63
V. HASIL DAN PEMBAHASAN
Harimau yang ditranslokasikan dan dipasangi kalung GPS selama penelitian adalah sebanyak enam ekor. Namun demikian, harimau yang ditranslokasikan ke TNKS (JD-4) ditemukan mati hanya tujuh hari setelah dilepas-liarkan akibat terperangkap jerat yang dipasang pemburu kambing hutan di dalam kawasan taman nasional tersebut. Harimau JD-1 dan JD-2 yang dilepas-liarkan di TNBBS, diamati pergerakannya selama masing-masing 224 hari (menghasilkan 3.469 data posisi) dan 253 hari (1.288 data posisi). Harimau BD-1 yang ditranslokasikan ke KHUM diobservasi selama 213 hari (6.680 data posisi), namun kemudian diketahui juga mati akibat terjerat di sebuah ladang di pinggir hutan setelah kalung GPS-nya bekerja selama 7 bulan. Harimau JD-3 di TNGL diamati pergerakannya selama 79 hari (1.486 data posisi), dan harimau JD-5 di TNKS dipantau selama 238 hari pengamatan (7.007 data posisi). Kalung GPS yang pasang pada harimau JD-1, JD-3 dan JD-5 rusak setelah masing-masing beroperasi selama 7,5 bulan, 2,5 bulan dan 8 bulan. Kalung GPS pada JD-2 terlepas secara otomatis sesuai rencana setelah 8,5 bulan beroperasi (Tabel 6).
Tabel 6. Jadwal operasi dan jumlah data posisi yang berhasil dikumpulkan melalui kerah GPS pada masing-masing harimau sumatera translokasi selama penelitian antara 2008-2011.
Harimau Lokasi Tanggal operasi
kerah GPS N hari operasi N data posisi Keterangan Nama Kls umur JD-1 Jantan dewasa TNBBS 22/072008-03/03/2009 224 3.469 GPS rusak JD-2 Jantan dewasa TNBBS 22/07/2008-31/03/2009 253 1.288 GPS lepas otomatis BD-1 Betina dewasa KHUM 21/12/2008-21/07/2009 213 6.680 Harimau mati JD-3 Jantan dewasa TNGL 27/12/2008-15/03/2009 79 1.486 GPS rusak JD-4 Jantan dewasa TNKS 05/06/2009-11/06/2009 7 71 Harimau mati JD-5 Jantan dewasa TNKS 20/12/2010-14/08/2011 238 7.007 GPS rusak
64
Sebelum ditranslokasikan, harimau jantan dewasa JD-1 dan JD-2 ditangkap oleh petugas Balai KSDA Aceh di lokasi yang sama, yaitu di satu desa pesisir Kabupaten Aceh Selatan. Harimau JD-1 dan JD-2 masing-masing ditangkap pada 9 Nopember 2007 dan 25 Nopember 2007, dengan alasan bahwa kedua harimau tersebut sering memasuki perkampungan hingga meresahkan masyarakat. Habitat di tempat asal JD-1 dan JD-2 merupakan hutan dataran rendah kering, yang umumnya merupakan hutan bekas HPH yang bertopografi perbukitan. Kawasan tersebut terletak di kaki Pegunungan Bukit Barisan sebelah barat. Setelah tertangkap, baik JD-1 maupun JD-2 dirawat di dalam kandang berbentuk kotak berjeruji besi di halaman belakang kantor Balai KSDA Aceh selama sekitar 7 bulan. Kemudian, keduanya dipindahkan ke karantina pemulihan di dekat lokasi translokasi di dalam kawasan TNBBS di Lampung. JD-1 dan JD-2 ditranslokasikan dengan jarak sekitar 1.350 km dari tempat asalnya di Aceh Selatan, ke kawasan hutan TNBBS di Lampung.
Harimau jantan JD-3 ditangkap 15 Nopember 2008 di satu desa tepi pantai di Aceh Barat. JD-3 juga ditangkap akibat sering memasuki desa dan diduga telah sering memangsa hewan-hewan ternak milik masyarakat desa. Tempat hidup asal JD-3 di Aceh Barat merupakan hutan dataran rendah kering dan hutan-hutan bekas HPH yang topografinya berbukit-bukit di kaki Pegunungan Bukit Barisan sebelah barat. Setelah mendapat perawatan selama 42 hari, JD-3 ditranslokasikan dengan jarak sekitar 200 km dari tempat asalnya di kawasan pantai barat Aceh ke kawasan hutan di TNGL yang berbatasan dengan kawasan bekas HPH di kaki bagian timur Pegunungan Bukit Barisan.
Harimau betina dewasa BD-1 ditangkap pada 3 Desember 2008 di satu desa di Kabupaten Aceh Utara, di wilayah timur laut Provinsi Nanggroe Aceh Darussalam. BD-1 evakuasi karena sering memasuki pemukiman dan memangsa hewan ternak masyarakat. Habitat asal BD-1 di Aceh Utara merupakan hutan perbukitan dan pegunungan rendah. Kawasan tersebut merupakan sisi timur dari Pegunungan Bukit Barisan. Setelah 18 hari
65
dikarantina, BD-1 langsung ditranslokasikan ke wilayah terpencil di kawasan hutan Ulu Masen, yang berjarak sekitar 70 km dari tempat dimana BD-1 ditangkap.
Harimau jantan dewasa JD-5 berhasil diselamatkan petugas Balai KSDA Sumatera Barat pada 24 Nopember 2010. Harimau ini terjebak selama beberapa hari dalam lubang perangkap yang dibuat masyarakat untuk menangkap rusa, di hutan dekat sebuah desa di Kabupaten Pesisir Selatan, Sumatera Barat. Habitat JD-5 di tempat asalnya adalah hutan dataran rendah yang berbukit-bukit, di sisi barat Pegunungan Bukit Barisan. Sekitar tiga minggu setelah penangkapannya (20 Desember 2010), JD-5 di translokasikan ke satu areal hutan di TNKS yang berbatasan dengan perkebunan sawit, dengan jarak sekitar 74 km dari tempat asalnya ditangkap.
5.1 Pola Penggunaan Ruang 5.1.1 Pergerakan
5.1.1.1 Panjang dan Bentuk Lintasan Pergerakan
Menurut (Ahearn et al. 2001) pola lintasan pergerakan harimau dicirikan melalui jarak dan arah pergerakannya. Dengan menggunakan data dari hari-hari observasi lengkap, diketahui bahwa rata-rata jarak pergerakan harian yang ditempuh oleh harimau sumatera translokasi berkisar antara 2,80 hingga 4,00 km. Rata-rata jarak pergerakan dari seluruh harimau adalah 3,52 km (Tabel 7). Jarak pergerakan harian betina dan jantan secara signifikan berbeda (U = 44473; P= 0,000). Harimau betina menempuh rata-rata jarak harian lebih panjang dibandingkan dengan jantan.
Rata-rata jarak tempuh harimau jantan JD-1 adalah 3,51 km/hari (kisaran 0,06-13,92 km/hari), sedangkan harimau jantan JD-3 dan JD-5 memiliki rata-rata jarak pergerakan masing-masing 2,80 km (kisaran 0,05-8,00 km/hari untuk JD-3) dan 3,32 km/hari (kisaran 0,14-18,99 km/hari untuk JD-5). Satu-satunya harimau betina BD-1 yang juga menggunakan data satu lokasi setiap 0,5 jam memiliki rata-rata jarak tempuh 4,00 km/hari (kisaran 0,20-11,33 km/hari). Hasil penelitian Smith (1993) di TN Chitwan, Nepal,
66
menyatakan sebaliknya dimana harimau jantan mampu menjelajah tiga kali lebih jauh daripada harimau betina. Selain untuk pencarian hewan mangsa, panjangnya penjelajahan harimau jantan lebih dikarenakan untuk menjaga wilayah teritori serta pencarian betina pasangan kawin. Sunquist (2010) berpendapat bahwa luasnya daerah jelajah jantan lebih disebabkan untuk penguasaan betina daripada penguasaan sumber pakan. Menurut observasi Valen (2011) harimau jantan akan memberikan tanda dengan cara menyemprotkan urin serta sekresi dari kelenjar anal lebih sering pada wilayah jelajahnya ketika datang masa-masa estrus harimau betina. Harimau jantan mengunjungi betina tiga sampai lima kali per bulan di dalam daerah jelajahnya dan akan bergerak lebih lambat bila sedang bersama atau mencari pasangan betina untuk kawin (Ahearn et al. 2001).
Tabel 7. Rata-rata jarak pergerakan harian dan jarak tempuh maksimum hari-mau sumatera translokasi.
Harimau Lokasi N hari observasi
Rata-rata jarak tempuh (km) Jarak tempuh maks/hari (km) hari + SD siang + SD malam + SD
JD-1 TNBBS 223 3,51 + 3,01 1,74 + 1,96 1,77 + 2,06 13,92 JD-3 TNGL 68 2,80 + 2,19 1,43 + 1,22 1,37 + 1,22 8,00 JD-5 TNKS 236 3,32 + 2,25 1,54 + 1,56 1,78 + 1,45 18,99 BD-1 EUM 208 4,00 + 2,41 2,27 + 1,74 1,74 + 1,26 11,33 Jantan 527 3,33 + 2,60 1,61 + 1,70 1,72 + 1,72 18,99 Betina 208 4,00 + 2,41 2,27 + 1,74 1,74 + 1,26 11,33 Rata-rata 735 3,52 + 2,56 1,80 + 1,74 1,73 + 1,60 18,99
Adanya perbedaan jarak jelajah harian pada setiap individu harimau translokasi di Sumatera ini sangat dimungkinkan akibat perbedaan tipe habitat utama dan kondisi topografi di masing-masing areal pelepas-liarannya. Lokasi pelepas-liaran harimau jantan JD-1 dan JD-2 di TNBBS serta JD-5 di TNKS, didominasi oleh belukar/hutan sekunder muda dan hutan dataran rendah dengan topografi umumnya datar. Lokasi translokasi jantan JD-3 di TNGL didominasi oleh hutan pegunungan rendah dan dataran rendah dengan tingkat kelerengan umumnya curam hingga sangat curam. Lokasi Ulu Masen (tempat BD-1 dilepas-liarkan) didominasi oleh hutan pegunungan rendah dan
67
belukar/sekunder muda dengan kelerengan datar-landai hingga curam. Konsekuensi dari perbedaan ketinggian tempat, medan dan tipe habitat adalah terjadinya perbedaan dalam keragaman dan kelimpahan hewan mangsa. Menurut Griffiths (1994), keanekaragaman dan kelimpahan hewan mangsa harimau di hutan dataran rendah dengan ketinggian 100-600 meter dpl lebih tinggi dibandingkan dengan di hutan pegunungan rendah dengan ketinggian 900-1.700 meter dpl. Dengan demikian, harimau BD-1 di Ulu Masen membutuhkan usaha yang lebih besar dengan melakukan perjalanan yang lebih panjang untuk mendapatkan hewan mangsanya. Data hasil observasi juga menunjukkan bahwa KHUM (tempat BD-1 ditranslokasikan), memiliki kelimpahan relatif hewan mangsa utama (rusa, kijang dan babi hutan) yang paling rendah dibandingkan dengan lokasi pelepas-liaran harimau lainnya (Lampiran 1). Menurut Sunquist (2010) sebagian besar waktu harimau di alam dihabiskan untuk mencari pakan dan umumnya mereka menjelajah areal yang luas untuk memenuhi kebutuhan pakannya. Pendapat lain (Nowak 1991) menyatakan bahwa di Rusia timur jauh, dimana hewan mangsa tersebar secara luas, harimau siberia melakukan pergerakan hingga 60 km per hari.
