradasi habitat tersebut diakibatkan oleh praktek-praktek pertanian, deforestasi dan pembangunan infrastruktur termasuk pemukiman penduduk, yang ketiganya masih berlangsung hingga saat ini (IUCN 2006).
Ironisnya, dengan status genting seperti ini, M. teijsmannii tidak digolongkan ke dalam tumbuhan langka Indonesia (Mogea et al, 2001) ataupun jenis-jenis yang dilindungi undang-undang di Indonesia berdasarkan PP No. 7 dan PP No. 8 tahun 1999 (Nurdjito dan Maryanto 2001). Dalam kedua peraturan tersebut, hanya empat jenis Myristica yang digolongkan langka dan dilindungi undang-undang berdasarkan SK Menteri Kehutanan No. 54/Kpts/Um/2/1972, yaitu
M. papuana Scheff., M. paucifolia var. lancifolia, M. sphaerosperma S.C. Smith.
11
Hingga saat ini belum pernah dilaporkan upaya perlindungan bagi
M. teijsmannii, baik secara ex-situ maupun in-situ. Berdasarkan data dari
Herbarium Bogoriense dan NHN Leiden, salah satu spesimen M. teijsmannii dikoleksi dari Hortus Botanicus (Kebun Raya Bogor). Namun setelah melalui pengecekan berulang termasuk pada spesies sinonimnya baik di lapangan baik di database maupun dari data katalog kartu mati, aksesi dari spesies ini tidak pernah ada, yang berarti belum pernah dikoleksi di Kebun Raya Bogor. Dari seluruh lokasi populasi M. teijsmannii, hanya satu yang merupakan kawasan perlindungan yang berada di bawah Departemen Kehutanan, yaitu P. Sempu yang berstatus cagar alam, sehingga penentuan status populasi terkini di kawasan tersebut untuk menentukan langkah konservasinya merupakan langkah awal yang sangat diperlukan bagi pelestarian M. teijsmannii.
2.1.10 Aspek Pemanfaatan
M. teijsmannii dilaporkan dapat bermanfaat sebagai bahan pewarna merah
dari getah pada batang kayu yang dikeluarkannya (Heyne 1987). Hasil survey pendahuluan yang telah dilakukan di Desa Parang Kecamatan Banyakan di kawasan Gunung Wilis, Kediri, mendukung data tersebut. Pada awal abad ke-20, getah dari buah dan kayu M. teijsmannii digunakan untuk bahan pewarna kertas payung. Selain getahnya, kayunya juga cukup sering digunakan untuk bahan bangunan pada tiang dan penyangga rumah serta untuk gandar cangkul. Di samping itu, daun dan buahnya dipakai sebagai bahan campuran ramuan obat-obatan, namun khasiatnya tidak diketahui secara khusus.
Hegnauer (2001) melakukan uji biokimia pada biji M. teijsmannii dan menemukan kandungan minyak sebesar 3,5% vol. Akan tetapi jenis minyak yang terkandung dalam biji M. teijsmannii yang diuji tersebut tidak dijelaskan jenisnya.
2.2 Kategori Kelangkaan
Kelangkaan merupakan sebuah konsep yang kompleks karena memiliki sejumlah batasan yang berlainan dan suatu kombinasi antara faktor-faktor dinamis yang berkaitan dengan penyebaran spesies. Kelangkaan spesies ini bersifat kompleks secara fenomenal: temporal, spasial dan taksonomik (Widyatmoko
12 2001). Ditinjau dari fenomena spasialnya, kelangkaan suatu spesies sangat bergantung pada distribusi spesies dalam area tertentu (Cropper 1993).
Berdasarkan penyebabnya yang berupa proses alami dan non-alami, kelangkaan spesies tumbuhan dikategorikan ke dalam tiga kelompok (Cropper 1993). Kelompok pertama, spesies yang memiliki beberapa populasi besar dan tidak dianggap memiliki risiko kepunahan. Biasanya kelangkaan spesies ini diakibatkan oleh perusakan atau degradasi habitatnya. Kelompok kedua, spesies endemik, di mana jumlah populasi maupun jumlah individu dalam setiap populasinya sangat rendah. Spesies ini memiliki risiko kepunahan karena adanya proses alami seperti serangan hama, kekeringan atau bencana alam. Kelompok ketiga, tumbuhan langka yang terancam karena kegiatan-kegiatan yang baik secara langsung maupun tidak langsung mempengaruhi kesintasan spesies tersebut. Penyebab utamanya adalah konversi habitat seperti praktek-praktek agrikultur, pembangunan jalan, pertambangan.
