www.elsevier.com / locate / bres
Research report
Estrogen replacement does not prevent the loss of choline
acetyltransferase-positive cells in the basal forebrain following either
neurochemical or mechanical lesions
*
Payal Aggarwal, Robert B. Gibbs
Department of Pharmaceutical Sciences, University of Pittsburgh School of Pharmacy, 1004 Salk Hall, Pittsburgh, PA 15261, USA
Accepted 8 August 2000
Abstract
Recent studies have shown that estrogen replacement can enhance the functional status of basal forebrain cholinergic neurons. Studies have also shown that estrogen has neuroprotective effects both in vitro and in vivo on a variety of cells and against a variety of insults. The present study examined the ability of estrogen replacement to protect basal forebrain cholinergic neurons from the effects of neurochemical and mechanical injury. Ovariectomized Sprague–Dawley rats received either estrogen replacement or sham surgery, and then received either a unilateral injection of ibotenic acid into the nucleus basalis magnocellularis, or unilateral transection of the fimbria fornix. Cholinergic neurons in the medial septum and nucleus basalis were detected and quantified using immunohistochemical techniques. The data show that neither 3 weeks nor 13 weeks of continuous estrogen replacement prevented the loss of choline acetyltransferase (ChAT)-containing cells in the nucleus basalis following a unilateral injection of ibotenic acid. Likewise, estrogen replacement did not prevent a decrease in ChAT-positive cells detected in the medial septum following unilateral transection of the fimbria fornix. Notably, increased numbers of ChAT-positive cells were detected in the contralateral nucleus basalis, and in the ipsilateral and contralateral medial septum, at 2 weeks following a unilateral injection of ibotenic acid into the nucleus basalis; however, these effects were not related to hormone treatment. These data suggest that estrogen replacement does not protect cholinergic neurons in the medial septum and nucleus basalis from the effects of excitotoxic or mechanical injury. 2000 Elsevier Science B.V. All rights reserved.
Theme: Disorders of the nervous system
Topic: Trauma
Keywords: Hormone replacement; Cholinergic neuron; Neuroprotection; Ibotenic acid
1. Introduction cognitive processes continues to unfold (see Ref. [11] for review).
Several recent prospective studies suggest that es- Recent studies have show that estrogen has significant trogen replacement can help to reduce the risk of Alz- neuroprotective effects in vitro on a wide variety of heimer’s-related dementia in postmenopausal women cultured cells exposed to a variety of toxic insults (see Ref. [3,34,62]. The mechanisms for this effect are still largely [39]). For example, estrogen has been shown to reduce cell unknown, although evidence for significant estrogen ef- death associated with serum deprivation, growth factor fects on neuronal anatomy, biochemistry, and function deprivation, b-amyloid toxicity, excitatory amino acid within regions of the brain that play an important role in toxicity, and various inducers of oxidative stress, in cells ranging from primary neuronal cultures to neuroblastoma and non-neuronal cell lines. In some cases estrogen-me-diated neuroprotection in culture is blocked with selective estrogen receptor antagonists [5,55], suggesting that es-trogen supports survival via the activation of eses-trogen
*Corresponding author. Fax:11-412-624-1850.
E-mail address: gibbsr[email protected] (R.B. Gibbs). receptor regulated genes (e.g. bcl-2 [10]). In other cases,
neuroprotection in culture is related to the structure of the 2. Materials and methods estrogen molecule and not to any specific estrogenicity of
the molecule or to the activation of estrogen receptors 2.1. Animals [4,27–29]. Neuroprotective effects of estrogen in vivo
have also been described. Specifically, several studies have Young female Sprague–Dawley rats were purchased reported the ability of estrogen replacement to reduce from Hilltop Laboratories and housed in pairs on a 12:12-h infarct size and neuronal damage following experimentally light / dark cycle with food and water available ad libitum. induced transient forebrain ischemia [7,53,61,64]. In one All animals were housed for a minimum of 2 weeks before study, the reduction in infarct size was associated with the use. All animals then underwent bilateral ovariectomy and prevention of injury-induced down-regulation of bcl-2 and 2 weeks later, received either 3-mm silastic capsules estrogen receptor beta gene expression [7]. Ischemic injury containing 17b-estradiol crystals (implanted s.c.) or blank was also shown to dramatically upregulate estrogen re- capsules. Studies have shown that these estrogen capsules ceptor alpha gene expression on the injured side. Estrogen produce mean circulating levels of estradiol in the range of has also been shown to reduce kainic acid-induced cell loss 60–120 pg / ml serum.