Selain itu, mengingat BD-1 adalah harimau betina, panjangnya perjalanan harian yang ditempuhnya mungkin juga ada kaitannya dengan upaya menemukan jantan siap kawin karena masa estrus harimau betina terjadi pada setiap tiga sampai sembilan minggu sekali (Nowak 1991, Ahearn et al. 2001). Lamanya masa estrus adalah tiga sampai enam hari pada setiap periode (Nowak 1991). Pada masa estrusnya harimau betina sering mengeluarkan suara auman untuk menarik harimau jantan (Sunquist 1981). Jackson (1996) menemukan bukti bahwa pada musim kawin, macan salju betina melakukan pergerakan satu setengah kali lebih panjang daripada biasanya.
Barlow et al. (2011) melakukan studi menggunakan kalung GPS memperoleh rata-rata jarak tempuh harimau betina di hutan bakau Sundarbans, Bangladesh adalah sekitar 2,9 km/hari dengan jarak pergerakan
68
maksimum 10,8 km. Adanya perbedaan jarak tempuh antara harimau betina di dua lokasi yang sangat berjauhan ini kemungkinan disebabkan oleh perbedaan tipe habitat, dimana Sundarbans merupakan kawasan hutan bakau yang didominasi lahan basah, sementara KHUM merupakan kawasan yang didominasi oleh hutan pegunungan, yang secara alamiah memiliki kelimpahan hewan mangsa lebih rendah dibandingkan kawasan dataran rendah (Sunquist et al. 1999). Menurut Sunquist (2010) panjang atau pendeknya jarak tempuh harimau ada hubungannya dengan kelimpahan hewan mangsa, dimana di kawasan yang kelimpahan hewan mangsanya tinggi harimau tidak melakukan pencarian mangsa secara aktif. Namun, jarak pergerakan harimau sangat bervariasi tergantung lokasi dan habitat.
Hasil pengamatan menunjukkan bahwa secara umum bentuk lintasan pergerakan semua harimau translokasi dilakukan secara zig-zag ketika melakukan eksplorasi di dalam daerah jelajahnya. Hal ini kemungkinan ada kaitannya dengan pemilihan jalan lintasan dalam pergerakannya. Sunquist (2010) berpendapat bahwa harimau sering ditemukan menggunakan jalan-jalan bekas logging, jalan-jalan setapak dan alur-alur sungai di dalam hutan, dalam melakukan pergerakan di antara lokasi perburuan hewan mangsa. Hasil pengamatan pada saat observasi lapangan, juga menunjukkan bahwa jejak-jekak harimau sangat sering ditemukan di jalan setapak yang biasa digunakan manusia di dalam hutan.
Data pergerakan juga menunjukkan bahwa kadang-kadang sesekali harimau translokasi terlihat melakukan perjalanan panjang membentuk garis lurus, yang langsung menuju ke suatu tempat. Pergerakan seperti ini sepertinya berhubungan dengan perilaku kawin terutama pada harimau jantan, kemungkinan harimau akan langsung bergerak menuju daerah jelajah betina ketika mendapat tanda dari harimau betina yang siap kawin. Ahearn et al. (2001) berpendapat bahwa harimau jantan akan melakukan perjalanan langsung menuju sasaran ketika mencari hewan mangsa dan pasangan kawin, dan biasanya bergerak lebih lambat ketika berada pada daerah jelajah betina.
69
Harimau juga akan tinggal selama beberapa hari pada lokasi yang sama setelah mendapatkan hewan mangsa.
Selain itu, harimau-harimau translokasi juga melakukan pergerakan memutar kembali mengunjungi tempat-tempat yang sebelumnya didatangi, setelah menjelajah selama beberapa hari atau beberapa minggu. Menurut Sunquist (2010), dalam pencarian hewan mangsanya harimau jarang sekali melakukan pengembaraan, tetapi pergerakan mereka sangat terarah. Harimau juga dapat mengingat dengan baik areal-areal tempat berburu hewan mangsa di dalam daerah jelajahnya, dan mereka juga hafal akan jalur-jalur yang terbaik di antara dua lokasi berburu mangsanya tersebut.
Pergerakan harimau-harimau translokasi selalu diarahkan menuju batas-batas dua tipe habitat yang berbeda dan ke tepi-tepi hutan antara hutan dataran rendah dan vegetasi belukar/hutan sekunder muda. Pola pergerakan ini sepertinya erat kaitannya dengan kebiasaan hewan mangsa harimau yang selalu mencari makan di tepi-tepi atau batas hutan dengan vegetasi belukar. Karanth & Sunquist (1992) menyatakan bahwa mosaik lansekap yang terdiri atas hamparan hutan dan padang rumput, merupakan habitat yang sangat mendukung kehidupan hewan ungulata. Dalam penjelajahannya, terdapat kecenderungan bahwa harimau bergerak mengikuti kontur topografi dan punggungan bukit pada areal perbukitan dan pegunungan. Harimau umumnya menghindari daerah yang sangat terjal, dan akan memilih punggung bukit yang terendah apabila ingin melintasinya. Bentuk serta pola lintasan harimau translokasi setiap bulan disajikan pada Lampiran 2, 3, 4, 5 dan 6.
5.1.1.2 Panjang Pergerakan pada Siang dan Malam
Rata-rata jarak pergerakan harimau pada siang hari berkisar antara 1,43-2,27 km, sedangkan pada malam hari antara 1,37-1,78 km. Secara individu, hasil uji menunjukkan bahwa pada dua harimau jantan, yaitu JD-1 (Z= -0,184; P= 0,854) dan JD-3 (Z= -0,706; P= 0,480), tidak ada perbedaan yang signifikan antara rata-rata jarak pergerakan pada siang dengan malam
70
harinya. Pada jantan JD-5 (Z= -2,667; P= 0,008) terdapat perbedaan jarak pergerakan antara siang dan malam hari, dimana pada malam hari harimau ini menempuh jarak lebih panjang dibandingkan siang hari. Hal ini kemungkinan diakibatkan oleh kondisi lokasi pelepas-liaran JD-5 di TNKS, yang mana daerah jelajah JD-5 didominasi oleh habitat belukar/sekunder muda (47,0%) serta mayoritas elevasinya adalah 0-500 meter dpl (59,8%). Pada kondisi tersebut dapat diprediksi bahwa suhu udara lingkungan umumnya panas pada siang hari, sehingga pergerakan atau penjelajahan akan lebih nyaman dilakukan pada malam hari yang suhu udara lingkungannya lebih dingin. Namun, secara umum tidak ada perbedaan yang signifikan antara rata-rata jarak pergerakan siang dan malam hari pada harimau jantan (Z= -1,348; P= 0,178).
Rata-rata jarak pergerakan siang hari dan malam hari pada harimau betina secara signifikan berbeda (Z= -3,711; P= 0,000), dimana pada harimau betina rata-rata jarak jelajah pada siang hari (2,27 km) lebih panjang daripada malam hari (1,74 km). Lokasi pelepas-liaran harimau BD-1 di Ulu Masen didominasi oleh habitat hutan pegunungan rendah (37,0%) dan elevasi umumnya diatas 1.500 meter dpl (55,1%). Secara alamiah suhu udara lingkungan di dalam daerah jelajah BD-1 sejuk meskipun pada siang hari, sehingga pergerakan dan penjelajahan tetap nyaman meskipun dilakukan pada siang hari. Hamilton (1976) dan Sunquist (1981) melaporkan bahwa aktivitas macan tutul dan harimau erat kaitannya dengan temperatur udara, mereka umumnya tidak beraktivitas pada saat suhu udara panas di siang hari. Menurut Sunquist (2010) meskipun harimau merupakan satwa yang cenderung nokturnal, pada beberapa kasus perburuan hewan mangsa juga terjadi pada siang hari.
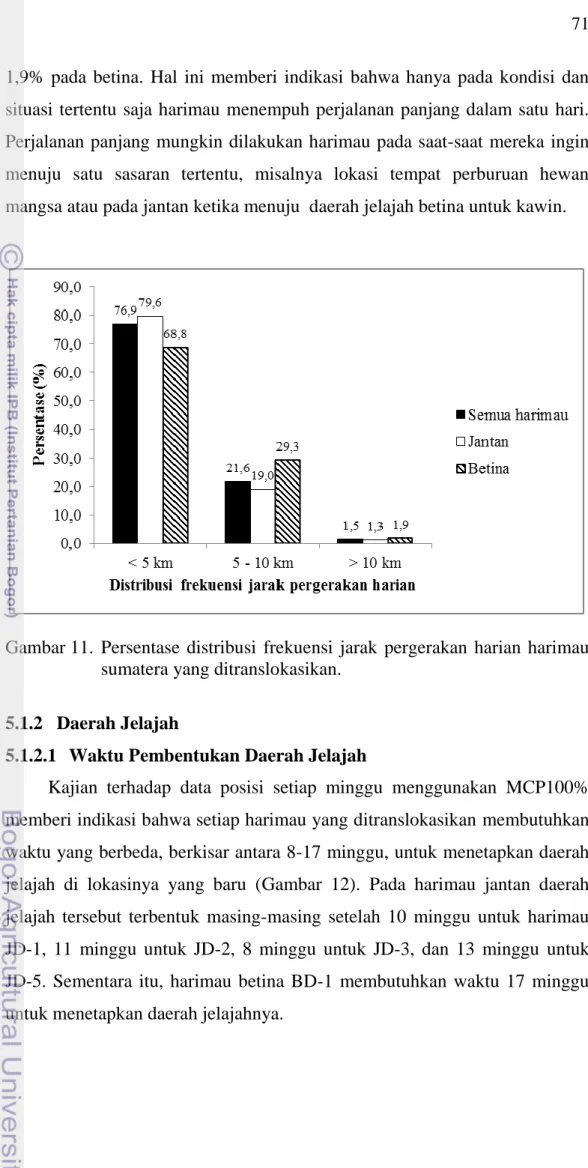
Berdasarkan distribusi frekuensi jarak tempuh hariannya, ternyata umumnya harimau menempuh jarak < 5 km dalam sehari (76,9%) (Gambar 11). Sementara itu, masing-masing 19,0% dan 29,3% frekuensi rata-rata jarak tempuh harian harimau jantan dan betina adalah antara 5-10 km, sedangkan frekuensi untuk jarak tempuh diatas 10 km/hari hanya 1,3% pada jantan dan
71
1,9% pada betina. Hal ini memberi indikasi bahwa hanya pada kondisi dan situasi tertentu saja harimau menempuh perjalanan panjang dalam satu hari. Perjalanan panjang mungkin dilakukan harimau pada saat-saat mereka ingin menuju satu sasaran tertentu, misalnya lokasi tempat perburuan hewan mangsa atau pada jantan ketika menuju daerah jelajah betina untuk kawin.
Gambar 11. Persentase distribusi frekuensi jarak pergerakan harian harimau sumatera yang ditranslokasikan.