Kelangkaan spesies dapat berujung pada kerentanan terhadap kepunahan. Spesies-spesies yang rentan terhadap kepunahan memiliki paling tidak satu dari 10 karakter tertentu yang dirangkum dalam Primack et al. (1998). Spesies-spesies tersebut perlu mendapat perhatian agar keberadaannya dapat terselamatkan dari kepunahan.
Untuk melindungi keberadaan suatu species, The IUCN Species Survival Commission telah menetapkan penggolongan spesies berdasarkan risiko kepunahannya dalam Red Data Book, yang direvisi secara kontinyu. Sejak pertama kali diperkenalkan, penggunaan kategori ini telah diakui dan dipakai secara internasional. Ada delapan kategori kelangkaan yang ditetapkan IUCN (Hilton-Taylor 2000), yaitu:
1. Extinct (EX) / punah
Suatu taksa dikategorikan Extinct jika individu terakhir telah mati. 2. Extinct in the wild (EW)/ punah di alam
Suatu jenis dikategorikan EW jika taksa tersebut diketahui hanya ada dalam budidaya, di karantina atau sebagai suatu populasi yang telah mengalami naturalisasi di luar daerahnya. Suatu taksa diduga punah di alam jika survey-survei sudah dilakukan pada daerah yang diduga /diketahui sebagai habitatnya,
13 pada waktu tertentu (diurnal, musiman, tahunan) dalam sepanjang daur hidupnya, tetapi taksa tersebut tidak ditemukan lagi.
3. Critically endangered (CR)
Suatu takson dikategorikan CR jika taksa tersebut menghadapi risiko kepunahan yang sangat ekstrim di alam dalam waktu yang sangat dekat, dengan kriteria-kriteria yang telah ditetapkan.
4. Endangered (EN) / genting
Suatu takson dikategorikan EN jika taksa tersebut tidak termasuk kategori CR tetapi mengalami risiko kepunahan yang sangat tinggi di alam dalam waktu dekat, yang didefinisikan dengan kriteria-kriteria tertentu.
5. Vulnerable (VU) / rentan
Suatu taksa dikategorikan VU jika taksa tersebut tidak termasuk kategori CR atau EN tetapi mengalami risiko kepunahan yang tinggi di alam dalam waktu dekat, seperti yang didefinisikan dengan kriteria-kriteria tertentu.
6. Lower Risk (LR) / risiko rendah
Suatu taksa dikategorikan LR jika taksa tersebut setelah dievaluasi, tidak memenuhi criteria-kriteria CR, EN atau VU. Taksa yang termasuk kategori ini dibagi menjadi tiga sub kategori:
1). Conservation dependent (cd) / tergantung upaya konservasi 2). Near threatened (nt) / Nyaris terancam
3). Least concern (lc) / Kekhawatiran minimal 7. Data deficient (DD)
Suatu taksa termasuk kategori ini jika informasi distribusi dan atau status populasinya tidak memadai untuk membuat dugaan risiko kepunahan suatu taksa baik secara langsung atau tidak langsung.
8. Not evaluated (NE)
Suatu taksa termasuk kategori NE jika taksa tersebut belum dapat dimasukkan ke dalam kategori-kategori No. 1-7 karena belum dievaluasi.
Berdasarkan IUCN (2008), ada beberapa kriteria khusus untuk kategori CR, EN dan VU (Tabel 2). M. teijsmannii dikategorikan ke dalam EN pada kriteria B1+2C berdasarkan data kualitatif spesimen herbarium (IUCN 2006), artinya spesies ini diperkirakan memiliki daerah sebaran yang sempit di mana luas daerah sebarannya (extent of occurrence) <5000 km2 dan luas daerah yang ditempatinya
14 (area of occupancy) <500 km2, dengan jumlah perkiraan < 2500 individu dewasa. Kemungkinan kepunahan dari spesies ini juga tergolong tinggi. Primack et al. (1998) menyatakan bahwa tingginya tingkat risiko kepunahan spesies disebabkan oleh kemerosotan populasi yang besar, area geografi dan luas area yang ditempatinya sempit serta ukuran populasi kecil.