in the dentate gyrus [2]. These studies suggest that at least
some neuroprotective effects of estrogen in vivo may 2.2. Neurochemical lesions involve the selective regulation of estrogen receptors and
estrogen receptor target genes. Seven to ten days following capsule implantation, One of the consistent biochemical features of Alzheim- animals (n541) were anesthetized with a combination of er’s disease is the loss of cholinergic neurons in the medial ketamine (14 mg / 100 g body wt.) and xylazine (2.8 septum, the diagonal band of Broca, and the nucleus mg / 100 g body wt.) and placed into a standard stereotaxic basalis magnocellularis (see Ref. [33] for review). These device. Twenty animals (ten estrogen (E)-treated, ten non-neurons are the major source of cholinergic innervation to E-treated) received unilateral lesions of the nucleus basalis the hippocampus and neocortex [69] and play an important magnocellularis (NBM) by intracerebroventricular infusion role in learning, memory, and attentional processes of ibotenic acid. Briefly, the skull was exposed and a hole [6,8,36,47,63,66]. Recent studies have shown that estrogen was drilled 1.3 mm posterior to Bregma and 2.4 mm to the replacement can enhance the functional status of basal left of the midline. A stainless steel cannula (28 gauge; forebrain cholinergic projections as indicated by increases Small Parts, Inc.) containing ibotenic acid (10 mg / ml in in choline acetyltransferase mRNA and protein in the sterile filtered saline) was lowered 7.5 mm from the skull medial septum and nucleus basalis magnocellularis into the NBM. One microliter (10 mg) of ibotenic acid [16,12,38], and by increases in high affinity choline uptake solution was infused into the NBM at a rate of 12 ml / h. [46,56] and acetylcholine release [24] in the hippocampus Following infusion, the cannula was left in place for 5 min and overlying cortex. Treatment with estrogen and with and then withdrawn very slowly. The skin overlying the estrogen plus progesterone have also been shown to skull was sutured shut and the animals were placed onto a increase the expression of trkA (nerve growth factor warm heating pad during recovery. Two separate control receptor) mRNA [18,40] in the medial septum and to groups were prepared. Cannula controls (consisting of five increase relative levels of brain-derived neurotrophic factor E-treated and five non-E-treated animals) had an empty mRNA in the hippocampus [18,20,57], suggesting that infusion cannula lowered into the NBM to control for estrogen may enhance basal forebrain cholinergic projec- damage produced by lowering the cannula. Sham controls tions in part by increasing trophic support for the choliner- (consisting of six E-treated and five non-E-treated animals) gic neurons. were anesthetized and placed into the stereotaxic ap-These findings suggest that one mechanism by which paratus, the skull was exposed and then the skin was estrogen replacement can help to reduce the risk of sutured shut. Following surgery, all animals were given Alzheimer’s-related dementia in postmenopausal women is Torbugesic (0.02 cc / 100 g) twice a day for 3 days to by increasing the functional status of cholinergic projec- reduce discomfort associated with the surgical procedures. tions to the hippocampus and neocortex. Whether estrogen
replacement also reduces the loss of cholinergic neurons 2.3. Fimbria /fornix transections associated with Alzheimer’s disease, and / or protects the
incision was made through the dura to expose the brain. method as previously described [23] to verify that the An ultrathin microknife (Tieman) was lowered 6.0 mm unilateral transections were complete.
into the brain at midline. Care was taken to avoid the
midsagittal sinus which was gently pushed to the side. The 2.5. ChAT immunostaining knife was moved laterally along the trench to bone and
then removed. The trench was packed with gelfoam soaked ChAT was detected as previously described [12] using with sterile saline. The skin was sutured shut and the an affinity purified goat polyclonal antibody raised against animals were placed onto a warm heating pad during rat ChAT (Chemicon Pharmaceuticals, AP144P; diluted
recovery. 1:3500 in 50 mM PBS (pH 7.2) containing 0.05% Triton
X-100 and 1% normal rabbit serum). Briefly, sections were 2.4. Tissue preparation placed in the primary antibody solution for 3 days at 48C. The sections were rinsed well with PBS and then incubated Ten of the ibotenate-treated animals (five E-treated and with a biotinylated rabbit-anti-goat antibody (Vector Lab-five non-E-treated), 18 of the fimbria / fornix transected oratories, diluted 1:1000) for 1 h at room temperature. animals (nine E-treated, nine non-E-treated), and all of the Sections were rinsed again with PBS and then incubated controls were killed 2 weeks post-surgery. The remaining with an avidin–HRP complex (Vectastain Elite kit; Vector ten ibotenate-treated animals (five E-treated and five non- Laboratories) for 1 h at room temperature. Sections were E-treated) and 12 fimbria / fornix transected animals (six rinsed with PBS, placed in 50 mM Tris–HCl (pH 7.6) E-treated, six non-E-treated) were killed 12 weeks post- containing 3,39-diaminobenzidine (DAB; 0.5 mg / ml), and surgery. For euthanasia, the animals were anesthetized then reacted with 50 mM Tris containing DAB (0.5 mg / with pentobarbital (100 mg / kg) and perfused with saline ml), H O (0.01%), and NiCl (0.032%) for 7–10 min.2 2 2
followed by 4% paraformaldehyde in 50 mM sodium The sections were then rinsed with PBS, mounted onto acetate (pH 6.5), and then 4% paraformaldehyde in 50 mM Superfrost plus slides (Fisher Scientific), counterstained Tris (pH 9.0). Blood was collected by cardiac puncture with cresyl violet (Sigma), dehydrated, and coverslipped. from two E-treated animals killed after 3 weeks of
estradiol treatment to verify that the estrogen capsules 2.6. Data analysis were producing the expected levels of circulating estradiol.
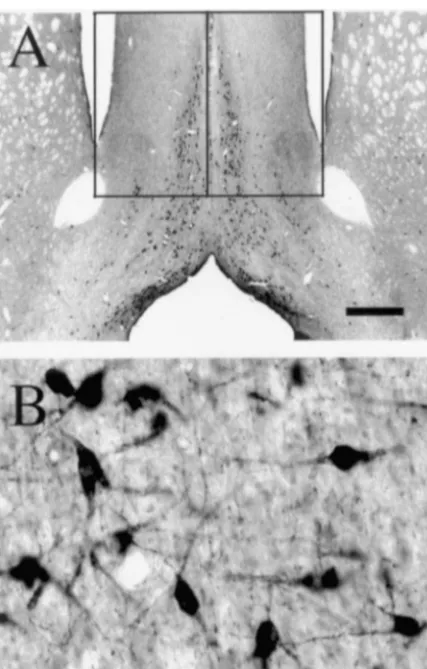
Fig. 1. Bright-field photomicrographs showing ChAT-positive profiles detected in the MS at low (A) and high (B) power. The borders used for stereology are also outlined in A. Scale bars: A51 mm; B562.5mm.
forbidden lines and two inclusive lines. Objects completely inside the counting frame were counted whereas objects completely outside the counting frame were excluded. Objects which crossed through any of the three forbidden planes (bottom, front, and left sides of the frame) were excluded. Objects which crossed through any of the remaining planes (top, right, and back sides of the frame) were included if they either came into focus or went out of focus when focusing through the dissector [68]. The nucleus of each labeled cell was used as the reference point for counting. An estimate of the total number of
Fig. 2. Bright-field photomicrographs showing ChAT staining detected in
ChAT-positive neurons in each region was then calculated the NBM contralateral (A) and ipsilateral (B) to the ibotenate infusion. with the following formula: Total ChAT-positive cells / Arrows in A and B indicate the borders used for stereology. (C)
region5Total profiles counted35(1 / percentage of total High-power magnification of ChAT-positive profiles in the NBM con-tralateral to the lesion. Scale bars: A,B51 mm; C562.5mm.
area)3separation factor (3 for the NBM and 2 for the MS). Statistical analyses were performed using SYSTAT 5.2
for Macintosh. The effects of surgical manipulation (ibote- relative to the controls was analyzed by one-way ANOVA. nate vs. cannula vs. sham) and hormone treatment (E vs. Individual group comparisons were made using the Tukey no E) on the number of ChAT-positive cells in the MS and test.