5.1.2 Daerah Jelajah
5.1.2.1 Waktu Pembentukan Daerah Jelajah
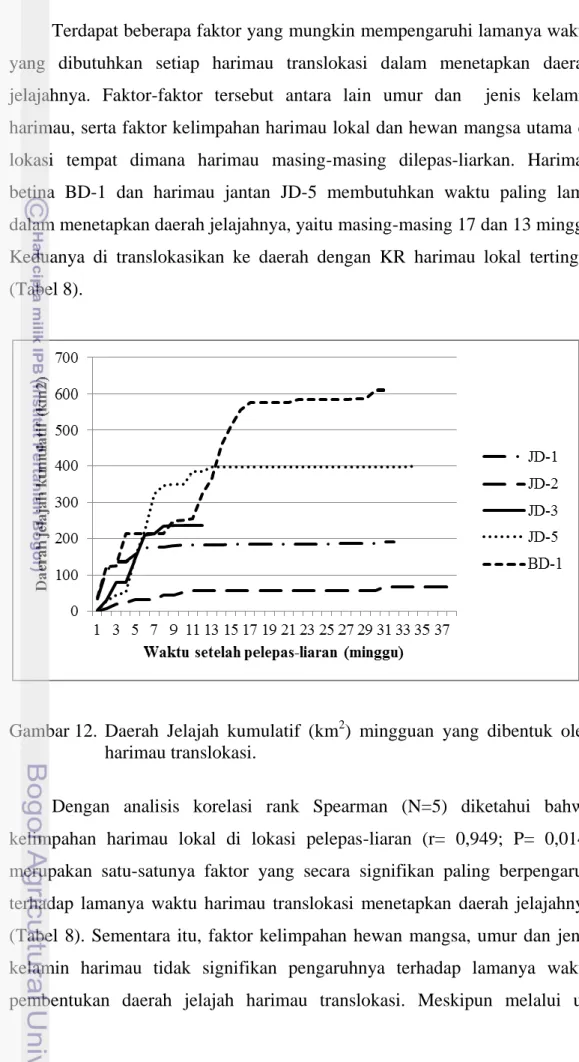
Kajian terhadap data posisi setiap minggu menggunakan MCP100% memberi indikasi bahwa setiap harimau yang ditranslokasikan membutuhkan waktu yang berbeda, berkisar antara 8-17 minggu, untuk menetapkan daerah jelajah di lokasinya yang baru (Gambar 12). Pada harimau jantan daerah jelajah tersebut terbentuk masing-masing setelah 10 minggu untuk harimau JD-1, 11 minggu untuk JD-2, 8 minggu untuk JD-3, dan 13 minggu untuk JD-5. Sementara itu, harimau betina BD-1 membutuhkan waktu 17 minggu untuk menetapkan daerah jelajahnya.
72
Terdapat beberapa faktor yang mungkin mempengaruhi lamanya waktu yang dibutuhkan setiap harimau translokasi dalam menetapkan daerah jelajahnya. Faktor-faktor tersebut antara lain umur dan jenis kelamin harimau, serta faktor kelimpahan harimau lokal dan hewan mangsa utama di lokasi tempat dimana harimau masing-masing dilepas-liarkan. Harimau betina BD-1 dan harimau jantan JD-5 membutuhkan waktu paling lama dalam menetapkan daerah jelajahnya, yaitu masing-masing 17 dan 13 minggu. Keduanya di translokasikan ke daerah dengan KR harimau lokal tertinggi (Tabel 8).
Gambar 12. Daerah Jelajah kumulatif (km2) mingguan yang dibentuk oleh harimau translokasi.
Dengan analisis korelasi rank Spearman (N=5) diketahui bahwa kelimpahan harimau lokal di lokasi pelepas-liaran (r= 0,949; P= 0,014) merupakan satu-satunya faktor yang secara signifikan paling berpengaruh terhadap lamanya waktu harimau translokasi menetapkan daerah jelajahnya (Tabel 8). Sementara itu, faktor kelimpahan hewan mangsa, umur dan jenis kelamin harimau tidak signifikan pengaruhnya terhadap lamanya waktu pembentukan daerah jelajah harimau translokasi. Meskipun melalui uji
73
statistik tidak berpengaruh, namun data menunjukkan adanya kecenderungan bahwa lamanya waktu harimau translokasi membangun daerah jelajah tetapnya, dipengaruhi juga oleh faktor kelimpahan hewan mangsa di lokasi pelepas-liaran serta umur harimau ketika dilepas-liarkan. Harimau JD-1 dan JD-2 ternyata relatif cepat dalam menetapkan daerah jelajah, yaitu 10 dan 11 minggu. Keduanya ditranslokasikan ke areal dengan KR hewan mangsa paling tinggi (yaitu 0,8 tanda/km) dibanding areal translokasi lainnya (Lampiran 1). Baik harimau JD-1 maupun JD-2, keduanya juga berumur paling tua (6 dan 4 tahun) ketika dilepas-liarkan, sehingga sepertinya mereka lebih siap untuk menempati areal kosong di antara daerah jelajah harimau yang telah ada lebih dahulu di sekitar lokasi translokasinya.
Tabel 8. Waktu dibutuhkan harimau translokasi dalam menetapkan daerah jelajahnya serta nilai KR harimau lokal dan mangsa utama pada masing -masing lokasi pelepas-liaran.
ID Lokasi Waktu penetapan (minggu) Kelamin (1=jantan; 2=betina) Umur (thn) KR harimau lokal (tanda/km) KR mangsa utama (tanda/km) JD-1 TNBBS 10 1 6 0,05 0,80 JD-2 TNBBS 11 1 4 0,05 0,80 BD-1 KHUM 17 2 2 0,09 0,25 JD-3 TNGL 8 1 4 0,01 0,45 JD-5 TNKS 13 1 2 0,09 0,31
Menurut laporan Smith et al. (1987), selain harimau jantan soliter yang memiliki wilayah teritori, setiap harimau betina juga memiliki daerah jelajah. Meskipun ukuran daerah jelajahnya lebih kecil dibandingkan jantan, namun daerah jelajah betina lebih stabil. Smith et al. (1987) dalam penelitiannya menemukan beberapa kasus perkelahian antara dua harimau betina yang bertemu pada satu lokasi di dalam hutan. Dengan adanya sistem teritorialitas baik pada harimau jantan maupun betina, maka jika satu harimau ditranslokasikan ke satu wilayah dimana terdapat harimau lokal yang lebih dahulu menghuni wilayah tersebut, maka akan dibutuhkan waktu untuk
74
mencari wilayah kosong yang dapat dijadikan daerah jelajah oleh harimau yang ditranslokasikan tersebut.
5.1.2.2 Luas Daerah Jelajah
Setiap individu harimau yang ditranslokasi dan dilepas-liarkan di kawasan yang berbeda di Sumatera, menetapkan daerah jelajah dengan yang berbeda-beda. Dengan banyaknya data yang dikumpulkan melalui dari kalung GPS memungkinkan dilakukannya pendugaan luas daerah jelajah harimau dengan metode Fixed Kernel (FK). Perhitungan dengan FK95% memberikan variasi luas jelajah harimau jantan antara 37,5-188,1 km2 dan betina 376,8 km2 (Tabel 9). Perkiraan ukuran daerah jelajah dengan metode FK memberikan hasil yang akurat namun membutuhkan sampel data yang besar (Seaman & Powell 1996, Mitchell 2007).
Tabel 9. Luas daerah jelajah harimau yang diamati dengan kalung GPS yang dianalisis berdasarkan Minimum Convex Polygon dan Fixed Kernel.
Harimau Lokasi N hari pengamatan N data posisi Daerah jelajah (km2) MCP (100%) FK (95%) FK (50%) JD-1 TNBBS 224 3.469 191,2 140,9 27,9 JD-2 TNBBS 253 1.288 67,1 37,5 4,9 JD-3 TNGL 79 1.486 236 141,2 28,9 JD-5 TNKS 238 7.007 400 188,1 42,2 BD-1 KHUM 213 6.880 610,3 376,8 80,2
Luas daerah jelajah yang dibangun oleh masing-masing harimau translokasi tidak dipengaruhi oleh umur (r= -0,580; P= 0,306), jenis kelamin (r= 0,000; P= 1,000), kelimpahan harimau lokal (r= 0,264; P= 0,668) serta kelimpahan hewan mangsa (r= 0,667; P= 0,219). Namun demikian terdapat kecenderungan bahwa luas daerah jelajah yang dibentuk oleh setiap harimau translokasi berhubungan dengan kelimpahan hewan mangsa di masing-masing lokasi translokasi. Ahearn et al. (2001) melaporkan bahwa kelimpahan spesies hewan mangsa memainkan peran yang sangat penting dalam menentukan daerah jelajah harimau. Smith et al. (1987) juga
75
berpendapat bahwa ukuran daerah jelajah harimau betina dipengaruhi oleh kelimpahan hewan mangsa dan kualitas habitat.
Harimau jantan JD-1 dan JD-2 membentuk daerah jelajah dengan ukuran terkecil dibanding harimau translokasi lainnya. Kedua harimau tersebut dilepas-liarkan di TNBBS yang kelimpahan relatif hewan mangsa utamanya tertinggi. Sementara itu, harimau betina BD-1 membentuk daerah jelajah dengan ukuran terluas dibandingkan dengan harimau translokasi lainnya. BD-1 dilepas-liarkan di KHUM yang memiliki kelimpahan hewan mangsa utama terendah (Lampiran 1).
Lebih jauh, Griffiths (1994) menyatakan bahwa salah satu faktor yang menentukan daerah jelajah harimau adalah ketersediaan hewan mangsa, dimana luas daerah jelajah harimau meningkat seiring dengan berkurangnya kepadatan hewan mangsa. Studi Franklin et al. (1999) menunjukkan bahwa semakin tinggi kelimpahan hewan mangsa utamanya, maka semakin kecil pula daerah jelajah satu individu harimau. Menurut (Sunquist 2010), luasnya daerah jelajah pada harimau jantan lebih disebabkan untuk penguasaan betina daripada penguasaan sumber pakan.
Rata-rata luas daerah jelajah MCP100% yang dihasilkan dari JD-1 dan JD-2 yang dilepas-liarkan dalam waktu dan lokasi yang sama, di hutan dataran rendah di TNBBS adalah 129 km2. Perkiraan ini mirip dengan perkiraan terdahulu dengan metode yang sama (Franklin et al. 1999), yang menyatakan bahwa luas daerah jelajah harimau jantan dewasa di hutan dataran rendah TN Way Kambas adalah sekitar 110 km2. Luas daerah jelajah MCP100% harimau betina BD-1 yang dilepas-liarkan di KHUM dan harimau jantan yang dilepas-liarkan di TNKS memiliki luas jelajah yang sangat luas masing-masing 610,3 km2 dan 400 km2. Ukuran daerah jelajah tersebut lebih luas jika dibandingkan dengan perkiraan luas jelajah harimau sumatera liar yang pernah ada sebelumnya. Hal ini diakibatkan oleh perbedaan data yang digunakan dalam menduga ukuran daerah jelajah dengan metode MCP. Pendugaan-pendugaan luas daerah jelajah harimau yang ada sebelumnya (Tabel 10) umumnya cenderung underestimate karena
76
dilakukan menggunakan data dari camera-trapping dan radio-tracking. Kedua metode tersebut memiliki keterbatasan baik dalam hal penempatan camera trapping maupun dalam pelacakan signal radio-tracking, sehingga harimau studi sulit dilacak keberadaannya ketika menjauh dari wilayah studi. Estimasi yang dilakukan pada penelitian ini menggunakan data posisi dari kalung GPS, sehingga hasilnya jauh lebih akurat dari hasil-hasil studi yang pernah ada di Sumatera, karena harimau-harimau studi selalu terlacak posisinya dimana pun mereka berada.