Tabel 2 Kriteria status kelangkaan untuk kategori kritis, genting dan rentan Kriteria Kritis (CR) Genting (EN) Rentan (VU) A Penurunan populasi Pengukuran dilakukan 10 tahun atau 3 generasi
A1 >90% >70% >50%
A2, A3, A4 >80% >50% >30% B Kisaran sebaran geografis:
B1 : luas daerah sebaran <100 km 2 <5000 km2 <20000 km2 B2 : luas daerah ditempati, atau 2 dari 3 kriteria berikut <10 km2 <500 km2 <2000 km2
(a) Jumlah lokasi =1 ≤5 ≤10
(b) penurunan terus menerus (c) fluktuasi ekstrim
C Populasi kecil dan menurun
Jumlah individu
dewasa (C1 dan C2) < 250 < 2500 < 10000
C1: Penurunan populasi terus menerus s.d. 100 tahun 25% dalam 3 tahun atau 1 generasi 20% dalam 5 tahun atau 2 generasi 10% dalam 10 tahun atau 3 generasi C2:
(ai) jumlah individu dewasa pada sub populasi terbesar <50 <250 <1000 (aii) % individu dewasa dalam 1 subpopulasi 90-100% 95-100% 100%
(b) fluktuasi ekstrim dalam jumlah individu dewasa
D Populasi sangat kecil atau populasi terbatas (restricted)
D1 : jumlah individu dewasa < 50 < 250 <1000 D2 : daerah ditempati terbatas - - <20 km2 atau ≤5 lokasi E Analisis kuantitatif; Kemungkinan punah di alam ≥50% dalam 10
tahun atau 3 generasi (maks 100 tahun)
≥20% dalam 20 tahun atau 5 generasi
(maks 100 tahun)
≥10% dalam 100 tahun
15
2.3 Istilah-istilah yang digunakan
Autekologi adalah studi ekologi yang memfokuskan pada satu spesies tertentu
beserta interaksi dengan habitat atau lingkungannya, meliputi dinamika dan status populasi, demografi, preferensi habitat, interaksi dengan komponen abiotik, asosiasi serta ekofisiologi (Widyatmoko dan Irawati 2007).
Cagar Alam. Kawasan suaka alam yang karena keadaan alamnya mempunyai
kekhasan tumbuhan, satwa dan ekosistemnya atau ekosistem tertentu yang perlu dilindungi dan perkembangannya berlangsung secara alami. Di dalam cagar alam dapat dilakukan kegiatan untuk kepentingan penelitian dan pengembangan, ilmu pengetahuan, pendidikan dan kegiatan lainnya yang menunjang budidaya (UU No. 5 Tahun 1990).
Endangered atau genting. Suatu takson digolongkan genting bila takson tersebut
tidak tergolong kritis namun mengalami resiko kepunahan yang sangat tinggi di alam dalam waktu dekat. Tingkat risiko kepunahan yang tinggi disebabkan oleh kemerosotan populasi yang besar, area geografi dan area okupasi yang sempit serta ukuran populasi yang kecil (IUCN 2001).
Fase semai. Semai didefinisikan sebagai individu yang memiliki dbh (diameter
setinggi dada pada batang bebas cabang ± 1,3 m dari atas permukaan tanah) <3 cm dan tinggi ≤1,5 m (Indriyanto 2006).
Fase sapling. Sapling didefinisikan sebagai individu yang memiliki dbh <10 cm
dan tinggi >1,5 m (Indriyanto 2006).
Fase tiang. Tiang diartikan sebagai individu yang memiliki dbh 10 – 20 cm
(Indriyanto 2006).
Fase pohon. Pohon diartikan sebagai individu yang memiliki dbh >20 cm
(Indriyanto 2006).