NBM at 2 weeks post-surgery were analyzed by
three-factor (Hormone Treatment3Surgical Condition3Side) 2.6.2. Fimbria /fornix transected animals
as previously described [12,17,25]. Briefly, the average treated animals. Significant interactions between Surgical number of ChAT-positive profiles on each side of the Condition3Side (F(2,25)5112.7, P,0.001), and Hor-MS / section was determined for each animal by counting mone Treatment3Surgical Condition3Side (F(2,25)5
the number of labeled profiles with an identifiable nucleus 7.01, P,0.005) were also detected.
in each of three matched sections / animal. Group means Subsequent post hoc analyses revealed no significant were calculated and the effects of hormone treatment and interaction between Hormone Treatment3Surgical Con-fimbria transection were compared using analysis of dition on either the intact (F(2,25)52.60, P50.10) or the variance with repeated measures on ‘side’. lesioned (F(2,25)50.26, P50.78) side, contrary to the three-way interaction detected in the ANOVA. Additional post hoc analyses revealed a significant effect of Surgical 3. Results Condition on both the intact (F(2,25)57.30, P,0.005) and
the lesioned (F(2,25)528.62, P,0.005) side.
3.1. Serum estradiol levels A one-way ANOVA comparing the effects of ibotenate in animals killed 2 weeks and 3 months following injection Blood was collected from two E-treated animals and the relative to controls revealed a significant effect of surgical serum levels of 17b-E2 were determined by radioim- condition on the number of ChAT-positive profiles de-munoassay. The serum estradiol levels in these animals tected in the NBM on the lesioned side (F(3,37)530.40, were 85.6 and 115.0 pg / ml, which is consistent with our P,0.001). A significant effect of ibotenate on the number recent studies which show that these capsules produce of ChAT-positive cells detected in the NBM contralateral serum levels of estradiol in the range of 60–120 pg / ml to the lesion was also detected (F(3,37)54.6, P,0.01). ([21]; R.B. Gibbs, unpublished observations). Post hoc analysis revealed a significant decrease in the number of ChAT-positive cells ipsilateral to the lesion at 3.2. Neurochemical lesions both 2 weeks and 3 months following surgery relative to the sham (P,0.001) and cannula (P,0.001) controls (Fig. 3.2.1. Effects on ChAT-positive profiles in the NBM 3). No significant difference between the sham and cannula The number of ChAT-positive cells detected in the controls with respect to the number of ChAT-positive cells NBM on each side of the brain in the ibotenate-injected detected in the NBM ipsilateral to the cannula placement animals and controls is summarized in Table 1. A signifi- was detected (P51.0).
cant loss (approximately 68%) of ChAT positive profiles Post hoc analysis also revealed a significant increase in was detected in the NBM ipsilateral to the ibotenate the number of ChAT-positive cells detected in the NBM injection at 2 weeks post-injection. A similar loss (approxi- contralateral to the lesion in animals killed 2 weeks (P,
mately 63%) was likewise detected at 3 months post- 0.01), but not 3 months (P50.52) post-surgery. More injection. Analysis of the data from animals killed 2 weeks ChAT-positive cells were also detected in the NBM post-surgery revealed a significant main effect of ‘Side’ contralateral to the cannula placement in the cannula (F(1,25)5297.9, P,0.001), but no significant effect of controls than in the sham controls; however, this difference Hormone Treatment (F(1,25)52.6, P50.12) or Surgical was not statistically significant (P50.37).
Condition (F(2,25)51.6, P50.22). No significant
inter-action between Hormone Treatment3Surgical Condition 3.2.2. Effects on chat-positive profiles in the ms
was detected (F(2,25)50.96, P50.40); however, a signifi- The number of ChAT-positive cells detected in the MS cant interaction between Hormone Treatment3Side on each side of the brain in the ibotenate-injected animals (F(1,25)510.8, P,0.005) was detected, reflecting the and controls is summarized in Table 2. Significantly more presence of significantly more ChAT-positive cells in the ChAT-positive cells were detected in the MS both ipsila-NBM contralateral to the lesion in E-treated vs. non-E- teral and contralateral to the NBM lesion at 2 weeks
Table 1
a Mean numbers of ChAT positive profiles in the nucleus basalis magnocellularis
No estrogen Estrogen
Contralateral Ipsilateral Contralateral Ipsilateral
Sham: 2 weeks 8886 84.3 8226 69.0 9286 79.7 7986 46.4
Cannula injury: 2 weeks 10386 99.6 7656100.9 11376 91.5 8586108.5
[
Ibotenate: 2 weeks 10686121.0 2526 28.9 15576154.0 2676118.6*
[
Ibotenate: 12 weeks 9846 79.6 2676 64.0 11376 37.8 3246 92.2*
a
Values indicate mean numbers of profiles6S.E.M. ipsilateral and contralateral to the NBM infusions. n55 in each group with the exception of the estrogen-treated shams where n56. *P,0.001 relative to the intact side of the cannula lesioned and sham controls and to the intact side of estrogen-treated,
[
tralateral (F(3,37)53.3, P50.03) to the NBM lesion. A Tukey test revealed significantly more ChAT-positive cells detected in the MS ipsilateral to the NBM lesion in the 2-week ibotenate group relative to the cannula (P,0.001) and sham (P,0.001) controls. Significantly more ChAT-positive profiles were also detected in the MS ipsilateral to the injection in animals killed 2 weeks vs. 3 months following injection (P,0.001) Contralateral to the NBM lesion, significantly more ChAT-positive cells were de-tected in the MS of animals killed 2 weeks following ibotenate injection relative to the sham controls (P,0.05). Notably, more ChAT-positive cells were detected in the MS both ipsilateral and contralateral to the cannula place-ment in the cannula controls than in the sham controls; however, these differences were not statistically signifi-cant.