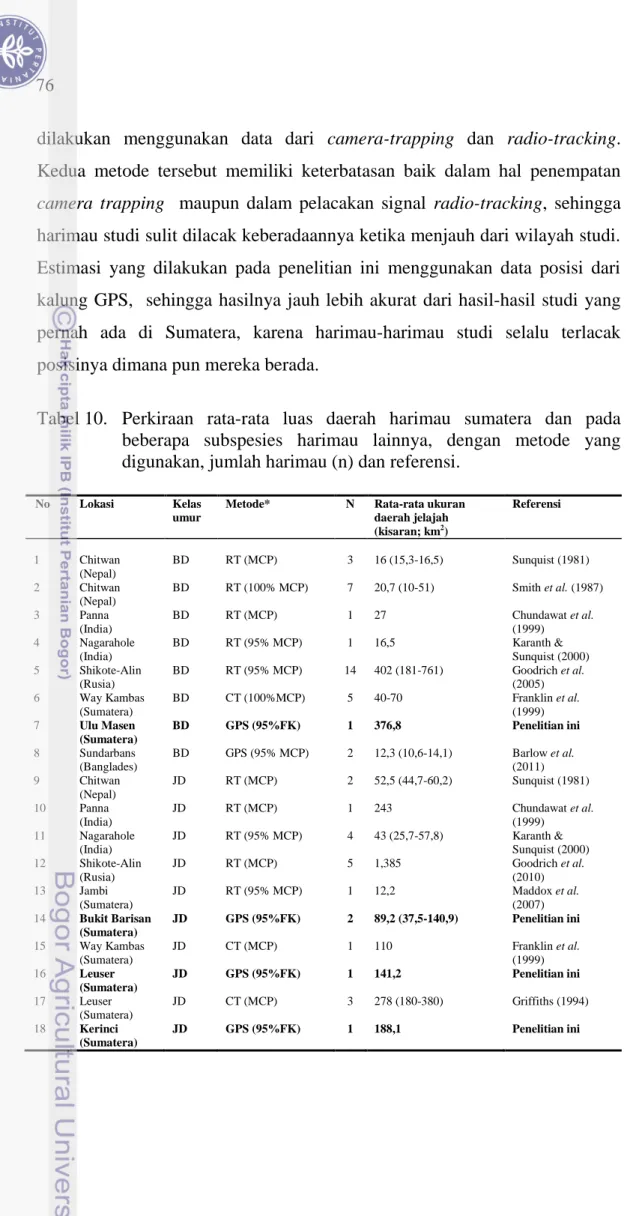
Tabel 10. Perkiraan rata-rata luas daerah harimau sumatera dan pada beberapa subspesies harimau lainnya, dengan metode yang digunakan, jumlah harimau (n) dan referensi.
No Lokasi Kelas
umur
Metode* N Rata-rata ukuran
daerah jelajah (kisaran; km2) Referensi 1 Chitwan (Nepal) BD RT (MCP) 3 16 (15,3-16,5) Sunquist (1981) 2 Chitwan (Nepal) BD RT (100% MCP) 7 20,7 (10-51) Smith et al. (1987) 3 Panna (India) BD RT (MCP) 1 27 Chundawat et al. (1999) 4 Nagarahole (India) BD RT (95% MCP) 1 16,5 Karanth & Sunquist (2000) 5 Shikote-Alin (Rusia) BD RT (95% MCP) 14 402 (181-761) Goodrich et al. (2005) 6 Way Kambas (Sumatera) BD CT (100%MCP) 5 40-70 Franklin et al. (1999) 7 Ulu Masen (Sumatera) BD GPS (95%FK) 1 376,8 Penelitian ini 8 Sundarbans (Banglades) BD GPS (95% MCP) 2 12,3 (10,6-14,1) Barlow et al. (2011) 9 Chitwan (Nepal) JD RT (MCP) 2 52,5 (44,7-60,2) Sunquist (1981) 10 Panna (India) JD RT (MCP) 1 243 Chundawat et al. (1999) 11 Nagarahole (India) JD RT (95% MCP) 4 43 (25,7-57,8) Karanth & Sunquist (2000) 12 Shikote-Alin (Rusia) JD RT (MCP) 5 1,385 Goodrich et al. (2010) 13 Jambi (Sumatera) JD RT (95% MCP) 1 12,2 Maddox et al. (2007) 14 Bukit Barisan (Sumatera) JD GPS (95%FK) 2 89,2 (37,5-140,9) Penelitian ini 15 Way Kambas (Sumatera) JD CT (MCP) 1 110 Franklin et al. (1999) 16 Leuser (Sumatera) JD GPS (95%FK) 1 141,2 Penelitian ini 17 Leuser (Sumatera) JD CT (MCP) 3 278 (180-380) Griffiths (1994) 18 Kerinci (Sumatera) JD GPS (95%FK) 1 188,1 Penelitian ini
77
Apabila data memungkinkan, sebaiknya metode FK digunakan dalam memprediksi ukuran daerah jelajah satwaliar. Penggunaan metode MCP akan selalu overestimate, karena poligon daerah jelajah yang dibentuk dengan metode ini didasarkan pada titik-titik terluar posisi harimau. Namun, pada kenyataanya, sebetulnya banyak areal di dalam daerah jelajah harimau yang dibentuk dengan MCP, sama sekali tidak digunakan oleh harimau dalam penjelajahannya. Menurut Nilsen et al. (2008) sebaiknya penggunaan metode MCP dihindari pada studi-studi ekologi. Mitchell (2007) menyatakan pendugaan luas daerah jelajah dengan metode FK membutuhkan banyak data, sehingga sulit dipenuhi oleh data-data yang dihasilkan dari camera-trapping atau radio-tracking. FK merupakan salah satu metode yang direkomendasikan untuk digunakan dalam menganalisis ukuran daerah jelajah satwa liar (Mitchell 2007).
Pada Tabel 10 juga dapat dilihat bahwa ukuran daerah jelajah harimau sangat bervariasi. Hal ini mengindikasikan bahwa terdapat variasi yang tinggi pada penggunaan wilayah jelajah pada harimau, baik sesama subspesies maupun di antara subspesies. Selain itu, harimau yang ditraslokasikan ke kawasan hutan yang baru akan membutuhkan areal yang lebih luas, sehubungan dengan adanya masa orientasi sebelum mereka membangun daerah jelajah tetapnya. Masa orientasi ini dibutuhkan mengingat pada saat harimau-harimau tersebut dilepas-liarkan di kawasan yang baru, di tempat tersebut sudah terdapat populasi harimau lokal. Oleh karena itu, pengukuran luas daerah jelajah harimau yang ditranslokasikan, sebaiknya dilakukan pada saat harimau tersebut sudah merasa “nyaman” tinggal di lokasinya yang baru.
Kajian yang dilakukan oleh Franklin et al. (1999) menunjukkan bahwa daerah jelajah harimau sumatera betina dewasa berkisar antara 40-70 km2, sedangkan Griffiths (1994) melaporkan bahwa daerah jelajah harimau sumatera jantan dewasa sangat bervariasi, tergantung pada ketinggian habitat dari permukaan laut (dpl). Berdasarkan perkiraannya, luas daerah jelajah harimau jantan dewasa sekitar 180 km2 pada kisaran ketinggian antara 100-600 meter dpl, 274 km2 pada ketinggian antara 600-1.700 meter dpl, dan 380
78
km2 pada ketinggian diatas 1.700 meter dpl. Berdasarkan studi kamera-trap jangka panjang, Maddox et al. (2004) melaporkan bahwa luas daerah jelajah seekor harimau sumatera jantan pada kawasan hutan dataran rendah di Jambi adalah 14,2 km2 sedangkan betina dewasa rata-rata 7,9 km2.
Jantan dewasa JD-1 dan JD-2 ditranslokasikan ke kawasan yang sama pada waktu yang juga bersamaan di hutan dataran rendah TNBBS. Namun, yang menarik adalah bahwa daerah jelajah FK95% yang dibangun oleh kedua harimau jantan tersebut sangat berbeda, dimana daerah jelajah yang dibentuk oleh JD-1 (140,9 km2) hampir empat kali lebih luas dari luas daerah jelajah JD-2 (37,5 km2). Selain itu, selama 7,5 bulan pengamatan, diketahui bahwa hampir 100% daerah jelajah JD-2 tumpang-tindih dan berada di dalam daerah jelajah JD-1. Hal ini memberi indikasi bahwa daerah jelajah harimau jantan juga tidak eksklusif, yakni suatu areal yang menjadi bagian dari daerah jelajah seekor harimau jantan, mungkin juga dapat digunakan oleh jantan lain pada waktu yang berlainan. Selain itu, pada kasus ini ada kemungkinan juga bahwa jantan JD-1 lebih dominan. Pada saat dilepas-liarkan, JD-1 berumur sekitar 6 tahun dengan berat tubuh 122 kg; sedangkan JD-2 berumur sekitar 4 tahun dengan berat tubuh 73 kg. Sebagai harimau jantan yang lebih dominan, JD-1 dapat menggunakan kawasan yang lebih luas, sehingga selain memiliki akses yang lebih besar terhadap hewan mangsa, JD-1 juga memiliki peluang yang lebih besar untuk mendapatkan harimau betina.
Fixed Kernel 50% digunakan untuk menentukan areal inti (core areal) yang paling sering digunakan oleh satwa liar di dalam daerah jelajahnya (Aliaga-Rossel et al. 2008). Menggunakan metode FK50% (Edwards et al. 2001, Barlow et al. 2011) dapat diprediksi bahwa luas daerah jelajah inti individu harimau jantan translokasi adalah masing-masing JD-1= 27,9 km2, JD-2= 4,9 km2, JD-3= 28,9 km2 dan JD-5= 42,2 km2 (Lampiran 7, 8, 9 dan 10); sedangkan daerah jelajah inti harimau betina BD-1= 80,2 km2 (Lampiran 11). Areal inti yang sering digunakan harimau sumatera translokasi berkisar antara 13,1% - 22,4% dari ukuran daerah jelajahnya masing-masing. Dari
79
data yang terkumpul, ada kecenderungan bahwa semakin besar luas daerah jelajah harimau translokasi, semakin besar pula persentase luas areal intinya.
Selanjutnya memperhatikan pada bentuk dari setiap daerah jelajah yang dibangun dengan metode FK95%, terlihat bahwa daerah jelajah harimau jantan JD-3 yang ditranslokasikan di kawasan hutan TNGL bentuknya memanjang (Lampiran 9). Hal ini mengindikasikan bahwa sebetulnya sampai akhir masa pengamatan (selama 97 hari), harimau JD-3 ini masih belum menetapkan daerah jelajahnya. Kemungkinan besar di lokasi dimana JD-3 dilepas-liarkan sudah ada harimau jantan dewasa lain yang mengokupasi wilayah tersebut, sehingga JD-3 masih menjadi individu harimau pelintas (transient atau floater) yang belum memiliki daerah jelajah yang tetap (Karanth & Chundawat 2002).
5.1.2.3 Bentuk Daerah Jelajah
Pada Lampiran 7 dapat dilihat bahwa selama masa pengamatan daerah jelajah harimau jantan dewasa JD-1 yang ditranslokasikan ke TNBBS berbentuk menyerupai trapesium terbalik dengan panjang 20,08 km dan lebar 10,95 km. Namun, tidak semua areal di dalam poligon daerah jelajah digunakan oleh 1. Pada tingkat FK95% terlihat bentuk daerah jelajah JD-1 menyerupai lingkaran lonjong (oval) dengan jarak terpanjang JD-16,33 km dan terlebar 10,95 km.