Global Strategy for Plant Conservation (GSPC). Strategi dunia untuk konservasi
tumbuhan yang disusun oleh BGCI (Botanic Gardens Conservation International) beserta CBD (Convention on Biological Diversity) dan UNEP (United Nation for Environmental Program). Strategi dalam GSPC ini mencakup kerangka kerja
16 global untuk konservasi tumbuhan di seluruh dunia, yang terangkum dalam 16 target yang akan dicapai pada tahun 2010 (CBD 2002).
Reintroduksi adalah pelepasan dan pengelolaan suatu spesies ke suatu daerah di
mana spesies tersebut dulu pernah ada namun telah punah atau dipercaya telah punah dari area tersebut dengan tujuan memulihkan keberadaannya secara global atau lokal di habitat alaminya di dalam kisaran area geografiknya (Widyatmoko dan Irawati 2007).
Studi populasi adalah penelitian mengenai populasi spesies tertentu yang meliputi
kemelimpahan, pola penyebaran, struktur populasi setiap fase pertumbuhannya serta demografi yang mencakup regenerasi, kematian maupun perpindahan (Widyatmoko dan Irawati 2007).
17
III.
METODOLOGI PENELITIAN
3.1 Waktu dan Tempat Penelitian
Penelitian autekologi Myristica teijsmannii dilakukan di kawasan hutan campuran dataran rendah Cagar Alam Pulau Sempu (CAPS), Jawa Timur. Studi herbarium dilakukan di herbarium Kebun Raya Bogor dan Herbarium Bogoriense LIPI, analisis tanah dilakukan di Balai Penelitian Tanah Bogor. Kegiatan penelitian dilakukan mulai Maret 2007 sampai dengan Januari 2008, sejak survey pendahuluan hingga diselesaikannya tahap identifikasi spesimen herbarium.
Enam lokasi penelitian dipilih dalam kawasan cagar alam, yaitu Telaga Lele, Telaga Sat, Waru-waru, Air Tawar, Gua Macan dan Teluk Semut (Gambar 4). Seluruh lokasi penelitian merupakan hutan tropis dataran rendah dengan dominasi vegetasi bervariasi berdasarkan estimasi visual pada tahap observasi umum pada awal penelitian.
3.2 Bahan dan Alat
Objek utama dalam penelitian ini adalah Myristica teijsmannii. Peralatan yang digunakan adalah:
1). Perlengkapan sampling vegetasi: Global Positioning System (GPS) Garmin GPSMAP 60CSx, peta lokasi, peta kontur Pulau Sempu, meteran, kompas, tambang plastik;
2). Perlengkapan koleksi herbarium dan koleksi tumbuhan hidup termasuk buku identifikasi tumbuhan;
3). Peralatan pencatatan data lingkungan: altimeter, clinometer, termohigrograf, soil tester;
4). Peralatan sampling tanah: ring sample diameter 2 inci, Munsell’s color chart, pisau, sekop, label bersegel dan kertas label;
5). Perlengkapan entri data dan analisis data: software program Statistical Ecology (Ludwig & Reynolds 1988), Ecological Methodology (Krebs 2002), MapSource untuk upload data dari GPS ke komputer, STATISTICA 6 untuk analisis statistik, serta ArcView 3.2 (ESRI) untuk kepentingan pemetaan.
18
3.3 Metode Penelitian
3.3.1 Autekologi Myristica teijsmannii
Data yang dikumpulkan meliputi data biotik dan abiotik pada lokasi penelitian. Data faktor biotik meliputi kemelimpahan spesies, komposisi vegetasi, pencatatan individu M. teijsmannii reproduktif untuk data struktur populasi dan asosiasi serta spesies agen dispersalnya. Data abiotik yang dicatat meliputi karakteristik habitat berupa faktor edafik, topografi dan iklim mikro.