Fig. 3. Graph illustrating the effects of surgical condition on the number
3.3. Mechanical lesions
of ChAT-positive profiles in the NBM detected ipsilateral and contralater-al to the ibotenate lesion. Bars represent the margincontralater-al means of ChAT1
profiles6S.E.M. n510 in cannula, ibo-2 and ibo-12 groups; n511 in the An example of AChE staining in the ipsilateral and sham-operated group. *P,0.001 relative to the contralateral side of the
contralateral hippocampus of an animal with a complete
[
sham-treated controls. P,0.001 relative to the ipsilateral side of the
unilateral transection of the fimbria / fornix is shown in Fig.
cannula lesion and sham-treated controls.
4. Of the 30 animals that received unilateral transection of post-injection. Analysis of the data from animals killed 2 the fimbria / fornix, five were excluded from the data weeks post-surgery revealed a significant main effect of analysis (three killed 2 weeks following injury and two Side (F(1,25)515.8, P,0.001), a significant main effect killed 3 months following injury) due to incomplete of Surgical Condition (F(2,25)532.2, P,0.001) and a transections. The number of ChAT-positive cells detected significant Surgical Condition3Side interaction (F(2,25)5 in the MS of the remaining animals ipsilateral and con-8.5, P50.001). Post hoc analysis revealed that the effect of tralateral to the lesion is summarized in Table 3. Fimbria / Surgical Condition on the number of ChAT-positive fornix transection resulted in a significant decrease profiles detected in the MS was significant both ipsilateral (41.166.7% in E-treated animals, 52.367.9% in non-E-(F(2,25)531.80, P,0.001) and contralateral (F(2,25)5 treated animals) in the number of ChAT-positive cells 5.47, P50.01) to the ibotenate injection. No significant detected in the MS ipsilateral to the lesion in animals main effect of Hormone Treatment was detected killed 2 weeks following injury (P,0.01). At 3 months (F(1,25)51.3, P50.28); however, the interaction between following injury, no significant decrease in the number of Hormone Treatment3Side was very close to statistical ChAT-positive cells was detected in the MS on the significance (F(1,25)54.0, P50.06). No significant inter- lesioned vs. the unlesioned side in either E-treated or actions between Hormone Treatment3Surgical Condition non-E-treated animals. Analysis of variance revealed a (F(2,25)50.84, P50.45) or Hormone Treatment3Surgical significant effect of Side (F(1,20)538.1, P,0.001), a Treatment3Side (F(2,25)50.23, P50.80) were detected. significant effect of Time (F(1,20)514.6, P,0.005), and a A one-way ANOVA comparing the effects of ibotenate significant interaction between Side3Time (F(1,20)523.8, in animals killed 2 weeks and 3 months following injection P,0.001). No significant effect of Hormone Treatment relative to controls revealed a significant effect of surgical (F(1,20)53.1, P50.09) and no significant interactions condition ipsilateral (F(3,37)525.9, P,0.001) and con- between Hormone Treatment3Side (F(1,20)52.1, P5
Table 2
a Mean numbers of ChAT-positive profiles in the medial septum
No estrogen Estrogen
Contralateral Ipsilateral Contralateral Ipsilateral
Sham: 2 weeks 7266 38.4 6486 52.5 7956 66.1 8606100.9
Cannula injury: 2 weeks 7926 51.6 7926 67.5 7806 49.3 10986166.1
Ibotenate: 2 weeks 11946187.4* 16686160.9* 9726156.1* 17886176.9*
Ibotenate: 12 weeks 8106109.4 7206100.0 9966159.2 8106121.1
a
cell loss. In addition, no recovery in the number of ChAT-IR cells was detected on the injected side at 3 months following injection in either E-treated or non-E-treated animals, suggesting that estrogen treatment did not reduce cell death or enhance recovery of the cholinergic pheno-type following ibotenic acid injection.
The data also demonstrate that estrogen replacement did not prevent the loss of ChAT-positive cells in the MS following mechanical injury to the fimbria / fornix. This is demonstrated by the fact that unilateral transection of the fimbria / fornix produced a significant decrease in the number of ChAT-positive cells detected in the MS on the injured side at 2 weeks post-injury, and that estrogen treatment had no significant effect on the magnitude of the cell loss. Unlike the effects of ibotenate on ChAT-positive cells in the NBM, the number of ChAT-positive cells in the MS recovered to control levels within 3 months following fimbria / fornix transection. This is consistent with previous reports [31,42,49] and suggests that the decrease in ChAT-positive cells in the MS following fimbria / fornix transection is due to an injury-related decrease in ChAT expression as opposed to a loss of cells. ChAT expression then recovers over time resulting in an increase in the number of ChAT-positive cells to control levels. Estrogen replacement had no apparent effect on the
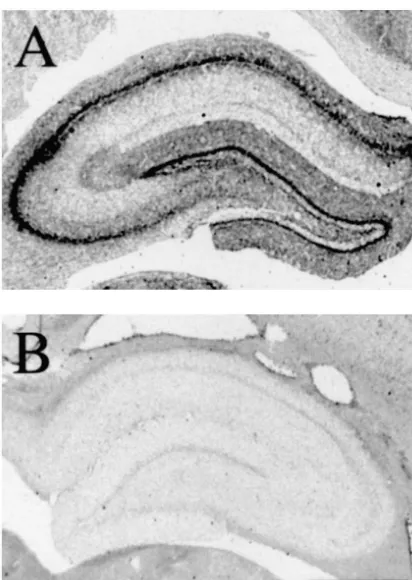
Fig. 4. Photomicrographs showing AchE staining in the hippocampus number of ChAT-positive cells detected in the MS on the
contralateral (A) and ipsilateral (B) to the fimbria / fornix transection injured side at 3 months following injury. Therefore, found in an animal killed 2 weeks following transection. Note the
despite the evidence that estrogen replacement can enhance
presence of AchE-positive staining through the hippocampus contralateral
the functional status of cholinergic neurons in the MS and
to the lesion, compared with the absence of staining ipsilateral to the
NBM [14], these data suggest that hormone replacement
lesion. Scale bar51 mm in both.