Lampiran 8 memperlihatkan bahwa bentuk daerah jelajah harimau jantan JD-2, yang lokasi translokasinya sama dengan JD-1 yaitu di TNBBS, terlihat seperti layang-layang terbalik dengan panjang 13,30 km dan lebar 8,58 km. Di dalam poligon daerah jelajah JD-2 juga terlihat areal-areal yang tidak digunakan oleh JD-2 selama masa pengamatan. Bentuk daerah jelajah JD-2 yang dihasilkan oleh FK95% adalah seperti lingkaran lonjong (oval) dengan jarak terpanjang 6,89 km dan terlebar 5,28 km.
Lain halnya dengan JD-3 yang ditranslokasi ke TNGL, harimau jantan dewasa ini membangun daerah jelajah yang berbentuk seperti jajar genjang dengan panjang 31,47 km dan lebar 13,88 (Lampiran 9). Harimau JD-3 ini
80
diduga belum menentukan daerah jelajah tetapnya hingga akhir masa 2,5 bulan observasi, karena daerah jelajah hasil analisis FK95% masih berbentuk seperti koridor dengan jarak terpanjang 31,47 km dan terlebar 7,69 km.
Pada Lampiran 10 diperlihatkan bahwa harimau jantan JD-5 selama 8 bulan pengamatan di TNKS membangun daerah jelajah yang bentuknya seperti persegi panjang 32,16 km x 14,33 km. Sama halnya dengan harimau lainnya yang mana di dalam daerah jelajahnya ini terdapat wilayah-wilayah yang sebetulnya tidak digunakan oleh JD-5 selama penjelajahannya. Dengan metode FK95% diketahui bahwa bentuk daerah jelajah JD-5 adalah belah ketupat berukuran 13,79 km x 13,09 km.
Satu-satunya harimau betina dewasa yang ditranslokasi ke kawasan Ulu Masen membentuk daerah jelajah seperti lingkaran lonjong (oval) dengan panjang 37,14 km dan panjang 25,61 km (Lampiran 11). Dalam wilayah jelajah yang terbangun ini juga sebetulnya banyak areal yang tidak digunakan BD-1 dalam pergerakannya. Melalui FK95%, daerah jelajah harimau betina BD-1 berbentuk seperti layang-layang yang berukuran 35,80 km x 23,01 km. Dari hasil pengamatan diketahui harimau jantan JD-1 dan betina BD-1 menggunakan lokasi pelepas-liarannya sebagai bagian dari daerah jelajahnya. Harimau jantan JD-2 dan JD-5 sepertinya meninggalkan areal pelepas-liaran dan membangun daerah jelajah pada jarak masing-masing 4,06 km dan 14,64 km dari tempat mereka dilepas-liarkan. Harimau jantan JD-3 meninggalkan lokasi translokasi sejauh 31,52 km dan belum menetapkan daerah jelajahnya hingga akhir masa pengamatan. Menjauhnya JD-2, JD-5 dan JD-3 dari lokasi pelepas-liarannya mungkin akibat sudah adanya harimau lain yang mendiami lokasi pelepas-liarannya, sehingga mereka harus berusaha mencari wilayah kosong untuk dijadikan daerah jelajahnya. Contohnya, bergesernya harimau jantan JD-2 dari lokasi peliarannya akibat wilayah tersebut telah dijadikan bagian dari daerah jelajah harimau jantan JD-1, yang diliarkan pada lokasi dan waktu yang sama dengan JD-2.
Berdasarkan survey transek sign (Lampiran 1) diketahui juga bahwa lokasi pelepas-liaran harimau jantan JD-3 dan JD-5 telah kuasai oleh harimau
81
lokal yang ada sebelumnya, namun kedua harimau ini kemungkinan kalah bersaing sehingga mereka harus pergi meninggalkan lokasi pelepas-liarannya. Faktor lain yang juga mungkin turut mendorong harimau JD-3 dan JD-5 untuk pergi meninggalkan lokasi pelepas-liarannya adalah tipe vegetasi dan topografi wilayah. JD-3 dilepas-liarkan di areal yang didominasi oleh hutan pegunungan rendah dan tinggi dengan topografi mayoritas sangat curam, sedangkan JD-5 dilepas-liarkan di areal yang didominasi oleh perkebunan sawit skala besar. Kedua harimau tersebut (JD-3 dan JD-5) diketahui bermigrasi ke areal yang didominasi oleh hutan dataran rendah dan vegetasi belukar/hutan sekunder muda.
5.1.2.4 Karakteristik Daerah Jelajah
Lima harimau sumatera yang diamati dilepas-liarkan di empat lokasi yang berbeda, yaitu TN Bukit Barisan Selatan, TN Gunung Leuser, TN Kerinci Seblat dan kawasan hutan Ulu Masen. Hasil overlay antara data posisi dengan peta tutupan vegetasi MODIS (Moderate-resolution Imaging Spectroradiometer), menunjukkan bahwa areal yang dijadikan daerah jelajah harimau jantan JD-1 di TNBBS didominasi oleh vegetasi belukar/hutan sekunder muda (83,62%) dan hutan dataran rendah (13,95%). Daerah jelajah harimau jantan JD-2, yang juga dilepas-liarkan di TNBBS, juga umumya merupakan vegetasi belukar/hutan sekunder muda (90,00%) dengan sebagain kecil hutan dataran rendah (9,24%) (Lampiran 12).
Elevasi di dalam daerah jelajah JD-1 dan JD-2 seluruhnya merupakan dataran rendah dengan ketinggian antara 0-500 meter dpl (Lampiran 13), yang mayoritas topografi datar (kelas slope 0-8 persen) (87,46%) dengan sedikit landai (kelas slope 8-15 persen) (12,04%) (Lampiran 14). Selama pengamatan, kedua harimau baik JD-1 maupun JD-2 menghabiskan seluruh waktunya (100%) pada areal dataran rendah dengan ketinggian 0-500 meter dpl, dan seluruh waktunya (100%) pada areal-areal yang datar dengan kelerengan 0-8 persen. Sesuai dengan pernyataan O‟Brien et. al. (2003) bahwa medan pada ketinggian 0-500 meter dpl di TNBBS didominasi oleh
82
dataran dan perbukitan. Kawasan yang dijadikan daerah jelajah 1 dan JD-2 merupakan kaki Pegunungan Bukit Barisan dengan kisaran curah hujan antara 3.000 mm hingga lebih dari 4.000 mm per tahun, dan memiliki suhu udara 22 hingga 35°C. Hasil penelitian O‟Brien et. al. (2003) menunjukkan bahwa di kawasan tempat dimana JD-1 dan JD-2 ditranslokasikan memiliki kepadatan harimau lokal 1,6 harimau/100 km2. Hewan mangsa utama yang umum dijumpai di dalam daerah jelajah JD-1 dan JD-2 adalah babi hutan (Sus scrofa) and rusa sambar (Rusa unicolor). Hasil survey transek sign sepanjang 92 km di lokasi translokasi TNBBS, menghasilkan kelimpahan relatif (KR) harimau lokal sebesar 0,05 jejak/km dan KR hewan mangsa utama (babi hutan, rusa sambar dan kijang) sebesar 0,80 jejak/km (Lampiran 1).
Areal yang menjadi daerah jelajah harimau jantan JD-3 di TNGL secara umum didominasi oleh vegetasi hutan pegunungan rendah (50,22%) dan hutan dataran rendah (36,87%), dengan sebagian kecil hutan pegunungan tinggi (7,54%) serta vegetasi belukar/hutan sekunder muda (5,36%) (Lampiran 12). Ketinggian elevasi pada wilayah jelajah JD-3 bervariasi mulai dari kurang dari 100 meter hingga diatas 2.000 meter dpl, namun kelas ketinggian yang mendominasi wilayah tersebut adalah berturut-turut hutan antara 500-1.000 meter dpl (34,18%), 0-500 meter dpl (33,82%), dan 1.000-1.500 meter dpl (26,24%) (Lampiran 13). Kondisi medan (terrain) sangat curam (kelas slope > 40 persen) (60,38%) medominasi lokasi translokasi JD-3, dimana topografi yang lainnya adalah curam (17,37%) dan agak curam (10,74%). Sementara itu areal dengan topografi datar dan landai (kelas slope 0-15 persen) hanya 10,16% dari total luas areal (Lampiran 14). Namun, selama 2,5 bulan observasi harimau jantan JD-3 menghabiskan 38% waktunya untuk menjelajah pada areal dengan ketinggian 0-500 meter dpl dan 44% waktunya mengeksplorasi areal dengan elevasi 500-1.000 meter dpl. Selain itu, 42% waktu JD-3 digunakan untuk menjelajah pada daerah-daerah yang datar, dan 41% waktu lainnya digunakan untuk berkelana pada daerah-daerah dengan topografi sangat curam.
83
Seperti daerah-daerah lain di Sumatera, kawasan sekitar daerah jelajah JD-3 memiliki kisaran curah hujan antara 2.000 mm hingga 3.000 mm per tahun, dengan fluktuasi suhu udara mulai 21-28 °C. Wibisono & Pusparini (2011) melaporkan bahwa kepadatan harimau lokal minimum di sekitar lokasi translokasi JD-3 adalah 0,3 harimau/100 km2. Mereka juga mendapati bahwa rusa sambar (Rusa unicolor), babi hutan (Sus scrofa) dan kijang (Muntiacus muntjac) merupakan hewan-hewan mangsa utama harimau di lokasi tersebut. Berdasarkan hasil transek sign sepanjang 83 km di sekitar lokasi translokasi JD-3 di TNGL diketahui bahwa KR harimau lokal sebesar 0,01 jejak/km dan KR hewan mangsa sebesar 0,45 jejak/km (Lampiran 1).
Tutupan vegetasi mayoritas di dalam daerah jelajah harimau jantan JD-5 di TNKS adalah belukar/hutan sekunder muda (40,37%) dan hutan dataran rendah (37,11%). Selain itu, di dalam daerah jelajah JD-5 juga terdapat vegetasi hutan pegunungan rendah (17,99%), hutan pegunungan tinggi (3,93%) dan perkebunan sawit skala besar (5,01%) (Lampiran 12). Daerah jelajah JD-5 juga didominasi oleh areal dengan ketinggian 0-500 meter dpl (62,59,80%) dan 500-1.000 meter dpl (26,72%) (Lampiran 13), serta mayoritas topografinya datar (61,53%) dan landai (19,57%) (Lampiran 14). Harimau jantan JD-5 menghabiskan sebagian besar waktunya (65%) menjelajah pada areal yang datar dan landai (12%). Harimau JD-5 juga memilih wilayah pada ketinggian 0-500 meter dpl (64%) dan 500-1.000 meter dpl (34%) sebagai tempat mencari hewan mangsa dan beraktivitas lainnya. Menurut Linkie et al. (2006), rata-rata curah hujan tahunannya sekitar 3.000 mm dan kisaran suhu udara antara 7 hingga 28°C. Lebih jauh Linkie et al. (2006) memperkirakan bahwa kepadatan harimau di areal translokasi JD-5 di TNKS adalah sekitar 1,5-3,3 harimau/100 km2. Dinata & Sugadrjito (2008) mengidentifikasi bahwa hewan mangsa utama harimau di kawasan ini adalah babi hutan (Sus scrofa), rusa sambar (Rusa unicolor) dan kijang (Muntiacus muntjac). Survey transek sign sepanjang 136 km di TNKS menghasilkan KR harimau lokal 0,09 jejak/km dan KR hewan mangsa 0,31 jejak/km (Lampiran 1).