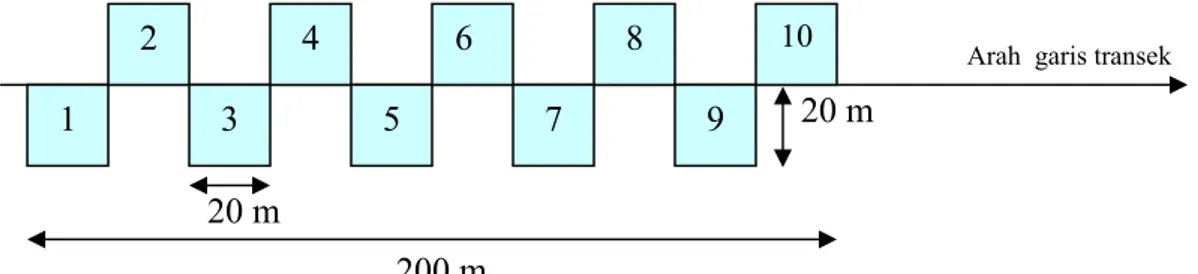
Untuk mengetahui struktur populasi M. teijsmannii dan struktur komunitas di lokasi penelitian, empat fase pertumbuhan (semai, sapling, tiang dan pohon) seluruh spesies yang ditemui dalam plot dihitung dan diukur diameternya pada setinggi dada (DBH / diameter at breast height). Pengambilan data populasi dilakukan dengan menggunakan metode sampling garis paralel sistematis (Cropper 1993; Krebs 1989) dalam sejumlah plot sepanjang garis transek (Gambar 1). Plot yang dibangun berbentuk bujursangkar yang berlainan ukurannya untuk setiap fase vegetasi dengan menggunakan metode plot bersarang (nested plot method). Untuk fase semai, plot yang dibangun berukuran 2 x 2 m, fase sapling 5 x 5 m, fase tiang 10 x 10 m, dan fase pohon 20 x 20 m (Gambar 2). Definisi untuk setiap fase telah dijelaskan pada bagian istilah-istilah bab II halaman 15. Transek yang dibangun berjumlah 15 buah dengan total luas plot penelitian 150 x 20 x 20 m yang meliputi kawasan seluas 0,68% dari total luas Cagar Alam Pulau Sempu (CAPS). Setiap transek dan plot pengamatan ditandai lokasinya (ditentukan koordinatnya) dengan menggunakan GPS.
3.3.2 Pengambilan Data Karakteristik Habitat 3.3.2.1 Penentuan Lokasi Penelitian
Langkah pertama penentuan lokasi penelitian adalah dengan melakukan penjelajahan di kawasan CAPS. Pada observasi umum ini dilakukan estimasi visual terhadap fisiognomi dan asosiasi floristik yang dominan. Penempatan transek-transek ditentukan berdasarkan asosiasi tegakan dan kondisi hutan yang cukup berbeda dengan sedapat mungkin mewakili tipe-tipe asosiasi dan kondisi topografi yang ada di kawasan CAPS.
19
3.3.2.2 Pengukuran di Lapangan
Karakteristik habitat yang diukur di lapangan meliputi faktor topografis, klimatik dan faktor edafik. Faktor topografis yang diukur adalah ketinggian menggunakan GPS dan altimeter, kemiringan tempat diukur dengan clinometer Suunto, serta arah lereng dengan kompas. Arah lereng diukur dari garis transek ke arah tepi terluar plot 20 x 20 m yang sejajar dengan garis transek. Arah lereng juga diperoleh dari titik koordinat transek dalam GPS yang ditumpangsusunkan ke dalam peta kontur yang telah dianalisis lereng dan arah lerengnya dengan program ArcView.
Data klimatik (iklim mikro) yang dicatat meliputi kelembaban dan suhu udara menggunakan termohigrograf serta curah hujan. Data curah hujan diperoleh dari data sekunder di lokasi yang bersumber dari instansi terkait. Data tanah yang diukur di lokasi penelitian adalah pH, kelembaban dan suhu tanah yang diukur pada permukaan tanah menggunakan Soil Tester. Warna tanah dicatat dan dibandingkan dengan Munsell’s Soil Color Chart (Anonim 2005).
Arah garis transek 20 m
20 m
200 m
Gambar 1 Skema pembuatan plot pada sampling.
Gambar 2 Skema plot bersarang pada plot 20 x 20 m.
Keterangan: Plot 1 = 2 x 2 m untuk semai; Plot 2 = 5 x 5 m untuk sapihan; Plot 3 = 10 x 10 m untuk tiang; Plot 4 = 20 x 20 m untuk pohon.