does not prevent the loss of cholinergic neurons or the loss of ChAT expression following either neurochemical or 0.16) or Hormone Treatment3Side3Time (F(1,20)50.05, mechanical injury. It should be noted that this does not
P50.82) were detected. preclude the possibility that estrogen enhances other measures of cholinergic function within the remaining cholinergic neurons such as increasing high affinity choline 4. Discussion uptake or acetylcholine production and release.
One possibility is that estrogen failed to reduce the toxic The data demonstrate that estrogen replacement did not effects of ibotenic acid because either the dose of estrogen prevent the loss of cholinergic neurons in the NBM was too low, or the dose of ibotenic acid was too high. following injections of ibotenic acid. This is demonstrated Previous studies, including additional data collected from by the fact that unilateral injection of ibotenic acid into the over one hundred animals (unpublished observations), have NBM produced a significant decrease in the number of shown that the estrogen capsules that were used produce ChAT-positive cells detected in the NBM on the injected mean circulating levels of estradiol in the high physiologi-side within 2 weeks post-injection, and that estrogen cal range (60–120 pg / ml). This was confirmed in two treatment had no significant effect on the magnitude of the E-treated animals that had serum levels of estradiol of 85.6
Table 3
a Mean numbers of ChAT-positive profiles in the medial septum following fimbria / fornix transection
Survival time No estrogen Estrogen
following transection
Contralateral Ipsilateral Contralateral Ipsilateral
2 weeks 55.7611.3 29.9614.2* 54.76 7.7 22.569.9*
12 weeks 64.6614.3 65.6621.7 56.4611.2 48.6613.4
a
and 115.0 pg / ml. This suggests that estrogen levels were direct estrogen effects on the cholinergic neurons mediated sufficient to elicit any neuroprotective effect that would be by the activation of classical estrogen receptors. Recent associated with physiological levels of circulating es- studies have shown that a significant percentage of the tradiol. Furthermore, a preliminary study (unpublished cholinergic neurons in the MS contain estrogen receptor observations) showed that half the dose of ibotenic acid alpha mRNA and protein and bind 17b-estradiol produced much less cell loss overall, and much less of a [15,41,52], suggesting that estrogen may directly effect loss of ChAT-positive cells (less than half the degree of ChAT expression via ligand-mediated activation of the cell loss) than was obtained in the present study. This estrogen receptor. It is possible that mechanical injury suggests that the extent to which the dose–response curve induces cellular responses that prevent estrogen receptor-would need to be affected in order to elicit a significant mediated activation of the ChAT gene. One interesting neuroprotective effect was not so great as to account for study would be to administer estrogen 3 months following the lack of an effect of estrogen. In addition, Singer et al. transection of the fimbria / fornix when the injury response [55] reported that estrogen significantly protected cortical has subsided and the number of ChAT-positive cells in the neurons from glutamate-induced cell death in culture using MS has recovered. An estrogen-mediated induction of glutamate levels as high as 0.1 mM. Azcoitia et al. [2] ChAT expression at this time could indicate that the reported that estrogen reduced cell loss in the dentate gyrus inability to induce ChAT expression 2 weeks following following systemic injection of 7 mg / kg kainic acid. These injury is due to an injury-induced response which blocks data suggest that the concentration of ibotenate used in the estrogen-mediated transcriptional activation as opposed to present study was not so overwhelming as to preclude the disconnection from the hippocampus.
ability to detect a neuroprotective effect of estrogen. Estrogen has also been shown to exert neuroprotective Nevertheless, the possibility that estrogen has neuroprotec- effects via non-classical mechanisms, possibly involving tive effects on the cholinergic neurons when administered activation of specific second messenger systems such as at different doses or for different periods of time, or that ERK and MEK. Several studies have documented rapid estrogen protects the cholinergic neurons from other types membrane effects of estrogen in cells which lack estrogen of neurotoxic or mechanical injury, cannot be excluded. receptors, resulting in neuroprotection or in the activation Notably, estrogen treatment did increase the number of of ERK and MEK [54,58,59,65]. Whether estrogen acts on ChAT-positive cells detected in the NBM contralateral to basal forebrain cholinergic neurons via non-classical path-the ibotenate injection, and in path-the MS ipsilateral to path-the ways and whether those effects are altered following NBM injection, at 2 weeks post-injury. We interpret these neurochemical or mechanical injury are currently un-increases as un-increases in ChAT protein within cholinergic known.
growth factor expression after chronic neurotoxic injury — I.
Ref. [13]). Therefore, it is reasonable to suspect that
Degeneration and plastic response of basal forebrain neurons,
injury-induced increases in NGF contribute at least in part
Neuroscience 65 (1995) 633–645.
to the increases in ChAT-positive cells detected in the MS [2] I. Azcoitia, A. Sierra, L.M. Garcia-Segura, Estradiol prevents kainic and in the contralateral NBM. acid-induced neuronal loss in the rat dentate gyrus, Neuroreport 9
Even so, the increases in the numbers of ChAT-positive (1998) 3075–3079.