84
Kawasan yang dijadikan daerah jelajah harimau betina BD-1 di Ulu Masen didominasi oleh tiga tipe tutupan vegetasi, yaitu belukar/hutan sekunder muda (33,29%), hutan pegunungan rendah (30,84%), dan hutan pegunungan tinggi (17,73%); sedangkan hutan dataran rendah hanya 11,56% dari luas total daerah jelajah BD-1 (Lampiran 12). Mayoritas kawasan tersebut merupakan dataran tinggi pada elevasi > 2.000 meter dpl (29,22%) dan antara 1.500-2.000 meter dpl (25,88%). Selebihnya adalah areal-areal dengan ketinggian 1.000-15.00 meter dpl (18,32%) dan 0-500 meter dpl (19,76%) (Lampiran 13). Areal-areal dengan topografi sangat curam (kelas slope > 40 persen) dan curam umum dijumpai di sekitar daerah jelajah BD-1 (Lampiran 14). Namun, harimau betina BD-1 menghabiskan lebih dari separuh waktunya (64%) beraktivitas pada daerah dengan topografi datar dan landai. BD-1 juga menghabiskan 64% waktunya dihabiskan di areal-areal dengan ketinggian 0-500 meter dpl dan 500-1.000 m dpl (15%). Pada Lampiran 1 dapat dilihat bahwa survey transek sepanjang 129 km di sekitar lokasi translokasi BD-1 di Ulu Masen menghasilkan KR harimau lokal 0,09 jejak/km dan KR hewan mangsa 0,25 jejak/km.
5.2 Pola Aktivitas
Harimau jantan JD-1 terdeteksi aktif pada 1.483 (42,8%) dari 3.469 data aktivitas yang terbaca, sedangkan jantan JD-5 terdeteksi aktif pada 3.465 (49,5%) dari 7.007 data aktivitasnya. Hasil penelitian serupa pada jenis Felidae lainnya di Thailand mengungkapkan bahwa macan dahan (Neofelis nebulosa) jantan dan betina aktif selama masing-masing 57% dan 59% selama masa pengamatan (Grassman et al. 2005). Hasil penelitian ini juga menunjukkan bahwa harimau sumatera jantan pada malam hari (antara pukul 18:00-06:00) menghabiskan 45,5% waktunya untuk beraktivitas dan bergerak. Menurut Sunquist (1981) harimau bengal di Nepal menghabiskan 42% waktu malam harinya untuk beraktivitas dan melakukan pergerakan. Aktivitas harimau pada malam hari mungkin ada hubungannya dengan suhu udara dan aktivitas hewan mangsa harimau karena harimau umumnya melakukan
85
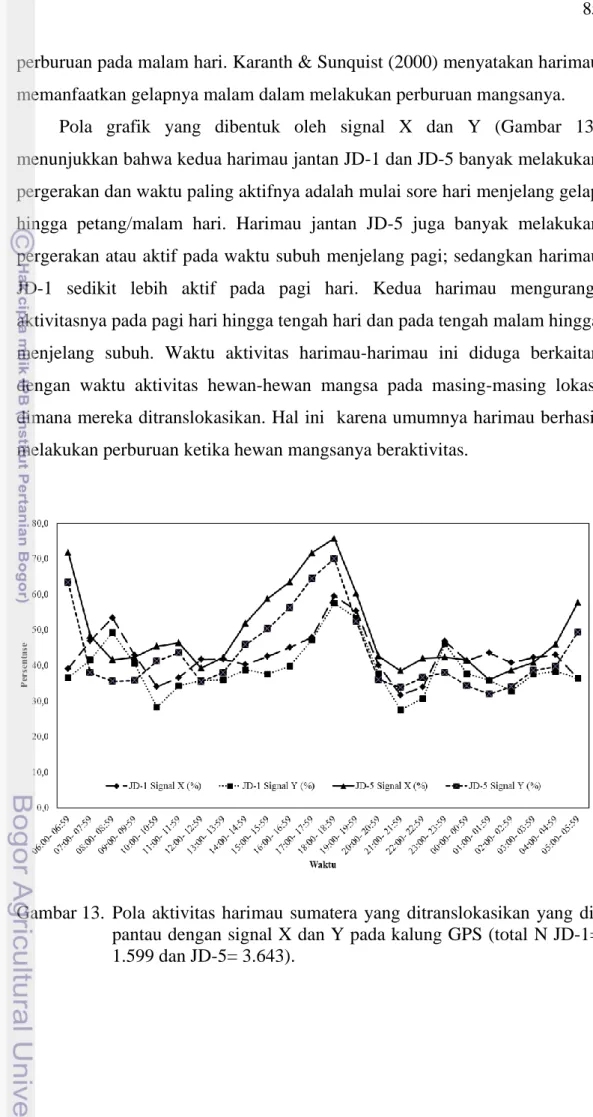
perburuan pada malam hari. Karanth & Sunquist (2000) menyatakan harimau memanfaatkan gelapnya malam dalam melakukan perburuan mangsanya.
Pola grafik yang dibentuk oleh signal X dan Y (Gambar 13) menunjukkan bahwa kedua harimau jantan JD-1 dan JD-5 banyak melakukan pergerakan dan waktu paling aktifnya adalah mulai sore hari menjelang gelap hingga petang/malam hari. Harimau jantan JD-5 juga banyak melakukan pergerakan atau aktif pada waktu subuh menjelang pagi; sedangkan harimau JD-1 sedikit lebih aktif pada pagi hari. Kedua harimau mengurangi aktivitasnya pada pagi hari hingga tengah hari dan pada tengah malam hingga menjelang subuh. Waktu aktivitas harimau-harimau ini diduga berkaitan dengan waktu aktivitas hewan-hewan mangsa pada masing-masing lokasi dimana mereka ditranslokasikan. Hal ini karena umumnya harimau berhasil melakukan perburuan ketika hewan mangsanya beraktivitas.
Gambar 13. Pola aktivitas harimau sumatera yang ditranslokasikan yang di-pantau dengan signal X dan Y pada kalung GPS (total N JD-1= 1.599 dan JD-5= 3.643).
86
Sunquist (1981) menyatakan bahwa waktu aktivitas harimau tidak ada hubungannya dengan waktu terbit dan terbenamnya matahari, namun memang cenderung lebih nokturnal karena kadang-kadang ditemukan aktif pada siang hari. Pada Gambar 13 dapat dilihat bahwa pada siang hari harimau translokasi meningkat aktivitasnya dimulai pada pukul 14:00 WIB dan terus meningkat hingga malam. Pada malam hari, peningkatan aktivitas harimau translokasi terjadi hingga pukul 22:00 WIB dan meninkat kembali pada waktu subuh hingga awal pagi. Sunquist (1981) menjelaskan bahwa harimau memiliki pola aktivitas musiman yang sepertinya dipengaruhi oleh beberapa faktor lingkungan, termasuk jadwal waktu aktivitas hewan mangsa dan suhu udara. Lebih jauh dijelaskan juga bahwa manakala tidak ada perbedaan dalam jumlah waktu aktivitas antara jantan dan betina, mungkin dipengaruhi oleh status sosial atau kondisi reproduktif setiap individu.
Hasil pengamatan menunjukkan bahwa periode waktu paling aktif harimau jantan adalah pada petang/malam hari, yaitu antara pukul 18:00-22:00 WIB (w= 1,19; Lampiran 15). Secara individual ada sedikit perbedaan dimana harimau jantan JD-1 yang ditranslokasikan ke TNBBS memilih waktu paling aktifnya pada petang/malam (pukul 18:00-22:00) dan pagi (pukul 06:00-10:00) hari (Gambar 14). Hal ini mengindikasikan bahwa terdapat perbedaan yang signifikan dalam penggunaan waktu aktif harimau (χ2
hitung = 91,96; χ2 0,05; 5 = 11,07). Pola waktu aktif harimau JD-1 ini
sepertinya sangat erat kaitannya dengan waktu aktivitas hewan mangsanya, karena di hutan-hutan Sumatera hewan-hewan mangsa utama harimau seperti rusa sambar, kijang dan babi hutan, umumnya mencari pakan pada awal pagi hari dan pada sore hari hingga menjelang malam. Selain itu, dengan berkurangnya waktu aktivitas harimau pada siang hari mengindikasikan bahwa harimau sumatera translokasi juga menghindari panasnya sinar matahari. Penelitian-penelitian sebelumnya baik di Nepal maupun di Sumatera telah membuktikan bahwa pola aktivitas harimau sangat erat kaitannya dengan waktu aktif hewan mangsanya. Hal ini karena harimau akan lebih mudah melakukan perburuan hewan mangsanya pada saat
87
aktivitas hewan mangsa tinggi, yaitu ketika matahari mulai terbenam hingga menjelang tengah malam serta ketika matahari mulai terbit hingga awal pagi hari (Sunquist 1981, Fata 2011, Linkie & Ridout 2011).
Gambar 14. Grafik pola persentase pola aktivitas harimau sumatera yang di-translokasikan dalam 24 jam berdasarkan interval waktu setiap 4 jam.
Temuan waktu aktif harimau ini hampir serupa dengan hasil penelitian sebelumnya menggunakan camera-trapping (Fata 2011) yang melaporkan bahwa peningkatan aktivitas harimau sumatera terjadi pada selang waktu menjelang subuh hingga awal pagi, akhir siang menjelang sore, serta senja hingga menjelang tengah malam. Meskipun Sunquist (1981) dan Sunquist (2010) menyatakan bahwa harimau bengal di cenderung nokturnal, namun dalam penelitian ini ditemukan bahwa ternyata harimau sumatera yang ditraslokasikan tidak sepenuhnya nokturnal, bahkan ada kecenderungan merupakan satwa krepuskular, yakni hanya aktif pada senja menjelang malam hari saja. Temuan ini didukung oleh Perry (1964 diacu dalam Sunquist 1981) yang menyatakan bahwa harimau adalah satwa yang nokturnal dan krepuskular.
88
Perbedaan waktu aktif harimau sumatera translokasi dengan dengan harimau bengal di Nepal, dimana mereka umumnya aktif dan bergerak mulai matahari terbenam hingga menjelang pagi (Sunquist 1981), kemungkinan diakibatkan oleh adanya perbedaan waktu perilaku makan pada hewan-hewan ungulata mangsa harimau di kedua tempat tersebut. Schaller (1967) melaporkan bahwa harimau bengal di India waktu paling aktifnya adalah pada malam hari dan beristirahat mulai pagi menjelang siang hingga menjelang sore, namun kadang-kadang ada juga harimau yang berburu pada siang hari apabila harimau tersebut gagal menangkap hewan mangsa pada malam hari sebelumnya. Sunquist (1981) telah membuktikan bahwa waktu aktif harimau erat kaitannya dengan temperatur udara, dimana harimau bengal di Nepal umumnya beristirahat pada areal dengan tutupan vegetasi rapat di sepanjang aliran sungai pada siang hari di musim panas. Hamilton (1976) melaporkan hal yang sama, dimana macan tutul di Afrika umumnya tidak beraktivitas pada saat suhu udara panas di siang hari.
5.3 Pemilihan Habitat
Seleksi habitat merupakan satu proses dimana individu-individu satwa liar yang secara preferensial memanfaatkan habitat-habitat yang tersedia pada satu lansekap (Morris 2003). Khan & Chivers (2007) menyatakan bahwa ada indikasi bahwa harimau memiliki kesukaan (preferensi) terhadap satu habitat tertentu yang disesuaikan dengan beberapa aktivitasnya.