4 3 2 1 1 2 3 4 5 6 7 8 9 10
20
3.3.2.3 Pengambilan Contoh Tanah
Contoh tanah untuk sifat fisika diambil dengan menggunakan ring sample berdiameter 2 inci sesuai dengan prosedur standar (Anonim 2005). Untuk kepentingan pengujian sifat kimia, contoh tanah diambil dan disimpan dalam plastik bersegel. Pengambilan contoh dilakukan di setiap lokasi penelitian dari lapisan top soil (0-20 cm) dan lapisan sub soil (>20 cm) masing-masing sebanyak 2 ulangan.
3.3.2.4 Analisis Contoh Tanah
Contoh tanah yang diambil dari lokasi penelitian dianalisis sifat fisika dan kimianya di Balai Penelitian Tanah, Balai Besar Penelitian Sumberdaya Lahan Pertanian di Bogor, melalui tahap pengeringan suhu 1050C. Faktor fisika tanah yang dianalisis meliputi tekstur tanah (pasir, debu dan liat), kadar air, kerapatan lindak (bulk density), dan ruang pori total, sedangkan faktor kimiawi meliputi pH, kandungan bahan organik yang dinyatakan dalam rasio C/N, serta kandungan bahan organik meliputi Ca, Mg, K dan Na, serta nilai kapasitas tukar kation (KTK).
Analisis tekstur tanah dilakukan dengan pemisahan partikel liat menggunakan ultrasonik dan sedimentasi, sedangkan partikel pasir dipisahkan dan diukur dengan metode penyaringan dan pencucian. pH tanah diukur pada ekstraksi campuran tanah dan air dengan perbandingan 1:5, KTK diekstraksi dengan NH4 asetat 1 N dan pH 7, kandungan C dianalisis dengan metode Walkley & Black, sedangkan N total dideterminasi dengan metode Kjeldahl.
Untuk mendapatkan nilai total setiap variabel tanah dari dua lapisan topsoil dan subsoil, setiap parameter hasil analisis dihitung dengan formula:
(
)
(
)
S T S X T X X topsoil subsoil + × + × =di mana : X = nilai pada parameter i
T = kedalaman topsoil
21 Apabila kadar unsur hara dalam tanah dibandingkan dengan kebutuhan unsur hara bagi tanaman, maka akan diketahui apakah kadar unsur-unsur hara dalam tanah tersebut kurang, rendah, sedang atau tinggi. Kriteria penilaian hasil analisis tanah disajikan pada Tabel 3.
Tabel 3 Kriteria penilaian sifat kimia tanah Sifat Tanah Sangat
rendah Rendah Sedang Tinggi Sangat tinggi
C (%) < 1,00 1,00 – 2,00 2,01 – 3,00 3,01 – 5,00 > 5,00 N (%) < 0,10 0,10 – 0,20 0,21 – 0,50 0,51 – 0,75 > 0,75 C/N < 5 5 –10 11 – 15 16 – 25 > 25 Sangat masam Masam Agak masam Netral Agak alkalis Alkalis pH H2O < 4,5 4,5 – 5,5 5,6 – 6,5 6,6 – 7,5 7,5 – 8,5 > 8,5 Sumber: Hardjowigeno (1992). 3.4 Analisis Data
3.4.1 Autekologi dan Populasi Myristica teijsmannii Serta Struktur Komunitas
Data populasi yang meliputi kemelimpahan spesies dalam komunitas dihitung mengikuti formulasi dari Cox (2002). Keragaman spesies, kesamaan (similarity) antar lokasi penelitian serta pola penyebaran M. teijsmannii dihitung berdasarkan Krebs (1989) dengan menggunakan program ECOLOGICAL METHODOLOGY (Krebs 2002). Khusus untuk uji asosiasi interspesifik, perhitungan dilakukan berdasarkan metode dan software STATISTICAL ECOLOGY (Ludwig & Reynolds 1988).