[3] M. Baldereschi, A. Di Carlo, V. Lepore, L. Bracco, S. Maggi, F.
cells detected in the ipsilateral MS of E-treated and
non-E-Grigoletto, G. Scarlato, L. Amaducci, Estrogen-replacement therapy
treated animals 2 weeks following an injection of ibotenate
and Alzheimer’s disease in the Italian Longitudinal Study on Aging,
into the NBM were quite striking (2–3 fold). This
indi-Neurology 50 (1998) 996–1002.
cates a transient, but powerful, effect of the ibotenate [4] C. Behl, T. Skutella, F. Lezoualch, A. Post, M. Widmann, C.J. injections on cholinergic neurons in the ipsilateral MS. Newton, F. Holsboer, Neuroprotection against oxidative stress by This may be caused by the leakage of some ibotenate up estrogens: structure–activity relationship, Mol. Pharmacol. 51
(1997) 535–541.
the cannula tract into the hippocampus, resulting in
[5] J.A. Chowen, I. Torres-Aleman, L.M. Garcia-Segura, Trophic
excitation of the hippocampal neurons. Previous studies
effects of estradiol on fetal rat hypothalamic neurons,
Neuro-have shown that increased electrical activity in the
hip-endocrinology 56 (1992) 895–901.
pocampus results in the up-regulation of both NGF and [6] J.A.M. Dekker, D.J. Connor, L.J. Thal, The role of cholinergic BDNF within hippocampal neurons [9,32,35,70]. Any projections from the nucleus basalis in memory, Neurosci. leakage of ibotenate up the cannula tract and into the Biobehav. Rev. 15 (1991) 299–317.
[7] D.B. Dubal, P.J. Shughrue, M.E. Wilson, I. Merchenthaler, P.M.
hippocampus would undoubtedly produce increased
neuro-Wise, Estradiol modulates bcl-2 in cerebral ischemia: a potential
nal activity within the hippocampus, which could result in
role for estrogen receptors, J. Neurosci. 19 (1999) 6385–6393.
increased NGF and BDNF expression and increased
[8] H.C. Fibiger, Cholinergic mechanisms in learning, memory and
trophic support for cholinergic neurons in the MS. This dementia: a review of recent evidence, Trends Neurosci. 14 (1991) could account for the dramatic and transient increase in the 220–223.
number of ChAT-positive cells detected in the MS ipsila- [9] C.M. Gall, P.J. Isackson, Limbic seizures increase neuronal pro-duction of messenger RNA for nerve growth factor, Science 245
teral to the ibotenate injections. Additional studies which
(1989) 758–761.
examine changes in hippocampal and cortical levels of
[10] L.M. Garcia-Segura, P. Cardona-Gomez, F. Naftolin, J.A. Chowen,
NGF and BDNF following unilateral ibotenic acid
in-Estradiol upregulates Bcl-2 expression in adult brain neurons,
jections into the NBM would reveal the extent to which NeuroReport 9 (1998) 593–597.
changes in neurotrophin expression may contribute to the [11] R.B. Gibbs, Effects of estrogen on basal forebrain cholinergic increased numbers of ChAT-positive cells detected in the neurons and cognition: Implications for brain aging and dementia in women, in: M. Morrison (Ed.), Hormones, Aging, and Mental
MS.
Disorders, Cambridge University Press, Cambridge, 2000, pp. 183–
In conclusion, the present studies failed to provide
222.
evidence for an ability of estrogen replacement to prevent
[12] R.B. Gibbs, Effects of estrogen on basal forebrain cholinergic
the loss of ChAT-positive neurons in the MS and NBM neurons vary as a function of dose and duration of treatment, Brain following excitatory amino acid toxicity or mechanical Res. 757 (1997) 10–16.
injury. Whether estrogen is neuroprotective for the [13] R.B. Gibbs, Estrogen and nerve growth factor-related systems in brain: effects on basal forebrain cholinergic neurons and
implica-cholinergic cells against other types of injury such as
tions for learning and memory processes and aging, in: V.N. Luine,
b-amyloid-induced toxicity or cortical impact injury, and
C.F. Harding (Eds.), Hormonal Restructuring of the Adult Brain,
whether estrogen can reduce the loss of cholinergic
Basic and Clinical Perspectives, Vol. 743, New York Academy of
neurons associated with normal aging, are still unknown. Science, New York, 1994, pp. 165–199.
Hence, the possibility that neuroprotective effects of [14] R.B. Gibbs, Oestrogen and the cholinergic hypothesis: Implications
estrogen on basal forebrain cholinergic neurons contribute for oestrogen replacement therapy in postmenopausal women, in: J. Goode (Ed.), Neuronal and Cognitive Effects of Oestrogens,
Novar-to the ability of estrogen replacement Novar-to reduce the risk
tis Found. Symposium 230, Wiley, Chichester, 2000, pp. 94–111.
for developing Alzheimer’s-related dementia in
post-[15] R.B. Gibbs, Expression of estrogen receptor-like immunoreactivity
menopausal women cannot be excluded. by different subgroups of basal forebrain cholinergic neurons in
gonadectomized male and female rats, Brain Res. 720 (1996) 61–68.
[16] R.B. Gibbs, Fluctuations in relative levels of choline
acetyltrans-Acknowledgements
ferase mRNA in different regions of the rat basal forebrain across the estrous cycle: effects of estrogen and progesterone, J. Neurosci.
Estradiol radioimmunoassays were performed by the 16 (1996) 1049–1055.
Assay Core of the Center for Reproductive Physiology. [17] R.B. Gibbs, Impairment of basal forebrain cholinergic neurons
This work was supported by NIH Grant NS28896. associated with aging and long-term loss of ovarian function, Exp. Neurol. 151 (1998) 289–302.
[18] R.B. Gibbs, Levels of trkA and BDNF mRNA, but not NGF mRNA, fluctuate across the estrous cycle and increase in response to acute
References hormone replacement, Brain Res. 787 (1998) 259–268.
