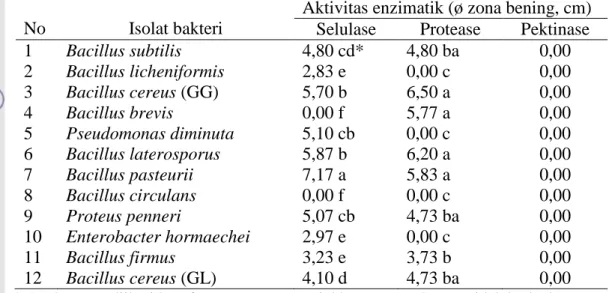
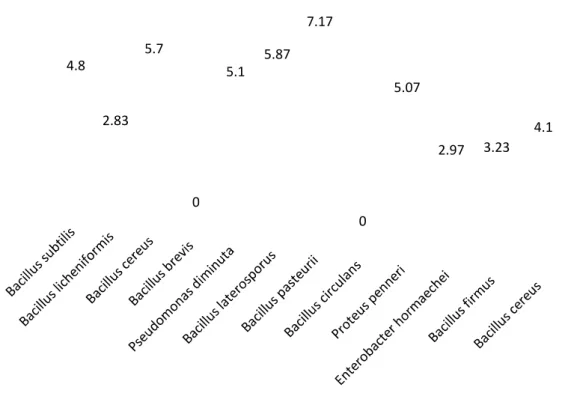
DIVERSITAS BAKTERI ASAL SPORA FUNGI MIKORIZA ARBUSKULA GIGASPORA SP. DAN GLOMUS SP. SERTA POTENSINYA SEBAGAI MYCORRHIZA HELPER BACTERIA
Teks penuh
Gambar




Dokumen terkait
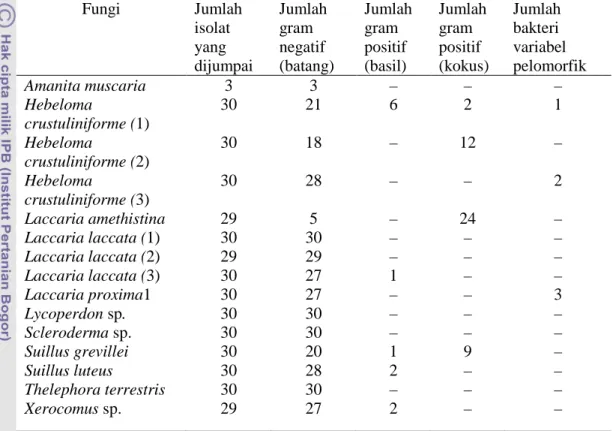
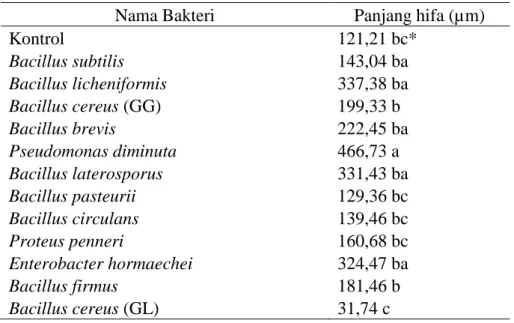
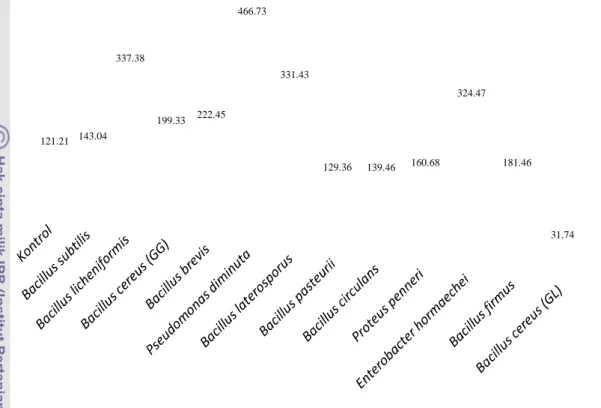
Hasil penelitian menunjukkan bahwa terjadi peningkatan rata-rata kepadatan spora dari lapangan pada hasil trapping , untuk persentase kolonisasi fungi mikoriza arbuskula pada
Hasil penelitian status dan keanekaragaman fungi mikoriza arbuskula (FMA) pada tanah bekas letusan Gunung Sinabung di Kabupaten Karo memiliki 2 genus spora yaitu, Acalauspora 3
Penelitian ini bertujuan untuk Mengidentifikasi jenis fungi mikoriza arbuskula (FMA) berdasarkan ukuran, warna, ornamen dari spora FMA yang di temukan pada sampel tanah pada
Tujuan dari penelitian ini adalah untuk mengetahui kepadatan spora fungi mikoriza arbuskula, persentase kolonisasi akar, dan tipe spora pada perkebunan kopi.. Sampel tanah dan
YUDHA PRANATA: Status dan Keanekaragaman Fungi Mikoriza Arbuskula (FMA) pada Tanah Bekas Letusan Gunung Sinabung di Kabupaten Karo, dibimbing oleh DENI ELFIATI dan DELVIAN..
Pemanfaatan Fungi Mikoriza Arbuskula dan Fungi Pelarut Fosfat untuk Meningkatkan Pertumbuhan Bibit Glodokan (Polyalthia longifolia) pada Tanah Marginal.. Dibimbing
Keanekaragaman fungi mikoriza arbuskula (FMA) di bawah tegakan tanaman jabon Pemalang Jawa Tengah diantaranya terdapat 3 genus yaitu Glomus, Acaulospora dan
Penelitian ini bertujuan untuk mengetahui kepadatan spora, persentase kolonisasi, dan tipe fungi mikoriza arbuskula (FMA) pada areal tanaman karet di PTPN III Kebun Batang Toru