3.4.1.1 Kerapatan (K)
Kerapatan adalah jumlah individu suatu spesies dalam suatu luasan tertentu. Parameter ini dihitung dengan persamaan:
Ki = ni / A [3.1]
di mana Ki adalah kerapatan untuk spesies i, ni adalah jumlah total individu spesies
22 Kerapatan relatif (KR) adalah jumlah individu spesies i (ni) proporsional
terhadap jumlah total individu seluruh spesies yang dijumpai dalam plot (Σn). KR dihitung dengan persamaan:
KR = (ni / Σn) x 100% [3.2]
3.4.1.2 Frekuensi (F)
Frekuensi adalah kesempatan menjumpai spesies dalam suatu sampel, yang diperoleh dari persamaan [3.3].
Fi = jumlah plot yang diduduki spesies [3.3] jumlah total plot yang dibangun
Frekuensi relatif (FR) adalah frekuensi dari individu spesies i (Fi) sebagai
proporsi dari jumlah frekuensi seluruh spesies yang dijumpai dalam plot. FR dihitung dengan persamaan [3.4].
FRi = (Fi / ΣFi) x 100% [3.4]
3.4.1.3 Dominasi (D)
Dominasi adalah proporsi dari permukaan tanah yang diduduki oleh proyeksi vertikal bagian tajuk tumbuhan ke permukaan tanah. Dominasi dihitung dengan persamaan:
Di = ai / A [3.5]
di mana ai merupkan total area yang ditutupi oleh spesies i (perkiraan melalui basal
area, penutupan tajuk) dan A merupakan total area habitat yang disampling.
Dominansi Relatif untuk spesies i (DRi) adalah adalah penutupan atau
kelindungan untuk spesies tersebut (Di) yang diekspresikan sebagai proporsi dari
total dominansi seluruh spesies. DRi dihitung dengan persamaan:
DRi = (Di / ΣD) x 100% [3.6]
3.4.1.4 Indeks Nilai Penting
Jumlah dari ketiga pengukuran relatif untuk spesies i merupakan sebuah indeks yang disebut indeks nilai penting (INP).
23
INP = KR + FR + DR [3.7]
3.4.1.5 Indeks Keragaman
Nilai keragaman spesies dalam habitat dinyatakan dengan indeks keragaman atau diversity index. Indeks keragaman yang digunakan dalam penelitian ini dan juga paling umum digunakan dalam metode ekologi adalah indeks keragaman Shannon-Wiener. Persamaannya adalah sebagai berikut.
H’ = – Σ pi log2 pi [3.8]
di mana H’ menyatakan indeks keragaman menurut Shannon-Wiener (Krebs 1989), sedangkan pi menyatakan proporsi spesies i terhadap keseluruhan jumlah
spesies yang dijumpai pada plot di dalam komunitas yang diteliti. Proporsi ini diperoleh dari perbandingan jumlah spesiesnya di mana ni merupakan jumlah
individu spesies i dan N adalah jumlah seluruh individu spesies yang dijumpai dalam plot.
pi = ni / N [3.9]
3.4.1.6 Indeks Kesamaan Komunitas
Tingkat kesamaan atau kemiripan komunitas ditentukan berdasarkan indeks kesamaan Morisita (Morisita 1951 dalam Krebs 1989). Indeks ini memiliki kisaran 0 (tidak memiliki kesamaan) hingga mendekati 1 (kesamaan sempurna). Krebs (1989) menyatakan bahwa indeks kesamaan Morisita hampir tidak dipengaruhi oleh ukuran sampel, kecuali oleh ukuran sampel yang sangat kecil sehingga direkomendasikan sebagai metode pengukuran kemiripan komunitas yang terbaik untuk ekologi (Wolda 1981 dalam Krebs 1989).
(
)
j k ik ij n N N X X C 2 1 2λ
λ
λ = ∑+ [3.10]di mana Cλ = indeks kesamaan Morisita antara sampel j dan k
Xij,Xik = jumlah individu spesies i dalam sampel j dan sampel k
Nj = Σ Xij = total jumlah individu dalam sampel j
24
(
)
[
]
(
1)
1 1 − − Σ = j j ij ij n N N X X λ [3.11](
)
[
]
(
1)
1 2 − − Σ = k k ik ik n N N X X λ [3.12] 3.4.1.7 Pola PenyebaranUntuk mengetahui pola penyebaran populasi M. teijsmannii di lokasi penelitian digunakan indeks penyebaran Morisita (Morisita 1962 dalam Krebs 1989). Indeks tersebut dihitung dengan [3.13].