[19] R.B. Gibbs, Long-term treatment with estrogen and progesterone ¨
[20] R.B. Gibbs, Treatment with estrogen and progesterone affects [39] B.S. McEwen, S.E. Alves, Estrogen actions in the central nervous relative levels of brain-derived neurotrophic factor mRNA and system, Endocr. Rev. 20 (1999) 279–307.
protein in different regions of the adult rat brain, Brain Res. 844 [40] P.J. McMillan, C.A. Singer, D.M. Dorsa, The effects of ovariectomy (1999) 20–27. and estrogen replacement on trkA and choline acetyltransferase [21] R.B. Gibbs, A.M. Burke, D.A. Johnson, Estrogen replacement mRNA expression in the basal forebrain of adult female Sprague–
attenuates effects of scopolamine and lorazepam on memory acqui- Dawley rat, J. Neurosci. 16 (1996) 1860–1865.
sition and retention, Horm. Behav. 34 (1998) 112–125. [41] E.J. Mufson, W.J. Cai, S. Jaffar, E.Y. Chen, G. Stebbins, T. Sendera, [22] R.B. Gibbs, M.V. Chao, D.W. Pfaff, Effects of fimbria-fornix and J.H. Kordower, Estrogen receptor immunoreactivity within
subreg-NGFR
angular bundle transection on expression of the p75 mRNA by ions of the rat forebrain: neuronal distribution and association with cells in the medial septum and diagonal band of Broca: correlations perikarya containing choline acetyltransferase, Brain Res. 849 with cell survival, synaptic reorganization and sprouting, Mol. Brain (1999) 253–274.
Res. 11 (1991) 207–219. [42] T. Naumann, G.M. Peterson, M. Frotscher, Fine structure of rat [23] R.B. Gibbs, E.W. Harris, C.W. Cotman, Replacement of damaged septohippocampal neurons: II. A time course analysis following
cortical projections by homotypic transplants of entorhinal cortex, J. axotomy, J. Comp. Neurol. 325 (1992) 219–242.
Comp. Neurol. 237 (1985) 47–64. [43] E.A. Nimchinsky, P.R. Hof, W.G. Young, F.E. Bloom, J.H. Mor-[24] R.B. Gibbs, A. Hashash, D.A. Johnson, Effects of estrogen on
rison, NeuroZoom software: development, validation, and neuro-potassium-evoked acetylcholine release in the hippocampus and
biological application, FASEB J. 12 (1998) A628. overlying cortex of adult rats, Brain Res. 749 (1997) 143–146.
[44] A. Nitta, Y. Furukawa, K. Hayashi, T. Hasegawa, T. Nabeshima, [25] R.B. Gibbs, D.W. Pfaff, Effects of estrogen and fimbria / fornix
NGFR Nerve growth factor content of rat brain increases following basal transection on p75 and ChAT expression in the medial septum
forebrain lesions induced by ibotenic acid, but not by electrolysis, and diagonal band of Broca, Exp. Neurol. 116 (1992) 23–39.
Biol. Pharm. Bull. 17 (1994) 34–38. [26] J.R. Goss, M.E. O’Malley, L. Zou, S.D. Styren, P.M. Kochanek, S.T.
[45] T. Nonomura, H. Hatanaka, Neurotrophic effect of brain-derived DeKosky, Astrocytes are the major source of nerve growth factor
neurotrophic factor on basal forebrain cholinergic neurons in culture upregulation following traumatic brain injury in the rat, Exp. Neurol.
from postnatal rats, Neurosci. Res. 14 (1992) 226–233. 149 (1998) 301–309.
[46] C.A. O’Malley, R.D. Hautamaki, M. Kelley, E.M. Meyer, Effects of [27] P.S. Green, J. Bishop, J.W. Simpkins, 17-alpha-estradiol exerts
ovariectomy and estradiol benzoate on high affinity choline uptake, neuroprotective effects on SK-N-SH cells, J. Neurosci. 17 (1997)
ACh synthesis, and release from rat cerebral cortical synaptosomes, 511–515.
Brain Res. 403 (1987) 389–392. [28] P.S. Green, K. Gordon, J.W. Simpkins, Phenolic A ring requirement
[47] D.S. Olton, Dementia: animal models of the cognitive impairments for the neuroprotective effects of steroids, J. Steroid Biochem. Mol.
following damage to the basal forebrain cholinergic system, Brain Biol. 63 (1997) 229–235.
Res. Bull. 25 (1990) 499–502. [29] P.S. Green, K.E. Gridley, J.W. Simpkins, Nuclear estrogen
receptor-independent neuroprotection by estratrienes — a novel interaction [48] G. Paxinos, C. Watson, The Rat Brain in Stereotaxic Coordinates, with glutathione, Neuroscience 84 (1998) 7–10. 2nd ed., Academic Press, London, 1986.
[30] H.J. Gundersen, P. Bagger, T.F. Bendtsen, S.M. Evans, L. Korbo, N. [49] G.M. Peterson, T. Naumann, M. Frotscher, Identified septohip-Marcussen, A. Moller, K. Nielsen, J.R. Nyengaard, B. Pakkenberg et pocampal neurons survive axotomy: a fine-structural analysis in the al., The new stereological tools: disector, fractionator, nucleator and rat, Neurosci. Lett. 138 (1992) 81–85.
point sampled intercepts and their use in pathological research and [50] J.P. Schwartz, N. Nishiyama, Neurotrophic factor gene expression in diagnosis, APMIS 96 (1988) 857–881. astrocytes during development and following injury, Brain Res. [31] C.A. Haas, T. Deller, T. Naumann, M. Frotscher, Selective expres- Bull. 35 (1994) 403–407.
sion of the immediate early gene c-jun in axotomized rat medial [51] L.W. Shaughnessy, S.J. Barone, Damage to the NBM leads to a septal neurons is not related to neuronal degeneration, J. Neurosci. sustained lesion-induced increase in functional NGF in the cortex,
16 (1996) 1894–1903. NeuroReport 8 (1997) 2767–2774.