Hasil overlay data posisi dengan peta tutupan vegetasi memperlihatkan bahwa semua harimau yang ditranslokasikan terbukti secara signifikan mempunyai preferensi terhadap tipe habitat tertentu di masing-masing lokasi dimana mereka dilepas-liarkan (Tabel 11). Analisis preferensi dengan metode Neu juga mempertegas bahwa setiap harimau translokasi memilih tipe tutupan vegetasi tertentu sebagai habitat utamanya.
89
Tabel 11. Tipe habitat yang paling disukai oleh harimau translokasi di masing -masing lokasi pelepas-liaran.
Harimau Lokasi Hasil uji Chi-square Habitat yang paling disukai (nilai indeks Neu/W)
JD-1 TNBBS χ2hitung= 304,04 >
χ2
0,05;5= 11,07
Belukar/hutan sekunder muda (W= 1,18)
JD-2 TNBBS χ2hitung= 2.840,72 >
χ2
0,05;5= 11,07
Belukar/hutan sekunder muda (W= 1,21)
JD-3 TNGL χ2hitung= 306,96 >
χ2
0,05;8= 15,51
Hutan pegunungan rendah (W= 1,41)
JD-5 TNKS χ2hitung= 3.551,99 >
χ2
0,05;9= 16,92
Hutan dataran rendah (W= 1,98)
BD-1 EUM χ2hitung= 3.234,16 >
χ2
0,05;6= 12,59
Belukar/hutan sekunder muda (W= 2,17)
Harimau JD-1 dan JD-2 yang diliarkan di TNBBS serta JD-5 di TNKS, mereka menggunakan tutupan vegetasi belukar/hutan sekunder muda dengan intensitas yang sangat tinggi (93,4%, 96,3% dan 58,6%). Hal ini terjadi karena memang tipe tutupan vegetasi belukar/hutan sekunder muda mendominasi lansekap dimana mereka dilepas-liarkan (79,4% di TNBBS dan 41,6% di TNKS). Tingginya intensitas penggunaan vegetasi belukar/hutan sekunder muda sangat erat kaitannya dengan kesukaan hewan mangsa utama harimau (ungulata/herbivora) mencari makan pada areal-areal tersebut. Namun, harimau-harimau tersebut juga menggunakan hutan dataran rendah sebagai habitat yang juga digunakan dengan intensitas tinggi kedua. Demikian pula dengan harimau JD-3 yang dilepas-liarkan di TNGL, mengkombinasikan penggunaan hutan pegunungan rendah (42,8%) dengan hutan dataran rendah (30,1% ) sebagai habitat utamanya. Hal ini juga terjadi karena kedua tipe tutupan vegetasi tersebut memang mendominasi lansekap dimana JD-3 ditranslokasikan. Dengan demikian ada kecenderungan bahwa setiap harimau memang memiliki preferensi terhadap tipe habitat tertentu, namun pemilihan ini selalu didasarkan atas tipe habitat alami yang mendominasi kawasan tempat mereka dilepas-liarkan (Gambar 15, 16, 17,18 dan 19). Dinata & Sugardjito (2008) menyatakan bahwa harimau sumatera cenderung lebih menyukai hutan dataran rendah sebagai habitatnya, karena
90
hutan dataran rendah dapat mendukung biomassa hewan-hewan ungulata besar (Santiapillai & Ramono 1993), seperti babi hutan (Sus scrofa), rusa sambar (Rusa unicolor) dan kijang (Muntiacus muntjak) yang merupakan hewan mangsa utamanya. Rajapandian (2009) menginformasikan bahwa di kawasan Terai Arc, India, harimau menyenangi habitat-habitat hutan lebat yang kesesuaiannya tinggi dengan hewan mangsa utama mereka.
Selain itu, terbukti juga bahwa keberadaan tutupan habitat hutan tetap penting bagi kehidupan harimau. Meskipun Sunquist et al. (1999) menyatakan bahwa secara global harimau menghuni berbagai tipe habitat dan mampu beradaptasi pada berbagai kondisi lingkungan, namun hasil penelitian Sunarto et al. (2012) telah membuktikan bahwa harimau di Sumatera merupakan satwa yang sangat bergantung pada dan lebih menyukai kawasan hutan alam. Menurut mereka, harimau juga menggunakan kawasan perkebunan sawit dan hutan akasia, namun proporsinya sangat kecil dibandingkan dengan luas kawasan yang tersedia. Hasil studi ini di lokasi translokasi TNKS mendukung pernyataan tersebut, dimana meskipun perkebunan sawit menutupi 18,5% lansekap pelepas-liaran di TNKS, harimau JD-5 hanya menggunakan 0,6% frekuensi waktunya di kawasan perkebunan sawit. Selain itu, Maddox et al. (2007) juga berpendapat bahwa harimau sumatera sering menggunakan lahan belukar/hutan sekunder muda di kawasan perkebunan sawit, namun tidak memasuki wilayah interior dari areal yang sudah ditanami pohon sawit. Sunarto (2011) mengemukakan bahwa harimau sumatera umumnya terdeteksi di areal-areal yang memiliki kerapatan vegetasi bawah yang rapat.
91
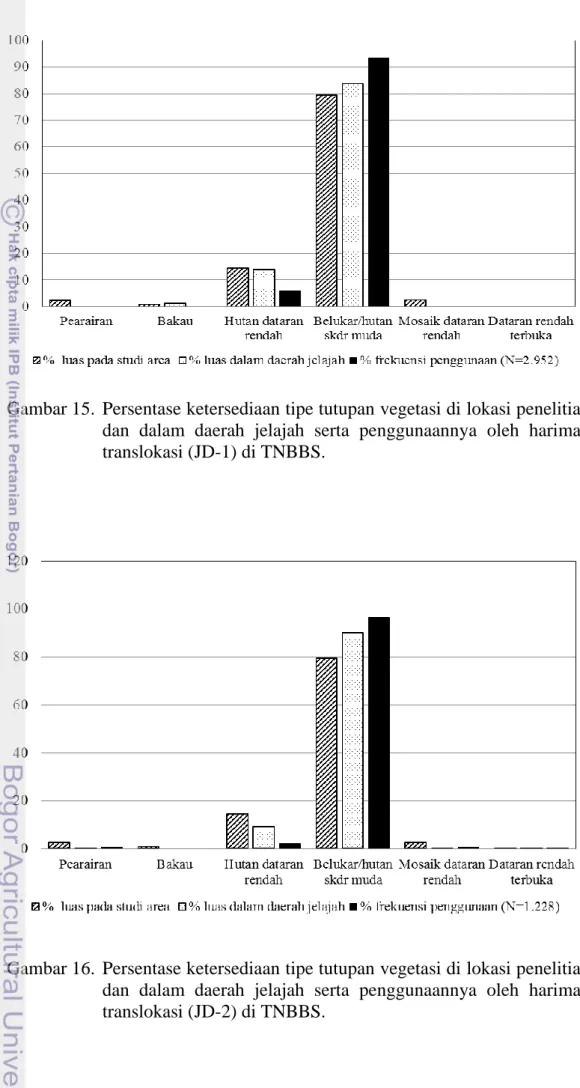
Gambar 15. Persentase ketersediaan tipe tutupan vegetasi di lokasi penelitian dan dalam daerah jelajah serta penggunaannya oleh harimau translokasi (JD-1) di TNBBS.
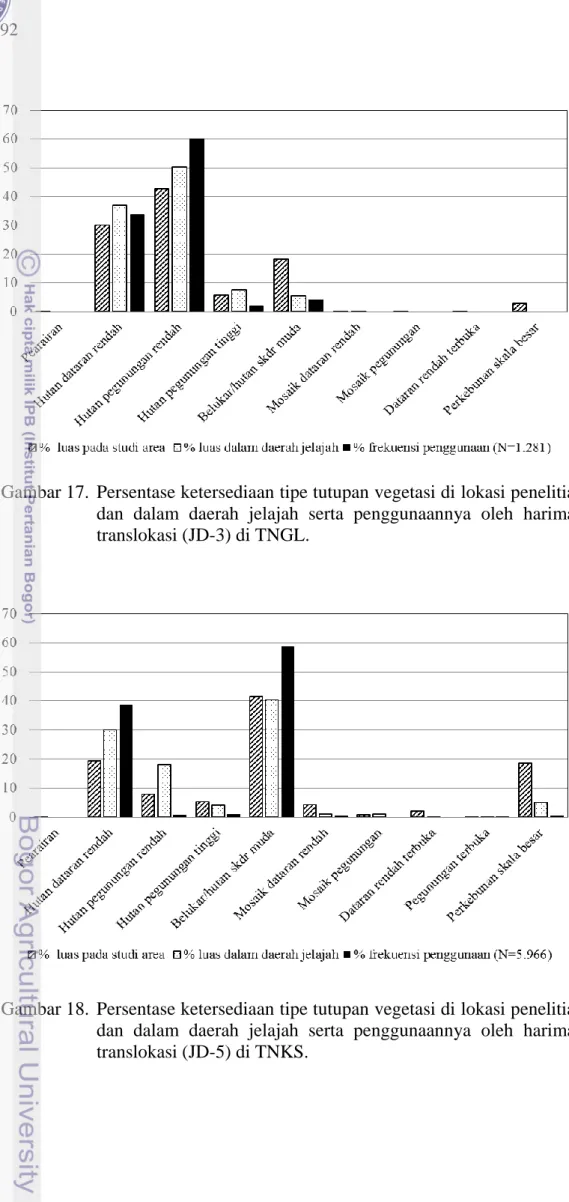
Gambar 16. Persentase ketersediaan tipe tutupan vegetasi di lokasi penelitian dan dalam daerah jelajah serta penggunaannya oleh harimau translokasi (JD-2) di TNBBS.
92
Gambar 17. Persentase ketersediaan tipe tutupan vegetasi di lokasi penelitian dan dalam daerah jelajah serta penggunaannya oleh harimau translokasi (JD-3) di TNGL.
Gambar 18. Persentase ketersediaan tipe tutupan vegetasi di lokasi penelitian dan dalam daerah jelajah serta penggunaannya oleh harimau translokasi (JD-5) di TNKS.
93
Gambar 19. Persentase ketersediaan tipe tutupan vegetasi di lokasi penelitian dan dalam daerah jelajah serta penggunaannya oleh harimau translokasi (BD-1) di KHUM.
5.3.1 Penggunaan Habitat pada Siang dan Malam
Hasil pengamatan menunjukkan bahwa penggunaan setiap tipe habitat utama pada siang hari dan malam hari oleh masing-masing harimau di lokasi translokasi ada sedikit perbedaan. Harimau jantan JD-1 yang translokasikan ke TNBBS menggunakan habitat belukar/hutan sekunder muda sebesar 93,6% pada siang hari dan 93,3% pada malam hari. Harimau JD-3 di TNGL menghabiskan 62,2% waktu siangnya dan 58,2% waktu malamnya di habitat hutan pegunungan rendah. Sementara itu, harimau JD-5 menggunakan 54,1% waktu siangnya dan 62,3% waktu malamnya di habitat belukar/hutan sekunder muda. Sebaliknya, harimau betina BD-1 menggunakan 56,7% waktu siangnya dan 55,1% waktu malamnya menjelajah habitat semak/hutan sekunder muda (Tabel 12).
94
Tabel 12. Persentase penggunaan habitat/tutupan vegetasi oleh harimau pada siang (S) dan malam (M) hari di masing-masing lokasi translokasi.