( )
⎥
⎥
⎦
⎤
⎢
⎢
⎣
⎡
∑
−
∑
∑
−
∑
=
x
x
x
x
n
d 2 2I
[3.13]di mana : Id = indeks dispersi Morisita
n = jumlah plot Σx = jumlah total plot
Σx2 = jumlah kuadrat untuk total plot
Jika : Id = 1, maka pola penyebaran spesies acak (random)
Id = 0, maka pola penyebaran spesies seragam (uniform)
Id = n, maka pola penyebaran spesies mengelompok (clumped)
Untuk mengetahui signifikansi dari nilai tersebut maka dilakukan uji hipotesis dengan tes distribusi χ2 melalui rumus sebagai berikut.
χ2 = (
∑
− 1) + −∑
(d.f.= −1) dI x n x n
[3.14] Hipotesis null (H0) yang diajukan adalah spesies menyebar secara acak. Apabila
χ2
hitung > χ2tabel maka H0 ditolak, berarti spesies menyebar tidak secara acak.
Meskipun banyak digunakan dalam penelitian ekologi, terdapat indeks dispersi yang lebih baik dengan melakukan standarisasi Indeks Morisita. Dalam sebuah simulasi studi, Indeks Morisita yang distandarisasi ini (standardized
Morisita’s index) dinilai sebagai pengukuran dispersi populasi yang terbaik karena
sifatnya independen terhadap kerapatan populasi dan tidak bergantung pada ukuran sample (Myers 1978 dalam Krebs 2002). Namun demikian direkomendasikan bahwa ukuran sampel minimum untuk pengujian pola dispersi populasi ini adalah 50 kuadrat (Green 1966 dalam Krebs 2002).
25
Indeks Morisita yang diperoleh selanjutnya dicari dua titik kritisnya
melalui uji χ2 untuk mencari derajat pengelompokannya.
χ − + = −
∑
∑
2 .975 Uniform index = ( ) 1 i u i n x M x [3.15] di mana χ2.975= nilai dari tabel dengan df (n-1) yang memiliki 97,5% area ke sebelah kanan kurva
ΣXi = jumlah organisme dalam kuadrat i (i = 1,...n) n = jumlah kuadrat χ − + = −
∑
∑
2 .025 Clumped index = ( ) 1 i c i n x M x [3.16] di mana χ2.025 = nilai dari tabel dengan df (n-1) yang memiliki 2,5% area ke sebelah kanan kurva
Berdasarkan hasil indeks Mc atau Mu di atas maka Indeks Morisita standar
(IP) dihitung berdasarkan salah satu dari empat persamaan berikut ini.
1. Jika Id ≥ Mc > 1,0 : ⎟⎟ ⎠ ⎞ ⎜⎜ ⎝ ⎛ − − + = c c d P M n M I I 0.5 0.5 [3.17] 2. Jika Mc > Id ≥ 0 : ⎟⎟ ⎠ ⎞ ⎜⎜ ⎝ ⎛ − − = 1 1 5 . 0 u d P M I I [3.18] 3. Jika 1,0 > Id > Mu : ⎟⎟ ⎠ ⎞ ⎜⎜ ⎝ ⎛ − − − = 1 1 5 . 0 u d P M I I [3.19] 4. Jika 1,0 > Mu > Id : ⎟⎟ ⎠ ⎞ ⎜⎜ ⎝ ⎛ − + − = u u d P M M I I 0.5 0.5 [3.20]
Indeks Morisita yang distandarkan (IP) ini berkisar antara -1,0 hingga +1,0.
Jika IP = 0 pola penyebaran acak, IP < 0 penyebaran seragam dan IP > 0
penyebaran mengelompok.
3.4.1.8 Peta Penyebaran M. teijsmannii di Cagar Alam Pulau Sempu
Penyebaran populasi M. teijsmannii digambarkan pada peta digital Pulau Sempu dengan mentransfer data koordinat hasil penandaan GPS di setiap plot