[32] P.J. Isackson, M.M. Huntsman, K.D. Murray, C.M. Gall, BDNF [52] P.J. Shughrue, P.J. Scrimo, I. Merchenthaler, Estrogen bonding and mRNA expression is increased in adult rat forebrain after limbic estrogen receptor characterization (ERaand ERb) in the cholinergic seizures: temporal patterns of induction distinct from NGF, Neuron neurons of the rat basal forebrain, Neuroscience 96 (2000) 41–49. 6 (1991) 937–948. [53] J.W. Simpkins, G. Rajakamar, Y.Q. Zhang, C.E. Simpkins, D. [33] P. Kasa, Z. Rakonczay, K. Gulya, The cholinergic system in Greenwald, C.J. Yu, N. Bodor, A.L. Day, Estrogens may reduce Alzheimer’s disease, Prog. Neurobiol. 52 (1997) 511–535. mortality and ischemic damage caused by middle cerebral artery [34] C. Kawas, S. Resnick, A. Morrison, R. Brookmeyer, M. Corrada, A. occlusion in the female rat, J. Neurosurg. 87 (1997) 724–730.
Zonderman, C. Bacal, D.L. Lingle, E. Metter, A prospective study of [54] C.A. Singer, X.A. Figueroa-Masot, R.H. Batchelor, D.M. Dorsa, The estrogen replacement therapy and the risk of developing Alzheim- mitogen-activated protein kinase pathway mediates estrogen neuro-er’s disease: The Baltimore Longitudinal Study of Aging, Neurology protection after glutamate toxicity in primary cortical neurons, J.
48 (1997) 1517–1521. Neurosci. 19 (1999) 2455–2463.
[35] J.C. Lauterborn, P.J. Isackson, C.M. Gall, Seizure-induced increases [55] C.A. Singer, K.L. Rogers, T.M. Strickland, D.M. Dorsa, Estrogen in NGF mRNA exhibit different time-courses across forebrain protects primary cortical neurons from glutamate toxicity, Neurosci. regions and are biphasic in hippocampus, Exp. Neurol. 125 (1994) Lett. 212 (1996) 13–16.
22–40. [56] M. Singh, E.M. Meyer, W.J. Millard, J.W. Simpkins, Ovarian steroid [36] A.D. Lawrence, B.J. Sahakian, Alzheimer disease, attention, and the deprivation results in a reversible learning impairment and compro-cholinergic system, Alzheimer Dis. Assoc. Disord. 9 (Suppl. 2) mised cholinergic function in female Sprague–Dawley rats, Brain
(1995) 43–49. Res. 644 (1994) 305–312.
[37] R.M. Lindsay, P.C. Barber, M.R. Sherwood, J. Zimmer, G. Raisman, [57] M. Singh, E.M. Meyer, J.W. Simpkins, The effects of ovariectomy Astrocyte cultures from adult rat brain. Derivation, characterization and estradiol replacement on brain-derived neurotrophic factor and neurotrophic properties of pure astroglial cells from corpus messenger ribonucleic acid expression in cortical and hippocampal callosum, Brain Res. 243 (1982) 329–343. brain regions of female Sprague–Dawley rats, Endocrinology 136 [38] V.N. Luine, Estradiol increases choline acetyltransferase activity in (1995) 2320–2324.
cerebral cortical explants: convergence of estrogen and neurotrophin perfusion-independent mechanisms in rats, Stroke 30 (1998) 630– signaling pathways, J. Neurosci. 19 (1999) 1179–1188. 637.
[59] M. Singh, G. Setalo, X.P. Guan, D.E. Frail, C.D. ToranAllerand, [65] J.J. Watters, J.S. Campbell, M.J. Cunningham, E.G. Krebs, D.M. Estrogen-induced activation of the mitogen-activated protein kinase Dorsa, Rapid membrane effects of steroids in neuroblastoma cells: cascade in the cerebral cortex of estrogen receptor-alpha knock-out effects of estrogen on mitogen activated protein signaling cascade mice, J. Neurosci. 20 (2000) 1694–1700. and c-fos immediate early gene transcription, Endocrinology 138 [60] S. Strauss, U. Otten, B. Joggerst, K. Pluss, B. Volk, Increased levels (1997) 4030–4033.
of nerve growth factor (NGF) protein and mRNA and reactive [66] G.L. Wenk, The nucleus basalis magnocellularis cholinergic system: gliosis following kainic acid injection into the rat striatum, Neurosci. one hundred years of progress, Neurobiol. Learn. Mem. 67 (1997)
Lett. 168 (1994) 193–196. 85–95.
¨
[61] S. Sudo, T.C. Wen, J. Desaki, S. Matsuda, J. Tanaka, T. Arai, N. [67] H.R. Widmer, B. Knusel, F. Hefti, BDNF protection of basal Maeda, M. Sakanaka, Beta-estradiol protects hippocampal CA1 forebrain cholinergic neurons after axotomy: complete protection of neurons against transient forebrain ischemia in gerbil, Neurosci. Res. p75NGFR-positive cells, NeuroReport 4 (1993) 363–366. 29 (1997) 345–354. [68] R.W. Williams, P. Rakic, Three-dimensional counting: an accurate [62] M.-X. Tang, D. Jacobs, Y. Stern, K. Marder, P. Schofield, B. and direct method to estimate numbers of cells in sectioned material, Gurland, H. Andrews, R. Mayeux, Effect of oestrogen during J. Comp. Neurol. 278 (1988) 344–352, published erratum appears in menopause on risk and age at onset of Alzheimer’s disease, Lancet J. Comp. Neurol. 281 (1989) 335).
348 (1996) 429–432. [69] N.J. Woolf, Cholinergic systems in mammalian brain and spinal [63] M.L. Voytko, Cognitive functions of the basal forebrain cholinergic cord, Prog. Neurobiol. 37 (1991) 475–524.
system in monkeys: memory or attention? Behav. Brain Res. 75 [70] F. Zafra, B. Hengerer, J. Leibrock, H. Thoenen, D. Lindholm, (1996) 13–25. Activity dependent regulation of BDNF and NGF mRNAs in the rat [64] Q. Wang, R. Santizo, V.L. Baughman, D.A. Pelligrino, Estrogen hippocampus is mediated by non-NMDA glutamate receptors,