Tipe habitat/tutupan vegetasi
Persentase frekuensi penggunaan (%) Harimau JD-1 Harimau JD-3 Harimau JD-5 Harimau BD-1 S M S M S M S M Pearairan 0,0 0,4 0,0 0,0 0,0 0,0 Bakau 0,1 0,2
Hutan rawa gambut
Hutan dataran rendah 6,4 5,7 32,2 35,2 43,9 34,2 9,2 8,7
Hutan pegunungan rendah 62,2 58,2 1,0 0,7 17,8 17,8 Hutan pegunungan tinggi 2,0 1,9 0,6 1,1 7,5 9,2 Belukar/hutan sekunder muda 93,6 93,3 3,6 4,6 54,1 62,3 56,7 55,1
Mosaik dataran rendah 0,0 0,1 0,0 0,0 0,1 0,7 8,7 8,9
Mosaik pegunungan 0,0 0,0 0,0 0,0 0,1 0,3 Dataran rendah terbuka 0,0 0,3 0,0 0,0 0,0 0,0 0,0 0,0 Pegunungan terbuka 0,3 0,0 Areal urban Perkebunan skala besar 0,0 0,0 0,0 1,0 Total (%) 100 100 100 100 100 100 100 100 N data posisi 1.351 1.601 608 673 2.679 3.287 2.949 3.167
Tidak ada perbedaan yang signifikan dalam penggunaan habitat pada waktu siang dan malam hari diantara individu harimau (Tabel 13). Tidak adanya perbedaan ini mungkin disebabkan oleh sifat harimau yang cenderung krepuskular, yakni aktif pada waktu peralihan antara terang dan gelap. Selain itu, hal ini mungkin juga terjadi karena harimau merupakan satwa top predator di hutan Sumatera yang tidak memiliki kompetitor, sehingga ada alasan bagi harimau untuk memilih waktu dalam penggunaan habitat utama. Simcharoen et al. (2008) yang mendapati bahwa macan tutul di Thailand menggunakan berbagai tipe habitat dengan proporsi yang berbeda antara siang dan malam. Hal ini terjadi kemungkinan diakibatkan oleh adanya harimau yang hidup simpatrik dengan macan tutul, dimana macan tutul menggunakan satu tipe habitat ketika harimau tidak menggunakan habitat
95
tersebut. Sunquist (1981) menyatakan bahwa macan tutul di TN Chitwan, Nepal, aktif pada malam hari karena pola aktivitasnya mungkin dipengaruhi oleh keberadaan harimau. Begitu juga dengan macan tutul di TN Tsavo, Kenya, yang juga aktif pada malam hari akibat adanya singa yang hidup simpatrik (Hamilton 1976).
Tabel 13. Hasil uji Wilcoxon untuk melihat perbedaan penggunaan habitat/tutupan vegetasi oleh harimau pada siang (S) dan malam (M) hari di masing-masing lokasi translokasi.
Harimau Lokasi Hasil uji Wilcoxon Habitat utama (% penggunaan
siang dan malam)
JD-1 TNBBS Z= -0,211; P= 0,883 Belukar/hutan sekunder muda (siang= 93,6: malam= 93,3) JD-3 TNGL Z= 0,000; P= 1,000 Hutan pegunungan rendah (siang=
62,2: malam= 58,2)
JD-5 TNKS Z= -0,315; P= 0,752 Hutan dataran rendah (siang= 54,1: malam= 62,3)
BD-1 EUM Z= -0,135; P= 0,892 Belukar/hutan sekunder muda (siang= 56,7: malam= 55,1)
5.4 Model Kesesuaian Habitat
5.4.1 Penentuan Titik Presence dan Pseudo-absence
Jumlah total titik posisi koordinat yang berhasil dikumpulkan (titik presence) melalui kalung GPS dari harimau BD-1 yang ditranslokasikan di kawasan hutan Ulu Masen (KHUM) adalah 6.880 titik posisi. Namun, setelah diseleksi ternyata jumlah posisi harimau yang memiliki akurasi tinggi adalah 6.116 titik. Sebanyak 3.058 titik posisi presence (50% dari jumlah total titik) digunakan untuk membangun model, sedangkan 50% lainnya digunakan untuk validasi model (Gambar 20).
Penyusunan model regresi logistik biner membutuhkan titik posisi kehadiran harimau translokasi (presence = 1) dan titik yang diduga tidak dikunjungi harimau (pseudo-absence = 0). Penetapan titik pseudo-absence untuk penyusunan model ataupun validasi model dilakukan secara acak menggunakan grid 30m x30m, luar poligon batas kawasan hutan Ulu Masen dan di luar poligon daerah jelajah harimau studi yang telah diberi buffer. Jumlah titik pseudo-absence yang ditetapkan untuk menyusun model dan
96
validasi model masing-masing 3.058 titik. Menurut Putri (2010) pemberian buffer pada batas kawasan hutan dalam penentuan titik pseudo-absence penting untuk menghindari termasukkannya areal-areal di luar hutan yang juga masih digunakan oleh harimau.
Gambar 20. Peta sebaran titik presence harimau translokasi yang digunakan untuk menyusun model dan validasi model.
5.4.2 Uji Multikolinearitas
Dalam analisis regresi, untuk melihat ada atau tidak adanya multikolinearitas salah satunya dapat dilihat melalui nilai Variance Inflation Factor (VIF) setiap variabel bebas yang diuji (Tabel 14).
Hasil pengujian menunjukkan bahwa ada dua variabel bebas yang memiliki nilai toleransi < 0,1 dan nilai VIF > 10 yaitu “jarak dari jalan” (TOL=0,052; VIF=19,064) dan “jarak dari pemukiman” (TOL=0,042; VIF=23,807). Hal ini memberikan gambaran adanya multikolinearitas antara variabel bebas tersebut dengan variabel bebas lainnya. Permasalahan
97
multikolinearitas (keterhubungan) antar variabel bebas harus ditanggulangi sebelum analisis data dapat dilanjutkan.
Tabel 14. Hasil diagnosa multikolinearitas variabel bebas dengan VIF.
Model Collinearity Statistics
Tolerance (TOL) VIF
1 (Constant)
Ketinggian 0,116 8,587
Jarak dari jalan 0,052 19,064
Jarak dari pemukiman 0,042 23,807
Jarak dari sungai 0,903 1,107
Jarak dari tepi hutan 0,701 1,426
NDVI 0,886 1,129
Kelerengan/slope 0,713 1,402
Multikolinearitas dapat ditanggulangi dengan berbagai cara antara lain penggunaan informasi apriori, analisis komponen utama, analisis faktor, penambahan data serta prosedur mengeluarkan variabel bebas (eliminasi) yang berkolinear ganda. Prosedur penanggulangan multikolinearitas yang digunakan dalam penelitian ini adalah prosedur eliminasi (Ambagau 2010). Berdasarkan prosedur tersebut, maka ada dua variabel bebas yang harus dikeluarkan yaitu “jarak dari jalan” dan “jarak dari pemukiman”. Dengan demikian, hanya lima variabel bebas yang dapat dianalisis lebih lanjut untuk mendapatkan model kesesuaian habitat harimau translokasi berdasarkan persamaan regresi logistik. Variabel-variabel bebas tersebut adalah ketinggian/elevasi, jarak dari sungai, jarak dari tepi hutan, NDVI serta kelerengan/slope.
5.4.3 Analisis Regresi Logistik
Variabel bebas yang digunakan untuk membangun model regresi logistik adalah lima variabel bebas yang tidak memiliki kolinearitas ganda berdasarkan nilai tolerasi dan nilai VIF yaitu ketinggian, jarak dari sungai, jarak dari tepi hutan, NDVI, serta kelerengan. Hasil analisis regresi logistik
98
biner dengan metode Enter pada SPSS 17 dengan taraf kepercayaan 95% disajikan pada Tabel 15.
Pada Tabel 15 ditunjukkan bahwa kelima variabel bebas yang dianalisis seluruhnya memiliki taraf nyata secara statistik (Sig < 0,05). Berdasarkan perhitungan tersebut dapat dilihat nilai-nilai sebagai berikut:
Konstanta = -1,149
Konstanta variabel ketinggian (elv) = -1,563 Konstanta variabel jarak sungai (jsg) = -0,205 Konstanta variabel jarak tepi hutan (jth) = -1,021 Konstanta variabel NDVI (ndvi) = 3,724 Konstanta variabel slope (slp) = 0,062
Dengan demikian persamaan regresi logistik kesesuaian habitat harimau translokasi yang terbentuk adalah sebagai berikut:
Z = -1,149 - (1,563*elv)-(0,205*jsg)-(1,021*jth)+(3,724*ndv)+(0,062*slp)
Tabel 15. Hasil analisis regresi logistik biner dengan metode Enter terhadap variabel bebas.
B S.E. Wald df Sig. Exp(B)
Step 1(a) Ketinggian -1,563 0,064 595,028 1 0,000 ,210 Sungai -,205 0,010 435,361 1 0,000 ,815 Edge/tepi -1,021 0,178 32,947 1 0,000 ,360 NDVI 3,724 0,277 181,124 1 0,000 41,437
Slope ,062 0,004 285,582 1 0,000 1,064
Constant -1,149 0,171 45,010 1 0,000 ,317
Selanjutnya, melalui persamaan yang terbentuk dapat diprediksi kehadiran (presence) harimau sumatera translokasi pada wilayah studi sebagai berikut:
1
P =
1 + e
-(-1,149 - (1,563*elv)-(0,205*jsg)-(1,021*jth)+(3,724*ndv)+(0,062*slp)) Keterangan: P= probabilitas; e adalah bilangan alam = 2,718281899
Secara umum hasil analisis yang disajikan pada Tabel 15 menunjukkan bahwa variabel ketinggian, jarak dari sungai dan jarak dari tepi hutan memberikan pengaruh yang negatif terhadap model regresi yang terbentuk. Semakin tinggi, semakin jauh dari sungai serta semakin jauh satu areal dari tepi hutan, maka semakin tidak sesuai bagi habitat harimau. Sementara, variabel NDVI dan kelerengan memberikan pengaruh positif terhadap model, yang menunjukkan bahwa semakin rapat satu vegetasi (semakin berhutan) dan semakin curam satu areal maka semakin sesuai bagi habitat harimau. Melalui nilai koefisien regresinya, dapat ditentukan bahwa NDVI merupakan variabel lingkungan yang paling berpengaruh pada model, sedangkan kelerengan/slope merupakan variabel yang memberi pengaruh paling kecil terhadap model yang disusun.
Areal studi di hutan Blangraweu, Ulu Masen, merupakan kawasan daratan rendah yang berpadu dengan perbukitan hingga pegunungan dengan variasi ketinggian mulai 0 meter sampai 2.771 meter dpl (Lampiran 16). Pada Tabel 15 dapat dilihat bahwa nilai koefisien regresi ketinggian tempat atau elevasi adalah sebesar -1,563. Ini berarti semakin tinggi satu lokasi maka semakin kecil kemungkinannya bagi kehadiran harimau. Pemodelan yang dilakukan Wibisono et al. (2011) menunjukkan bahwa harimau sumatera umumnya ditemukan pada kawasan dataran rendah. Ketinggian merupakan salah satu faktor lingkungan yang berpengaruh terhadap distribusi dan bentuk tumbuhan yang hidup di daerah pegunungan (Jin et al. 2008). Adanya perbedaan ketinggian menyebabkan terjadinya variasi iklim yang berpengaruh pada keragaman jenis tumbuhan. Kawasan hutan dataran rendah memiliki keragaman tumbuhan yang lebih tinggi dibandingkan dengan dataran tinggi. Keragaman jenis tumbuhan pada satu kawasan berpengaruh terhadap keragaman jenis satwa yang ada di dalamnya. Satwa mangsa utama harimau merupakan herbivora yang memerlukan tumbuhan sebagai sumber pakan.